
bright_id
stringlengths 11
257
| bright_doc
stringlengths 25
9.18M
| bright_split
stringclasses 8
values | scandi_id
stringlengths 3
7
| scandi_url
stringlengths 31
143
| scandi_title
stringlengths 1
79
| scandi_text
stringlengths 15
118k
| scandi_language
stringclasses 4
values | scandi_dist
float64 0.2
1.75
|
|---|---|---|---|---|---|---|---|---|
vegetables_bitter/9-bitter-foods-that-are-good-for-you.txt | * Download MyVinmec now
* Patient guide
* Our Hospitals
* Customer service portal
English
English Tiếng Việt
#### Book an appointment easily with the MyVinmec application
INSTALL
* Ha Noi
* Da Nang
* Nha Trang
* Hai Phong
* Ha Long
* Phu Quoc
* Ho Chi Minh
Book visit
via MyVinmec
Make an appointment
* Vinmec Healthcare System
* Vinmec Central Park (Ho Chi Minh City)
* Vinmec Times City (Ha noi)
* Vinmec Da Nang (Da Nang)
* Vinmec Nha Trang (Khanh Hoa)
* Vinmec Hai Phong (Hai Phong)
* Vinmec Ha Long (Quang Ninh)
* Vinmec Phu Quoc (Kien Giang)
* Hospitals & Clinics
* Facilities
* Key Specialties
* Obstetrics & Gynecology and Assisted reproductive technology
* Cardiology
* Pediatrics
* Oncology- Radiotherapy
* General health check
* Gastroenterology - Hepatology
* Rheumatology
* Stem cell and Gene technology
* Vinmec Tissue Bank
* Traditional Medicine
* Doctors
* Packages
* Patient guide
* Working hours
* Hospital charges
* Instruction for appointment
* Rights and responsibilities of patients and their families
* Outpatient examination procedure
* Guideline for inpatient treatment
* Visting hours and regulations
* Insurance
* Others
* About us
* News
* Search
* Pharmaceutical Information
* Tissue bank
* Vinmec Research Institute of Stem Cell and Gene Technology
* Recruiment
* Packages
* Video
* Online store
* ESG
* Download MyVinmec now
* Patient guide
* Customer service portal
* Our Hospitals
*
/news/health-news/nutrition/
# 9 Bitter Foods That Are Good For You
Share:
This is an automatically translated article.
Bitter foods sometimes resonate in culinary circles because of their intense
flavor. However, they can also be frustrating for picky eaters. However,
bitter foods are extremely nutritious and contain a variety of phytochemicals
that have health benefits, including: reducing the risk of diseases: Cancer,
cardiovascular disease and diabetes.
## 1\. Bitter gourd
Bitter melon is a green, rough, cucumber-shaped melon with an extremely bitter
taste. It is commonly used in Asian, African and Caribbean countries but less
commonly in other regions. Bitter gourd contains many phytochemicals such as
triterpenoids, polyphenols, and flavonoids that have been shown to slow the
growth of many types of cancer.
Bitter gourd is one of those bitter foods that is also used in natural
medicine to help lower blood sugar levels in people with diabetes.
A study conducted over a 4-week period showed that consuming 2,000 mg of
dried, powdered bitter gourd daily significantly reduced blood sugar levels in
people with diabetes - but not as much as a type of bitter gourd. common
diabetes medication.
A meta-review study reviewed mixed results in humans and determined that the
evidence was insufficient to recommend bitter melon supplementation for people
with diabetes.
Similar to most other bitter vegetables, bitter melon is rich in antioxidants,
which help prevent cell damage caused by free radicals and may reduce the risk
of heart disease and diabetes.
## 2\. Cruciferous vegetables
The cruciferous family has many bitter vegetables including broccoli, Brussels
sprouts, cabbage, kale, turnips and arugula. These foods contain compounds
called glucosinolates, which give them their bitter taste and provide many of
their health benefits.
Studies have shown that glucosinolate can slow the growth and spread of cancer
cells, but the results obtained have not been consistently repeated in studies
performed in humans.
Nhiều loại rau họ cải có vị đắng và đem lại nhiều lợi ích cho sức khỏe
Some researchers have believed that the difference between people who consume
this vegetable and those who don't may be due to genetic differences between
each person, as well as natural differences in glucosinolate content. due to
growing conditions and cooking methods. However, more in-depth studies are
still needed to prove this.
In addition to the potential anti-cancer effects of cruciferous vegetables,
the glucosinolates in these vegetables help liver enzymes process toxins more
effectively, reducing negative effects on the body.
Although there is no official recommendation for consuming this vegetable,
some studies show that eating at least five servings of cruciferous vegetables
per week is beneficial. You can combine eating spicy food with this vegetable
to increase its attractive taste.
## 3\. Green dandelion
Dandelions are garden weeds, but their leaves are edible and have high
nutritional value.
Blue dandelion is medium sized, vibrant green leaves with irregular edges.
They can be eaten raw with salads, sautéed as a side dish or included in soups
and pastas.
In the composition of green dandelion contains very bitter compounds, so
dandelion greens are often used to balance with other flavors such as garlic
or lemon.
Although the specific health benefits of dandelion greens are poorly studied,
they are rich in vitamins and minerals, including calcium, manganese, iron,
vitamin A, vitamin C, and vitamin K.
Furthermore, green dandelion also contains the carotenoid compounds lutein and
zeaxanthin, which may help protect your eyes from cataracts and macular
degeneration. And, dandelion greens are an excellent source of the prebiotics
inulin and oligofructose, which promote the growth of healthy gut bacteria.
## 4\. Citrus peels
Citrus fruits such as lemons, oranges and grapefruits have a sweet or sour
taste in the flesh, while the outer skin and white flesh are quite bitter.
This can be explained by the presence of flavonoid compounds, which help
protect the fruit from being eaten by insects but have many health benefits.
In fact, the compounds in the citrus peel in particular contain higher levels
of flavonoids than any other component of the fruit. Two of the most abundant
citrus flavonoids are hesperidin and naringin - both of which are powerful
antioxidants.
Test-tube and animal studies suggest that flavonoids in citrus may help fight
cancer by reducing inflammation, improving detoxification, and slowing the
growth and spread of cancer cells, but it's still needed human research.
Hàm lượng flavonoid trong vỏ cam quýt là chất chống oxy hóa mạnh
## 5\. Cranberries
Cranberries are red berries with a sour, bitter taste that can be enjoyed raw,
cooked, dried or juiced. Cranberries contain a class of polyphenols called
type A proanthocyanidins, which can prevent bacteria from attaching to
surfaces, such as body tissues.
This can be beneficial for reducing bacterial tooth decay, reducing the risk
of H. pylori infection in the stomach, and even preventing E.coli infections
in the intestinal tract and urinary tract.
Although many of these studies were done in test tubes or animals, the results
from human-based studies still need further study.
A 90-day study found that drinking about two cups (500ml) of cranberry juice
daily was more than three times more effective at clearing stomach infections
caused by H. pylori than a placebo. Some other studies have also found that a
daily dose of cranberry pills containing at least 36 mg of proanthocyanidins
can significantly reduce urinary tract infections.
In addition to the antibacterial properties of cranberries, this fruit is also
rich in antioxidants. In fact, they contain the highest concentrations of the
24 most commonly eaten fruits. Because of this, drinking cranberry juice
regularly is linked to better heart health, including reduced inflammation,
blood sugar, blood pressure, and triglyceride levels.
## 6\. Cocoa
Cocoa powder is made from the seeds of the cocoa tree and tastes very bitter
when unsweetened. Cocoa is often used in desserts, it is also mixed with cocoa
butter, cocoa liqueur, vanilla and sugar to make chocolate. The study found
that people who ate chocolate at least five times per week had a 56% lower
risk of heart disease, compared with those who didn't use chocolate at all.
Ca cao thường được sử dụng trực tiếp hoặc dùng trong các món tráng miệng
It can be explained by polyphenols and antioxidants found in cocoa, which can
widen blood vessels and reduce inflammation, protect the heart.
Cocoa is also a good source of several trace minerals, including copper,
manganese, magnesium and iron.
Unsweetened cocoa powder, dark chocolate contains the most antioxidants and
the least amount of sugar. Therefore, using cocoa will help to make a healthy
addition to the diet.
## 7\. Coffee
Coffee is one of the most widely consumed beverages in the world and a leading
source of antioxidants in the American diet. As with most bitter foods, coffee
is packed with polyphenols that give the coffee its unique flavor.
One of the most abundant polyphenol compounds in coffee is chlorogenic acid, a
powerful antioxidant that has the potential to provide many of the health
benefits of coffee, including reducing oxidative damage and reducing the risk
of disease. heart disease and diabetes.
Studies show that drinking 3-4 cups of coffee a day can reduce the risk of
death, cancer and heart disease by 17%, 15% and 18% respectively compared to
not drinking coffee at all.
A separate analysis found that each cup of coffee consumed per day reduces the
risk of type 2 diabetes by 7%. Other studies have also found that caffeinated
coffee may help prevent neurological disorders, including Alzheimer's and
Parkinson's disease.
## 8\. Green tea
Green tea is another popular beverage consumed around the world. Green tea is
characterized by its natural bitter taste due to the presence of catechins and
polyphenols. The most well-known catechin compounds are epigallocatechin
gallate, or EGCG.
Studies have found that EGCG can slow the growth of cancer cells, but it is
not yet clear if it has the same effect in humans.
Trà xanh có hương vị đắng tự nhiên do chứa catechin và polyphenol
While some other studies have found that regular consumption of green tea
reduces the risk of some cancers.
Green tea also contains many polyphenol compounds in an active state such as:
Antioxidant and anti-inflammatory.
In fact, drinking just one cup of green tea daily can reduce your risk of
having a heart attack by almost 20%. And choose green tea over black or white
teas for the maximum dose of antioxidants.
## 9\. Red wine
Red wine contains two main polyphenol compounds: Proanthocyanidins and
tannins, which give the wine its rich color and bitter taste. The combination
of alcohol and polyphenol compounds may reduce the risk of heart disease by
reducing cholesterol oxidation, reducing blood clotting, and widening blood
vessels.
Several recent studies have shown that red wine can be good for your gut. In
another study, drinking two glasses of red wine daily for a month increased
the number of healthy gut bacteria. What's more, changes in gut bacteria
counts are directly linked to lower cholesterol levels and reduced
inflammation in the body.
Other benefits of drinking red wine include: Increased life expectancy and
reduced risk of diabetes and osteoporosis. However, keep in mind that
excessive drinking can lead to liver damage and other health problems, so
moderation is important.
Each bitter-tasting food has unique health benefits, including protection
against cancer, heart disease and diabetes, as well as reduced inflammation
and oxidative stress. Most of the benefits from red wine must be attributed to
the wide range of polyphenols, which act as antioxidants, anti-inflammatory
and even prebiotics.
Please dial HOTLINE for more information or register for an appointment
HERE . Download MyVinmec app to make appointments faster and to manage your
bookings easily.
Reference source: healthline.com
XEM THÊM:
* 9 bitter foods good for you
* Dexpanthenol side effects
* What are the benefits of eating gac seeds?
This article is written for readers from Sài Gòn, Hà Nội, Hồ Chí Minh, Phú
Quốc, Nha Trang, Hạ Long, Hải Phòng, Đà Nẵng.
159 reads
Tags: Quả đắng Giảm nguy cơ mắc bệnh Tiểu đường Giảm viêm Thực phẩm đắng
Chống oxy hóa Tim mạch Rau đắng
Relating articles
* 14 benefits and uses of rosemary essential oil
Cây hương thảo (Rosemary) là một loại cây bụi xanh với lá giống như cây kim và
mùi thơm của gỗ. Mặc dù được biết đến như một loại gia vị thực phẩm, cây hương
thảo còn là một trong ...
Readmore
* Uses of the drug Chymoric
Chymorich is a pain reliever and antipyretic drug whose main ingredient is
Alphachymotrypsin. To ensure effective use and avoid unwanted side effects,
patients need to take the drug exactly as directed.
Readmore
* Is it good to eat bitter vegetables?
Hiện nay rau đắng đang dần trở nên rất phổ biến trong bữa ăn gia đình người
việt, khi loại rau này được đánh giá mang đến nhiều công dụng cho sức khỏe.
Vậy ăn rau đắng có tốt không ...
Readmore
* Uses of Dexlacyl
Dexlacyl drug is used to reduce inflammation, anti-allergic in diseases such
as arthritis, nephrotic syndrome, allergic skin diseases, Sarcoid disease, ...
So to find out what is Dexlacyl drug? What should be noted when using? Let's
learn the necessary information ...
Readmore
* Nutritional value of chestnuts
Hạt dẻ (còn có tên khoa học là Aesculus hippocastanum) và các chiết xuất từ
hạt dẻ đều đem lại rất nhiều lợi ích sức khỏe. Thành phần hoạt chất chính
trong hạt dẻ là aescin giúp cải thiện sức ...
Readmore
Maybe you are interested
* 9 bitter foods good for you
* Dexpanthenol side effects
* What are the benefits of eating gac seeds?
* What is Hadocort?
* Uses of Hydrocortisone 10mg
### Follow Us
### About us
* Vinmec System
* Our doctors
* News
* Recruitment
* Sitemap
* Privacy policy
### Vinmec Services
* About us
* Make an appointment
* For customers
* Vinmec Research Institute of Stem Cell and Gene Technology
### Download MyVinmec now
App Store Google Play
#### PARTNER
Đặt câu hỏi cho bác sĩ
Copyright © 2019 Vinmec. All rights reserved
The information on the www.vinmec.com is ONLY for references. Customers SHOULD
NOT arbitrarily apply it at any circumstances. Vinmec is not responsible for
any cases of self-application without a doctor's prescription.
#### Vinmec International Hospital
Joint Stock Company
Contact us
Business Registration Number:0106050554
First registration on November 30th 2012, issued by Hanoi authority for
Planning and Investment
Address: 458 Minh Khai, Vinh Tuy Ward, Hai Ba Trung District, Hanoi, Vietnam
Ask a question to doctor
Loading...
* Vinmec Times City 0243 9743 556
* Vinmec Central Park 0283 6221 166
* Vinmec Đà Nẵng 0236 3711 111
* Vinmec Nha Trang 0258 3900 168
* Vinmec Hải Phòng 0225 7309 888
* Vinmec Hạ Long 0203 3828 188
* Vinmec Phú Quốc 0297 3985 588
* Book visit via MyVinmec Choose the doctor and the appointment date at home
Install
* Customer Service Portal 24/07
Send
Subscribed
Thank you for subscribing. Vinmec is happy to send you the latest news by
email.
OK
### Subscribe for more healthcare information
#### Topic
* General information
* Pediatrics
* Obstetrics
* Cardiology
* Vaccine
* Nutrition
Register for special offer at Vinmec
All
* All
* Vinmec Times City
* Vinmec Central Park
* Vinmec Đà Nẵng
* Vinmec Nha Trang
* Vinmec Hải Phòng
* Vinmec Hạ Long
* Vinmec Phú Quốc
submit
By clicking the Sign Up button or the submit button, I confirm that I have
read and agree to the Privacy Policy Rules
1
| biology | 13707 | https://da.wikipedia.org/wiki/Vitamin | Vitamin | Et vitamin er en organisk forbindelse og et livsnødvendigt næringsstof for en organisme. En organisk kemisk forbindelse (eller et relateret sæt af forbindelser) kaldes et vitamin, når en organisme ikke på egen hånd kan syntetisere forbindelsen i de nødvendige mængder, og det derfor bliver nødt til at få gennem kosten; derfor afhænger definitionen af et "vitamin" af omstændighederne og den pågældende organisme. Fx er ascorbinsyre (en variant af C-vitamin) et vitamin for mennesker, men ikke for mange andre. Mange vitaminer findes som kosttilskud, og mens kosttilskud er vigtige for behandlingen af bestemte helbredsproblemer, findes der ikke beviser på næringsmæssige fordele, når de bruges af sunde og raske mennesker.
Opfattelsen er, at begrebet vitamin hverken inkluderer andre essentielle næringsstoffer som mineraler, fedtsyrer eller aminosyrer, som behøves i større mængder end vitaminer, eller det store antal andre næringsstoffer, der er sunde for kroppen, men som kun sjældnere kræves for at vedligeholde helbredet. I øjeblikket anerkendes 13 vitaminer universelt. Vitaminer klassificeres ved deres biologiske og kemiske aktivitet, ikke deres struktur. Derfor henviser hvert "vitamin" til et antal vitamer-forbindelser, der alle viser den biologiske aktivitet associeret med et bestemt vitamin. Sådan et sæt kemiske stoffer er grupperet under en alfabetiseret "generisk deskriptor"-titel som "A-vitamin" med forbindelserne retinal, retinol og fire kendte karotenoider. Vitamere kan pr. definition omdannes til den aktive form af vitaminet i kroppen og kan somme tider også omdannes til hinanden.
Vitaminer har forskellige biokemiske funktioner: Nogle som D-vitamin har hormonlignende funktioner som regulatorer af mineralmetabolisme eller regulatorer af celle- og vævsvækst og -differentiering som nogle former for A-vitamin. Andre fungerer som antioxidanter som E-vitamin og visse C-vitaminer. Det største antal vitaminer, de komplekse B-vitaminer, fungerer som enzym-cofaktorer (coenzymer) eller udgangsstoffer for dem; coenzymer hjælper enzymer som katalysatorer i metabolisme. I denne rolle kan vitaminer bindes stramt til enzymer som en del af prostetiske grupper: Fx er biotin en del af enzymer involveret i dannelsen af fedtsyrer. De kan også være mindre stramt bundet til enzymkatalysatorer som coenzymer, adskillelige molekyler, hvis funktion er at transportere kemiske grupper eller elektroner mellem molekyler. For eksempel kan folsyre transportere methyl, aldehyd og methylen-grupper i cellen. Selvom disse roller i at assistere enzymsubstratreaktioner er vitaminernes bedst-kendte funktion, er de andre vitaminfunktioner lige så vigtige.
Frem til midten af 1930'erne, da de første kommercielle B-vitaminer af gærekstrakt og semi-syntetiserede C-vitaminkosttilskudspiller blev solgt, blev vitaminer udelukkende indtaget gennem kosten, og forandringer i kosten (som kunne ske på en bestemt årstid) ændrede ofte voldsomt hvilke typer og mængder af vitaminer man indtog. Vitaminer er dog produceret som råt kemikalie og gjort meget tilgængeligt som billige semisyntetiserede og syntetiserede multivitamin-tilskud siden midten af det 20. århundrede. Studier i strukturel aktivitet, funktion og deres rolle i at vedligeholde helbredet kaldes vitaminologi.
Ordet vitamin er en sammensætning og sammentrækning af vita (liv) og amin. Det blev første gang formuleret af Casimir Funk i 1912.
Tilstedeværelsen af vitaminer kan være påkrævet for at optage andre stoffer fra maden. Fx optages mineralet kalk bedst ved tilstedeværelsen af D-vitamin.
Liste over vitaminer
Hvert vitamin bruges typisk i flere reaktioner, og derfor har de fleste flere funktioner.
Helbredsmæssige effekter
Vitaminer er essentielle for en multicellet organismes normale vækst og udvikling. Et foster begynder, fra det øjeblik, det undfanges, at udvikles ud fra sine forældres genetiske model ved hjælp af de næringsstoffer, det absorberer. Det kræver, at bestemte vitaminer og mineraler er til stede på bestemte tidspunkter. Disse næringsstoffer faciliterer de kemiske reaktioner, som blandt andet producerer hud, knogler og muskler. Hvis der er en alvorlig mangel på et eller flere af disse næringsstoffer, kan et barn udvikle en mangelsygdom. Selv mindre mangler kan forårsage permanent skade.
For størstedelens vedkommende fås vitaminer gennem føden, men nogle få opnås på andre måder. Fx producerer mikroorganismer i tarmene — normalt kaldet "tarmflora" — K-vitamin og biotin, mens en type D-vitamin syntetiseres i huden ved hjælp af den naturlige ultraviolette bølgelængde fra sollys. Mennesker kan producere nogle vitaminer fra udgangsstoffer, som de indtager som A-vitamin produceret fra betakaroten, og niacin fra aminosyren tryptophan.
Når vækst og udvikling er fuldendt, forbliver vitaminer essentielle næringsstoffer for en sund vedligeholdelse af cellerne, vævet og organismerne i en multicellet organisme; de lader også en multicellet livsform benytte den kemiske energi fra føden, den indtager, mere effektivt og hjælper med at bearbejde proteiner, kulhydrater og fedt, der kræves for respiration.
Tilskud
Hos individer, der ellers lever sundt, er der ikke meget bevis for, at tilskud har nogen gavnlig effekt, hvad angår kræft eller hjertekarsygdomme. Tilskud af A- og E-vitamin kan sågar øge dødeligheden hos sunde individer, omend de to store studier, som understøttede denne konklusion, medtog rygere, for hvem det allerede var kendt at betakaroten-tilskud kan være skadelige. Andre studier peger hen mod, at E-vitaminforgiftning er begrænset til overdosis af en bestemt type.
Den Europæiske Union og andre europæiske lande har regelsæt, som definerer begrænsningerne af vitamin- (og mineral-) indtag, der kan bruges sikkert som kosttilskud. De fleste vitaminer, der sælges som kosttilskud, må ikke overstige en maksimal daglig dosis. Vitaminprodukter over disse foreskrevne grænser betragtes ikke som kosttilskud og skal registreres som enten receptpligtig medicin eller håndkøbslægemiddel pga. deres potentielle bivirkninger. Som følge heraf betragtes de fleste tilskud af de fedtopløselige vitaminer (som A-, D-, E- og K-vitamin) med mængder over det daglige indtag som medicin. Den daglige dosis for et vitamintilskud må for eksempel ikke overstige 300 % af det anbefalede daglige indtag, og for A-vitamin er denne begrænsning endnu lavere (200 %). Denne type reguleringerfinder sted i de fleste europæiske lande.
Kosttilskud indeholder ofte vitaminer men kan også indeholde mineraler og urter. Videnskabelige undersøgelser bakker op om, at der er helbredsmæssige fordele ved kosttilskud til personer med bestemte helbredsmæssige problemer. I nogle tilfælde kan vitamintilskud have uønskede effekter, særligt hvis de tages før kirurgi sammen med andre kosttilskud eller medicin, eller hvis personen, der tager dem, har bestemte sundhedsmæssige forhold. De kan også indeholde vitaminniveauer, der er mange gange større, og i andre former, end man kan indtage gennem kost.
Mangel
Mennesker skal indtage vitaminer periodisk, men efter forskellige tidsplaner for at undgå vitaminmangel. Menneskekroppens depoter for de forskellige vitaminer varierer bredt; A- og B-vitamin og B12 opbevares i betragtelige mængder i menneskes legeme, hovedsageligt i leveren, og et voksent menneske kan undvære A- og D-vitamin i mange måneder, og i B12's tilfælde i flere år, før de udvikler mangeltilstand. Herimod opbevares vitamin B3 (niacin og niacinamid) ikke i menneskekroppen i særligt store mængder, så depoter kan ofte kun vare nogle uger. For C-vitamin er de første symptomer på skørbug i eksperimentelle studier med total C-vitamindeprivation varieret voldsomt, fra en måned til mere end seks måneder afhængig af den tidligere kostmæssige historie, som afgjorde kroppens depoter.
Vitaminmangel klassificeres som enten primær eller sekundær.
En primær mangel sker, når en organisme ikke får nok af vitaminet i sin kost.
En sekundær mangel kan skyldes en dybereliggende tilstand, som forhindrer eller begrænser optagelsen eller brugen af vitaminet pga. en "livsstilsfaktor" som rygning, alkoholmisbrug eller brug af medicin, som forstyrrer optagelse eller brug af vitaminet. Folk, som indtager en varieret kost, udvikler kun meget sjældent en alvorlig primær vitaminmangel. I modsætning hertil kan restriktive kostplaner have potentiale til at skabe forlænget vitaminmangel, der kan føre til alvorlige sygdomme.
Velkendte menneskelige vitaminmangler er tiamin (beriberi), niacin (pellagra), C-vitamin (skørbug) og D-vitamin (engelsk syge). I meget af den udviklede verden er sådanne mangelsygdomme sjældne; dette skyldes (1) et tilstrækkeligt forråd af mad og (2) tilsættelsen af vitaminer og mineraler til udbredte fødevarer, hvilket ofte kaldes berigelse. Udover disse klassiske mangelsygdomme har der også været tegn på forbindelse mellem vitaminmangel og en række andre lidelser.
Bivirkninger
I store doser har nogle vitaminer bivirkninger, som normalt bliver alvorligere, jo højere dosen er. Risikoen for at indtage for meget af et vitamin fra kosten er meget lille, men overdosis (vitaminforgiftning) fra vitamintilskud er set. Ved høje nok doser giver nogle vitaminer bivirkninger som kvalme, diarré og opkast. Når der fremkommer bivirkninger, kan man sædvanligvis komme problemet til livs ved at reducere dosis. Vitamindoserne er forskellige, da individuelle toleranceniveauer kan variere meget og lader til at være relaterede til alder og almen helbredstilstand.
Farmakologi
Vitaminer klassificeres enten som vandopløselige eller fedtopløselige. Der findes 13 vitaminer i mennesker: fire fedtopløselige (A, D, E og K) og ni vandopløselige (8 B-vitaminer og C-vitamin). Vandopløselige vitaminer opløses let i vand og udskilles generelt let fra kroppen, i en grad så niveau i urinen er en stærk indikator af vitaminindtagelse. Da de ikke lagres særlig nemt i kroppen, er det vigtigt at have et konstant indtag af dem. Mange typer af vandopløselige vitaminer syntetiseres af bakterier. Fedtopløselige vitaminer absorberes gennem mavetarmkanalen ved hjælp af lipider (fedt). De har en større risiko for at føre til hypervitaminose end de vandopløselige, da de har større tendens til at blive lagret i kroppen. Fedtopløselig vitaminregulering af særlig betydning i cystisk fibrose.
Historie
Værdien af at spise bestemte fødevarer for at vedligeholde et godt helbred blev anerkendt længe før vitaminerne blev identificeret. Man vidste allerede i det gamle Egypten, at ved at spise lever kunne man hjælpe til at kurere natteblindhed, en sygdom som det nu vides skyldes mangel på vitamin A. Havsejlads under Renæssancen resulterede i lange perioder uden adgang til frugter og grøntsager og gjorde vitaminmangelsygdomme udbredte i skibsbesætningerne.
I 1747 opdagede den skotske kirurg James Lind, at citrusfrugter hjalp til at forhindre skørbug, en dødelig sygdom, hvor kollagen ikke dannes ordentligt, hvilket fører til dårlig sårheling, blødende tandkød, stærk smerte og død. I 1753 udgav Lind sin Treatise on the Scurvy, som anbefalede at bruge citroner og limefrugter for at undgå skørbug. Anbefalingen blev fulgt af den britiske Royal Navy, hvilket førte til udtrykket "limey" om sømænd. Linds opdagelse blev ikke bredt accepteret i Royal Navy's Arktis-ekspeditioner i det 19. århundrede, hvor mange mente, at skørbug kunne undgås ved god hygiejne, regelmæssig motion og ved at holde mandskabets gejst høj, snarere end ved frisk frugt. Som følge heraf fortsatte de arktiske ekspeditioner med at være plaget af skørbug og andre mangelsygdomme. I begyndelsen af det 20. århundrede, da Robert Falcon Scott foretog sine to ekspeditioner til Antarktis, var den fremherskende medicinske teori, at skørbug skyldtes "fordærvet" dåsemad.
I slutningen af det 18. og begyndelsen af det 19. århundrede lod brugen af deprivationsstudier videnskabsfolk isolere og identificere en række vitaminer. Lipider fra fiskeolie blev brugt til at kurere engelsk syge (rakitis) hos rotter, og det fedtopløselige næringsstof blev oprindeligt kaldt "antirakitis A". På den måde blev den første "vitamin"-bioaktivitet, der nogensinde blev isoleret og kurerede rakitis, oprindeligt kaldt "A-vitamin"; i dag kaldes denne forbindelses bioaktivitet dog D-vitamin.
I 1881 studerede den russiske kirurg Nikolai Lunin skørbugens effekter, mens han var ved Tartu Universitet (nu i Estland. Han fodrede mus med en kunstig mikstur af alle de separate bestanddele, der på daværende tidspunkt var identificeret i mælk, navnlig proteinerne, fedttyperne, kulhydraterne og saltene. De mus, der kun fik de individuelle bestanddele døde, mens de mus der fik mælk udviklede sig normalt. Han konkluderede, at "en naturlig føde som mælk derfor skal, udover disse kendte centrale bestanddele, indeholde små mængder ukendte substanser, som er essentielle for liv." Hans konklusioner blev dog afvist af hans rådgiver, Gustav von Bunge, selv efter andre studenter reproducerede hans resultater. Et lignende resultat af Cornelius Pekelharing blev beskrevet i en hollandsk medicinsk journal i 1905, men blev ikke publiceret særlig vidt.
I Østasien, hvor polerede hvide ris var normal kost for middelklassen, var beriberi fra mangel på vitamin B1 endemisk. I 1884 observerede Takaki Kanehiro, en britisk-trænet læge fra den kejserlige japanske flåde, at beriberi var endemisk blandt lavtrangerende mandskab, som ofte ikke spiste andet end ris, men ikke blandt officerer som indtog en vestligt-inspireret kost. Med støtte fra den japanske flåde eksperimenterede han med besætningen på to krigsskibe: en besætning fik udelukkende hvide ris at spise, mens den anden fik kød, fisk, byg, ris og bønner. Den gruppe, som kun spiste hvide ris havde 161 besætningsmedlemmer med beriberi og 25 dødsfald, mens den anden gruppe kun havde 14 tilfælde af beriberi og ingen dødsfald. Det overbeviste Takaki og den japanske flåde om, at årsagen til beriberi skulle findes i kosten, men konkluderede fejlagtigt, at passende mængder protein ville forhindre det. At sygdomme kunne opstå fra kostmæssige mangler blev yderligere undersøgt af Christiaan Eijkman, som i 1897 opdagede, at man kunne forhindre beriberi i kyllinger ved at fodre dem med upolerede ris i stedet for polerede ris. Det følgende år postulerede Frederick Hopkins, at nogle madvarer indeholdt ekstra faktorer — udover proteiner, kulhydrater, fedt, etc. — som er nødvendige for menneskekroppens funktion. Hopkins og Eijkman blev tildelt Nobelprisen i fysiologi eller medicin i 1929 for deres opdagelser.
I 1910 blev det første vitaminkompleks isoleret af den japanske videnskabsmand Umetaro Suzuki, som havde held med at udtrække et vandopløseligt kompleks af mikronæringsstoffer fra risklid og navngav det aberisk syre (senere Orizanin). Han offentliggjorde denne opdagelse i en japansk videnskabelig journal. Da artiklen blev oversat til tysk glemte man i oversættelsen at gøre opmærksom på, at det var et nyligt opdaget næringsstof, hvilket blev påstået i den oprindelige japanske artikel, og opdagelsen gik derfor relativt ubemærket hen. I 1912 isolerede den polskfødte biokemiker Casimir Funk i London det samme kompleks af mikronæringsstoffer og fremsatte forslag om, at man navngav komplekset "vitamin". Det blev senere kendt som B-vitamin3 (niacin), selv om han selv beskrev det som "anti-beri-beri-faktor" (som i dag ville blive kaldt tiamin eller vitamin B1). Funk fremsatte en hypotese om, at andre sygdomme, såsom engelsk syge, pellagra, cøliaki og skørbug også ville kunne kureres med vitaminer. Max Nierenstein foreslog angiveligt navnet "vitamin" (fra "vital amin").). Navnet blev snart efter synonymt med Hopkins' ekstra faktorer, og da det endelig blev påvist, at ikke alle vitaminer er aminer, sad navnet allerede fast.
I 1930 afdækkede Paul Karrer betakarotens korrekte struktur, og identificerede andre karotenoider. Karrer og Norman Haworth bekræftede Albert Szent-Györgyis opdagelse af askorbinsyre og kom med anseelige bidrag til flaviners kemi, hvilket førte til identificeringen af laktoflavin. De modtog begge Nobelprisen i kemi i 1937 for deres undersøgelser af karotenoider, flaviner og vitamin A og B2.
I 1931 havde Albert Szent-Györgyi og hans kollega Joseph Svirbely mistanke om, at "hexuronisk syre" faktisk var vitamin C, og gav en prøve til Charles Glen King, som beviste dets anti-skørbug-effekt. I 1937 blev Szent-Györgyi tildelt Nobelprisen i fysiologi eller medicin for sin opdagelse. I 1943 blev Edward Adelbert Doisy og Henrik Dam tildelt Nobelprisen i fysiologi eller medicin for deres opdagelse af vitamin K og dets kemiske struktur. I 1967 blev George Wald tildelt nobelprisen (sammen med Ragnar Granit og Haldan Keffer Hartline) for opdagelsen af at vitamin A kan deltage direkte i en fysiologisk proces.
Navngivning
Grunden til, at vitaminsættet springer direkte fra E til K, er, at vitaminerne der svarede til bogstaverne F–J enten blev omklassificeret, droppet som falske eller omdøbt på grund af deres forhold til vitamin B, som blev et kompleks af vitaminer.
De tysktalende forskere, som isolerede og beskrev (og navngav) K-vitamin, valgte navnet fordi vitaminet er tæt involveret i koagulationen af blod efter at man pådrager sig sår (det tyske ord er, som det danske, Koagulation). På det tidspunkt var de fleste bogstaver fra F til J allerede brugt, så brugen af bogstavet K blev betragtet som fornuftig.
Der findes andre manglende B-vitaminer som blev omklassificeret eller vurderet til ikke at være vitaminer. For eksempel er B9 folsyre og fem af folaterne er i rækken B11 til B16, former af vitaminer der allerede er opdaget, ikke krævet som et næringsstof af hele befolkningen (som B10, PABA til internt brug), biologisk inaktive, giftige, eller med uklassificerbare effekter i mennesker, eller ikke generelt anset som vitaminer af videnskaben, såsom det højest-nummererede, som nogle naturopater kalder B21 og B22. Der findes også ni B-kompleksvitaminer med bogstaver (såsom Bm). Der er andre D-vitaminer, som nu anerkendes som andre substanser, hvilket nogle kilder af samme type nummererer op til D7. Den kontroversielle kræftbehandling laetril var på et tidspunkt navngivet som vitamin B17. Der lader ikke til at være nogen konsensus om vitamin Q, R, T, V, W, X, Y eller Z, ligesom der heller ikke er substanser, der officielt er designeret vitamin N eller I.
Noter
Litteratur
Henrik Dilling, Klar besked om vitaminer og mineraler, Aschehoug, .
Eksterne henvisninger
DTU Fødevareinstituttet: Fødevaredata
Netdoktor: oversigt vitaminer
Se også
Bioforstærkning
Mikrobiom
Ernæring | danish | 0.384841 |
sex_duration/i-climax-after-30-seconds-sex-what-should-i-do.txt | Skip to main content Skip to navigation
Close dialogue 1 / 1 Next image Previous image Toggle caption
Skip to navigation
Print subscriptions
Sign in
Search jobs
Search
International edition
* International edition
* UK edition
* US edition
* Australia edition
* Europe edition
The Guardian - Back to home The Guardian
* News
* Opinion
* Sport
* Culture
* Lifestyle
Show More Show More
* News
* View all News
* World news
* UK news
* Climate crisis
* Ukraine
* Environment
* Science
* Global development
* Football
* Tech
* Business
* Obituaries
* Opinion
* View all Opinion
* The Guardian view
* Columnists
* Cartoons
* Opinion videos
* Letters
* Sport
* View all Sport
* Football
* Cricket
* Rugby union
* Tennis
* Cycling
* F1
* Golf
* US sports
* Culture
* View all Culture
* Books
* Music
* TV & radio
* Art & design
* Film
* Games
* Classical
* Stage
* Lifestyle
* View all Lifestyle
* Fashion
* Food
* Recipes
* Love & sex
* Health & fitness
* Home & garden
* Women
* Men
* Family
* Travel
* Money
* Search input
google-search
Search
* Support us
* Print subscriptions
* International edition
* UK edition
* US edition
* Australia edition
* Europe edition
* * Search jobs
* Holidays
* Digital Archive
* Guardian Licensing
* About Us
* The Guardian app
* Video
* Podcasts
* Pictures
* Newsletters
* Today's paper
* Inside the Guardian
* The Observer
* Guardian Weekly
* Crosswords
* Wordiply
* Corrections
* * Search jobs
* Holidays
* Digital Archive
* Guardian Licensing
* About Us
* Fashion
* Food
* Recipes
* Love & sex
* Health & fitness
* Home & garden
* Women
* Men
* Family
* Travel
* Money
‘I love sex but I find myself stressing about it.’ Composite: Getty
Images/Guardian Design Team
View image in fullscreen
‘I love sex but I find myself stressing about it.’ Composite: Getty
Images/Guardian Design Team
Sexual healing Life and style
This article is more than 1 year old
# I climax after 30 seconds of sex. What should I do?
This article is more than 1 year old
I have a problem with premature ejaculation, and numbing spray only spoils the
mood
Pamela Stephenson Connolly
Tue 3 Jan 2023 08.00 GMT Last modified on Thu 5 Jan 2023 06.39 GMT
Share
I am a single male in my 20s who often goes on dates, so usually when I have
sex it is with somebody new. I climax way too early and often ejaculate
prematurely. Is there anything I can do about it? I love sex but I find
myself stressing about it because of performance anxiety. I sometimes
pretend during intercourse that I haven’t ejaculated yet just to avoid the
shame . At times, I only last for 30 seconds. I have a numbing spray, but
it is very awkward to use and it can spoil the mood when I stop to apply it.
How do I approach this problem, especially when getting with new people?
While many women and men say they want athletic lovemaking with prolonged
intercourse or penetration, many others say the opposite. The latter group
either prefer a short amount of penetration with less chance of discomfort or
irritation – or no penetration at all. This is especially understandable for
women, since female pleasure and the means to orgasm are best achieved via
direct stimulation of the clitoris, which is located outside the vagina. Early
ejaculation is a treatable issue, but try not to stress about it. Being
anxious about your “performance” will only exacerbate the situation. Instead,
work on improving your ability to please your partner with skills that do not
involve your penis, such as oral and manual stimulation. There are many highly
erotic, non-penetrative techniques and it’s time you stopped worrying about
how your penis performs and think about what else would directly please your
partner. Combine good communication skills with your personal seductive style
to elicit information about his or her preferences, and practice sharing what
you need with them. Even people who do not worry about early ejaculation would
be wise to adopt this course of action.
* Pamela Stephenson Connolly is a US-based psychotherapist who specialises in treating sexual disorders.
* If you would like advice from Pamela on sexual matters, send us a brief description of your concerns to [email protected] (please don’t send attachments). Each week, Pamela chooses one problem to answer, which will be published online. She regrets that she cannot enter into personal correspondence. Submissions are subject to our terms and conditions .
* Comments on this piece are premoderated to ensure discussion remains on topics raised by the writer. Please be aware there may be a short delay in comments appearing on the site.
Explore more on these topics
* Life and style
* Sexual healing
* Sex
* Relationships
* features
Share
Reuse this content
## Comments (…)
Sign in or create your Guardian account to join the discussion
## Most viewed
## Most viewed
* Fashion
* Food
* Recipes
* Love & sex
* Health & fitness
* Home & garden
* Women
* Men
* Family
* Travel
* Money
* News
* Opinion
* Sport
* Culture
* Lifestyle
Original reporting and incisive analysis, direct from the Guardian every
morning
Sign up for our email
* Help
* Complaints & corrections
* SecureDrop
* Work for us
* * Privacy policy
* Cookie policy
* Terms & conditions
* Contact us
* All topics
* All writers
* Digital newspaper archive
* Facebook
* YouTube
* Instagram
* LinkedIn
* Twitter
* Newsletters
* Advertise with us
* Search UK jobs
Back to top
© 2024 Guardian News & Media Limited or its affiliated companies. All rights
reserved. (dcr)
| biology | 175578 | https://da.wikipedia.org/wiki/Stephen%20Glass | Stephen Glass | Stephen Glass, født 15. september 1972 i Highland Park, Illinois, USA, er en amerikansk advokat, forfatter og tidligere journalist ved magasinet The New Republic, hvorfra han blev fyret, da det blev afsløret, at han havde baseret flere af sine artikler på opdigtede citater, falske kilder og fiktive hændelser. Historien om Stephen Glass og hans personlige deroute dannede senere grundlag for filmen Shattered Glass fra 2003.
Historien
Stephen Glass havde studeret jura ved University of Pennsylvania, hvor han var redaktør og senere fra 1993 ansvarshavende redaktør for universitetets studenteravis The Daily Pennsylvanian. Han vakte opsigt med en række afslørende artikler, der handlede om United Way of America, der er en sammenslutning af velgørenhedsorganisationer i USA, samt en new journalism artikel han skrev om en nat, han havde tilbragt med en hjemløs narkoman nær Penn's campus, området ved University of Pennsylvania.
Stephen Glass afbrød i 1994 studierne ved University of Pennsylvania for at skrive for The New Republic, hvor han allerede som 23 årig opnåede national anerkendelse i USA som politisk journalist. En anerkendelse, han dog skulle miste igen.
Stephen Glass blev fyret fra The New Republic i maj 1998, efter at det blev afsløret, at han havde begået flere tilfælde af journalistisk bedrageri. Senere færdiggjorde han sine jurastudier ved Georgetown University Law Center.
Bedraget
Historien, der udløste fyringen, var "Hack Heaven", der handlede om en 15 årig hacker, som angiveligt blev ansat som it-sikkerhedskonsulent i et større amerikansk firma, Jukt Micronics, efter at have hacket sig ind i deres computersystemer, afsløret deres svagheder, og afpresset ledelsen.
Artiklen havde følgende åbning:
Ian Restil, a 15-year-old computer hacker who looks like an even more adolescent version of Bill Gates, is throwing a tantrum. "I want more money. I want a Miata. I want a trip to Disney World. I want X-Men comic [book] number one. I want a lifetime subscription to Playboy, and throw in Penthouse. Show me the money! Show me the money!"...
Across the table, executives from a California software firm called Jukt Micronics are listening – and trying ever so delicately to oblige. "Excuse me, sir," one of the suits says, tentatively, to the pimply teenager. "Excuse me. Pardon me for interrupting you, sir. We can arrange more money for you ..."
Kort efter offentliggørelsen af "Hack Heaven", kunne journalisten Adam Penenberg ved Forbes Magazines online udgave, Forbes.com dokumentere, at historien var fri fantasi, og at firmaet Jukt Micronics slet ikke eksisterede. En intern undersøgelse hos The New Republic afslørede, at Stephen Glass havde fabrikeret en falsk hjemmeside og telefonsvarer, der skulle snyde The New Republics fact checkers, altså de medarbejdere, hvis opgave det var at kontrollere dokumentationen og kilderne i alle magasinets artikler inden offentliggørelsen af dem.
The New Republic kunne efter den interne undersøgelse konstatere, at 27 ud af de 41 artikler som Stephen Glass havde skrevet for magasinet, var opdigtede. Tre andre magasiner som Stephen Glass på freelancebasis havde leveret artikler til, Rolling Stone, George og Harper's Magazine, iværksatte også interne undersøgelser. Rolling Stone og Harper's fandt ikke noget, der kunne tyde på opdigtede historier, men de kunne ikke få efterprøvet anonyme kildeangivelser. Magasinet George opdagede at Stephen Glass havde fabrikeret citater i forbindelse med en artikel om Vernon Jordan, en af Bill Clintons rådgivere.
Forbes.com høstede stor anerkendelse for afsløringen af Stephen Glass, og var medvirkende til, at internetjournalistikken i højere grad blev taget alvorligt som et seriøst alternativ til den trykte presse.
Efterspillet
Efter at være blevet fyret fra The New Republic, færdiggjorde Stephen Glass sine jurastudier i 2000 ved Georgetown University Law Center. I 2003 var han gæst hos det amerikanske nyhedsmagasin hos CBS, 60 Minutes, hvor han promoverede sin nyskrevne selvbiografiske roman "The Fabulist".
Stephen Glass løftede lidt af sløret for motivet til sine bedragerier med udtalelsen: "I wanted them to think I was a good journalist, a good person. I wanted them to love the story so they would love me"
I 2004 trak han sin ansøgning om om tilladelse til at praktisere som advokat i New York efter han fik nys om den ikke ville bilve tildelt. I 2009 blev han nægtet at praktisere som advokat i Californien. Glass ankede og sagen blev endelig afgjort med stadfæstelse i Californiens Højesteret i 2014.
I 2003 udkom filmen Shattered Glass, der handler om Stephen Glass' storhed og fald. Filmen beskriver samtidigt den hektiske verden inden for den politiske journalistik og det daglige arbejde hos et politisk magasin. I filmen portrætteres Stephen Glass af skuespilleren Hayden Christensen.
Baggrundsviden
The New Republic er et politisk moderat liberalistisk magasin, der ukommer to gange om måneden i USA og har mellem 40.000 og 65.000 læsere. Ansvarshavende redaktør er Martin Peretz. Redaktør er Franklin Foer.
The Daily Pennsylvanian er en uafhængig studenteravis ved University of Pennsylvania. Det udkommer fem gange om ugen, og har en medarbejderstab der udgøres af mere end 250 studerende. Man udgiver også et ugentligt underholdningsmagsin ved navn 34th Street Magazine.
New Journalism er et begreb skabt i 1973 af forfatteren Tom Wolfe i en serie artikler, der foruden ham selv blev skrevet af bl.a. Truman Capote, Hunter Stockton Thompson, Norman Mailer og Robert Christgau. New Journalism bygger på fire grundpiller:
historien skal bygges op omkring scenarier frem for resuméer
dialogen skal være fuldstændig frem for bygget op om citater
fortællingen skal komme fra personernes synsvinkel
der være beskrivelser af detaljer, der fortæller om personernes liv og miljø.
Forbes magazine er flagskibet hos det amerikanske forlags- og medieselskab Forbes, og henvender sig hovedsageligt til forretningsverdenen, med artikler om den merkantile og finansielle sektor.
Rolling Stone udkommer to gange om måneden, og koncentrerer sig om musik, politik og popkulturen. Det blev grundlagt i 1966 af Jann Wenner, der stadig er redaktør og udgiver, og musikkritikeren Ralph J. Gleason.
George var et lige dele politisk magasin som et livsstilsmagsin der udkom mellem 1995 og 2001. Det blev grundlagt af John F. Kennedy Jr. og Michael J. Berman. Magasinets debut vakte en del opsigt, idet det havde modellen Cindy Crawford på forsiden – klædt på, eller klædt af om man vil, som George Washington.
Harper's Magazine er et amerikansk månedsmagasin, der dækker litteratur, politik, kultur, økonomi og kunst fra et mere progressivt og lettere venstreorienteret synspunkt. Det er efter Scientific American det ældste magasin i USA, og har godt 220.000 læsere.
Stephen Glass' artikler online
Her følger et udvalg af de artikler, Stephen Glass menes at have forfalsket til The New Republic:
"Mrs. Colehill Thanks God For Private Social Security", publiceret juni 1997 i Policy Review Magazine
"Probable Claus" , publiceret d. 6. og d. 13. januar 1997
"Don't You D.A.R.E.", publiceret d. 3. marts 1997
"Writing on the Wall", publiceret d. 24. marts 1997
"The Young and the Feckless", publiceret d. 15. september 1997
"Washington Scene: Hack Heaven", publiceret d. 18. maj 1998
Kildehenvisning
Forbes smokes out fake New Republic story on hackers, 11. maj 1998
Forbes.com: The New York Times Scandal Recalls Glass Episode
CBS NEWS: Stephen Glass: I Lied For Esteem
NFIA – National Families in Action: Letter to D.A.R.E. from Stephen Glass
Journalist who made up “Hack Heaven” story barred from legal profession
Eksterne henvisninger
Romanforfattere fra USA
Journalister fra USA | danish | 1.116054 |
sex_duration/Handicap_principle.txt |
The handicap principle is a disputed hypothesis proposed by the Israeli biologist Amotz Zahavi in 1975. It is meant to explain how sexual selection may lead to "honest" or reliable signalling between male and female animals which have an obvious motivation to bluff or deceive each other. The handicap principle suggests that secondary sexual characteristics are costly signals which must be reliable, as they cost the signaller resources that individuals with less of a particular trait could not afford. The handicap principle further proposes that animals of greater biological fitness signal this through handicapping behaviour, or morphology that effectively lowers overall fitness. The central idea is that sexually selected traits function like conspicuous consumption, signalling the ability to afford to squander a resource. Receivers then know that the signal indicates quality, because inferior-quality signallers are unable to produce such wastefully extravagant signals.
The handicap principle is supported by game theory modelling representing situations such as nestlings begging for food, predator-deterrent signalling, and threat displays. However, honest signals are not necessarily costly, undermining the theoretical basis for the handicap principle, which remains unconfirmed by empirical evidence.
History[edit]
Origins[edit]
The handicap principle was proposed in 1975 by the Israeli biologist Amotz Zahavi. He argued that sexual selection may lead to "honest" or reliable signalling between male and female animals, even though they have an interest in bluffing or deceiving each other. The handicap principle asserts that secondary sexual characteristics are costly signals, which are reliable indicators of the signaller's quality, since they cost the signaller resources that lower-quality individuals could not afford. The generality of the phenomenon is a matter of some debate and disagreement, and Zahavi's views on the scope and importance of handicaps in biology have not been accepted by the mainstream. Nevertheless, the idea has been very influential, with most researchers in the field believing that the theory explains some aspects of animal communication.
Grafen's signaling game model[edit]
Further information: Signaling game
Graph based on Johnstone's 1997 graphical representation of a Zahavian handicap. Where
C
L
{\displaystyle C_{L}}
is cost to a low-quality signaller and
C
H
{\displaystyle C_{H}}
is cost to a high-quality signaller. Optimal signalling levels are
S
L
∗
{\displaystyle S_{L}^{*}}
for a low-quality signaller, and
S
H
∗
{\displaystyle S_{H}^{*}}
for a high-quality signaller.
The handicap principle was initially controversial; The British biologist John Maynard Smith was a notable early critic of Zahavi's ideas.
However, the handicap principle gained wider acceptance because it is supported by game theory models, most notably the Scottish biologist Alan Grafen's signalling game model. This was essentially a rediscovery of the Canadian-American economist Michael Spence's job market signalling model, where the job applicant signals their quality by declaring a costly education. In Grafen's model, the courting male's quality is signalled by investment in an extravagant trait—similar to the peacock's tail. The signal is reliable if the cost to the signaller of producing it is proportionately lower for higher-quality signallers than for lower-quality ones.
A series of papers by the American biologist Thomas Getty showed that Grafen's proof of the handicap principle depends on the critical, simplifying assumption that signallers trade off costs for benefits in an additive fashion, analogous to the way humans invest money to increase income in the same currency. This is illustrated in the figures from Johnstone 1997, which show that the optimum signalling levels are different for low- and high-quality signallers. The validity of the assumption that costs and benefits are additive has been contested, in its application to the evolution of sexually selected signals. It can be reasoned that since fitness depends on the production of offspring, this is a multiplicative rather than additive function of reproductive success.
Further game theoretical models demonstrated the evolutionary stability of handicapped signals in nestlings' begging calls, in predator-deterrent signals and in threat-displays. In the classic handicap models of begging in game theory, all players are assumed to pay the same amount to produce a signal of a given level of intensity, but differ in the relative value of eliciting the desired response (donation) from the receiver. The hungrier the baby bird, the more food is of value to it, and the higher the optimal signalling level (the louder its chirping).
Cheap talk models[edit]
Further information: Cheap talk
Counter-examples to handicap models predate handicap models themselves. Models of signals (such as threat displays) without any handicapping costs show that what biologists call cheap talk may be an evolutionarily stable form of communication. Analysis of some begging models shows that non-communication strategies are not only evolutionarily stable, but lead to higher payoffs for both players. In human mate choice, mathematical analyses including Monte Carlo simulations suggest that costly traits ought to be more attractive to the other sex and much rarer than non-costly traits.
It was soon discovered that honest signals need not be costly at the honest equilibrium, even under conflict of interest. This conclusion was first shown in discrete models and then in continuous models. Similar results were obtained in conflict models: threat displays need not be handicaps to be honest and evolutionarily stable.
Dustin J. Penn and Szabolcs Számadó stated in 2019 that there was still no empirical evidence for evolutionary pressure for wasteful biology or acts, and proposed that the handicap principle should be abandoned.
Predictions and interpretations[edit]
Luxury cars and other "Veblen goods" may be an example of the handicap principle in humans
The handicap principle predicts that a sexual ornament, or any other signal such as visibly risky behavior, must be costly if it is to accurately advertise a trait of relevance to an individual with conflicting interests. Typical examples of handicapped signals include bird songs, the peacock's tail, courtship dances, and bowerbird bowers. American scientist Jared Diamond has proposed that certain risky human behaviours, such as bungee jumping, may be expressions of instincts that have evolved through the operation of the handicap principle. Zahavi has invoked the gift-giving potlatch ceremony as a human example of the handicap principle in action: the conspicuous generosity is costly. This interpretation of potlatch can be traced to Thorstein Veblen's use of the ceremony in his book Theory of the Leisure Class as an example of "conspicuous consumption".
The handicap principle gains further support by providing interpretations for behaviours that fit into a single unifying gene-centered view of evolution and making earlier explanations based on group selection obsolete. A classic example is that of stotting in gazelles. This behaviour consists in the gazelle initially running slowly and jumping high when threatened by a predator such as a lion or cheetah. The explanation based on group selection was that such behaviour might be adapted to alerting other gazelle to a cheetah's presence or might be part of a collective behaviour pattern of the group of gazelle to confuse the cheetah. Instead, Zahavi proposed that each gazelle was communicating that it was a fitter individual than its fellows.
Signals to members of the same species[edit]
Zahavi studied in particular the Arabian babbler, a highly social bird, with a life-length of 30 years, which appears to behave altruistically. Its helping-at-the-nest behavior, where non-parent birds assist in feeding, guarding, and caring for nestlings, often occurs among unrelated individuals. This, therefore, cannot be explained by kin selection, natural selection acting on genes that close relatives share with the altruistic individual. Zahavi reinterpreted these behaviors according to his signalling theory and its correlative, the handicap principle. The altruistic act is costly to the donor, but may improve its attractiveness to potential mates. The evolution of this condition may be explained by competitive altruism.
French biologist Patrice David showed that in the stalk-eyed fly species Cyrtodiopsis dalmanni, genetic variation underlies the response to environmental stress, such as variable food quality, of a male sexual ornament, eye span. He showed that some male genotypes always develop large eye spans, but others reduce eye span in proportion to environmental worsening. David inferred that female mate choice yields genetic benefits for offspring.
Signals to other species[edit]
Further information: Anti-predator adaptation
Impala stotting, a behavior that may serve as a pursuit deterrence signal to predators
Signals may be directed at predators, with the function of showing that pursuit will probably be unprofitable. Stotting, for instance, is a form of energetic jumping that certain gazelles do when they sight a predator. As this behavior gives no evident benefit and would seem to waste resources (diminishing the gazelle's head start if chased by the predator), it appeared likely to be selected against. However, it made sense when seen as a pursuit deterrence signal to predators. By investing a little energy to show a lion that it has the fitness necessary to avoid capture, a gazelle reduces the likelihood that it will have to evade the lion in an actual pursuit. The lion, faced with the demonstration of fitness, might decide that it would fail to catch this gazelle, and thus choose to avoid a probably wasted pursuit. The benefit to the gazelle is twofold. First, for the small amount of energy invested in the stotting, the gazelle might not have to expend the tremendous energy required to evade the lion. Second, if the lion is in fact capable of catching this gazelle, the gazelle's bluff may lead to its survival that day (in the event the bluff succeeds). However, the mathematical biologist John Maynard Smith commented that other explanations were possible, such as that it was an honest signal of fitness, or an honest signal that the predator had been detected, and it was hard to see how stotting could be a handicap.
Another example is provided by larks, some of which discourage merlins by sending a similar message: they sing while being chased, telling their predator that they will be difficult to capture.
Immunocompetence handicaps[edit]
The theory of immunocompetence handicaps suggests that androgen-mediated traits accurately signal condition due to the immunosuppressive effects of androgens. This immunosuppression may be either because testosterone alters the allocation of limited resources between the development of ornamental traits and other tissues, including the immune system, or because heightened immune system activity has a propensity to launch autoimmune attacks against gametes, such that suppression of the immune system enhances fertility. Healthy individuals can afford to suppress their immune system by raising their testosterone levels, at the same time augmenting secondary sexual traits and displays. A review of empirical studies into the various aspects of this theory found weak support.
See also[edit]
Aposematism
Multiple sexual ornaments
Parasite-stress theory
Sacrifice | biology | 998 | https://da.wikipedia.org/wiki/Art | Art | Arten (species, forkortet sp., flertal: spp.) er den grundlæggende systematiske enhed inden for biologien. Arten defineres ofte som en naturlig gruppe af populationer, hvor udveksling af gener finder sted (eller kan finde sted) og som i forhold til forplantning er isoleret fra andre grupper. Det vil sige at kun individer inden for samme art kan parre sig og få forplantningsdygtigt afkom. Dette kaldes det biologiske artsbegreb. For organismer, der formerer sig ukønnet eller ved selvbestøvning, må arter afgrænses ud fra ligheder og forskelle mellem forskellige individer. Nogle dyrearter kan i fangenskab hybridisere og få fertilt afkom, men da dette ikke vil ske i naturen, selv om de mødes her, betragtes de som forskellige arter.
Eksempel
To heste kan parre sig og få et føl, der igen kan få føl med andre heste – hestene tilhører derfor samme art. En hest og et æsel kan også parre sig og deres unger kaldes enten muldyr eller mulæsel, afhængig af hvem der er moren, men muldyret eller mulæselet kan (normalt) ikke få unger, da de oftest er sterile. Af den grund regnes hest og æsel som to forskellige arter. Det samme princip gælder også for planterne. Denne naturskabte afgrænsning mellem to arter kaldes en artsbarriere. Den kan af og til gennembrydes, når ellers sterile krydsninger spontant eller kunstigt får gennemført en kromosomfordobling. Se f.eks. Vadegræs (Spartina pectinata).
Arter over for hybrider
Man kan dog godt komme ud for, at arter kan krydses og får blandet afkom, men hybriden vil kun kunne bestå på steder, hvor ingen af forældrearterne kan klare sig. Dette er et særligt udpræget problem med Rododendron (Rhododendron) og Tjørn (Crataegus), fordi disse slægter breder sig voldsomt efter skovbrand eller stormfald. Da hybriderne bliver frugtbare i en yngre alder end arterne, kan de dominere i en periode, men når skoven lukker sig, så fortrænges hybriderne og kun de specialiserede arter kan overleve i skovens dybe skygge eller ude i lyset i sumpe, på ur og i kalksten, m.m.
Flere artsbegreber
Fordi det biologiske artsbegreb kan være besværligt at anvende i praksis, er der efterhånden skabt en række andre artsbegreber:
Morfologisk artsbegreb Arterne adskiller sig fra hinanden ved deres bygning. Dette begreb er blevet meget anvendt gennem tiden.
Økologisk artsbegreb Definerer en art som en gruppe af organismer, der udfylder samme niche. Krydsninger mellem to nærtstående arter vil ikke være optimalt tilpasset til forældrearternes nicher og vil ikke klare sig i konkurrencen.
Evolutionære artsbegreb Også kaldet det kladistiske eller fylogenetiske artsbegreb. Naturen er dynamisk, ikke statisk - alle arter ændrer sig med tiden og bliver, hvis de ikke uddør som følge af konkurrence, naturkatastrofer m.v., til én eller flere nye arter. Det evolutionære artsbegreb minder om det biologiske, men inddrager tidsdimensionen, det vil sige at en art udvikler sig over tid og at nye arter opstår ved artsdannelse. Individer der fylogenetisk har samme stamfader tilhører samme art.
Pluralistisk artsbegreb En art er et samfund af populationer, der formerer sig og lever inden for en bestemt niche i naturen.
Se også
Systematik
Evolutionsteori
Kilder
Lars Skipper: Hvad er en art? Citat: "...Arten er den eneste [klassifikations-kategori] der eksisterer i virkeligheden, alle andre (slægter, familier, ordener m.v.) er indført for overskuelighedens skyld..."
Eksterne henvisninger
2003-12-31, ScienceDaily: Working On The 'Porsche Of Its Time': New Model For Species Determination Offered Citat: "...two species of dinosaur that are members of the same genera varied from each other by just 2.2 percent. Translation of the percentage into an actual number results in an average of just three skeletal differences out of the total 338 bones in the body. Amazingly, 58 percent of these differences occurred in the skull alone. "This is a lot less variation than I'd expected," said Novak..."
2003-08-08, ScienceDaily: Cross-species Mating May Be Evolutionarily Important And Lead To Rapid Change, Say Indiana University Researchers Citat: "...the sudden mixing of closely related species may occasionally provide the energy to impel rapid evolutionary change..."
2004-01-09 ScienceDaily: Mayo Researchers Observe Genetic Fusion Of Human, Animal Cells; May Help Explain Origin Of AIDS Citat: "...The researchers have discovered conditions in which pig cells and human cells can fuse together in the body to yield hybrid cells that contain genetic material from both species..."What we found was completely unexpected," says Jeffrey Platt, M.D..."
2000-09-18, ScienceDaily: Scientists Unravel Ancient Evolutionary History Of Photosynthesis Citat: "...gene-swapping was common among ancient bacteria early in evolution..."
2004-06-07, Sciencedaily: Parting Genomes: University Of Arizona Biologists Discover Seeds Of Speciation Citat: "...There's a huge amount of biodiversity out there, and we don't know where it comes from. Evolutionary biologists are excited to figure out what causes what we see out there--the relative forces of selection and drift--whether things are adapting to their environment or variation is random..."
2005-07-05, Sciencedaily: Trees, Vines And Nets -- Microbial Evolution Changes Its Face Citat: "... EBI researchers have changed our view of 4 billion years of microbial evolution...In all, more than 600,000 vertical transfers are observed, coupled with 90,000 gene loss events and approximately 40,000 horizontal gene transfers...A few species, including beneficial nitrogen-fixing soil bacteria, appear to be 'champions'of horizontal gene transfer; "it's entirely possible that apparently harmless organisms are quietly spreading antibiotic resistance under our feet," concludes Christos Ouzounis..."
2005-11-11, Sciencedaily: Lateral Thinking Produces First Map Of Gene Transmission Citat: "...Their results clearly show genetic modification of organisms by lateral transfer is a widespread natural phenomenon, and it can occur even between distantly related organisms... it was assumed that transfer of genes could only be vertical, i.e. from parents to offspring..."
Økologi
Biologi | danish | 0.791178 |
sex_duration/showthread.php.txt | Ask a question Ask
Sign in Sign up
* Home
* Main topics
Study help discussion
* All study help
* GCSEs
* A-level
University discussion
* Uni applications
* University and HE colleges
* University help and courses
* University student life
* Postgraduate
* Medicine
Careers discussion
* Careers and jobs
* Teacher training
* Finance and accountancy
Lifestyle discussion
* Chat
* Relationships
* Fashion
* Fitness
* Sexual health
* GCSE and A-level
Study level discussion
* GCSE
* A-level
* Btec
* T-level
* Scottish qualifications
* IB
* EPQ
Study help discussion (by subject)
* Maths
* English
* Geography
* History
* Biology
* Chemistry
* Physics
* Foreign languages
* All study help
GCSE and A-level articles
* GCSE articles
* A-level articles
* Exam and revision articles
* What to do after GCSEs
* What to do after A-levels
* When is A-level results day 2024?
* When is GCSE results day 2024?
More study help
* Studying, revision and exam support
* Grow your Grades
* Revision and exams
Revision and exam discussion
* A-levels
* GCSEs
* Btecs
* All A-level exams
* All GCSE exams
Revision tips
* Top vloggers for revision help
* Best video resources for revising GCSE or A-level English
* Best places online for maths revision
* How to use past exam papers for revision
Preparing for an exam
* How to study effectively for your exams
* Handling revision and exam stress
* What to expect from an exam
* Seven things to do in the last week before an exam
* How to handle exam nightmares
Finding motivation
* Getting past procrastination
* It's not too late: 10 ways to kick-start your revision
* How to revise when you're feeling lazy
* University
University discussion
* All universities
* Applying through Ucas
* Student finance
* Personal statement
* Postgraduate study
* Uni accommodation
* University life
University course discussion
* All uni courses
* Apprenticeships
* Medicine
* Arts and humanities courses
* Stem courses
* Social science courses
Universities by region
* North of England
* Midlands
* South of England
* Greater London
* Scotland
* Wales
* Northern Ireland
* Distance learning
* International study
University guides and articles
* All university articles
* Applying to uni articles
* Personal statements
* Personal statement examples
* Freshers
* University open days
* Studying law at university
* Student life at university
* Careers and jobs
Careers discussion
* Careers and jobs discussion
* Apprenticeships discussion
* Part-time and temp jobs
Industry forums
* Career forums by sector
* Armed forces careers
* Consultancy careers
* Finance careers
* Legal careers
More industry forums
* Marketing careers
* Medicine and healthcare careers
* Public sector careers
* Stem careers
* Teaching careers
* Life and relationships
Life chat
* General chat
* Relationships
* Friends, family and colleagues
* Advice on everyday issues
* Fashion
Health chat
* Life and style
* General health
* Sexual health
* Mental health
* Fitness
News and entertainment chat
* Debate and current affairs
* Entertainment
* UK politics
* News
* Educational debate
* Football
* Sport
* Student finance
Undergraduate
* Full time
* Part time
Postgraduate
* Postgraduate Master's Loan
* Postgraduate Doctoral Loan
* Disabled Students' Allowances
* Taking a break or withdrawing from your course
Further information
* Parents and partners
* Repayment
* Advanced Learner Loan
What would you like to say?
Your discussion will live here...
(Start typing, we will pick a forum for you)
Post
1. Forums
2. 3. 4. 5. ###### My boyfriend lasts 20 seconds :(
This discussion is now closed.
Check out other Related discussions
* House viewing, Should I go alone?
* My boyfriend dumped me over these reasons
* issues (??) in my relationship
* Stuck on Past Paper
* Does body count matter ?
* have a fear and insecurity of oral sex
* If I got average mark in my second yr could I still get a first class
* Is my friend toxic?
* i should stop thinking about him
* URGENT !!
* struggling with dating
* Tired of never being anyone’s first choice
* Need help !
* Why do so many girls have a boyfriend? Am I really unlucky?
* How to cope with the cost of living as a student
* Normal body count
* veterinary medicine as a second degree
* I'm clueless
* Boyfriend going on a lads holiday!!
* Need some advice
Watch
11 years ago
# My boyfriend lasts 20 seconds :(
Anonymous #1
My boyfriend and I have been together for 3 years, and we
generally have a great relationship apart from how things are in
the bedroom. Ever since we first slept together, he only lasts in
between 20-40 seconds when having sex (if having sex for the first
time that day and at a normal pace, eg. not too fast but not
slow). If we've already had sex once that day and we do it a
second time, it might reach a maximum of 2 minutes.
He says that he's been like this his whole life and even with
girls before me. I know it's common for boys to ejaculate quickly
when they're young, but he's 22 and we've been together for 3
years so he's had plenty of practice, but the problem is not
improving in the slightest. He'll last longer if masturbating, but
again it is still quick, around 2 minutes. It has got to the
point where we'll have sex at a ridiculous slow pace, or he'll
just keep stopping during it to try and last longer; it makes
everything so awkward.
I know I probably sound insensitive and that there's more than sex
to a relationship, but this has been going on for so long and I
really feel it's beginning to take a toll on our relationship
because it frustrates me so, so much, and then that makes me feel
bad because it's not his fault. I told him the other day that it
was affecting me, and he looked so upset and said that I deserved
a better boyfriend, which made me feel awful for even saying
anything in the first place. I've told him to go to the doctors
and that I would accompany him, but he says he's too embarrassed.
I know that he can do 'stuff' to me, but I don't enjoy it all
that much and would prefer to just have sex. I now feel as though
I don't even want to have sex with him, because it just feels
like there's no point to it. Even though it's over quickly for
him, he's still getting some pleasure out of it whereas I get
absolutely nothing. I know that it must make him feel really
inadequate which is why I have tried not to make an issue out of
it for three years, but it's really getting me down now.
Does anyone have any advice on what we could do to improve the
problem, or any tablets we can buy? Anything, basically!
1 Report
Scroll to see replies
* 1
* 2
* Next
Reply 1
11 years ago
unprinted
19
He's not the only one and there are plenty of threads about this
here.
What he needs to do is become aware of when he's getting close to
coming and then do something to delay it.
0 Report
Reply 2
11 years ago
Idle
2
If you don't use condoms maybe try them, if you do then thicker
ones to make the feeling less for him. A lot of guys find if
they are going to cum to soon that thinking about something random
can take you away from the edge.
0 Report
Reply 3
11 years ago
username391622
11
It won't happen over night but there are methods and he can do
things to last longer. I'm not gonna go into detail just google
it.
0 Report
Reply 4
11 years ago
The_Procrastinator
4
http://www.durex.com/en-GB/Products/condoms/Pages/Durex-Performa.aspx
1 Report
Reply 5
11 years ago
Classical Liberal
19
Original post by Anonymous
x
Has he tried "reverse kegels"?
I am under the impression, I don't actually know and rarely talk
to guys about things like this (for all guy bravado with regards
to sex they will not talk about it), that most involuntary
ejactulation is caused by the "PC" muscle (the muscle that controls
peeing, pooing and ejactulation) is too tense. The muscle being
tensed up stimulates the male "g spot" (no idea where this actually
is but I can definitely feel the sensation myself) which causes a
guy to ejactulate.
You can tense up the PC muscle when peeing. Try and stop the flow
of pee. You can relax the PC muscle by doing the opposite. By
trying to force out your pee as hard as is possible. You can also
loosen up the PC muscle by simulating taking a poo.
I know it sounds wierd but get your boyfriend to try simulate
having a ****/farting during sex/masturbating. This should release the
tension on the PC muscle (stretch it out) and reduce the
stimulation that leads to ejactulation. Ideally he should try to
simulate trying to take a **** and then simulate trying to pee
hard, however the latter is quite tricky without any urine. He will
also probably find it hard to breathe doing this and should feel
his abs flexing. Ideally with practice he should be able to learn
how to breathe and not flex his abs.
I suspect that when guys start feeling that sensation of just being
about to ejactulate, they desperately tense up the PC muscle to try
and block the semen (just like trying to stop the flow of pee)
however this just makes everything worse because it just stimulates
you even more.
As an added bonus to being able to last longer by stretching out
(removing the tension) the PC muscle he will also be able to have
more forceful orgasms. Take your arm and imagine you do not have a
tricep (only a bicep). If you tense up your bicep your arm bends.
Without your tricep the only way to straighten out your arm would
be to remove the tension from your bicep. Your PC muscle works
like this (girls have them too). If you had your bicep without any
tension and then you suddenly tensed it as much as possible, your
arm would go through a large range of motion with a lot of force.
Compare this to if you had your bicep already a bit tense and
then tensed it as much as possible. There would be less force and
a smaller range of motion compared to if there was no tension in
this bicep. It is this tensing up of the PC muscle that forces
out the semen. And the larger the range of motion of the PC
muscle the more forceful the ejactulation. So keeping your PC muscle
loose allows you to have a more forceful ejactulation when you do
orgasm
Note : This is just stuff I have a read and found out
experimenting with myself, I have no formal medical or sex training.
Also you should be able to feel your PC muscle. When you go for
wee put your finger inbetween your anus/testis and push the pee out
hard, you should feel the PC muscle drop, then cut off the flow
of pee and the PC muscle will tense up.
If you need any more proof, when guys pee they will loosen their
PC muscle, howevever when it comes to the last few drops they try
to squeeze out, they will tense up their PC muscle. The motion of
the penis when doing this will be very similar to when a guy
ejactulates, notice how the penis seems to jump up.
Anyway there is my ramble on the topic, I really hope this helps.
(edited 11 years ago)
3 Report
Reply 6
11 years ago
Anonymous #2
I know how he feels.
i had this problem but well into the relationship. Me and my
Fiancee have a good sex life now, but i had a problem with not
lasting very long. There were times whenever i lasted about 20
seconds.
I wanna stress \- a wise guy once said that women should take it
as a compliment that he comes early.
If you want to help him \- you have to make the end of his
penis less sensitive. I know premature ejaculation can be due to
the head of the penis being quite over-sensitive \- but this isn't
the only thing that causes it.
Premature ejaculation can be caused by numerous factors \- both due
to mental and physical problems.
Physical being \- oversensitivity.
Mental being \- confidence problems, nervousness etc.
The best thing for you to do is to make him feel relaxed and
keep reassuring him that it doesn't bother you.
Do you give oral? that sometimes can help over-sensitivity but
obviously go slow on him and then work it faster... pardon me if
this sounds disgusting but if he ejaculates in your mouth \- keep
sucking cos this is when it is the most sensitive... If you keep
sucking it \- it will sort of condition him to be less sensitive.
Hope this helps you out.
0 Report
Reply 7
11 years ago
Anonymous #3
This has been said before, but if you don't use condoms then try
using them, at the rate he comes, probably the extra thick ones
would be best. Personal anecdote:
Without condoms I usually come around 10 or 15 minutes, but with
condoms I can't come at all (which is actually extremely frustrating
because I am trying to have safe sex but my partner never wants
to use anything else). They can make a huge difference for me,
even the relatively thin ones (I have never tried super-thin), as
they massively reduce sensitivity for the guy.
0 Report
Reply 8
11 years ago
Anonymous #4
Count your blessings, i only last 10\.
1 Report
Reply 9
11 years ago
Anonymous #5
I went to doctors recently and asked about my premature ejaculation
problem, obviously after my main question (concerning Hernia). I did
explain to the doctor that I was sexually active, and I have tried
all these keggle and shiz exercises. She simply said its through
experience and practice that it gets better.
0 Report
Reply 10
11 years ago
Clare~Bear
15
Delay condoms and lube?
0 Report
Reply 11
11 years ago
ilem
17
Have him take just about any SSRI class antidepressant. They're
literally anti-orgasm pills
2 Report
Reply 12
11 years ago
Sagga
5
There are special condoms with anaesthetic on te heads to delay
ejaculation.try those. It came up on the joys of teen sex last
year.
0 Report
Reply 13
11 years ago
leeds85
What happens if you have a break of 5mins after sex then give him
a bj to get him going again. A few of those and he'll be able
to control himself during sex, if he's really interested in
controlling himself that is?
0 Report
Reply 14
11 years ago
meloncoly
11
0 Report
Reply 15
11 years ago
BambieWambie
1
Wow 20 seconds? My ex lasted 2 minutes. I thought that was bad.
0 Report
Reply 16
11 years ago
Happydude
11
Original post by Classical Liberal
Has he tried "reverse kegels"?
I am under the impression, I don't actually know and rarely talk
to guys about things like this (for all guy bravado with regards
to sex they will not talk about it), that most involuntary
ejactulation is caused by the "PC" muscle (the muscle that controls
peeing, pooing and ejactulation) is too tense. The muscle being
tensed up stimulates the male "g spot" (no idea where this actually
is but I can definitely feel the sensation myself) which causes a
guy to ejactulate.
You can tense up the PC muscle when peeing. Try and stop the flow
of pee. You can relax the PC muscle by doing the opposite. By
trying to force out your pee as hard as is possible. You can also
loosen up the PC muscle by simulating taking a poo.
I know it sounds wierd but get your boyfriend to try simulate
having a ****/farting during sex/masturbating. This should release the
tension on the PC muscle (stretch it out) and reduce the
stimulation that leads to ejactulation. Ideally he should try to
simulate trying to take a **** and then simulate trying to pee
hard, however the latter is quite tricky without any urine. He will
also probably find it hard to breathe doing this and should feel
his abs flexing. Ideally with practice he should be able to learn
how to breathe and not flex his abs.
I suspect that when guys start feeling that sensation of just being
about to ejactulate, they desperately tense up the PC muscle to try
and block the semen (just like trying to stop the flow of pee)
however this just makes everything worse because it just stimulates
you even more.
As an added bonus to being able to last longer by stretching out
(removing the tension) the PC muscle he will also be able to have
more forceful orgasms. Take your arm and imagine you do not have a
tricep (only a bicep). If you tense up your bicep your arm bends.
Without your tricep the only way to straighten out your arm would
be to remove the tension from your bicep. Your PC muscle works
like this (girls have them too). If you had your bicep without any
tension and then you suddenly tensed it as much as possible, your
arm would go through a large range of motion with a lot of force.
Compare this to if you had your bicep already a bit tense and
then tensed it as much as possible. There would be less force and
a smaller range of motion compared to if there was no tension in
this bicep. It is this tensing up of the PC muscle that forces
out the semen. And the larger the range of motion of the PC
muscle the more forceful the ejactulation. So keeping your PC muscle
loose allows you to have a more forceful ejactulation when you do
orgasm
Note : This is just stuff I have a read and found out
experimenting with myself, I have no formal medical or sex training.
Also you should be able to feel your PC muscle. When you go for
wee put your finger inbetween your anus/testis and push the pee out
hard, you should feel the PC muscle drop, then cut off the flow
of pee and the PC muscle will tense up.
If you need any more proof, when guys pee they will loosen their
PC muscle, howevever when it comes to the last few drops they try
to squeeze out, they will tense up their PC muscle. The motion of
the penis when doing this will be very similar to when a guy
ejactulates, notice how the penis seems to jump up.
Anyway there is my ramble on the topic, I really hope this helps.
Those are muscles I didn't even know I had!
0 Report
Reply 17
11 years ago
Tedaus
8
I have the same problem My gf finishes so early.
0 Report
Reply 18
11 years ago
shirley7
11
Just before he's about to climax, tell him to think of his Granny
having a dump with crusty bogeys around her nose. Should give him
an extra minute or two.
2 Report
Reply 19
11 years ago
shirley7
11
Original post by Classical Liberal
Has he tried "reverse kegels"?
I am under the impression, I don't actually know and rarely talk
to guys about things like this (for all guy bravado with regards
to sex they will not talk about it), that most involuntary
ejactulation is caused by the "PC" muscle (the muscle that controls
peeing, pooing and ejactulation) is too tense. The muscle being
tensed up stimulates the male "g spot" (no idea where this actually
is but I can definitely feel the sensation myself) which causes a
guy to ejactulate.
You can tense up the PC muscle when peeing. Try and stop the flow
of pee. You can relax the PC muscle by doing the opposite. By
trying to force out your pee as hard as is possible. You can also
loosen up the PC muscle by simulating taking a poo.
I know it sounds wierd but get your boyfriend to try simulate
having a ****/farting during sex/masturbating. This should release the
tension on the PC muscle (stretch it out) and reduce the
stimulation that leads to ejactulation. Ideally he should try to
simulate trying to take a **** and then simulate trying to pee
hard, however the latter is quite tricky without any urine. He will
also probably find it hard to breathe doing this and should feel
his abs flexing. Ideally with practice he should be able to learn
how to breathe and not flex his abs.
I suspect that when guys start feeling that sensation of just being
about to ejactulate, they desperately tense up the PC muscle to try
and block the semen (just like trying to stop the flow of pee)
however this just makes everything worse because it just stimulates
you even more.
As an added bonus to being able to last longer by stretching out
(removing the tension) the PC muscle he will also be able to have
more forceful orgasms. Take your arm and imagine you do not have a
tricep (only a bicep). If you tense up your bicep your arm bends.
Without your tricep the only way to straighten out your arm would
be to remove the tension from your bicep. Your PC muscle works
like this (girls have them too). If you had your bicep without any
tension and then you suddenly tensed it as much as possible, your
arm would go through a large range of motion with a lot of force.
Compare this to if you had your bicep already a bit tense and
then tensed it as much as possible. There would be less force and
a smaller range of motion compared to if there was no tension in
this bicep. It is this tensing up of the PC muscle that forces
out the semen. And the larger the range of motion of the PC
muscle the more forceful the ejactulation. So keeping your PC muscle
loose allows you to have a more forceful ejactulation when you do
orgasm
Note : This is just stuff I have a read and found out
experimenting with myself, I have no formal medical or sex training.
Also you should be able to feel your PC muscle. When you go for
wee put your finger inbetween your anus/testis and push the pee out
hard, you should feel the PC muscle drop, then cut off the flow
of pee and the PC muscle will tense up.
If you need any more proof, when guys pee they will loosen their
PC muscle, howevever when it comes to the last few drops they try
to squeeze out, they will tense up their PC muscle. The motion of
the penis when doing this will be very similar to when a guy
ejactulates, notice how the penis seems to jump up.
Anyway there is my ramble on the topic, I really hope this helps.
That's the biggest load of *******s I've ever heard.
1 Report
### Related discussions
* House viewing, Should I go alone?
* My boyfriend dumped me over these reasons
* issues (??) in my relationship
* Stuck on Past Paper
* Does body count matter ?
* have a fear and insecurity of oral sex
* If I got average mark in my second yr could I still get a first class
* Is my friend toxic?
* i should stop thinking about him
* URGENT !!
* struggling with dating
* Tired of never being anyone’s first choice
* Need help !
* Why do so many girls have a boyfriend? Am I really unlucky?
* How to cope with the cost of living as a student
* Normal body count
* veterinary medicine as a second degree
* I'm clueless
* Boyfriend going on a lads holiday!!
* Need some advice
Show 10 more
### Latest
### Trending
Last reply 1 day ago
Scared to get drunk 🥴
Health
10
Last reply 2 days ago
Acne
Health
16
Last reply 3 days ago
How much sleep do you get?
Health
97
Last reply 3 days ago
Knee injury - advice pls
Health
10
Last reply 4 days ago
will i still grow
Health
21
Last reply 6 days ago
Thinking to stop using spf
Health
5
Last reply 1 week ago
Annoying Red Cheeks
Health
12
Last reply 1 week ago
Toilet water splashback; is it deep?
Health
4
Last reply 1 week ago
Arthritis - can anyone help?
Health
11
Last reply 2 weeks ago
Bought a Rabbit but doesn’t fit, help?
Health
4
Last reply 2 weeks ago
More tired after drinking coffee / energy drinks
Health
7
Last reply 2 weeks ago
How am I gaining weight
Health
22
Last reply 3 weeks ago
Need help with sleep routine and time
Health
5
Last reply 3 weeks ago
Tired of feeling *****y with chronic symptoms for the last month
Health
6
Last reply 3 weeks ago
i'm done
Health
1
Last reply 3 weeks ago
How bad is it to take 30-50mg of codeine a day
Health
82
Last reply 3 weeks ago
IBS-D ruins my life
Health
13
Last reply 4 weeks ago
Always tired when studying
Health
6
Last reply 1 month ago
I feel like people shame me for exercising?
Health
13
Last reply 1 month ago
Eating with braces/ flossing
Health
16
### Trending
Last reply 1 day ago
Scared to get drunk 🥴
Health
10
Last reply 2 days ago
Acne
Health
16
Last reply 3 days ago
How much sleep do you get?
Health
97
Last reply 3 days ago
Knee injury - advice pls
Health
10
Last reply 4 days ago
will i still grow
Health
21
Last reply 6 days ago
Thinking to stop using spf
Health
5
Last reply 1 week ago
Annoying Red Cheeks
Health
12
Last reply 1 week ago
Toilet water splashback; is it deep?
Health
4
Last reply 1 week ago
Arthritis - can anyone help?
Health
11
Last reply 2 weeks ago
Bought a Rabbit but doesn’t fit, help?
Health
4
Last reply 2 weeks ago
More tired after drinking coffee / energy drinks
Health
7
Last reply 2 weeks ago
How am I gaining weight
Health
22
Last reply 3 weeks ago
Need help with sleep routine and time
Health
5
Last reply 3 weeks ago
Tired of feeling *****y with chronic symptoms for the last month
Health
6
Last reply 3 weeks ago
i'm done
Health
1
Last reply 3 weeks ago
How bad is it to take 30-50mg of codeine a day
Health
82
Last reply 3 weeks ago
IBS-D ruins my life
Health
13
Last reply 4 weeks ago
Always tired when studying
Health
6
Last reply 1 month ago
I feel like people shame me for exercising?
Health
13
Last reply 1 month ago
Eating with braces/ flossing
Health
16
### Articles for you
* What can you do with an arts degree?
* Can I change my choices when applying to university?
* Help! I’m having second thoughts about my uni choice
* What my law degree was like - from the students themselves
The Student Room and The Uni Guide are both part of The Student Room Group.
##### Get Started
* Today's posts
* Unanswered posts
* Community guidelines
##### Using TSR
* TSR help centre
* Cookies & online safety
* Terms & conditions
* Privacy policy
* Ad Privacy Settings
##### Info
* About TSR
* Advertise
* Contact us
##### Connect with TSR
* Twitter
* Facebook
* YouTube
* Instagram
* TikTok
© Copyright The Student Room 2024 all rights reserved
The Student Room and The Uni Guide are trading names of The Student Room Group
Ltd.
Register Number: 04666380 (England and Wales), VAT No. 806 8067 22 Registered
Office: Imperial House, 2nd Floor, 40-42 Queens Road, Brighton, East Sussex,
BN1 3XB
| biology | 3824512 | https://sv.wikipedia.org/wiki/Hister%20planepunctatus | Hister planepunctatus | Hister planepunctatus är en skalbaggsart som beskrevs av Desbordes 1914. Hister planepunctatus ingår i släktet Hister och familjen stumpbaggar. Inga underarter finns listade i Catalogue of Life.
Källor
Stumpbaggar
planepunctatus | swedish | 1.535072 |
sex_duration/how-long-does-sex-last.txt | * Health Conditions
* Health Conditions
* Alzheimer's & Dementia
* Anxiety
* Asthma & Allergies
* Atopic Dermatitis
* Breast Cancer
* Cancer
* Cardiovascular Health
* COVID-19
* Diabetes
* Environment & Sustainability
* Exercise & Fitness
* Eye Health
* Headache & Migraine
* Health Equity
* HIV & AIDS
* Human Biology
* Leukemia
* LGBTQIA+
* Men's Health
* Mental Health
* Multiple Sclerosis (MS)
* Nutrition
* Parkinson's Disease
* Psoriasis
* Psoriatic Arthritis
* Sexual Health
* Ulcerative Colitis
* Women's Health
* Health Products
* Health Products
* Nutrition & Fitness
* Vitamins & Supplements
* CBD
* Sleep
* Mental Health
* At-Home Testing
* Men’s Health
* Women’s Health
* Discover
* News
* Latest News
* Original Series
* Medical Myths
* Honest Nutrition
* Through My Eyes
* New Normal Health
* Podcasts
* 2023 in medicine
* Why exercise is key to living a long and healthy life
* What do we know about the gut microbiome in IBD?
* My podcast changed me
* Can 'biological race' explain disparities in health?
* Why Parkinson's research is zooming in on the gut
* Tools
* General Health
* Drugs A-Z
* Health Hubs
* Health Tools
* Find a Doctor
* BMI Calculators and Charts
* Blood Pressure Chart: Ranges and Guide
* Breast Cancer: Self-Examination Guide
* Sleep Calculator
* Quizzes
* RA Myths vs Facts
* Type 2 Diabetes: Managing Blood Sugar
* Ankylosing Spondylitis Pain: Fact or Fiction
* Connect
* About Medical News Today
* Who We Are
* Our Editorial Process
* Content Integrity
* Conscious Language
* Newsletters
* Sign Up
* Follow Us
Medical News Today
* Health Conditions
* Health Products
* Discover
* Tools
* Connect
Subscribe
# Sex: How long does it last?
Medically reviewed by Janet Brito, Ph.D., LCSW, CST — By Zawn Villines —
Updated on November 9, 2023
* How long is average?
* Variables
* Strategies
* Summary
There is no set time for how long sex should last. It can vary in length
depending on personal preference and other factors, such as what a person
considers sex to be.
People define sex differently. One person might only consider it to entail
penetrative intercourse, while another might consider sex to start with the
beginning of foreplay and last beyond each partner’s orgasm.
Evidence of how long sex tends to last is difficult to obtain, even
anecdotally. A person might feel pressure to be untruthful if the duration of
sex for them differs from perceived cultural norms.
There is also a distinction between how long sex actually lasts and how long
some people feel it should. Some studies show that penile-vaginal
intercourse lasts, on average, less time than women in monogamous, “stable”
heterosexual relationships need to have an orgasm, for example.
### How long does sex last? Here is what we know so far
There is little research into how long sex lasts, and many factors can affect
it, including what counts as “sex” and whether it follows orgasm time or
activities. Some studies have attempted to use self-reported data to determine
a general length. These studies are limited, but an older 2005 study found a
self-reported average to be 5.4 minutes. More research is needed to fully
understand it.
Was this helpful?
##
What does the research say?
Share on Pinterest Ibai Acevedo/Stocksy
There is very little published research into how long sex usually lasts.
Existing studies have significant limitations — most, for example, considered
sex only to be penile-vaginal intercourse or only included heterosexual
couples.
A 2005 multinational study of heterosexual couples that defined sex as
penile-vaginal intercourse asked participants to time sex from penetration
through to male ejaculation.
Within their very limited parameters, the team found that reports ranged from
33 seconds to 44 minutes, with the average session lasting 5.4 minutes.
Other researchers have attempted to ascertain a “normal” duration by asking
people who diagnose and treat sexual disorders.
A 2008 study asked sex therapists in the United States and Canada to
estimate the average duration of sex within different categories.
The therapists responded that sex:
* lasting under 3 minutes warrants clinical concern
* lasting 3–7 minutes is “adequate”
* lasting 7–13 minutes is “desirable”
* lasting 10–30 minutes is “too long”
However, a 2020 study assessing the time to orgasm in heterosexual women
found that the average was 13.41 minutes. This suggests that the therapists
polled in 2008 would consider sex that satisfies many heterosexual females to
last “too long.”
Cultural norms, including guidelines from medical professionals, can influence
sexual expectations and play a role in sexual dissatisfaction.
The findings of a 2010 study also suggest that a vaginal orgasm correlates
with having penile-vaginal penetrative sex for a longer period.
##
What can affect how long sex lasts?
A range of factors can contribute to the duration of sex, including:
* What counts as sex: Some people define any sexually stimulating contact as sex. Overall, people with a broader definition may consider their sex to last longer.
* Sexual practices, goals, and norms: The goal of sex, such as one orgasm for each partner, can influence the duration.
* External constraints: For example, new parents might have a quick sex session while their baby takes a nap, or they might arrange for a night away when they can spend hours on foreplay.
* Sexual orientation: A 2014 study found that females in same-sex couples have sex that lasts longer compared with the sex of other types of couples.
* Sexual function and overall health: Pain during or after sex and premature ejaculation are just a few examples of issues that can limit the duration of sex.
* Age: The duration may decrease with age due to factors affecting health and stamina.
* Geography: The multinational 2005 study found that heterosexual couples in Turkey have the shortest sex, lasting, on average, 3.7 minutes. This may speak to cultural norms, conditioning, and the study’s very limited definition of sex.
##
Strategies
There are several ways people can make sex longer or finish more quickly.
### Ways to make sex last longer
If the aim is to make sex last longer, people may want to consider the
following strategies:
* When one partner is approaching orgasm, they might take a break and focus on pleasure in other ways. Some toys and tools , medications, and techniques can also be used to delay orgasm .
* Continually communicate about sexual needs and desires. This can help each partner enjoy sex more.
* If a person with a vagina is involved, a partner can make sure to consider their orgasm by offering manual clitoral stimulation or oral sex, for example.
* Focus on foreplay before penetration. This may extend the length of the interaction and boost pleasure for everyone involved.
* Use visualization exercises and deep breathing to delay orgasm.
### Ways to make sex finish more quickly
If people want sex to finish more quickly, perhaps due to time constraints,
there are ways to speed it up. For example, by:
* considering using stimulation tools and toys and lubricant to help partners orgasm more quickly
* exploring ways to increase stimulation, such as considering specific interests or kinks and using media that can increase arousal
* optimizing clean-up techniques when time is short
* considering prescheduling some alone time to make sure there is enough time to enjoy the session
##
Summary
There is no right or wrong regarding the duration of sex — no standard
definition of what is typical.
People should communicate what makes sex feel good and satisfying for them and
structure the encounter around this. Having continual open communication and
focusing on each individual’s pleasure can generally make sex better.
Last medically reviewed on March 21, 2022
* Men's Health
* Sexual Health / STDs
* Women's Health / Gynecology
### How we reviewed this article:
Sources
Medical News Today has strict sourcing guidelines and draws only from peer-
reviewed studies, academic research institutions, and medical journals and
associations. We avoid using tertiary references. We link primary sources —
including studies, scientific references, and statistics — within each article
and also list them in the resources section at the bottom of our articles. You
can learn more about how we ensure our content is accurate and current by
reading our editorial policy .
* Bhat, G. S., et al . (2020). Time to orgasm in women in a monogamous stable heterosexual relationship.
https://www.sciencedirect.com/science/article/abs/pii/S1743609520300308
* Blair, K. L., et al . (2014). Can less be more? Comparing duration vs. frequency of sexual encounters in same-sex and mixed-sex relationships.
https://psycnet.apa.org/record/2014-43312-008
* Brody, S., et al . (2010). Vaginal orgasm is associated with vaginal (not clitoral) sex education, focusing mental attention on vaginal sensations, intercourse duration, and a preference for a longer penis.
https://www.sciencedirect.com/science/article/abs/pii/S1743609515331520
* Corty, E. W., et al . (2008). Canadian and American sex therapists' perceptions of normal and abnormal ejaculatory latencies: How long should intercourse last?
https://pubmed.ncbi.nlm.nih.gov/18331255
* Waldinger, M. D., et al . (2005). A multinational population survey of intravaginal ejaculation latency time.
https://pubmed.ncbi.nlm.nih.gov/16422843
Share this article
Medically reviewed by Janet Brito, Ph.D., LCSW, CST — By Zawn Villines —
Updated on November 9, 2023
## Latest news
* Wearable tech uses AI to detect cardiac arrhythmia 30 minutes before onset
* Artificial sweetener neotame may have potential to damage gut, lead to IBS
* AI tool may help detect cancer in a few minutes with a drop of blood
* Regularly eating avocado linked to lower diabetes risk in women
* Misplaced your keys? How to distinguish dementia from normal age-related memory loss
### Related Coverage
* How often should couples have sex?
There is no specific number of times a couple should have sex. Learn more
about how often couples have sex here.
READ MORE
* * How does sex affect your brain?
Sex can truly be the spice of life, but what does it do to the brain, and how
does it impact our hormonal activity? This Spotlight investigates.
READ MORE
* What to know about cleaning up after sex
What are the best ways to clean up after sex and prevent infections? Read on
for tips and suggestions for cleaning up after different types of sex.
READ MORE
* Everything you need to know about tantric sex
What is tantric sex? Read on discover the definition, processes, breathing
techniques, and positions of tantric sex, as well as how to prepare for it.
READ MORE
* About Us
* Contact Us
* Terms of Use
* Privacy Policy
* Privacy Settings
* Advertising Policy
* Health Topics
* Health Hubs
* Medical Affairs
* Content Integrity
* Newsletters
* © 2024 Healthline Media UK Ltd, Brighton, UK. All rights reserved. MNT is the registered trade mark of Healthline Media. Any medical information published on this website is not intended as a substitute for informed medical advice and you should not take any action before consulting with a healthcare professional. See additional information .
© 2024 Healthline Media UK Ltd, Brighton, UK. All rights reserved. MNT is the
registered trade mark of Healthline Media. Any medical information published
on this website is not intended as a substitute for informed medical advice
and you should not take any action before consulting with a healthcare
professional. See additional information .
About Careers Advertise with us
OUR BRANDS
Healthline Medical News Today Greatist Psych Central Bezzy
| biology | 846423 | https://no.wikipedia.org/wiki/Hemmet%20seksuell%20opphisselse%20hos%20kvinner | Hemmet seksuell opphisselse hos kvinner | Hemmet seksuell opphisselse hos kvinner (FSAD), ofte referert til som frigiditet , er en lidelse preget av en vedvarende eller gjentatt manglende evne til å oppnå seksuell opphisselse eller å opprettholde opphisselse til fullføringen av en seksuell aktivitet eller en tilstrekkelig lubrikasjon – fuktighets-oppsvulmings-reaksjon som ellers er stede under opphisselse og seksuell aktivitet. Tilstanden bør skilles fra et generelt tap av interesse for seksuell aktivitet og fra andre seksuell dysfunksjoner, slik som kvinnelig orgasmeforstyrrelse (anorgasmi) og hypoaktiv seksuell dysfunksjon (manglende lyst) som karakteriseres som mangel eller fravær på seksuelle fantasier og begjær for seksuell aktivitet over en viss periode.
Selv om kvinnelig seksuell dysfunksjon for tiden er en omdiskutert diagnose, begynner farmasøytiske selskaper å markedsføre produkter til behandling av hypoaktiv seksuell dysfunksjon (lystforstyrrelse) ofte med lave doser av testosteron.
Inndeling
Det finnes flere inndelinger på nedsatt seksuell respons hos kvinner. Disse kan indikere utbruddet: primær (livslang) eller sekundær (ervervet). De kan være basert på kontekst: det kan være i enhver sammenheng (generalisert) eller situasjonsbetinget. For eksempel kan problemet være tilstede med en ektefelle, mens det ikke er noe problem med en annen partner.
Hvor lenge tilstanden har vart, og i hvilken grad den er partner- eller situasjonsspesifikk, kan være et resultat av ulike årsaksfaktorer og kan påvirke behandlingen for tilstanden. Det kan skyldes psykologiske faktorer eller en kombinasjon av faktorer.
Diagnostiske trekk
I DSM-IV (American Psychiatric Association 1994) er diagnostiske kriterier for kvinnelig seksuell opphisselse lidelser:
Vedvarende eller tilbakevendende sviktende evne til å oppnå tilstrekkelig lubrikasjon (fuktighets-oppsvulmingsreaksjon på seksuell opphisselse) til gjennomføring av seksuell aktivitet.
Forstyrrelsen forårsaker merkbart ubehag eller mellommenneskelige problemer
Den seksuelle dysfunksjon beskrives ikke bedre av en annen Akse I-lidelse (utenom andre seksuelle dysfunksjoner) og skyldes ikke utelukkende de direkte fysiologiske virkninger av en substans (for eksempel legemiddel eller stoffmisbruk) eller en underliggende medisinsk tilstand.
Marita P. McCabe har merket seg at
"Problemer oppstår med denne definisjonen i forhold til hva som utgjør tilstrekkelig lubrikasjon. Det finnes ingen "gullstandard" på hvor lang tid det skal ta å bli opphisset eller på grad av opphisselse som skal oppnås. Disse responsene kan variere fra kvinne til kvinne og er avhengig av en rekke faktorer, inkludert humøret generelt når seksuell stimulering begynner og partners ferdigheter i å stimulere henne. Det kan også være forskjeller mellom fysiologisk og subjektiv grad av opphisselse. Noen kvinner sier de ikke føler seg seksuelt opphisselse tross økt fysiologisk respons som blodgjennomstrømning i skjedeveggen, mens andre sier de opplever seg seksuelt opphisset uten at det er påvisbar fysiologisk respons. Forventninger og tidligere erfaringer hos klinikere og klienter kan også lede dem til å klassifisere samme symptomene som sviktende seksuell respons hos en kvinne, men ikke hos en annen. "
Årsaker
En rekke studier har undersøkt hvilke faktorer som bidrar til hemmet seksuell respons og orgasmeforstyrrelse hos kvinner. Det omfatter både psykologiske og fysiske faktorer. Psykologisk sett kan følger av erfaringer fra barndom- og oppvekst, samt aktuelle hendelser – både hos kvinnen selv og i den aktuelle parrelasjonen være mulige årsaker til problemet.
Følger av hendelser i barndoms- og ungdomstiden
De fleste studier som har vurdert følgene av opplevelser i oppveksten på kvinnelig seksuell dysfunksjon blir vurdert som potensielt metodisk svake. Studiene stoler på retrospektiv hukommelse, noe som er spesielt problematisk når følelsesmessige reaksjoner på hendelsen blir rapportert så vel som den faktiske hendelsen . Det ser imidlertid ut til å være koblinger mellom seksuelle overgrep i barndommen og senere seksuelle problemer.
Enkeltfaktorer hos kvinnen
Det har vært lite undersøkelser av følgen av enkeltfaktorer på kvinnelig seksuell dysfunksjon. Slike faktorer er stress, tretthetsgrad, kjønnsidentitet, helse og andre individuelle egenskaper og erfaringer som kan påvirke seksuell lyst eller respons. Overeksponering for pornografilignende media antas å føre til dårlig kroppsbilde, selvbilde og nedsatt selvfølelse. En enkeltpersons seksuelle aktivitet kan forstyrres av overveldende følelsesmessig ubehag som resulterer i manglende evne til å oppnå seksuell nytelse.
Faktorer i parforholdet
En betydelig mengde vitenskapelige studier har utforsket rollen mellommenneskelige faktorer spiller på kvinnelig seksuell dysfunksjon, spesielt i forhold til orgasmerespons. Disse studiene har i stor grad fokusert på følger av kvaliteten på forholdet når det gjelder den seksuelle funksjonen til partnerne. Noen studier har vurdert betydningen av spesifikke relasjonsvariabler, mens andre har undersøkt generell tilfredshet i forholdet. Noen studier har utforsket hendelser, mens andre har fokusert på holdninger som et empirisk mål på hvordan forholdet fungerer. Populasjoner som har blitt undersøkt har variert fra ulykkelige par og seksuelt dysfunksjonelle klienter til par med høy grad av tilfredshet i forholdet.
Fysiske faktorer
Anslag over andelen kvinnelig seksuell dysfunksjon som kan tilskrives fysiske faktorer har variert fra 30 % til 80 %. Lidelsene som med størst sannsynlighet kan føre til seksuell dysfunksjon er de som fører til forstyrrelser i kar- eller nervefunksjon. Disse faktorene har blitt mer omfattende undersøkt blant menn enn hos kvinner. Fysiske årsaker som for eksempel nevrologiske og hjerte- og karsykdommer har vært direkte innblandet i både tidlig og forsinket utløsning, samt erektil dysfunksjon (Hawton 1993), men bidraget av fysiologiske faktorer til kvinnelig seksuelle dysfunksjoner er ikke like tydelig. Men nyere litteratur tyder på at det kan være en svekkelse i opphisselsesfasen hos kvinner med diabetes. Gitt at kvinner med diabetes viser en betydelig variasjon i sine svar på denne medisinske lidelsen, er det ikke overraskende at sykdommens påvirkning av opphisselse også er svært variabel. Faktisk tyder mangelen på en klar sammenheng mellom medisinske lidelser og seksuell funksjon på at psykologiske faktorer spiller en betydelig rolle i virkningen av disse lidelsene på seksuell funksjon (Melman et al. 1988).
Kenneth Maravilla, professor i radiologi og nevrologisk kirurgi og direktør for MR Research Laboratory ved University of Washington, Seattle, presenterte forskningsresultater basert på bildediagnostikk av kvinners seksuelle funksjon. I en liten pilotstudie på fire kvinner med hemmet seksuell opphisselse, rapporterte Maravilla at det var mindre hjerneaktivisering sett i denne gruppen, blant annet svært lite aktivering i amygdala. Disse kvinnene viste også økt aktivering i temporale områder, i motsetning til kvinner uten seksuelle problemer som viste deaktivering i lignende områder. Dette kan tyde på økt grad av hemming ved opphisselsesstimuli hos denne lille gruppen av kvinner med FSAD.
Behandling
Selv om det er stor variasjon i hvordan sviktende seksuell respons og orgasmeforstyrrelser uttrykkes hos kvinner, er det ingen dokumentasjon som tyder på at forskjellige faktorer bidrar til de to tilstandene, eller at ulike behandlingsstrategier bør brukes. Faktisk anvendes generelt de samme behandlingsstrategiene for begge tilstandene. Disse strategiene må kanskje suppleres med ekstra teknikker for å løse spesifikke problemer for enkelte kvinner, men de er generelt gode utgangspunkter for å løse problemene som bidrar til utvikling og vedlikehold av det seksuelle problemet. Fordi forholdet mellom kvinnen og hennes partner har vist seg å spille en betydelig rolle i både utvikling og vedlikehold av seksuelle problemer, er de fleste programmer laget slik at de skal gjennomføres av paret. Det finnes imidlertid også andre strategier som fokuserer på individet. Det er også viktig å skille tilstanden fra manglende seksuell lyst.
Kritiske bemerkninger
Begrepet hemmet seksuell respons hos kvinner er ikke uten kritiske bemerkninger.
Kilder
Female Sexual Arousal Disorder and Female Orgasmic Disorder''Se også
Hemmet seksuell opphisselse
Seksuell dysfunksjon
Seksuell funksjon
Anorgasmi
Referanser
Anbefalt lesing
Barlow David H (1986): The causes of sexual dysfunction: the role of anxiety and cognitive interference. Journal of Consulting and Clinical Psychology, 54, 140-148
Beck J Gayle og Barlow David H (1984): Current conceptualizations of sexual dysfunction: a review and an alternative perspective.'' Clinical Psychology Review, 9, 37-47.
Cahill C, Llewelyn SP, Pearson C: Long term effects of sexual abuse which occurred in childhood: a review. Br J Clin psychol 30:117-130, 1991
Delaney SM, McCabe MP: Secondary inorgasmia in women: a treatment program and case study. Sexual and Marital Therapy 3:165-190, 1988
Hallström T, Samuelsson S: Changes in women's sexual desire in middle life: the longitudinal study of women in Gothenburg. Arch Sex Behav 19:259-268, 1990
Hawton K: Sex Therapy. Oxford, UK, Oxford University Press, 1993
Heiman JR, Gladue BA, Roberts CW, et al:. Historical and current factors discriminating sexually functional from sexually dysfunctional married couples. J Marital Fam Ther 12:163-174, 1986
Hoch Z, Safir MP, Peres G, et al: An evaluation of sexual performance – comparison between sexually dysfunctional and functional couples. J Sex Marital Ther 7:195-206, 1981
The sexual genogram. J Marital Fam Ther 12:39-47, 1986
Hulbert DF. The role of assertiveness in female sexuality: a comparative study between sexually assertive and sexually non-assertive women. J Sex Marital Ther 17:183-190, 1991
Kilpatrick AC. Some correlates of women's childhood sexual experiences: a retrospective study. J Sex Res 22:221-242, 1986
Salamonsen LA: Hormonal activity in the endometrium: tissue remodelling and uterine bleeding, in Progress in the Management of Menopause. Redigert av Wren BG. London, Parthenon, 1997, pp 212–216
Laks UJ, Geist SH: The effects of androgens upon libido in women. Journal of Clinical Endocrinology 3:235-238, 1943
Segraves RT, Segraves KB. Human sexuality and aging. Journal of Sex Education and Therapy 21:88-102, 1995
Spector IP, Carey P: Incidence and prevalence of the sexual dysfunctions: a critical review of the empirical literature. Arch Sex Behav 19:389-408, 1990
Spector KR, Boyle M: The prevalence and perceived aetiology of male sexual problems in a non-clinical sample. Br J Med psychol 59:351-358, 1986
Eksterne lenker
Helsebibliotekets sider for kjønn og seksualitet
Nedsatt seksuell tenning hos kvinner på Norsk Helseinformatikk AS
Kvinnelig seksuell dysfunksjon i oppslagsverket BMJ Best Practice
Sexual Problems Overview Seksuelle problemer hos kvinner i Emedicine
Useful Addresses – The Sexual Dysfunction Association
A brief review that explores issues of the medicalization of the female orgasm
Our Bodies Ourselves chapter on Female Sexual Dysfunction: A Feminist View
Det kvinnelige reproduktive system
Seksuelle dysfunksjoner og kjønnsidentitetsforstyrrelser
Seksuell helse
Seksuell opphisselse
Kvinnehelse | norwegian_bokmål | 0.603964 |
sex_duration/how-to-last-longer-in-bed.txt | * Health Conditions
* Featured
* Breast Cancer
* IBD
* Migraine
* Multiple Sclerosis (MS)
* Rheumatoid Arthritis
* Type 2 Diabetes
* Sponsored Topics
* Articles
* Acid Reflux
* ADHD
* Allergies
* Alzheimer's & Dementia
* Bipolar Disorder
* Cancer
* Crohn's Disease
* Chronic Pain
* Cold & Flu
* COPD
* Depression
* Fibromyalgia
* Heart Disease
* High Cholesterol
* HIV
* Hypertension
* IPF
* Osteoarthritis
* Psoriasis
* Skin Disorders and Care
* STDs
* Discover
* Wellness Topics
* Nutrition
* Fitness
* Skin Care
* Sexual Health
* Women's Health
* Mental Well-Being
* Sleep
* Product Reviews
* Vitamins & Supplements
* Sleep
* Mental Health
* Nutrition
* At-Home Testing
* CBD
* Men’s Health
* Original Series
* Fresh Food Fast
* Diagnosis Diaries
* You’re Not Alone
* Present Tense
* Video Series
* Youth in Focus
* Healthy Harvest
* Through An Artist's Eye
* Future of Health
* Plan
* Health Challenges
* Mindful Eating
* Sugar Savvy
* Move Your Body
* Gut Health
* Mood Foods
* Align Your Spine
* Find Care
* Primary Care
* Mental Health
* OB-GYN
* Dermatologists
* Neurologists
* Cardiologists
* Orthopedists
* Lifestyle Quizzes
* Weight Management
* Am I Depressed? A Quiz for Teens
* Are You a Workaholic?
* How Well Do You Sleep?
* Tools & Resources
* Health News
* Find a Diet
* Find Healthy Snacks
* Drugs A-Z
* Health A-Z
* Connect
* Find Your Bezzy Community
* Breast Cancer
* Inflammatory Bowel Disease
* COPD
* Migraine
* Multiple Sclerosis
* Psoriasis
* Follow us on social media
Healthline
* Health Conditions
* Discover
* Plan
* Connect
Subscribe
#### Sexual Health
* Birth control
* STIs
* HIV
* HSV
* Activity
* Relationships
# How to Delay Ejaculation and Last Longer in Bed
Medically reviewed by Matt Coward, MD, FACS — By Jenna Fletcher and
Anna Schaefer — Updated on February 27, 2024
* Biofeedback
* Edging
* Kegels
* Desensitizers
* Circumcision
* Masturbation
* Weight
* Diet
* Stimulation
* Squeeze
* Mindfulness
* Arousal
* Condoms
* Medication
* Position
* FAQ
* Takeaway
Certain therapies may help extend the time before ejaculation during
masturbation and partner sex, including biofeedback, pelvic floor exercises
like Kegels, certain condoms, and more.
When it comes to partner sex, there are a lot of areas where doubt can creep
in. One common topic, especially for people with penises, is lasting long
enough in bed.
You may just want to increase how long your sexual encounters last. Or maybe
you experience premature ejaculation, which causes you to climax faster than
you’d like.
Either way, the following tips and tricks may help you enjoy a longer sexual
encounter.
##
1\. Biofeedback
Biofeedback is a process to retrain your brain and is used in a few different
therapeutic areas. When it comes to premature ejaculation , biofeedback
involves measuring electrical signals in the brain during masturbation or
other forms of stimulation.
If you want to try biofeedback, you’ll need to visit a physical therapist or
doctor’s office that specializes in or has experience with the technique. They
will use the measurements to help guide you in performing visualizations,
exercises, or other techniques to help you last longer during sex.
Dr. Philip Werthman , urologist and director of the Center for Male
Reproductive Medicine and Vasectomy Reversal in Los Angeles, CA, says this is
a viable solution for some people.
##
2\. Edging
Edging is the non-medical version of biofeedback. With edging, you masturbate
to bring yourself to just about ejaculation (or the “edge”) and then stop all
stimulating activity.
The idea is for you to become comfortable and knowledgeable about your sexual
arousal so you can prevent unintended ejaculation.
Edging may also enhance your sexual experience once you do ejaculate.
##
3\. Pelvic floor exercises
Your pelvic floor muscles help support both your bladder and ejaculation.
Exercises that help strengthen this muscle group may help improve your
ability to delay ejaculation.
According to Dr. Christopher Asandra , chief medical officer at NuMale
Medical Center, Kegel exercises may be able to help people with penises as
much as they do people with vulvas. But they require a consistent commitment.
They work by strengthening the pubococcygeus muscle.
“To do Kegel exercises, flex the same muscle you would use to stop the flow of
urine,” he said. “Clench this muscle for 10 seconds, then release, aiming for
at least three sets of 10 reps each day.”
##
4\. Desensitizers
Desensitizing creams, gels, and other products work to lessen the sensations
in your penis.
These products may also affect your sexual partner, so be sure to wipe or
clean off the product from the penis before stimulation.
Asandra says there’s one product he recommends, called Promescent , that
absorbs well enough so your partner isn’t affected.
Before you use a desensitizer, it’s important to note that these products
typically contain local anesthetics, which can prove dangerous if used too
frequently or in excess.
Always follow the recommendations on the product, or better yet, consult with
a doctor before use.
##
5\. Adult circumcision
Adult circumcision , or the removal of the foreskin, may also be an option.
A 2015 study showed promising results for people who got circumcised. They
noted that many of the people in the study found an increased control over
when they orgasm or a positive effect on premature ejaculation once
circumcised.
Consulting a doctor with experience in circumcisions may help you decide if
this method is right for you.
##
6\. Masturbation
Masturbation may help with preventing premature ejaculation. If you masturbate
shortly before partner sex, you may find that you can delay ejaculation during
partner play.
You also might find that you can use masturbation as a therapeutic massage.
According to a small 2019 study, researchers found that penis-root
masturbation may help you delay ejaculation.
They found that stimulating the root of the penis until ready to ejaculate and
then backing off helped to increase the time it took to ejaculate during
penetrative sex.
To perform penis-root masturbation, place both thumbs near the base of your
penis and rub both in a circular motion or up and down. You need to continue
the motion until you are just about to ejaculate and then back off.
##
7\. Find your optimal weight
Maintaining a moderate weight may improve your performance in bed.
According to a 2017 study, researchers found that people with three or more
of the following conditions have an increased likelihood of premature
ejaculation:
* high blood pressure
* low levels of “good” cholesterol
* waist circumference of 40 or more inches
* elevated blood sugar levels
* elevated blood triglycerides
“We know that obesity, high blood pressure, and diabetes are major causes of
erectile dysfunction, so treating the underlying problem may be beneficial,”
said Werthman.
##
8\. Change up your diet
Certain dietary changes may also help you delay ejaculation.
According to Werthman, some specific foods may increase the amount of free
testosterone in your body, which is necessary for healthy erectile function.
“Certain foods, like fenugreek (an herb), can increase free testosterone,”
he said. “As a matter of fact, fenugreek is the source of the compounds called
Testofen, found in many over-the-counter supplements.”
Note: An increase in free testosterone does not definitively correlate to
lasting longer in bed (or “longer ejaculatory latency”), so this method may
not prove as fruitful as others.
As with any dietary change, it’s a good idea to run your decision by your
doctor first, as they may know if any specific herbs or supplements could
interact with medications you’re already taking.
##
9\. Wait longer before penetration or skip it altogether
You may think of penile-vaginal or penile-anal sex as your ultimate goal when
you start to become intimate with your partner — but rushing to the goal may
be part of the problem.
If you find you prematurely ejaculate when engaging in penetrative sex,
focusing on other activities, like oral sex or sensual touch, may help.
You also may want to skip penis-in-vagina sex altogether. According to a 2017
study , about 18% of women experience orgasm from penetration. The rest
needed direct clitoris stimulation to orgasm, which means other sexual
activities may help both you and your partner feel satisfied.
##
10\. Squeeze technique
The squeeze technique may help you prevent ejaculation when simply stopping
stimulation will no longer work.
To perform this technique, you need to:
* discontinue manual stimulation or withdraw your penis from your partner
* use your hand to apply firm pressure to the tip of your penis
* return to sexual activity when the need to ejaculate passes
You can do this once or several times during sexual activity to help delay
ejaculation.
If you plan to do this during partner sex, let them know what you’re doing.
You may also want to figure out additional ways to stimulate them so they
remain aroused.
##
11\. Stay in the moment
You may have heard friends or others talking about focusing on non-sexual
things to prevent ejaculation or orgasm.
Instead, Amanda Holmberg , LMFT, clinic director at the Sexual Wellness
Institute, suggests you should “stay present in the moment with your body .”
“This will help you decide what you need,” she said. “Maybe you need to go a
little slower; maybe you need to take a little bit of a break completely… you
can’t do any of that if you’re thinking about baseball stats.”
In other words, staying focused on the task may help you find the solution to
managing premature ejaculation.
##
12\. Watch your arousal level going into partner sex
As part of being more in tune with your body, you should consider how aroused
you’re going into sex.
“If you are trying to last longer during penetrative sex, don’t penetrate at a
higher arousal level,” Holmberg said. “This is not setting you up for success.
Penetrate at a medium arousal level to give yourself some room to grow and
enjoy the pleasure.”
If you’re overly aroused when you start being intimate, your chances of
premature ejaculation increase greatly.
##
13\. Use condoms
Condoms can help desensitize the penis. If your usual brand isn’t making a
difference, you could try desensitizing condoms .
Desensitizing condoms often contain extra material or some of the ingredients
in desensitizing sprays and creams.
As an added benefit, condoms remain one of the best ways to reduce the risk
of sexually transmitted infections and unintended pregnancy.
##
14\. Reevaluate your medications
In certain cases, antidepressant medications known as selective serotonin
reuptake inhibitors (SSRIs) can sometimes be used to treat premature
ejaculation, says Asandra.
“Medications like SSRIs, such as Prozac, Paxil, and Zoloft , can delay
orgasm in men, but they can also cause problems,” he explains. “These drugs
take hours before they’re effective and can sometimes cause unwanted side
effects like dizziness, drowsiness, erectile dysfunction , and decreased
libido.”
While a 2016 study found that most of the antidepressants had either limited
effect or not enough evidence to support their use for premature ejaculation,
other studies have shown some benefits.
A 2019 review of studies featuring Paroxetine, an SSRI, as a treatment for
premature ejaculation found that the drug performed better than placebo and
had relatively mild side effects.
Because the research around using SSRIs to treat premature ejaculation is
mixed, and side effects can occur, if you’re working with a doctor to treat
this condition, medication may be one of your last options to try.
##
15\. Switch up your position
If all else fails, you can try switching up the position you’re in. This can
be enjoyable, confidence-boosting , and informative.
For example, you may find that having your partner on top may help with
reducing stimulation. You can experiment until you find a position that helps
prolong your climax and provides pleasure for your partner.
##
Frequently asked questions
There’s no one-size-fits-all answer to how long sexual activity should last.
What matters most is that all partners feel satisfied and comfortable.
Open communication with your partner can lead to a more fulfilling sexual
experience. Focus on pleasure and intimacy rather than a specific timeframe.
### Does drinking water make you last longer in bed?
Drinking water is important for overall health and well-being, including
sexual health. Still, there is no direct evidence to suggest that it can delay
ejaculation or boost your overall endurance.
### Does ibuprofen make you last longer in bed?
Ibuprofen is a pain reliever and anti-inflammatory medication. While it may
help with discomfort or inflammation, it is not known to have any significant
impact on sexual performance or duration.
##
The bottom line
Premature ejaculation is a fairly common condition, with about 30–75% of
people with male anatomy reportedly affected by it.
There’s no shame in consulting a professional to discuss your symptoms and how
to manage them. Premature ejaculation could be a sign of an underlying
condition, such as a hormonal imbalance or other sexual dysfunction, that
requires treatment.
Last medically reviewed on February 27, 2024
### How we reviewed this article:
Sources
History
Healthline has strict sourcing guidelines and relies on peer-reviewed studies,
academic research institutions, and medical associations. We avoid using
tertiary references. You can learn more about how we ensure our content is
accurate and current by reading our editorial policy .
* Abdel-Hamid IA, et al. (2016). The drug treatment of delayed ejaculation.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5001980/
* Asandra C. (2019). Personal interview.
* Bolat D, et al. (2017). The relationship between acquired premature ejaculation and metabolic syndrome: a prospective, comparative study.
https://www.nature.com/articles/ijir20173
* Crowdis M, et al. (2023). Premature ejaculation.
https://www.ncbi.nlm.nih.gov/books/NBK546701/
* Gao J, et al. (2015). Effects of adult male circumcision on premature ejaculation: Results from a prospective study in China.
https://www.hindawi.com/journals/bmri/2015/417846/
* Herbenick D, et al. (2017). Women's experiences with genital touching, sexual pleasure, and orgasm: results from a u.s. probability sample of women ages 18 to 94.
https://www.tandfonline.com/doi/abs/10.1080/0092623X.2017.1346530
* Holmberg A. (2019). Personal interview.
* Pastore AL, et al. (2014). Pelvic floor muscle rehabilitation for patients with lifelong premature ejaculation: a novel therapeutic approach.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4003840/
* Pastore AL, et al. (2018). Pelvic muscle floor rehabilitation as a therapeutic option in lifelong premature ejaculation: long-term outcomes.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6219291/
* Ma G-C, et al. (2019). Regular penis-root masturbation, a novel behavioral therapy in the treatment of primary premature ejaculation.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6859670/
* Saleh R, et al. (2021). An update on the treatment of premature ejaculation: A systematic review.
https://www.ncbi.nlm.nih.gov/labs/pmc/articles/PMC8451625/
* Werthman P. (2019). Personal interview.
* Zhang D, et al. (2019). Paroxetine in the treatment of premature ejaculation: a systematic review and meta-analysis.
https://bmcurol.biomedcentral.com/articles/10.1186/s12894-018-0431-7
Our experts continually monitor the health and wellness space, and we update
our articles when new information becomes available.
Current Version
Feb 27, 2024
Written By
Jenna Fletcher, Anna Schaefer
Edited By
Tess Catlett
Medically Reviewed By
Matt Coward, MD, FACS
Copy Edited By
Copy Editors
Feb 3, 2022
Written By
Jenna Fletcher, Anna Schaefer
Edited By
Jessica DiGiacinto
Medically Reviewed By
Joseph Brito, MD
Copy Edited By
Connor Rice
VIEW ALL HISTORY
Share this article
Medically reviewed by Matt Coward, MD, FACS — By Jenna Fletcher and
Anna Schaefer — Updated on February 27, 2024
### Read this next
* How Long Should Sex Really Last?
Medically reviewed by Janet Brito, Ph.D., LCSW, CST
It's hard to quantify how long sex typically lasts — or "should" last —
because there are countless ways to have sex. Limited research does offer
some…
READ MORE
* * Premature Ejaculation Wipes: Everything You Need to Know
Medically reviewed by Debra Sullivan, Ph.D., MSN, R.N., CNE, COI
Do you come earlier than you'd like to during sex? You may try benzocaine
wipes. They work by numbing the head and shaft of the penis. Read on to see…
READ MORE
* Managing Erectile Dysfunction: Tips for Sexually Frustrated Couples
Medically reviewed by University of Illinois
Even if you have erectile dysfunction and can’t maintain an erection, you
don’t have to give up on sex. Learn strategies to keep the spark alive.
READ MORE
* Best Online Services for Erectile Dysfunction (ED) Pills in 2024
For those experiencing the symptoms of erectile dysfunction, there are several
different medications available. We took a closer look at some of the…
READ MORE
* Is There a Link Between COVID-19 and Erectile Dysfunction?
Medically reviewed by Matt Coward, MD, FACS
COVID-19 can cause endothelial damage, which could contribute to erectile
dysfunction. Neurological effects and mental health changes may also play a…
READ MORE
* What Is the Most Effective Pill for Erectile Dysfunction (ED)?
The most effective ED pill for you will vary depending on your needs, health,
and other medications you may already be taking.
READ MORE
* What Is the Safest Drug for Erectile Dysfunction?
All FDA-approved prescription ED drugs are considered safe. However, there are
a few things to keep in mind before taking them.
READ MORE
* Can Semaglutide Cause or Increase the Risk of Erectile Dysfunction?
While most of the side effects from semaglutide are GI related, there are
concerns that some males may also experience side effects like ED.
READ MORE
* How to Get Harder Erections Without Medication
Medically reviewed by the Healthline Medical Network
There are several things you can do to get harder erections. Talking with your
partner, making a few lifestyle adjustments, and forming new habits may…
READ MORE
* Can Too Much Exercise Cause Erectile Dysfunction?
Medically reviewed by Matt Coward, MD, FACS
Does excessive exercise cause erectile dysfunction or increase certain health
risks? Here's what you need to know about the connection between them.
READ MORE
* About Us
* Contact Us
* Privacy Policy
* Privacy Settings
* Advertising Policy
* Health Topics
* Sitemap
* Medical Affairs
* Content Integrity
* Newsletters
* © 2024 Healthline Media LLC. All rights reserved. Our website services, content, and products are for informational purposes only. Healthline Media does not provide medical advice, diagnosis, or treatment. See additional information. See additional information .
© 2024 Healthline Media LLC. All rights reserved. Our website services,
content, and products are for informational purposes only. Healthline Media
does not provide medical advice, diagnosis, or treatment. See additional
information. See additional information .
About Careers Advertise with us
OUR BRANDS
Healthline Medical News Today Greatist Psych Central Bezzy
| biology | 825734 | https://no.wikipedia.org/wiki/Seksuell%20dysfunksjon | Seksuell dysfunksjon | Seksuell dysfunksjon (se også seksuell funksjon) refererer til vanskeligheter som oppleves av en person eller et par i alle stadier av en normal seksuell aktivitet, inkludert lyst, opphisselse eller orgasme.
Kategorier
Seksuell dysfunksjon kan klassifiseres i fire kategorier: nedsatt seksuell lyst, sviktende genital respons, orgastisk dysfunksjon og smerter i forbindelse med samleie.
Nedsatt seksuell lyst
Nedsatt seksuell lyst, eller nedsatt libido, er karakterisert ved manglende lyst på seksuell aktivitet eller fravær av seksuelle fantasier. Nedsettelsen kan vare en kort periode eller over lengre tid. Tilstanden varierer fra en generell mangel på seksuell lyst til mangel på seksuell lyst for den nåværende partner. Tilstanden kan ha startet etter en periode med normal seksuell funksjon, eller personen kan alltid ha hatt liten eller ingen seksuell lyst.
Årsakene varierer betydelig, men inkluderer en mulig nedgang i produksjonen av normalt østrogen hos kvinner eller testosteron hos både menn og kvinner. Andre årsaker kan være aldring, tretthet, graviditet, medisiner (for eksempel SSRI) eller psykiatriske tilstander, som depresjon og angst. Tap av libido fra SSRI reverseres vanligvis etter at SSRI er seponnert, men i noen tilfeller gjør det ikke. Selv om en rekke årsaker til lav seksuell lyst nevnes, har kun noen av disse vært gjenstand for empirisk forskning. Mange av årsakene er basert på terapeuters inntrykk.
Sviktende genital respons
Sviktende genital respons var tidligere kjent som frigiditet hos kvinner og impotens hos menn, men disse har nå blitt erstattet med mindre fordømmende betingelser. Impotens er nå kjent som erektil dysfunksjon, og frigiditet har blitt erstattet med en rekke begreper som beskriver spesifikke problemer med for eksempel lyst eller opphisselse.
For både menn og kvinner kan disse forholdene manifestere seg som en aversjon mot, og unngåelse av, seksuell kontakt med en partner. Hos menn kan det være at en (delvis eller fullstendig) ikke klarer å oppnå eller opprettholde en ereksjon, eller det kan være mangel på seksuell opphisselse og nytelse av seksuell aktivitet.
Det kan være medisinske årsaker til disse lidelsene, som redusert blodtilførsel eller mangel på vaginal smøring. Kronisk sykdom eller hvordan forholdet mellom partnerne er, kan også bidra. I motsetning til orgasmeforstyrrelser, skyldes erektil dysfunksjon hos menn i første rekke fysiske forhold. Suksessen til Viagra (sildenafilsitrat) vitner om dette.
Erektil dysfunksjon
Erektil dysfunksjon, eller impotens, er en seksuell dysfunksjon preget av manglende evne til å utvikle eller opprettholde en ereksjon av penis. Det finnes ulike bakenforliggende årsaker, som for eksempel skader på Nervi erigentes som hindrer eller forsinker ereksjon, eller diabetes, som bare reduserer blodstrømmen til vev i penis. Mange av disse årsakene til erektil dysfunksjon er reversible.
Årsakene til erektil dysfunksjon kan være psykiske eller fysiske. Psykologisk impotens kan ofte bli bedret av nesten alt som pasienten tror på, det er en veldig sterk placeboeffekt. Fysisk skade er mye mer alvorlig. En viktig fysisk årsak til ED er kontinuerlig eller alvorlig skade på Nervi erigentes. Disse nervene passerer ved siden av prostata og springer ut fra det sakrale plexus, og de kan bli skadet ved prostata- og colo-rektale operasjoner.
På grunn av deres pinlige natur og skammen pasientene føler, var temaet tabu i lang tid, og er gjenstand for mange myter. Folkelige remedier har lenge vært brukt. Innføringen av kanskje det første farmakologisk effektive middel for impotens, sildenafil (varenavn Viagra) på 1990-tallet førte til en bølge av offentlig oppmerksomhet, drevet delvis av nyhetsverdien av historiene, delvis av tung annonsering.
Det latinske ordet impotentia coeundi beskriver ganske enkelt manglende evne til å sette inn penis inn i skjeden. Det er nå stort sett erstattet av mer presise betegnelser.
Orgastisk dysfunksjon
Orgastisk dysfunksjon (orgasmeforstyrrelser) er vedvarende forsinkelser eller fravær av orgasme etter en normal seksuell opphisselsesfase. Lidelsen kan ha fysiske, psykiske, eller farmakologiske årsaker. SSRIer er en vanlig farmasøytisk årsak. SSRIer kan utsette orgasme eller fjerne den helt.
Smerter ved samleie
Smerte ved samleie påvirker nesten utelukkende kvinner og er kjent som dyspareuni (smertefullt samleie) eller vaginisme (en ufrivillig kramper i musklene i skjedeveggen som forstyrrer samleiet).
Dyspareuni kan være forårsaket av utilstrekkelig smøring (vaginal tørrhet) hos kvinner. Dårlig smøring kan skyldes utilstrekkelig spenning og stimulering, eller hormonelle endringer forårsaket av menopause, graviditet eller amming. Irritasjon fra prevensjonskremer og skum kan også forårsake tørrhet, det samme kan frykt og angst for sex gjøre.
Det er uklart nøyaktig hva som forårsaker vaginisme, men det er antatt at tidligere seksuelle traumer (for eksempel voldtekt eller overgrep) kan spille en rolle. En annen kvinnelig seksuell smertelidelse kalles vulvodynia eller vulva vestibulitt. I denne tilstanden opplever kvinner brennende smerter under sex som synes å være relatert til problemer med huden i vulva og vagina. Årsaken er ukjent.
Mindre vanlige seksuelle dysfunksjoner
Erektil dysfunksjon forårsaket av vaskulær sykdom er vanligvis sett bare blant eldre personer som har åreforkalkning. Vaskulær sykdom er vanlig hos personer som har diabetes, perifer vaskulær sykdom, hypertensjon og blant røykere. Når blodstrømmen til penis er svekket, er erektil dysfunksjon sluttresultatet.
Hormonmangel er en relativt sjelden årsak til erektil dysfunksjon. Hos menn med testikkelsvikt som Klinefelter syndrom eller de som har fått strålebehandling, kjemoterapi eller ble eksponert for kusmavirus i barndommen, kan testiklenes produksjon av testosteron svikte. Andre hormonelle årsaker til ereksjonssvikt er hjernesvulster, hypertyreose, hypotyreose eller sykdommer i binyrene.
Strukturelle abnormiteter i penis som Peyronies sykdom kan gjøre samleie vanskelig. Sykdommen er preget av tykke fibrøse bånd i penis som fører til en deformert utseende penis.
Narkotika er også en årsak til erektil dysfunksjon. Personer som tar medisiner for å senke blodtrykket, bruker antipsykotika, antidepressiva, beroligende midler, narkotika, antacida eller alkohol kan ha problemer med seksuell funksjon og tap av libido.
Priapisme er en smertefull ereksjon som forekommer i flere timer og forekommer i fravær av seksuell stimulering. Denne tilstanden oppstår når blodet blir fanget i penis, og ikke er i stand til å strømme ut igjen. Dersom tilstanden ikke umiddelbart behandles, kan det føre til alvorlige arr og permanente tap av erektil funksjon. Lidelsen forekommer hos unge menn og barn. Personer med sigdcelleanemi og de som misbruker visse medisiner kan ofte utvikle denne lidelsen.
Årsaker
Det er mange faktorer som kan resultere i at en person opplever en seksuell dysfunksjon.
Seksuell dysfunksjon kan oppstå fra emosjonelle faktorer, blant annet mellommenneskelige eller psykologiske problemer. Mangel på tillit og åpen kommunikasjon mellom partnerne kan være en årsak. Psykiske problemer som depresjon, seksuell frykt eller skyldfølelse, tidligere seksuelle traumer, seksuelle forstyrrelser kan også virke inn.
Seksuell dysfunksjon er spesielt vanlig blant folk som har angstlidelser. Ordinær engstelse kan selvsagt føre til erektil dysfunksjon hos menn uten psykiatriske problemer, men klinisk diagnostiserte sykdommer som panikklidelse er vanlige årsaker til prematur ejakulasjon og unngåelse av samleie. Smerter under samleie forekommer ofte sammen med angstlidelser hos kvinner.
Seksuell aktivitet kan også være påvirket av fysiske faktorer. Disse vil inkludere bruk av rusmidler, som alkohol, nikotin, narkotika, sentralstimulerende midler, antihypertensiva, antihistaminer, og noen psykofarmaka. For kvinner kan nesten alle fysiologiske endringer som påvirker det reproduktive systemet - premenstruelt syndrom, graviditet, postpartum, menopause - ha en negativ effekt på libido. Skader i ryggen kan også påvirke seksuell aktivitet, det samme gjelder problemer med en forstørret prostata, problemer med blodtilførsel, nerveskader (som ved ryggmargsskader). Sykdom, for eksempel diabetisk nevropati, multippel sklerose, tumorer og (sjelden) tertiær syfilis, kan også ha innvirkning på seksuell aktivitet. Det samme gjelder svikt i ulike organsystemer (som hjerte og lunger), endokrine sykdommer i skjoldbruskkjertelen, hypofysen eller binyrene, hormonelle mangler (lavt testosteron, østrogen eller androgener) og noen medfødte defekter.
Symptomer
Psykologiske seksuelle forstyrrelser
Den fjerde utgaven av Diagnostic and Statistical Manual of Mental Disorders lister følgende psykiske seksuelle forstyrrelser:
Hemmet seksuell lyst (se også aseksualitet som ikke er klassifisert som en lidelse)
Seksuell aversjon (unngåelse av eller mangel på ønske om samleie)
Hemmet seksuell opphisselse hos kvinner (svikt i normal smørerespons)
Erektil dysfunksjon
Kvinnelig orgasmeforstyrrelse (se anorgasmi)
Mannlig orgasmeforstyrrelse (se anorgasmi)
Prematur ejakulasjon
Dyspareuni
Vaginisme
Seksuelle avvik
PTSD som følge av kjønnslemlestelse eller seksuelle overgrep i barndommen
Anhedoni
Andre seksuelle problemer
Seksuell misnøye (ikke-spesifisert)
Erektil dysfunksjon
Seksuelt overførbare sykdommer
Manglende evne til å slappe av vaginale muskler nok til at samleie kan finne sted
Ulykkelighet eller forvirring knyttet til seksuell orientering
Transpersoner kan ha seksuelle problemer (før eller etter kirurgi), men det å faktisk være en transperson er ikke et seksuelt problem i seg selv.
Vedvarende seksuell opphisselse syndrom
Seksuell dysfunksjon etter bruk av SSRI
Seksuell avhengighet
Hyperseksualitet
Kjønnslemlestelse
Andre relaterte problemer
Pedofili
Nekrofili
Zoofili
Sterilitet
Parafili
Behandling for menn
Siden årsaken til seksuell dysfunksjon hos mange menn er relatert til angst om ytelse, kan psykoterapi hjelpe. Situasjonell angst oppstår fra en tidligere dårlig hendelse eller mangel på erfaring. Denne angsten fører ofte til utvikling av frykt for seksuell aktivitet og unngåelse. Unngåelse fører til en syklus av økt angst og desensitivisering av penis. I noen tilfeller kan erektil dysfunksjon skyldes disharmoni i parforholdet. Ekteskapsrådgiving anbefales i denne situasjonen.
Livsstilsendringer som for eksempel røykeslutt, samt avvenning fra narkotika- eller alkoholmisbruk kan også hjelpe i noen typer av erektil dysfunksjon.
Flere medisiner som Viagra, Cialis og Levitra har blitt tilgjengelige for å hjelpe menn med erektil dysfunksjon. Disse medikamentene virker for ca 60 % av pasientene.
Behandling for kvinner
Selv om det ikke er noen godkjente legemidler for å behandle kvinnelige seksuelle lidelser, drives det nå forskning på flere legemidler for å undersøke om de har effekt. En vakuumpumpe er det eneste godkjente medisinske utstyret for opphisselses- og orgasme forstyrrelser hos kvinner. Den er utformet for å øke blodtilførselen til klitoris og de ytre kjønnsorganer. Kvinner som opplever smerte med samleie, får ofte foreskrevet smertestillende eller desensitiviserende midler. Andre får foreskrevet smøreolje og / eller hormonbehandling. Mange pasienter med kvinnelig seksuell dysfunksjon blir ofte også henvist til psykolog for psykososial rådgivning.
En manuell fysioterapi, Wurn-teknikk som er utformet for å redusere bekken- og vaginal sammenklistring, kan også være gunstig for kvinner som opplever seksuell smerte og dysfunksjon. I en kontrollert studie, Increasing orgasm and decreasing intercourse pain by a manual physical therapy technique , fikk tjuetre (23) kvinner som hadde klagd over smertefullt samleie og / eller seksuell dysfunksjon et 20-timers program med manipulerende fysioterapi. Resultatene ble sammenliknet med den validerte Female Sexual Function Index, med post-test vs forundersøkelse skåre. Resultatene av behandlingen viste forbedringer i alle seks anerkjente områder av seksuell dysfunksjon. Resultatene var signifikante (P </ = 0,003) på alle mål, med individuelle mål og P-verdier som følger: begjær (P <0.001), opphisselse (P = 0.0033), smøring (P <0.001), orgasme (P <0.001), tilfredshet (P <0.001), og smerter (P <0.001). En annen studie for å forbedre seksuell funksjon hos pasienter med endometriose viste tilsvarende statistiske resultater.
Kliniske studier
De tidligste forsøkene på å behandle seksuelle dysfunksjoner, spesielt erektil dysfunksjon ble utført av muslimske leger og farmasøyter i middelalderens islamske verden. De var de første til å foreskrive medisiner for behandling av dette problemet, og de utviklet flere metoder for terapi for dette problemet, inkludert en metode hvor ettmedikament ble foreskrevet, og en kombinert metode der pasienten fikk enten et medikament eller mat. De fleste av disse medikamentene ble gitt oralt (gjennom munnen), men noen pasienter ble også behandlet med lokale metoder og gjennom urinrøret. Seksuelle dysfunksjoner ble behandlet med klinisk testede medikamenter i den islamske verden fra det 9. århundre til det 16. århundre av en rekke muslimske leger og farmasøyter, deriblant Muhammad ibn Zakariya Razi, Thabit bin Qurra, Ibn Al-Jazzar, Avicenna (The Canon of medicine) , Averroes, Ibn al-Baitar og Ibn al-Nafis (The Comprehensive Book on Medicine).
I moderne tid regner vi at egentlige kliniske studier av seksuelle problemer startet først i 1970 da Masters og Johnson's Human Sexual Inadequacy ble publisert. Det var resultatet av over ti års arbeid med 790 pasienter ved Reproductive Biology Research Foundation i St. Louis. Dette arbeidet sprang ut fra Masters og Johnson's tidligere Human Sexual Response (1966).
Før Masters og Johnson var den kliniske tilnærmingen til seksuelle problemer i hovedsak hentet fra tenkningen til Freud. Seksuelle problemer ble regnet som psykopatologi, og man så på sjansen for hjelp eller bedring med en viss pessimisme. Seksuelle problemer var bare symptomer på en underliggende sykdom, og den diagnostiske tilnærmingen var psykopatologisk. Det var lite skille mellom funksjonsproblemer og variasjoner, og man skilte heller ikke mellom perversjon og problemer. Til tross for arbeidet til psykoterapeuter som Balint ble seksuelle dysfunksjoner grovt delt inn i frigiditet eller impotens, tilstander som tidlig vekket negative konnotasjoner i populærkulturen.
Det Human Sexual Inadequacy oppnådde, var å flytte tenkning fra psykopatologi til læring , bare hvis et problem ikke ble bedre av opplæring, ville psykopatologiske problemer bli vurdert. Dessuten var behandlingen rettet mot par, mens partnerne tidligere ville bli behandlet individuelt. Masters og Johnson så at sex var en felles handling. De mente at seksuell kommunikasjon og ikke individuelle problemer, var det viktigste ved seksuelle problemer. De foreslo også samterapi, et matchende par av terapeuter til pasientene. De argumenterte med at en enslig mannlig terapeut ikke fullt ut kunne forstå kvinnelige problemer.
Den grunnleggende behandlingsprogrammet til Masters og Johnson var et intensivt to ukers program for å utvikle effektiv seksuell kommunikasjon. Programmet begynte med diskusjon og fortsatte deretter med fokus på sansing mellom paret for å utvikle felles erfaringer. Utfra erfaringene kunne man konstatere spesifikke vansker og tilnærmet seg disse med en spesifikk terapi. I et begrenset antall tilfeller med bare menn (41) hadde Masters og Johnson utviklet bruken av en kvinnelig surrogat, en tilnærming de snart oppga på grunn av de etiske, juridiske og andre problemstillinger det forårsaket.
Da de anslo omfanget av seksuelle problemer, hadde Masters og Johnson definert en grense mellom dysfunksjon og avvik. Dysfunksjoner var forbigående, og oppleves av de fleste mennesker, hos menn omfattet dysfunksjoner primær eller sekundær impotens, prematur ejakulasjon, manglende ejakulasjon, hos kvinner primær orgasmisk dysfunksjon og situasjonsbestemt orgasmisk dysfunksjon, smerter under samleie (dyspareuni) og vaginisme. Ifølge Masters og Johnson er seksuell opphisselse og klimaks en normal fysiologisk prosess hos alle funksjonelt intakte voksne, men selv om det er autonomt, kan det bli hemmet. Masters og Johnson behandlingsprogram for dysfunksjon var 81,1 % vellykket.
Til tross for arbeidet til Masters og Johnson, ble feltet i USA raskt gjenstand for tilnærminger som var entusiastiske snarere enn systematiske, noe som visket ut skillet mellom "berikelse" og terapi. Det har blitt hevdet at virkningen av arbeidet var slik at det ville være umulig å gjenta et slik rent eksperiment.
Referanser
Litteratur
Kaplan, Helen Singer, The New Sex Therapy: Active treatment of sexual dysfunctions , New York, Brunner / Mazel, 1974. ISBN 0876300832
Eksterne lenker
Erektil dysfunksjon hos Legemiddelhåndboka
Seksuell dysfunksjon hos menn i Tidsskrift for Den norske legeforening
Kvinnelig seksuell dysfunksjon i oppslagsverket Best Practice
Erektil dysfunksjon (impotens) i oppslagsverket Best Practice
Helsebibliotekets sider om kjønn og seksualitet
Helsenetts sider om seksualitet
International Society for Sexual Medicine
NIH site on sexual problems
100 FAQs about sexual dysfunction
"Premature E"
"The many mysteries of the female orgasm" – En lederartikkel om fremskritt i seksuelle farmakologi
Traumatice masturbatory syndro and other masturbation issues
Sexual Medicine Society of North America's hjemmeside: SexHealthMatters.org
Persistent Sexual Arousal Syndrome – Språk: Nederlandsk og engelsk
Seksualitet
Psykiske lidelser og atferdsforstyrrelser | norwegian_bokmål | 0.610565 |
induce_sleep/Adenosinediphosphate.txt | Jump to content
Main menu
Main menu
move to sidebar hide
Navigation
* [ Main page ](/wiki/Main_Page "Visit the main page \[z\]")
* [ Contents ](/wiki/Wikipedia:Contents "Guides to browsing Wikipedia")
* [ Current events ](/wiki/Portal:Current_events "Articles related to current events")
* [ Random article ](/wiki/Special:Random "Visit a randomly selected article \[x\]")
* [ About Wikipedia ](/wiki/Wikipedia:About "Learn about Wikipedia and how it works")
* [ Contact us ](//en.wikipedia.org/wiki/Wikipedia:Contact_us "How to contact Wikipedia")
* [ Donate ](https://donate.wikimedia.org/wiki/Special:FundraiserRedirector?utm_source=donate&utm_medium=sidebar&utm_campaign=C13_en.wikipedia.org&uselang=en "Support us by donating to the Wikimedia Foundation")
Contribute
* [ Help ](/wiki/Help:Contents "Guidance on how to use and edit Wikipedia")
* [ Learn to edit ](/wiki/Help:Introduction "Learn how to edit Wikipedia")
* [ Community portal ](/wiki/Wikipedia:Community_portal "The hub for editors")
* [ Recent changes ](/wiki/Special:RecentChanges "A list of recent changes to Wikipedia \[r\]")
* [ Upload file ](/wiki/Wikipedia:File_upload_wizard "Add images or other media for use on Wikipedia")
[ 
 
](/wiki/Main_Page)
[ Search ](/wiki/Special:Search "Search Wikipedia \[f\]")
Search
* [ Create account ](/w/index.php?title=Special:CreateAccount&returnto=Adenosine+diphosphate "You are encouraged to create an account and log in; however, it is not mandatory")
* [ Log in ](/w/index.php?title=Special:UserLogin&returnto=Adenosine+diphosphate "You're encouraged to log in; however, it's not mandatory. \[o\]")
Personal tools
* [ Create account ](/w/index.php?title=Special:CreateAccount&returnto=Adenosine+diphosphate "You are encouraged to create an account and log in; however, it is not mandatory")
* [ Log in ](/w/index.php?title=Special:UserLogin&returnto=Adenosine+diphosphate "You're encouraged to log in; however, it's not mandatory. \[o\]")
Pages for logged out editors [ learn more ](/wiki/Help:Introduction)
* [ Contributions ](/wiki/Special:MyContributions "A list of edits made from this IP address \[y\]")
* [ Talk ](/wiki/Special:MyTalk "Discussion about edits from this IP address \[n\]")
## Contents
move to sidebar hide
* (Top)
* 1 Bioenergetics
* 2 Cellular respiration Toggle Cellular respiration subsection
* 2.1 Catabolism
* 2.2 Glycolysis
* 2.3 Citric acid cycle
* 2.4 Oxidative phosphorylation
* 2.4.1 Mitochondrial ATP synthase complex
* 3 Blood platelet activation
* 4 See also
* 5 References
Toggle the table of contents
# Adenosine diphosphate
50 languages
* [ العربية ](https://ar.wikipedia.org/wiki/%D8%A3%D8%AF%D9%8A%D9%86%D9%88%D8%B3%D9%8A%D9%86_%D8%AB%D9%86%D8%A7%D8%A6%D9%8A_%D8%A7%D9%84%D9%81%D9%88%D8%B3%D9%81%D8%A7%D8%AA "أدينوسين ثنائي الفوسفات – Arabic")
* [ تۆرکجه ](https://azb.wikipedia.org/wiki/%D8%A2%D8%AF%D9%86%D9%88%D8%B2%DB%8C%D9%86_%D8%AF%DB%8C%D9%81%D9%88%D8%B3%D9%81%D8%A7%D8%AA "آدنوزین دیفوسفات – South Azerbaijani")
* [ Беларуская ](https://be.wikipedia.org/wiki/%D0%90%D0%B4%D1%8D%D0%BD%D0%B0%D0%B7%D1%96%D0%BD%D0%B4%D1%8B%D1%84%D0%B0%D1%81%D1%84%D0%B0%D1%82 "Адэназіндыфасфат – Belarusian")
* [ བོད་ཡིག ](https://bo.wikipedia.org/wiki/%E0%BD%A8%E0%BD%BA%E0%BC%8B%E0%BD%8C%E0%BD%B2%E0%BC%8B%E0%BD%94%E0%BD%B2%E0%BC%8B%E0%BD%A0%E0%BD%91%E0%BD%B4%E0%BD%A6%E0%BC%8B%E0%BD%A2%E0%BE%AB%E0%BD%A6%E0%BC%8D "ཨེ་ཌི་པི་འདུས་རྫས། – Tibetan")
* [ Bosanski ](https://bs.wikipedia.org/wiki/Adenozin-difosfat "Adenozin-difosfat – Bosnian")
* [ Català ](https://ca.wikipedia.org/wiki/Difosfat_d%27adenosina "Difosfat d'adenosina – Catalan")
* [ Čeština ](https://cs.wikipedia.org/wiki/Adenosindifosf%C3%A1t "Adenosindifosfát – Czech")
* [ Dansk ](https://da.wikipedia.org/wiki/Adenosindifosfat "Adenosindifosfat – Danish")
* [ Deutsch ](https://de.wikipedia.org/wiki/Adenosindiphosphat "Adenosindiphosphat – German")
* [ Eesti ](https://et.wikipedia.org/wiki/Adenosiindifosfaat "Adenosiindifosfaat – Estonian")
* [ Ελληνικά ](https://el.wikipedia.org/wiki/%CE%94%CE%B9%CF%86%CF%89%CF%83%CF%86%CE%BF%CF%81%CE%B9%CE%BA%CE%AE_%CE%B1%CE%B4%CE%B5%CE%BD%CE%BF%CF%83%CE%AF%CE%BD%CE%B7 "Διφωσφορική αδενοσίνη – Greek")
* [ Español ](https://es.wikipedia.org/wiki/Adenos%C3%ADn_difosfato "Adenosín difosfato – Spanish")
* [ Esperanto ](https://eo.wikipedia.org/wiki/Adenozina_pirofosfato "Adenozina pirofosfato – Esperanto")
* [ Euskara ](https://eu.wikipedia.org/wiki/Adenosina_difosfato "Adenosina difosfato – Basque")
* [ فارسی ](https://fa.wikipedia.org/wiki/%D8%A2%D8%AF%D9%86%D9%88%D8%B2%DB%8C%D9%86_%D8%AF%DB%8C%E2%80%8C%D9%81%D8%B3%D9%81%D8%A7%D8%AA "آدنوزین دیفسفات – Persian")
* [ Français ](https://fr.wikipedia.org/wiki/Ad%C3%A9nosine_diphosphate "Adénosine diphosphate – French")
* [ Gaeilge ](https://ga.wikipedia.org/wiki/D%C3%A9fhosf%C3%A1it_adan%C3%B3is%C3%ADn "Défhosfáit adanóisín – Irish")
* [ Galego ](https://gl.wikipedia.org/wiki/Adenos%C3%ADn_difosfato "Adenosín difosfato – Galician")
* [ 한국어 ](https://ko.wikipedia.org/wiki/%EC%95%84%EB%8D%B0%EB%85%B8%EC%8B%A0_%EC%9D%B4%EC%9D%B8%EC%82%B0 "아데노신 이인산 – Korean")
* [ Hrvatski ](https://hr.wikipedia.org/wiki/Adenozin_difosfat "Adenozin difosfat – Croatian")
* [ Bahasa Indonesia ](https://id.wikipedia.org/wiki/Adenosina_difosfat "Adenosina difosfat – Indonesian")
* [ Italiano ](https://it.wikipedia.org/wiki/Adenosina_difosfato "Adenosina difosfato – Italian")
* [ עברית ](https://he.wikipedia.org/wiki/ADP "ADP – Hebrew")
* [ Қазақша ](https://kk.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D0%B4%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозиндифосфат – Kazakh")
* [ Kreyòl ayisyen ](https://ht.wikipedia.org/wiki/ADP "ADP – Haitian Creole")
* [ Latina ](https://la.wikipedia.org/wiki/Adenosinum_diphosphoricum "Adenosinum diphosphoricum – Latin")
* [ Lietuvių ](https://lt.wikipedia.org/wiki/Adenozindifosfatas "Adenozindifosfatas – Lithuanian")
* [ Magyar ](https://hu.wikipedia.org/wiki/Adenozin-difoszf%C3%A1t "Adenozin-difoszfát – Hungarian")
* [ മലയാളം ](https://ml.wikipedia.org/wiki/%E0%B4%85%E0%B4%A1%E0%B4%BF%E0%B4%A8%E0%B5%8B%E0%B4%B8%E0%B4%BF%E0%B5%BB_%E0%B4%A1%E0%B5%88%E0%B4%AB%E0%B5%8B%E0%B4%B8%E0%B5%8D%E0%B4%AB%E0%B5%87%E0%B4%B1%E0%B5%8D%E0%B4%B1%E0%B5%8D "അഡിനോസിൻ ഡൈഫോസ്ഫേറ്റ് – Malayalam")
* [ Bahasa Melayu ](https://ms.wikipedia.org/wiki/Adenosina_difosfat "Adenosina difosfat – Malay")
* [ Nederlands ](https://nl.wikipedia.org/wiki/Adenosinedifosfaat "Adenosinedifosfaat – Dutch")
* [ 日本語 ](https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%87%E3%83%8E%E3%82%B7%E3%83%B3%E4%BA%8C%E3%83%AA%E3%83%B3%E9%85%B8 "アデノシン二リン酸 – Japanese")
* [ Occitan ](https://oc.wikipedia.org/wiki/Adenosina_difosfat "Adenosina difosfat – Occitan")
* [ Polski ](https://pl.wikipedia.org/wiki/Adenozyno-5%E2%80%B2-difosforan "Adenozyno-5′-difosforan – Polish")
* [ Português ](https://pt.wikipedia.org/wiki/Adenosina_difosfato "Adenosina difosfato – Portuguese")
* [ Română ](https://ro.wikipedia.org/wiki/Adenozin_difosfat "Adenozin difosfat – Romanian")
* [ Русский ](https://ru.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D0%B4%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозиндифосфат – Russian")
* [ Simple English ](https://simple.wikipedia.org/wiki/Adenosine_diphosphate "Adenosine diphosphate – Simple English")
* [ Slovenčina ](https://sk.wikipedia.org/wiki/Adenoz%C3%ADndifosf%C3%A1t "Adenozíndifosfát – Slovak")
* [ Српски / srpski ](https://sr.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD-%D0%B4%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозин-дифосфат – Serbian")
* [ Srpskohrvatski / српскохрватски ](https://sh.wikipedia.org/wiki/Adenozin_difosfat "Adenozin difosfat – Serbo-Croatian")
* [ Suomi ](https://fi.wikipedia.org/wiki/Adenosiinidifosfaatti "Adenosiinidifosfaatti – Finnish")
* [ Svenska ](https://sv.wikipedia.org/wiki/Adenosindifosfat "Adenosindifosfat – Swedish")
* [ Türkçe ](https://tr.wikipedia.org/wiki/Adenozin_difosfat "Adenozin difosfat – Turkish")
* [ Українська ](https://uk.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D0%B4%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозиндифосфат – Ukrainian")
* [ اردو ](https://ur.wikipedia.org/wiki/%D8%A7%DB%8C%DA%88%DB%8C%D9%86%D9%88%D8%B3%D8%A7%D8%A6%D9%86_%DA%88%D8%A7%D8%A6%DB%8C_%D9%81%D8%A7%D8%B3%D9%81%DB%8C%D9%B9 "ایڈینوسائن ڈائی فاسفیٹ – Urdu")
* [ Tiếng Việt ](https://vi.wikipedia.org/wiki/Adenosine_diphosphat "Adenosine diphosphat – Vietnamese")
* [ 吴语 ](https://wuu.wikipedia.org/wiki/%E4%BA%8C%E7%A3%B7%E9%85%B8%E8%85%BA%E8%8B%B7 "二磷酸腺苷 – Wu")
* [ 粵語 ](https://zh-yue.wikipedia.org/wiki/%E4%BA%8C%E7%A3%B7%E9%85%B8%E8%85%BA%E8%8B%B7 "二磷酸腺苷 – Cantonese")
* [ 中文 ](https://zh.wikipedia.org/wiki/%E4%BA%8C%E7%A3%B7%E9%85%B8%E8%85%BA%E8%8B%B7 "二磷酸腺苷 – Chinese")
[ Edit links
](https://www.wikidata.org/wiki/Special:EntityPage/Q185253#sitelinks-wikipedia
"Edit interlanguage links")
* [ Article ](/wiki/Adenosine_diphosphate "View the content page \[c\]")
* [ Talk ](/wiki/Talk:Adenosine_diphosphate "Discuss improvements to the content page \[t\]")
English
* [ Read ](/wiki/Adenosine_diphosphate)
* [ Edit ](/w/index.php?title=Adenosine_diphosphate&action=edit "Edit this page \[e\]")
* [ View history ](/w/index.php?title=Adenosine_diphosphate&action=history "Past revisions of this page \[h\]")
Tools
Tools
move to sidebar hide
Actions
* [ Read ](/wiki/Adenosine_diphosphate)
* [ Edit ](/w/index.php?title=Adenosine_diphosphate&action=edit "Edit this page \[e\]")
* [ View history ](/w/index.php?title=Adenosine_diphosphate&action=history)
General
* [ What links here ](/wiki/Special:WhatLinksHere/Adenosine_diphosphate "List of all English Wikipedia pages containing links to this page \[j\]")
* [ Related changes ](/wiki/Special:RecentChangesLinked/Adenosine_diphosphate "Recent changes in pages linked from this page \[k\]")
* [ Upload file ](/wiki/Wikipedia:File_Upload_Wizard "Upload files \[u\]")
* [ Special pages ](/wiki/Special:SpecialPages "A list of all special pages \[q\]")
* [ Permanent link ](/w/index.php?title=Adenosine_diphosphate&oldid=1176961502 "Permanent link to this revision of this page")
* [ Page information ](/w/index.php?title=Adenosine_diphosphate&action=info "More information about this page")
* [ Cite this page ](/w/index.php?title=Special:CiteThisPage&page=Adenosine_diphosphate&id=1176961502&wpFormIdentifier=titleform "Information on how to cite this page")
* [ Get shortened URL ](/w/index.php?title=Special:UrlShortener&url=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FAdenosine_diphosphate)
* [ Download QR code ](/w/index.php?title=Special:QrCode&url=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FAdenosine_diphosphate)
* [ Wikidata item ](https://www.wikidata.org/wiki/Special:EntityPage/Q185253 "Structured data on this page hosted by Wikidata \[g\]")
Print/export
* [ Download as PDF ](/w/index.php?title=Special:DownloadAsPdf&page=Adenosine_diphosphate&action=show-download-screen "Download this page as a PDF file")
* [ Printable version ](/w/index.php?title=Adenosine_diphosphate&printable=yes "Printable version of this page \[p\]")
In other projects
* [ Wikimedia Commons ](https://commons.wikimedia.org/wiki/Category:Adenosine_diphosphate)
From Wikipedia, the free encyclopedia
Adenosine diphosphate [ 
](/wiki/File:Adenosindiphosphat_protoniert.svg "Skeletal formula of ADP")
---
[ 
](/wiki/File:Adenosine-diphosphate-3D-balls.png "Ball-and-stick model of ADP
\(shown here as a 3- ion\)")
Names
[ IUPAC name ](/wiki/Chemical_nomenclature "Chemical nomenclature")
Adenosine 5′-(trihydrogen diphosphate)
[ Systematic IUPAC name ](/wiki/Chemical_nomenclature#Systematic_name
"Chemical nomenclature")
[(2 _R_ ,3 _S_ ,4 _R_ ,5 _R_ )-5-(6-Amino-9 _H_
-purin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl trihydrogen diphosphate
Other names
Adenosine 5′-diphosphate; Adenosine 5′-pyrophosphate; Adenosine pyrophosphate
Identifiers
[ CAS Number ](/wiki/CAS_Registry_Number "CAS Registry Number")
|
* [ 58-64-0 ](https://commonchemistry.cas.org/detail?cas_rn=58-64-0)  Y
3D model ( [ JSmol ](/wiki/JSmol "JSmol") )
|
* [ Interactive image ](https://chemapps.stolaf.edu/jmol/jmol.php?model=O%3DP%28O%29%28O%29OP%28%3DO%29%28O%29OC%5BC%40H%5D3O%5BC%40%40H%5D%28n2cnc1c%28ncnc12%29N%29%5BC%40H%5D%28O%29%5BC%40%40H%5D3O)
* [ Interactive image ](https://chemapps.stolaf.edu/jmol/jmol.php?model=c1nc%28c2c%28n1%29n%28cn2%29%5BC%40H%5D3%5BC%40%40H%5D%28%5BC%40%40H%5D%28%5BC%40H%5D%28O3%29COP%28%3DO%29%28O%29OP%28%3DO%29%28O%29O%29O%29O%29N)
[ ChEBI ](/wiki/ChEBI "ChEBI") |
* [ CHEBI:16761 ](https://www.ebi.ac.uk/chebi/searchId.do?chebiId=16761)  Y
[ ChEMBL ](/wiki/ChEMBL "ChEMBL") |
* [ ChEMBL14830 ](https://www.ebi.ac.uk/chembldb/index.php/compound/inspect/ChEMBL14830)  Y
[ ChemSpider ](/wiki/ChemSpider "ChemSpider") |
* [ 5800 ](https://www.chemspider.com/Chemical-Structure.5800.html)  Y
[ DrugBank ](/wiki/DrugBank "DrugBank") |
* [ DB03431 ](https://www.drugbank.ca/drugs/DB03431)  N
[ ECHA InfoCard ](/wiki/ECHA_InfoCard "ECHA InfoCard") | [ 100.000.356
](https://echa.europa.eu/substance-information/-/substanceinfo/100.000.356) [

](https://www.wikidata.org/wiki/Q185253#P2566 "Edit this at Wikidata")
[ EC Number ](/wiki/European_Community_number "European Community number") |
* 218-249-0
[ IUPHAR/BPS ](/wiki/IUPHAR/BPS "IUPHAR/BPS")
|
* [ 1712 ](http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?tab=summary&ligandId=1712)
[ KEGG ](/wiki/KEGG "KEGG") |
* [ C00008 ](https://www.kegg.jp/entry/C00008)  N
[ PubChem ](/wiki/PubChem "PubChem") CID
|
* [ 6022 ](https://pubchem.ncbi.nlm.nih.gov/compound/6022)
[ RTECS number ](/wiki/RTECS "RTECS") |
* AU7467000
[ UNII ](/wiki/Unique_Ingredient_Identifier "Unique Ingredient Identifier") |
* [ 61D2G4IYVH ](https://precision.fda.gov/uniisearch/srs/unii/61D2G4IYVH)  Y
[ CompTox Dashboard ](/wiki/CompTox_Chemicals_Dashboard "CompTox Chemicals
Dashboard") ( EPA )
|
* [ DTXSID60883210 ](https://comptox.epa.gov/dashboard/chemical/details/DTXSID60883210) [  ](https://www.wikidata.org/wiki/Q185253#P3117 "Edit this at Wikidata")
[ InChI ](/wiki/International_Chemical_Identifier "International Chemical
Identifier")
* InChI=1S/C10H15N5O10P2/c11-8-5-9(13-2-12-8)15(3-14-5)10-7(17)6(16)4(24-10)1-23-27(21,22)25-26(18,19)20/h2-4,6-7,10,16-17H,1H2,(H,21,22)(H2,11,12,13)(H2,18,19,20)/t4-,6-,7-,10-/m1/s1  Y
Key: XTWYTFMLZFPYCI-KQYNXXCUSA-N
 Y
* InChI=1/C10H15N5O10P2/c11-8-5-9(13-2-12-8)15(3-14-5)10-7(17)6(16)4(24-10)1-23-27(21,22)25-26(18,19)20/h2-4,6-7,10,16-17H,1H2,(H,21,22)(H2,11,12,13)(H2,18,19,20)/t4-,6-,7-,10-/m1/s1
Key: XTWYTFMLZFPYCI-KQYNXXCUBP
[ SMILES ](/wiki/Simplified_molecular-input_line-entry_system "Simplified
molecular-input line-entry system")
* O=P(O)(O)OP(=O)(O)OC[C@H]3O[C@@H](n2cnc1c(ncnc12)N)[C@H](O)[C@@H]3O
* c1nc(c2c(n1)n(cn2)[C@H]3[C@@H]([C@@H]([C@H](O3)COP(=O)(O)OP(=O)(O)O)O)O)N
Properties
[ Chemical formula ](/wiki/Chemical_formula "Chemical formula")
| C 10 H 15 N 5 O 10 P 2
[ Molar mass ](/wiki/Molar_mass "Molar mass") | 427.201 g/mol
[ Density ](/wiki/Density "Density") | 2.49 g/mL
[ log _P_ ](/wiki/Partition_coefficient "Partition coefficient") | -2.640
Hazards
[ Safety data sheet ](/wiki/Safety_data_sheet "Safety data sheet") (SDS) | [
MSDS
](http://www.sigmaaldrich.com/MSDS/MSDS/DisplayMSDSPage.do?country=PL&language=EN-
generic&productNumber=01905&brand=SIGMA&PageToGoToURL=http%3A//www.sigmaaldrich.com/catalog/product/sigma/01905%3Flang%3Dpl)
Except where otherwise noted, data are given for materials in their [ standard
state ](/wiki/Standard_state "Standard state") (at 25 °C [77 °F], 100 kPa).
 N [ verify
](https://en.wikipedia.org/w/index.php?title=Special:ComparePages&rev1=477242390&page2=Adenosine+diphosphate)
( [ what is ](/wiki/Wikipedia:WikiProject_Chemicals/Chembox_validation
"Wikipedia:WikiProject Chemicals/Chembox validation")
 Y
 N ?)
[ Infobox references ](/wiki/Wikipedia:Chemical_infobox#References
"Wikipedia:Chemical infobox")
Chemical compound
**Adenosine diphosphate** ( **ADP** ), also known as **adenosine
pyrophosphate** ( **APP** ), is an important [ organic compound
](/wiki/Organic_compound "Organic compound") in [ metabolism
](/wiki/Metabolism "Metabolism") and is essential to the flow of energy in
living [ cells ](/wiki/Cells_\(biology\) "Cells \(biology\)") . ADP consists
of three important structural components: a [ sugar ](/wiki/Sugar "Sugar")
backbone attached to [ adenine ](/wiki/Adenine "Adenine") and two [ phosphate
](/wiki/Phosphate "Phosphate") groups bonded to the 5 carbon atom of [ ribose
](/wiki/Ribose "Ribose") . The diphosphate group of ADP is attached to the 5’
carbon of the sugar backbone, while the adenine attaches to the 1’ carbon.
[1]
ADP can be interconverted to [ adenosine triphosphate
](/wiki/Adenosine_triphosphate "Adenosine triphosphate") (ATP) and [ adenosine
monophosphate ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
(AMP). ATP contains one more phosphate group than does ADP. AMP contains one
fewer phosphate group. Energy transfer used by all living things is a result
of [ dephosphorylation ](/wiki/Dephosphorylation "Dephosphorylation") of ATP
by enzymes known as [ ATPases ](/wiki/ATPase "ATPase") . The cleavage of a
phosphate group from ATP results in the coupling of energy to metabolic
reactions and a by-product of ADP. [1] ATP is continually reformed from
lower-energy species ADP and AMP. The biosynthesis of ATP is achieved
throughout processes such as [ substrate-level phosphorylation
](/wiki/Substrate-level_phosphorylation "Substrate-level phosphorylation") , [
oxidative phosphorylation ](/wiki/Oxidative_phosphorylation "Oxidative
phosphorylation") , and [ photophosphorylation ](/wiki/Photophosphorylation
"Photophosphorylation") , all of which facilitate the addition of a phosphate
group to ADP.
## Bioenergetics [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=1 "Edit
section: Bioenergetics") ]
ADP cycling supplies the [ energy ](/wiki/Energy "Energy") needed to do work
in a biological system, the [ thermodynamic ](/wiki/Thermodynamic
"Thermodynamic") process of transferring energy from one source to another.
There are two types of energy: [ potential energy ](/wiki/Potential_energy
"Potential energy") and [ kinetic energy ](/wiki/Kinetic_energy "Kinetic
energy") . Potential energy can be thought of as stored energy, or usable
energy that is available to do work. Kinetic energy is the energy of an object
as a result of its motion. The significance of ATP is in its ability to store
potential energy within the phosphate bonds. The energy stored between these
bonds can then be transferred to do work. For example, the transfer of energy
from ATP to the protein [ myosin ](/wiki/Myosin "Myosin") causes a
conformational change when connecting to [ actin ](/wiki/Actin "Actin") during
[ muscle contraction ](/wiki/Muscle_contraction "Muscle contraction") . [1]
[ 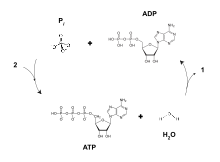 ](/wiki/File:ATP-ADP.svg) The cycle of synthesis and
degradation of ATP; 1 and 2 represent output and input of energy,
respectively.
It takes multiple reactions between myosin and actin to effectively produce
one muscle contraction, and, therefore, the availability of large amounts of
ATP is required to produce each muscle contraction. For this reason,
biological processes have evolved to produce efficient ways to replenish the
potential energy of ATP from ADP. [2]
Breaking one of ATP's phosphorus bonds generates approximately 30.5 [
kilojoules ](/wiki/Kilojoule "Kilojoule") per [ mole ](/wiki/Mole_\(unit\)
"Mole \(unit\)") of ATP (7.3 [ kcal ](/wiki/Calorie "Calorie") ). [3] ADP
can be converted, or powered back to ATP through the process of releasing the
chemical energy available in food; in humans, this is constantly performed via
[ aerobic respiration ](/wiki/Aerobic_respiration "Aerobic respiration") in
the [ mitochondria ](/wiki/Mitochondrion "Mitochondrion") . [2] Plants use [
photosynthetic ](/wiki/Photosynthetic "Photosynthetic") pathways to convert
and store energy from sunlight, also conversion of ADP to ATP. [3] Animals
use the energy released in the breakdown of glucose and other molecules to
convert ADP to ATP, which can then be used to fuel necessary growth and cell
maintenance. [2]
## Cellular respiration [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=2 "Edit
section: Cellular respiration") ]
### Catabolism [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=3 "Edit
section: Catabolism") ]
The ten-step [ catabolic ](/wiki/Catabolic "Catabolic") pathway of [
glycolysis ](/wiki/Glycolysis "Glycolysis") is the initial phase of free-
energy release in the breakdown of [ glucose ](/wiki/Glucose "Glucose") and
can be split into two phases, the preparatory phase and payoff phase. ADP and
[ phosphate ](/wiki/Phosphate "Phosphate") are needed as precursors to
synthesize ATP in the payoff reactions of the [ TCA cycle ](/wiki/TCA_cycle
"TCA cycle") and [ oxidative phosphorylation ](/wiki/Oxidative_phosphorylation
"Oxidative phosphorylation") mechanism. [4] During the payoff phase of
glycolysis, the enzymes phosphoglycerate kinase and pyruvate kinase facilitate
the addition of a phosphate group to ADP by way of [ substrate-level
phosphorylation ](/wiki/Substrate-level_phosphorylation "Substrate-level
phosphorylation") . [5]
[
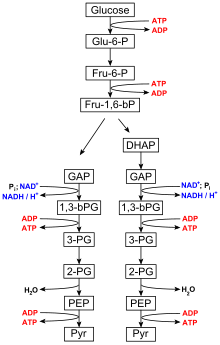 ](/wiki/File:Glycolysis_overview.svg) Glycolysis
overview
### Glycolysis [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=4 "Edit
section: Glycolysis") ]
Main article: [ glycolysis ](/wiki/Glycolysis "Glycolysis")
Glycolysis is performed by all living organisms and consists of 10 steps. The
net reaction for the overall process of [ glycolysis ](/wiki/Glycolysis
"Glycolysis") is: [6]
Glucose + 2 NAD+ + 2 P i \+ 2 ADP → 2 pyruvate + 2 ATP + 2 NADH + 2 H 2 O
Steps 1 and 3 require the input of energy derived from the hydrolysis of ATP
to ADP and P i (inorganic phosphate), whereas steps 7 and 10 require the
input of ADP, each yielding ATP. [7] The [ enzymes ](/wiki/Enzyme "Enzyme")
necessary to break down glucose are found in the [ cytoplasm ](/wiki/Cytoplasm
"Cytoplasm") , the viscous fluid that fills living cells, where the glycolytic
reactions take place. [1]
### Citric acid cycle [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=5 "Edit
section: Citric acid cycle") ]
Main article: [ citric acid cycle ](/wiki/Citric_acid_cycle "Citric acid
cycle")
The [ citric acid cycle ](/wiki/Citric_acid_cycle "Citric acid cycle") , also
known as the Krebs cycle or the TCA (tricarboxylic acid) cycle is an 8-step
process that takes the pyruvate generated by glycolysis and generates 4 NADH,
FADH2, and GTP, which is further converted to ATP. [8] It is only in step 5,
where GTP is generated, by succinyl-CoA synthetase, and then converted to ATP,
that ADP is used (GTP + ADP → GDP + ATP). [9]
### Oxidative phosphorylation [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=6 "Edit
section: Oxidative phosphorylation") ]
Main article: [ oxidative phosphorylation ](/wiki/Oxidative_phosphorylation
"Oxidative phosphorylation")
[ Oxidative phosphorylation ](/wiki/Oxidative_phosphorylation "Oxidative
phosphorylation") produces 26 of the 30 equivalents of ATP generated in
cellular respiration by transferring electrons from NADH or FADH2 to [ O 2
](/wiki/Oxygen "Oxygen") through electron carriers. [10] The energy released
when electrons are passed from higher-energy NADH or FADH2 to the lower-energy
O 2 is required to phosphorylate ADP and once again generate ATP. [11] It
is this energy coupling and phosphorylation of ADP to ATP that gives the
electron transport chain the name oxidative phosphorylation. [1]
[ 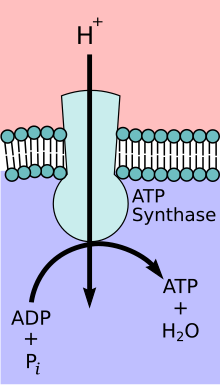 ](/wiki/File:ATP-Synthase.svg) ATP-
Synthase
#### Mitochondrial ATP synthase complex [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=7 "Edit
section: Mitochondrial ATP synthase complex") ]
Main article: [ ATP synthase ](/wiki/ATP_synthase "ATP synthase")
During the initial phases of [ glycolysis ](/wiki/Glycolysis "Glycolysis") and
the [ TCA cycle ](/wiki/TCA_cycle "TCA cycle") , [ cofactors
](/wiki/Cofactor_\(biochemistry\) "Cofactor \(biochemistry\)") such as [ NAD+
](/wiki/NAD%2B "NAD+") donate and accept electrons [12] that aid in the [
electron transport chain ](/wiki/Electron_transport_chain "Electron transport
chain") 's ability to produce a proton gradient across the inner mitochondrial
membrane. [13] The ATP synthase complex exists within the mitochondrial
membrane (F O portion) and protrudes into the matrix (F 1 portion). The
energy derived as a result of the chemical gradient is then used to synthesize
ATP by coupling the reaction of inorganic phosphate to ADP in the active site
of the [ ATP synthase ](/wiki/ATP_synthase "ATP synthase") enzyme; the
equation for this can be written as ADP + P i → ATP. [ _[ citation needed
](/wiki/Wikipedia:Citation_needed "Wikipedia:Citation needed") _ ]
## Blood platelet activation [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=8 "Edit
section: Blood platelet activation") ]
Under normal conditions, small disk-shape [ platelets ](/wiki/Platelet
"Platelet") circulate in the blood freely and without interaction with one
another. ADP is stored in [ dense bodies ](/wiki/Platelet "Platelet") inside [
blood ](/wiki/Blood "Blood") [ platelets ](/wiki/Platelet "Platelet") and is
released upon platelet activation. ADP interacts with a family of ADP
receptors found on platelets (P2Y1, [ P2Y12 ](/wiki/P2Y12 "P2Y12") , and
P2X1), which leads to platelet activation. [14]
* **P2Y1** receptors initiate platelet aggregation and shape change as a result of interactions with ADP.
* **P2Y12** receptors further amplify the response to ADP and draw forth the completion of aggregation.
ADP in the blood is converted to [ adenosine ](/wiki/Adenosine "Adenosine") by
the action of [ ecto-ADPases ](/wiki/Ecto-ADPase "Ecto-ADPase") , inhibiting
further platelet activation via [ adenosine receptors
](/wiki/Adenosine_receptor "Adenosine receptor") . [ _[ citation needed
](/wiki/Wikipedia:Citation_needed "Wikipedia:Citation needed") _ ]
## See also [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=9 "Edit
section: See also") ]
* [ Nucleoside ](/wiki/Nucleoside "Nucleoside")
* [ Nucleotide ](/wiki/Nucleotide "Nucleotide")
* [ DNA ](/wiki/DNA "DNA")
* [ RNA ](/wiki/RNA "RNA")
* [ Oligonucleotide ](/wiki/Oligonucleotide "Oligonucleotide")
* [ Apyrase ](/wiki/Apyrase "Apyrase")
* [ Phosphate ](/wiki/Phosphate "Phosphate")
* [ Adenosine diphosphate ribose ](/wiki/Adenosine_diphosphate_ribose "Adenosine diphosphate ribose")
## References [ [ edit
](/w/index.php?title=Adenosine_diphosphate&action=edit§ion=10 "Edit
section: References") ]
1. ^ _**a** _ _**b** _ _**c** _ _**d** _ _**e** _ Cox, Michael; Nelson, David R.; Lehninger, Albert L (2008). [ _Lehninger principles of biochemistry_ ](https://archive.org/details/lehningerprincip00lehn_1) . San Francisco: W.H. Freeman. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-7167-7108-1 ](/wiki/Special:BookSources/978-0-7167-7108-1 "Special:BookSources/978-0-7167-7108-1") .
2. ^ _**a** _ _**b** _ _**c** _ Nave, C.R. (2005). [ "Adenosine Triphosphate" ](http://hyperphysics.phy-astr.gsu.edu/hbase/biology/atp.html) . _Hyper Physics [serial on the Internet]_ . Georgia State University.
3. ^ _**a** _ _**b** _ Farabee, M.J. (2002). [ "The Nature of ATP" ](https://web.archive.org/web/20071201180511/http://www.emc.maricopa.edu/faculty/farabee/BIOBK/BioBookATP.html) . _ATP and Biological Energy [serial on the Internet]_ . Archived from [ the original ](http://www.emc.maricopa.edu/faculty/farabee/BIOBK/BioBookATP.html) on 2007-12-01.
4. ** ^ ** Jensen TE, Richter EA (March 2012). [ "Regulation of glucose and glycogen metabolism during and after exercise" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3381815) . _J. Physiol_ . **590** (Pt 5): 1069–76. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1113/jphysiol.2011.224972 ](https://doi.org/10.1113%2Fjphysiol.2011.224972) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 3381815 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3381815) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 22199166 ](https://pubmed.ncbi.nlm.nih.gov/22199166) .
5. ** ^ ** Liapounova NA, Hampl V, Gordon PM, Sensen CW, Gedamu L, Dacks JB (December 2006). [ "Reconstructing the mosaic glycolytic pathway of the anaerobic eukaryote Monocercomonoides" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1694820) . _Eukaryotic Cell_ . **5** (12): 2138–46. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1128/EC.00258-06 ](https://doi.org/10.1128%2FEC.00258-06) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 1694820 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1694820) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 17071828 ](https://pubmed.ncbi.nlm.nih.gov/17071828) .
6. ** ^ ** Medh, J.D. [ "Glycolysis" ](http://www.csun.edu/~jm77307/Glycolysis.pdf) (PDF) . CSUN.Edu. [ Archived ](https://ghostarchive.org/archive/20221009/http://www.csun.edu/~jm77307/Glycolysis.pdf) (PDF) from the original on 2022-10-09 . Retrieved 3 April 2013 .
7. ** ^ ** Bailey, Regina. [ "10 Steps of Glycolysis" ](https://web.archive.org/web/20130515102637/http://biology.about.com/od/cellularprocesses/a/aa082704a.htm) . Archived from [ the original ](http://biology.about.com/od/cellularprocesses/a/aa082704a.htm) on 2013-05-15 . Retrieved 2013-05-10 .
8. ** ^ ** [ "Citric Acid Cycle" ](https://web.archive.org/web/20120324072437/http://web.ku.edu/~crystal/taksnotes/Biol_638/notes/chp_16.pdf) (PDF) . Takusagawa’s Note. Archived from [ the original ](http://web.ku.edu/~crystal/taksnotes/Biol_638/notes/chp_16.pdf) (PDF) on 24 March 2012 . Retrieved 4 April 2013 .
9. ** ^ ** [ "Biochemistry" ](https://web.archive.org/web/20130228175004/http://www.uccs.edu/~sbraunsa/Images/482Notes/17-TCAcycle.pdf) (PDF) . UCCS.edu. Archived from [ the original ](http://www.uccs.edu/~sbraunsa/Images/482Notes/17-TCAcycle.pdf) (PDF) on 2013-02-28.
10. ** ^ ** [ "Oxidative phosphorylation" ](https://www.ncbi.nlm.nih.gov/books/NBK21208/) . W H Freeman, 2002 . Retrieved 4 April 2013 .
11. ** ^ ** Medh, J. D. [ "Electron Transport Chain (Overview)" ](http://www.csun.edu/~jm77307/Oxidative%20Phosphorylation.pdf) (PDF) . CSUN.edu. [ Archived ](https://ghostarchive.org/archive/20221009/http://www.csun.edu/~jm77307/Oxidative%20Phosphorylation.pdf) (PDF) from the original on 2022-10-09 . Retrieved 4 April 2013 .
12. ** ^ ** Belenky P, Bogan KL, Brenner C (January 2007). "NAD+ metabolism in health and disease". _Trends Biochem. Sci_ . **32** (1): 12–9. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/j.tibs.2006.11.006 ](https://doi.org/10.1016%2Fj.tibs.2006.11.006) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 17161604 ](https://pubmed.ncbi.nlm.nih.gov/17161604) .
13. ** ^ ** Murray, Robert F. (2003). _Harper's illustrated biochemistry_ . New York: McGraw-Hill. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 0-07-121766-5 ](/wiki/Special:BookSources/0-07-121766-5 "Special:BookSources/0-07-121766-5") .
14. ** ^ ** Murugappa S, Kunapuli SP (2006). [ "The role of ADP receptors in platelet function" ](http://www.bioscience.org/2006/v11/af/1939/fulltext.htm) . _Front. Biosci_ . **11** : 1977–86. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.2741/1939 ](https://doi.org/10.2741%2F1939) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 16368572 ](https://pubmed.ncbi.nlm.nih.gov/16368572) .
* [ v ](/wiki/Template:Nucleobases,_nucleosides,_and_nucleotides "Template:Nucleobases, nucleosides, and nucleotides")
* [ t ](/wiki/Template_talk:Nucleobases,_nucleosides,_and_nucleotides "Template talk:Nucleobases, nucleosides, and nucleotides")
* [ e ](/wiki/Special:EditPage/Template:Nucleobases,_nucleosides,_and_nucleotides "Special:EditPage/Template:Nucleobases, nucleosides, and nucleotides")
[ Nucleic acid ](/wiki/Nucleic_acid "Nucleic acid") constituents
---
[ Nucleobase ](/wiki/Nucleobase "Nucleobase") |
* [ Purine ](/wiki/Purine "Purine")
* [ Adenine ](/wiki/Adenine "Adenine")
* [ Guanine ](/wiki/Guanine "Guanine")
* [ Hypoxanthine ](/wiki/Hypoxanthine "Hypoxanthine")
* [ Xanthine ](/wiki/Xanthine "Xanthine")
* [ Purine analogue ](/wiki/Purine_analogue "Purine analogue")
* [ Pyrimidine ](/wiki/Pyrimidine "Pyrimidine")
* [ Uracil ](/wiki/Uracil "Uracil")
* [ Thymine ](/wiki/Thymine "Thymine")
* [ Cytosine ](/wiki/Cytosine "Cytosine")
* [ Pyrimidine analogue ](/wiki/Pyrimidine_analogue "Pyrimidine analogue")
* [ Unnatural base pair (UBP) ](/wiki/Base_pair#Unnatural_base_pair_\(UBP\) "Base pair")
[ Nucleoside ](/wiki/Nucleoside "Nucleoside") |
| [ Ribonucleoside ](/wiki/Ribonucleoside "Ribonucleoside") |
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* [ Guanosine ](/wiki/Guanosine "Guanosine")
* [ 5-Methyluridine ](/wiki/5-Methyluridine "5-Methyluridine")
* [ Uridine ](/wiki/Uridine "Uridine")
* [ 5-Methylcytidine ](/wiki/5-Methylcytidine "5-Methylcytidine")
* [ Cytidine ](/wiki/Cytidine "Cytidine")
* [ Pseudouridine ](/wiki/Pseudouridine "Pseudouridine")
* [ Inosine ](/wiki/Inosine "Inosine")
* [ _N_ 6 -Methyladenosine ](/wiki/N6-Methyladenosine "N6-Methyladenosine")
* [ Xanthosine ](/wiki/Xanthosine "Xanthosine")
* [ Wybutosine ](/wiki/Wybutosine "Wybutosine")
---|---
[ Deoxyribonucleoside ](/wiki/Deoxyribonucleoside "Deoxyribonucleoside") |
* [ Deoxyadenosine ](/wiki/Deoxyadenosine "Deoxyadenosine")
* [ Deoxyguanosine ](/wiki/Deoxyguanosine "Deoxyguanosine")
* [ Thymidine ](/wiki/Thymidine "Thymidine")
* [ Deoxyuridine ](/wiki/Deoxyuridine "Deoxyuridine")
* [ Deoxycytidine ](/wiki/Deoxycytidine "Deoxycytidine")
* [ Deoxyinosine ](/w/index.php?title=Deoxyinosine&action=edit&redlink=1 "Deoxyinosine \(page does not exist\)")
* [ Deoxyxanthosine ](/w/index.php?title=Deoxyxanthosine&action=edit&redlink=1 "Deoxyxanthosine \(page does not exist\)")
[ Nucleotide ](/wiki/Nucleotide "Nucleotide")
(Nucleoside monophosphate) |
| [ Ribonucleotide ](/wiki/Ribonucleotide "Ribonucleotide") |
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* [ GMP ](/wiki/Guanosine_monophosphate "Guanosine monophosphate")
* [ m 5 UMP ](/w/index.php?title=5-Methyluridine_monophosphate&action=edit&redlink=1 "5-Methyluridine monophosphate \(page does not exist\)")
* [ UMP ](/wiki/Uridine_monophosphate "Uridine monophosphate")
* [ CMP ](/wiki/Cytidine_monophosphate "Cytidine monophosphate")
* [ IMP ](/wiki/Inosinic_acid "Inosinic acid")
* [ XMP ](/wiki/Xanthosine_monophosphate "Xanthosine monophosphate")
---|---
[ Deoxyribonucleotide ](/wiki/Deoxyribonucleotide "Deoxyribonucleotide") |
* [ dAMP ](/wiki/Deoxyadenosine_monophosphate "Deoxyadenosine monophosphate")
* [ dGMP ](/wiki/Deoxyguanosine_monophosphate "Deoxyguanosine monophosphate")
* [ dTMP ](/wiki/Thymidine_monophosphate "Thymidine monophosphate")
* [ dUMP ](/wiki/Deoxyuridine_monophosphate "Deoxyuridine monophosphate")
* [ dCMP ](/wiki/Deoxycytidine_monophosphate "Deoxycytidine monophosphate")
* [ dIMP ](/wiki/Deoxyinosine_monophosphate "Deoxyinosine monophosphate")
* [ dXMP ](/w/index.php?title=Deoxyxanthosine_monophosphate&action=edit&redlink=1 "Deoxyxanthosine monophosphate \(page does not exist\)")
[ Cyclic nucleotide ](/wiki/Cyclic_nucleotide "Cyclic nucleotide") |
* [ cAMP ](/wiki/Cyclic_adenosine_monophosphate "Cyclic adenosine monophosphate")
* [ cGMP ](/wiki/Cyclic_guanosine_monophosphate "Cyclic guanosine monophosphate")
* [ c-di-GMP ](/wiki/Cyclic_di-GMP "Cyclic di-GMP")
* [ c-di-AMP ](/wiki/Cyclic_di-AMP "Cyclic di-AMP")
* [ cADPR ](/wiki/Cyclic_ADP-ribose "Cyclic ADP-ribose")
* [ cGAMP ](/wiki/Cyclic_guanosine_monophosphate%E2%80%93adenosine_monophosphate "Cyclic guanosine monophosphate–adenosine monophosphate")
Nucleoside diphosphate |
|
* ADP
* [ GDP ](/wiki/Guanosine_diphosphate "Guanosine diphosphate")
* [ m 5 UDP ](/w/index.php?title=5-Methyluridine_diphosphate&action=edit&redlink=1 "5-Methyluridine diphosphate \(page does not exist\)")
* [ UDP ](/wiki/Uridine_diphosphate "Uridine diphosphate")
* [ CDP ](/wiki/Cytidine_diphosphate "Cytidine diphosphate")
* [ Xanthosine diphosphate ](/w/index.php?title=Xanthosine_diphosphate&action=edit&redlink=1 "Xanthosine diphosphate \(page does not exist\)")
---
* [ dADP ](/wiki/Deoxyadenosine_diphosphate "Deoxyadenosine diphosphate")
* [ dGDP ](/wiki/Deoxyguanosine_diphosphate "Deoxyguanosine diphosphate")
* [ dTDP ](/wiki/Thymidine_diphosphate "Thymidine diphosphate")
* [ dUDP ](/w/index.php?title=Deoxyuridine_diphosphate&action=edit&redlink=1 "Deoxyuridine diphosphate \(page does not exist\)")
* [ dCDP ](/wiki/Deoxycytidine_diphosphate "Deoxycytidine diphosphate")
[ Nucleoside triphosphate ](/wiki/Nucleoside_triphosphate "Nucleoside
triphosphate") |
|
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
* [ GTP ](/wiki/Guanosine_triphosphate "Guanosine triphosphate")
* [ m 5 UTP ](/wiki/5-Methyluridine_triphosphate "5-Methyluridine triphosphate")
* [ UTP ](/wiki/Uridine_triphosphate "Uridine triphosphate")
* [ CTP ](/wiki/Cytidine_triphosphate "Cytidine triphosphate")
* [ ITP ](/wiki/Inosine_triphosphate "Inosine triphosphate")
* [ XTP ](/wiki/Xanthosine_triphosphate "Xanthosine triphosphate")
---
* [ dATP ](/wiki/Deoxyadenosine_triphosphate "Deoxyadenosine triphosphate")
* [ dGTP ](/wiki/Deoxyguanosine_triphosphate "Deoxyguanosine triphosphate")
* [ dTTP ](/wiki/Thymidine_triphosphate "Thymidine triphosphate")
* [ dUTP ](/w/index.php?title=Deoxyuridine_triphosphate&action=edit&redlink=1 "Deoxyuridine triphosphate \(page does not exist\)")
* [ dCTP ](/wiki/Deoxycytidine_triphosphate "Deoxycytidine triphosphate")
* [ dITP ](/w/index.php?title=Deoxyinosine_triphosphate&action=edit&redlink=1 "Deoxyinosine triphosphate \(page does not exist\)")
* [ dXTP ](/w/index.php?title=Deoxyxanthosine_triphosphate&action=edit&redlink=1 "Deoxyxanthosine triphosphate \(page does not exist\)")
* [ v ](/wiki/Template:Neurotransmitters "Template:Neurotransmitters")
* [ t ](/wiki/Template_talk:Neurotransmitters "Template talk:Neurotransmitters")
* [ e ](/wiki/Special:EditPage/Template:Neurotransmitters "Special:EditPage/Template:Neurotransmitters")
[ Neurotransmitters ](/wiki/Neurotransmitter "Neurotransmitter")
---
[ Amino acid ](/wiki/Amino_acid "Amino acid") -derived |
| Major excitatory /
inhibitory systems |
| Glutamate system |
* [ Agmatine ](/wiki/Agmatine "Agmatine")
* [ Aspartic acid (aspartate) ](/wiki/Aspartic_acid "Aspartic acid")
* [ Glutamic acid (glutamate) ](/wiki/Glutamate_\(neurotransmitter\) "Glutamate \(neurotransmitter\)")
* [ Glutathione ](/wiki/Glutathione "Glutathione")
* [ Glycine ](/wiki/Glycine "Glycine")
* [ GSNO ](/wiki/S-Nitrosoglutathione "S-Nitrosoglutathione")
* [ GSSG ](/wiki/Oxidized_glutathione "Oxidized glutathione")
* [ Kynurenic acid ](/wiki/Kynurenic_acid "Kynurenic acid")
* [ NAA ](/wiki/N-Acetylaspartic_acid "N-Acetylaspartic acid")
* [ NAAG ](/wiki/N-Acetylaspartylglutamic_acid "N-Acetylaspartylglutamic acid")
* [ Proline ](/wiki/Proline "Proline")
* [ Serine ](/wiki/Serine "Serine")
---|---
GABA system |
* [ GABA ](/wiki/Gamma-Aminobutyric_acid "Gamma-Aminobutyric acid")
* [ GABOB ](/wiki/Gamma-Amino-beta-hydroxybutyric_acid "Gamma-Amino-beta-hydroxybutyric acid")
* [ GHB ](/wiki/Gamma-Hydroxybutyric_acid "Gamma-Hydroxybutyric acid")
Glycine system |
* [ α-Alanine ](/wiki/Alanine "Alanine")
* [ β-Alanine ](/wiki/%CE%92-Alanine "Β-Alanine")
* [ Glycine ](/wiki/Glycine "Glycine")
* [ Hypotaurine ](/wiki/Hypotaurine "Hypotaurine")
* [ Proline ](/wiki/Proline "Proline")
* [ Sarcosine ](/wiki/Sarcosine "Sarcosine")
* [ Serine ](/wiki/Serine "Serine")
* [ Taurine ](/wiki/Taurine "Taurine")
[ GHB system ](/wiki/GHB_receptor "GHB receptor") |
* [ GHB ](/wiki/Gamma-Hydroxybutyric_acid "Gamma-Hydroxybutyric acid")
* [ T-HCA (GHC) ](/wiki/T-HCA "T-HCA")
[ Biogenic amines ](/wiki/Biogenic_amine "Biogenic amine") |
| Monoamines |
* [ 6-OHM ](/wiki/6-Hydroxymelatonin "6-Hydroxymelatonin")
* [ Dopamine ](/wiki/Dopamine "Dopamine")
* [ Epinephrine (adrenaline) ](/wiki/Adrenaline "Adrenaline")
* [ NAS (normelatonin) ](/wiki/N-Acetylserotonin "N-Acetylserotonin")
* [ Norepinephrine (noradrenaline) ](/wiki/Norepinephrine "Norepinephrine")
* [ Serotonin (5-HT) ](/wiki/Serotonin "Serotonin")
---|---
Trace amines |
* [ 3-Iodothyronamine ](/wiki/3-Iodothyronamine "3-Iodothyronamine")
* [ N-Methylphenethylamine ](/wiki/N-Methylphenethylamine "N-Methylphenethylamine")
* [ N-Methyltryptamine ](/wiki/N-Methyltryptamine "N-Methyltryptamine")
* [ _m_ -Octopamine ](/wiki/Norfenefrine "Norfenefrine")
* [ _p_ -Octopamine ](/wiki/Octopamine "Octopamine")
* [ Phenylethanolamine ](/wiki/Phenylethanolamine "Phenylethanolamine")
* [ Phenethylamine ](/wiki/Phenethylamine "Phenethylamine")
* [ Synephrine ](/wiki/Synephrine "Synephrine")
* [ Tryptamine ](/wiki/Tryptamine "Tryptamine")
* [ _m_ -Tyramine ](/wiki/Meta-Tyramine "Meta-Tyramine")
* [ _p_ -Tyramine ](/wiki/Tyramine "Tyramine")
Others |
* [ Histamine ](/wiki/Histamine "Histamine")
[ Neuropeptides ](/wiki/Neuropeptide "Neuropeptide") |
|
* See [ here ](/wiki/Template:Neuropeptides "Template:Neuropeptides") instead.
---
[ Lipid ](/wiki/Lipid "Lipid") -derived |
| Endocannabinoids |
* [ 2-AG ](/wiki/2-Arachidonoylglycerol "2-Arachidonoylglycerol")
* [ 2-AGE (noladin ether) ](/wiki/2-Arachidonyl_glyceryl_ether "2-Arachidonyl glyceryl ether")
* [ 2-ALPI ](/w/index.php?title=2-Arachidonoyl_lysophosphatidylinositol&action=edit&redlink=1 "2-Arachidonoyl lysophosphatidylinositol \(page does not exist\)")
* [ 2-OG ](/wiki/2-Oleoylglycerol "2-Oleoylglycerol")
* [ AA-5-HT ](/wiki/Arachidonoyl_serotonin "Arachidonoyl serotonin")
* [ Anandamide (AEA) ](/wiki/Anandamide "Anandamide")
* [ DEA ](/wiki/Docosatetraenoylethanolamide "Docosatetraenoylethanolamide")
* [ LPI ](/wiki/Lysophosphatidylinositol "Lysophosphatidylinositol")
* [ NADA ](/wiki/N-Arachidonoyl_dopamine "N-Arachidonoyl dopamine")
* [ NAGly ](/wiki/N-Arachidonylglycine "N-Arachidonylglycine")
* [ OEA ](/wiki/Oleoylethanolamide "Oleoylethanolamide")
* [ Oleamide ](/wiki/Oleamide "Oleamide")
* [ PEA ](/wiki/Palmitoylethanolamide "Palmitoylethanolamide")
* [ RVD-Hpα ](/wiki/RVD-Hp%CE%B1 "RVD-Hpα")
* [ SEA ](/wiki/Stearoylethanolamide "Stearoylethanolamide")
* [ Virodhamine (O-AEA) ](/wiki/Virodhamine "Virodhamine")
---|---
[ Neurosteroids ](/wiki/Neurosteroid "Neurosteroid") |
|
* See [ here ](/wiki/Template:Steroid_hormones "Template:Steroid hormones") instead.
---
[ Nucleobase ](/wiki/Nucleobase "Nucleobase") -derived |
| Nucleosides |
| [ Adenosine ](/wiki/Adenosine "Adenosine") system |
* ADP
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
---|---
[ Vitamin ](/wiki/Vitamin "Vitamin") -derived |
---|---
Miscellaneous |
| Cholinergic system |
* [ Acetylcholine ](/wiki/Acetylcholine "Acetylcholine")
---|---
Gasotransmitters |
* [ Carbon monoxide (CO) ](/wiki/Carbon_monoxide "Carbon monoxide")
* [ Hydrogen sulfide (H 2 S) ](/wiki/Hydrogen_sulfide "Hydrogen sulfide")
* [ Nitric oxide (NO) ](/wiki/Nitric_oxide "Nitric oxide")
---|---
Candidates |
* [ Acetaldehyde ](/wiki/Acetaldehyde "Acetaldehyde")
* [ Ammonia (NH 3 ) ](/wiki/Ammonia "Ammonia")
* [ Carbonyl sulfide (COS) ](/wiki/Carbonyl_sulfide "Carbonyl sulfide")
* [ Nitrous oxide (N 2 O) ](/wiki/Nitrous_oxide "Nitrous oxide")
* [ Sulfur dioxide (SO 2 ) ](/wiki/Sulfur_dioxide "Sulfur dioxide")
---|---
* [ v ](/wiki/Template:Purine_receptor_modulators "Template:Purine receptor modulators")
* [ t ](/wiki/Template_talk:Purine_receptor_modulators "Template talk:Purine receptor modulators")
* [ e ](/wiki/Special:EditPage/Template:Purine_receptor_modulators "Special:EditPage/Template:Purine receptor modulators")
[ Purine receptor ](/wiki/Purine_receptor "Purine receptor") [ modulators
](/wiki/Receptor_modulator "Receptor modulator")
---
[ Receptor ](/wiki/Receptor_\(biochemistry\) "Receptor \(biochemistry\)")
( [ ligands ](/wiki/Ligand_\(biochemistry\) "Ligand \(biochemistry\)") ) |
| [ P0 ](/wiki/P0_receptor "P0 receptor") ( [ adenine ](/wiki/Adenine
"Adenine") ) |
* **Agonists:** [ 8-Aminoadenine ](/w/index.php?title=8-Aminoadenine&action=edit&redlink=1 "8-Aminoadenine \(page does not exist\)")
* [ Adenine ](/wiki/Adenine "Adenine")
---|---
[ P1 ](/wiki/Adenosine_receptor "Adenosine receptor")
( [ adenosine ](/wiki/Adenosine "Adenosine") ) |
* **Agonists:** [ 2-(1-Hexynyl)- _N_ -methyladenosine ](/w/index.php?title=2-\(1-Hexynyl\)-N-methyladenosine&action=edit&redlink=1 "2-\(1-Hexynyl\)-N-methyladenosine \(page does not exist\)")
* [ 2-Cl-IB-MECA ](/w/index.php?title=2-Cl-IB-MECA&action=edit&redlink=1 "2-Cl-IB-MECA \(page does not exist\)")
* [ 2'-MeCCPA ](/w/index.php?title=2%27-MeCCPA&action=edit&redlink=1 "2'-MeCCPA \(page does not exist\)")
* [ 4'-O-β-D-Glucosyl-9-O-(6''-deoxysaccharosyl)olivil ](/w/index.php?title=4%27-O-%CE%B2-D-Glucosyl-9-O-\(6%27-deoxysaccharosyl\)olivil&action=edit&redlink=1 "4'-O-β-D-Glucosyl-9-O-\(6'-deoxysaccharosyl\)olivil \(page does not exist\)")
* [ 5'- _N_ -ethylcarboxamidoadenosine ](/w/index.php?title=5%27-N-ethylcarboxamidoadenosine&action=edit&redlink=1 "5'-N-ethylcarboxamidoadenosine \(page does not exist\)")
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* ADP
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* [ Apadenoson ](/w/index.php?title=Apadenoson&action=edit&redlink=1 "Apadenoson \(page does not exist\)")
* [ ATL-146e ](/wiki/ATL-146e "ATL-146e")
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
* [ BAY 60–6583 ](/wiki/BAY_60%E2%80%936583 "BAY 60–6583")
* [ Binodenoson ](/w/index.php?title=Binodenoson&action=edit&redlink=1 "Binodenoson \(page does not exist\)")
* [ Capadenoson ](/w/index.php?title=Capadenoson&action=edit&redlink=1 "Capadenoson \(page does not exist\)")
* [ CCPA ](/wiki/CCPA_\(biochemistry\) "CCPA \(biochemistry\)")
* [ CGS-21680 ](/wiki/CGS-21680 "CGS-21680")
* [ CP-532,903 ](/wiki/CP-532,903 "CP-532,903")
* [ Evodenoson ](/w/index.php?title=Evodenoson&action=edit&redlink=1 "Evodenoson \(page does not exist\)")
* [ GR 79236 ](/w/index.php?title=GR_79236&action=edit&redlink=1 "GR 79236 \(page does not exist\)")
* [ LUF-5835 ](/w/index.php?title=LUF-5835&action=edit&redlink=1 "LUF-5835 \(page does not exist\)")
* [ LUF-5845 ](/w/index.php?title=LUF-5845&action=edit&redlink=1 "LUF-5845 \(page does not exist\)")
* [ _N_ 6 -Cyclopentyladenosine ](/wiki/N6-Cyclopentyladenosine "N6-Cyclopentyladenosine")
* [ Namodenoson ](/wiki/Namodenoson "Namodenoson")
* [ Neladenoson dalanate ](/w/index.php?title=Neladenoson_dalanate&action=edit&redlink=1 "Neladenoson dalanate \(page does not exist\)")
* [ Piclidenoson ](/w/index.php?title=Piclidenoson&action=edit&redlink=1 "Piclidenoson \(page does not exist\)")
* [ Regadenoson ](/wiki/Regadenoson "Regadenoson")
* [ SDZ WAG 994 ](/w/index.php?title=SDZ_WAG_994&action=edit&redlink=1 "SDZ WAG 994 \(page does not exist\)")
* [ Selodenoson ](/w/index.php?title=Selodenoson&action=edit&redlink=1 "Selodenoson \(page does not exist\)")
* [ Sonedenoson ](/w/index.php?title=Sonedenoson&action=edit&redlink=1 "Sonedenoson \(page does not exist\)")
* [ Tecadenoson ](/w/index.php?title=Tecadenoson&action=edit&redlink=1 "Tecadenoson \(page does not exist\)")
* [ UK-432,097 ](/wiki/UK-432,097 "UK-432,097")
* **Antagonists:** [ 7-Methylxanthine ](/wiki/7-Methylxanthine "7-Methylxanthine")
* [ 8-Chlorotheophylline ](/wiki/8-Chlorotheophylline "8-Chlorotheophylline")
* [ 8-Phenyl-1,3-dipropylxanthine ](/w/index.php?title=8-Phenyl-1,3-dipropylxanthine&action=edit&redlink=1 "8-Phenyl-1,3-dipropylxanthine \(page does not exist\)")
* [ 8-Phenyltheophylline ](/wiki/8-Phenyltheophylline "8-Phenyltheophylline")
* [ Acefylline ](/wiki/Acefylline "Acefylline")
* [ Aminophylline ](/wiki/Aminophylline "Aminophylline")
* [ ATL-444 ](/wiki/ATL-444 "ATL-444")
* [ Bamifylline ](/wiki/Bamifylline "Bamifylline")
* [ Cafedrine ](/wiki/Cafedrine "Cafedrine")
* [ Caffeine ](/wiki/Caffeine "Caffeine")
* [ Caffeine citrate ](/wiki/Caffeine_citrate "Caffeine citrate")
* [ Cartazolate ](/wiki/Cartazolate "Cartazolate")
* [ CGH-2466 ](/w/index.php?title=CGH-2466&action=edit&redlink=1 "CGH-2466 \(page does not exist\)")
* [ CGS-15943 ](/wiki/CGS-15943 "CGS-15943")
* [ Choline theophyllinate ](/wiki/Choline_theophyllinate "Choline theophyllinate")
* [ Ciforadenant ](/w/index.php?title=Ciforadenant&action=edit&redlink=1 "Ciforadenant \(page does not exist\)")
* [ CPX ](/wiki/8-Cyclopentyl-1,3-dimethylxanthine "8-Cyclopentyl-1,3-dimethylxanthine")
* [ CVT-6883 ](/w/index.php?title=CVT-6883&action=edit&redlink=1 "CVT-6883 \(page does not exist\)")
* [ Dimethazan ](/wiki/Dimethazan "Dimethazan")
* [ DMPX ](/wiki/DMPX "DMPX")
* [ DPCPX ](/wiki/8-Cyclopentyl-1,3-dipropylxanthine "8-Cyclopentyl-1,3-dipropylxanthine")
* [ Dyphylline ](/wiki/Dyphylline "Dyphylline")
* [ Enprofylline ](/wiki/Enprofylline "Enprofylline")
* [ Etazolate ](/wiki/Etazolate "Etazolate")
* [ Fenethylline ](/wiki/Fenethylline "Fenethylline")
* [ IBMX ](/wiki/IBMX "IBMX")
* [ Isovaleric acid ](/wiki/Isovaleric_acid "Isovaleric acid")
* [ Istradefylline ](/wiki/Istradefylline "Istradefylline")
* [ KF-26777 ](/wiki/KF-26777 "KF-26777")
* [ MRE3008F20 ](/wiki/MRE3008F20 "MRE3008F20")
* [ MRS-1220 ](/w/index.php?title=MRS-1220&action=edit&redlink=1 "MRS-1220 \(page does not exist\)")
* [ MRS-1334 ](/w/index.php?title=MRS-1334&action=edit&redlink=1 "MRS-1334 \(page does not exist\)")
* [ MRS-1706 ](/wiki/MRS-1706 "MRS-1706")
* [ MRS-1754 ](/w/index.php?title=MRS-1754&action=edit&redlink=1 "MRS-1754 \(page does not exist\)")
* [ MRS-3777 ](/w/index.php?title=MRS-3777&action=edit&redlink=1 "MRS-3777 \(page does not exist\)")
* [ Paraxanthine ](/wiki/Paraxanthine "Paraxanthine")
* [ Pentoxifylline ](/wiki/Pentoxifylline "Pentoxifylline")
* [ Preladenant ](/wiki/Preladenant "Preladenant")
* [ Propentofylline ](/wiki/Propentofylline "Propentofylline")
* [ Proxyphylline ](/wiki/Proxyphylline "Proxyphylline")
* [ PSB-10 ](/wiki/PSB-10 "PSB-10")
* [ PSB-11 ](/w/index.php?title=PSB-11&action=edit&redlink=1 "PSB-11 \(page does not exist\)")
* [ PSB-36 ](/w/index.php?title=PSB-36&action=edit&redlink=1 "PSB-36 \(page does not exist\)")
* [ PSB-603 ](/w/index.php?title=PSB-603&action=edit&redlink=1 "PSB-603 \(page does not exist\)")
* [ PSB-788 ](/w/index.php?title=PSB-788&action=edit&redlink=1 "PSB-788 \(page does not exist\)")
* [ PSB-1115 ](/w/index.php?title=PSB-1115&action=edit&redlink=1 "PSB-1115 \(page does not exist\)")
* [ Reversine ](/wiki/Reversine "Reversine")
* [ Rolofylline ](/wiki/Rolofylline "Rolofylline")
* [ SCH-442,416 ](/wiki/SCH-442,416 "SCH-442,416")
* [ SCH-58261 ](/wiki/SCH-58261 "SCH-58261")
* [ Theacrine ](/wiki/Theacrine "Theacrine")
* [ Theobromine ](/wiki/Theobromine "Theobromine")
* [ Theodrenaline ](/wiki/Theodrenaline "Theodrenaline")
* [ Theophylline ](/wiki/Theophylline "Theophylline")
* [ Tozadenant ](/w/index.php?title=Tozadenant&action=edit&redlink=1 "Tozadenant \(page does not exist\)")
* [ Tracazolate ](/wiki/Tracazolate "Tracazolate")
* [ VUF-5574 ](/w/index.php?title=VUF-5574&action=edit&redlink=1 "VUF-5574 \(page does not exist\)")
* [ ZM-241,385 ](/wiki/ZM-241,385 "ZM-241,385")
[ P2 ](/wiki/P2_receptor "P2 receptor")
( [ nucleotide ](/wiki/Nucleotide "Nucleotide") ) |
| [ P2X ](/wiki/P2X_purinoreceptor "P2X purinoreceptor")
( [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate") Tooltip
Adenosine triphosphate ) |
* **Agonists:** [ 2-Me-SATP ](/w/index.php?title=2-Methylthio-ATP&action=edit&redlink=1 "2-Methylthio-ATP \(page does not exist\)")
* [ α,β-Me-ATP ](/w/index.php?title=%CE%91,%CE%B2-Methylene-ATP&action=edit&redlink=1 "Α,β-Methylene-ATP \(page does not exist\)")
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* ADP
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* [ Ap4A ](/wiki/Diadenosine_tetraphosphate "Diadenosine tetraphosphate")
* [ Ap5A ](/w/index.php?title=Diadenosine_pentaphosphate&action=edit&redlink=1 "Diadenosine pentaphosphate \(page does not exist\)")
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
* [ ATPγS ](/w/index.php?title=Adenosine-5%27-\(%CE%B3-thio\)-triphosphate&action=edit&redlink=1 "Adenosine-5'-\(γ-thio\)-triphosphate \(page does not exist\)")
* [ BzATP ](/w/index.php?title=BzATP&action=edit&redlink=1 "BzATP \(page does not exist\)")
* [ Cibacron blue ](/w/index.php?title=Cibacron_blue&action=edit&redlink=1 "Cibacron blue \(page does not exist\)")
* [ CTP ](/wiki/Cytidine_triphosphate "Cytidine triphosphate")
* [ D-β,γ-Me-ATP ](/w/index.php?title=D-%CE%B2,%CE%B3-Me-ATP&action=edit&redlink=1 "D-β,γ-Me-ATP \(page does not exist\)")
* [ GTP ](/wiki/Guanosine_triphosphate "Guanosine triphosphate")
* [ HT-AMP ](/w/index.php?title=HT-AMP&action=edit&redlink=1 "HT-AMP \(page does not exist\)")
* [ Ivermectin ](/wiki/Ivermectin "Ivermectin")
* [ L-β,γ-Me-ATP ](/w/index.php?title=L-%CE%B2,%CE%B3-Me-ATP&action=edit&redlink=1 "L-β,γ-Me-ATP \(page does not exist\)")
* [ MRS-2219 ](/w/index.php?title=MRS-2219&action=edit&redlink=1 "MRS-2219 \(page does not exist\)")
* [ PAPET-ATP ](/w/index.php?title=PAPET-ATP&action=edit&redlink=1 "PAPET-ATP \(page does not exist\)")
* [ UTP ](/wiki/Uridine_triphosphate "Uridine triphosphate")
* [ Zinc ](/wiki/Zinc "Zinc")
* **Antagonists:** [ 5-BDBD ](/w/index.php?title=5-BDBD&action=edit&redlink=1 "5-BDBD \(page does not exist\)")
* [ A-317491 ](/w/index.php?title=A-317491&action=edit&redlink=1 "A-317491 \(page does not exist\)")
* [ A-438079 ](/w/index.php?title=A-438079&action=edit&redlink=1 "A-438079 \(page does not exist\)")
* [ A-740003 ](/w/index.php?title=A-740003&action=edit&redlink=1 "A-740003 \(page does not exist\)")
* [ A-804598 ](/w/index.php?title=A-804598&action=edit&redlink=1 "A-804598 \(page does not exist\)")
* [ A-839977 ](/w/index.php?title=A-839977&action=edit&redlink=1 "A-839977 \(page does not exist\)")
* [ AF-353 ](/w/index.php?title=AF-353&action=edit&redlink=1 "AF-353 \(page does not exist\)")
* [ AZ-10606120 ](/w/index.php?title=AZ-10606120&action=edit&redlink=1 "AZ-10606120 \(page does not exist\)")
* [ AZ-11645373 ](/w/index.php?title=AZ-11645373&action=edit&redlink=1 "AZ-11645373 \(page does not exist\)")
* [ BBG ](/wiki/Brilliant_Blue_G "Brilliant Blue G")
* [ Calcium ](/wiki/Calcium "Calcium")
* [ Calmidazolium ](/w/index.php?title=Calmidazolium&action=edit&redlink=1 "Calmidazolium \(page does not exist\)")
* [ Chelerythrine ](/wiki/Chelerythrine "Chelerythrine")
* [ Copper ](/wiki/Copper "Copper")
* [ Emodin ](/wiki/Emodin "Emodin") ( _[ Rheum officinale ](/wiki/Rheum_officinale "Rheum officinale") _ )
* [ Evans blue ](/wiki/Evans_blue_\(dye\) "Evans blue \(dye\)")
* [ Gefapixant ](/wiki/Gefapixant "Gefapixant")
* [ GW-791343 ](/w/index.php?title=GW-791343&action=edit&redlink=1 "GW-791343 \(page does not exist\)")
* [ HMA ](/w/index.php?title=5-\(N,N-Hexamethylene\)amiloride&action=edit&redlink=1 "5-\(N,N-Hexamethylene\)amiloride \(page does not exist\)")
* [ Ip5I ](/w/index.php?title=Diinosine_pentaphosphate&action=edit&redlink=1 "Diinosine pentaphosphate \(page does not exist\)")
* [ isoPPADS ](/w/index.php?title=IsoPPADS&action=edit&redlink=1 "IsoPPADS \(page does not exist\)")
* [ JNJ-47965567 ](/w/index.php?title=JNJ-47965567&action=edit&redlink=1 "JNJ-47965567 \(page does not exist\)")
* [ KN-04 ](/w/index.php?title=KN-04&action=edit&redlink=1 "KN-04 \(page does not exist\)")
* [ KN-62 ](/wiki/KN-62 "KN-62")
* [ Magnesium ](/wiki/Magnesium "Magnesium")
* [ MRS-2159 ](/w/index.php?title=MRS-2159&action=edit&redlink=1 "MRS-2159 \(page does not exist\)")
* [ NF-023 ](/w/index.php?title=NF-023&action=edit&redlink=1 "NF-023 \(page does not exist\)")
* [ NF-110 ](/w/index.php?title=NF-110&action=edit&redlink=1 "NF-110 \(page does not exist\)")
* [ NF-157 ](/w/index.php?title=NF-157&action=edit&redlink=1 "NF-157 \(page does not exist\)")
* [ NF-279 ](/w/index.php?title=NF-279&action=edit&redlink=1 "NF-279 \(page does not exist\)")
* [ NF-449 ](/w/index.php?title=NF-449&action=edit&redlink=1 "NF-449 \(page does not exist\)")
* [ Opiranserin (VVZ-149) ](/wiki/Opiranserin "Opiranserin")
* [ Oxidized-ATP ](/w/index.php?title=Oxidized-ATP&action=edit&redlink=1 "Oxidized-ATP \(page does not exist\)")
* [ Phenol Red ](/wiki/Phenol_Red "Phenol Red")
* [ Phenolphthalein ](/wiki/Phenolphthalein "Phenolphthalein")
* [ PPADS ](/wiki/PPADS "PPADS")
* [ PPNDS ](/w/index.php?title=PPNDS&action=edit&redlink=1 "PPNDS \(page does not exist\)")
* [ PSB-12062 ](/w/index.php?title=PSB-12062&action=edit&redlink=1 "PSB-12062 \(page does not exist\)")
* [ Puerarin ](/wiki/Puerarin "Puerarin") ( _[ Radix puerariae ](/wiki/Radix_puerariae "Radix puerariae") _ )
* [ Purotoxin 1 ](/w/index.php?title=Purotoxin_1&action=edit&redlink=1 "Purotoxin 1 \(page does not exist\)")
* [ RB-2 ](/w/index.php?title=Reactive_Blue_2&action=edit&redlink=1 "Reactive Blue 2 \(page does not exist\)")
* [ Ro 0437626 ](/w/index.php?title=Ro_0437626&action=edit&redlink=1 "Ro 0437626 \(page does not exist\)")
* [ Ro 51 ](/w/index.php?title=Ro_51&action=edit&redlink=1 "Ro 51 \(page does not exist\)")
* [ RO-3 ](/w/index.php?title=RO-3&action=edit&redlink=1 "RO-3 \(page does not exist\)")
* [ Sodium ferulate ](/wiki/Sodium_ferulate "Sodium ferulate") ( _[ Angelica sinensis ](/wiki/Angelica_sinensis "Angelica sinensis") _ , _[ Ligusticum wallichii ](/wiki/Ligusticum_wallichii "Ligusticum wallichii") _ )
* [ Suramin ](/wiki/Suramin "Suramin")
* [ TC-P 262 ](/w/index.php?title=TC-P_262&action=edit&redlink=1 "TC-P 262 \(page does not exist\)")
* [ Tetramethylpyrazine (ligustrazine) ](/wiki/Tetramethylpyrazine "Tetramethylpyrazine") ( _[ Ligusticum wallichii ](/wiki/Ligusticum_wallichii "Ligusticum wallichii") _ )
* [ TNP-ATP ](/wiki/TNP-ATP "TNP-ATP")
* [ Zinc ](/wiki/Zinc "Zinc")
---|---
[ P2Y ](/wiki/P2Y_receptor "P2Y receptor") |
* **Agonists:** [ 2-Me-SADP ](/w/index.php?title=2-Methylthioladenosine_diphosphate&action=edit&redlink=1 "2-Methylthioladenosine diphosphate \(page does not exist\)")
* [ 2-Me-SATP ](/w/index.php?title=2-Methylthioadenosine_triphosphate&action=edit&redlink=1 "2-Methylthioadenosine triphosphate \(page does not exist\)")
* [ 2-Thio-UTP ](/w/index.php?title=2-Thio-UTP&action=edit&redlink=1 "2-Thio-UTP \(page does not exist\)")
* [ 5-Br-UDP ](/w/index.php?title=5-Bromouridine_5%E2%80%B2-diphosphate&action=edit&redlink=1 "5-Bromouridine 5′-diphosphate \(page does not exist\)")
* [ 5-OMe-UDP ](/w/index.php?title=5-OMe-UDP&action=edit&redlink=1 "5-OMe-UDP \(page does not exist\)")
* [ α,β-Me-ATP ](/w/index.php?title=%CE%91,%CE%B2-Methylene-ATP&action=edit&redlink=1 "Α,β-Methylene-ATP \(page does not exist\)")
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* ADP
* [ ADPβS ](/w/index.php?title=Adenosine-5%27-\(%CE%B2-thio\)-diphosphate&action=edit&redlink=1 "Adenosine-5'-\(β-thio\)-diphosphate \(page does not exist\)")
* [ Ap3A ](/w/index.php?title=Diadenosine_triphosphate&action=edit&redlink=1 "Diadenosine triphosphate \(page does not exist\)")
* [ AR-C 67085MX ](/w/index.php?title=AR-C_67085MX&action=edit&redlink=1 "AR-C 67085MX \(page does not exist\)")
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
* [ ATPγS ](/w/index.php?title=Adenosine-5%27-\(%CE%B3-thio\)-triphosphate&action=edit&redlink=1 "Adenosine-5'-\(γ-thio\)-triphosphate \(page does not exist\)")
* [ CTP ](/wiki/Cytidine_triphosphate "Cytidine triphosphate")
* [ dATP ](/wiki/Deoxyadenosine_triphosphate "Deoxyadenosine triphosphate")
* [ Denufosol ](/wiki/Denufosol "Denufosol")
* [ Diquafosol ](/wiki/Diquafosol "Diquafosol")
* [ IDP ](/w/index.php?title=Inosine_diphosphate&action=edit&redlink=1 "Inosine diphosphate \(page does not exist\)")
* [ ITP ](/wiki/Inosine_triphosphate "Inosine triphosphate")
* [ INS-365 ](/wiki/INS-365 "INS-365")
* [ INS-37217 ](/w/index.php?title=INS-37217&action=edit&redlink=1 "INS-37217 \(page does not exist\)")
* [ MRS-2365 ](/w/index.php?title=MRS-2365&action=edit&redlink=1 "MRS-2365 \(page does not exist\)")
* [ MRS-2690 ](/w/index.php?title=MRS-2690&action=edit&redlink=1 "MRS-2690 \(page does not exist\)")
* [ MRS-2693 ](/w/index.php?title=MRS-2693&action=edit&redlink=1 "MRS-2693 \(page does not exist\)")
* [ MRS-2768 ](/w/index.php?title=MRS-2768&action=edit&redlink=1 "MRS-2768 \(page does not exist\)")
* [ MRS-2957 ](/w/index.php?title=MRS-2957&action=edit&redlink=1 "MRS-2957 \(page does not exist\)")
* [ MRS-4062 ](/w/index.php?title=MRS-4062&action=edit&redlink=1 "MRS-4062 \(page does not exist\)")
* [ NF-546 ](/w/index.php?title=NF-546&action=edit&redlink=1 "NF-546 \(page does not exist\)")
* [ PAPET-ATP ](/w/index.php?title=PAPET-ATP&action=edit&redlink=1 "PAPET-ATP \(page does not exist\)")
* [ PSB-0474 ](/w/index.php?title=PSB-0474&action=edit&redlink=1 "PSB-0474 \(page does not exist\)")
* [ PSB-1114 ](/w/index.php?title=PSB-1114&action=edit&redlink=1 "PSB-1114 \(page does not exist\)")
* [ UDP ](/wiki/Uridine_diphosphate "Uridine diphosphate")
* [ UDPβS ](/w/index.php?title=Uridine_5%C2%B4-\(%CE%B2-thio\)-diphosphate&action=edit&redlink=1 "Uridine 5´-\(β-thio\)-diphosphate \(page does not exist\)")
* [ UDP-galactose ](/wiki/Uridine_diphosphate_galactose "Uridine diphosphate galactose")
* [ UDP-glucose ](/wiki/Uridine_diphosphate_glucose "Uridine diphosphate glucose")
* [ UDP-N-acetylglucosamine ](/wiki/Uridine_diphosphate_N-acetylglucosamine "Uridine diphosphate N-acetylglucosamine")
* [ Up3U ](/w/index.php?title=Diuridine_triphosphate&action=edit&redlink=1 "Diuridine triphosphate \(page does not exist\)")
* [ UTP ](/wiki/Uridine_triphosphate "Uridine triphosphate")
* [ UTPγS ](/w/index.php?title=Uridine-5%27-\(%CE%B3-thio\)-triphosphate&action=edit&redlink=1 "Uridine-5'-\(γ-thio\)-triphosphate \(page does not exist\)")
* **Antagonists:** [ 2-Me-SAMP ](/w/index.php?title=2-Methylthioladenosine_monophosphate&action=edit&redlink=1 "2-Methylthioladenosine monophosphate \(page does not exist\)")
* [ A3P5PS ](/w/index.php?title=A3P5PS&action=edit&redlink=1 "A3P5PS \(page does not exist\)")
* [ AMPαS ](/w/index.php?title=AMP%CE%B1S&action=edit&redlink=1 "AMPαS \(page does not exist\)")
* [ Ap4A ](/wiki/Diadenosine_tetraphosphate "Diadenosine tetraphosphate")
* [ AR-C 66096 ](/w/index.php?title=AR-C_66096&action=edit&redlink=1 "AR-C 66096 \(page does not exist\)")
* [ AR-C 67085MX ](/w/index.php?title=AR-C_67085MX&action=edit&redlink=1 "AR-C 67085MX \(page does not exist\)")
* [ AR-C 69931MX ](/w/index.php?title=AR-C_69931MX&action=edit&redlink=1 "AR-C 69931MX \(page does not exist\)")
* [ AR-C 118925XX ](/w/index.php?title=AR-C_118925XX&action=edit&redlink=1 "AR-C 118925XX \(page does not exist\)")
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
* [ BzATP ](/w/index.php?title=BzATP&action=edit&redlink=1 "BzATP \(page does not exist\)")
* [ C1330-7 ](/w/index.php?title=C1330-7&action=edit&redlink=1 "C1330-7 \(page does not exist\)")
* [ Cangrelor ](/wiki/Cangrelor "Cangrelor")
* [ Clopidogrel ](/wiki/Clopidogrel "Clopidogrel")
* [ Elinogrel ](/wiki/Elinogrel "Elinogrel")
* [ Ip5I ](/w/index.php?title=Diinosine_pentaphosphate&action=edit&redlink=1 "Diinosine pentaphosphate \(page does not exist\)")
* [ MRS-2179 ](/w/index.php?title=MRS-2179&action=edit&redlink=1 "MRS-2179 \(page does not exist\)")
* [ MRS-2211 ](/w/index.php?title=MRS-2211&action=edit&redlink=1 "MRS-2211 \(page does not exist\)")
* [ MRS-2279 ](/w/index.php?title=MRS-2279&action=edit&redlink=1 "MRS-2279 \(page does not exist\)")
* [ MRS-2395 ](/w/index.php?title=MRS-2395&action=edit&redlink=1 "MRS-2395 \(page does not exist\)")
* [ MRS-2500 ](/w/index.php?title=MRS-2500&action=edit&redlink=1 "MRS-2500 \(page does not exist\)")
* [ MRS-2578 ](/w/index.php?title=MRS-2578&action=edit&redlink=1 "MRS-2578 \(page does not exist\)")
* [ NF-157 ](/w/index.php?title=NF-157&action=edit&redlink=1 "NF-157 \(page does not exist\)")
* [ NF-340 ](/w/index.php?title=NF-340&action=edit&redlink=1 "NF-340 \(page does not exist\)")
* [ PIT ](/w/index.php?title=2,2%27-Pyridylisatogen_tosylate&action=edit&redlink=1 "2,2'-Pyridylisatogen tosylate \(page does not exist\)")
* [ PPADS ](/wiki/PPADS "PPADS")
* [ Prasugrel ](/wiki/Prasugrel "Prasugrel")
* [ PSB-0739 ](/w/index.php?title=PSB-0739&action=edit&redlink=1 "PSB-0739 \(page does not exist\)")
* [ RB-2 ](/w/index.php?title=Reactive_Blue_2&action=edit&redlink=1 "Reactive Blue 2 \(page does not exist\)")
* [ Regrelor ](/wiki/Regrelor "Regrelor")
* [ Suramin ](/wiki/Suramin "Suramin")
* [ Ticagrelor ](/wiki/Ticagrelor "Ticagrelor")
* [ Ticlopidine ](/wiki/Ticlopidine "Ticlopidine")
* [ UDP ](/wiki/Uridine_diphosphate "Uridine diphosphate")
[ Transporter ](/wiki/Membrane_transport_protein "Membrane transport protein")
( [ blockers ](/wiki/Reuptake_inhibitor "Reuptake inhibitor") ) |
| [ CNTs ](/wiki/Concentrative_nucleoside_transporters "Concentrative
nucleoside transporters") Tooltip Concentrative nucleoside transporters |
* [ 6-Hydroxy-7-methoxyflavone ](/w/index.php?title=6-Hydroxy-7-methoxyflavone&action=edit&redlink=1 "6-Hydroxy-7-methoxyflavone \(page does not exist\)")
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* [ dMeThPmR ](/w/index.php?title=DMeThPmR&action=edit&redlink=1 "DMeThPmR \(page does not exist\)")
* [ Estradiol ](/wiki/Estradiol "Estradiol")
* [ KGO-2142 ](/w/index.php?title=KGO-2142&action=edit&redlink=1 "KGO-2142 \(page does not exist\)")
* [ KGO-2173 ](/w/index.php?title=KGO-2173&action=edit&redlink=1 "KGO-2173 \(page does not exist\)")
* [ MeThPmR ](/w/index.php?title=MeThPmR&action=edit&redlink=1 "MeThPmR \(page does not exist\)")
* [ Phloridzin ](/wiki/Phloridzin "Phloridzin")
* [ Progesterone ](/wiki/Progesterone "Progesterone")
---|---
[ ENTs ](/wiki/Equilibrative_nucleoside_transporters "Equilibrative
nucleoside transporters") Tooltip Equilibrative nucleoside transporters |
* [ Barbiturates ](/wiki/Barbiturate "Barbiturate")
* [ Benzodiazepines ](/wiki/Benzodiazepine "Benzodiazepine")
* [ Cilostazol ](/wiki/Cilostazol "Cilostazol")
* [ Dilazep ](/wiki/Dilazep "Dilazep")
* [ Dipyridamole ](/wiki/Dipyridamole "Dipyridamole")
* [ Estradiol ](/wiki/Estradiol "Estradiol")
* [ Ethanol ](/wiki/Ethanol "Ethanol")
* [ Hexobendine ](/wiki/Hexobendine "Hexobendine")
* [ NBMPR ](/w/index.php?title=6-S-\(\(4-Nitrophenyl\)methyl\)-6-thioinosine&action=edit&redlink=1 "6-S-\(\(4-Nitrophenyl\)methyl\)-6-thioinosine \(page does not exist\)")
* [ Pentoxifylline ](/wiki/Pentoxifylline "Pentoxifylline")
* [ Progesterone ](/wiki/Progesterone "Progesterone")
* [ Propentofylline ](/wiki/Propentofylline "Propentofylline")
[ PMAT ](/wiki/Plasma_membrane_monoamine_transporter "Plasma membrane
monoamine transporter") Tooltip Plasma membrane monoamine transporter |
* [ Decynium-22 ](/wiki/Decynium-22 "Decynium-22")
[ Enzyme ](/wiki/Enzyme "Enzyme")
( [ inhibitors ](/wiki/Enzyme_inhibitor "Enzyme inhibitor") ) |
| [ XO ](/wiki/Xanthine_oxidase "Xanthine oxidase") Tooltip Xanthine oxidase
|
* [ Allopurinol ](/wiki/Allopurinol "Allopurinol")
* [ Amflutizole ](/wiki/Amflutizole "Amflutizole")
* [ Benzbromarone ](/wiki/Benzbromarone "Benzbromarone")
* [ Caffeic acid ](/wiki/Caffeic_acid "Caffeic acid")
* [ Cinnamaldehyde ](/wiki/Cinnamaldehyde "Cinnamaldehyde")
* _[ Cinnamomum osmophloeum ](/wiki/Cinnamomum_osmophloeum "Cinnamomum osmophloeum") _
* [ Febuxostat ](/wiki/Febuxostat "Febuxostat")
* [ Myo-inositol ](/wiki/Myo-inositol "Myo-inositol")
* [ Kaempferol ](/wiki/Kaempferol "Kaempferol")
* [ Myricetin ](/wiki/Myricetin "Myricetin")
* [ Niraxostat ](/w/index.php?title=Niraxostat&action=edit&redlink=1 "Niraxostat \(page does not exist\)")
* [ Oxipurinol ](/wiki/Oxipurinol "Oxipurinol")
* [ Phytic acid ](/wiki/Phytic_acid "Phytic acid")
* _[ Pistacia integerrima ](/wiki/Pistacia_integerrima "Pistacia integerrima") _
* [ Propolis ](/wiki/Propolis "Propolis")
* [ Quercetin ](/wiki/Quercetin "Quercetin")
* [ Tisopurine ](/wiki/Tisopurine "Tisopurine")
* [ Topiroxostat ](/wiki/Topiroxostat "Topiroxostat")
---|---
Others |
* [ Aminopterin ](/wiki/Aminopterin "Aminopterin")
* [ Azathioprine ](/wiki/Azathioprine "Azathioprine")
* [ Methotrexate ](/wiki/Methotrexate "Methotrexate")
* [ Mycophenolic acid ](/wiki/Mycophenolic_acid "Mycophenolic acid")
* [ Pemetrexed ](/wiki/Pemetrexed "Pemetrexed")
* [ Pralatrexate ](/wiki/Pralatrexate "Pralatrexate")
* _Many others_
Others |
* **Precursors:** [ Adenine ](/wiki/Adenine "Adenine")
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* ADP
* [ ATP ](/wiki/Adenosine_triphosphate "Adenosine triphosphate")
* [ Cytosine ](/wiki/Cytosine "Cytosine")
* [ Cytidine ](/wiki/Cytidine "Cytidine")
* [ CMP ](/wiki/Cytidine_monophosphate "Cytidine monophosphate")
* [ CDP ](/wiki/Cytidine_diphosphate "Cytidine diphosphate")
* [ CTP ](/wiki/Cytidine_triphosphate "Cytidine triphosphate")
* [ Guanine ](/wiki/Guanine "Guanine")
* [ Guanosine ](/wiki/Guanosine "Guanosine")
* [ GMP ](/wiki/Guanosine_monophosphate "Guanosine monophosphate")
* [ GDP ](/wiki/Guanosine_diphosphate "Guanosine diphosphate")
* [ GTP ](/wiki/Guanosine_triphosphate "Guanosine triphosphate")
* [ Hypoxanthine ](/wiki/Hypoxanthine "Hypoxanthine")
* [ Inosine ](/wiki/Inosine "Inosine")
* [ IMP ](/wiki/Inosine_monophosphate "Inosine monophosphate")
* [ IDP ](/w/index.php?title=Inosine_diphosphate&action=edit&redlink=1 "Inosine diphosphate \(page does not exist\)")
* [ ITP ](/wiki/Inosine_triphosphate "Inosine triphosphate")
* [ Ribose ](/wiki/Ribose "Ribose")
* [ Uracil ](/wiki/Uracil "Uracil")
* [ Uridine ](/wiki/Uridine "Uridine")
* [ UMP ](/wiki/Uridine_monophosphate "Uridine monophosphate")
* [ UDP ](/wiki/Uridine_diphosphate "Uridine diphosphate")
* [ UTP ](/wiki/Uridine_triphosphate "Uridine triphosphate")
* **Others:** [ Chrysophanol ](/wiki/Chrysophanol "Chrysophanol") ( [ rhubarb ](/wiki/Rhubarb "Rhubarb") )
_**See also:** [ Receptor/signaling modulators
](/wiki/Template:Receptor_modulators "Template:Receptor modulators") _
[ Authority control databases ](/wiki/Help:Authority_control "Help:Authority
control") : National [ 
](https://www.wikidata.org/wiki/Q185253#identifiers "Edit this at Wikidata") |
* [ Japan ](https://id.ndl.go.jp/auth/ndlna/00575814)
---|---

Retrieved from " [
https://en.wikipedia.org/w/index.php?title=Adenosine_diphosphate&oldid=1176961502
](https://en.wikipedia.org/w/index.php?title=Adenosine_diphosphate&oldid=1176961502)
"
[ Categories ](/wiki/Help:Category "Help:Category") :
* [ Adenosine receptor agonists ](/wiki/Category:Adenosine_receptor_agonists "Category:Adenosine receptor agonists")
* [ Neurotransmitters ](/wiki/Category:Neurotransmitters "Category:Neurotransmitters")
* [ Nucleotides ](/wiki/Category:Nucleotides "Category:Nucleotides")
* [ Cellular respiration ](/wiki/Category:Cellular_respiration "Category:Cellular respiration")
* [ Phosphate esters ](/wiki/Category:Phosphate_esters "Category:Phosphate esters")
* [ Purines ](/wiki/Category:Purines "Category:Purines")
* [ Purinergic signalling ](/wiki/Category:Purinergic_signalling "Category:Purinergic signalling")
* [ Pyrophosphates ](/wiki/Category:Pyrophosphates "Category:Pyrophosphates")
Hidden categories:
* [ Chemical articles with multiple compound IDs ](/wiki/Category:Chemical_articles_with_multiple_compound_IDs "Category:Chemical articles with multiple compound IDs")
* [ Multiple chemicals in an infobox that need indexing ](/wiki/Category:Multiple_chemicals_in_an_infobox_that_need_indexing "Category:Multiple chemicals in an infobox that need indexing")
* [ Articles with changed DrugBank identifier ](/wiki/Category:Articles_with_changed_DrugBank_identifier "Category:Articles with changed DrugBank identifier")
* [ ECHA InfoCard ID from Wikidata ](/wiki/Category:ECHA_InfoCard_ID_from_Wikidata "Category:ECHA InfoCard ID from Wikidata")
* [ Articles with changed KEGG identifier ](/wiki/Category:Articles_with_changed_KEGG_identifier "Category:Articles with changed KEGG identifier")
* [ Articles containing unverified chemical infoboxes ](/wiki/Category:Articles_containing_unverified_chemical_infoboxes "Category:Articles containing unverified chemical infoboxes")
* [ Chembox image size set ](/wiki/Category:Chembox_image_size_set "Category:Chembox image size set")
* [ Articles with short description ](/wiki/Category:Articles_with_short_description "Category:Articles with short description")
* [ Short description matches Wikidata ](/wiki/Category:Short_description_matches_Wikidata "Category:Short description matches Wikidata")
* [ All articles with unsourced statements ](/wiki/Category:All_articles_with_unsourced_statements "Category:All articles with unsourced statements")
* [ Articles with unsourced statements from April 2023 ](/wiki/Category:Articles_with_unsourced_statements_from_April_2023 "Category:Articles with unsourced statements from April 2023")
* [ Articles with NDL identifiers ](/wiki/Category:Articles_with_NDL_identifiers "Category:Articles with NDL identifiers")
* This page was last edited on 25 September 2023, at 01:54 (UTC) .
* Text is available under the [ Creative Commons Attribution-ShareAlike License 4.0 ](//en.wikipedia.org/wiki/Wikipedia:Text_of_the_Creative_Commons_Attribution-ShareAlike_4.0_International_License) [ ](//en.wikipedia.org/wiki/Wikipedia:Text_of_the_Creative_Commons_Attribution-ShareAlike_4.0_International_License) ; additional terms may apply. By using this site, you agree to the [ Terms of Use ](//foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Terms_of_Use) and [ Privacy Policy ](//foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Privacy_policy) . Wikipedia® is a registered trademark of the [ Wikimedia Foundation, Inc. ](//www.wikimediafoundation.org/) , a non-profit organization.
* [ Privacy policy ](https://foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Privacy_policy)
* [ About Wikipedia ](/wiki/Wikipedia:About)
* [ Disclaimers ](/wiki/Wikipedia:General_disclaimer)
* [ Contact Wikipedia ](//en.wikipedia.org/wiki/Wikipedia:Contact_us)
* [ Code of Conduct ](https://foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Universal_Code_of_Conduct)
* [ Developers ](https://developer.wikimedia.org)
* [ Statistics ](https://stats.wikimedia.org/#/en.wikipedia.org)
* [ Cookie statement ](https://foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Cookie_statement)
* [ Mobile view ](//en.m.wikipedia.org/w/index.php?title=Adenosine_diphosphate&mobileaction=toggle_view_mobile)
* [  ](https://wikimediafoundation.org/)
* [  ](https://www.mediawiki.org/)
* Toggle limited content width
*[
CID
]: Compound ID
*[
EPA
]: U.S. Environmental Protection Agency
*[
v
]: View this template
*[
t
]: Discuss this template
*[
e
]: Edit this template
*[
ATP
]: Adenosine triphosphate
*[
CNTs
]: Concentrative nucleoside transporters
*[
ENTs
]: Equilibrative nucleoside transporters
*[
PMAT
]: Plasma membrane monoamine transporter
*[
XO
]: Xanthine oxidase
| biology | 1622593 | https://sv.wikipedia.org/wiki/TFIIH | TFIIH | TFIIH är en basal transkriptionsfaktor vars roll är att bidra med helikas- och kinasaktivitet. TFIIH består av flera underenheter. Helikasaktiviteten särar på DNAsträngarna så att en transkriptionsbubbla bildas, detta görs av subenheterna XPB och XPD. Kinaset, som består av ett komplex av Cdk7/cyklin H, hyperfosforylerar den C-terminala domänen av RNA-polymeras II vilket startar transkriptionen. TFIIH kom först i dagen 1989 när generell transkriptionsfaktor-δ eller grundläggande transkriptionsfaktor 2 karakteriserades som en oumbärlig transkriptionsfaktor in vitro. Denna faktor isolerades 1992 också från jäst och gavs slutligen namnet TFIIH.
TFIIH består av tio underenheter, varav 7 (ERCC2/XPD, ERCC3 /XPB, GTF2H1/p62, GTF2H4/p52, GTF2H2/p44, GTF2H3/p34 ochGTF2H5/TTDA) utgör kärnkomplexet. Det cyklinaktiverande kinasunderkomplexet (CDK7, MAT1 och cyklin H) är kopplat till kärnan via XPD-proteinet. Två av underenheterna, ERCC2/XPD och ERCC3/XPB, har helikas- och ATPas-aktiviteter och hjälper till att skapa transkriptionsbubblan. I ett provrör krävs dessa underenheter endast för transkription om DNA-mallen inte redan är denaturerad eller om den är superlindad.
Två andra TFIIH-underenheter, CDK7 och cyklin H, fosforylat serinaminosyror på RNA-polymeras II C-terminaldomänen och möjligen andra proteiner medverkar i cellcykeln. Förutom en vital funktion vid transkriptionsinitiering är TFIIH också involverad i nukleotidexcisionsreparation.
Struktur hos TFIIH
TFIIH är ett tiounderenhetskomplex. Sju av dessa underenheter utgör "kärnan" medan tre innefattar den dissocierbara "CAK"-modulen (CDK Activating Kinase). Kärnan består av underenheter XPB, XPD, p62, p52, s. 44, p34 och p8 medan CAK är sammansatt av CDK7, cyklin H och MAT1.
Funktioner
Allmän funktion för TFIIH är
Initiering av transkription av proteinkodande gen.
DNA-nukleotidreparation.
(NER) TFIIH är en allmän transkriptionsfaktor som verkar för att rekrytera RNA Pol II till promotorerna av gener. Den fungerar som en helikas som lindar upp DNA. Den lindar också upp DNA efter att en DNA-skada har identifierats av antingen den globala genomreparationsvägen (GGR) eller den transkriptionskopplade reparationsvägen (TCR) av NER. Renad TFIIH har en roll i att stoppa ytterligare RNA-syntes genom att aktivera den cykliska peptiden α-amanitin.
Trikotiodystrofi
Mutation i generna ERCC3 (XPB), ERCC2 (XPD) eller GTF2H5 (TTDA) orsakar trikotiodystrofi, ett tillstånd som kännetecknas av ljuskänslighet, iktyos, skört hår och naglar, intellektuell funktionsnedsättning, minskad fertilitet och/eller kortväxthet.
Sjukdom
Genetiska polymorfismer av gener som kodar för subenheter av TFIIH är kända för att vara förenade med ökad cancerkänslighet i många vävnader, till exempel hudvävnad, bröstvävnad och lungvävnad. Mutationer i underenheterna (som XPD och XPB) kan leda till en mängd olika sjukdomar, såsom xeroderma pigmentosum (XP) eller XP kombinerat med Cockayne syndrom. Förutom genetiska variationer riktar sig viruskodade proteiner även mot TFIIH.
DNA-reparation
TFIIH deltar i nukleotidexcisionsreparation (NER) genom att öppna DNA-dubbelhelixen efter att skadan initialt har upptäckts. NER är en flerstegsväg som tar bort en lång rad olika skador som förvränger normal basparning, inklusive skrymmande kemiska skador och UV-orsakade skador. Individer med mutationsdefekter i gener som specificerar proteinkomponenter som katalyserar NER-vägen, inklusive TFIIH-komponenterna, visar ofta drag av för tidigt åldrande.
Inhibitorer
Potenta, bioaktiva naturliga produkter som triptolid som hämmar transkription hos däggdjur via hämning av XPB-subenheten av den allmänna transkriptionsfaktorn TFIIH har nyligen (2020) rapporterats som ett glukoskonjugat för att rikta in sig på hypoxiska cancerceller med ökat glukostransportöruttryck.
Referenser
Noter
Externa länkar
Enzymer
Genuttryck
Transkription (genetik) | swedish | 0.930366 |
induce_sleep/25063AATPAdenosineTr.txt | Skip to main content
Table of Contents menu
[ 
](https://bio.libretexts.org/)
search Search build_circle Toolbar fact_check Homework cancel Exit
Reader Mode
* [ school Campus Bookshelves ](https://bio.libretexts.org/Courses)
* [ menu_book Bookshelves ](https://bio.libretexts.org/Bookshelves)
* [ perm_media Learning Objects ](https://bio.libretexts.org/Learning_Objects)
* [ login Login ](https://bio.libretexts.org/Special:Userlogin?readerView#tab=login)
* [ how_to_reg Request Instructor Account ](https://register.libretexts.org/)
* [ hub Instructor Commons ](https://commons.libretexts.org/)
## Search
Search this book
Submit Search
* [ Downloads expand_more ](https://bio.libretexts.org/?downloads)
* [ Download Page (PDF) ](https://bio.libretexts.org/?downloadpage)
* [ Download Full Book (PDF) ](https://bio.libretexts.org/?downloadfull)
* [ Resources expand_more ](https://bio.libretexts.org/?resources)
* [ Periodic Table ](https://bio.libretexts.org/?pertable)
* [ Physics Constants ](https://bio.libretexts.org/?physconst)
* [ Scientific Calculator ](https://bio.libretexts.org/?scientificcal)
* [ Reference expand_more ](https://bio.libretexts.org/?references)
* [ Reference & Cite ](https://bio.libretexts.org/?refmodal)
* [ Tools expand_more ](https://bio.libretexts.org/?tools)
* [ Help expand_more ](https://bio.libretexts.org/?help)
* [ Get Help ](https://bio.libretexts.org/?helpmodal)
* [ Feedback ](https://bio.libretexts.org/?feedback)
* [ Readability ](https://bio.libretexts.org/?readability)
##
x
selected template will load here
## Error
This action is not available.
[ 
](https://bio.libretexts.org/)
chrome_reader_mode Enter Reader Mode
2.5: Organic Compounds
2: Chemistry
{ }
{ "2.5.01:_Carbohydrate_Molecules" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.5.02:_Lipid_Molecules" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.5.03:_DNA_and_RNA" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.5.04:_Amino_Acids" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.5.05:_Types_and_Functions_of_Protiens" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.5.06:_ATP-_Adenosine_Triphosphate" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()"
}
{ "2.01:_Atomic_Structure" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.02:_Chemical_Bonds" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.03:_Chemical_Reactions" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.04:_Inorganic_Compounds" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.05:_Organic_Compounds" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.06:_Energy" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
"2.07:_Enzymes" : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()"
}
Sat, 24 Dec 2022 23:20:31 GMT
2.5.6: ATP- Adenosine Triphosphate
8804
8804
Delmar Larsen
{ }
Anonymous
Anonymous
2
false
false
[ "article:topic", "authorname:boundless", "showtoc:no", "license:ccbysa",
"licenseversion:40" ]
[ "article:topic", "authorname:boundless", "showtoc:no", "license:ccbysa",
"licenseversion:40" ]
https://bio.libretexts.org/@app/auth/3/login?returnto=https%3A%2F%2Fbio.libretexts.org%2FBookshelves%2FMicrobiology%2FMicrobiology_(Boundless)%2F02%253A_Chemistry%2F2.05%253A_Organic_Compounds%2F2.5.06%253A_ATP-
_Adenosine_Triphosphate
[ 
](https://bio.libretexts.org "Biology LibreTexts")
1. Search site
Search Search
Go back to previous article
2. 1. Username
Password
Sign in
2. Sign in
* [ Sign in ](https://bio.libretexts.org/Special:UserLogin?returntotitle=Bookshelves%2FMicrobiology%2FMicrobiology_%28Boundless%29%2F02%3A_Chemistry%2F2.05%3A_Organic_Compounds%2F2.5.06%3A_ATP-_Adenosine_Triphosphate "Sign in")
* [ Forgot password ](https://bio.libretexts.org/Special:UserPassword "Retrieve lost password")
Expand/collapse global hierarchy
1. [ Home ](https://bio.libretexts.org/)
2. [ Bookshelves ](https://bio.libretexts.org/Bookshelves)
3. [ Microbiology ](https://bio.libretexts.org/Bookshelves/Microbiology)
4. [ Microbiology (Boundless) ](https://bio.libretexts.org/Bookshelves/Microbiology/Microbiology_\(Boundless\))
5. [ 2: Chemistry ](https://bio.libretexts.org/Bookshelves/Microbiology/Microbiology_\(Boundless\)/02%3A_Chemistry)
6. [ 2.5: Organic Compounds ](https://bio.libretexts.org/Bookshelves/Microbiology/Microbiology_\(Boundless\)/02%3A_Chemistry/2.05%3A_Organic_Compounds)
7. 2.5.6: ATP- Adenosine Triphosphate
Expand/collapse global location
# 2.5.6: ATP- Adenosine Triphosphate
1. Last updated
2. [ Save as PDF ](https://bio.libretexts.org/@api/deki/pages/8804/pdf/2.5.6%253A%2bATP-%2bAdenosine%2bTriphosphate.pdf "Export page as a PDF")
3.
* Page ID
8804
* 
* Boundless
* Boundless
\\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup}
{\mathbf{#1}} } \\) \\(
\newcommand{\vecd}[1]{\overset{-\\!-\\!\rightharpoonup}{\vphantom{a}\smash
{#1}}} \\)\\(\newcommand{\id}{\mathrm{id}}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\) \\(
\newcommand{\kernel}{\mathrm{null}\,}\\) \\(
\newcommand{\range}{\mathrm{range}\,}\\) \\(
\newcommand{\RealPart}{\mathrm{Re}}\\) \\(
\newcommand{\ImaginaryPart}{\mathrm{Im}}\\) \\(
\newcommand{\Argument}{\mathrm{Arg}}\\) \\( \newcommand{\norm}[1]{\| #1 \|}\\)
\\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\) \\(\newcommand{\id}{\mathrm{id}}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\) \\(
\newcommand{\kernel}{\mathrm{null}\,}\\) \\(
\newcommand{\range}{\mathrm{range}\,}\\) \\(
\newcommand{\RealPart}{\mathrm{Re}}\\) \\(
\newcommand{\ImaginaryPart}{\mathrm{Im}}\\) \\(
\newcommand{\Argument}{\mathrm{Arg}}\\) \\( \newcommand{\norm}[1]{\| #1 \|}\\)
\\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\)\\(\newcommand{\AA}{\unicode[.8,0]{x212B}}\\)
1. Learning Objectives
2. Key Points
3. Key Terms
4. ATP: Adenosine Triphosphate
5. Molecular Structure
6. ATP Hydrolysis and Synthesis
7. ATP and Energy Coupling
8. Energy Coupling in Sodium-Potassium Pumps
9. Energy Coupling in Metabolism
Cells couple the exergonic reaction of ATP hydrolysis with endergonic
reactions to harness the energy within the bonds of ATP.
##### Learning Objectives
* Explain the role of ATP as the currency of cellular energy
## Key Points
* Adenosine triphosphate is composed of the nitrogenous base adenine, the five-carbon sugar ribose, and three phosphate groups.
* ATP is hydrolyzed to ADP in the reaction ATP+H2O→ADP+Pi+ free energy; the calculated ∆G for the hydrolysis of 1 mole of ATP is -57 kJ/mol.
* ADP is combined with a phosphate to form ATP in the reaction ADP+Pi+free energy→ATP+H2O.
* The energy released from the hydrolysis of ATP into ADP is used to perform cellular work, usually by coupling the exergonic reaction of ATP hydrolysis with endergonic reactions.
* Sodium-potassium pumps use the energy derived from exergonic ATP hydrolysis to pump sodium and potassium ions across the cell membrane while phosphorylation drives the endergonic reaction.
## Key Terms
* **energy coupling** : Energy coupling occurs when the energy produced by one reaction or system is used to drive another reaction or system.
* **endergonic** : Describing a reaction that absorbs (heat) energy from its environment.
* **exergonic** : Describing a reaction that releases energy (heat) into its environment.
* **free energy** : Gibbs free energy is a thermodynamic potential that measures the useful or process-initiating work obtainable from a thermodynamic system at a constant temperature and pressure (isothermal, isobaric).
* **hydrolysis** : A chemical process of decomposition involving the splitting of a bond by the addition of water.
## ATP: Adenosine Triphosphate
Adenosine triphosphate (ATP) is the energy currency for cellular processes.
ATP provides the energy for both energy-consuming endergonic reactions and
energy-releasing exergonic reactions, which require a small input of
activation energy. When the chemical bonds within ATP are broken, energy is
released and can be harnessed for cellular work. The more bonds in a molecule,
the more potential energy it contains. Because the bond in ATP is so easily
broken and reformed, ATP is like a rechargeable battery that powers cellular
process ranging from DNA replication to protein synthesis.
## Molecular Structure
Adenosine triphosphate (ATP) is comprised of the molecule adenosine bound to
three phosphate groups. Adenosine is a nucleoside consisting of the
nitrogenous base adenine and the five-carbon sugar ribose. The three phosphate
groups, in order of closest to furthest from the ribose sugar, are labeled
alpha, beta, and gamma. Together, these chemical groups constitute an energy
powerhouse. The two bonds between the phosphates are equal high-energy bonds
(phosphoanhydride bonds) that, when broken, release sufficient energy to power
a variety of cellular reactions and processes. The bond between the beta and
gamma phosphate is considered “high-energy” because when the bond breaks, the
products [adenosine diphosphate (ADP) and one inorganic phosphate group (P i
)] have a lower free energy than the reactants (ATP and a water molecule). ATP
breakdown into ADP and P i is called hydrolysis because it consumes a water
molecule (hydro-, meaning “water”, and lysis, meaning “separation”).
 Figure:
**Adenosine Triphosphate (ATP)** : ATP is the primary energy currency of the
cell. It has an adenosine backbone with three phosphate groups attached.
## ATP Hydrolysis and Synthesis
ATP is hydrolyzed into ADP in the following reaction:
ATP+H 2 O→ADP+P i +free energy
Like most chemical reactions, the hydrolysis of ATP to ADP is reversible. The
reverse reaction combines ADP + P i to regenerate ATP from ADP. Since ATP
hydrolysis releases energy, ATP synthesis must require an input of free
energy.
ADP is combined with a phosphate to form ATP in the following reaction:
ADP+P i +free energy→ATP+H 2 O
## ATP and Energy Coupling
Exactly how much free energy (∆G) is released with the hydrolysis of ATP, and
how is that free energy used to do cellular work? The calculated ∆G for the
hydrolysis of one mole of ATP into ADP and P i is −7.3 kcal/mole (−30.5
kJ/mol). However, this is only true under standard conditions, and the ∆G for
the hydrolysis of one mole of ATP in a living cell is almost double the value
at standard conditions: 14 kcal/mol (−57 kJ/mol).
ATP is a highly unstable molecule. Unless quickly used to perform work, ATP
spontaneously dissociates into ADP + P i , and the free energy released
during this process is lost as heat. To harness the energy within the bonds of
ATP, cells use a strategy called energy coupling.
## Energy Coupling in Sodium-Potassium Pumps
 Figure:
**Energy Coupling** : Sodium-potassium pumps use the energy derived from
exergonic ATP hydrolysis to pump sodium and potassium ions across the cell
membrane.
Cells couple the exergonic reaction of ATP hydrolysis with the endergonic
reactions of cellular processes. For example, transmembrane ion pumps in nerve
cells use the energy from ATP to pump ions across the cell membrane and
generate an action potential. The sodium-potassium pump (Na \+ /K \+ pump)
drives sodium out of the cell and potassium into the cell. When ATP is
hydrolyzed, it transfers its gamma phosphate to the pump protein in a process
called phosphorylation. The Na \+ /K \+ pump gains the free energy and
undergoes a conformational change, allowing it to release three Na \+ to the
outside of the cell. Two extracellular K \+ ions bind to the protein,
causing the protein to change shape again and discharge the phosphate. By
donating free energy to the Na \+ /K \+ pump, phosphorylation drives the
endergonic reaction.
## Energy Coupling in Metabolism
During cellular metabolic reactions, or the synthesis and breakdown of
nutrients, certain molecules must be altered slightly in their conformation to
become substrates for the next step in the reaction series. In the very first
steps of cellular respiration, glucose is broken down through the process of
glycolysis. ATP is required for the phosphorylation of glucose, creating a
high-energy but unstable intermediate. This phosphorylation reaction causes a
conformational change that allows enzymes to convert the phosphorylated
glucose molecule to the phosphorylated sugar fructose. Fructose is a necessary
intermediate for glycolysis to move forward. In this example, the exergonic
reaction of ATP hydrolysis is coupled with the endergonic reaction of
converting glucose for use in the metabolic pathway.
LICENSES AND ATTRIBUTIONS
CC LICENSED CONTENT, SPECIFIC ATTRIBUTION
* dehydration reaction. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/dehydration_reaction. **License** : _CC BY-SA: Attribution-ShareAlike_
* OpenStax College, Biology. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...ol11448/latest ](http://cnx.org/content/m44400/latest/?collection=col11448/latest "http://cnx.org/content/m44400/latest/?collection=col11448/latest") . **License** : _CC BY: Attribution_
* isomer. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/isomer. **License** : _CC BY-SA: Attribution-ShareAlike_
* biopolymer. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/biopolymer. **License** : _CC BY-SA: Attribution-ShareAlike_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_07.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_04.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_01.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Biology. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...ol11448/latest ](http://cnx.org/content/m44401/latest/?collection=col11448/latest "http://cnx.org/content/m44401/latest/?collection=col11448/latest") . **License** : _CC BY: Attribution_
* hydrogenation. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/hydrogenation. **License** : _CC BY-SA: Attribution-ShareAlike_
* ester. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/ester. **License** : _CC BY-SA: Attribution-ShareAlike_
* carboxyl. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/carboxyl. **License** : _CC BY-SA: Attribution-ShareAlike_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_07.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_04.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_01.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_02.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_07.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_05.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Biology. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44403/latest...ol11448/latest ](http://cnx.org/content/m44403/latest/?collection=col11448/latest "http://cnx.org/content/m44403/latest/?collection=col11448/latest") . **License** : _CC BY: Attribution_
* genome. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/genome. **License** : _CC BY-SA: Attribution-ShareAlike_
* nucleotide. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/nucleotide. **License** : _CC BY-SA: Attribution-ShareAlike_
* monomer. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/monomer. **License** : _CC BY-SA: Attribution-ShareAlike_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_07.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_04.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_01.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_02.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_07.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_05.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg") . **License** : _CC BY: Attribution_
* OpenStax College, Nucleic Acids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44403/latest...e_03_05_01.jpg ](http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg "http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg") . **License** : _CC BY: Attribution_
* amino acid. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/amino_acid. **License** : _CC BY-SA: Attribution-ShareAlike_
* OpenStax College, Biology. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...ol11448/latest ](http://cnx.org/content/m44402/latest/?collection=col11448/latest "http://cnx.org/content/m44402/latest/?collection=col11448/latest") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* Boundless. **Provided by** : Boundless Learning. **Located at** : www.boundless.com//biology/definition/r-group. **License** : _CC BY-SA: Attribution-ShareAlike_
* polypeptide. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/polypeptide. **License** : _CC BY-SA: Attribution-ShareAlike_
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_07.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_04.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_01.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_02.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_07.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_05.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Nucleic Acids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44403/latest...e_03_05_01.jpg ](http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg "http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_03.jpg ](http://cnx.org/content/m44402/latest/Figure_03_04_03.jpg "http://cnx.org/content/m44402/latest/Figure_03_04_03.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_02.png ](http://cnx.org/content/m44402/latest/Figure_03_04_02.png "http://cnx.org/content/m44402/latest/Figure_03_04_02.png") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_01.jpg ](http://cnx.org/content/m44402/latest/Figure_03_04_01.jpg "http://cnx.org/content/m44402/latest/Figure_03_04_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* amino acid. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/amino_acid. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* OpenStax College, Biology. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...ol11448/latest ](http://cnx.org/content/m44402/latest/?collection=col11448/latest "http://cnx.org/content/m44402/latest/?collection=col11448/latest") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* Proteins. **Provided by** : Wikipedia. **Located at** : en.Wikipedia.org/wiki/Proteins. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* Structural Biochemistry/Proteins/Amino Acids. **Provided by** : Wikibooks. **Located at** : en.wikibooks.org/wiki/Structu...ns/Amino_Acids. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* catalyze. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/catalyze. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* polypeptide. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/polypeptide. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_07.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_04.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_01.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_02.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_07.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_05.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Nucleic Acids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44403/latest...e_03_05_01.jpg ](http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg "http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_03.jpg ](http://cnx.org/content/m44402/latest/Figure_03_04_03.jpg "http://cnx.org/content/m44402/latest/Figure_03_04_03.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_02.png ](http://cnx.org/content/m44402/latest/Figure_03_04_02.png "http://cnx.org/content/m44402/latest/Figure_03_04_02.png") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_01.jpg ](http://cnx.org/content/m44402/latest/Figure_03_04_01.jpg "http://cnx.org/content/m44402/latest/Figure_03_04_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* BPAE cells | Flickr - Photo Sharing!. **Provided by** : Flickr. **Located at** : [ http://www.flickr.com/photos/6345024...-dtyzGH-duUj14 ](http://www.flickr.com/photos/63450246@N03/6126222546/in/photolist-akmuwE-akiFR6-dxpvoj-duUjsi-dwLu92-dwh63u-dtyzGH-duUj14 "http://www.flickr.com/photos/63450246@N03/6126222546/in/photolist-akmuwE-akiFR6-dxpvoj-duUjsi-dwLu92-dwh63u-dtyzGH-duUj14") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* 1GZX Haemoglobin. **Provided by** : Wikipedia. **Located at** : en.Wikipedia.org/wiki/File:1G...aemoglobin.png. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* Induced fit diagram. **Provided by** : Wikipedia. **Located at** : en.Wikipedia.org/wiki/File:In...it_diagram.svg. **License** : _Public Domain: No Known Copyright_
* OpenStax College, Biology. October 21, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44427/latest...ol11448/latest ](http://cnx.org/content/m44427/latest/?collection=col11448/latest "http://cnx.org/content/m44427/latest/?collection=col11448/latest") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Biology. October 26, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44427/latest...ol11448/latest ](http://cnx.org/content/m44427/latest/?collection=col11448/latest "http://cnx.org/content/m44427/latest/?collection=col11448/latest") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* hydrolysis. **Provided by** : Wiktionary. **Located at** : [ http://en.wiktionary.org/wiki/hydrolysis ](http://en.wiktionary.org/wiki/hydrolysis "http://en.wiktionary.org/wiki/hydrolysis") . **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* exergonic. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/exergonic. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* endergonic. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/endergonic. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* Boundless. **Provided by** : Boundless Learning. **Located at** : www.boundless.com//biology/de...nergy-coupling. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* free energy. **Provided by** : Wiktionary. **Located at** : en.wiktionary.org/wiki/free_energy. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_07.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_07.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_04.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_04.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Carbohydrates. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44400/latest...e_03_02_01.jpg ](http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg "http://cnx.org/content/m44400/latest/Figure_03_02_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_02.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_02.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_07.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_07.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Lipids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44401/latest...e_03_03_05.jpg ](http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg "http://cnx.org/content/m44401/latest/Figure_03_03_05.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Nucleic Acids. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44403/latest...e_03_05_01.jpg ](http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg "http://cnx.org/content/m44403/latest/Figure_03_05_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_03.jpg ](http://cnx.org/content/m44402/latest/Figure_03_04_03.jpg "http://cnx.org/content/m44402/latest/Figure_03_04_03.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_02.png ](http://cnx.org/content/m44402/latest/Figure_03_04_02.png "http://cnx.org/content/m44402/latest/Figure_03_04_02.png") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Proteins. October 16, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44402/latest...e_03_04_01.jpg ](http://cnx.org/content/m44402/latest/Figure_03_04_01.jpg "http://cnx.org/content/m44402/latest/Figure_03_04_01.jpg") . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* BPAE cells | Flickr - Photo Sharing!. **Provided by** : Flickr. **Located at** : [ http://www.flickr.com/photos/63450246@N03/6126222546/in/photolist-akmuwE-akiFR6-dxpvoj-duUjsi-dwLu92-dwh63u-dtyzGH-duUj14 ](http://www.flickr.com/photos/63450246@N03/6126222546/in/photolist-akmuwE-akiFR6-dxpvoj-duUjsi-dwLu92-dwh63u-dtyzGH-duUj14) . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* 1GZX Haemoglobin. **Provided by** : Wikipedia. **Located at** : en.Wikipedia.org/wiki/File:1GZX_Haemoglobin.png. **License** : _[ CC BY-SA: Attribution-ShareAlike ](https://creativecommons.org/licenses/by-sa/4.0/) _
* Induced fit diagram. **Provided by** : Wikipedia. **Located at** : en.Wikipedia.org/wiki/File:Induced_fit_diagram.svg. **License** : _[ Public Domain: No Known Copyright ](https://creativecommons.org/about/pdm) _
* OpenStax College, Biology. October 26, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44427/latest/?collection=col11448/latest ](http://cnx.org/content/m44427/latest/?collection=col11448/latest) . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* OpenStax College, Biology. October 26, 2013. **Provided by** : OpenStax CNX. **Located at** : [ http://cnx.org/content/m44427/latest/?collection=col11448/latest ](http://cnx.org/content/m44427/latest/?collection=col11448/latest) . **License** : _[ CC BY: Attribution ](https://creativecommons.org/licenses/by/4.0/) _
* * *
This page titled [ 2.5.6: ATP- Adenosine Triphosphate
](https://bio.libretexts.org/Bookshelves/Microbiology/Microbiology_\(Boundless\)/02%3A_Chemistry/2.05%3A_Organic_Compounds/2.5.06%3A_ATP-
_Adenosine_Triphosphate) is shared under a [ CC BY-SA 4.0
](https://creativecommons.org/licenses/by-sa/4.0) license and was authored,
remixed, and/or curated by [ Boundless ]() .
1. Back to top
2. * [ 2.5.5: Types and Functions of Protiens ](https://bio.libretexts.org/Bookshelves/Microbiology/Microbiology_\(Boundless\)/02%3A_Chemistry/2.05%3A_Organic_Compounds/2.5.05%3A_Types_and_Functions_of_Protiens "2.5.5: Types and Functions of Protiens")
* [ 2.6: Energy ](https://bio.libretexts.org/Bookshelves/Microbiology/Microbiology_\(Boundless\)/02%3A_Chemistry/2.06%3A_Energy "2.6: Energy")
* Was this article helpful?
* Yes
* No
*
## Recommended articles
1. Article type
[ Section or Page ](https://bio.libretexts.org/Special:Search?tags=article%3Atopic)
Author
[ Boundless ](https://bio.libretexts.org/Special:Search?tags=authorname%3Aboundless)
License
[ CC BY-SA ](https://bio.libretexts.org/Special:Search?tags=license%3Accbysa)
License Version
[ 4.0 ](https://bio.libretexts.org/Special:Search?tags=licenseversion%3A40)
Show TOC
[ no ](https://bio.libretexts.org/Special:Search?tags=showtoc%3Ano)
2. Tags
This page has no tags.
1. © Copyright 2024 Biology LibreTexts
2. [ Powered by CXone Expert ® ](https://mindtouch.com/demo "MindTouch")
The LibreTexts libraries are [ Powered by NICE CXone Expert
](https://www.nice.com/resources/cxone-expert-knowledge-management) and are
supported by the Department of Education Open Textbook Pilot Project, the UC
Davis Office of the Provost, the UC Davis Library, the California State
University Affordable Learning Solutions Program, and Merlot. We also
acknowledge previous National Science Foundation support under grant numbers
1246120, 1525057, and 1413739. [ Legal
](https://libretexts.org/legal/index.html) . [ Accessibility Statement
](https://commons.libretexts.org/accessibility) For more information contact
us at [ [email protected] ](mailto:[email protected]) .
| biology | 4124264 | https://sv.wikipedia.org/wiki/Cycas%20ophiolitica | Cycas ophiolitica | Cycas ophiolitica är en kärlväxtart som beskrevs av Kenneth D. Hill. Cycas ophiolitica ingår i släktet Cycas, och familjen Cycadaceae. IUCN kategoriserar arten globalt som sårbar. Inga underarter finns listade i Catalogue of Life.
Bildgalleri
Källor
Externa länkar
Kärlväxter
ophiolitica | swedish | 1.315552 |
induce_sleep/Adenosinetriphosphat.txt | Jump to content
Main menu
Main menu
move to sidebar hide
Navigation
* [ Main page ](/wiki/Main_Page "Visit the main page \[z\]")
* [ Contents ](/wiki/Wikipedia:Contents "Guides to browsing Wikipedia")
* [ Current events ](/wiki/Portal:Current_events "Articles related to current events")
* [ Random article ](/wiki/Special:Random "Visit a randomly selected article \[x\]")
* [ About Wikipedia ](/wiki/Wikipedia:About "Learn about Wikipedia and how it works")
* [ Contact us ](//en.wikipedia.org/wiki/Wikipedia:Contact_us "How to contact Wikipedia")
* [ Donate ](https://donate.wikimedia.org/wiki/Special:FundraiserRedirector?utm_source=donate&utm_medium=sidebar&utm_campaign=C13_en.wikipedia.org&uselang=en "Support us by donating to the Wikimedia Foundation")
Contribute
* [ Help ](/wiki/Help:Contents "Guidance on how to use and edit Wikipedia")
* [ Learn to edit ](/wiki/Help:Introduction "Learn how to edit Wikipedia")
* [ Community portal ](/wiki/Wikipedia:Community_portal "The hub for editors")
* [ Recent changes ](/wiki/Special:RecentChanges "A list of recent changes to Wikipedia \[r\]")
* [ Upload file ](/wiki/Wikipedia:File_upload_wizard "Add images or other media for use on Wikipedia")
[ 
 
](/wiki/Main_Page)
[ Search ](/wiki/Special:Search "Search Wikipedia \[f\]")
Search
* [ Create account ](/w/index.php?title=Special:CreateAccount&returnto=Adenosine+triphosphate "You are encouraged to create an account and log in; however, it is not mandatory")
* [ Log in ](/w/index.php?title=Special:UserLogin&returnto=Adenosine+triphosphate "You're encouraged to log in; however, it's not mandatory. \[o\]")
Personal tools
* [ Create account ](/w/index.php?title=Special:CreateAccount&returnto=Adenosine+triphosphate "You are encouraged to create an account and log in; however, it is not mandatory")
* [ Log in ](/w/index.php?title=Special:UserLogin&returnto=Adenosine+triphosphate "You're encouraged to log in; however, it's not mandatory. \[o\]")
Pages for logged out editors [ learn more ](/wiki/Help:Introduction)
* [ Contributions ](/wiki/Special:MyContributions "A list of edits made from this IP address \[y\]")
* [ Talk ](/wiki/Special:MyTalk "Discussion about edits from this IP address \[n\]")
## Contents
move to sidebar hide
* (Top)
* 1 Structure Toggle Structure subsection
* 1.1 Metal cation binding
* 2 Chemical properties
* 3 Reactive aspects
* 4 Production from AMP and ADP Toggle Production from AMP and ADP subsection
* 4.1 Production, aerobic conditions
* 4.1.1 Glycolysis
* 4.1.1.1 Regulation
* 4.1.2 Citric acid cycle
* 4.1.2.1 Regulation
* 4.1.3 Beta oxidation
* 4.1.3.1 Regulation
* 4.1.4 Ketosis
* 4.2 Production, anaerobic conditions
* 4.2.1 ATP replenishment by nucleoside diphosphate kinases
* 4.3 ATP production during photosynthesis
* 4.4 ATP recycling
* 5 Biochemical functions Toggle Biochemical functions subsection
* 5.1 Intracellular signaling
* 5.2 DNA and RNA synthesis
* 5.3 Amino acid activation in protein synthesis
* 5.4 ATP binding cassette transporter
* 5.5 Extracellular signalling and neurotransmission
* 5.6 Protein solubility
* 6 Abiogenic origins
* 7 ATP analogues
* 8 Medical use
* 9 History
* 10 See also
* 11 References
* 12 External links
Toggle the table of contents
# Adenosine triphosphate
81 languages
* [ Afrikaans ](https://af.wikipedia.org/wiki/Adenosientrifosfaat "Adenosientrifosfaat – Afrikaans")
* [ العربية ](https://ar.wikipedia.org/wiki/%D8%A3%D8%AF%D9%8A%D9%86%D9%88%D8%B3%D9%8A%D9%86_%D8%AB%D9%84%D8%A7%D8%AB%D9%8A_%D8%A7%D9%84%D9%81%D9%88%D8%B3%D9%81%D8%A7%D8%AA "أدينوسين ثلاثي الفوسفات – Arabic")
* [ Azərbaycanca ](https://az.wikipedia.org/wiki/Adenozintrifosfat_tur%C5%9Fusu "Adenozintrifosfat turşusu – Azerbaijani")
* [ تۆرکجه ](https://azb.wikipedia.org/wiki/%D8%A2%D8%AF%D9%86%D9%88%D8%B2%DB%8C%D9%86_%D8%AA%D8%B1%DB%8C%D9%81%D9%88%D8%B3%D9%81%D8%A7%D8%AA "آدنوزین تریفوسفات – South Azerbaijani")
* [ বাংলা ](https://bn.wikipedia.org/wiki/%E0%A6%85%E0%A7%8D%E0%A6%AF%E0%A6%BE%E0%A6%A1%E0%A6%BF%E0%A6%A8%E0%A7%8B%E0%A6%B8%E0%A6%BF%E0%A6%A8_%E0%A6%9F%E0%A7%8D%E0%A6%B0%E0%A6%BE%E0%A6%87%E0%A6%AB%E0%A6%B8%E0%A6%AB%E0%A7%87%E0%A6%9F "অ্যাডিনোসিন ট্রাইফসফেট – Bangla")
* [ 閩南語 / Bân-lâm-gú ](https://zh-min-nan.wikipedia.org/wiki/ATP "ATP – Minnan")
* [ Башҡортса ](https://ba.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%BE%D1%80_%D0%BA%D0%B8%D1%81%D0%BB%D0%BE%D1%82%D0%B0%D2%BB%D1%8B_%E2%80%94_%D0%90%D0%A2%D0%A4 "Аденозинтрифосфор кислотаһы — АТФ – Bashkir")
* [ Беларуская ](https://be.wikipedia.org/wiki/%D0%90%D0%B4%D1%8D%D0%BD%D0%B0%D0%B7%D1%96%D0%BD%D1%82%D1%80%D1%8B%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%80%D0%BD%D0%B0%D1%8F_%D0%BA%D1%96%D1%81%D0%BB%D0%B0%D1%82%D0%B0 "Адэназінтрыфосфарная кіслата – Belarusian")
* [ Български ](https://bg.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозинтрифосфат – Bulgarian")
* [ བོད་ཡིག ](https://bo.wikipedia.org/wiki/%E0%BD%A8%E0%BD%BA%E0%BC%8B%E0%BD%8A%E0%BD%B2%E0%BC%8B%E0%BD%94%E0%BD%B2%E0%BC%8B%E0%BD%A0%E0%BD%91%E0%BD%B4%E0%BD%A6%E0%BC%8B%E0%BD%A2%E0%BE%A1%E0%BD%B4%E0%BD%A3%E0%BC%8D "ཨེ་ཊི་པི་འདུས་རྡུལ། – Tibetan")
* [ Bosanski ](https://bs.wikipedia.org/wiki/Adenozin-trifosfat "Adenozin-trifosfat – Bosnian")
* [ Català ](https://ca.wikipedia.org/wiki/Trifosfat_d%27adenosina "Trifosfat d'adenosina – Catalan")
* [ Čeština ](https://cs.wikipedia.org/wiki/Adenosintrifosf%C3%A1t "Adenosintrifosfát – Czech")
* [ Dansk ](https://da.wikipedia.org/wiki/Adenosintrifosfat "Adenosintrifosfat – Danish")
* [ Deutsch ](https://de.wikipedia.org/wiki/Adenosintriphosphat "Adenosintriphosphat – German")
* [ Eesti ](https://et.wikipedia.org/wiki/Adenosiintrifosfaat "Adenosiintrifosfaat – Estonian")
* [ Ελληνικά ](https://el.wikipedia.org/wiki/%CE%A4%CF%81%CE%B9%CF%86%CF%89%CF%83%CF%86%CE%BF%CF%81%CE%B9%CE%BA%CE%AE_%CE%B1%CE%B4%CE%B5%CE%BD%CE%BF%CF%83%CE%AF%CE%BD%CE%B7 "Τριφωσφορική αδενοσίνη – Greek")
* [ Español ](https://es.wikipedia.org/wiki/Adenos%C3%ADn_trifosfato "Adenosín trifosfato – Spanish")
* [ Esperanto ](https://eo.wikipedia.org/wiki/Adenozina_trifosfato "Adenozina trifosfato – Esperanto")
* [ Euskara ](https://eu.wikipedia.org/wiki/Adenosina_trifosfato "Adenosina trifosfato – Basque")
* [ فارسی ](https://fa.wikipedia.org/wiki/%D8%A2%D8%AF%D9%86%D9%88%D8%B2%DB%8C%D9%86_%D8%AA%D8%B1%DB%8C%E2%80%8C%D9%81%D8%B3%D9%81%D8%A7%D8%AA "آدنوزین تریفسفات – Persian")
* [ Français ](https://fr.wikipedia.org/wiki/Ad%C3%A9nosine_triphosphate "Adénosine triphosphate – French")
* [ Gaeilge ](https://ga.wikipedia.org/wiki/Tr%C3%ADfhosf%C3%A1it_adan%C3%B3is%C3%ADn "Trífhosfáit adanóisín – Irish")
* [ Galego ](https://gl.wikipedia.org/wiki/Adenos%C3%ADn_trifosfato "Adenosín trifosfato – Galician")
* [ 한국어 ](https://ko.wikipedia.org/wiki/%EC%95%84%EB%8D%B0%EB%85%B8%EC%8B%A0_%EC%82%BC%EC%9D%B8%EC%82%B0 "아데노신 삼인산 – Korean")
* [ Հայերեն ](https://hy.wikipedia.org/wiki/%D4%B1%D5%A4%D5%A5%D5%B6%D5%B8%D5%A6%D5%AB%D5%B6%D5%A5%D5%BC%D6%86%D5%B8%D5%BD%D6%86%D5%A1%D5%BF "Ադենոզինեռֆոսֆատ – Armenian")
* [ हिन्दी ](https://hi.wikipedia.org/wiki/%E0%A4%8F%E0%A4%A1%E0%A5%80%E0%A4%A8%E0%A5%8B%E0%A4%B8%E0%A4%BF%E0%A4%A8_%E0%A4%9F%E0%A5%8D%E0%A4%B0%E0%A4%BE%E0%A4%87%E0%A4%AB%E0%A5%89%E0%A4%B8%E0%A5%8D%E0%A4%AB%E0%A5%87%E0%A4%9F "एडीनोसिन ट्राइफॉस्फेट – Hindi")
* [ Hrvatski ](https://hr.wikipedia.org/wiki/Adenozin_trifosfat "Adenozin trifosfat – Croatian")
* [ Bahasa Indonesia ](https://id.wikipedia.org/wiki/Adenosina_trifosfat "Adenosina trifosfat – Indonesian")
* [ Íslenska ](https://is.wikipedia.org/wiki/Aden%C3%B3s%C3%ADn%C3%BEr%C3%ADfosfat "Adenósínþrífosfat – Icelandic")
* [ Italiano ](https://it.wikipedia.org/wiki/Adenosina_trifosfato "Adenosina trifosfato – Italian")
* [ עברית ](https://he.wikipedia.org/wiki/ATP "ATP – Hebrew")
* [ Jawa ](https://jv.wikipedia.org/wiki/Adenosin_Triphosphate "Adenosin Triphosphate – Javanese")
* [ ಕನ್ನಡ ](https://kn.wikipedia.org/wiki/%E0%B2%85%E0%B2%A1%E0%B3%86%E0%B2%A8%E0%B3%8B%E0%B2%B8%E0%B3%80%E0%B2%A8%E0%B3%8D_%E0%B2%9F%E0%B3%8D%E0%B2%B0%E0%B3%88%E0%B2%AB%E0%B2%BE%E0%B2%B8%E0%B3%8D%E0%B2%AB%E0%B3%87%E0%B2%9F%E0%B3%8D_\(%E0%B2%8E_%E0%B2%9F%E0%B2%BF_%E0%B2%AA%E0%B2%BF\) "ಅಡೆನೋಸೀನ್ ಟ್ರೈಫಾಸ್ಫೇಟ್ \(ಎ ಟಿ ಪಿ\) – Kannada")
* [ ქართული ](https://ka.wikipedia.org/wiki/%E1%83%90%E1%83%93%E1%83%94%E1%83%9C%E1%83%9D%E1%83%96%E1%83%98%E1%83%9C%E1%83%A2%E1%83%A0%E1%83%98%E1%83%A4%E1%83%9D%E1%83%A1%E1%83%A4%E1%83%9D%E1%83%A0%E1%83%9B%E1%83%9F%E1%83%90%E1%83%95%E1%83%90 "ადენოზინტრიფოსფორმჟავა – Georgian")
* [ Қазақша ](https://kk.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D2%AF%D1%88%D1%84%D0%BE%D1%81%D1%84%D0%BE%D1%80_%D2%9B%D1%8B%D1%88%D2%9B%D1%8B%D0%BB%D1%8B "Аденозинүшфосфор қышқылы – Kazakh")
* [ Kreyòl ayisyen ](https://ht.wikipedia.org/wiki/Adenozin_trifosfat "Adenozin trifosfat – Haitian Creole")
* [ Kurdî ](https://ku.wikipedia.org/wiki/Adenoz%C3%AEna_tr%C3%AEfosfat "Adenozîna trîfosfat – Kurdish")
* [ Кыргызча ](https://ky.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозинтрифосфат – Kyrgyz")
* [ Latina ](https://la.wikipedia.org/wiki/Adenosinum_triphosphoricum "Adenosinum triphosphoricum – Latin")
* [ Latviešu ](https://lv.wikipedia.org/wiki/Adenoz%C4%ABntrifosforsk%C4%81be "Adenozīntrifosforskābe – Latvian")
* [ Lëtzebuergesch ](https://lb.wikipedia.org/wiki/Adenosintriphosphat "Adenosintriphosphat – Luxembourgish")
* [ Lietuvių ](https://lt.wikipedia.org/wiki/ATP "ATP – Lithuanian")
* [ Lingua Franca Nova ](https://lfn.wikipedia.org/wiki/Trifosfato_de_adenosina "Trifosfato de adenosina – Lingua Franca Nova")
* [ Lombard ](https://lmo.wikipedia.org/wiki/Adenosina_trifosfad "Adenosina trifosfad – Lombard")
* [ Magyar ](https://hu.wikipedia.org/wiki/Adenozin-trifoszf%C3%A1t "Adenozin-trifoszfát – Hungarian")
* [ Македонски ](https://mk.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD_%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозин трифосфат – Macedonian")
* [ മലയാളം ](https://ml.wikipedia.org/wiki/%E0%B4%85%E0%B4%A1%E0%B4%BF%E0%B4%A8%E0%B5%8B%E0%B4%B8%E0%B4%BF%E0%B5%BB_%E0%B4%9F%E0%B5%8D%E0%B4%B0%E0%B5%88%E0%B4%AB%E0%B5%8B%E0%B4%B8%E0%B5%8D%E0%B4%AB%E0%B5%87%E0%B4%B1%E0%B5%8D%E0%B4%B1%E0%B5%8D "അഡിനോസിൻ ട്രൈഫോസ്ഫേറ്റ് – Malayalam")
* [ Bahasa Melayu ](https://ms.wikipedia.org/wiki/Adenosina_trifosfat "Adenosina trifosfat – Malay")
* [ Nederlands ](https://nl.wikipedia.org/wiki/Adenosinetrifosfaat "Adenosinetrifosfaat – Dutch")
* [ 日本語 ](https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%87%E3%83%8E%E3%82%B7%E3%83%B3%E4%B8%89%E3%83%AA%E3%83%B3%E9%85%B8 "アデノシン三リン酸 – Japanese")
* [ Nordfriisk ](https://frr.wikipedia.org/wiki/Adenosintriphosphat "Adenosintriphosphat – Northern Frisian")
* [ Norsk bokmål ](https://no.wikipedia.org/wiki/Adenosintrifosfat "Adenosintrifosfat – Norwegian Bokmål")
* [ Norsk nynorsk ](https://nn.wikipedia.org/wiki/Adenosintrifosfat "Adenosintrifosfat – Norwegian Nynorsk")
* [ Occitan ](https://oc.wikipedia.org/wiki/Adenosina_trifosfat "Adenosina trifosfat – Occitan")
* [ Oʻzbekcha / ўзбекча ](https://uz.wikipedia.org/wiki/Adenozintrifosfat_kislota "Adenozintrifosfat kislota – Uzbek")
* [ ਪੰਜਾਬੀ ](https://pa.wikipedia.org/wiki/%E0%A8%85%E0%A8%A1%E0%A9%88%E0%A8%A8%E0%A9%8B%E0%A8%B8%E0%A9%80%E0%A8%A8_%E0%A8%9F%E0%A8%B0%E0%A8%BE%E0%A8%88%E0%A8%AB%E0%A8%BC%E0%A9%8C%E0%A8%B8%E0%A8%AB%E0%A8%BC%E0%A9%87%E0%A8%9F "ਅਡੈਨੋਸੀਨ ਟਰਾਈਫ਼ੌਸਫ਼ੇਟ – Punjabi")
* [ Polski ](https://pl.wikipedia.org/wiki/Adenozyno-5%E2%80%B2-trifosforan "Adenozyno-5′-trifosforan – Polish")
* [ Português ](https://pt.wikipedia.org/wiki/Trifosfato_de_adenosina "Trifosfato de adenosina – Portuguese")
* [ Română ](https://ro.wikipedia.org/wiki/Adenozin_trifosfat "Adenozin trifosfat – Romanian")
* [ Русиньскый ](https://rue.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D1%96%D0%BD%D1%82%D1%80%D1%96%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозінтріфосфат – Rusyn")
* [ Русский ](https://ru.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозинтрифосфат – Russian")
* [ Seeltersk ](https://stq.wikipedia.org/wiki/Adenosintriphosphat "Adenosintriphosphat – Saterland Frisian")
* [ Shqip ](https://sq.wikipedia.org/wiki/Adenozintrifosfat "Adenozintrifosfat – Albanian")
* [ Simple English ](https://simple.wikipedia.org/wiki/Adenosine_triphosphate "Adenosine triphosphate – Simple English")
* [ Slovenčina ](https://sk.wikipedia.org/wiki/Adenoz%C3%ADntrifosf%C3%A1t "Adenozíntrifosfát – Slovak")
* [ Slovenščina ](https://sl.wikipedia.org/wiki/Adenozin_trifosfat "Adenozin trifosfat – Slovenian")
* [ Српски / srpski ](https://sr.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD-%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозин-трифосфат – Serbian")
* [ Srpskohrvatski / српскохрватски ](https://sh.wikipedia.org/wiki/Adenozin_trifosfat "Adenozin trifosfat – Serbo-Croatian")
* [ Sunda ](https://su.wikipedia.org/wiki/Ad%C3%A9nosin_trifosfat "Adénosin trifosfat – Sundanese")
* [ Suomi ](https://fi.wikipedia.org/wiki/Adenosiinitrifosfaatti "Adenosiinitrifosfaatti – Finnish")
* [ Svenska ](https://sv.wikipedia.org/wiki/Adenosintrifosfat "Adenosintrifosfat – Swedish")
* [ தமிழ் ](https://ta.wikipedia.org/wiki/%E0%AE%85%E0%AE%9F%E0%AE%BF%E0%AE%A9%E0%AF%8B%E0%AE%9A%E0%AE%BF%E0%AE%A9%E0%AF%8D_%E0%AE%AE%E0%AF%81%E0%AE%AA%E0%AF%8D%E0%AE%AA%E0%AF%8A%E0%AE%9A%E0%AF%81%E0%AE%AA%E0%AF%87%E0%AE%B1%E0%AF%8D%E0%AE%B1%E0%AF%81 "அடினோசின் முப்பொசுபேற்று – Tamil")
* [ ไทย ](https://th.wikipedia.org/wiki/%E0%B8%AD%E0%B8%B0%E0%B8%94%E0%B8%B5%E0%B9%82%E0%B8%99%E0%B8%8B%E0%B8%B5%E0%B8%99%E0%B9%84%E0%B8%95%E0%B8%A3%E0%B8%9F%E0%B8%AD%E0%B8%AA%E0%B9%80%E0%B8%9F%E0%B8%95 "อะดีโนซีนไตรฟอสเฟต – Thai")
* [ Türkçe ](https://tr.wikipedia.org/wiki/Adenozin_trifosfat "Adenozin trifosfat – Turkish")
* [ Українська ](https://uk.wikipedia.org/wiki/%D0%90%D0%B4%D0%B5%D0%BD%D0%BE%D0%B7%D0%B8%D0%BD%D1%82%D1%80%D0%B8%D1%84%D0%BE%D1%81%D1%84%D0%B0%D1%82 "Аденозинтрифосфат – Ukrainian")
* [ اردو ](https://ur.wikipedia.org/wiki/%D8%A7%DA%88%DB%8C%D9%86%D9%88_%D8%B3%D8%A7%D8%A6%D9%86_%D9%B9%D8%B1%D8%A7%D8%A6%DB%8C_%D9%81%D8%A7%D8%B3%D9%81%DB%8C%D9%B9 "اڈینو سائن ٹرائی فاسفیٹ – Urdu")
* [ Tiếng Việt ](https://vi.wikipedia.org/wiki/Adenosine_triphosphate "Adenosine triphosphate – Vietnamese")
* [ 吴语 ](https://wuu.wikipedia.org/wiki/%E4%B8%89%E7%A3%B7%E9%85%B8%E8%85%BA%E8%8B%B7 "三磷酸腺苷 – Wu")
* [ 粵語 ](https://zh-yue.wikipedia.org/wiki/%E4%B8%89%E7%A3%B7%E9%85%B8%E8%85%BA%E8%8B%B7 "三磷酸腺苷 – Cantonese")
* [ 中文 ](https://zh.wikipedia.org/wiki/%E4%B8%89%E7%A3%B7%E9%85%B8%E8%85%BA%E8%8B%B7 "三磷酸腺苷 – Chinese")
[ Edit links
](https://www.wikidata.org/wiki/Special:EntityPage/Q80863#sitelinks-wikipedia
"Edit interlanguage links")
* [ Article ](/wiki/Adenosine_triphosphate "View the content page \[c\]")
* [ Talk ](/wiki/Talk:Adenosine_triphosphate "Discuss improvements to the content page \[t\]")
English
* [ Read ](/wiki/Adenosine_triphosphate)
* [ Edit ](/w/index.php?title=Adenosine_triphosphate&action=edit "Edit this page \[e\]")
* [ View history ](/w/index.php?title=Adenosine_triphosphate&action=history "Past revisions of this page \[h\]")
Tools
Tools
move to sidebar hide
Actions
* [ Read ](/wiki/Adenosine_triphosphate)
* [ Edit ](/w/index.php?title=Adenosine_triphosphate&action=edit "Edit this page \[e\]")
* [ View history ](/w/index.php?title=Adenosine_triphosphate&action=history)
General
* [ What links here ](/wiki/Special:WhatLinksHere/Adenosine_triphosphate "List of all English Wikipedia pages containing links to this page \[j\]")
* [ Related changes ](/wiki/Special:RecentChangesLinked/Adenosine_triphosphate "Recent changes in pages linked from this page \[k\]")
* [ Upload file ](/wiki/Wikipedia:File_Upload_Wizard "Upload files \[u\]")
* [ Special pages ](/wiki/Special:SpecialPages "A list of all special pages \[q\]")
* [ Permanent link ](/w/index.php?title=Adenosine_triphosphate&oldid=1219758132 "Permanent link to this revision of this page")
* [ Page information ](/w/index.php?title=Adenosine_triphosphate&action=info "More information about this page")
* [ Cite this page ](/w/index.php?title=Special:CiteThisPage&page=Adenosine_triphosphate&id=1219758132&wpFormIdentifier=titleform "Information on how to cite this page")
* [ Get shortened URL ](/w/index.php?title=Special:UrlShortener&url=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FAdenosine_triphosphate)
* [ Download QR code ](/w/index.php?title=Special:QrCode&url=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FAdenosine_triphosphate)
* [ Wikidata item ](https://www.wikidata.org/wiki/Special:EntityPage/Q80863 "Structured data on this page hosted by Wikidata \[g\]")
Print/export
* [ Download as PDF ](/w/index.php?title=Special:DownloadAsPdf&page=Adenosine_triphosphate&action=show-download-screen "Download this page as a PDF file")
* [ Printable version ](/w/index.php?title=Adenosine_triphosphate&printable=yes "Printable version of this page \[p\]")
In other projects
* [ Wikimedia Commons ](https://commons.wikimedia.org/wiki/Category:Adenosine_triphosphate)
From Wikipedia, the free encyclopedia
Energy-carrying molecule in living cells
Adenosine-5'-triphosphate [

](/wiki/File:Adenosintriphosphat_protoniert.svg)
---
| [  ](/wiki/File:ATP-
xtal-3D-balls.png) | [
 ](/wiki/File:ATP-xtal-3D-vdW.png)
---|---
Names
[ IUPAC name ](/wiki/Chemical_nomenclature "Chemical nomenclature")
Adenosine 5′-(tetrahydrogen triphosphate)
[ Systematic IUPAC name ](/wiki/Chemical_nomenclature#Systematic_name
"Chemical nomenclature")
_O_ 1 -{[(2 _R_ ,3 _S_ ,4 _R_ ,5 _R_ )-5-(6-Amino-9 _H_
-purin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl} tetrahydrogen triphosphate
Identifiers
[ CAS Number ](/wiki/CAS_Registry_Number "CAS Registry Number")
|
* [ 56-65-5 ](https://commonchemistry.cas.org/detail?cas_rn=56-65-5) (free acid)  Y
* [ 34369-07-8 ](https://commonchemistry.cas.org/detail?cas_rn=34369-07-8) (disodium salt hydrate)  N
3D model ( [ JSmol ](/wiki/JSmol "JSmol") )
|
* [ Interactive image ](https://chemapps.stolaf.edu/jmol/jmol.php?model=O%3DP%28O%29%28O%29OP%28%3DO%29%28O%29OP%28%3DO%29%28O%29OC%5BC%40H%5D3O%5BC%40%40H%5D%28n2cnc1c%28ncnc12%29N%29%5BC%40H%5D%28O%29%5BC%40%40H%5D3O)
* [ Interactive image ](https://chemapps.stolaf.edu/jmol/jmol.php?model=c1nc%28c2c%28n1%29n%28cn2%29%5BC%40H%5D3%5BC%40%40H%5D%28%5BC%40%40H%5D%28%5BC%40H%5D%28O3%29COP%28%3DO%29%28O%29OP%28%3DO%29%28O%29OP%28%3DO%29%28O%29O%29O%29O%29N)
[ ChEBI ](/wiki/ChEBI "ChEBI") |
* [ CHEBI:15422 ](https://www.ebi.ac.uk/chebi/searchId.do?chebiId=15422)  Y
[ ChEMBL ](/wiki/ChEMBL "ChEMBL") |
* [ ChEMBL14249 ](https://www.ebi.ac.uk/chembldb/index.php/compound/inspect/ChEMBL14249)  Y
[ ChemSpider ](/wiki/ChemSpider "ChemSpider") |
* [ 5742 ](https://www.chemspider.com/Chemical-Structure.5742.html)  Y
[ DrugBank ](/wiki/DrugBank "DrugBank") |
* [ DB00171 ](https://www.drugbank.ca/drugs/DB00171)  Y
[ ECHA InfoCard ](/wiki/ECHA_InfoCard "ECHA InfoCard") | [ 100.000.258
](https://echa.europa.eu/substance-information/-/substanceinfo/100.000.258) [

](https://www.wikidata.org/wiki/Q80863#P2566 "Edit this at Wikidata")
[ IUPHAR/BPS ](/wiki/IUPHAR/BPS "IUPHAR/BPS")
|
* [ 1713 ](http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?tab=summary&ligandId=1713)
[ KEGG ](/wiki/KEGG "KEGG") |
* [ C00002 ](https://www.kegg.jp/entry/C00002)  Y
[ PubChem ](/wiki/PubChem "PubChem") CID
|
* [ 5957 ](https://pubchem.ncbi.nlm.nih.gov/compound/5957)
[ UNII ](/wiki/Unique_Ingredient_Identifier "Unique Ingredient Identifier") |
* [ 8L70Q75FXE ](https://precision.fda.gov/uniisearch/srs/unii/8L70Q75FXE)  Y
[ CompTox Dashboard ](/wiki/CompTox_Chemicals_Dashboard "CompTox Chemicals
Dashboard") ( EPA )
|
* [ DTXSID6022559 ](https://comptox.epa.gov/dashboard/chemical/details/DTXSID6022559) [  ](https://www.wikidata.org/wiki/Q80863#P3117 "Edit this at Wikidata")
[ InChI ](/wiki/International_Chemical_Identifier "International Chemical
Identifier")
* InChI=1S/C10H16N5O13P3/c11-8-5-9(13-2-12-8)15(3-14-5)10-7(17)6(16)4(26-10)1-25-30(21,22)28-31(23,24)27-29(18,19)20/h2-4,6-7,10,16-17H,1H2,(H,21,22)(H,23,24)(H2,11,12,13)(H2,18,19,20)/t4-,6-,7-,10-/m1/s1  Y
Key: ZKHQWZAMYRWXGA-KQYNXXCUSA-N
 Y
* Key: ZKHQWZAMYRWXGA-KQYNXXCUBG
[ SMILES ](/wiki/Simplified_molecular-input_line-entry_system "Simplified
molecular-input line-entry system")
* O=P(O)(O)OP(=O)(O)OP(=O)(O)OC[C@H]3O[C@@H](n2cnc1c(ncnc12)N)[C@H](O)[C@@H]3O
* c1nc(c2c(n1)n(cn2)[C@H]3[C@@H]([C@@H]([C@H](O3)COP(=O)(O)OP(=O)(O)OP(=O)(O)O)O)O)N
Properties
[ Chemical formula ](/wiki/Chemical_formula "Chemical formula")
| C 10 H 16 N 5 O 13 P 3
[ Molar mass ](/wiki/Molar_mass "Molar mass") | 507.18 g/mol
[ Density ](/wiki/Density "Density") | 1.04 g/cm 3 (disodium salt)
[ Melting point ](/wiki/Melting_point "Melting point") | 187 °C (369 °F; 460
K) disodium salt; decomposes
[ Acidity ](/wiki/Acid_dissociation_constant "Acid dissociation constant") (p
_K_ a ) | 0.9, 1.4, 3.8, 6.5
[ UV-vis ](/wiki/Ultraviolet%E2%80%93visible_spectroscopy "Ultraviolet–visible
spectroscopy") (λ max ) | 259 nm [1]
[ Absorbance ](/wiki/Absorbance "Absorbance") | [ ε 259
](/wiki/Molar_attenuation_coefficient "Molar attenuation coefficient") = 15.4
mM −1 cm −1 [1]
Except where otherwise noted, data are given for materials in their [ standard
state ](/wiki/Standard_state "Standard state") (at 25 °C [77 °F], 100 kPa).
 Y [ verify
](https://en.wikipedia.org/w/index.php?title=Special:ComparePages&rev1=477228486&page2=Adenosine+triphosphate)
( [ what is ](/wiki/Wikipedia:WikiProject_Chemicals/Chembox_validation
"Wikipedia:WikiProject Chemicals/Chembox validation")
 Y
 N ?)
[ Infobox references ](/wiki/Wikipedia:Chemical_infobox#References
"Wikipedia:Chemical infobox")
Chemical compound
[ 
](/wiki/File:AdenosineTriphosphate.qutemol.svg) Interactive animation of the
structure of ATP
**Adenosine triphosphate** ( **ATP** ) is a [ nucleotide ](/wiki/Nucleotide
"Nucleotide") [2] that provides [ energy ](/wiki/Energy "Energy") to drive
and support many processes in living [ cells ](/wiki/Cell_\(biology\) "Cell
\(biology\)") , such as [ muscle contraction ](/wiki/Muscle_contraction
"Muscle contraction") , [ nerve impulse ](/wiki/Nerve_impulse "Nerve impulse")
propagation, condensate dissolution, and [ chemical synthesis
](/wiki/Chemical_synthesis "Chemical synthesis") . Found in all known forms of
[ life ](/wiki/Life "Life") , it is often referred to as the "molecular unit
of [ currency ](/wiki/Currency "Currency") " of intracellular [ energy
transfer ](/wiki/Energy_transfer "Energy transfer") . [3]
When consumed in a [ metabolic ](/wiki/Metabolism "Metabolism") processes, ATP
converts either to [ adenosine diphosphate ](/wiki/Adenosine_diphosphate
"Adenosine diphosphate") (ADP) or to [ adenosine monophosphate
](/wiki/Adenosine_monophosphate "Adenosine monophosphate") (AMP). Other
processes regenerate ATP. It is also a [ precursor
](/wiki/Precursor_\(chemistry\) "Precursor \(chemistry\)") to [ DNA
](/wiki/DNA "DNA") and [ RNA ](/wiki/RNA "RNA") , and is used as a [ coenzyme
](/wiki/Coenzyme "Coenzyme") . An average human adult processes around 50
kilograms daily. [4]
From the perspective of [ biochemistry ](/wiki/Biochemistry "Biochemistry") ,
ATP is classified as a [ nucleoside triphosphate
](/wiki/Nucleoside_triphosphate "Nucleoside triphosphate") , which indicates
that it consists of three components: a nitrogenous base ( [ adenine
](/wiki/Adenine "Adenine") ), the sugar [ ribose ](/wiki/Ribose "Ribose") ,
and the [ triphosphate ](/wiki/Polyphosphate "Polyphosphate") .
## Structure [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=1 "Edit
section: Structure") ]
ATP consists of an [ adenine ](/wiki/Adenine "Adenine") attached by the
9-nitrogen atom to the 1′ [ carbon ](/wiki/Carbon "Carbon") [ atom
](/wiki/Atom "Atom") of a sugar ( [ ribose ](/wiki/Ribose "Ribose") ), which
in turn is attached at the 5' carbon atom of the sugar to a triphosphate
group. In its many reactions related to metabolism, the adenine and sugar
groups remain unchanged, but the triphosphate is converted to di- and
monophosphate, giving respectively the derivatives [ ADP
](/wiki/Adenosine_diphosphate "Adenosine diphosphate") and [ AMP
](/wiki/Adenosine_monophosphate "Adenosine monophosphate") . The three
phosphoryl groups are labeled as alpha (α), beta (β), and, for the terminal
phosphate, gamma (γ). [5]
In neutral solution, ionized ATP exists mostly as ATP 4− , with a small
proportion of ATP 3− . [6]
### Metal cation binding [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=2 "Edit
section: Metal cation binding") ]
Polyanionic and featuring a potentially [ chelating ](/wiki/Chelation
"Chelation") polyphosphate group, ATP binds metal cations with high affinity.
The [ binding constant ](/wiki/Binding_constant "Binding constant") for [ Mg
2+
](/wiki/Magnesium "Magnesium") is ( 9 554 ). [7] The binding of a [
divalent ](/wiki/Divalent "Divalent") [ cation ](/wiki/Cation "Cation") ,
almost always [ magnesium ](/wiki/Magnesium "Magnesium") , strongly affects
the interaction of ATP with various proteins. Due to the strength of the ATP-
Mg 2+ interaction, ATP exists in the cell mostly as a complex with Mg 2+
bonded to the phosphate oxygen centers. [6] [8]
A second magnesium ion is critical for ATP binding in the kinase domain. [9]
The presence of Mg 2+ regulates kinase activity. [10] It is interesting
from an RNA world perspective that ATP can carry a Mg ion which catalyzes RNA
polymerization. [ _[ citation needed ](/wiki/Wikipedia:Citation_needed
"Wikipedia:Citation needed") _ ]
## Chemical properties [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=3 "Edit
section: Chemical properties") ]
Salts of ATP can be isolated as colorless solids. [11]
[  ](/wiki/File:ATP-ADP.svg) The cycles of synthesis and
degradation of ATP; 2 and 1 represent input and output of energy,
respectively.
ATP is stable in aqueous solutions between [ pH ](/wiki/PH "PH") 6.8 and 7.4
(in the absence of catalysts). At more extreme pH levels, it rapidly [
hydrolyses ](/wiki/Hydrolyses "Hydrolyses") to ADP and phosphate. Living cells
maintain the ratio of ATP to ADP at a point ten orders of magnitude from
equilibrium, with ATP concentrations fivefold higher than the concentration of
ADP. [12] [13] In the context of biochemical reactions, the P-O-P bonds are
frequently referred to as [ _high-energy bonds_ ](/wiki/High-energy_phosphate
"High-energy phosphate") . [14]
## Reactive aspects [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=4 "Edit
section: Reactive aspects") ]
The hydrolysis of ATP into ADP and inorganic phosphate
ATP 4- (aq) + H 2 O (l) = ADP 3- (aq) + HPO 2- (aq) + H \+ (aq)
releases 20.5 kilojoules per mole (4.9 kcal/mol) of [ enthalpy
](/wiki/Enthalpy "Enthalpy") . This may differ under physiological conditions
if the reactant and products are not exactly in these ionization states. [15]
The values of the free energy released by cleaving either a phosphate (P i )
or a pyrophosphate (PP i ) unit from ATP at [ standard state
](/wiki/Standard_state "Standard state") concentrations of 1 mol/L at pH 7
are: [16]
ATP + H
2 O → ADP + P i Δ _G_ °' = −30.5 kJ/mol (−7.3 kcal/mol)
ATP + H
2 O → AMP + PP i Δ _G_ °' = −45.6 kJ/mol (−10.9 kcal/mol)
These abbreviated equations at a pH near 7 can be written more explicitly (R =
[ adenosyl ](/wiki/Adenosine "Adenosine") ):
[RO-P(O) 2 -O-P(O) 2 -O-PO 3 ] 4− \+ H
2 O → [RO-P(O) 2 -O-PO 3 ] 3− \+ [HPO 4 ] 2− \+ H \+
[RO-P(O) 2 -O-P(O) 2 -O-PO 3 ] 4− \+ H
2 O → [RO-PO 3 ] 2− \+ [HO 3 P-O-PO 3 ] 3− \+ H \+
At cytoplasmic conditions, where the ADP/ATP ratio is 10 orders of magnitude
from equilibrium, the Δ _G_ is around −57 kJ/mol. [12]
Along with pH, the free energy change of ATP hydrolysis is also associated
with Mg 2+ concentration, from ΔG°' = −35.7 kJ/mol at a Mg 2+
concentration of zero, to ΔG°' = −31 kJ/mol at [Mg 2+ ] = 5 mM. Higher
concentrations of Mg 2+ decrease free energy released in the reaction due to
binding of Mg 2+ ions to negatively charged oxygen atoms of ATP at pH 7.
[17]
[
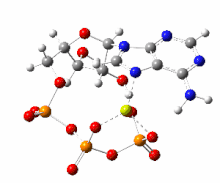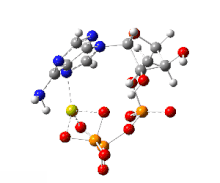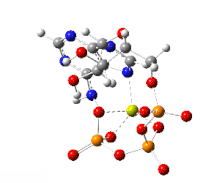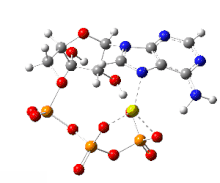 ](/wiki/File:MgATP2-small.gif) This image shows a 360-degree
rotation of a single, gas-phase [ magnesium ](/wiki/Magnesium "Magnesium")
-ATP chelate with a charge of −2. The anion was optimized at the
UB3LYP/6-311++G(d,p) theoretical level and the atomic connectivity modified by
the human optimizer to reflect the probable electronic structure.
## Production from AMP and ADP [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=5 "Edit
section: Production from AMP and ADP") ]
### Production, aerobic conditions [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=6 "Edit
section: Production, aerobic conditions") ]
A typical intracellular [ concentration ](/wiki/Concentration "Concentration")
of ATP may be 1–10 μmol per gram of tissue in a variety of eukaryotes. [18]
The dephosphorylation of ATP and rephosphorylation of ADP and AMP occur
repeatedly in the course of aerobic metabolism. [19]
ATP can be produced by a number of distinct cellular processes; the three main
pathways in [ eukaryotes ](/wiki/Eukaryote "Eukaryote") are (1) [ glycolysis
](/wiki/Glycolysis "Glycolysis") , (2) the [ citric acid cycle
](/wiki/Citric_acid_cycle "Citric acid cycle") / [ oxidative phosphorylation
](/wiki/Oxidative_phosphorylation "Oxidative phosphorylation") , and (3) [
beta-oxidation ](/wiki/Beta-oxidation "Beta-oxidation") . The overall process
of oxidizing [ glucose ](/wiki/Glucose "Glucose") to [ carbon dioxide
](/wiki/Carbon_dioxide "Carbon dioxide") , the combination of pathways 1 and
2, known as [ cellular respiration ](/wiki/Cellular_respiration "Cellular
respiration") , produces about 30 equivalents of ATP from each molecule of
glucose. [20]
ATP production by a non- [ photosynthetic ](/wiki/Photosynthetic
"Photosynthetic") aerobic eukaryote occurs mainly in the [ mitochondria
](/wiki/Mitochondria "Mitochondria") , which comprise nearly 25% of the volume
of a typical cell. [21]
#### Glycolysis [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=7 "Edit
section: Glycolysis") ]
Main article: [ Glycolysis ](/wiki/Glycolysis "Glycolysis")
In glycolysis, glucose and glycerol are metabolized to [ pyruvate
](/wiki/Pyruvate "Pyruvate") . Glycolysis generates two equivalents of ATP
through [ substrate phosphorylation ](/wiki/Substrate-level_phosphorylation
"Substrate-level phosphorylation") catalyzed by two enzymes, [
phosphoglycerate kinase ](/wiki/Phosphoglycerate_kinase "Phosphoglycerate
kinase") (PGK) and [ pyruvate kinase ](/wiki/Pyruvate_kinase "Pyruvate
kinase") . Two equivalents of [ nicotinamide adenine dinucleotide
](/wiki/Nicotinamide_adenine_dinucleotide "Nicotinamide adenine dinucleotide")
(NADH) are also produced, which can be oxidized via the [ electron transport
chain ](/wiki/Electron_transport_chain "Electron transport chain") and result
in the generation of additional ATP by [ ATP synthase ](/wiki/ATP_synthase
"ATP synthase") . The pyruvate generated as an end-product of glycolysis is a
substrate for the [ Krebs Cycle ](/wiki/Citric_acid_cycle "Citric acid cycle")
. [22]
Glycolysis is viewed as consisting of two phases with five steps each. In
phase 1, "the preparatory phase", glucose is converted to 2
d-glyceraldehyde-3-phosphate (g3p). One ATP is invested in Step 1, and another
ATP is invested in Step 3. Steps 1 and 3 of glycolysis are referred to as
"Priming Steps". In Phase 2, two equivalents of g3p are converted to two
pyruvates. In Step 7, two ATP are produced. Also, in Step 10, two further
equivalents of ATP are produced. In Steps 7 and 10, ATP is generated from ADP.
A net of two ATPs is formed in the glycolysis cycle. The glycolysis pathway is
later associated with the Citric Acid Cycle which produces additional
equivalents of ATP. [ _[ citation needed ](/wiki/Wikipedia:Citation_needed
"Wikipedia:Citation needed") _ ]
##### Regulation [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=8 "Edit
section: Regulation") ]
In glycolysis, [ hexokinase ](/wiki/Hexokinase "Hexokinase") is directly
inhibited by its product, glucose-6-phosphate, and [ pyruvate kinase
](/wiki/Pyruvate_kinase "Pyruvate kinase") is inhibited by ATP itself. The
main control point for the glycolytic pathway is [ phosphofructokinase
](/wiki/Phosphofructokinase "Phosphofructokinase") (PFK), which is
allosterically inhibited by high concentrations of ATP and activated by high
concentrations of AMP. The inhibition of PFK by ATP is unusual since ATP is
also a substrate in the reaction catalyzed by PFK; the active form of the
enzyme is a [ tetramer ](/wiki/Tetramer_protein "Tetramer protein") that
exists in two conformations, only one of which binds the second substrate
fructose-6-phosphate (F6P). The protein has two [ binding sites
](/wiki/Binding_site "Binding site") for ATP – the [ active site
](/wiki/Active_site "Active site") is accessible in either protein
conformation, but ATP binding to the inhibitor site stabilizes the
conformation that binds F6P poorly. [22] A number of other small molecules
can compensate for the ATP-induced shift in equilibrium conformation and
reactivate PFK, including [ cyclic AMP ](/wiki/Cyclic_AMP "Cyclic AMP") , [
ammonium ](/wiki/Ammonium "Ammonium") ions, inorganic phosphate, and
fructose-1,6- and -2,6-biphosphate. [22]
#### Citric acid cycle [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=9 "Edit
section: Citric acid cycle") ]
Main articles: [ Citric acid cycle ](/wiki/Citric_acid_cycle "Citric acid
cycle") and [ Oxidative phosphorylation ](/wiki/Oxidative_phosphorylation
"Oxidative phosphorylation")
In the [ mitochondrion ](/wiki/Mitochondrion "Mitochondrion") , pyruvate is
oxidized by the [ pyruvate dehydrogenase complex
](/wiki/Pyruvate_dehydrogenase_complex "Pyruvate dehydrogenase complex") to
the [ acetyl ](/wiki/Acetyl "Acetyl") group, which is fully oxidized to carbon
dioxide by the [ citric acid cycle ](/wiki/Citric_acid_cycle "Citric acid
cycle") (also known as the [ Krebs ](/wiki/Hans_Krebs_\(biochemist\) "Hans
Krebs \(biochemist\)") cycle). Every "turn" of the citric acid cycle produces
two molecules of carbon dioxide, one equivalent of ATP [ guanosine
triphosphate ](/wiki/Guanosine_triphosphate "Guanosine triphosphate") (GTP)
through [ substrate-level phosphorylation ](/wiki/Substrate-
level_phosphorylation "Substrate-level phosphorylation") catalyzed by [
succinyl-CoA synthetase ](/wiki/Succinyl-CoA_synthetase "Succinyl-CoA
synthetase") , as succinyl-CoA is converted to succinate, three equivalents of
NADH, and one equivalent of [ FADH 2 ](/wiki/Flavin_group "Flavin group") .
NADH and FADH 2 are recycled (to NAD \+ and [ FAD
](/wiki/Flavin_adenine_dinucleotide "Flavin adenine dinucleotide") ,
respectively) by [ oxidative phosphorylation ](/wiki/Oxidative_phosphorylation
"Oxidative phosphorylation") , generating additional ATP. The oxidation of
NADH results in the synthesis of 2–3 equivalents of ATP, and the oxidation of
one FADH 2 yields between 1–2 equivalents of ATP. [20] The majority of
cellular ATP is generated by this process. Although the citric acid cycle
itself does not involve molecular [ oxygen ](/wiki/Oxygen "Oxygen") , it is an
obligately [ aerobic ](/wiki/Aerobic_glycolysis "Aerobic glycolysis") process
because O 2 is used to recycle the NADH and FADH 2 . In the absence of
oxygen, the citric acid cycle ceases. [21]
The generation of ATP by the mitochondrion from cytosolic NADH relies on the [
malate-aspartate shuttle ](/wiki/Malate-aspartate_shuttle "Malate-aspartate
shuttle") (and to a lesser extent, the [ glycerol-phosphate shuttle
](/wiki/Glycerol-phosphate_shuttle "Glycerol-phosphate shuttle") ) because the
inner mitochondrial membrane is impermeable to NADH and NAD \+ . Instead of
transferring the generated NADH, a [ malate dehydrogenase
](/wiki/Malate_dehydrogenase "Malate dehydrogenase") enzyme converts [
oxaloacetate ](/wiki/Oxaloacetate "Oxaloacetate") to [ malate ](/wiki/Malate
"Malate") , which is translocated to the mitochondrial matrix. Another malate
dehydrogenase-catalyzed reaction occurs in the opposite direction, producing
oxaloacetate and NADH from the newly transported malate and the
mitochondrion's interior store of NAD \+ . A [ transaminase
](/wiki/Transaminase "Transaminase") converts the oxaloacetate to [ aspartate
](/wiki/Aspartate "Aspartate") for transport back across the membrane and into
the intermembrane space. [21]
In oxidative phosphorylation, the passage of electrons from NADH and FADH 2
through the electron transport chain releases the energy to pump [ protons
](/wiki/Proton "Proton") out of the mitochondrial matrix and into the
intermembrane space. This pumping generates a [ proton motive force
](/wiki/Proton_motive_force "Proton motive force") that is the net effect of a
pH gradient and an [ electric potential ](/wiki/Electric_potential "Electric
potential") gradient across the inner mitochondrial membrane. Flow of protons
down this potential gradient – that is, from the intermembrane space to the
matrix – yields ATP by ATP synthase. [23] Three ATP are produced per turn.
Although oxygen consumption appears fundamental for the maintenance of the
proton motive force, in the event of oxygen shortage ( [ hypoxia
](/wiki/Hypoxia_\(medical\) "Hypoxia \(medical\)") ), intracellular acidosis
(mediated by enhanced glycolytic rates and [ ATP hydrolysis
](/wiki/ATP_hydrolysis "ATP hydrolysis") ), contributes to mitochondrial
membrane potential and directly drives ATP synthesis. [24]
Most of the ATP synthesized in the mitochondria will be used for cellular
processes in the cytosol; thus it must be exported from its site of synthesis
in the mitochondrial matrix. ATP outward movement is favored by the membrane's
electrochemical potential because the cytosol has a relatively positive charge
compared to the relatively negative matrix. For every ATP transported out, it
costs 1 H \+ . Producing one ATP costs about 3 H \+ . Therefore, making
and exporting one ATP requires 4H +. The inner membrane contains an [
antiporter ](/wiki/Antiporter "Antiporter") , the ADP/ATP translocase, which
is an [ integral membrane protein ](/wiki/Integral_membrane_protein "Integral
membrane protein") used to exchange newly synthesized ATP in the matrix for
ADP in the intermembrane space. [25]
##### Regulation [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=10 "Edit
section: Regulation") ]
The citric acid cycle is regulated mainly by the availability of key
substrates, particularly the ratio of NAD \+ to NADH and the concentrations
of [ calcium ](/wiki/Calcium "Calcium") , inorganic phosphate, ATP, ADP, and
AMP. [ Citrate ](/wiki/Citrate "Citrate") – the ion that gives its name to the
cycle – is a feedback inhibitor of [ citrate synthase ](/wiki/Citrate_synthase
"Citrate synthase") and also inhibits PFK, providing a direct link between the
regulation of the citric acid cycle and glycolysis. [22]
#### Beta oxidation [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=11 "Edit
section: Beta oxidation") ]
Main article: [ Beta-oxidation ](/wiki/Beta-oxidation "Beta-oxidation")
In the presence of air and various cofactors and enzymes, fatty acids are
converted to [ acetyl-CoA ](/wiki/Acetyl-CoA "Acetyl-CoA") . The pathway is
called [ beta-oxidation ](/wiki/Beta-oxidation "Beta-oxidation") . Each cycle
of beta-oxidation shortens the fatty acid chain by two carbon atoms and
produces one equivalent each of acetyl-CoA, NADH, and FADH 2 . The acetyl-
CoA is metabolized by the citric acid cycle to generate ATP, while the NADH
and FADH 2 are used by oxidative phosphorylation to generate ATP. Dozens of
ATP equivalents are generated by the beta-oxidation of a single long acyl
chain. [26]
##### Regulation [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=12 "Edit
section: Regulation") ]
In oxidative phosphorylation, the key control point is the reaction catalyzed
by [ cytochrome c oxidase ](/wiki/Cytochrome_c_oxidase "Cytochrome c oxidase")
, which is regulated by the availability of its substrate – the reduced form
of [ cytochrome c ](/wiki/Cytochrome_c "Cytochrome c") . The amount of reduced
cytochrome c available is directly related to the amounts of other substrates:
1 2 NADH \+ cyt c ox \+ ADP \+ P i ⇌ 1 2 NAD \+ \+ cyt c red \+ ATP {\displaystyle {\frac {1}{2}}{\ce {NADH}}+{\ce {cyt}}\ {\ce {c_{ox}}}+{\ce {ADP}}+{\ce {P_{i}}}\rightleftharpoons {\frac {1}{2}}{\ce {NAD^+}}+{\ce {cyt}}\ {\ce {c_{red}}}+{\ce {ATP}}} 
which directly implies this equation:
[ c y t c r e d ] [ c y t c o x ] = ( [ N A D H ] [ N A D ] \+ ) 1 2 ( [ A D P ] [ P i ] [ A T P ] ) K e q {\displaystyle {\frac {[\mathrm {cyt~c_{red}} ]}{[\mathrm {cyt~c_{ox}} ]}}=\left({\frac {[\mathrm {NADH} ]}{[\mathrm {NAD} ]^{+}}}\right)^{\frac {1}{2}}\left({\frac {[\mathrm {ADP} ][\mathrm {P_{i}} ]}{[\mathrm {ATP} ]}}\right)K_{\mathrm {eq} }} ![{\\displaystyle {\\frac {\[\\mathrm {cyt~c_{red}} \]}{\[\\mathrm {cyt~c_{ox}} \]}}=\\left\({\\frac {\[\\mathrm {NADH} \]}{\[\\mathrm {NAD} \]^{+}}}\\right\)^{\\frac {1}{2}}\\left\({\\frac {\[\\mathrm {ADP} \]\[\\mathrm {P_{i}} \]}{\[\\mathrm {ATP} \]}}\\right\)K_{\\mathrm {eq} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/baed008d658c507750599ae155a9a1c11da873e7)
Thus, a high ratio of [NADH] to [NAD \+ ] or a high ratio of [ADP] [P i ]
to [ATP] imply a high amount of reduced cytochrome c and a high level of
cytochrome c oxidase activity. [22] An additional level of regulation is
introduced by the transport rates of ATP and NADH between the mitochondrial
matrix and the cytoplasm. [25]
#### Ketosis [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=13 "Edit
section: Ketosis") ]
Main article: [ Ketone bodies ](/wiki/Ketone_bodies "Ketone bodies")
Ketone bodies can be used as fuels, yielding 22 ATP and 2 [ GTP
](/wiki/Guanosine_triphosphate "Guanosine triphosphate") molecules per
acetoacetate molecule when oxidized in the mitochondria. Ketone bodies are
transported from the [ liver ](/wiki/Liver "Liver") to other tissues, where [
acetoacetate ](/wiki/Acetoacetate "Acetoacetate") and [ _beta_
-hydroxybutyrate ](/wiki/Beta-Hydroxybutyric_acid "Beta-Hydroxybutyric acid")
can be reconverted to acetyl-CoA to produce reducing equivalents (NADH and
FADH 2 ), via the citric acid cycle. Ketone bodies cannot be used as fuel by
the liver, because the liver lacks the enzyme β-ketoacyl-CoA transferase, also
called [ thiolase ](/wiki/Thiolase "Thiolase") . [ Acetoacetate
](/wiki/Acetoacetate "Acetoacetate") in low concentrations is taken up by the
liver and undergoes detoxification through the methylglyoxal pathway which
ends with lactate. Acetoacetate in high concentrations is absorbed by cells
other than those in the liver and enters a different pathway via [
1,2-propanediol ](/wiki/1,2-propanediol "1,2-propanediol") . Though the
pathway follows a different series of steps requiring ATP, 1,2-propanediol can
be turned into pyruvate. [27]
### Production, anaerobic conditions [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=14 "Edit
section: Production, anaerobic conditions") ]
[ Fermentation ](/wiki/Fermentation_\(biochemistry\) "Fermentation
\(biochemistry\)") is the metabolism of organic compounds in the absence of
air. It involves [ substrate-level phosphorylation ](/wiki/Substrate-
level_phosphorylation "Substrate-level phosphorylation") in the absence of a
respiratory [ electron transport chain ](/wiki/Electron_transport_chain
"Electron transport chain") . The equation for the reaction of glucose to form
[ lactic acid ](/wiki/Lactic_acid "Lactic acid") is:
C
6 H
12 O
6 \+ 2 ADP + 2 P i → 2 CH
3 CH(OH)COOH \+ 2 ATP + 2 H
2 O
[ Anaerobic respiration ](/wiki/Anaerobic_respiration "Anaerobic respiration")
is respiration in the absence of [ O
2 ](/wiki/Oxygen "Oxygen") . Prokaryotes can utilize a variety of electron
acceptors. These include [ nitrate ](/wiki/Nitrate "Nitrate") , [ sulfate
](/wiki/Sulfate "Sulfate") , and carbon dioxide.
#### ATP replenishment by nucleoside diphosphate kinases [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=15 "Edit
section: ATP replenishment by nucleoside diphosphate kinases") ]
ATP can also be synthesized through several so-called "replenishment"
reactions catalyzed by the enzyme families of [ nucleoside diphosphate kinases
](/wiki/Nucleoside_diphosphate_kinase "Nucleoside diphosphate kinase") (NDKs),
which use other nucleoside triphosphates as a high-energy phosphate donor, and
the [ ATP:guanido-phosphotransferase
](/wiki/ATP:guanido_phosphotransferase_family "ATP:guanido phosphotransferase
family") family. [ _[ citation needed ](/wiki/Wikipedia:Citation_needed
"Wikipedia:Citation needed") _ ]
### ATP production during photosynthesis [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=16 "Edit
section: ATP production during photosynthesis") ]
In plants, ATP is synthesized in the [ thylakoid membrane
](/wiki/Thylakoid_membrane "Thylakoid membrane") of the [ chloroplast
](/wiki/Chloroplast "Chloroplast") . The process is called [
photophosphorylation ](/wiki/Photophosphorylation "Photophosphorylation") .
The "machinery" is similar to that in mitochondria except that light energy is
used to pump protons across a membrane to produce a proton-motive force. ATP
synthase then ensues exactly as in oxidative phosphorylation. [28] Some of
the ATP produced in the chloroplasts is consumed in the [ Calvin cycle
](/wiki/Calvin_cycle "Calvin cycle") , which produces [ triose ](/wiki/Triose
"Triose") sugars.
### ATP recycling [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=17 "Edit
section: ATP recycling") ]
The total quantity of ATP in the human body is about 0.1 [ mol/L
](/wiki/Molar_concentration "Molar concentration") . [29] The majority of
ATP is recycled from ADP by the aforementioned processes. Thus, at any given
time, the total amount of ATP + ADP remains fairly constant.
The energy used by human cells in an adult requires the hydrolysis of 100 to
150 mol/L of ATP daily, which means a human will typically use their body
weight worth of ATP over the course of the day. [30] Each equivalent of ATP
is recycled 1000–1500 times during a single day ( 150 / 0.1 = 1500 ), [29]
at approximately 9×10 20 molecules/s. [29]
[  ](/wiki/File:Rossmann-
fold-1g5q.png) An example of the Rossmann fold, a [ structural domain
](/wiki/Structural_domain "Structural domain") of a [ decarboxylase
](/wiki/Decarboxylase "Decarboxylase") enzyme from the bacterium _[
Staphylococcus epidermidis ](/wiki/Staphylococcus_epidermidis "Staphylococcus
epidermidis") _ ( [ PDB ](/wiki/Protein_Data_Bank "Protein Data Bank") : [
1G5Q ](https://www.rcsb.org/structure/1G5Q) ) with a bound [ flavin
mononucleotide ](/wiki/Flavin_mononucleotide "Flavin mononucleotide") cofactor
## Biochemical functions [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=18 "Edit
section: Biochemical functions") ]
### Intracellular signaling [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=19 "Edit
section: Intracellular signaling") ]
ATP is involved in [ signal transduction ](/wiki/Signal_transduction "Signal
transduction") by serving as substrate for kinases, enzymes that transfer
phosphate groups. Kinases are the most common ATP-binding proteins. They share
a small number of common folds. [31] [ Phosphorylation
](/wiki/Phosphorylation "Phosphorylation") of a protein by a kinase can
activate a cascade such as the [ mitogen-activated protein kinase
](/wiki/Mitogen-activated_protein_kinase "Mitogen-activated protein kinase")
cascade. [32]
ATP is also a substrate of [ adenylate cyclase ](/wiki/Adenylate_cyclase
"Adenylate cyclase") , most commonly in [ G protein-coupled receptor
](/wiki/G_protein%E2%80%93coupled_receptor "G protein–coupled receptor")
signal transduction pathways and is transformed to [ second messenger
](/wiki/Second_messenger "Second messenger") , cyclic AMP, which is involved
in triggering calcium signals by the release of calcium from intracellular
stores. [33] This form of signal transduction is particularly important in
brain function, although it is involved in the regulation of a multitude of
other cellular processes. [34]
### DNA and RNA synthesis [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=20 "Edit
section: DNA and RNA synthesis") ]
ATP is one of four monomers required in the synthesis of [ RNA ](/wiki/RNA
"RNA") . The process is promoted by [ RNA polymerases ](/wiki/RNA_polymerase
"RNA polymerase") . [35] A similar process occurs in the formation of DNA,
except that ATP is first converted to the [ deoxyribonucleotide
](/wiki/Deoxyribonucleotide "Deoxyribonucleotide") dATP. Like many
condensation reactions in nature, [ DNA replication ](/wiki/DNA_replication
"DNA replication") and [ DNA transcription ](/wiki/DNA_transcription "DNA
transcription") also consume ATP.
### Amino acid activation in protein synthesis [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=21 "Edit
section: Amino acid activation in protein synthesis") ]
Main article: [ Amino acid activation ](/wiki/Amino_acid_activation "Amino
acid activation")
[ Aminoacyl-tRNA synthetase ](/wiki/Aminoacyl-tRNA_synthetase "Aminoacyl-tRNA
synthetase") enzymes consume ATP in the attachment tRNA to amino acids,
forming aminoacyl-tRNA complexes. Aminoacyl transferase binds AMP-amino acid
to tRNA. The coupling reaction proceeds in two steps:
1. aa + ATP ⟶ aa-AMP + [ PP i ](/wiki/Pyrophosphate "Pyrophosphate")
2. aa-AMP + tRNA ⟶ aa-tRNA + AMP
The amino acid is coupled to the penultimate nucleotide at the 3′-end of the
tRNA (the A in the sequence CCA) via an ester bond (roll over in
illustration).
### ATP binding cassette transporter [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=22 "Edit
section: ATP binding cassette transporter") ]
Transporting chemicals out of a cell against a gradient is often associated
with ATP hydrolysis. Transport is mediated by [ ATP binding cassette
transporters ](/wiki/ATP_binding_cassette_transporter "ATP binding cassette
transporter") . The human genome encodes 48 ABC transporters, that are used
for exporting drugs, lipids, and other compounds. [36]
### Extracellular signalling and neurotransmission [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=23 "Edit
section: Extracellular signalling and neurotransmission") ]
Cells secrete ATP to communicate with other cells in a process called [
purinergic signalling ](/wiki/Purinergic_signalling "Purinergic signalling") .
ATP serves as a [ neurotransmitter ](/wiki/Neurotransmitter
"Neurotransmitter") in many parts of the nervous system, modulates ciliary
beating, affects vascular oxygen supply etc. ATP is either secreted directly
across the cell membrane through channel proteins [37] [38] or is pumped
into vesicles [39] which then [ fuse ](/wiki/Exocytosis "Exocytosis") with
the membrane. Cells detect ATP using the [ purinergic receptor
](/wiki/Purinergic_receptor "Purinergic receptor") proteins P2X and P2Y.
### Protein solubility [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=24 "Edit
section: Protein solubility") ]
ATP has recently been proposed to act as a biological [ hydrotrope
](/wiki/Hydrotrope "Hydrotrope") [40] and has been shown to affect proteome-
wide solubility. [41]
## Abiogenic origins [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=25 "Edit
section: Abiogenic origins") ]
Acetyl phosphate (AcP), a precursor to ATP, can readily be synthesized at
modest yields from thioacetate in pH 7 and 20 °C and pH 8 and 50 °C, although
acetyl phosphate is less stable in warmer temperatures and alkaline conditions
than in cooler and acidic to neutral conditions. It is unable to promote
polymerization of ribonucleotides and amino acids and was only capable of
phosphorylation of organic compounds. It was shown that it can promote
aggregation and stabilization of AMP in the presence of Na \+ , aggregation
of nucleotides could promote polymerization above 75 °C in the absence of Na
\+ . It is possible that polymerization promoted by AcP could occur at
mineral surfaces. [42] It was shown that ADP can only be phosphorylated to
ATP by AcP and other nucleoside triphosphates were not phosphorylated by AcP.
This might explain why all lifeforms use ATP to drive biochemical reactions.
[43]
## ATP analogues [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=26 "Edit
section: ATP analogues") ]
Biochemistry laboratories often use _[ in vitro ](/wiki/In_vitro "In vitro") _
studies to explore ATP-dependent molecular processes. ATP analogs are also
used in [ X-ray crystallography ](/wiki/X-ray_crystallography "X-ray
crystallography") to determine a [ protein structure ](/wiki/Protein_structure
"Protein structure") in complex with ATP, often together with other
substrates. [ _[ citation needed ](/wiki/Wikipedia:Citation_needed
"Wikipedia:Citation needed") _ ]
[ Enzyme inhibitors ](/wiki/Enzyme_inhibitor "Enzyme inhibitor") of ATP-
dependent enzymes such as [ kinases ](/wiki/Kinase "Kinase") are needed to
examine the [ binding sites ](/wiki/Binding_site "Binding site") and [
transition states ](/wiki/Transition_state "Transition state") involved in
ATP-dependent reactions. [ _[ citation needed
](/wiki/Wikipedia:Citation_needed "Wikipedia:Citation needed") _ ]
Most useful ATP analogs cannot be hydrolyzed as ATP would be; instead, they
trap the enzyme in a structure closely related to the ATP-bound state.
Adenosine 5′-(γ-thiotriphosphate) is an extremely common ATP analog in which
one of the gamma-phosphate oxygens is replaced by a [ sulfur ](/wiki/Sulfur
"Sulfur") atom; this anion is hydrolyzed at a dramatically slower rate than
ATP itself and functions as an inhibitor of ATP-dependent processes. In
crystallographic studies, hydrolysis transition states are modeled by the
bound [ vanadate ](/wiki/Vanadate "Vanadate") ion.
Caution is warranted in interpreting the results of experiments using ATP
analogs, since some enzymes can hydrolyze them at appreciable rates at high
concentration. [44]
## Medical use [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=27 "Edit
section: Medical use") ]
ATP is used intravenously for some heart related conditions. [45]
## History [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=28 "Edit
section: History") ]
ATP was discovered in 1929 by [ Karl Lohmann
](https://de.wikipedia.org/wiki/Karl_Lohmann_\(Biochemiker\) "de:Karl Lohmann
\(Biochemiker\)") [46] and Jendrassik [47] and, independently, by Cyrus
Fiske and [ Yellapragada Subba Rao ](/wiki/Yellapragada_Subba_Rao
"Yellapragada Subba Rao") of [ Harvard Medical School
](/wiki/Harvard_Medical_School "Harvard Medical School") , [48] both teams
competing against each other to find an assay for phosphorus.
It was proposed to be the intermediary between energy-yielding and energy-
requiring reactions in cells by [ Fritz Albert Lipmann
](/wiki/Fritz_Albert_Lipmann "Fritz Albert Lipmann") in 1941. [49]
It was first synthesized in the laboratory by [ Alexander Todd
](/wiki/Alexander_R._Todd,_Baron_Todd "Alexander R. Todd, Baron Todd") in
1948, [50] and he was awarded the [ Nobel Prize in Chemistry
](/wiki/Nobel_Prize_in_Chemistry "Nobel Prize in Chemistry") in 1957 partly
for this work.
The 1978 [ Nobel Prize in Chemistry ](/wiki/Nobel_Prize_in_Chemistry "Nobel
Prize in Chemistry") was awarded to [ Peter Dennis Mitchell
](/wiki/Peter_D._Mitchell "Peter D. Mitchell") for the discovery of the [
chemiosmotic ](/wiki/Chemiosmosis "Chemiosmosis") mechanism of ATP synthesis.
The 1997 Nobel Prize in Chemistry was divided, one half jointly to [ Paul D.
Boyer ](/wiki/Paul_D._Boyer "Paul D. Boyer") and [ John E. Walker
](/wiki/John_E._Walker "John E. Walker") "for their elucidation of the
enzymatic mechanism underlying the synthesis of adenosine triphosphate (ATP)"
and the other half to [ Jens C. Skou ](/wiki/Jens_C._Skou "Jens C. Skou") "for
the first discovery of an ion-transporting enzyme, Na \+ , K \+ -ATPase."
[51]
## See also [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=29 "Edit
section: See also") ]
* [ Adenosine-tetraphosphatase ](/wiki/Adenosine-tetraphosphatase "Adenosine-tetraphosphatase")
* [ Adenosine methylene triphosphate ](/wiki/NDPCP "NDPCP")
* [ ATPases ](/wiki/ATPases "ATPases")
* [ ATP test ](/wiki/ATP_test "ATP test")
* [ Creatine ](/wiki/Creatine "Creatine")
* [ Cyclic adenosine monophosphate ](/wiki/Cyclic_adenosine_monophosphate "Cyclic adenosine monophosphate") (cAMP)
* [ Nucleotide exchange factor ](/wiki/Nucleotide_exchange_factor "Nucleotide exchange factor")
* [ Phosphagen ](/wiki/Phosphagen "Phosphagen")
## References [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=30 "Edit
section: References") ]
1. ^ _**a** _ _**b** _ [ "Adenosine 5'-triphosphate disodium salt Product Information" ](https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Product_Information_Sheet/a7699pis.pdf) (PDF) . Sigma. [ Archived ](https://web.archive.org/web/20190323054630/https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Product_Information_Sheet/a7699pis.pdf) (PDF) from the original on 2019-03-23 . Retrieved 2019-03-22 .
2. ** ^ ** Dunn, Jacob; Grider, Michael H. (2023). [ "Physiology, Adenosine Triphosphate" ](https://www.ncbi.nlm.nih.gov/books/NBK553175/) . _StatPearls_ . StatPearls Publishing. [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 31985968 ](https://pubmed.ncbi.nlm.nih.gov/31985968) . Retrieved 13 November 2023 .
3. ** ^ ** Knowles, J. R. (1980). "Enzyme-catalyzed phosphoryl transfer reactions". _Annu. Rev. Biochem_ . **49** : 877–919. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1146/annurev.bi.49.070180.004305 ](https://doi.org/10.1146%2Fannurev.bi.49.070180.004305) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 6250450 ](https://pubmed.ncbi.nlm.nih.gov/6250450) .
4. ** ^ ** "An average individual with a daily diet of 8000 kJ and a 30% efficiency of turning foodstuffs into chemical energy will synthesize (and hydrolyze) about 50 kg of ATP during 1 day." Wilkens, Stephan (2008). "<SCP>ATP</SCP> Synthesis, Chemistry of". _Wiley Encyclopedia of Chemical Biology_ . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1002/9780470048672.wecb648 ](https://doi.org/10.1002%2F9780470048672.wecb648) . [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 9780471754770 ](/wiki/Special:BookSources/9780471754770 "Special:BookSources/9780471754770") .
5. ** ^ ** Dunn, Jacob; Grider, Michael H. (2023), [ "Physiology, Adenosine Triphosphate" ](http://www.ncbi.nlm.nih.gov/books/NBK553175/) , _StatPearls_ , Treasure Island (FL): StatPearls Publishing, [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 31985968 ](https://pubmed.ncbi.nlm.nih.gov/31985968) , retrieved 2023-09-28
6. ^ _**a** _ _**b** _ Storer, A.; [ Cornish-Bowden, A. ](/wiki/Athel_Cornish-Bowden "Athel Cornish-Bowden") (1976). [ "Concentration of MgATP 2− and other ions in solution. Calculation of the true concentrations of species present in mixtures of associating ions" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1164030) . _Biochem. J_ . **159** (1): 1–5. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1042/bj1590001 ](https://doi.org/10.1042%2Fbj1590001) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 1164030 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1164030) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 11772 ](https://pubmed.ncbi.nlm.nih.gov/11772) .
7. ** ^ ** Wilson, J.; Chin, A. (1991). "Chelation of divalent cations by ATP, studied by titration calorimetry". _Anal. Biochem_ . **193** (1): 16–19. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/0003-2697(91)90036-S ](https://doi.org/10.1016%2F0003-2697%2891%2990036-S) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 1645933 ](https://pubmed.ncbi.nlm.nih.gov/1645933) .
8. ** ^ ** Garfinkel, L.; Altschuld, R.; Garfinkel, D. (1986). "Magnesium in cardiac energy metabolism". _J. Mol. Cell. Cardiol_ . **18** (10): 1003–1013. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/S0022-2828(86)80289-9 ](https://doi.org/10.1016%2FS0022-2828%2886%2980289-9) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 3537318 ](https://pubmed.ncbi.nlm.nih.gov/3537318) .
9. ** ^ ** Saylor, P.; Wang, C.; Hirai, T.; Adams, J. (1998). "A second magnesium ion is critical for ATP binding in the kinase domain of the oncoprotein v-Fps". _Biochemistry_ . **37** (36): 12624–12630. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1021/bi9812672 ](https://doi.org/10.1021%2Fbi9812672) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 9730835 ](https://pubmed.ncbi.nlm.nih.gov/9730835) .
10. ** ^ ** Lin, X.; Ayrapetov, M; Sun, G. (2005). [ "Characterization of the interactions between the active site of a protein tyrosine kinase and a divalent metal activator" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1316873) . _BMC Biochem_ . **6** : 25. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1186/1471-2091-6-25 ](https://doi.org/10.1186%2F1471-2091-6-25) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 1316873 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1316873) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 16305747 ](https://pubmed.ncbi.nlm.nih.gov/16305747) .
11. ** ^ ** Budavari, Susan, ed. (2001), _[ The Merck Index: An Encyclopedia of Chemicals, Drugs, and Biologicals ](/wiki/Merck_Index "Merck Index") _ (13th ed.), Merck, [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 0911910131 ](/wiki/Special:BookSources/0911910131 "Special:BookSources/0911910131")
12. ^ _**a** _ _**b** _ Ferguson, S. J.; Nicholls, David; Ferguson, Stuart (2002). _Bioenergetics 3_ (3rd ed.). San Diego, CA: Academic. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-12-518121-1 ](/wiki/Special:BookSources/978-0-12-518121-1 "Special:BookSources/978-0-12-518121-1") .
13. ** ^ ** Berg, J. M.; Tymoczko, J. L.; Stryer, L. (2003). [ _Biochemistry_ ](https://archive.org/details/biochemistry200100jere) . New York, NY: W. H. Freeman. p. [ 376 ](https://archive.org/details/biochemistry200100jere/page/376) . [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-7167-4684-3 ](/wiki/Special:BookSources/978-0-7167-4684-3 "Special:BookSources/978-0-7167-4684-3") .
14. ** ^ ** Chance, B.; Lees, H.; Postgate, J. G. (1972). "The Meaning of "Reversed Electron Flow" and "High Energy Electron" in Biochemistry". _Nature_ . **238** (5363): 330–331. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 1972Natur.238..330C ](https://ui.adsabs.harvard.edu/abs/1972Natur.238..330C) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1038/238330a0 ](https://doi.org/10.1038%2F238330a0) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 4561837 ](https://pubmed.ncbi.nlm.nih.gov/4561837) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 4298762 ](https://api.semanticscholar.org/CorpusID:4298762) .
15. ** ^ ** Gajewski, E.; Steckler, D.; Goldberg, R. (1986). [ "Thermodynamics of the hydrolysis of adenosine 5′-triphosphate to adenosine 5′-diphosphate" ](https://doi.org/10.1016%2FS0021-9258%2818%2967153-4) . _J. Biol. Chem_ . **261** (27): 12733–12737. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/S0021-9258(18)67153-4 ](https://doi.org/10.1016%2FS0021-9258%2818%2967153-4) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 3528161 ](https://pubmed.ncbi.nlm.nih.gov/3528161) .
16. ** ^ ** Berg, Jeremy M.; Tymoczko, John L.; Stryer, Lubert (2007). _Biochemistry_ (6th ed.). New York, NY: W. H. Freeman. p. 413. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-7167-8724-2 ](/wiki/Special:BookSources/978-0-7167-8724-2 "Special:BookSources/978-0-7167-8724-2") .
17. ** ^ ** Garrett, Reginald H.; Grisham, Charles M. (2016). _Biochemistry_ (6th ed.). Cengage Learning. p. 68. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-1305577206 ](/wiki/Special:BookSources/978-1305577206 "Special:BookSources/978-1305577206") .
18. ** ^ ** Beis, I.; Newsholme, E. A. (October 1, 1975). [ "The contents of adenine nucleotides, phosphagens and some glycolytic intermediates in resting muscles from vertebrates and invertebrates" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1172435) . _Biochem. J_ . **152** (1): 23–32. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1042/bj1520023 ](https://doi.org/10.1042%2Fbj1520023) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 1172435 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1172435) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 1212224 ](https://pubmed.ncbi.nlm.nih.gov/1212224) .
19. ** ^ ** [ "Adenosine triphosphate" ](https://www.britannica.com/science/adenosine-triphosphate) . Britannica. 11 November 2023 . Retrieved 1 December 2023 .
20. ^ _**a** _ _**b** _ Rich, P. R. (2003). "The molecular machinery of Keilin's respiratory chain". _Biochem. Soc. Trans_ . **31** (6): 1095–1105. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1042/BST0311095 ](https://doi.org/10.1042%2FBST0311095) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 14641005 ](https://pubmed.ncbi.nlm.nih.gov/14641005) .
21. ^ _**a** _ _**b** _ _**c** _ Lodish, H.; Berk, A.; Matsudaira, P.; Kaiser, C. A.; Krieger, M.; Scott, M. P.; Zipursky, S. L.; Darnell, J. (2004). [ _Molecular Cell Biology_ ](https://archive.org/details/molecularcellbio00harv) (5th ed.). New York, NY: W. H. Freeman. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-7167-4366-8 ](/wiki/Special:BookSources/978-0-7167-4366-8 "Special:BookSources/978-0-7167-4366-8") .
22. ^ _**a** _ _**b** _ _**c** _ _**d** _ _**e** _ Voet, D.; Voet, J. G. (2004). _Biochemistry_ . Vol. 1 (3rd ed.). Hoboken, NJ: Wiley. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-471-19350-0 ](/wiki/Special:BookSources/978-0-471-19350-0 "Special:BookSources/978-0-471-19350-0") .
23. ** ^ ** Abrahams, J.; Leslie, A.; Lutter, R.; Walker, J. (1994). "Structure at 2.8 Å resolution of F1-ATPase from bovine heart mitochondria". _Nature_ . **370** (6491): 621–628. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 1994Natur.370..621A ](https://ui.adsabs.harvard.edu/abs/1994Natur.370..621A) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1038/370621a0 ](https://doi.org/10.1038%2F370621a0) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 8065448 ](https://pubmed.ncbi.nlm.nih.gov/8065448) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 4275221 ](https://api.semanticscholar.org/CorpusID:4275221) .
24. ** ^ ** Devaux, JBL; Hedges, CP; Hickey, AJR (January 2019). [ "Acidosis Maintains the Function of Brain Mitochondria in Hypoxia-Tolerant Triplefin Fish: A Strategy to Survive Acute Hypoxic Exposure?" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6346031) . _Front Physiol_ . 9, 1914: 1941. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.3389/fphys.2018.01941 ](https://doi.org/10.3389%2Ffphys.2018.01941) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 6346031 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6346031) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 30713504 ](https://pubmed.ncbi.nlm.nih.gov/30713504) .
25. ^ _**a** _ _**b** _ Dahout-Gonzalez, C.; Nury, H.; Trézéguet, V.; Lauquin, G.; Pebay-Peyroula, E.; Brandolin, G. (2006). "Molecular, functional, and pathological aspects of the mitochondrial ADP/ATP carrier". _Physiology_ . **21** (4): 242–249. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1152/physiol.00005.2006 ](https://doi.org/10.1152%2Fphysiol.00005.2006) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 16868313 ](https://pubmed.ncbi.nlm.nih.gov/16868313) .
26. ** ^ ** Ronnett, G.; Kim, E.; Landree, L.; Tu, Y. (2005). "Fatty acid metabolism as a target for obesity treatment". _Physiol. Behav_ . **85** (1): 25–35. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/j.physbeh.2005.04.014 ](https://doi.org/10.1016%2Fj.physbeh.2005.04.014) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 15878185 ](https://pubmed.ncbi.nlm.nih.gov/15878185) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 24865576 ](https://api.semanticscholar.org/CorpusID:24865576) .
27. ** ^ ** [ "Integrated Risk Information System" ](http://www.epa.gov/iris/toxreviews/0128tr.pdf) (PDF) . 2013-03-15. [ Archived ](https://web.archive.org/web/20150924074331/http://www.epa.gov/iris/toxreviews/0128tr.pdf) (PDF) from the original on 2015-09-24 . Retrieved 2019-02-01 .
28. ** ^ ** Allen, J. (2002). [ "Photosynthesis of ATP-electrons, proton pumps, rotors, and poise" ](https://doi.org/10.1016%2FS0092-8674%2802%2900870-X) . _Cell_ . **110** (3): 273–276. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/S0092-8674(02)00870-X ](https://doi.org/10.1016%2FS0092-8674%2802%2900870-X) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 12176312 ](https://pubmed.ncbi.nlm.nih.gov/12176312) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 1754660 ](https://api.semanticscholar.org/CorpusID:1754660) .
29. ^ _**a** _ _**b** _ _**c** _ Fuhrman, Bradley P.; Zimmerman, Jerry J. (2011). [ _Pediatric Critical Care_ ](https://www.sciencedirect.com/science/article/pii/B9780323073073100746#s0025) . Elsevier. p. 1061. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-323-07307-3 ](/wiki/Special:BookSources/978-0-323-07307-3 "Special:BookSources/978-0-323-07307-3") . Retrieved 16 May 2020 .
30. ** ^ ** Fuhrman, Bradley P.; Zimmerman, Jerry J. (2011). [ _Pediatric Critical Care_ ](https://www.sciencedirect.com/science/article/pii/B9780323073073100746#s0025) . Elsevier. pp. 1058–1072. [ ISBN ](/wiki/ISBN_\(identifier\) "ISBN \(identifier\)") [ 978-0-323-07307-3 ](/wiki/Special:BookSources/978-0-323-07307-3 "Special:BookSources/978-0-323-07307-3") . Retrieved 16 May 2020 .
31. ** ^ ** Scheeff, E.; Bourne, P. (2005). [ "Structural evolution of the protein kinase-like superfamily" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1261164) . _PLOS Comput. Biol_ . **1** (5): e49. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 2005PLSCB...1...49S ](https://ui.adsabs.harvard.edu/abs/2005PLSCB...1...49S) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1371/journal.pcbi.0010049 ](https://doi.org/10.1371%2Fjournal.pcbi.0010049) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 1261164 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1261164) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 16244704 ](https://pubmed.ncbi.nlm.nih.gov/16244704) .
32. ** ^ ** Mishra, N.; Tuteja, R.; Tuteja, N. (2006). "Signaling through MAP kinase networks in plants". _Arch. Biochem. Biophys_ . **452** (1): 55–68. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/j.abb.2006.05.001 ](https://doi.org/10.1016%2Fj.abb.2006.05.001) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 16806044 ](https://pubmed.ncbi.nlm.nih.gov/16806044) .
33. ** ^ ** Kamenetsky, M.; Middelhaufe, S.; Bank, E.; Levin, L.; Buck, J.; Steegborn, C. (2006). [ "Molecular details of cAMP generation in mammalian cells: a tale of two systems" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3662476) . _J. Mol. Biol_ . **362** (4): 623–639. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1016/j.jmb.2006.07.045 ](https://doi.org/10.1016%2Fj.jmb.2006.07.045) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 3662476 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3662476) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 16934836 ](https://pubmed.ncbi.nlm.nih.gov/16934836) .
34. ** ^ ** Hanoune, J.; Defer, N. (2001). "Regulation and role of adenylyl cyclase isoforms". _Annu. Rev. Pharmacol. Toxicol_ . **41** (1): 145–174. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1146/annurev.pharmtox.41.1.145 ](https://doi.org/10.1146%2Fannurev.pharmtox.41.1.145) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 11264454 ](https://pubmed.ncbi.nlm.nih.gov/11264454) .
35. ** ^ ** Joyce, C. M.; Steitz, T. A. (1995). [ "Polymerase structures and function: variations on a theme?" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC177480) . _J. Bacteriol_ . **177** (22): 6321–6329. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1128/jb.177.22.6321-6329.1995 ](https://doi.org/10.1128%2Fjb.177.22.6321-6329.1995) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 177480 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC177480) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 7592405 ](https://pubmed.ncbi.nlm.nih.gov/7592405) .
36. ** ^ ** Borst, P.; Elferink, R. Oude (2002). [ "Mammalian ABC transporters in health and disease" ](https://pure.uva.nl/ws/files/3499814/42885_202387y.pdf) (PDF) . _Annual Review of Biochemistry_ . **71** : 537–592. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1146/annurev.biochem.71.102301.093055 ](https://doi.org/10.1146%2Fannurev.biochem.71.102301.093055) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 12045106 ](https://pubmed.ncbi.nlm.nih.gov/12045106) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 34707074 ](https://api.semanticscholar.org/CorpusID:34707074) . [ Archived ](https://web.archive.org/web/20180421032744/https://pure.uva.nl/ws/files/3499814/42885_202387y.pdf) (PDF) from the original on 2018-04-21 . Retrieved 2018-04-20 .
37. ** ^ ** Romanov, Roman A.; Lasher, Robert S.; High, Brigit; Savidge, Logan E.; Lawson, Adam; Rogachevskaja, Olga A.; Zhao, Haitian; Rogachevsky, Vadim V.; Bystrova, Marina F.; Churbanov, Gleb D.; Adameyko, Igor; Harkany, Tibor; Yang, Ruibiao; Kidd, Grahame J.; Marambaud, Philippe; Kinnamon, John C.; Kolesnikov, Stanislav S.; Finger, Thomas E. (2018). [ "Chemical synapses without synaptic vesicles: Purinergic neurotransmission through a CALHM1 channel-mitochondrial signaling complex" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5966022) . _Science Signaling_ . **11** (529): eaao1815. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1126/scisignal.aao1815 ](https://doi.org/10.1126%2Fscisignal.aao1815) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 1945-0877 ](https://www.worldcat.org/issn/1945-0877) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 5966022 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5966022) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 29739879 ](https://pubmed.ncbi.nlm.nih.gov/29739879) .
38. ** ^ ** Dahl, Gerhard (2015). [ "ATP release through pannexon channels" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4455760) . _Philosophical Transactions of the Royal Society B: Biological Sciences_ . **370** (1672): 20140191. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1098/rstb.2014.0191 ](https://doi.org/10.1098%2Frstb.2014.0191) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 0962-8436 ](https://www.worldcat.org/issn/0962-8436) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 4455760 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4455760) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 26009770 ](https://pubmed.ncbi.nlm.nih.gov/26009770) .
39. ** ^ ** Larsson, Max; Sawada, Keisuke; Morland, Cecilie; Hiasa, Miki; Ormel, Lasse; Moriyama, Yoshinori; Gundersen, Vidar (2012). [ "Functional and Anatomical Identification of a Vesicular Transporter Mediating Neuronal ATP Release" ](https://doi.org/10.1093%2Fcercor%2Fbhr203) . _Cerebral Cortex_ . **22** (5): 1203–1214. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1093/cercor/bhr203 ](https://doi.org/10.1093%2Fcercor%2Fbhr203) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 1460-2199 ](https://www.worldcat.org/issn/1460-2199) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 21810784 ](https://pubmed.ncbi.nlm.nih.gov/21810784) .
40. ** ^ ** Hyman, Anthony A.; Krishnan, Yamuna; Alberti, Simon; Wang, Jie; Saha, Shambaditya; Malinovska, Liliana; Patel, Avinash (2017-05-19). "ATP as a biological hydrotrope". _Science_ . **356** (6339): 753–756. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 2017Sci...356..753P ](https://ui.adsabs.harvard.edu/abs/2017Sci...356..753P) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1126/science.aaf6846 ](https://doi.org/10.1126%2Fscience.aaf6846) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 0036-8075 ](https://www.worldcat.org/issn/0036-8075) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 28522535 ](https://pubmed.ncbi.nlm.nih.gov/28522535) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 24622983 ](https://api.semanticscholar.org/CorpusID:24622983) .
41. ** ^ ** Savitski, Mikhail M.; Bantscheff, Marcus; Huber, Wolfgang; Dominic Helm; Günthner, Ina; Werner, Thilo; Kurzawa, Nils; Sridharan, Sindhuja (2019-03-11). [ "Proteome-wide solubility and thermal stability profiling reveals distinct regulatory roles for ATP" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6411743) . _Nature Communications_ . **10** (1): 1155. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 2019NatCo..10.1155S ](https://ui.adsabs.harvard.edu/abs/2019NatCo..10.1155S) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1038/s41467-019-09107-y ](https://doi.org/10.1038%2Fs41467-019-09107-y) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 2041-1723 ](https://www.worldcat.org/issn/2041-1723) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 6411743 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6411743) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 30858367 ](https://pubmed.ncbi.nlm.nih.gov/30858367) .
42. ** ^ ** Whicher, Alexandra; Camprubi, Eloi; Pinna, Silvana; Herschy, Barry; Lane, Nick (2018-06-01). [ "Acetyl Phosphate as a Primordial Energy Currency at the Origin of Life" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6061221) . _Origins of Life and Evolution of Biospheres_ . **48** (2): 159–179. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 2018OLEB...48..159W ](https://ui.adsabs.harvard.edu/abs/2018OLEB...48..159W) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1007/s11084-018-9555-8 ](https://doi.org/10.1007%2Fs11084-018-9555-8) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 1573-0875 ](https://www.worldcat.org/issn/1573-0875) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 6061221 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6061221) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 29502283 ](https://pubmed.ncbi.nlm.nih.gov/29502283) .
43. ** ^ ** [ "Ancient chemistry may explain why living things use ATP as the universal energy currency: An early step in metabolic evolution set the stage for emergence of ATP as the universal energy carrier" ](https://www.sciencedaily.com/releases/2022/10/221004151228.htm) . _ScienceDaily_ . Retrieved 2023-08-27 .
44. ** ^ ** Resetar, A. M.; Chalovich, J. M. (1995). "Adenosine 5′-(gamma-thiotriphosphate): an ATP analog that should be used with caution in muscle contraction studies". _Biochemistry_ . **34** (49): 16039–16045. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1021/bi00049a018 ](https://doi.org/10.1021%2Fbi00049a018) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 8519760 ](https://pubmed.ncbi.nlm.nih.gov/8519760) .
45. ** ^ ** Pelleg, Amir; Kutalek, Steven P.; Flammang, Daniel; Benditt, David (February 2012). [ "ATPace: injectable adenosine 5′-triphosphate" ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3265710) . _Purinergic Signalling_ . **8** (Suppl 1): 57–60. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1007/s11302-011-9268-1 ](https://doi.org/10.1007%2Fs11302-011-9268-1) . [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 1573-9538 ](https://www.worldcat.org/issn/1573-9538) . [ PMC ](/wiki/PMC_\(identifier\) "PMC \(identifier\)") [ 3265710 ](https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3265710) . [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 22057692 ](https://pubmed.ncbi.nlm.nih.gov/22057692) .
46. ** ^ ** Lohmann, K. (August 1929). "Über die Pyrophosphatfraktion im Muskel" [On the pyrophosphate fraction in muscle]. _Naturwissenschaften_ (in German). **17** (31): 624–625. [ Bibcode ](/wiki/Bibcode_\(identifier\) "Bibcode \(identifier\)") : [ 1929NW.....17..624. ](https://ui.adsabs.harvard.edu/abs/1929NW.....17..624.) . [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1007/BF01506215 ](https://doi.org/10.1007%2FBF01506215) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 20328411 ](https://api.semanticscholar.org/CorpusID:20328411) .
47. ** ^ ** Vaughan, Martha; Hill, Robert L.; Simoni, Robert D. (2002). [ "The Determination of Phosphorus and the Discovery of Phosphocreatine and ATP: the Work of Fiske and SubbaRow" ](http://www.jbc.org/content/277/32/e21) . _Journal of Biological Chemistry_ . **277** (32): e21. [ PMID ](/wiki/PMID_\(identifier\) "PMID \(identifier\)") [ 12161449 ](https://pubmed.ncbi.nlm.nih.gov/12161449) . [ Archived ](https://web.archive.org/web/20170808062708/http://www.jbc.org/content/277/32/e21) from the original on 2017-08-08 . Retrieved 2017-10-24 .
48. ** ^ ** Maruyama, K. (March 1991). "The discovery of adenosine triphosphate and the establishment of its structure". _J. Hist. Biol_ . **24** (1): 145–154. [ doi ](/wiki/Doi_\(identifier\) "Doi \(identifier\)") : [ 10.1007/BF00130477 ](https://doi.org/10.1007%2FBF00130477) . [ S2CID ](/wiki/S2CID_\(identifier\) "S2CID \(identifier\)") [ 87425890 ](https://api.semanticscholar.org/CorpusID:87425890) .
49. ** ^ ** Lipmann, F. (1941). "Metabolic generation and utilization of phosphate bond energy". _Adv. Enzymol_ . **1** : 99–162. [ ISSN ](/wiki/ISSN_\(identifier\) "ISSN \(identifier\)") [ 0196-7398 ](https://www.worldcat.org/issn/0196-7398) .
50. ** ^ ** [ "History: ATP first discovered in 1929" ](http://nobelprize.org/nobel_prizes/chemistry/laureates/1997/illpres/history.html) . _The Nobel Prize in Chemistry 1997_ . [ Nobel Foundation ](/wiki/Nobel_Foundation "Nobel Foundation") . [ Archived ](https://web.archive.org/web/20100123061355/http://nobelprize.org/nobel_prizes/chemistry/laureates/1997/illpres/history.html) from the original on 2010-01-23 . Retrieved 2010-05-26 .
51. ** ^ ** [ "The Nobel Prize in Chemistry 1997" ](https://www.nobelprize.org/nobel_prizes/chemistry/laureates/1997/) . _Nobel Prize_ . [ Archived ](https://web.archive.org/web/20171024205633/https://www.nobelprize.org/nobel_prizes/chemistry/laureates/1997/) from the original on 24 October 2017 . Retrieved 21 January 2018 .
## External links [ [ edit
](/w/index.php?title=Adenosine_triphosphate&action=edit§ion=31 "Edit
section: External links") ]

Wikimedia Commons has media related to [ Adenosine triphosphate
](https://commons.wikimedia.org/wiki/Category:Adenosine_triphosphate
"commons:Category:Adenosine triphosphate") .
* [ ATP bound to proteins ](http://www.ebi.ac.uk/pdbe-srv/PDBeXplore/ligand/?ligand=ATP) in the [ PDB ](/wiki/Protein_Data_Bank "Protein Data Bank")
* [ ScienceAid: Energy ATP and Exercise ](https://web.archive.org/web/20160310134410/http://www.scienceaid.co.uk/biology/biochemistry/atp.html)
* [ PubChem entry for Adenosine Triphosphate ](https://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?cid=5957)
* [ KEGG entry for Adenosine Triphosphate ](http://www.genome.jp/dbget-bin/www_bget?cpd:C00002)
* [ v ](/wiki/Template:Nucleobases,_nucleosides,_and_nucleotides "Template:Nucleobases, nucleosides, and nucleotides")
* [ t ](/wiki/Template_talk:Nucleobases,_nucleosides,_and_nucleotides "Template talk:Nucleobases, nucleosides, and nucleotides")
* [ e ](/wiki/Special:EditPage/Template:Nucleobases,_nucleosides,_and_nucleotides "Special:EditPage/Template:Nucleobases, nucleosides, and nucleotides")
[ Nucleic acid ](/wiki/Nucleic_acid "Nucleic acid") constituents
---
[ Nucleobase ](/wiki/Nucleobase "Nucleobase") |
* [ Purine ](/wiki/Purine "Purine")
* [ Adenine ](/wiki/Adenine "Adenine")
* [ Guanine ](/wiki/Guanine "Guanine")
* [ Hypoxanthine ](/wiki/Hypoxanthine "Hypoxanthine")
* [ Xanthine ](/wiki/Xanthine "Xanthine")
* [ Purine analogue ](/wiki/Purine_analogue "Purine analogue")
* [ Pyrimidine ](/wiki/Pyrimidine "Pyrimidine")
* [ Uracil ](/wiki/Uracil "Uracil")
* [ Thymine ](/wiki/Thymine "Thymine")
* [ Cytosine ](/wiki/Cytosine "Cytosine")
* [ Pyrimidine analogue ](/wiki/Pyrimidine_analogue "Pyrimidine analogue")
* [ Unnatural base pair (UBP) ](/wiki/Base_pair#Unnatural_base_pair_\(UBP\) "Base pair")
[ Nucleoside ](/wiki/Nucleoside "Nucleoside") |
| [ Ribonucleoside ](/wiki/Ribonucleoside "Ribonucleoside") |
* [ Adenosine ](/wiki/Adenosine "Adenosine")
* [ Guanosine ](/wiki/Guanosine "Guanosine")
* [ 5-Methyluridine ](/wiki/5-Methyluridine "5-Methyluridine")
* [ Uridine ](/wiki/Uridine "Uridine")
* [ 5-Methylcytidine ](/wiki/5-Methylcytidine "5-Methylcytidine")
* [ Cytidine ](/wiki/Cytidine "Cytidine")
* [ Pseudouridine ](/wiki/Pseudouridine "Pseudouridine")
* [ Inosine ](/wiki/Inosine "Inosine")
* [ _N_ 6 -Methyladenosine ](/wiki/N6-Methyladenosine "N6-Methyladenosine")
* [ Xanthosine ](/wiki/Xanthosine "Xanthosine")
* [ Wybutosine ](/wiki/Wybutosine "Wybutosine")
---|---
[ Deoxyribonucleoside ](/wiki/Deoxyribonucleoside "Deoxyribonucleoside") |
* [ Deoxyadenosine ](/wiki/Deoxyadenosine "Deoxyadenosine")
* [ Deoxyguanosine ](/wiki/Deoxyguanosine "Deoxyguanosine")
* [ Thymidine ](/wiki/Thymidine "Thymidine")
* [ Deoxyuridine ](/wiki/Deoxyuridine "Deoxyuridine")
* [ Deoxycytidine ](/wiki/Deoxycytidine "Deoxycytidine")
* [ Deoxyinosine ](/w/index.php?title=Deoxyinosine&action=edit&redlink=1 "Deoxyinosine \(page does not exist\)")
* [ Deoxyxanthosine ](/w/index.php?title=Deoxyxanthosine&action=edit&redlink=1 "Deoxyxanthosine \(page does not exist\)")
[ Nucleotide ](/wiki/Nucleotide "Nucleotide")
(Nucleoside monophosphate) |
| [ Ribonucleotide ](/wiki/Ribonucleotide "Ribonucleotide") |
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* [ GMP ](/wiki/Guanosine_monophosphate "Guanosine monophosphate")
* [ m 5 UMP ](/w/index.php?title=5-Methyluridine_monophosphate&action=edit&redlink=1 "5-Methyluridine monophosphate \(page does not exist\)")
* [ UMP ](/wiki/Uridine_monophosphate "Uridine monophosphate")
* [ CMP ](/wiki/Cytidine_monophosphate "Cytidine monophosphate")
* [ IMP ](/wiki/Inosinic_acid "Inosinic acid")
* [ XMP ](/wiki/Xanthosine_monophosphate "Xanthosine monophosphate")
---|---
[ Deoxyribonucleotide ](/wiki/Deoxyribonucleotide "Deoxyribonucleotide") |
* [ dAMP ](/wiki/Deoxyadenosine_monophosphate "Deoxyadenosine monophosphate")
* [ dGMP ](/wiki/Deoxyguanosine_monophosphate "Deoxyguanosine monophosphate")
* [ dTMP ](/wiki/Thymidine_monophosphate "Thymidine monophosphate")
* [ dUMP ](/wiki/Deoxyuridine_monophosphate "Deoxyuridine monophosphate")
* [ dCMP ](/wiki/Deoxycytidine_monophosphate "Deoxycytidine monophosphate")
* [ dIMP ](/wiki/Deoxyinosine_monophosphate "Deoxyinosine monophosphate")
* [ dXMP ](/w/index.php?title=Deoxyxanthosine_monophosphate&action=edit&redlink=1 "Deoxyxanthosine monophosphate \(page does not exist\)")
[ Cyclic nucleotide ](/wiki/Cyclic_nucleotide "Cyclic nucleotide") |
* [ cAMP ](/wiki/Cyclic_adenosine_monophosphate "Cyclic adenosine monophosphate")
* [ cGMP ](/wiki/Cyclic_guanosine_monophosphate "Cyclic guanosine monophosphate")
* [ c-di-GMP ](/wiki/Cyclic_di-GMP "Cyclic di-GMP")
* [ c-di-AMP ](/wiki/Cyclic_di-AMP "Cyclic di-AMP")
* [ cADPR ](/wiki/Cyclic_ADP-ribose "Cyclic ADP-ribose")
* [ cGAMP ](/wiki/Cyclic_guanosine_monophosphate%E2%80%93adenosine_monophosphate "Cyclic guanosine monophosphate–adenosine monophosphate")
Nucleoside diphosphate |
|
* [ ADP ](/wiki/Adenosine_diphosphate "Adenosine diphosphate")
* [ GDP ](/wiki/Guanosine_diphosphate "Guanosine diphosphate")
* [ m 5 UDP ](/w/index.php?title=5-Methyluridine_diphosphate&action=edit&redlink=1 "5-Methyluridine diphosphate \(page does not exist\)")
* [ UDP ](/wiki/Uridine_diphosphate "Uridine diphosphate")
* [ CDP ](/wiki/Cytidine_diphosphate "Cytidine diphosphate")
* [ Xanthosine diphosphate ](/w/index.php?title=Xanthosine_diphosphate&action=edit&redlink=1 "Xanthosine diphosphate \(page does not exist\)")
---
* [ dADP ](/wiki/Deoxyadenosine_diphosphate "Deoxyadenosine diphosphate")
* [ dGDP ](/wiki/Deoxyguanosine_diphosphate "Deoxyguanosine diphosphate")
* [ dTDP ](/wiki/Thymidine_diphosphate "Thymidine diphosphate")
* [ dUDP ](/w/index.php?title=Deoxyuridine_diphosphate&action=edit&redlink=1 "Deoxyuridine diphosphate \(page does not exist\)")
* [ dCDP ](/wiki/Deoxycytidine_diphosphate "Deoxycytidine diphosphate")
[ Nucleoside triphosphate ](/wiki/Nucleoside_triphosphate "Nucleoside
triphosphate") |
|
* ATP
* [ GTP ](/wiki/Guanosine_triphosphate "Guanosine triphosphate")
* [ m 5 UTP ](/wiki/5-Methyluridine_triphosphate "5-Methyluridine triphosphate")
* [ UTP ](/wiki/Uridine_triphosphate "Uridine triphosphate")
* [ CTP ](/wiki/Cytidine_triphosphate "Cytidine triphosphate")
* [ ITP ](/wiki/Inosine_triphosphate "Inosine triphosphate")
* [ XTP ](/wiki/Xanthosine_triphosphate "Xanthosine triphosphate")
---
* [ dATP ](/wiki/Deoxyadenosine_triphosphate "Deoxyadenosine triphosphate")
* [ dGTP ](/wiki/Deoxyguanosine_triphosphate "Deoxyguanosine triphosphate")
* [ dTTP ](/wiki/Thymidine_triphosphate "Thymidine triphosphate")
* [ dUTP ](/w/index.php?title=Deoxyuridine_triphosphate&action=edit&redlink=1 "Deoxyuridine triphosphate \(page does not exist\)")
* [ dCTP ](/wiki/Deoxycytidine_triphosphate "Deoxycytidine triphosphate")
* [ dITP ](/w/index.php?title=Deoxyinosine_triphosphate&action=edit&redlink=1 "Deoxyinosine triphosphate \(page does not exist\)")
* [ dXTP ](/w/index.php?title=Deoxyxanthosine_triphosphate&action=edit&redlink=1 "Deoxyxanthosine triphosphate \(page does not exist\)")
* [ v ](/wiki/Template:Enzyme_cofactors "Template:Enzyme cofactors")
* [ t ](/wiki/Template_talk:Enzyme_cofactors "Template talk:Enzyme cofactors")
* [ e ](/wiki/Special:EditPage/Template:Enzyme_cofactors "Special:EditPage/Template:Enzyme cofactors")
[ Enzyme ](/wiki/Enzyme "Enzyme") [ cofactors
](/wiki/Cofactor_\(biochemistry\) "Cofactor \(biochemistry\)")
---
Active forms |
| [ vitamins ](/wiki/Vitamin "Vitamin") |
* [ TPP / ThDP ](/wiki/Thiamine_pyrophosphate "Thiamine pyrophosphate") (B 1 )
* [ FMN ](/wiki/Flavin_mononucleotide "Flavin mononucleotide") , [ FAD ](/wiki/Flavin_adenine_dinucleotide "Flavin adenine dinucleotide") (B 2 )
* [ NAD \+ ](/wiki/Nicotinamide_adenine_dinucleotide "Nicotinamide adenine dinucleotide") , [ NADH ](/wiki/Nicotinamide_adenine_dinucleotide "Nicotinamide adenine dinucleotide") , [ NADP \+ ](/wiki/Nicotinamide_adenine_dinucleotide_phosphate "Nicotinamide adenine dinucleotide phosphate") , [ NADPH ](/wiki/Nicotinamide_adenine_dinucleotide_phosphate "Nicotinamide adenine dinucleotide phosphate") (B 3 )
* [ Coenzyme A ](/wiki/Coenzyme_A "Coenzyme A") (B 5 )
* [ PLP / P5P ](/wiki/Pyridoxal_phosphate "Pyridoxal phosphate") (B 6 )
* [ Biotin ](/wiki/Biotin "Biotin") (B 7 )
* [ THFA / H 4 FA ](/wiki/Tetrahydrofolic_acid "Tetrahydrofolic acid") , [ DHFA / H 2 FA ](/wiki/Dihydrofolic_acid "Dihydrofolic acid") , [ MTHF ](/wiki/5,10-Methylenetetrahydrofolate "5,10-Methylenetetrahydrofolate") (B 9 )
* [ AdoCbl ](/wiki/Adenosylcobalamin "Adenosylcobalamin") , [ MeCbl ](/wiki/Methylcobalamin "Methylcobalamin") (B 12 )
* [ Ascorbic acid ](/wiki/Vitamin_C "Vitamin C") (C)
* [ Phylloquinone ](/wiki/Phytomenadione "Phytomenadione") (K 1 ), [ Menaquinone ](/wiki/Vitamin_K "Vitamin K") (K 2 )
* [ Coenzyme F420 ](/wiki/Coenzyme_F420 "Coenzyme F420")
---|---
[ non-vitamins ](/wiki/Cofactor_\(biochemistry\)#Non-vitamins "Cofactor
\(biochemistry\)") |
* ATP
* [ CTP ](/wiki/Cytidine_triphosphate "Cytidine triphosphate")
* [ SAMe ](/wiki/S-Adenosyl_methionine "S-Adenosyl methionine")
* [ PAPS ](/wiki/3%27-Phosphoadenosine-5%27-phosphosulfate "3'-Phosphoadenosine-5'-phosphosulfate")
* [ GSH ](/wiki/Glutathione "Glutathione")
* [ Coenzyme B ](/wiki/Coenzyme_B "Coenzyme B")
* [ Cofactor F430 ](/wiki/Cofactor_F430 "Cofactor F430")
* [ Coenzyme M ](/wiki/Coenzyme_M "Coenzyme M")
* [ Coenzyme Q ](/wiki/Coenzyme_Q10 "Coenzyme Q10")
* [ Heme / Haem ](/wiki/Heme "Heme") ( [ A ](/wiki/Heme_A "Heme A") , [ B ](/wiki/Heme_B "Heme B") , [ C ](/wiki/Heme_C "Heme C") , [ O ](/wiki/Heme_O "Heme O") )
* [ Lipoic Acid ](/wiki/Lipoic_acid "Lipoic acid")
* [ Methanofuran ](/wiki/Methanofuran "Methanofuran")
* [ Molybdopterin ](/wiki/Molybdopterin "Molybdopterin")
* [ Mycofactocin ](/wiki/Mycofactocin "Mycofactocin")
* [ PQQ ](/wiki/Pyrroloquinoline_quinone "Pyrroloquinoline quinone")
* [ THB / BH 4 ](/wiki/Tetrahydrobiopterin "Tetrahydrobiopterin")
* [ THMPT / H 4 MPT ](/wiki/Tetrahydromethanopterin "Tetrahydromethanopterin")
[ metal ions ](/wiki/Mineral_\(nutrient\) "Mineral \(nutrient\)") |
* [ Ca 2+ ](/wiki/Calcium "Calcium")
* [ Cu 2+ ](/wiki/Copper "Copper")
* [ Fe 2+ , Fe 3+ ](/wiki/Iron "Iron")
* [ Mg 2+ ](/wiki/Magnesium "Magnesium")
* [ Mn 2+ ](/wiki/Manganese "Manganese")
* [ Mo ](/wiki/Molybdenum "Molybdenum")
* [ Ni 2+ ](/wiki/Nickel "Nickel")
* [ Zn 2+ ](/wiki/Zinc "Zinc")
Base forms |
* _[ vitamins ](/wiki/Vitamin "Vitamin") _ : _see[ vitamins ](/wiki/Template:Vitamins "Template:Vitamins") _
* [ v ](/wiki/Template:Neurotransmitters "Template:Neurotransmitters")
* [ t ](/wiki/Template_talk:Neurotransmitters "Template talk:Neurotransmitters")
* [ e ](/wiki/Special:EditPage/Template:Neurotransmitters "Special:EditPage/Template:Neurotransmitters")
[ Neurotransmitters ](/wiki/Neurotransmitter "Neurotransmitter")
---
[ Amino acid ](/wiki/Amino_acid "Amino acid") -derived |
| Major excitatory /
inhibitory systems |
| Glutamate system |
* [ Agmatine ](/wiki/Agmatine "Agmatine")
* [ Aspartic acid (aspartate) ](/wiki/Aspartic_acid "Aspartic acid")
* [ Glutamic acid (glutamate) ](/wiki/Glutamate_\(neurotransmitter\) "Glutamate \(neurotransmitter\)")
* [ Glutathione ](/wiki/Glutathione "Glutathione")
* [ Glycine ](/wiki/Glycine "Glycine")
* [ GSNO ](/wiki/S-Nitrosoglutathione "S-Nitrosoglutathione")
* [ GSSG ](/wiki/Oxidized_glutathione "Oxidized glutathione")
* [ Kynurenic acid ](/wiki/Kynurenic_acid "Kynurenic acid")
* [ NAA ](/wiki/N-Acetylaspartic_acid "N-Acetylaspartic acid")
* [ NAAG ](/wiki/N-Acetylaspartylglutamic_acid "N-Acetylaspartylglutamic acid")
* [ Proline ](/wiki/Proline "Proline")
* [ Serine ](/wiki/Serine "Serine")
---|---
GABA system |
* [ GABA ](/wiki/Gamma-Aminobutyric_acid "Gamma-Aminobutyric acid")
* [ GABOB ](/wiki/Gamma-Amino-beta-hydroxybutyric_acid "Gamma-Amino-beta-hydroxybutyric acid")
* [ GHB ](/wiki/Gamma-Hydroxybutyric_acid "Gamma-Hydroxybutyric acid")
Glycine system |
* [ α-Alanine ](/wiki/Alanine "Alanine")
* [ β-Alanine ](/wiki/%CE%92-Alanine "Β-Alanine")
* [ Glycine ](/wiki/Glycine "Glycine")
* [ Hypotaurine ](/wiki/Hypotaurine "Hypotaurine")
* [ Proline ](/wiki/Proline "Proline")
* [ Sarcosine ](/wiki/Sarcosine "Sarcosine")
* [ Serine ](/wiki/Serine "Serine")
* [ Taurine ](/wiki/Taurine "Taurine")
[ GHB system ](/wiki/GHB_receptor "GHB receptor") |
* [ GHB ](/wiki/Gamma-Hydroxybutyric_acid "Gamma-Hydroxybutyric acid")
* [ T-HCA (GHC) ](/wiki/T-HCA "T-HCA")
[ Biogenic amines ](/wiki/Biogenic_amine "Biogenic amine") |
| Monoamines |
* [ 6-OHM ](/wiki/6-Hydroxymelatonin "6-Hydroxymelatonin")
* [ Dopamine ](/wiki/Dopamine "Dopamine")
* [ Epinephrine (adrenaline) ](/wiki/Adrenaline "Adrenaline")
* [ NAS (normelatonin) ](/wiki/N-Acetylserotonin "N-Acetylserotonin")
* [ Norepinephrine (noradrenaline) ](/wiki/Norepinephrine "Norepinephrine")
* [ Serotonin (5-HT) ](/wiki/Serotonin "Serotonin")
---|---
Trace amines |
* [ 3-Iodothyronamine ](/wiki/3-Iodothyronamine "3-Iodothyronamine")
* [ N-Methylphenethylamine ](/wiki/N-Methylphenethylamine "N-Methylphenethylamine")
* [ N-Methyltryptamine ](/wiki/N-Methyltryptamine "N-Methyltryptamine")
* [ _m_ -Octopamine ](/wiki/Norfenefrine "Norfenefrine")
* [ _p_ -Octopamine ](/wiki/Octopamine "Octopamine")
* [ Phenylethanolamine ](/wiki/Phenylethanolamine "Phenylethanolamine")
* [ Phenethylamine ](/wiki/Phenethylamine "Phenethylamine")
* [ Synephrine ](/wiki/Synephrine "Synephrine")
* [ Tryptamine ](/wiki/Tryptamine "Tryptamine")
* [ _m_ -Tyramine ](/wiki/Meta-Tyramine "Meta-Tyramine")
* [ _p_ -Tyramine ](/wiki/Tyramine "Tyramine")
Others |
* [ Histamine ](/wiki/Histamine "Histamine")
[ Neuropeptides ](/wiki/Neuropeptide "Neuropeptide") |
|
* See [ here ](/wiki/Template:Neuropeptides "Template:Neuropeptides") instead.
---
[ Lipid ](/wiki/Lipid "Lipid") -derived |
| Endocannabinoids |
* [ 2-AG ](/wiki/2-Arachidonoylglycerol "2-Arachidonoylglycerol")
* [ 2-AGE (noladin ether) ](/wiki/2-Arachidonyl_glyceryl_ether "2-Arachidonyl glyceryl ether")
* [ 2-ALPI ](/w/index.php?title=2-Arachidonoyl_lysophosphatidylinositol&action=edit&redlink=1 "2-Arachidonoyl lysophosphatidylinositol \(page does not exist\)")
* [ 2-OG ](/wiki/2-Oleoylglycerol "2-Oleoylglycerol")
* [ AA-5-HT ](/wiki/Arachidonoyl_serotonin "Arachidonoyl serotonin")
* [ Anandamide (AEA) ](/wiki/Anandamide "Anandamide")
* [ DEA ](/wiki/Docosatetraenoylethanolamide "Docosatetraenoylethanolamide")
* [ LPI ](/wiki/Lysophosphatidylinositol "Lysophosphatidylinositol")
* [ NADA ](/wiki/N-Arachidonoyl_dopamine "N-Arachidonoyl dopamine")
* [ NAGly ](/wiki/N-Arachidonylglycine "N-Arachidonylglycine")
* [ OEA ](/wiki/Oleoylethanolamide "Oleoylethanolamide")
* [ Oleamide ](/wiki/Oleamide "Oleamide")
* [ PEA ](/wiki/Palmitoylethanolamide "Palmitoylethanolamide")
* [ RVD-Hpα ](/wiki/RVD-Hp%CE%B1 "RVD-Hpα")
* [ SEA ](/wiki/Stearoylethanolamide "Stearoylethanolamide")
* [ Virodhamine (O-AEA) ](/wiki/Virodhamine "Virodhamine")
---|---
[ Neurosteroids ](/wiki/Neurosteroid "Neurosteroid") |
|
* See [ here ](/wiki/Template:Steroid_hormones "Template:Steroid hormones") instead.
---
[ Nucleobase ](/wiki/Nucleobase "Nucleobase") -derived |
| Nucleosides |
| [ Adenosine ](/wiki/Adenosine "Adenosine") system |
* [ ADP ](/wiki/Adenosine_diphosphate "Adenosine diphosphate")
* [ AMP ](/wiki/Adenosine_monophosphate "Adenosine monophosphate")
* ATP
---|---
[ Vitamin ](/wiki/Vitamin "Vitamin") -derived |
---|---
Miscellaneous |
| Cholinergic system |
* [ Acetylcholine ](/wiki/Acetylcholine "Acetylcholine")
---|---
Gasotransmitters |
* [ Carbon monoxide (CO) ](/wiki/Carbon_monoxide "Carbon monoxide")
* [ Hydrogen sulfide (H 2 S) ](/wiki/Hydrogen_sulfide "Hydrogen sulfide")
* [ Nitric oxide (NO) ](/wiki/Nitric_oxide "Nitric oxide")
---|---
Candidates |
* [ Acetaldehyde ](/wiki/Acetaldehyde "Acetaldehyde")
* [ Ammonia (NH 3 ) ](/wiki/Ammonia "Ammonia")
* [ Carbonyl sulfide (COS) ](/wiki/Carbonyl_sulfide "Carbonyl sulfide")
* [ Nitrous oxide (N 2 O) ](/wiki/Nitrous_oxide "Nitrous oxide")
* [ Sulfur dioxide (SO 2 ) ](/wiki/Sulfur_dioxide "Sulfur dioxide")
---|---
* [ v ](/wiki/Template:Food_science "Template:Food science")
* [ t ](/wiki/Template_talk:Food_science "Template talk:Food science")
* [ e ](/wiki/Special:EditPage/Template:Food_science "Special:EditPage/Template:Food science")
[ Food science ](/wiki/Food_science "Food science")
---
General |
* [ Allergy ](/wiki/Food_allergy "Food allergy")
* [ Engineering ](/wiki/Food_engineering "Food engineering")
* [ Microbiology ](/wiki/Food_microbiology "Food microbiology")
* [ Nutrition ](/wiki/Nutrition "Nutrition")
* [ Diet ](/wiki/Diet_\(nutrition\) "Diet \(nutrition\)")
* [ clinical ](/wiki/Clinical_nutrition "Clinical nutrition")
* [ Processing ](/wiki/Food_processing "Food processing")
* [ Processing aids ](/wiki/Processing_aid "Processing aid")
* [ Psychology ](/wiki/Food_psychology "Food psychology")
* [ Quality ](/wiki/Food_quality "Food quality")
* [ Sensory analysis ](/wiki/Sensory_analysis "Sensory analysis")
* [ Discrimination testing ](/wiki/Discrimination_testing "Discrimination testing")
* [ Rheology ](/wiki/Food_rheology "Food rheology")
* [ Storage ](/wiki/Food_storage "Food storage")
* [ Technology ](/wiki/Food_technology "Food technology")
|
|
* [ v ](/wiki/Template:Food_chemistry "Template:Food chemistry")
* [ t ](/wiki/Template_talk:Food_chemistry "Template talk:Food chemistry")
* [ e ](/wiki/Special:EditPage/Template:Food_chemistry "Special:EditPage/Template:Food chemistry")
[ Food chemistry ](/wiki/Food_chemistry "Food chemistry")
---
* [ Additives ](/wiki/Food_additive "Food additive")
* [ Carbohydrates ](/wiki/Carbohydrate "Carbohydrate")
* [ Coloring ](/wiki/Food_coloring "Food coloring")
* [ Enzymes ](/wiki/Enzyme "Enzyme")
* [ Essential fatty acids ](/wiki/Essential_fatty_acid "Essential fatty acid")
* [ Flavors ](/wiki/Flavoring "Flavoring")
* [ Fortification ](/wiki/Food_fortification "Food fortification")
* [ Lipids ](/wiki/Lipid "Lipid")
* [ "Minerals" (Chemical elements) ](/wiki/Mineral_\(nutrient\) "Mineral \(nutrient\)")
* [ Proteins ](/wiki/Protein "Protein")
* [ Vitamins ](/wiki/Vitamin "Vitamin")
* [ Water ](/wiki/Water "Water")
|
* [ v ](/wiki/Template:Food_preservation "Template:Food preservation")
* [ t ](/wiki/Template_talk:Food_preservation "Template talk:Food preservation")
* [ e ](/wiki/Special:EditPage/Template:Food_preservation "Special:EditPage/Template:Food preservation")
[ Food preservation ](/wiki/Food_preservation "Food preservation")
---
* [ Biopreservation ](/wiki/Biopreservation "Biopreservation")
* [ Canning ](/wiki/Canning "Canning")
* [ Cold chain ](/wiki/Cold_chain "Cold chain")
* [ Curing ](/wiki/Curing_\(food_preservation\) "Curing \(food preservation\)")
* [ Drying ](/wiki/Food_drying "Food drying")
* [ Fermentation ](/wiki/Fermentation_in_food_processing "Fermentation in food processing")
* [ Freeze-drying ](/wiki/Freeze-drying "Freeze-drying")
* [ Freezing ](/wiki/Frozen_food "Frozen food")
* [ Hurdle technology ](/wiki/Hurdle_technology "Hurdle technology")
* [ Irradiation ](/wiki/Food_irradiation "Food irradiation")
* [ Jamming ](/wiki/Fruit_preserves "Fruit preserves")
* [ Jellying ](/wiki/Aspic "Aspic")
* [ Jugging ](/wiki/Jugging "Jugging")
* [ Modified atmosphere ](/wiki/Modified_atmosphere "Modified atmosphere")
* [ Pascalization ](/wiki/Pascalization "Pascalization")
* [ Pickling ](/wiki/Pickling "Pickling")
* [ Potting ](/wiki/Potted_meat "Potted meat")
* [ Confit ](/wiki/Confit "Confit")
* [ Potjevleesch ](/wiki/Potjevleesch "Potjevleesch")
* [ Salting ](/wiki/Salting_\(food\) "Salting \(food\)")
* [ Smoking ](/wiki/Smoking_\(cooking\) "Smoking \(cooking\)")
* [ Sugaring ](/wiki/Sugaring "Sugaring")
* [ Tyndallization ](/wiki/Tyndallization "Tyndallization")
* [ Vacuum packing ](/wiki/Vacuum_packing "Vacuum packing")
* [  ](/wiki/File:Foodlogo2.svg) [ Food portal ](/wiki/Portal:Food "Portal:Food")
*  [ Category: Food preservation ](/wiki/Category:Food_preservation "Category:Food preservation")
[ Food industry ](/wiki/Food_industry "Food industry") |
* [ Manufacturing ](/wiki/Food_manufacturing "Food manufacturing")
* [ Packaging ](/wiki/Food_packaging "Food packaging")
* [ Marketing ](/wiki/Food_marketing "Food marketing")
* [ Foodservice ](/wiki/Foodservice "Foodservice")
* [ Fortification ](/wiki/Food_fortification "Food fortification")
|
|
* [ v ](/wiki/Template:Consumer_food_safety "Template:Consumer food safety")
* [ t ](/wiki/Template_talk:Consumer_food_safety "Template talk:Consumer food safety")
* [ e ](/wiki/Special:EditPage/Template:Consumer_food_safety "Special:EditPage/Template:Consumer food safety")
[ Consumer food safety ](/wiki/Food_safety "Food safety")
---
[ Adulterants ](/wiki/Adulterant "Adulterant") , [ food contaminants
](/wiki/Food_contaminant "Food contaminant") |
* [ 3-MCPD ](/wiki/3-MCPD "3-MCPD")
* [ Aldicarb ](/wiki/Aldicarb "Aldicarb")
* [ Antibiotic use in livestock ](/wiki/Antibiotic_use_in_livestock "Antibiotic use in livestock")
* [ Cyanide ](/wiki/Cyanide "Cyanide")
* [ Formaldehyde ](/wiki/Formaldehyde "Formaldehyde")
* [ HGH controversies ](/wiki/HGH_controversies "HGH controversies")
* [ Lead poisoning ](/wiki/Lead_poisoning "Lead poisoning")
* [ Melamine ](/wiki/Melamine "Melamine")
* [ Mercury in fish ](/wiki/Mercury_in_fish "Mercury in fish")
* [ Sudan I ](/wiki/Sudan_I "Sudan I")
[ Food additives ](/wiki/Food_additive "Food additive") |
* [ Flavorings ](/wiki/Flavoring "Flavoring")
* [ Monosodium glutamate (MSG) ](/wiki/Monosodium_glutamate "Monosodium glutamate")
* [ Salt ](/wiki/Sodium_chloride "Sodium chloride")
* [ Sugar ](/wiki/Sugar "Sugar")
* [ High-fructose corn syrup ](/wiki/High-fructose_corn_syrup "High-fructose corn syrup")
[ Intestinal parasites ](/wiki/Intestinal_parasite_infection "Intestinal
parasite infection") , [ parasitic disease ](/wiki/Parasitic_disease
"Parasitic disease") |
* [ Amoebiasis ](/wiki/Amoebiasis "Amoebiasis")
* [ Anisakiasis ](/wiki/Anisakiasis "Anisakiasis")
* [ Cryptosporidiosis ](/wiki/Cryptosporidiosis "Cryptosporidiosis")
* [ Cyclosporiasis ](/wiki/Cyclosporiasis "Cyclosporiasis")
* [ Diphyllobothriasis ](/wiki/Diphyllobothriasis "Diphyllobothriasis")
* [ Enterobiasis ](/wiki/Pinworm_infection "Pinworm infection")
* [ Fasciolopsiasis ](/wiki/Fasciolopsiasis "Fasciolopsiasis")
* [ Fasciolosis ](/wiki/Fasciolosis "Fasciolosis")
* [ Giardiasis ](/wiki/Giardiasis "Giardiasis")
* [ Gnathostomiasis ](/wiki/Gnathostomiasis "Gnathostomiasis")
* [ Paragonimiasis ](/wiki/Paragonimiasis "Paragonimiasis")
* [ Toxocariasis ](/wiki/Toxocariasis "Toxocariasis")
* [ Toxoplasmosis ](/wiki/Toxoplasmosis "Toxoplasmosis")
* [ Trichinosis ](/wiki/Trichinosis "Trichinosis")
* [ Trichuriasis ](/wiki/Trichuriasis "Trichuriasis")
[ Microorganisms ](/wiki/Microorganism "Microorganism") |
* [ Botulism ](/wiki/Botulism "Botulism")
* _[ Campylobacter jejuni ](/wiki/Campylobacter_jejuni "Campylobacter jejuni") _
* _[ Clostridium perfringens ](/wiki/Clostridium_perfringens "Clostridium perfringens") _
* _[ Cronobacter ](/wiki/Cronobacter "Cronobacter") _
* _[ Enterovirus ](/wiki/Enterovirus "Enterovirus") _
* [ _Escherichia coli_ O104:H4 ](/wiki/Escherichia_coli_O104:H4 "Escherichia coli O104:H4")
* [ _Escherichia coli_ O157:H7 ](/wiki/Escherichia_coli_O157:H7 "Escherichia coli O157:H7")
* [ Hepatitis A ](/wiki/Hepatitis_A "Hepatitis A")
* [ Hepatitis E ](/wiki/Hepatitis_E "Hepatitis E")
* _[ Listeria ](/wiki/Listeria "Listeria") _
* [ Norovirus ](/wiki/Norovirus "Norovirus")
* [ Rotavirus ](/wiki/Rotavirus "Rotavirus")
* _[ Salmonella ](/wiki/Salmonella "Salmonella") _
* _[ Vibrio cholerae ](/wiki/Vibrio_cholerae "Vibrio cholerae") _
[ Pesticides ](/wiki/Pesticide "Pesticide") |
* [ Chlorpyrifos ](/wiki/Chlorpyrifos "Chlorpyrifos")
* [ DDT ](/wiki/DDT "DDT")
* [ Lindane ](/wiki/Lindane "Lindane")
* [ Malathion ](/wiki/Malathion "Malathion")
* [ Methamidophos ](/wiki/Methamidophos "Methamidophos")
[ Preservatives ](/wiki/Preservative "Preservative") |
* [ Benzoic acid ](/wiki/Benzoic_acid "Benzoic acid")
* [ Ethylenediaminetetraacetic acid (EDTA) ](/wiki/Ethylenediaminetetraacetic_acid "Ethylenediaminetetraacetic acid")
* [ Sodium benzoate ](/wiki/Sodium_benzoate "Sodium benzoate")
[ Sugar substitutes ](/wiki/Sugar_substitute "Sugar substitute") |
* [ Acesulfame potassium ](/wiki/Acesulfame_potassium "Acesulfame potassium")
* [ Aspartame ](/wiki/Aspartame "Aspartame")
* [ controversy ](/wiki/Aspartame_controversy "Aspartame controversy")
* [ Saccharin ](/wiki/Saccharin "Saccharin")
* [ Sodium cyclamate ](/wiki/Sodium_cyclamate "Sodium cyclamate")
* [ Sorbitol ](/wiki/Sorbitol "Sorbitol")
* [ Sucralose ](/wiki/Sucralose "Sucralose")
[ Toxins ](/wiki/Toxin "Toxin") , [ poisons ](/wiki/Poison "Poison") , [
environment pollution ](/wiki/Pollution "Pollution") |
* [ Aflatoxin ](/wiki/Aflatoxin "Aflatoxin")
* [ Arsenic contamination of groundwater ](/wiki/Arsenic_contamination_of_groundwater "Arsenic contamination of groundwater")
* [ Benzene in soft drinks ](/wiki/Benzene_in_soft_drinks "Benzene in soft drinks")
* [ Bisphenol A ](/wiki/Bisphenol_A "Bisphenol A")
* [ Dieldrin ](/wiki/Dieldrin "Dieldrin")
* [ Diethylstilbestrol ](/wiki/Diethylstilbestrol "Diethylstilbestrol")
* [ Dioxin ](/wiki/Dioxin "Dioxin")
* [ Mycotoxins ](/wiki/Mycotoxin "Mycotoxin")
* [ Nonylphenol ](/wiki/Nonylphenol "Nonylphenol")
* [ Shellfish poisoning ](/wiki/Shellfish_poisoning "Shellfish poisoning")
[ Food processing ](/wiki/Food_processing "Food processing") |
* [ 4-Hydroxynonenal ](/wiki/4-Hydroxynonenal "4-Hydroxynonenal")
* [ Acid-hydrolyzed vegetable protein ](/wiki/Acid-hydrolyzed_vegetable_protein "Acid-hydrolyzed vegetable protein")
* [ Acrylamide ](/wiki/Acrylamide "Acrylamide")
* [ Creutzfeldt–Jakob disease ](/wiki/Creutzfeldt%E2%80%93Jakob_disease "Creutzfeldt–Jakob disease")
* [ Food additives ](/wiki/List_of_food_additives "List of food additives")
* [ Food irradiation ](/wiki/Food_irradiation "Food irradiation")
* [ Heterocyclic amines ](/wiki/Heterocyclic_amine "Heterocyclic amine")
* [ Modified starch ](/wiki/Modified_starch "Modified starch")
* [ Nitrosamines ](/wiki/Nitrosamine "Nitrosamine")
* [ Polycyclic aromatic hydrocarbon ](/wiki/Polycyclic_aromatic_hydrocarbon "Polycyclic aromatic hydrocarbon")
* [ Shortening ](/wiki/Shortening "Shortening")
* [ Trans fat ](/wiki/Trans_fat "Trans fat")
* [ Water fluoridation controversy ](/wiki/Water_fluoridation_controversy "Water fluoridation controversy")
[ Food contamination incidents ](/wiki/List_of_food_contamination_incidents
"List of food contamination incidents") |
* [ Devon colic ](/wiki/Devon_colic "Devon colic")
* [ Swill milk scandal ](/wiki/Swill_milk_scandal "Swill milk scandal")
* [ Esing Bakery incident ](/wiki/Esing_Bakery_incident "Esing Bakery incident")
* [ 1858 Bradford sweets poisoning ](/wiki/1858_Bradford_sweets_poisoning "1858 Bradford sweets poisoning")
* [ 1900 English beer poisoning ](/wiki/1900_English_beer_poisoning "1900 English beer poisoning")
* [ Morinaga Milk arsenic poisoning incident ](/wiki/Morinaga_Milk_arsenic_poisoning_incident "Morinaga Milk arsenic poisoning incident")
* [ Minamata disease ](/wiki/Minamata_disease "Minamata disease")
* [ 1971 Iraq poison grain disaster ](/wiki/1971_Iraq_poison_grain_disaster "1971 Iraq poison grain disaster")
* [ Toxic oil syndrome ](/wiki/Toxic_oil_syndrome "Toxic oil syndrome")
* [ 1985 Austrian diethylene glycol wine scandal ](/wiki/1985_Austrian_diethylene_glycol_wine_scandal "1985 Austrian diethylene glycol wine scandal")
* [ United Kingdom BSE outbreak ](/wiki/United_Kingdom_BSE_outbreak "United Kingdom BSE outbreak")
* [ Australian meat substitution scandal ](/wiki/Australian_meat_substitution_scandal "Australian meat substitution scandal")
* [ Jack in the Box _E. coli_ outbreak ](/wiki/1993_Jack_in_the_Box_E._coli_outbreak "1993 Jack in the Box E. coli outbreak")
* [ 1996 Odwalla _E. coli_ outbreak ](/wiki/1996_Odwalla_E._coli_outbreak "1996 Odwalla E. coli outbreak")
* [ 2006 North American _E. coli_ outbreaks ](/wiki/2006_North_American_E._coli_O157:H7_outbreaks "2006 North American E. coli O157:H7 outbreaks")
* [ ICA meat repackaging controversy ](/wiki/ICA_meat_repackaging_controversy "ICA meat repackaging controversy")
* [ 2008 Canada listeriosis outbreak ](/wiki/2008_Canada_listeriosis_outbreak "2008 Canada listeriosis outbreak")
* [ 2008 Chinese milk scandal ](/wiki/2008_Chinese_milk_scandal "2008 Chinese milk scandal")
* [ 2008 Irish pork crisis ](/wiki/2008_Irish_pork_crisis "2008 Irish pork crisis")
* [ 2008 United States salmonellosis outbreak ](/wiki/2008_United_States_salmonellosis_outbreak "2008 United States salmonellosis outbreak")
* [ 2011 Germany _E. coli_ outbreak ](/wiki/2011_Germany_E._coli_O104:H4_outbreak "2011 Germany E. coli O104:H4 outbreak")
* [ 2011 United States listeriosis outbreak ](/wiki/2011_United_States_listeriosis_outbreak "2011 United States listeriosis outbreak")
* [ Bihar school meal poisoning ](/wiki/Bihar_school_meal_poisoning_incident "Bihar school meal poisoning incident")
* [ 2013 horse meat scandal ](/wiki/2013_horse_meat_scandal "2013 horse meat scandal")
* [ 2015 Mozambique funeral beer poisoning ](/wiki/Mozambique_funeral_beer_poisoning "Mozambique funeral beer poisoning")
* [ 2017 Brazil Operation Weak Meat ](/wiki/Operation_Weak_Meat "Operation Weak Meat")
* [ 2017–2018 South African listeriosis outbreak ](/wiki/2017%E2%80%932018_South_African_listeriosis_outbreak "2017–2018 South African listeriosis outbreak")
* [ 2018 Australian strawberry contamination ](/wiki/2018_Australian_strawberry_contamination "2018 Australian strawberry contamination")
* [ Food safety incidents in China ](/wiki/Food_safety_incidents_in_China "Food safety incidents in China")
* [ Food safety incidents in Taiwan ](/wiki/Food_safety_incidents_in_Taiwan "Food safety incidents in Taiwan")
* [ Foodborne illness ](/wiki/Foodborne_illness "Foodborne illness")
* [ outbreaks ](/wiki/List_of_foodborne_illness_outbreaks "List of foodborne illness outbreaks")
* [ death toll ](/wiki/List_of_foodborne_illness_outbreaks_by_death_toll "List of foodborne illness outbreaks by death toll")
* [ United States ](/wiki/List_of_foodborne_illness_outbreaks_in_the_United_States "List of foodborne illness outbreaks in the United States")
[ Regulation ](/wiki/Regulation "Regulation") , [ standards
](/wiki/Standards_organization "Standards organization") , [ watchdogs
](/wiki/List_of_food_safety_organisations "List of food safety organisations")
|
* [ Acceptable daily intake ](/wiki/Acceptable_daily_intake "Acceptable daily intake")
* [ E number ](/wiki/E_number "E number")
* [ Food labeling regulations ](/wiki/List_of_food_labeling_regulations "List of food labeling regulations")
* [ Food libel laws ](/wiki/Food_libel_laws "Food libel laws")
* [ Food safety in Australia ](/wiki/Food_safety_in_Australia "Food safety in Australia")
* [ International Food Safety Network ](/wiki/International_Food_Safety_Network "International Food Safety Network")
* [ ISO 22000 ](/wiki/ISO_22000 "ISO 22000")
* [ Nutrition facts label ](/wiki/Nutrition_facts_label "Nutrition facts label")
* [ Organic certification ](/wiki/Organic_certification "Organic certification")
* [ Quality Assurance International ](/wiki/Quality_Assurance_International "Quality Assurance International")
* [ United Kingdom food information regulations ](/wiki/United_Kingdom_food_information_regulations "United Kingdom food information regulations")
[ Institutions ](/wiki/List_of_food_safety_organisations "List of food safety
organisations") |
* [ Centre for Food Safety ](/wiki/Centre_for_Food_Safety "Centre for Food Safety") (Hong Kong)
* [ European Food Safety Authority ](/wiki/European_Food_Safety_Authority "European Food Safety Authority")
* [ Food and Drug Administration ](/wiki/Food_and_Drug_Administration "Food and Drug Administration")
* [ Food Information and Control Agency ](/wiki/Food_Information_and_Control_Agency "Food Information and Control Agency") (Spain)
* [ Food Standards Agency ](/wiki/Food_Standards_Agency "Food Standards Agency") (United Kingdom)
* [ Institute for Food Safety and Health ](/wiki/Institute_for_Food_Safety_and_Health "Institute for Food Safety and Health")
* [ International Food Safety Network ](/wiki/International_Food_Safety_Network "International Food Safety Network")
* [ Ministry of Food and Drug Safety ](/wiki/Ministry_of_Food_and_Drug_Safety "Ministry of Food and Drug Safety") (South Korea)
* [ Spanish Agency for Food Safety and Nutrition ](/wiki/Spanish_Agency_for_Food_Safety_and_Nutrition "Spanish Agency for Food Safety and Nutrition")
Related topics |
* [ Curing (food preservation) ](/wiki/Curing_\(food_preservation\) "Curing \(food preservation\)")
* [ Food and drink prohibitions ](/wiki/Food_and_drink_prohibitions "Food and drink prohibitions")
* [ Food fraud ](/wiki/Food_fraud "Food fraud")
* [ Food marketing ](/wiki/Food_marketing "Food marketing")
* [ Food politics ](/wiki/Food_politics "Food politics")
* [ Food preservation ](/wiki/Food_preservation "Food preservation")
* [ Food quality ](/wiki/Food_quality "Food quality")
* [ Genetically modified food ](/wiki/Genetically_modified_food "Genetically modified food")
* [ Conspiracy theories ](/wiki/Conspiracy_theories "Conspiracy theories")
* [  ](/wiki/File:Foodlogo2.svg) [ Food portal ](/wiki/Portal:Food "Portal:Food") 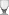 [ Drink portal ](/wiki/Portal:Drink "Portal:Drink")
*  [ Category ](/wiki/Category:Food_safety "Category:Food safety")
*  [ Commons ](https://commons.wikimedia.org/wiki/Category:Food_safety "commons:Category:Food safety")
* [  ](/wiki/File:Wikibooks-logo.svg "Wikibooks page") [ Cookbook ](https://en.wikibooks.org/wiki/Cookbook "wikibooks:Cookbook")
*  [ WikiProject ](/wiki/Wikipedia:WikiProject_Food_and_drink "Wikipedia:WikiProject Food and drink")
|
* [ v ](/wiki/Template:Artificial_foods "Template:Artificial foods")
* [ t ](/wiki/Template_talk:Artificial_foods "Template talk:Artificial foods")
* [ e ](/wiki/Special:EditPage/Template:Artificial_foods "Special:EditPage/Template:Artificial foods")
Artificial foods
---
Artificial [ fat substitutes ](/wiki/Fat_substitute "Fat substitute") |
* [ Olestra ](/wiki/Olestra "Olestra")
Artificial protein substitutes |
* [ Acid-hydrolyzed vegetable protein ](/wiki/Acid-hydrolyzed_vegetable_protein "Acid-hydrolyzed vegetable protein")
Artificial [ sugar substitutes ](/wiki/Sugar_substitute "Sugar substitute") |
* [ Acesulfame potassium ](/wiki/Acesulfame_potassium "Acesulfame potassium")
* [ Alitame ](/wiki/Alitame "Alitame")
* [ Aspartame ](/wiki/Aspartame "Aspartame")
* [ Aspartame-acesulfame salt ](/wiki/Aspartame-acesulfame_salt "Aspartame-acesulfame salt")
* [ Dulcin ](/wiki/Dulcin "Dulcin")
* [ Glucin ](/wiki/Glucin "Glucin")
* [ Hydrogenated starch hydrolysates ](/wiki/Hydrogenated_starch_hydrolysates "Hydrogenated starch hydrolysates")
* [ Neohesperidin dihydrochalcone ](/wiki/Neohesperidin_dihydrochalcone "Neohesperidin dihydrochalcone")
* [ Neotame ](/wiki/Neotame "Neotame")
* [ NutraSweet ](/wiki/NutraSweet "NutraSweet")
* [ Nutrinova ](/wiki/Nutrinova "Nutrinova")
* [ Saccharin ](/wiki/Saccharin "Saccharin")
* [ Sodium cyclamate ](/wiki/Sodium_cyclamate "Sodium cyclamate")
* [ Sucralose ](/wiki/Sucralose "Sucralose")
Natural food substitutes |
* [ Cheese analogues ](/wiki/Cheese_analogue "Cheese analogue")
* [ Coffee substitutes ](/wiki/Coffee_substitute "Coffee substitute")
* [ Egg substitutes ](/wiki/Egg_substitutes "Egg substitutes")
* [ Meat analogues ](/wiki/Meat_analogue "Meat analogue")
* [ bacon ](/wiki/List_of_bacon_substitutes "List of bacon substitutes")
* [ list ](/wiki/List_of_meat_substitutes "List of meat substitutes")
* [ Milk substitutes ](/wiki/Milk_substitute "Milk substitute")
* [ Phyllodulcin ](/wiki/Phyllodulcin "Phyllodulcin")
* [ Salt substitutes ](/wiki/Salt_substitute "Salt substitute")
Brands |
* [ Tang ](/wiki/Tang_\(drink_mix\) "Tang \(drink mix\)")
Related topics |
* [ Food safety ](/wiki/Food_safety "Food safety")
* [ List of food additives ](/wiki/List_of_food_additives "List of food additives")
[ Food politics ](/wiki/Food_politics "Food politics") |
* [ Food power ](/wiki/Food_power "Food power")
* [ Food security ](/wiki/Food_security "Food security")
* [ Famine ](/wiki/Famine "Famine")
* [ Malnutrition ](/wiki/Malnutrition "Malnutrition")
* [ Overnutrition ](/wiki/Overnutrition "Overnutrition")
Institutions |
* [ International Association for Food Protection ](/wiki/International_Association_for_Food_Protection "International Association for Food Protection")
* [ Food and Drug Administration ](/wiki/Food_and_Drug_Administration "Food and Drug Administration")
* [ Food and Agriculture Organization ](/wiki/Food_and_Agriculture_Organization "Food and Agriculture Organization")
* [ National Agriculture and Food Research Organization ](/wiki/National_Agriculture_and_Food_Research_Organization "National Agriculture and Food Research Organization")
* [ National Food and Drug Authority ](/wiki/National_Food_and_Drug_Authority "National Food and Drug Authority")
* [ v ](/wiki/Template:Metabolism "Template:Metabolism")
* [ t ](/wiki/Template_talk:Metabolism "Template talk:Metabolism")
* [ e ](/wiki/Special:EditPage/Template:Metabolism "Special:EditPage/Template:Metabolism")
[ Metabolism ](/wiki/Metabolism "Metabolism") , [ catabolism
](/wiki/Catabolism "Catabolism") , [ anabolism ](/wiki/Anabolism "Anabolism")
---
General |
* [ Metabolic pathway ](/wiki/Metabolic_pathway "Metabolic pathway")
* [ Metabolic network ](/wiki/Metabolic_network "Metabolic network")
* [ Primary nutritional groups ](/wiki/Primary_nutritional_groups "Primary nutritional groups")
[ Energy
metabolism ](/wiki/Bioenergetics "Bioenergetics") |
| [ Aerobic respiration ](/wiki/Aerobic_respiration "Aerobic respiration") |
* [ Glycolysis ](/wiki/Glycolysis "Glycolysis") → [ Pyruvate decarboxylation ](/wiki/Pyruvate_dehydrogenase "Pyruvate dehydrogenase") → [ Citric acid cycle ](/wiki/Citric_acid_cycle "Citric acid cycle") → [ Oxidative phosphorylation ](/wiki/Oxidative_phosphorylation "Oxidative phosphorylation") ( [ electron transport chain ](/wiki/Electron_transport_chain "Electron transport chain") \+ [ ATP synthase ](/wiki/ATP_synthase "ATP synthase") )
---|---
[ Anaerobic respiration ](/wiki/Anaerobic_respiration "Anaerobic respiration")
|
* Electron acceptors other than oxygen
[ Fermentation ](/wiki/Fermentation "Fermentation") |
* [ Glycolysis ](/wiki/Glycolysis "Glycolysis") → [ Substrate-level phosphorylation ](/wiki/Substrate-level_phosphorylation "Substrate-level phosphorylation")
* [ ABE ](/wiki/Acetone%E2%80%93butanol%E2%80%93ethanol_fermentation "Acetone–butanol–ethanol fermentation")
* [ Ethanol ](/wiki/Ethanol_fermentation "Ethanol fermentation")
* [ Lactic acid ](/wiki/Lactic_acid_fermentation "Lactic acid fermentation")
Specific
paths |
| [ Protein metabolism ](/wiki/Protein_metabolism "Protein metabolism") |
* [ Protein synthesis ](/wiki/Protein_biosynthesis "Protein biosynthesis")
* [ Catabolism ](/wiki/Protein_catabolism "Protein catabolism") (protein→peptide→amino acid)
| [ Amino acid ](/wiki/Amino_acid "Amino acid") |
* [ Amino acid synthesis ](/wiki/Amino_acid_synthesis "Amino acid synthesis")
* [ Amino acid degradation ](/wiki/Protein_catabolism#Amino_acid_degradation "Protein catabolism") (amino acid→pyruvate, acetyl CoA, or TCA intermediate)
* [ Urea cycle ](/wiki/Urea_cycle "Urea cycle")
---|---
[ Nucleotide
metabolism ](/wiki/Nucleic_acid_metabolism "Nucleic acid metabolism") |
* [ Purine metabolism ](/wiki/Purine_metabolism "Purine metabolism")
* [ Nucleotide salvage ](/wiki/Nucleotide_salvage "Nucleotide salvage")
* [ Pyrimidine metabolism ](/wiki/Pyrimidine_metabolism "Pyrimidine metabolism")
* [ Purine nucleotide cycle ](/wiki/Purine_nucleotide_cycle "Purine nucleotide cycle")
[ Carbohydrate metabolism ](/wiki/Carbohydrate_metabolism "Carbohydrate
metabolism")
( [ carbohydrate catabolism ](/wiki/Carbohydrate_catabolism "Carbohydrate
catabolism")
and [ anabolism ](/wiki/Anabolism "Anabolism") ) |
| Human |
|
* [ Glycolysis ](/wiki/Glycolysis "Glycolysis") ⇄ [ Gluconeogenesis ](/wiki/Gluconeogenesis "Gluconeogenesis")
---
* [ Glycogenolysis ](/wiki/Glycogenolysis "Glycogenolysis") ⇄ [ Glycogenesis ](/wiki/Glycogenesis "Glycogenesis")
* [ Pentose phosphate pathway ](/wiki/Pentose_phosphate_pathway "Pentose phosphate pathway")
* [ Fructolysis ](/wiki/Fructolysis "Fructolysis")
* [ Polyol pathway ](/wiki/Polyol_pathway "Polyol pathway")
* [ Galactolysis ](/wiki/Galactolysis "Galactolysis")
* [ Leloir pathway ](/wiki/Leloir_pathway "Leloir pathway")
* [ Glycosylation ](/wiki/Glycosylation "Glycosylation")
* [ N-linked ](/wiki/N-linked_glycosylation "N-linked glycosylation")
* [ O-linked ](/wiki/O-linked_glycosylation "O-linked glycosylation")
Nonhuman |
|
* [ Photosynthesis ](/wiki/Photosynthesis "Photosynthesis")
* [ Anoxygenic photosynthesis ](/wiki/Anoxygenic_photosynthesis "Anoxygenic photosynthesis")
* [ Chemosynthesis ](/wiki/Chemosynthesis "Chemosynthesis")
* [ Carbon fixation ](/wiki/Carbon_fixation "Carbon fixation")
* [ DeLey-Doudoroff pathway ](/w/index.php?title=DeLey-Doudoroff_pathway&action=edit&redlink=1 "DeLey-Doudoroff pathway \(page does not exist\)")
* [ Entner-Doudoroff pathway ](/wiki/Entner-Doudoroff_pathway "Entner-Doudoroff pathway")
---
* [ Xylose metabolism ](/wiki/Xylose_metabolism "Xylose metabolism")
* [ Radiotrophism ](/wiki/Radiotrophic_fungus "Radiotrophic fungus")
[ Lipid metabolism ](/wiki/Lipid_metabolism "Lipid metabolism")
( [ lipolysis ](/wiki/Lipolysis "Lipolysis") , [ lipogenesis
](/wiki/Lipogenesis "Lipogenesis") ) |
| [ Fatty acid metabolism ](/wiki/Fatty_acid_metabolism "Fatty acid
metabolism") |
* [ Fatty acid degradation ](/wiki/Fatty_acid_degradation "Fatty acid degradation") ( [ Beta oxidation ](/wiki/Beta_oxidation "Beta oxidation") )
* [ Fatty acid synthesis ](/wiki/Fatty_acid_synthesis "Fatty acid synthesis")
---|---
Other |
* [ Steroid metabolism ](/wiki/Steroid "Steroid")
* [ Sphingolipid metabolism ](/wiki/Sphingolipid_metabolism "Sphingolipid metabolism")
* [ Eicosanoid metabolism ](/wiki/Eicosanoid_metabolism "Eicosanoid metabolism")
* [ Ketosis ](/wiki/Ketosis "Ketosis")
* [ Reverse cholesterol transport ](/wiki/Reverse_cholesterol_transport "Reverse cholesterol transport")
Other |
* [ Metal metabolism ](/wiki/Bioinorganic_chemistry "Bioinorganic chemistry")
* [ Iron metabolism ](/wiki/Human_iron_metabolism "Human iron metabolism")
* [ Ethanol metabolism ](/wiki/Ethanol_metabolism "Ethanol metabolism")
* [ Phospagen system (ATP-PCr) ](/wiki/Phosphagen "Phosphagen")
* [ v ](/wiki/Template:MetabolismMap "Template:MetabolismMap")
* [ t ](/wiki/Template_talk:MetabolismMap "Template talk:MetabolismMap")
* [ e ](/wiki/Special:EditPage/Template:MetabolismMap "Special:EditPage/Template:MetabolismMap")
[ Metabolism ](/wiki/Metabolism "Metabolism") map
---
|
[ 
](/wiki/File:Metabolic_Metro_Map.svg "File:Metabolic Metro Map.svg")
_[ Carbon
fixation ](/wiki/Carbon_fixation "Carbon fixation") _
_[ Photo-
respiration ](/wiki/Photorespiration "Photorespiration") _
_[ Pentose
phosphate
pathway ](/wiki/Pentose_phosphate_pathway "Pentose phosphate pathway") _
_[ Citric
acid cycle ](/wiki/Citric_acid_cycle "Citric acid cycle") _
_[ Glyoxylate
cycle ](/wiki/Glyoxylate_cycle "Glyoxylate cycle") _
_[ Urea
cycle ](/wiki/Urea_cycle "Urea cycle") _
_[ Fatty
acid
synthesis ](/wiki/Fatty_acid_synthesis "Fatty acid synthesis") _
_[ Fatty
acid
elongation ](/wiki/Fatty_acid_synthesis "Fatty acid synthesis") _
_[ Beta
oxidation ](/wiki/Beta_oxidation "Beta oxidation") _
_[ Peroxisomal ](/wiki/Beta_oxidation#Peroxisomal_beta-oxidation "Beta
oxidation") _
_[ beta
oxidation ](/wiki/Beta_oxidation#Peroxisomal_beta-oxidation "Beta oxidation")
_
_**[ Glyco-
genolysis ](/wiki/Glycogenolysis "Glycogenolysis") ** _
_**[ Glyco-
genesis ](/wiki/Glycogenesis "Glycogenesis") ** _
_**[ Glyco-
lysis ](/wiki/Glycolysis "Glycolysis") ** _
_**[ Gluconeo-
genesis ](/wiki/Gluconeogenesis "Gluconeogenesis") ** _
_**[ Pyruvate
decarb-
oxylation ](/wiki/Pyruvate_dehydrogenase "Pyruvate dehydrogenase") ** _
_**[ Fermentation ](/wiki/Lactic_acid_fermentation "Lactic acid
fermentation") ** _
_**[ Keto-
lysis ](/wiki/Ketone_bodies "Ketone bodies") ** _
_**[ Keto-
genesis ](/wiki/Ketogenesis "Ketogenesis") ** _
_**[ feeders to
gluconeo-
genesis ](/wiki/Gluconeogenesis "Gluconeogenesis") ** _
_**[ Direct / C4 / CAM
carbon intake ](/wiki/Carbon_fixation#Carbon_concentrating_mechanisms "Carbon
fixation") ** _
_**[ Light reaction ](/wiki/Light-dependent_reactions "Light-dependent
reactions") ** _
_**[ Oxidative
phosphorylation ](/wiki/Oxidative_phosphorylation "Oxidative
phosphorylation") ** _
_**[ Amino acid
deamination ](/wiki/Deamination "Deamination") ** _
_**[ Citrate
shuttle
](/wiki/Fatty_acid_synthesis#Glycolytic_end_products_are_used_in_the_conversion_of_carbohydrates_into_fatty_acids
"Fatty acid synthesis") ** _
_**[ Lipogenesis ](/wiki/Lipogenesis "Lipogenesis") ** _
_**[ Lipolysis ](/wiki/Lipolysis "Lipolysis") ** _
_**[ Steroidogenesis ](/wiki/Steroid#Steroidogenesis "Steroid") ** _
_**[ MVA pathway ](/wiki/Mevalonate_pathway "Mevalonate pathway") ** _
_**[ MEP pathway ](/wiki/Non-mevalonate_pathway "Non-mevalonate pathway") **
_
_**[ Shikimate
pathway ](/wiki/Shikimate_pathway "Shikimate pathway") ** _
_**[ Transcription & ](/wiki/Transcription_\(genetics\) "Transcription
\(genetics\)")
[ replication ](/wiki/DNA_replication "DNA replication") ** _
_**[ Translation ](/wiki/Translation_\(biology\) "Translation \(biology\)")
** _
_**[ Proteolysis ](/wiki/Proteolysis "Proteolysis") ** _
_**[ Glycosyl-
ation ](/wiki/Glycosylation "Glycosylation") ** _
[ Sugar
acids ](/wiki/Sugar_acid "Sugar acid")
[ Double ](/wiki/Disaccharide "Disaccharide") / [ multiple
sugars ](/wiki/Polysaccharide "Polysaccharide") & [ glycans ](/wiki/Glycan
"Glycan")
[ Simple
sugars ](/wiki/Monosaccharide "Monosaccharide")
[ Inositol-P ](/wiki/Inositol_phosphate "Inositol phosphate")
[ Amino sugars ](/wiki/Amino_sugar "Amino sugar")
& [ sialic acids ](/wiki/Sialic_acid "Sialic acid")
[ Nucleotide sugars ](/wiki/Nucleotide_sugar "Nucleotide sugar")
[ Hexose-P ](/wiki/Glucose_6-phosphate "Glucose 6-phosphate")
[ Triose-P ](/wiki/Glyceraldehyde_3-phosphate "Glyceraldehyde 3-phosphate")
[ Glycerol ](/wiki/Glycerol "Glycerol")
[ P-glycerates ](/wiki/Phosphoglycerate "Phosphoglycerate")
[ Pentose-P ](/wiki/Ribose_5-phosphate "Ribose 5-phosphate")
[ Tetrose-P ](/wiki/Erythrose_4-phosphate "Erythrose 4-phosphate")
[ Propionyl
-CoA ](/wiki/Propionyl-CoA "Propionyl-CoA")
[ Succinate ](/wiki/Succinic_acid "Succinic acid")
[ Acetyl
-CoA ](/wiki/Acetyl-CoA "Acetyl-CoA")
[ Pentose-P ](/wiki/Ribose_5-phosphate "Ribose 5-phosphate")
[ P-glycerates ](/wiki/Phosphoglycerate "Phosphoglycerate")
[ Glyoxylate ](/wiki/Glyoxylic_acid "Glyoxylic acid")
[ Photosystems ](/wiki/Photosystem "Photosystem")
[ Pyruvate ](/wiki/Pyruvic_acid "Pyruvic acid")
[ Lactate ](/wiki/Lactic_acid "Lactic acid")
[ Acetyl
-CoA ](/wiki/Acetyl-CoA "Acetyl-CoA")
[ Citrate ](/wiki/Citric_acid "Citric acid")
[ Oxalo-
acetate ](/wiki/Oxaloacetic_acid "Oxaloacetic acid")
[ Malate ](/wiki/Malic_acid "Malic acid")
[ Succinyl
-CoA ](/wiki/Succinyl-CoA "Succinyl-CoA")
[ α-Keto-
glutarate ](/wiki/Alpha-Ketoglutaric_acid "Alpha-Ketoglutaric acid")
[ Ketone
bodies ](/wiki/Ketone_bodies "Ketone bodies")
[ Respiratory
chain ](/wiki/Electron_transport_chain "Electron transport chain")
[ Serine group ](/wiki/Serine "Serine")
[ Alanine ](/wiki/Alanine "Alanine")
[ Branched-chain
amino acids ](/wiki/Branched-chain_amino_acid "Branched-chain amino acid")
[ Aspartate
group ](/wiki/Aspartic_acid "Aspartic acid")
[ Homoserine
group ](/wiki/Homoserine "Homoserine")
& [ lysine ](/wiki/Lysine "Lysine")
[ Glutamate
group ](/wiki/Glutamic_acid "Glutamic acid")
& [ proline ](/wiki/Proline "Proline")
[ Arginine ](/wiki/Arginine "Arginine")
[ Creatine ](/wiki/Creatine "Creatine")
& [ polyamines ](/wiki/Polyamine "Polyamine")
[ Ketogenic ](/wiki/Ketogenic_amino_acid "Ketogenic amino acid") &
[ glucogenic
amino acids ](/wiki/Glucogenic_amino_acid "Glucogenic amino acid")
[ Amino acids ](/wiki/Amino_acid "Amino acid")
[ Shikimate ](/wiki/Shikimic_acid "Shikimic acid")
[ Aromatic amino
acids ](/wiki/Aromatic_amino_acid "Aromatic amino acid") & [ histidine
](/wiki/Histidine "Histidine")
[ Ascorbate ](/wiki/Ascorbate "Ascorbate")
( [ vitamin C ](/wiki/Vitamin_C "Vitamin C") )
[ δ-ALA ](/wiki/Aminolevulinic_acid "Aminolevulinic acid")
[ Bile
pigments ](/wiki/Bile_pigment "Bile pigment")
[ Hemes ](/wiki/Heme "Heme")
[ Cobalamins ](/wiki/Cobalamin "Cobalamin") ( [ vitamin B 12
](/wiki/Vitamin_B "Vitamin B") )
[ Various
vitamin Bs ](/wiki/Vitamin_B "Vitamin B")
[ Calciferols ](/wiki/Calciferol "Calciferol")
( [ vitamin D ](/wiki/Vitamin_D "Vitamin D") )
[ Retinoids ](/wiki/Retinoid "Retinoid")
( [ vitamin A ](/wiki/Vitamin_A "Vitamin A") )
[ Quinones ](/wiki/Quinone "Quinone") ( [ vitamin K ](/wiki/Vitamin_K "Vitamin
K") )
& [ tocopherols ](/wiki/Tocopherol "Tocopherol") ( [ vitamin E
](/wiki/Vitamin_E "Vitamin E") )
[ Cofactors ](/wiki/Cofactor_\(biochemistry\) "Cofactor \(biochemistry\)")
[ Vitamins ](/wiki/Vitamin "Vitamin")
& [ minerals ](/wiki/Metalloprotein "Metalloprotein")
[ Antioxidants ](/wiki/Antioxidant "Antioxidant")
[ PRPP ](/wiki/Phosphoribosyl_pyrophosphate "Phosphoribosyl pyrophosphate")
[ Nucleotides ](/wiki/Nucleotide "Nucleotide")
[ Nucleic
acids ](/wiki/Nucleic_acid "Nucleic acid")
[ Proteins ](/wiki/Protein "Protein")
[ Glycoproteins ](/wiki/Glycoprotein "Glycoprotein")
& [ proteoglycans ](/wiki/Proteoglycan "Proteoglycan")
[ Chlorophylls ](/wiki/Chlorophyll "Chlorophyll")
[ MEP ](/wiki/Methylerythritol_phosphate "Methylerythritol phosphate")
[ MVA ](/wiki/Mevalonic_acid "Mevalonic acid")
[ Acetyl
-CoA ](/wiki/Acetyl-CoA "Acetyl-CoA")
[ Polyketides ](/wiki/Polyketide "Polyketide")
[ Terpenoid
backbones ](/wiki/Isopentenyl_pyrophosphate "Isopentenyl pyrophosphate")
[ Terpenoids ](/wiki/Terpenoid "Terpenoid")
& [ carotenoids ](/wiki/Carotenoid "Carotenoid") ( [ vitamin A
](/wiki/Vitamin_A "Vitamin A") )
[ Cholesterol ](/wiki/Cholesterol "Cholesterol")
[ Bile acids ](/wiki/Bile_acid "Bile acid")
[ Glycero-
phospholipids ](/wiki/Glycerophospholipid "Glycerophospholipid")
[ Glycerolipids ](/wiki/Glycerolipid "Glycerolipid")
[ Acyl-CoA ](/wiki/Acyl-CoA "Acyl-CoA")
[ Fatty
acids ](/wiki/Fatty_acid "Fatty acid")
[ Glyco-
sphingolipids ](/wiki/Glycosphingolipid "Glycosphingolipid")
[ Sphingolipids ](/wiki/Sphingolipid "Sphingolipid")
[ Waxes ](/wiki/Wax "Wax")
[ Polyunsaturated
fatty acids ](/wiki/Polyunsaturated_fatty_acid "Polyunsaturated fatty acid")
[ Neurotransmitters ](/wiki/Neurotransmitters "Neurotransmitters")
& [ thyroid hormones ](/wiki/Thyroid_hormone "Thyroid hormone")
[ Steroids ](/wiki/Steroid "Steroid")
[ Endo-
cannabinoids ](/wiki/Endocannabinoid_system "Endocannabinoid system")
[ Eicosanoids ](/wiki/Eicosanoid "Eicosanoid")
---

Major [ metabolic pathways ](/wiki/Metabolic_pathway "Metabolic pathway") in [
metro-style map ](/wiki/Transit_map "Transit map") . Click any text (name of
pathway or metabolites) to link to the corresponding article.
Single lines: pathways common to most lifeforms. Double lines: pathways not in
humans (occurs in e.g. plants, fungi, prokaryotes).
 Orange nodes: [ carbohydrate metabolism
](/wiki/Carbohydrate_metabolism "Carbohydrate metabolism") .
 Violet nodes: [ photosynthesis
](/wiki/Photosynthesis "Photosynthesis") .
 Red nodes: [ cellular respiration
](/wiki/Cellular_respiration "Cellular respiration") .
 Pink nodes: [ cell signaling
](/wiki/Cell_signaling "Cell signaling") .
 Blue nodes: [ amino acid metabolism
](/wiki/Amino_acid_metabolism "Amino acid metabolism") .
 Grey nodes: [ vitamin ](/wiki/Vitamin "Vitamin")
and [ cofactor ](/wiki/Cofactor_\(biochemistry\) "Cofactor \(biochemistry\)")
metabolism.
 Brown nodes: [ nucleotide
](/wiki/Nucleic_acid_metabolism "Nucleic acid metabolism") and [ protein
](/wiki/Protein_metabolism "Protein metabolism") metabolism.
 Green nodes: [ lipid metabolism
](/wiki/Lipid_metabolism "Lipid metabolism") .
[ Authority control databases ](/wiki/Help:Authority_control "Help:Authority
control") [ 
](https://www.wikidata.org/wiki/Q80863#identifiers "Edit this at Wikidata")
---
International |
* [ FAST ](http://id.worldcat.org/fast/796573/)
National |
* [ France ](https://catalogue.bnf.fr/ark:/12148/cb122660296)
* [ BnF data ](https://data.bnf.fr/ark:/12148/cb122660296)
* [ Germany ](https://d-nb.info/gnd/4143345-2)
* [ Israel ](http://olduli.nli.org.il/F/?func=find-b&local_base=NLX10&find_code=UID&request=987007292950205171)
* [ United States ](https://id.loc.gov/authorities/sh85000850)
* [ Japan ](https://id.ndl.go.jp/auth/ndlna/00560059)
* [ Czech Republic ](https://aleph.nkp.cz/F/?func=find-c&local_base=aut&ccl_term=ica=ph880313&CON_LNG=ENG)

Retrieved from " [
https://en.wikipedia.org/w/index.php?title=Adenosine_triphosphate&oldid=1219758132
](https://en.wikipedia.org/w/index.php?title=Adenosine_triphosphate&oldid=1219758132)
"
[ Categories ](/wiki/Help:Category "Help:Category") :
* [ Adenosine receptor agonists ](/wiki/Category:Adenosine_receptor_agonists "Category:Adenosine receptor agonists")
* [ Cellular respiration ](/wiki/Category:Cellular_respiration "Category:Cellular respiration")
* [ Coenzymes ](/wiki/Category:Coenzymes "Category:Coenzymes")
* [ Ergogenic aids ](/wiki/Category:Ergogenic_aids "Category:Ergogenic aids")
* [ Exercise physiology ](/wiki/Category:Exercise_physiology "Category:Exercise physiology")
* [ Neurotransmitters ](/wiki/Category:Neurotransmitters "Category:Neurotransmitters")
* [ Nucleotides ](/wiki/Category:Nucleotides "Category:Nucleotides")
* [ Phosphate esters ](/wiki/Category:Phosphate_esters "Category:Phosphate esters")
* [ Purinergic signalling ](/wiki/Category:Purinergic_signalling "Category:Purinergic signalling")
* [ Purines ](/wiki/Category:Purines "Category:Purines")
* [ Substances discovered in the 1920s ](/wiki/Category:Substances_discovered_in_the_1920s "Category:Substances discovered in the 1920s")
Hidden categories:
* [ CS1: long volume value ](/wiki/Category:CS1:_long_volume_value "Category:CS1: long volume value")
* [ CS1 German-language sources (de) ](/wiki/Category:CS1_German-language_sources_\(de\) "Category:CS1 German-language sources \(de\)")
* [ Articles with short description ](/wiki/Category:Articles_with_short_description "Category:Articles with short description")
* [ Short description is different from Wikidata ](/wiki/Category:Short_description_is_different_from_Wikidata "Category:Short description is different from Wikidata")
* [ Chemical articles with multiple compound IDs ](/wiki/Category:Chemical_articles_with_multiple_compound_IDs "Category:Chemical articles with multiple compound IDs")
* [ Multiple chemicals in an infobox that need indexing ](/wiki/Category:Multiple_chemicals_in_an_infobox_that_need_indexing "Category:Multiple chemicals in an infobox that need indexing")
* [ Chemical articles with multiple CAS registry numbers ](/wiki/Category:Chemical_articles_with_multiple_CAS_registry_numbers "Category:Chemical articles with multiple CAS registry numbers")
* [ Articles without InChI source ](/wiki/Category:Articles_without_InChI_source "Category:Articles without InChI source")
* [ Articles with changed CASNo identifier ](/wiki/Category:Articles_with_changed_CASNo_identifier "Category:Articles with changed CASNo identifier")
* [ ECHA InfoCard ID from Wikidata ](/wiki/Category:ECHA_InfoCard_ID_from_Wikidata "Category:ECHA InfoCard ID from Wikidata")
* [ Articles containing unverified chemical infoboxes ](/wiki/Category:Articles_containing_unverified_chemical_infoboxes "Category:Articles containing unverified chemical infoboxes")
* [ Chembox image size set ](/wiki/Category:Chembox_image_size_set "Category:Chembox image size set")
* [ All articles with unsourced statements ](/wiki/Category:All_articles_with_unsourced_statements "Category:All articles with unsourced statements")
* [ Articles with unsourced statements from December 2023 ](/wiki/Category:Articles_with_unsourced_statements_from_December_2023 "Category:Articles with unsourced statements from December 2023")
* [ Articles with unsourced statements from April 2023 ](/wiki/Category:Articles_with_unsourced_statements_from_April_2023 "Category:Articles with unsourced statements from April 2023")
* [ Commons category link is on Wikidata ](/wiki/Category:Commons_category_link_is_on_Wikidata "Category:Commons category link is on Wikidata")
* [ Articles with FAST identifiers ](/wiki/Category:Articles_with_FAST_identifiers "Category:Articles with FAST identifiers")
* [ Articles with BNF identifiers ](/wiki/Category:Articles_with_BNF_identifiers "Category:Articles with BNF identifiers")
* [ Articles with BNFdata identifiers ](/wiki/Category:Articles_with_BNFdata_identifiers "Category:Articles with BNFdata identifiers")
* [ Articles with GND identifiers ](/wiki/Category:Articles_with_GND_identifiers "Category:Articles with GND identifiers")
* [ Articles with J9U identifiers ](/wiki/Category:Articles_with_J9U_identifiers "Category:Articles with J9U identifiers")
* [ Articles with LCCN identifiers ](/wiki/Category:Articles_with_LCCN_identifiers "Category:Articles with LCCN identifiers")
* [ Articles with NDL identifiers ](/wiki/Category:Articles_with_NDL_identifiers "Category:Articles with NDL identifiers")
* [ Articles with NKC identifiers ](/wiki/Category:Articles_with_NKC_identifiers "Category:Articles with NKC identifiers")
* This page was last edited on 19 April 2024, at 17:08 (UTC) .
* Text is available under the [ Creative Commons Attribution-ShareAlike License 4.0 ](//en.wikipedia.org/wiki/Wikipedia:Text_of_the_Creative_Commons_Attribution-ShareAlike_4.0_International_License) [ ](//en.wikipedia.org/wiki/Wikipedia:Text_of_the_Creative_Commons_Attribution-ShareAlike_4.0_International_License) ; additional terms may apply. By using this site, you agree to the [ Terms of Use ](//foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Terms_of_Use) and [ Privacy Policy ](//foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Privacy_policy) . Wikipedia® is a registered trademark of the [ Wikimedia Foundation, Inc. ](//www.wikimediafoundation.org/) , a non-profit organization.
* [ Privacy policy ](https://foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Privacy_policy)
* [ About Wikipedia ](/wiki/Wikipedia:About)
* [ Disclaimers ](/wiki/Wikipedia:General_disclaimer)
* [ Contact Wikipedia ](//en.wikipedia.org/wiki/Wikipedia:Contact_us)
* [ Code of Conduct ](https://foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Universal_Code_of_Conduct)
* [ Developers ](https://developer.wikimedia.org)
* [ Statistics ](https://stats.wikimedia.org/#/en.wikipedia.org)
* [ Cookie statement ](https://foundation.wikimedia.org/wiki/Special:MyLanguage/Policy:Cookie_statement)
* [ Mobile view ](//en.m.wikipedia.org/w/index.php?title=Adenosine_triphosphate&mobileaction=toggle_view_mobile)
* [  ](https://wikimediafoundation.org/)
* [  ](https://www.mediawiki.org/)
* Toggle limited content width
*[
CID
]: Compound ID
*[
EPA
]: U.S. Environmental Protection Agency
*[
v
]: View this template
*[
t
]: Discuss this template
*[
e
]: Edit this template
*[
v
]: View this template
*[
t
]: Discuss this template
*[
e
]: Edit this template
| biology | 8401598 | https://sv.wikipedia.org/wiki/Lista%20%C3%B6ver%20f%C3%A5gelarter%20beskrivna%20under%202010-talet | Lista över fågelarter beskrivna under 2010-talet | Detta är en lista över fågelarter som beskrivits som nya för vetenskapen under perioden 2010−2019.
Sammanfattande statistik
Antal beskrivna arter per år
Länder med större antal nya beskrivna arter
Brasilien
Peru
Filippinerna
Indonesien
Arter, år för år
2010
Karstsångare (Phylloscopus calciatilis):
Urraomyrpitta (Grallaria fenwickorum):
Sokotravråk (Buteo socotraensis):
Willardbusktörnskata (Laniarius willardi):
Klipptapakul (Scytalopus petrophilus):
2011
Tsingyskogsrall (Canirallus beankaensis):
Bryanlira (Puffinus bryani):
Várzeatrast (Turdus sanchezorum):
2012
Altaflorestamyrpitta (Hylopezus whittakeri):
Caucagärdsmyg (Thryophilus sernai):
Sirabarbett (Capito fitzpatricki):
†Bermudabusksparv (Pipilo naufragus):
Cinclodes espinhacensis: . Sammanslagen med långstjärtad cinklod (Cinclodes pabsti) år 2013
Camiguinspökuggla (Ninox leventisi):. Fördes tidigare arten till luzonspökuggla (Ninox philippensis).
Cebuspökuggla (Ninox rumseyi): Fördes tidigare arten till luzonspökuggla (Ninox philippensis).
2013
Lombokdvärguv (Otus jolandae):
Pincoyastormsvala (Oceanites pincoyae):
Deltaamacuromjukstjärt (Thripophaga amacurensis):
†Bermudaspett (Colaptes oceanicus):
†Azordvärguv (Otus frutuosi):
"Seramtornuggla" (Tyto almae):
Juníntapakul (Scytalopus gettyae):
Kambodjaskräddarfågel (Orthotomus chaktomuk):
Tropeirofröfink (Sporophila beltoni):
Sierramadregräsfågel (Robsonius thompsoni):
Arremon kuehnerii:
Strix omanensis: Kategoriseras idag som synonym till Strix butleri.
†Nyakaledonienbeckasin (Coenocorypha neocaledonica):
Följande 15 brasilianska arter beskrevs i den 17:e volymen av Handbook of the Birds of the World:
Obamatrögfågel (Nystalus obamai)
Dendrocolaptes retentus - förs ofta till vattrad trädklättrare
Inambariträdklättrare (Lepidocolaptes fatimalimae)
Campylorhamphus gyldenstolpei - förs oftast till amazonskärnäbb
Campylorhamphus cardosoi - förs oftast till amazonskärnäbb
Madeiramyrsmyg (Epinecrophylla dentei)
Myrmotherula oreni - först oftast till iheringmyrsmyg
Purusmyrsmyg (Herpsilochmus praedictus)
Aripuanãmyrsmyg (Herpsilochmus stotzi)
Manicorédrillmyrfågel (Hypocnemis rondoni)
Chicodvärgtyrann (Zimmerius chicomendesi)
Acretodityrann (Hemitriccus cohnhafti)
Zimmerflatnäbb (Tolmomyias sucunduri)
Inambarimyggsnappare (Polioptila attenboroughi)
"Campinaskrika" (Cyanocorax hafferi)
2014
Formicivora paludicola:
Dicaeum kuehni: Förs ofta till sulawesiblomsterpickare.
Alagoasträdletare (Cichlocolaptes mazarbarnetti)
Boanovatapakul (Scytalopus gonzagai):
Sodhiflugsnappare (Muscicapa sodhii):
2015
Västlig klippuggla (Strix hadorami):
Perijátapakul (Scytalopus perijanus):
Sichuansmygsångare (Locustella chengi):
2016
Himalayatrast (Zoothera salimalii):
Stiphrornis dahomeyensis: Voelker, G.; Tobler, M.; Prestridge, H. L.; Duijm, E.; Groenenberg, D.; Hutchinson, M. R.; Martin, A. D.; Nieman, A.; Roselaar, C. S.; Huntley, J. W. (2016). "Three new species of Stiphrornis (Aves: Muscicapidae) from the Afro-tropics, with a molecular phylogenetic assessment of the genus". Systematics and Biodiversity. doi:10.1080/14772000.2016.1226978 Tidigare förd till skogsskvätta.
Stiphrornis inexpectatus: Voelker, G.; Tobler, M.; Prestridge, H. L.; Duijm, E.; Groenenberg, D.; Hutchinson, M. R.; Martin, A. D.; Nieman, A.; Roselaar, C. S.; Huntley, J. W. (2016). "Three new species of Stiphrornis (Aves: Muscicapidae) from the Afro-tropics, with a molecular phylogenetic assessment of the genus". Systematics and Biodiversity. doi:10.1080/14772000.2016.1226978. Tidigare förd till skogsskvätta.
Stiphrornis rudderi: Voelker, G.; Tobler, M.; Prestridge, H. L.; Duijm, E.; Groenenberg, D.; Hutchinson, M. R.; Martin, A. D.; Nieman, A.; Roselaar, C. S.; Huntley, J. W. (2016). "Three new species of Stiphrornis (Aves: Muscicapidae) from the Afro-tropics, with a molecular phylogenetic assessment of the genus". Systematics and Biodiversity. doi:10.1080/14772000.2016.1226978 Tidigare förd till Stiphrornis xanthogaster
"Iberafrötangara" (Sporophila iberaensis): Adrian Di Giacomo, Bernabe López-Lanús and Cecilia Kopuchian. 2017. A New Species of Seedeater (Emberizidae: Sporophila) from the Iberá grasslands, in northeast Argentina. bioRxiv. 046318. doi:10.1101/046318
2017
Tatamátapakul (Scytalopus alvarezlopezi): Stiles, F. Gary; Laverde-R., Oscar; Cadena, Carlos Daniel (2017). "A new species of tapaculo (Rhinocryptidae: Scytalopus) from the Western Andes of Colombia". The Auk. 134 (2): 377–392. doi:10.1642/AUK-16-205.1.
Sholicola ashambuensis: Robin, V., Vishnudas, C.K., Gupta, P. et al. Two new genera of songbirds represent endemic radiations from the Shola Sky Islands of the Western Ghats, India. BMC Evol Biol 17, 31 (2017) doi:10.1186/s12862-017-0882-6. Kategoriseras idag som underart till keralaflugsnappare.
Amazona gomezgarzai: Silva, Tony; Guzmán, Antonio; Urantówka, Adam D.; Mackiewicz, Paweł (2017). "A new parrot taxon from the Yucatán Peninsula, Mexico—its position within genus Amazona based on morphology and molecular phylogeny". PeerJ. 5: e3475. doi:10.7717/peerj.3475
Brokmanakin (Machaeropterus eckelberryi): Daniel F. Lane, Andrew W. Kratter, John P. O’Neill. A new species of manakin (Aves: Pipridae; Machaeropterus) from Peru with a taxonomic reassessment of the Striped Manakin (M. regulus) complex. Zootaxa, 2017; 4320 (2): 379 DOI: 10.11646/zootaxa.4320.2.11
Karstsabelvinge (Campylopterus calcirupicola: Lopes, Leonardo; Ferreira de Vasconcelos, Marcelo; Gonzaga, Luiz (2017-05-15). "A cryptic new species of hummingbird of the Campylopterus largipennis complex (Aves: Trochilidae)". Zootaxa. 4268: 1–33. doi:10.11646/zootaxa.4268.1.1
†Graciosadomherre (Pyrrhula crassa): J. C. Rando, H. Pieper, Storrs L. Olson, F. Pereira and J. A. Alcover. 2017. A New Extinct Species of Large Bullfinch (Aves: Fringillidae: Pyrrhula) from Graciosa Island (Azores, North Atlantic Ocean). Zootaxa. 4282(3); 567–583. DOI: 10.11646/zootaxa.4282.3.9
Rotemyzomela (Myzomela irianawidodoae): Dewi Malia Prawiradilaga, Pratibha Baveja, Suparno, Hidayat Ashari, Nathaniel Sheng Rong Ng, Chyi Yin Gwee, Philippe Verbelen and Frank Erwin Rheindt (2017). A colorful new species of Myzomela Honeyeater from Rote Island in Eastern Indonesia. Treubia. Number: 44. Pages: 77-100. December. http://e-journal.biologi.lipi.go.id/index.php/treubia/article/view/3414
Santamartaskrikuv (Megascops gilesi): Krabbe, N. K. (2017). "A new species of Megascops (Strigidae) from the Sierra Nevada de Santa Marta, Colombia, with notes on voices of New World screech-owls". Ornitología Colombiana. 16: 1–27.
2018
Gråbrynad myrfågel (Myrmoderus eowilsoni): Andre E. Moncrieff, Oscar Johnson, Daniel F. Lane, Josh R. Beck, Fernando Angulo and Jesse Fagan. 2018. A New Species of Antbird (Passeriformes: Thamnophilidae) from the Cordillera Azul, San Martín, Peru [Una nueva especie de hormiguero (Passeriformes: Thamnophilidae) de la Cordillera Azul, San Martín, Perú]. The Auk. 135(1); 114-126. DOI: 10.1642/AUK-17-97.1
Pelecanoides whenuahouensis: Fischer, Johannes H.; Debski, Igor; Miskelly, Colin M.; Bost, Charles A.; Fromant, Aymeric; Tennyson, Alan J. D.; Tessler, Jake; Cole, Rosalind; Hiscock, Johanna H. (2018-06-27). "Analyses of phenotypic differentiations among South Georgian Diving Petrel (Pelecanoides georgicus) populations reveal an undescribed and highly endangered species from New Zealand". PLOS ONE. 13(6): e0197766. doi:10.1371/journal.pone.0197766. ISSN 1932-6203
Västafrikansk drongo (Dicrurus occidentalis): Fuchs, Jérôme; DOUNO, MORY; Bowie, Rauri; Fjeldså, Jon (2018-06-20). "Taxonomic revision of the Square-tailed Drongo species complex (Passeriformes: Dicruridae) with description of a new species from western Africa". Zootaxa. 4438: 105. doi:10.11646/zootaxa.4438.1.4
Newtonia lavarambo: Younger, Jane & Strozier, Lynika & Maddox, J. Dylan & Nyári, Árpád & Bonfitto, Matthew & Raherilalao, Marie Jeanne & Goodman, Steven & Reddy, Sushma. (2018). Hidden diversity of forest birds in Madagascar revealed using integrative taxonomy. Molecular Phylogenetics and Evolution. 124. doi: 10.1016/j.ympev.2018.02.017. Behandlas oftast som underart till mörk newtonia
Blåstrupig bergstjärna (Oreotrochilus cyanolaemus): Francisco Sornoza-Molina, Juan F. Freile, Jonas Nilsson, Niels Krabbe and Elisa Bonaccorso. 2018. A Striking, Critically Endangered, New Species of Hillstar (Trochilidae: Oreotrochilus) from the southwestern Andes of Ecuador [Una sorprendente y críticamente amenazada especie nueva de estrella (Trochilidae: Oreotrochilus) de los Andes suroccidentales de Ecuador]. The Auk. 135(4);1146-1171. doi: 10.1642/AUK-18-58.1
Rotesångare (Phylloscopus rotiensis): Ng, Nathaniel. S. R.; Prawiradilaga, Dewi. M.; Ng, Elize. Y. X.; Suparno; Ashari, Hidayat; Trainor, Colin; Verbelen, Philippe; Rheindt, Frank. E. (2018-10-23). "A striking new species of leaf warbler from the Lesser Sundas as uncovered through morphology and genomics". Scientific Reports. 8 (1). doi:10.1038/s41598-018-34101-7. ISSN 2045-2322.
2019
Gräddögd bulbyl (Pycnonotus pseudosimplex): Subir B. Shakya, Haw Chuan Lim, Robert G. Moyle, Mustafa Abdul Rahman, Maklarin Lakim and Frederick H. Sheldon. 2019. A Cryptic New Species of Bulbul from Borneo. Bulletin of the British Ornithologists’ Club. 139(1) DOI 10.25226/bboc.v139i1.2019.a3 ISSN 0007-1595
Cercococcyx lemaireae: Peter Boesman and N. J. Collar "Two undescribed species of bird from West Africa," Bulletin of the British Ornithologists’ Club 139(2), 147-159, (17 June 2019). https://doi.org/10.25226/bboc.v139i2.2019.a7
Buccanodon dowsetti: Peter Boesman and N. J. Collar "Two undescribed species of bird from West Africa," Bulletin of the British Ornithologists’ Club 139(2), 147-159, (17 June 2019). https://doi.org/10.25226/bboc.v139i2.2019.a7
Alormyzomela (Myzomela prawiradilagae): Mohammad Irham, Hidayat Ashari, Suparno, Colin R. Trainor, Philippe Verbelen, Meng Yue Wu and Frank E. Rheindt. 2019. A New Myzomela Honeyeater (Meliphagidae) from the Highlands of Alor Island, Indonesia. Journal of Ornithology. DOI: 10.1007/s10336-019-01722-2 Dajakblomsterpickare (Dicaeum dayakorum): Jacob R. Saucier, Christopher M. Milensky, Marcos A. Caraballo-Ortiz, Roslina Ragai, N. Faridah Dahlan and David P. Edwards. 2019. A Distinctive New Species of Flowerpecker (Passeriformes: Dicaeidae) from Borneo. Zootaxa. 4686(4); 451–464. DOI: 10.11646/zootaxa.4686.4.1 Beskrivna under denna period, men som ej längre behandlas som egna arter
Cinclodes espinhacensis beskriven 2012. Behandlas som underart till långstjärtad cinklod
Strix omanensis, beskriven 2013. Kategoriseras idag som synonym till Strix butleri.
Amazona gomezgarzai, beskriven 2017. Anses idag utgör en hybrid.
Sholicola ashambuensis, beskriven 2017. Kategoriseras idag som underart till keralaflugsnappare
Newtonia lavarambo, beskriven 2018. Kategoriseras oftast som underart till mörk newtonia
Noter Texten bygger på en översättning av engelskspråkiga wikipedias artikel List of bird species described in the 2010s, läst 8 januari 2020''
Se även
Lista över fågelarter beskrivna under 2020-talet
Fåglar
Listor med anknytning till biologi | swedish | 0.824024 |
induce_sleep/adenosinetriphosphat.txt | Skip to Main content
Elsevier logo
Journals & Books
Adenosine Triphosphate
ATP is a nucleotide consisting of an adenine, a 5-carbon sugar (ribose), and three phosphate groups.
From: The Scientist's Guide to Cardiac Metabolism, 2016
Add to MendeleySet alert
About this page
Contents
Definition
Chapters and Articles
Related Terms
Recommended Publications
Chapters and Articles
Adenosine Triphosphate
Charles Kennedy, in xPharm: The Comprehensive Pharmacology Reference, 2007
Adenosine Triphosphate; ATP; adenosine sodiumtriphosphate; adenosine 5 triphosphate; adenosine 5′ triphosphate; adenosinetriphosphate; adenosine 5 triphosphate disodium; adenosinetriphosphate disodium; adenosine triphosphate disodium salt; adenosinetriphosphate potassium; adenosine 5′ triphosphate sodium; adenosinetriphosphate sodium; adenosine triphosphate sodium potassium; adenosinetriphosphate sodium salt; adenosine triphosphate tetrasodium; adenosinetriphosphoric acid; adenosinetriphosphoric acid; adenosine triphosphoric acidsodium salt; adenylpyrophosphoric acid; adephos; adetol; arteriotonin; atepodin; atipi; 5′ atp; a.t.p.; ATP; atp disodium; atriphos; disodiumadenosine 5 triphosphate; disodium adenosine triphosphate; epsilon atp; fosfobion; glucobasin; levo atp; phosphobion; potassium atp; sodium atp; sodium potassium adenosine triphosphate; striadyne; tetrasodium atp; thriphosaden; triadenyl; triphosphoadenosine; ATP (trade); 5′ atp; adenosine 5′ triphosphate; adenosine 5′ triphosphate sodium; adenosine sodium triphosphate; adenosine triphosphate disodium; adenosine triphosphate potassium; adenosine triphosphate sodium; adenosine triphosphate sodium salt; adenosine triphosphoric acid; adenosine triphosphoric acid sodium salt; disodium adenosine 5 triphosphate; stryadine
Cellular Respiration
Jerry J. Zimmerman, ... Jerry McLaughlin, in Pediatric Critical Care (Fourth Edition), 2011
Adenosine Triphosphate
ATP is considered the molecular unit of intracellular energy currency. ATP derives its inherent energy secondary to anhydride bonds connecting adjacent phosphate functional groups. Hydrolysis of ATP energy generates energy for all cellular processes. In addition ATP also serves as a cofactor for signal transduction reactions using a variety of kinases as well as adenyl cyclase. Normally cellular ATP concentration is maintained in the range of 1 to 10 mmol/L, with a normal ratio of ATP/ADP of approximately 1000. Totally quantity of ATP in an adult is approximately 0.10 mol/L. Approximately 100 to 150 mol/L of ATP are required daily, which means that each ATP molecule is recycled some 1000 to 1500 times per day. Basically, the human body turns over its weight in ATP daily.39 Transmembrane proton flux through the mitochondrial ATPase synthase complex occurs at an estimated rate of 3 × 1021 protons per second. This corresponds to ATP reformed at a rate of 9 × 1020 molecules/sec, or approximately 65 kg ATP recycled per day in a normal resting adult (Figure 74-5).24
Adenosine Triphosphate (ATP) as a Neurotransmitter
A. Verkhratsky, O. Krishtal, in Encyclopedia of Neuroscience, 2009
Adenosine triphosphate (ATP) is an important extracellular signaling molecule. ATP acts as a neurotransmitter in both peripheral and central nervous systems. In the peripheral nervous system, ATP is involved in chemical transmission in sensory and autonomic ganglia. In the central nervous system, ATP, released from synaptic terminals, induces fast excitatory postsynaptic currents. Postsynaptic action of ATP is mediated by a plethora of ionotropic and metabotropic receptors. Furthermore, ATP also acts as an important mediator in neuronal–glial and glial–glial signaling. All types of glial cells are endowed with diverse ATP receptors, which trigger Ca2+ signaling events and membrane currents. ATP can also act as a ‘glio’transmitter released from astroglial cells via regulated exocytosis or through plasmalemmal channels.
RNA Polymerases
Hyone-Myong Eun, in Enzymology Primer for Recombinant DNA Technology, 1996
i. Nucleotides.
ATP is by far the preferred nucleotide substrate. Both CTP and UTP are polymerized at less than 5% of the rate obtained with ATP (6). The enzyme does not show any activity with GTP, ADP, and dATP. Cordycepin triphosphate (3′-deoxy-ATP) serves as a chain-terminating substrate.
Eukaryotic poly (A) polymerases can utilize NTPs other than ATP, either singly or in the presence of ATP, at less than 1% of the rate for ATP (3). The same is true for rADP and dATP.
Purinergic neurotransmission and nucleotide receptors
Samuel J. Fountain, in Primer on the Autonomic Nervous System (Fourth Edition), 2023
Abstract
Adenosine 5′-triphosphate (ATP) is a neurotransmitter of postganglionic sympathetic nerves and nonadrenergic, noncholinergic nerves of the enteric nervous system. ATP can also modulate autonomic reflexes through actions at sensory afferent nerves. Mechanisms exist within nerves for the vesicular storage and exocytotic release of ATP. Once released, the availability and duration of ATP is regulated by cell surface ectonucleotidases that hydrolyze ATP, producing metabolites including ADP and adenosine in the process. The effects of ATP are mediated via activation of cell surface purinergic receptors, namely, P2X and P2Y receptors. P2X receptors are ligand-gated nonselective cation channels activated by ATP and causing membrane depolarization and elevation in intracellular Ca2+. P2Y receptors are G protein-coupled receptors activated by ATP and ADP. Signal transduction mechanisms of P2Y receptors including intracellular Ca2+ signaling and inhibition of adenylate cyclase activity. Postjunctional P2X and P2Y receptors effect neurotransmission by ATP, whereas prejunctional receptors are capable of modulating neurotransmitter release.
Cell Metabolism
Merri Lynn Casem BA, PhD, in Case Studies in Cell Biology, 2016
Measurement of ATP levels
ATP concentrations were measured using a bioluminescence assay based on the ability of luciferase to produce light in the presence of its substrate luciferin and ATP. There is a linear relationship between the amount of ATP present in the sample and the amount of light produced. Luminescence of a sample is compared with a standard curve of known ATP concentrations. At each time point, 12.5 μL of the cell sample was added to an equal volume of 10% trichloroacetic acid and vortexed vigorously for 1 min to extract ATP. The mixtures were then neutralized with 1 mL of neutralization buffer, and 10 μL of sample was reacted with 100 μL of the luciferin/luciferase mixture. ATP concentrations were normalized and expressed as the ratio of ATP levels in prestarved cells.
Mitochondria, 3rd Edition
Giampaolo Morciano, ... Paolo Pinton, in Methods in Cell Biology, 2020
Abstract
Adenosine 5′-triphosphate (ATP) is the central metabolite in the energy metabolism of cells and is hydrolyzed to ADP and inorganic phosphate to provide free energy in various cellular processes. ATP also functions as an intracellular signaling molecule. Thus, it is important to know the ATP concentration within cells to understand cellular activities. Here, we describe two methods to detect ATP concentrations in the cytoplasm and mitochondrial matrix using genetically encoded luminescent or fluorescent biosensors. These methods enable quantitative investigation of ATP concentration dynamics in living cells, single cells and cell populations.
Orientia
Daniel H. Paris, ... Nicholas P.J. Day, in Molecular Medical Microbiology (Second Edition), 2015
Energy Metabolism
ATP is the universal energy source in all biological systems, and can be synthesized by glycolysis and oxidative phosphorylation. The majority of the genes encoding the complexes of the oxidative phosphorylation pathways are present in Orientia; including three proton pumps, the succinate dehydrogenase complex, and the ATP synthase complex [158]. Orientia have five copies of the ATP/ADP translocases, thus they may first exploit host cell ATP available in the cytoplasm via these translocases and subsequently produce ATP self-sufficiently via aerobic respiration when the host ATP pool is depleted [158].
Ligases
Hyone-Myong Eun, in Enzymology Primer for Recombinant DNA Technology, 1996
(a) ATP.
ATP is the specific cofactor for the T4 RNA ligase. The Km for ATP is 0.2 μM in circularization reactions (1) or 12 μM in ATP-PP; exchange reactions (6). Of the common nucleoside triphosphates, only dATP can substitute for ATP but with only 10–20% efficacy.
Increasing the ATP concentrations up to 100 μM leads to an increase of ligation efficiency up to 50%. A higher concentration of ATP inhibits the reaction, resulting in excessive accumulation of the AMP-RNA (donor) intermediate. Eventually, however, this condition leads to a higher product yield after a long incubation time (7). At high enzyme and ATP concentrations, adenylylation at the 3′-P of both donor and product occurs as a side reaction. For practical purposes, an ATP concentration of 20 μM provides the best compromise between ligation and adenylylation (5).
Presynaptic: Mitochondrial and Presynaptic Function☆
Umut Cagin, in Reference Module in Neuroscience and Biobehavioral Psychology, 2017
Signaling Roles of ATP
ATP is present in synaptic vesicles (∼0.2 M) and is co-released with ACh. In cultured neuromuscular junctions, the retrograde effects of this co-released ATP potentiate spontaneous vesicular ACh release, and might play a role in synaptic development. It has been suggested that endogenously released ATP modulates intracellular [Ca2+] in perisynaptic Schwann cells of amphibian neuromuscular junctions. It has also been suggested that presynaptic actions of adenosine, derived from hydrolysis of released ATP, contribute to the depression of transmitter release in frog neuromuscular junctions. Although exogenously applied ATP depresses transmitter release in mammalian neuromuscular junctions, the endogenous release of ATP (or the accumulation of adenosine) might not be sufficient to alter evoked transmitter release under physiological conditions.
Related terms:
Eicosanoid Receptor
Adenosine Diphosphate
Reactive Oxygen Species
Mitochondrion
Oxidative Phosphorylation
In Vitro
Glutamic Acid
Adenosine
Astrocyte
Cell Membrane
View all Topics
Recommended publications
New Scientist
New Scientist
Journal
Enzymes (Second Edition)
Enzymes (Second Edition)
Book • 2007
Elsevier logo with wordmark
About ScienceDirect
Remote access
Shopping cart
Advertise
Contact and support
Terms and conditions
Privacy policy
Cookies are used by this site. Cookie settings | Your Privacy Choices
All content on this site: Copyright © 2024 Elsevier B.V., its licensors, and contributors. All rights are reserved, including those for text and data mining, AI training, and similar technologies. For all open access content, the Creative Commons licensing terms apply.
RELX group home page
Feedback | biology | 212255 | https://no.wikipedia.org/wiki/Adenosintrifosfat | Adenosintrifosfat | Adenosin-5'-trifosfat (ATP, etter det engelske navnet Adenosine triphosphate) er et multifunksjonelt nukleotid som brukes i celler som et koenzym. ATP blir ofte kalt «molekylær valutaenhet» for intracellulær energioverføring. ATP frakter kjemisk energi i cellene for metabolismen. ATP dannes under fotofosforylering og cellulær respirasjon, og brukes av enzymer og strukturelle proteiner i mange cellulære prosesser, herunder biosyntetiske reaksjoner, motilitet og celledeling. Ett ATP-molekyl inneholder tre fosfatgrupper, og det blir produsert fra uorganisk fosfat og adenosindifosfat (ADP) eller adenosinmonofosfat (AMP) ved hjelp av enzymet ATP-syntase.
Metabolske prosesser som bruker ATP som en energikilde omdanner det tilbake til dets forløpere. ATP er derfor i kontinuerlig resirkulasjon i levende organismer. Menneskekroppen inneholder i gjennomsnitt 250 gram ATP.
ATP blir brukt som et substrat i signaltransduksjonsveiene av kinaser som fosforylerer proteiner og lipider, samt av adenylat syklase, som bruker ATP til å produsere det sekundære budbringermolekylet syklisk AMP. Forholdet mellom ATP og AMP brukes av en celle til å oppfatte hvor mye energi som er tilgjengelig og kontrollere de metabolske veiene som produserer og forbruker ATP etter behov. I tillegg til sin rolle i energimetabolismen og signalisering, blir også ATP inkorporert i nukleinsyrer av polymeraser under DNA-replikasjon -og transkripsjon.
Strukturelt sett består dette molekylet av en purinbase (adenin) festet til et 1'-karbonatom i et pentosesukker (ribose). Tre fosfatgrupper er festet på 5'-karbonatomet i pentosesukkeret. Det er påhekting og avhekting av disse fosfatgruppene som hele tiden står for interkonvertering av ATP, ADP og AMP. Når ATP brukes i DNA-syntese, blir monosakkaridet ribose først omdannet til deoksyribose av ribonukleotid reduktase.
ATP ble oppdaget av Karl Lohmann i 1929, men dets korrekte struktur ble ikke bestemt før noen år senere. Det ble i 1941 foreslått av Fritz Albert Lipmann at ATP var det viktigste energioverføringsmolekylet i cellen. ATP ble kunstig syntetisert for første gang av Alexander Todd i 1948.
Referanser
Eksterne lenker
Oppføring på PubChem
Oppføring på KEGG
Adenosintrifosfat på HyperPhysics
ATP: The Perfect Energy Currency for the Cell
Stoffskiftet
Enzymer
Nukleinsyrer | norwegian_bokmål | 0.210941 |
induce_sleep/Adenosine_A1_receptor.txt |
The adenosine A1 receptor (A1AR) is one member of the adenosine receptor group of G protein-coupled receptors with adenosine as endogenous ligand.
Biochemistry[edit]
A1 receptors are implicated in sleep promotion by inhibiting wake-promoting cholinergic neurons in the basal forebrain. A1 receptors are also present in smooth muscle throughout the vascular system.
The adenosine A1 receptor has been found to be ubiquitous throughout the entire body.
Signaling[edit]
Activation of the adenosine A1 receptor by an agonist causes binding of Gi1/2/3 or Go protein. Binding of Gi1/2/3 causes an inhibition of adenylate cyclase and, therefore, a decrease in the cAMP concentration. An increase of the inositol triphosphate/diacylglycerol concentration is caused by an activation of phospholipase C, whereas the elevated levels of arachidonic acid are mediated by DAG lipase, which cleaves DAG to form arachidonic acid.
Several types of potassium channels are activated but N-, P-, and Q-type calcium channels are inhibited.
Effect[edit]
This receptor has an inhibitory function on most of the tissues in which it rests. In the brain, it slows metabolic activity by a combination of actions. At the neuron's synapse, it reduces synaptic vesicle release.
Ligands[edit]
Caffeine, as well as theophylline, has been found to antagonize both A1 and A2A receptors in the brain.
Agonists[edit]
2-Chloro-N(6)-cyclopentyladenosine (CCPA).
N6-Cyclopentyladenosine
N(6)-cyclohexyladenosine
Tecadenoson ((2R,3S,4R)-2-(hydroxymethyl)-5-(6-
((R)-tetrahydrofuran-3-ylamino)-9H-purin-9-yl)-tetrashydrofuran3,4-diol)
Selodenoson ((2S,3S,4R)-5-(6-(cyclopentylamino)-9Hpurin-9-yl)-N-ethyl-3,4-dihydroxytetrahydrofuran-2-carboxamide)
Capadenoson (BAY68-4986)
Benzyloxy-cyclopentyladenosine (BnOCPA) is an A1R selective agonist.
PAMs[edit]
2‑Amino-3-(4′-chlorobenzoyl)-4-substituted-5-arylethynyl thiophene # 4e
Antagonists[edit]
Non-selective
Caffeine
Theophylline
CGS-15943
Selective
8-Cyclopentyl-1,3-dimethylxanthine (CPX / 8-cyclopentyltheophylline)
8-Cyclopentyl-1,3-dipropylxanthine (DPCPX)
8-Phenyl-1,3-dipropylxanthine
Bamifylline
BG-9719
BG-9928
FK-453
FK-838
Rolofylline (KW-3902)
N-0861
ISAM-CV202
In the heart[edit]
In the heart, A1 receptors play roles in electrical pacing (chronotropy and dromotropy), fluid balance, local sympathetic regulation, and metabolism.
When bound by adenosine, A1 receptors inhibit impulses generated in supraventricular tissue (SA node, AV node) and the Bundle of His/Purkinje system, leading to negative chronotropy (slowing of the heart rate). Specifically, A1 receptor activation leads to inactivation of the inwardly rectifying K current and inhibition of the inward Ca current (ICa) and
the 'funny' hyperpolarization-activated current (If). Adenosine agonism of A1ARs also inhibits release of norepinephrine from cardiac nerves. Norepinephrine is a positive chronotrope, inotrope, and dromotrope, through its agonism of β adrenergic receptors on pacemaker cells and ventricular myocytes.
Collectively, these mechanisms lead to an myocardial depressant effect by decreasing the conduction of electrical impulses and suppressing pacemaker cells function, resulting in a decrease in heart rate. This makes adenosine a useful medication for treating and diagnosing tachyarrhythmias, or excessively fast heart rates. This effect on the A1 receptor also explains why there is a brief moment of cardiac standstill when adenosine is administered as a rapid IV push during cardiac resuscitation. The rapid infusion causes a momentary myocardial stunning effect.
In normal physiological states, this serves as protective mechanisms. However, in altered cardiac function, such as hypoperfusion caused by hypotension, heart attack or cardiac arrest caused by nonperfusing bradycardias, adenosine has a negative effect on physiological functioning by preventing necessary compensatory increases in heart rate and blood pressure that attempt to maintain cerebral perfusion.
Metabolically, A1AR activation by endogenous adenosine across the body reduces plasma glucose, lactate, and insulin levels, however A2aR activation increased glucose and lactate levels to an extent greater than the A1AR effect on glucose and lactate. Thus, intravascular administration of adenosine increases the amount of glucose and lactate available in the blood for cardiac myocytes. A1AR activation also partially inhibits glycolysis, slowing its rate to align with oxidative metabolism, which limits post-ischemic damage through reduced H generation.
In the state of myocardial hypertrophy and remodeling, interstitial adenosine and the expression of the A1AR receptor are both increased. After transition to heart failure however, overexpression of A1AR is no longer present. Excess A1AR expression can induce cardiomyopathy, cardiac dilatation, and cardiac hypertrophy. Cardiac failure may involve increased A1AR expression and decreased adenosine in physical models of cardiac overload and in dysfunction induced by TNFα. Heart failure often involves secretion of atrial natriuretic peptide to compensate for reduced renal perfusion and thus, secretion of electrolytes. A1AR activation also increases secretion of atrial natriuretic peptide from atrial myocytes. | biology | 214153 | https://da.wikipedia.org/wiki/Allosterisk%20regulering | Allosterisk regulering | Indenfor biokemien betegner allosterisk regulering kontrol af enzymer eller proteiners aktivitet ved binding af stoffer eller proteiner til et andet område af proteinet end dets aktive site; dette andet område kaldes et allosterisk site, og allosteriske proteiner kan have et eller flere allosteriske sites.
Allosterisk regulation af enzymer er centralt indenfor regulering af cellers metabolisme, og allosteri har også en vigtig effekt på blods evne til at binde og frigive oxygen og carbondioxid effektivt.
I farmakologi er en allosterisk modulator (allo- fra den græske betydning "anden") et stof, som indirekte påvirker virkningerne af en agonist eller omvendt agonist ved et målprotein, for eksempel en receptor. Allosteriske modulatorer binder til et sted, der er forskelligt fra det for orthosteriske agonist-bindingssted (dvs aktive sæde (aktive site). Normalt inducerer de en konformationsændring inden for protein-strukturen.
En positiv allosterisk modulator (PAM) eller allosterisk forstærker inducerer en amplifikation af den orthosteriske agonists virkning enten ved at forøge bindings-affiniteten eller den funktionelle effekt af den orthosteriske agonist for målproteinet. En negativ modulator (NAM) reducerer virkningerne af den orthosteriske ligand, men er inaktiv i fravær af den orthosteriske ligand. Stoffer, der optager det allosteriske bindingssted og er funktionelt neutrale, kaldes tavse allosteriske modulatorer (SAMer). Klassiske benzodiazepiner er velkendte PAM'er.
Den modulerende aktivitet kan være førsteorden eller andenorden (eller begge). Andenordens modulatorer ændrer modulator-aktiviteten af førstordens modulatorer. Epigallocatechin-3-gallat, en plante sekundær metabolit og den mest rigelige catechin i te, er et sådant eksempel på en andenordens-modulator ved GABAA-receptorer.
Allosteriske agonister skal skelnes fra rene allosteriske modulatorer. De er defineret som ligandere, der er i stand til direkte at aktivere en receptor ved at binde til et receptorsted, der er forskelligt fra det primære (orthosteriske) sted, det allosteriske sted. De er i stand til at binde målproteinet og udøve deres virkning i fravær af en orthosterisk ligand, medens tilstedeværelsen af en orthosterisk ligand typisk kræves for at observere de indirekte virkninger af rene allosteriske modulatorer.
Ago-allosteriske modulatorer er både allosteriske agonister og allosteriske modulatorer. En ago-allosterisk modulator virker som en agonist og en forstærker for endogene agonister i stigende agoniststyrke (det dosisområde, over hvilket et respons produceres) og tilvejebringer "superagonisme". Superagonisme resultater, når effekten er større end 100 procent. Ago-allosteriske modulatorer kan være neutrale, negative eller positive. Neutrale ago-allosteriske modulatorer øger effektiviteten, men har ingen effekt på styrke. En negativ-sidet allosterisk modulator har en negativ effekt på styrken, men en positiv effekt på effekten af en agonist. En positiv sidet-allosterisk modulator øger både effektivitet og styrke.
Det Endocannabinoide System
I 2005 blev de første beviser for et allosterisk bindingssted ved G-protein-koblede cannabinoid CB1-receptorer tilvejebragt ved identifikation af tre indoler af firmaet Organon. Allosteriske modulatorer af cannabinoid CB1-receptorer har potentialet til at behandle CNS og perifere forstyrrelser, samtidig med undgåelse af bivirkninger forbundet med orthosterisk agonisme eller antagonisme af disse receptorer.
Pepcan-12 (RVD-Hpα) er hoved-peptiden af en familie af endogene peptid-endocannabinoider (pepcans), der påvises at virke som negative allosteriske modulatorer (NAM) af cannabinoid CB1-receptorer. Det er vist, at pepcan-12 modsat virker som en potent CB2 cannabinoid-receptor positiv allosterisk modulator (PAMer). RVD-Hpα forstærkede også signifikant virkningerne af CB2-receptor-agonister, herunder endocannabinoiden 2-arakidonoylglycerol (2-AG: C23H38O4), til GTPγS-binding og cAMP-hæmning (5-10 gange). Den formodede forløber pepcan-23 blev identificeret med pepcan-12 i hjerne, lever og nyre hos mus.
Pepcan-12 blev forøget ved endotoxæmi (tilstedeværelsen af endotoxiner (lipopolysaccharid eller LPS) i blodet) og iskæmi-reperfusions-skade (vævsskade forårsaget, når blodtilførslen vender tilbage til væv efter en periode med iskæmi eller mangel på ilt (anoxi eller hypoxi), hvor CB2-receptorer spiller en beskyttende rolle. Den brede forekomst af denne endogene hormon-lignende CB2-receptor PAM, med uforudsete modsatte allosteriske virkninger på cannabinoid-receptorer, antyder dens potentielle rolle i perifere pato-fysiologiske processer.
Plante-cannabinoiden cannabidiol (CBD) er fundet at opføre sig som en ikke-konkurrencedygtig negativ allosterisk modulator (NAM) af CB1-receptorer (enzymhæmning, hvor CBD reducerer enzymets aktivitet og binder lige så godt til enzymet, uanset om det allerede har bundet substratet). Den allosteriske aktivitet af cannabidiol afhænger af, at polære rester er til stede i positionerne 98 og 107 i CB1-receptorens ekstracellulære (ydre del) amino-terminal (N-terminus) og derved viste reduceret effektivitet og styrke af 2-arakidonylglycerol og Δ9-tetrahydrocannabinol (THC) ved denne receptor. Allosterisk modulering i forbindelse med virkninger der ikke medieres af CB1-receptorer, kan forklare in vivo effekterne af cannabidiol.
Nye familier strukturelt baseret på urinstof og på 3-fenyltropan-analoger af kokain, udviklet til at reducere kokain-afhængighed og afhængighed, er også blevet opdaget som CB1-receptor-negative allosteriske modulatorer (NAMer). Andre endogene allosteriske ligandere af forskellig art for CB1 er blevet yderligere identificeret. Således er den terapeutiske neuro-beskyttende påføring af lipoxin A4, en arakidonsyre metabolisk afledning, som en allosterisk forstærker (PAM) af CB1-receptor-aktivitet blevet bekræftet in vivo.
Det er også tilfældet med steroid-hormonet pregnenolon, hvis negative allosteriske virkninger (NAM) på Δ9- tetrahydrocannabinol (Δ9 -THC) blev reproduceret in vivo i en adfærdsmæssig tetrad-model og i prøver med fødeindtag og dæmpning af hukommelse. Også den peroxisom-proliferator-aktiveret receptor gamma (PPAR-y eller PPARG) agonist fenofibrat og polypeptider har vist sig, at virke ved det endocannabinoide system via CB1-allosterisk modulering. Derfor kan allosterisk modulering udbrede nye tilgange til finjustering af signalvejene i CB1- og CB2-receptorerne.
Dertil er fundet, at cannabinoid CB1-receptoren er involveret i de anxiolytiske (antipanik; antiangst), beroligende og amnesiske (hukommelsestab) virkninger af benzodiazepiner (PAM'er) og at denne receptor, spiller en afgørende rolle i de farmakologiske virkninger af benzodiazepiner. Desuden, at blokade eller dæmpning af cannabinoid CB1-receptorer derfor kan være nyttig til behandling af patienter med problemer i forbindelse med forbruget af benzodiazepiner.
Anden forskning fremført i 2012 påviser desuden også, at både eksogene og endogene cannabinoider kan modulere glycin (C2H5NO2)-receptorer (GlyRs) allosterisk. Men lidt er kendt om det molekylære grundlag for cannabinoid-GlyR interaktioner, hvorfor der vises, at vedvarende inkubation med endocannabinoiden anandamide (AEA: C22H37NO2) øger amplituden væsentligt af den glycin (α-aminosyre)-aktiverede strøm i både rotte dyrkede neuroner i rygmarven og i HEK-293-celler, der udtrykker human α1, rotte α2 og α3 GlyRs.
Størrelsen af AEA potensering faldt med fjernelse af enten hydroxyl-eller oxygen-grupper på AEA. Mens desoxy-AEA var signifikant mindre effektiv i potensering af I (Gly), hæmmede desoxy-AEA potensering fremstillet af både Δ(9)-tetrahydrocannabinol (THC: C21H30O2), en betydelig psykoaktiv bestanddel af marihuana, og AEA. Tilsvarende påvirkede didesoxy-THC, en modificeret THC med fjernelse af både hydroxyl/oxygengrupper, ikke I (Gly), når de anvendes alene, men inhiberede potensering af I (Gly) induceret af AEA og THC. Disse resultater tyder på, at eksogene og endogene cannabinoider potenserer GlyRs via hydrogenbindings-lignende interaktion. En sådan specifik interaktion stammer sandsynligvis fra et fælles molekylær grundlag.
Se også
Enzymkinetik
Henvisninger
En fælles molekylær basis for eksogen og endogen cannabinoid potensering af glycin-receptorer - J Neurosci. 2012 April.
Noter
Biokemi | danish | 0.428539 |
temperature_sensed/Temperature.txt | Temperature is a physical quantity that quantitatively expresses the attribute of hotness or coldness. Temperature is measured with a thermometer. It reflects the average kinetic energy of the vibrating and colliding atoms making up a substance.
Thermometers are calibrated in various temperature scales that historically have relied on various reference points and thermometric substances for definition. The most common scales are the Celsius scale with the unit symbol °C (formerly called centigrade), the Fahrenheit scale (°F), and the Kelvin scale (K), the latter being used predominantly for scientific purposes. The kelvin is one of the seven base units in the International System of Units (SI).
Absolute zero, i.e., zero kelvin or −273.15 °C, is the lowest point in the thermodynamic temperature scale. Experimentally, it can be approached very closely but not actually reached, as recognized in the third law of thermodynamics. It would be impossible to extract energy as heat from a body at that temperature.
Temperature is important in all fields of natural science, including physics, chemistry, Earth science, astronomy, medicine, biology, ecology, material science, metallurgy, mechanical engineering and geography as well as most aspects of daily life.
Effects[edit]
Average daily variation in human body temperature
Many physical processes are related to temperature; some of them are given below:
the physical properties of materials including the phase (solid, liquid, gaseous or plasma), density, solubility, vapor pressure, electrical conductivity, hardness, wear resistance, thermal conductivity, corrosion resistance, strength
the rate and extent to which chemical reactions occur
the amount and properties of thermal radiation emitted from the surface of an object
air temperature affects all living organisms
the speed of sound, which in a gas is proportional to the square root of the absolute temperature
Scales[edit]
Main article: Scale of temperature
Two thermometers showing temperature in Celsius and Fahrenheit
This section needs additional citations for verification. Please help improve this article by adding citations to reliable sources in this section. Unsourced material may be challenged and removed. (January 2021) (Learn how and when to remove this template message)
Temperature scales need two values for definition: the point chosen as zero degrees and the magnitudes of the incremental unit of temperature.
The Celsius scale (°C) is used for common temperature measurements in most of the world. It is an empirical scale that developed historically, which led to its zero point 0 °C being defined as the freezing point of water, and 100 °C as the boiling point of water, both at atmospheric pressure at sea level. It was called a centigrade scale because of the 100-degree interval. Since the standardization of the kelvin in the International System of Units, it has subsequently been redefined in terms of the equivalent fixing points on the Kelvin scale, so that a temperature increment of one degree Celsius is the same as an increment of one kelvin, though numerically the scales differ by an exact offset of 273.15.
The Fahrenheit scale is in common use in the United States. Water freezes at 32 °F and boils at 212 °F at sea-level atmospheric pressure.
Absolute zero[edit]
At the absolute zero of temperature, no energy can be removed from matter as heat, a fact expressed in the third law of thermodynamics. At this temperature, matter contains no macroscopic thermal energy, but still has quantum-mechanical zero-point energy as predicted by the uncertainty principle, although this does not enter into the definition of absolute temperature. Experimentally, absolute zero can be approached only very closely; it can never be reached (the lowest temperature attained by experiment is 38 pK). Theoretically, in a body at a temperature of absolute zero, all classical motion of its particles has ceased and they are at complete rest in this classical sense. The absolute zero, defined as 0 K, is exactly equal to −273.15 °C, or −459.67 °F.
Absolute scales[edit]
Referring to the Boltzmann constant, to the Maxwell–Boltzmann distribution, and to the Boltzmann statistical mechanical definition of entropy, as distinct from the Gibbs definition, for independently moving microscopic particles, disregarding interparticle potential energy, by international agreement, a temperature scale is defined and said to be absolute because it is independent of the characteristics of particular thermometric substances and thermometer mechanisms. Apart from the absolute zero, it does not have a reference temperature. It is known as the Kelvin scale, widely used in science and technology. The kelvin (the unit name is spelled with a lower-case 'k') is the unit of temperature in the International System of Units (SI). The temperature of a body in a state of thermodynamic equilibrium is always positive relative to the absolute zero.
Besides the internationally agreed Kelvin scale, there is also a thermodynamic temperature scale, invented by Lord Kelvin, also with its numerical zero at the absolute zero of temperature, but directly relating to purely macroscopic thermodynamic concepts, including the macroscopic entropy, though microscopically referable to the Gibbs statistical mechanical definition of entropy for the canonical ensemble, that takes interparticle potential energy into account, as well as independent particle motion so that it can account for measurements of temperatures near absolute zero. This scale has a reference temperature at the triple point of water, the numerical value of which is defined by measurements using the aforementioned internationally agreed Kelvin scale.
Kelvin scale[edit]
Many scientific measurements use the Kelvin temperature scale (unit symbol: K), named in honor of the physicist who first defined it. It is an absolute scale. Its numerical zero point, 0 K, is at the absolute zero of temperature. Since May, 2019, the kelvin has been defined through particle kinetic theory, and statistical mechanics. In the International System of Units (SI), the magnitude of the kelvin is defined in terms of the Boltzmann constant, the value of which is defined as fixed by international convention.
Statistical mechanical versus thermodynamic temperature scales[edit]
Since May 2019, the magnitude of the kelvin is defined in relation to microscopic phenomena, characterized in terms of statistical mechanics. Previously, but since 1954, the International System of Units defined a scale and unit for the kelvin as a thermodynamic temperature, by using the reliably reproducible temperature of the triple point of water as a second reference point, the first reference point being 0 K at absolute zero.
Historically, the temperature of the triple point of water was defined as exactly 273.16 K. Today it is an empirically measured quantity. The freezing point of water at sea-level atmospheric pressure occurs at very close to 273.15 K (0 °C).
Classification of scales[edit]
There are various kinds of temperature scale. It may be convenient to classify them as empirically and theoretically based. Empirical temperature scales are historically older, while theoretically based scales arose in the middle of the nineteenth century.
Empirical scales[edit]
Empirically based temperature scales rely directly on measurements of simple macroscopic physical properties of materials. For example, the length of a column of mercury, confined in a glass-walled capillary tube, is dependent largely on temperature and is the basis of the very useful mercury-in-glass thermometer. Such scales are valid only within convenient ranges of temperature. For example, above the boiling point of mercury, a mercury-in-glass thermometer is impracticable. Most materials expand with temperature increase, but some materials, such as water, contract with temperature increase over some specific range, and then they are hardly useful as thermometric materials. A material is of no use as a thermometer near one of its phase-change temperatures, for example, its boiling-point.
In spite of these limitations, most generally used practical thermometers are of the empirically based kind. Especially, it was used for calorimetry, which contributed greatly to the discovery of thermodynamics. Nevertheless, empirical thermometry has serious drawbacks when judged as a basis for theoretical physics. Empirically based thermometers, beyond their base as simple direct measurements of ordinary physical properties of thermometric materials, can be re-calibrated, by use of theoretical physical reasoning, and this can extend their range of adequacy.
Theoretical scales[edit]
Theoretically based temperature scales are based directly on theoretical arguments, especially those of kinetic theory and thermodynamics. They are more or less ideally realized in practically feasible physical devices and materials. Theoretically based temperature scales are used to provide calibrating standards for practical empirically based thermometers.
Microscopic statistical mechanical scale[edit]
In physics, the internationally agreed conventional temperature scale is called the Kelvin scale. It is calibrated through the internationally agreed and prescribed value of the Boltzmann constant, referring to motions of microscopic particles, such as atoms, molecules, and electrons, constituent in the body whose temperature is to be measured. In contrast with the thermodynamic temperature scale invented by Kelvin, the presently conventional Kelvin temperature is not defined through comparison with the temperature of a reference state of a standard body, nor in terms of macroscopic thermodynamics.
Apart from the absolute zero of temperature, the Kelvin temperature of a body in a state of internal thermodynamic equilibrium is defined by measurements of suitably chosen of its physical properties, such as have precisely known theoretical explanations in terms of the Boltzmann constant. That constant refers to chosen kinds of motion of microscopic particles in the constitution of the body. In those kinds of motion, the particles move individually, without mutual interaction. Such motions are typically interrupted by inter-particle collisions, but for temperature measurement, the motions are chosen so that, between collisions, the non-interactive segments of their trajectories are known to be accessible to accurate measurement. For this purpose, interparticle potential energy is disregarded.
In an ideal gas, and in other theoretically understood bodies, the Kelvin temperature is defined to be proportional to the average kinetic energy of non-interactively moving microscopic particles, which can be measured by suitable techniques. The proportionality constant is a simple multiple of the Boltzmann constant. If molecules, atoms, or electrons are emitted from material and their velocities are measured, the spectrum of their velocities often nearly obeys a theoretical law called the Maxwell–Boltzmann distribution, which gives a well-founded measurement of temperatures for which the law holds. There have not yet been successful experiments of this same kind that directly use the Fermi–Dirac distribution for thermometry, but perhaps that will be achieved in the future.
The speed of sound in a gas can be calculated theoretically from the gas's molecular character, temperature, pressure, and the Boltzmann constant. For a gas of known molecular character and pressure, this provides a relation between temperature and the Boltzmann constant. Those quantities can be known or measured more precisely than can the thermodynamic variables that define the state of a sample of water at its triple point. Consequently, taking the value of the Boltzmann constant as a primarily defined reference of exactly defined value, a measurement of the speed of sound can provide a more precise measurement of the temperature of the gas.
Measurement of the spectrum of electromagnetic radiation from an ideal three-dimensional black body can provide an accurate temperature measurement because the frequency of maximum spectral radiance of black-body radiation is directly proportional to the temperature of the black body; this is known as Wien's displacement law and has a theoretical explanation in Planck's law and the Bose–Einstein law.
Measurement of the spectrum of noise-power produced by an electrical resistor can also provide accurate temperature measurement. The resistor has two terminals and is in effect a one-dimensional body. The Bose-Einstein law for this case indicates that the noise-power is directly proportional to the temperature of the resistor and to the value of its resistance and to the noise bandwidth. In a given frequency band, the noise-power has equal contributions from every frequency and is called Johnson noise. If the value of the resistance is known then the temperature can be found.
Macroscopic thermodynamic scale[edit]
Historically, till May 2019, the definition of the Kelvin scale was that invented by Kelvin, based on a ratio of quantities of energy in processes in an ideal Carnot engine, entirely in terms of macroscopic thermodynamics. That Carnot engine was to work between two temperatures, that of the body whose temperature was to be measured, and a reference, that of a body at the temperature of the triple point of water. Then the reference temperature, that of the triple point, was defined to be exactly 273.16 K. Since May 2019, that value has not been fixed by definition but is to be measured through microscopic phenomena, involving the Boltzmann constant, as described above. The microscopic statistical mechanical definition does not have a reference temperature.
Ideal gas[edit]
A material on which a macroscopically defined temperature scale may be based is the ideal gas. The pressure exerted by a fixed volume and mass of an ideal gas is directly proportional to its temperature. Some natural gases show so nearly ideal properties over suitable temperature range that they can be used for thermometry; this was important during the development of thermodynamics and is still of practical importance today. The ideal gas thermometer is, however, not theoretically perfect for thermodynamics. This is because the entropy of an ideal gas at its absolute zero of temperature is not a positive semi-definite quantity, which puts the gas in violation of the third law of thermodynamics. In contrast to real materials, the ideal gas does not liquefy or solidify, no matter how cold it is. Alternatively thinking, the ideal gas law, refers to the limit of infinitely high temperature and zero pressure; these conditions guarantee non-interactive motions of the constituent molecules.
Kinetic theory approach[edit]
The magnitude of the kelvin is now defined in terms of kinetic theory, derived from the value of the Boltzmann constant.
Kinetic theory provides a microscopic account of temperature for some bodies of material, especially gases, based on macroscopic systems' being composed of many microscopic particles, such as molecules and ions of various species, the particles of a species being all alike. It explains macroscopic phenomena through the classical mechanics of the microscopic particles. The equipartition theorem of kinetic theory asserts that each classical degree of freedom of a freely moving particle has an average kinetic energy of kBT/2 where kB denotes the Boltzmann constant. The translational motion of the particle has three degrees of freedom, so that, except at very low temperatures where quantum effects predominate, the average translational kinetic energy of a freely moving particle in a system with temperature T will be 3kBT/2.
Molecules, such as oxygen (O2), have more degrees of freedom than single spherical atoms: they undergo rotational and vibrational motions as well as translations. Heating results in an increase of temperature due to an increase in the average translational kinetic energy of the molecules. Heating will also cause, through equipartitioning, the energy associated with vibrational and rotational modes to increase. Thus a diatomic gas will require more energy input to increase its temperature by a certain amount, i.e. it will have a greater heat capacity than a monatomic gas.
As noted above, the speed of sound in a gas can be calculated from the gas's molecular character, temperature, pressure, and the Boltzmann constant. Taking the value of the Boltzmann constant as a primarily defined reference of exactly defined value, a measurement of the speed of sound can provide a more precise measurement of the temperature of the gas.
It is possible to measure the average kinetic energy of constituent microscopic particles if they are allowed to escape from the bulk of the system, through a small hole in the containing wall. The spectrum of velocities has to be measured, and the average calculated from that. It is not necessarily the case that the particles that escape and are measured have the same velocity distribution as the particles that remain in the bulk of the system, but sometimes a good sample is possible.
Thermodynamic approach[edit]
Temperature is one of the principal quantities in the study of thermodynamics. Formerly, the magnitude of the kelvin was defined in thermodynamic terms, but nowadays, as mentioned above, it is defined in terms of kinetic theory.
The thermodynamic temperature is said to be absolute for two reasons. One is that its formal character is independent of the properties of particular materials. The other reason is that its zero is, in a sense, absolute, in that it indicates absence of microscopic classical motion of the constituent particles of matter, so that they have a limiting specific heat of zero for zero temperature, according to the third law of thermodynamics. Nevertheless, a thermodynamic temperature does in fact have a definite numerical value that has been arbitrarily chosen by tradition and is dependent on the property of particular materials; it is simply less arbitrary than relative "degrees" scales such as Celsius and Fahrenheit. Being an absolute scale with one fixed point (zero), there is only one degree of freedom left to arbitrary choice, rather than two as in relative scales. For the Kelvin scale since May 2019, by international convention, the choice has been made to use knowledge of modes of operation of various thermometric devices, relying on microscopic kinetic theories about molecular motion. The numerical scale is settled by a conventional definition of the value of the Boltzmann constant, which relates macroscopic temperature to average microscopic kinetic energy of particles such as molecules. Its numerical value is arbitrary, and an alternate, less widely used absolute temperature scale exists called the Rankine scale, made to be aligned with the Fahrenheit scale as Kelvin is with Celsius.
The thermodynamic definition of temperature is due to Kelvin. It is framed in terms of an idealized device called a Carnot engine, imagined to run in a fictive continuous cycle of successive processes that traverse a cycle of states of its working body. The engine takes in a quantity of heat Q1 from a hot reservoir and passes out a lesser quantity of waste heat Q2 < 0 to a cold reservoir. The net heat energy absorbed by the working body is passed, as thermodynamic work, to a work reservoir, and is considered to be the output of the engine. The cycle is imagined to run so slowly that at each point of the cycle the working body is in a state of thermodynamic equilibrium. The successive processes of the cycle are thus imagined to run reversibly with no entropy production. Then the quantity of entropy taken in from the hot reservoir when the working body is heated is equal to that passed to the cold reservoir when the working body is cooled. Then the absolute or thermodynamic temperatures, T1 and T2, of the reservoirs are defined such that
T
1
T
2
=
−
Q
1
Q
2
.
{\displaystyle {\frac {T_{1}}{T_{2}}}=-{\frac {Q_{1}}{Q_{2}}}.}
(1)
The zeroth law of thermodynamics allows this definition to be used to measure the absolute or thermodynamic temperature of an arbitrary body of interest, by making the other heat reservoir have the same temperature as the body of interest.
Kelvin's original work postulating absolute temperature was published in 1848. It was based on the work of Carnot, before the formulation of the first law of thermodynamics. Carnot had no sound understanding of heat and no specific concept of entropy. He wrote of 'caloric' and said that all the caloric that passed from the hot reservoir was passed into the cold reservoir. Kelvin wrote in his 1848 paper that his scale was absolute in the sense that it was defined "independently of the properties of any particular kind of matter". His definitive publication, which sets out the definition just stated, was printed in 1853, a paper read in 1851.
Numerical details were formerly settled by making one of the heat reservoirs a cell at the triple point of water, which was defined to have an absolute temperature of 273.16 K. Nowadays, the numerical value is instead obtained from measurement through the microscopic statistical mechanical international definition, as above.
Intensive variability[edit]
In thermodynamic terms, temperature is an intensive variable because it is equal to a differential coefficient of one extensive variable with respect to another, for a given body. It thus has the dimensions of a ratio of two extensive variables. In thermodynamics, two bodies are often considered as connected by contact with a common wall, which has some specific permeability properties. Such specific permeability can be referred to a specific intensive variable. An example is a diathermic wall that is permeable only to heat; the intensive variable for this case is temperature. When the two bodies have been connected through the specifically permeable wall for a very long time, and have settled to a permanent steady state, the relevant intensive variables are equal in the two bodies; for a diathermal wall, this statement is sometimes called the zeroth law of thermodynamics.
In particular, when the body is described by stating its internal energy U, an extensive variable, as a function of its entropy S, also an extensive variable, and other state variables V, N, with U = U (S, V, N), then the temperature is equal to the partial derivative of the internal energy with respect to the entropy:
T
=
(
∂
U
∂
S
)
V
,
N
.
{\displaystyle T=\left({\frac {\partial U}{\partial S}}\right)_{V,N}.}
(2)
Likewise, when the body is described by stating its entropy S as a function of its internal energy U, and other state variables V, N, with S = S (U, V, N), then the reciprocal of the temperature is equal to the partial derivative of the entropy with respect to the internal energy:
1
T
=
(
∂
S
∂
U
)
V
,
N
.
{\displaystyle {\frac {1}{T}}=\left({\frac {\partial S}{\partial U}}\right)_{V,N}.}
(3)
The above definition, equation (1), of the absolute temperature, is due to Kelvin. It refers to systems closed to the transfer of matter and has a special emphasis on directly experimental procedures. A presentation of thermodynamics by Gibbs starts at a more abstract level and deals with systems open to the transfer of matter; in this development of thermodynamics, the equations (2) and (3) above are actually alternative definitions of temperature.
Local thermodynamic equilibrium[edit]
Real-world bodies are often not in thermodynamic equilibrium and not homogeneous. For the study by methods of classical irreversible thermodynamics, a body is usually spatially and temporally divided conceptually into 'cells' of small size. If classical thermodynamic equilibrium conditions for matter are fulfilled to good approximation in such a 'cell', then it is homogeneous and a temperature exists for it. If this is so for every 'cell' of the body, then local thermodynamic equilibrium is said to prevail throughout the body.
It makes good sense, for example, to say of the extensive variable U, or of the extensive variable S, that it has a density per unit volume or a quantity per unit mass of the system, but it makes no sense to speak of the density of temperature per unit volume or quantity of temperature per unit mass of the system. On the other hand, it makes no sense to speak of the internal energy at a point, while when local thermodynamic equilibrium prevails, it makes good sense to speak of the temperature at a point. Consequently, the temperature can vary from point to point in a medium that is not in global thermodynamic equilibrium, but in which there is local thermodynamic equilibrium.
Thus, when local thermodynamic equilibrium prevails in a body, the temperature can be regarded as a spatially varying local property in that body, and this is because the temperature is an intensive variable.
Basic theory[edit]
Conjugate variablesof thermodynamics
Pressure Volume (Stress) (Strain) Temperature Entropy Chemical potential Particle number
Temperature is a measure of a quality of a state of a material. The quality may be regarded as a more abstract entity than any particular temperature scale that measures it, and is called hotness by some writers. The quality of hotness refers to the state of material only in a particular locality, and in general, apart from bodies held in a steady state of thermodynamic equilibrium, hotness varies from place to place. It is not necessarily the case that a material in a particular place is in a state that is steady and nearly homogeneous enough to allow it to have a well-defined hotness or temperature. Hotness may be represented abstractly as a one-dimensional manifold. Every valid temperature scale has its own one-to-one map into the hotness manifold.
When two systems in thermal contact are at the same temperature no heat transfers between them. When a temperature difference does exist heat flows spontaneously from the warmer system to the colder system until they are in thermal equilibrium. Such heat transfer occurs by conduction or by thermal radiation.
Experimental physicists, for example Galileo and Newton, found that there are indefinitely many empirical temperature scales. Nevertheless, the zeroth law of thermodynamics says that they all measure the same quality. This means that for a body in its own state of internal thermodynamic equilibrium, every correctly calibrated thermometer, of whatever kind, that measures the temperature of the body, records one and the same temperature. For a body that is not in its own state of internal thermodynamic equilibrium, different thermometers can record different temperatures, depending respectively on the mechanisms of operation of the thermometers.
Bodies in thermodynamic equilibrium[edit]
For experimental physics, hotness means that, when comparing any two given bodies in their respective separate thermodynamic equilibria, any two suitably given empirical thermometers with numerical scale readings will agree as to which is the hotter of the two given bodies, or that they have the same temperature. This does not require the two thermometers to have a linear relation between their numerical scale readings, but it does require that the relation between their numerical readings shall be strictly monotonic. A definite sense of greater hotness can be had, independently of calorimetry, of thermodynamics, and of properties of particular materials, from Wien's displacement law of thermal radiation: the temperature of a bath of thermal radiation is proportional, by a universal constant, to the frequency of the maximum of its frequency spectrum; this frequency is always positive, but can have values that tend to zero. Thermal radiation is initially defined for a cavity in thermodynamic equilibrium. These physical facts justify a mathematical statement that hotness exists on an ordered one-dimensional manifold. This is a fundamental character of temperature and thermometers for bodies in their own thermodynamic equilibrium.
Except for a system undergoing a first-order phase change such as the melting of ice, as a closed system receives heat, without a change in its volume and without a change in external force fields acting on it, its temperature rises. For a system undergoing such a phase change so slowly that departure from thermodynamic equilibrium can be neglected, its temperature remains constant as the system is supplied with latent heat. Conversely, a loss of heat from a closed system, without phase change, without change of volume, and without a change in external force fields acting on it, decreases its temperature.
Bodies in a steady state but not in thermodynamic equilibrium[edit]
While for bodies in their own thermodynamic equilibrium states, the notion of temperature requires that all empirical thermometers must agree as to which of two bodies is the hotter or that they are at the same temperature, this requirement is not safe for bodies that are in steady states though not in thermodynamic equilibrium. It can then well be that different empirical thermometers disagree about which is hotter, and if this is so, then at least one of the bodies does not have a well-defined absolute thermodynamic temperature. Nevertheless, any one given body and any one suitable empirical thermometer can still support notions of empirical, non-absolute, hotness, and temperature, for a suitable range of processes. This is a matter for study in non-equilibrium thermodynamics.
Bodies not in a steady state[edit]
When a body is not in a steady-state, then the notion of temperature becomes even less safe than for a body in a steady state not in thermodynamic equilibrium. This is also a matter for study in non-equilibrium thermodynamics.
Thermodynamic equilibrium axiomatics[edit]
For the axiomatic treatment of thermodynamic equilibrium, since the 1930s, it has become customary to refer to a zeroth law of thermodynamics. The customarily stated minimalist version of such a law postulates only that all bodies, which when thermally connected would be in thermal equilibrium, should be said to have the same temperature by definition, but by itself does not establish temperature as a quantity expressed as a real number on a scale. A more physically informative version of such a law views empirical temperature as a chart on a hotness manifold. While the zeroth law permits the definitions of many different empirical scales of temperature, the second law of thermodynamics selects the definition of a single preferred, absolute temperature, unique up to an arbitrary scale factor, whence called the thermodynamic temperature. If internal energy is considered as a function of the volume and entropy of a homogeneous system in thermodynamic equilibrium, thermodynamic absolute temperature appears as the partial derivative of internal energy with respect the entropy at constant volume. Its natural, intrinsic origin or null point is absolute zero at which the entropy of any system is at a minimum. Although this is the lowest absolute temperature described by the model, the third law of thermodynamics postulates that absolute zero cannot be attained by any physical system.
Heat capacity[edit]
See also: Heat capacity and Calorimetry
When an energy transfer to or from a body is only as heat, the state of the body changes. Depending on the surroundings and the walls separating them from the body, various changes are possible in the body. They include chemical reactions, increase of pressure, increase of temperature and phase change. For each kind of change under specified conditions, the heat capacity is the ratio of the quantity of heat transferred to the magnitude of the change.
For example, if the change is an increase in temperature at constant volume, with no phase change and no chemical change, then the temperature of the body rises and its pressure increases. The quantity of heat transferred, ΔQ, divided by the observed temperature change, ΔT, is the body's heat capacity at constant volume:
C
V
=
Δ
Q
Δ
T
.
{\displaystyle C_{V}={\frac {\Delta Q}{\Delta T}}.}
If heat capacity is measured for a well-defined amount of substance, the specific heat is the measure of the heat required to increase the temperature of such a unit quantity by one unit of temperature. For example, raising the temperature of water by one kelvin (equal to one degree Celsius) requires 4186 joules per kilogram (J/kg).
Measurement[edit]
A typical Celsius thermometer measures a winter day temperature of −17 °C
See also: Timeline of temperature and pressure measurement technology, International Temperature Scale of 1990, and Comparison of temperature scales
Temperature measurement using modern scientific thermometers and temperature scales goes back at least as far as the early 18th century, when Daniel Gabriel Fahrenheit adapted a thermometer (switching to mercury) and a scale both developed by Ole Christensen Rømer. Fahrenheit's scale is still in use in the United States for non-scientific applications.
Temperature is measured with thermometers that may be calibrated to a variety of temperature scales. In most of the world (except for Belize, Myanmar, Liberia and the United States), the Celsius scale is used for most temperature measuring purposes. Most scientists measure temperature using the Celsius scale and thermodynamic temperature using the Kelvin scale, which is the Celsius scale offset so that its null point is 0 K = −273.15 °C, or absolute zero. Many engineering fields in the US, notably high-tech and US federal specifications (civil and military), also use the Kelvin and Celsius scales. Other engineering fields in the US also rely upon the Rankine scale (a shifted Fahrenheit scale) when working in thermodynamic-related disciplines such as combustion.
Units[edit]
The basic unit of temperature in the International System of Units (SI) is the kelvin. It has the symbol K.
For everyday applications, it is often convenient to use the Celsius scale, in which 0 °C corresponds very closely to the freezing point of water and 100 °C is its boiling point at sea level. Because liquid droplets commonly exist in clouds at sub-zero temperatures, 0 °C is better defined as the melting point of ice. In this scale, a temperature difference of 1 degree Celsius is the same as a 1kelvin increment, but the scale is offset by the temperature at which ice melts (273.15 K).
By international agreement, until May 2019, the Kelvin and Celsius scales were defined by two fixing points: absolute zero and the triple point of Vienna Standard Mean Ocean Water, which is water specially prepared with a specified blend of hydrogen and oxygen isotopes. Absolute zero was defined as precisely 0 K and −273.15 °C. It is the temperature at which all classical translational motion of the particles comprising matter ceases and they are at complete rest in the classical model. Quantum-mechanically, however, zero-point motion remains and has an associated energy, the zero-point energy. Matter is in its ground state, and contains no thermal energy. The temperatures 273.16 K and 0.01 °C were defined as those of the triple point of water. This definition served the following purposes: it fixed the magnitude of the kelvin as being precisely 1 part in 273.16 parts of the difference between absolute zero and the triple point of water; it established that one kelvin has precisely the same magnitude as one degree on the Celsius scale; and it established the difference between the null points of these scales as being 273.15 K (0 K = −273.15 °C and 273.16 K = 0.01 °C). Since 2019, there has been a new definition based on the Boltzmann constant, but the scales are scarcely changed.
In the United States, the Fahrenheit scale is the most widely used. On this scale the freezing point of water corresponds to 32 °F and the boiling point to 212 °F. The Rankine scale, still used in fields of chemical engineering in the US, is an absolute scale based on the Fahrenheit increment.
Historical scales[edit]
See also: Conversion of scales of temperature
The following temperature scales are in use or have historically been used for measuring temperature:
Kelvin scale
Celsius scale
Fahrenheit scale
Rankine scale
Delisle scale
Newton scale
Réaumur scale
Rømer scale
Plasma physics[edit]
The field of plasma physics deals with phenomena of electromagnetic nature that involve very high temperatures. It is customary to express temperature as energy in a unit related to the electronvolt or kiloelectronvolt (eV/kB or keV/kB). The corresponding energy, which is dimensionally distinct from temperature, is then calculated as the product of the Boltzmann constant and temperature,
E
=
k
B
T
{\displaystyle E=k_{\text{B}}T}
. Then, 1 eV/kB is 11605 K. In the study of QCD matter one routinely encounters temperatures of the order of a few hundred MeV/kB, equivalent to about 10 K.
Continuous or discrete[edit]
When one measures the variation of temperature across a region of space or time, do the temperature measurements turn out to be continuous or discrete? There is a widely held misconception that such temperature measurements must always be continuous. This misconception partly originates from the historical view associated with the continuity of classical physical quantities, which states that physical quantities must assume every intermediate value between a starting value and a final value. However, the classical picture is only true in the cases where temperature is measured in a system that is in equilibrium, that is, temperature may not be continuous outside these conditions. For systems outside equilibrium, such as at interfaces between materials (e.g., a metal/non-metal interface or a liquid-vapour interface) temperature measurements may show steep discontinuities in time and space. For instance, Fang and Ward were some of the first authors to successfully report temperature discontinuities of as much as 7.8 K at the surface of evaporating water droplets. This was reported at inter-molecular scales, or at the scale of the mean free path of molecules which is typically of the order of a few micrometers in gases at room temperature. Generally speaking, temperature discontinuities are considered to be norms rather than exceptions in cases of interfacial heat transfer. This is due to the abrupt change in the vibrational or thermal properties of the materials across such interfaces which prevent instantaneous transfer of heat and the establishment of thermal equilibrium (a prerequisite for having a uniform equilibrium temperature across the interface). Further, temperature measurements at the macro-scale (typical observational scale) may be too coarse-grained as they average out the microscopic thermal information based on the scale of the representative sample volume of the control system, and thus it is likely that temperature discontinuities at the micro-scale may be overlooked in such averages. Such an averaging may even produce incorrect or misleading results in many cases of temperature measurements, even at macro-scales, and thus it is prudent that one examines the micro-physical information carefully before averaging out or smoothing out any potential temperature discontinuities in a system as such discontinuities cannot always be averaged or smoothed out. Temperature discontiuities, rather than merely being anomalies, have actually substantially improved our understanding and predictive abilities pertaining to heat transfer at small scales.
Theoretical foundation[edit]
See also: Thermodynamic temperature
Historically, there are several scientific approaches to the explanation of temperature: the classical thermodynamic description based on macroscopic empirical variables that can be measured in a laboratory; the kinetic theory of gases which relates the macroscopic description to the probability distribution of the energy of motion of gas particles; and a microscopic explanation based on statistical physics and quantum mechanics. In addition, rigorous and purely mathematical treatments have provided an axiomatic approach to classical thermodynamics and temperature. Statistical physics provides a deeper understanding by describing the atomic behavior of matter and derives macroscopic properties from statistical averages of microscopic states, including both classical and quantum states. In the fundamental physical description, the temperature may be measured directly in units of energy. However, in the practical systems of measurement for science, technology, and commerce, such as the modern metric system of units, the macroscopic and the microscopic descriptions are interrelated by the Boltzmann constant, a proportionality factor that scales temperature to the microscopic mean kinetic energy.
The microscopic description in statistical mechanics is based on a model that analyzes a system into its fundamental particles of matter or into a set of classical or quantum-mechanical oscillators and considers the system as a statistical ensemble of microstates. As a collection of classical material particles, the temperature is a measure of the mean energy of motion, called translational kinetic energy, of the particles, whether in solids, liquids, gases, or plasmas. The kinetic energy, a concept of classical mechanics, is half the mass of a particle times its speed squared. In this mechanical interpretation of thermal motion, the kinetic energies of material particles may reside in the velocity of the particles of their translational or vibrational motion or in the inertia of their rotational modes. In monatomic perfect gases and, approximately, in most gas and in simple metals, the temperature is a measure of the mean particle translational kinetic energy, 3/2 kBT. It also determines the probability distribution function of energy. In condensed matter, and particularly in solids, this purely mechanical description is often less useful and the oscillator model provides a better description to account for quantum mechanical phenomena. Temperature determines the statistical occupation of the microstates of the ensemble. The microscopic definition of temperature is only meaningful in the thermodynamic limit, meaning for large ensembles of states or particles, to fulfill the requirements of the statistical model.
Kinetic energy is also considered as a component of thermal energy. The thermal energy may be partitioned into independent components attributed to the degrees of freedom of the particles or to the modes of oscillators in a thermodynamic system. In general, the number of these degrees of freedom that are available for the equipartitioning of energy depends on the temperature, i.e. the energy region of the interactions under consideration. For solids, the thermal energy is associated primarily with the vibrations of its atoms or molecules about their equilibrium position. In an ideal monatomic gas, the kinetic energy is found exclusively in the purely translational motions of the particles. In other systems, vibrational and rotational motions also contribute degrees of freedom.
Kinetic theory of gases[edit]
A theoretical understanding of temperature in a hard-sphere model of a gas can be obtained from the Kinetic theory.
Maxwell and Boltzmann developed a kinetic theory that yields a fundamental understanding of temperature in gases.
This theory also explains the ideal gas law and the observed heat capacity of monatomic (or 'noble') gases.
Plots of pressure vs temperature for three different gas samples extrapolated to absolute zero
The ideal gas law is based on observed empirical relationships between pressure (p), volume (V), and temperature (T), and was recognized long before the kinetic theory of gases was developed (see Boyle's and Charles's laws). The ideal gas law states:
p
V
=
n
R
T
,
{\displaystyle pV=nRT,}
where n is the number of moles of gas and R = 8.314462618... J⋅mol⋅K is the gas constant.
This relationship gives us our first hint that there is an absolute zero on the temperature scale, because it only holds if the temperature is measured on an absolute scale such as Kelvin's. The ideal gas law allows one to measure temperature on this absolute scale using the gas thermometer. The temperature in kelvins can be defined as the pressure in pascals of one mole of gas in a container of one cubic meter, divided by the gas constant.
Although it is not a particularly convenient device, the gas thermometer provides an essential theoretical basis by which all thermometers can be calibrated. As a practical matter, it is not possible to use a gas thermometer to measure absolute zero temperature since the gases condense into a liquid long before the temperature reaches zero. It is possible, however, to extrapolate to absolute zero by using the ideal gas law, as shown in the figure.
The kinetic theory assumes that pressure is caused by the force associated with individual atoms striking the walls, and that all energy is translational kinetic energy. Using a sophisticated symmetry argument, Boltzmann deduced what is now called the Maxwell–Boltzmann probability distribution function for the velocity of particles in an ideal gas. From that probability distribution function, the average kinetic energy (per particle) of a monatomic ideal gas is
E
k
=
1
2
m
v
rms
2
=
3
2
k
B
T
,
{\displaystyle E_{\text{k}}={\frac {1}{2}}mv_{\text{rms}}^{2}={\frac {3}{2}}k_{\text{B}}T,}
where the Boltzmann constant kB is the ideal gas constant divided by the Avogadro number, and
v
rms
=
⟨
v
2
⟩
=
⟨
v
⋅
v
⟩
{\textstyle v_{\text{rms}}={\sqrt {\langle v^{2}\rangle }}={\sqrt {\langle \mathbf {v\cdot v} \rangle }}}
is the root-mean-square speed. This direct proportionality between temperature and mean molecular kinetic energy is a special case of the equipartition theorem, and holds only in the classical limit of a perfect gas. It does not hold exactly for most substances.
Zeroth law of thermodynamics[edit]
Main article: Zeroth law of thermodynamics
When two otherwise isolated bodies are connected together by a rigid physical path impermeable to matter, there is the spontaneous transfer of energy as heat from the hotter to the colder of them. Eventually, they reach a state of mutual thermal equilibrium, in which heat transfer has ceased, and the bodies' respective state variables have settled to become unchanging.
One statement of the zeroth law of thermodynamics is that if two systems are each in thermal equilibrium with a third system, then they are also in thermal equilibrium with each other.
This statement helps to define temperature but it does not, by itself, complete the definition. An empirical temperature is a numerical scale for the hotness of a thermodynamic system. Such hotness may be defined as existing on a one-dimensional manifold, stretching between hot and cold. Sometimes the zeroth law is stated to include the existence of a unique universal hotness manifold, and of numerical scales on it, so as to provide a complete definition of empirical temperature. To be suitable for empirical thermometry, a material must have a monotonic relation between hotness and some easily measured state variable, such as pressure or volume, when all other relevant coordinates are fixed. An exceptionally suitable system is the ideal gas, which can provide a temperature scale that matches the absolute Kelvin scale. The Kelvin scale is defined on the basis of the second law of thermodynamics.
Second law of thermodynamics[edit]
Main article: Second law of thermodynamics
As an alternative to considering or defining the zeroth law of thermodynamics, it was the historical development in thermodynamics to define temperature in terms of the second law of thermodynamics which deals with entropy. The second law states that any process will result in either no change or a net increase in the entropy of the universe. This can be understood in terms of probability.
For example, in a series of coin tosses, a perfectly ordered system would be one in which either every toss comes up heads or every toss comes up tails. This means the outcome is always 100% the same result. In contrast, many mixed (disordered) outcomes are possible, and their number increases with each toss. Eventually, the combinations of ~50% heads and ~50% tails dominate, and obtaining an outcome significantly different from 50/50 becomes increasingly unlikely. Thus the system naturally progresses to a state of maximum disorder or entropy.
As temperature governs the transfer of heat between two systems and the universe tends to progress toward a maximum of entropy, it is expected that there is some relationship between temperature and entropy. A heat engine is a device for converting thermal energy into mechanical energy, resulting in the performance of work. An analysis of the Carnot heat engine provides the necessary relationships. According to energy conservation and energy being a state function that does not change over a full cycle, the work from a heat engine over a full cycle is equal to the net heat, i.e. the sum of the heat put into the system at high temperature, qH > 0, and the waste heat given off at the low temperature, qC < 0.
The efficiency is the work divided by the heat input:
efficiency
=
w
cy
q
H
=
q
H
+
q
C
q
H
=
1
−
|
q
C
|
q
H
,
{\displaystyle {\text{efficiency}}={\frac {w_{\text{cy}}}{q_{\text{H}}}}={\frac {q_{\text{H}}+q_{\text{C}}}{q_{\text{H}}}}=1-{\frac {|q_{\text{C}}|}{q_{\text{H}}}},}
(4)
where wcy is the work done per cycle. The efficiency depends only on |qC|/qH. Because qC and qH correspond to heat transfer at the temperatures TC and TH, respectively, |qC|/qH should be some function of these temperatures:
|
q
C
|
q
H
=
f
(
T
H
,
T
C
)
.
{\displaystyle {\frac {|q_{\text{C}}|}{q_{\text{H}}}}=f\left(T_{\text{H}},T_{\text{C}}\right).}
(5)
Carnot's theorem states that all reversible engines operating between the same heat reservoirs are equally efficient. Thus, a heat engine operating between T1 and T3 must have the same efficiency as one consisting of two cycles, one between T1 and T2, and the second between T2 and T3. This can only be the case if
q
13
=
q
1
q
2
q
2
q
3
,
{\displaystyle q_{13}={\frac {q_{1}q_{2}}{q_{2}q_{3}}},}
which implies
q
13
=
f
(
T
1
,
T
3
)
=
f
(
T
1
,
T
2
)
f
(
T
2
,
T
3
)
.
{\displaystyle q_{13}=f\left(T_{1},T_{3}\right)=f\left(T_{1},T_{2}\right)f\left(T_{2},T_{3}\right).}
Since the first function is independent of T2, this temperature must cancel on the right side, meaning f(T1, T3) is of the form g(T1)/g(T3) (i.e. f(T1, T3) = f(T1, T2)f(T2, T3) = g(T1)/g(T2) · g(T2)/g(T3) = g(T1)/g(T3)), where g is a function of a single temperature. A temperature scale can now be chosen with the property that
|
q
C
|
q
H
=
T
C
T
H
.
{\displaystyle {\frac {|q_{\text{C}}|}{q_{\text{H}}}}={\frac {T_{\text{C}}}{T_{\text{H}}}}.}
(6)
Substituting (6) back into (4) gives a relationship for the efficiency in terms of temperature:
efficiency
=
1
−
|
q
C
|
q
H
=
1
−
T
C
T
H
.
{\displaystyle {\text{efficiency}}=1-{\frac {|q_{\text{C}}|}{q_{\text{H}}}}=1-{\frac {T_{\text{C}}}{T_{\text{H}}}}.}
(7)
For TC = 0 K the efficiency is 100% and that efficiency becomes greater than 100% below 0 K. Since an efficiency greater than 100% violates the first law of thermodynamics, this implies that 0 K is the minimum possible temperature. In fact, the lowest temperature ever obtained in a macroscopic system was 20 nK, which was achieved in 1995 at NIST. Subtracting the right hand side of (5) from the middle portion and rearranging gives
q
H
T
H
+
q
C
T
C
=
0
,
{\displaystyle {\frac {q_{\text{H}}}{T_{\text{H}}}}+{\frac {q_{\text{C}}}{T_{\text{C}}}}=0,}
where the negative sign indicates heat ejected from the system. This relationship suggests the existence of a state function, S, whose change characteristically vanishes for a complete cycle if it is defined by
d
S
=
d
q
rev
T
,
{\displaystyle dS={\frac {dq_{\text{rev}}}{T}},}
(8)
where the subscript indicates a reversible process. This function corresponds to the entropy of the system, which was described previously. Rearranging (8) gives a formula for temperature in terms of fictive infinitesimal quasi-reversible elements of entropy and heat:
T
=
d
q
rev
d
S
.
{\displaystyle T={\frac {dq_{\text{rev}}}{dS}}.}
(9)
For a constant-volume system where entropy S(E) is a function of its energy E, dE = dqrev and (9) gives
T
−
1
=
d
d
E
S
(
E
)
,
{\displaystyle T^{-1}={\frac {d}{dE}}S(E),}
(10)
i.e. the reciprocal of the temperature is the rate of increase of entropy with respect to energy at constant volume.
Definition from statistical mechanics[edit]
Statistical mechanics defines temperature based on a system's fundamental degrees of freedom. Eq.(10) is the defining relation of temperature, where the entropy
S
{\displaystyle S}
is defined (up to a constant) by the logarithm of the number of microstates of the system in the given macrostate (as specified in the microcanonical ensemble):
S
=
k
B
ln
(
W
)
{\displaystyle S=k_{\mathrm {B} }\ln(W)}
where
k
B
{\displaystyle k_{\mathrm {B} }}
is the Boltzmann constant and W is the number of microstates with the energy E of the system (degeneracy).
When two systems with different temperatures are put into purely thermal connection, heat will flow from the higher temperature system to the lower temperature one; thermodynamically this is understood by the second law of thermodynamics: The total change in entropy following a transfer of energy
Δ
E
{\displaystyle \Delta E}
from system 1 to system 2 is:
Δ
S
=
−
(
d
S
/
d
E
)
1
⋅
Δ
E
+
(
d
S
/
d
E
)
2
⋅
Δ
E
=
(
1
T
2
−
1
T
1
)
Δ
E
{\displaystyle \Delta S=-(dS/dE)_{1}\cdot \Delta E+(dS/dE)_{2}\cdot \Delta E=\left({\frac {1}{T_{2}}}-{\frac {1}{T_{1}}}\right)\Delta E}
and is thus positive if
T
1
>
T
2
{\displaystyle T_{1}>T_{2}}
From the point of view of statistical mechanics, the total number of microstates in the combined system 1 + system 2 is
N
1
⋅
N
2
{\displaystyle N_{1}\cdot N_{2}}
, the logarithm of which (times the Boltzmann constant) is the sum of their entropies; thus a flow of heat from high to low temperature, which brings an increase in total entropy, is more likely than any other scenario (normally it is much more likely), as there are more microstates in the resulting macrostate.
Generalized temperature from single-particle statistics[edit]
It is possible to extend the definition of temperature even to systems of few particles, like in a quantum dot. The generalized temperature is obtained by considering time ensembles instead of configuration-space ensembles given in statistical mechanics in the case of thermal and particle exchange between a small system of fermions (N even less than 10) with a single/double-occupancy system. The finite quantum grand canonical ensemble, obtained under the hypothesis of ergodicity and orthodicity, allows expressing the generalized temperature from the ratio of the average time of occupation
τ
1
{\displaystyle \tau _{1}}
and
τ
2
{\displaystyle \tau _{2}}
of the single/double-occupancy system:
T
=
E
−
E
F
(
1
+
3
2
N
)
k
B
ln
(
2
τ
2
τ
1
)
,
{\displaystyle T={\frac {E-E_{\text{F}}\left(1+{\frac {3}{2N}}\right)}{k_{\text{B}}\ln \left(2{\frac {\tau _{2}}{\tau _{1}}}\right)}},}
where EF is the Fermi energy. This generalized temperature tends to the ordinary temperature when N goes to infinity.
Negative temperature[edit]
Main article: Negative temperature
On the empirical temperature scales that are not referenced to absolute zero, a negative temperature is one below the zero-point of the scale used. For example, dry ice has a sublimation temperature of −78.5 °C which is equivalent to −109.3 °F. On the absolute Kelvin scale this temperature is 194.6 K. No body can be brought to exactly 0 K (the temperature of the ideally coldest possible body) by any finite practicable process; this is a consequence of the third law of thermodynamics.
The international kinetic theory temperature of a body cannot take negative values. The thermodynamic temperature scale, however, is not so constrained.
For a body of matter, there can sometimes be conceptually defined, in terms of microscopic degrees of freedom, namely particle spins, a subsystem, with a temperature other than that of the whole body. When the body is in its own state of internal thermodynamic equilibrium, the temperatures of the whole body and of the subsystem must be the same. The two temperatures can differ when, by work through externally imposed force fields, energy can be transferred to and from the subsystem, separately from the rest of the body; then the whole body is not in its own state of internal thermodynamic equilibrium. There is an upper limit of energy such a spin subsystem can attain.
Considering the subsystem to be in a temporary state of virtual thermodynamic equilibrium, it is possible to obtain a negative temperature on the thermodynamic scale. Thermodynamic temperature is the inverse of the derivative of the subsystem's entropy with respect to its internal energy. As the subsystem's internal energy increases, the entropy increases for some range, but eventually attains a maximum value and then begins to decrease as the highest energy states begin to fill. At the point of maximum entropy, the temperature function shows the behavior of a singularity, because the slope of the entropy as a function of energy decreases to zero and then turns negative. As the subsystem's entropy reaches its maximum, its thermodynamic temperature goes to positive infinity, switching to negative infinity as the slope turns negative. Such negative temperatures are hotter than any positive temperature. Over time, when the subsystem is exposed to the rest of the body, which has a positive temperature, energy is transferred as heat from the negative temperature subsystem to the positive temperature system. The kinetic theory temperature is not defined for such subsystems.
Examples[edit]
An illustration of the range of cosmic temperatures
Main article: Orders of magnitude (temperature)
Comparisons of temperatures in various scales
Temperature
Peak emittance wavelength of black-body radiation
Kelvin
Celsius
Absolute zero(precisely by definition)
0 K
−273.15 °C
Infinity
Blackbody temperature of the black hole atthe centre of our galaxy, Sagittarius A*
15 fK
−273.149999999999985 °C
2.5×10 km (1.7 AU)
Lowest temperatureachieved
100 pK
−273.149999999900 °C
29000 km
Coldest Bose–Einstein condensate
450 pK
−273.14999999955 °C
6400 km
One millikelvin(precisely by definition)
0.001 K
−273.149 °C
2.89777 m(radio, FM band)
Cosmic microwave background(2013 measurement)
2.7260 K
−270.424 °C
0.00106301 m(millimeter-wavelength microwave)
Water triple point(previously by definition)
273.16 K
0.01 °C
10608.3 nm(long-wavelength IR)
Water boiling point
373.1339 K
99.9839 °C
7766.03 nm(mid-wavelength IR)
Iron melting point
1811 K
1538 °C
1600 nm(far infrared)
Incandescent lamp
2500 K
≈2200 °C
1160 nm(near infrared)
Sun's visible surface
5778 K
5505 °C
501.5 nm(green-blue light)
Lightning boltchannel
28 kK
28000 °C
100 nm(far ultraviolet light)
Sun's core
16 MK
16 million °C
0.18 nm(X-rays)
Thermonuclear weapon(peak temperature)
350 MK
350 million °C
8.3×10 nm(gamma rays)
Sandia National Labs'Z machine
2 GK
2 billion °C
1.4×10 nm(gamma rays)
Core of a high-massstar on its last day
3 GK
3 billion °C
1×10 nm(gamma rays)
Merging binary neutronstar system
350 GK
350 billion °C
8×10 nm(gamma rays)
Relativistic HeavyIon Collider
1 TK
1 trillion °C
3×10 nm(gamma rays)
CERN's proton vs nucleus collisions
10 TK
10 trillion °C
3×10 nm(gamma rays)
For Vienna Standard Mean Ocean Water at one standard atmosphere (101.325 kPa) when calibrated strictly per the two-point definition of thermodynamic temperature.
The 2500 K value is approximate. The 273.15 K difference between K and °C is rounded to 300 K to avoid false precision in the Celsius value.
For a true black-body (which tungsten filaments are not). Tungsten filament emissivity is greater at shorter wavelengths, which makes them appear whiter.
Effective photosphere temperature. The 273.15 K difference between K and °C is rounded to 273 K to avoid false precision in the Celsius value.
The 273.15 K difference between K and °C is within the precision of these values.
For a true black-body (which the plasma was not). The Z machine's dominant emission originated from 40 MK electrons (soft x-ray emissions) within the plasma.
See also[edit]
Atmospheric temperature – Physical quantity that expresses hot and cold in the atmosphere
Body temperature – Ability of an organism to keep its body temperature within certain boundaries (thermoregulation)
Color temperature – Property of light sources related to black-body radiation
Dry-bulb temperature – Temperature of air as measured by a thermometer shielded from radiation and moisture
Thermal conduction – Process by which heat is transferred within an object
Convective heat transfer – Heat transfer due to combined effects of advection and diffusionPages displaying short descriptions of redirect targets
Instrumental temperature record – In situ measurements that provide the temperature of Earth's climate system
ISO 1 – ISO standard temperature, 20°C
International Temperature Scale of 1990 (ITS-90) – Practical temperature scale
Laser schlieren deflectometry
List of cities by average temperature
Maxwell's demon – Thought experiment of 1867
Orders of magnitude (temperature) – Range of temperatures from absolute zero to very high
Outside air temperature
Planck temperature – Units defined only by physical constantsPages displaying short descriptions of redirect targets
Rankine scale – Absolute temperature scale using Fahrenheit degrees
Relativistic heat conduction – Model compatible with special relativity
Satellite temperature measurements – Measurements of atmospheric, land surface or sea temperature by satellites.Pages displaying short descriptions of redirect targets
Scale of temperature – Method to measure temperature quantitatively
Sea surface temperature – Water temperature close to the ocean's surface
Stagnation temperature
Thermal radiation – Electromagnetic radiation generated by the thermal motion of particles
Thermoception – Sensation and perception of temperature
Thermodynamic (absolute) temperature – Measure of absolute temperature
Thermography – Infrared imaging used to reveal temperature
Thermometer – Device to measure temperature
Virtual temperature – Virtual temperature of a moist air parcel
Wet-bulb globe temperature – Apparent temperature estimating how humans are affected
Wet-bulb temperature – Temperature read by a thermometer covered in water-soaked cloth
Notes and references[edit]
Notes
^ The cited emission wavelengths are for black bodies in equilibrium. CODATA 2006 recommended value of 2.8977685(51)×10 m K used for Wien displacement law constant b.
^ A temperature of 450 ±80 pK in a Bose–Einstein condensate (BEC) of sodium atoms was achieved in 2003 by researchers at MIT. Citation: Cooling Bose–Einstein Condensates Below 500 Picokelvin, A.E. Leanhardt et al., Science 301, 12 Sept. 2003, p. 1515. This record's peak emittance black-body wavelength of 6,400 kilometers is roughly the radius of Earth.
^ The peak emittance wavelength of 2.89777 m is a frequency of 103.456 MHz
^ Since 2019, Kelvin is now defined on the Boltzmann constant, so that the triple point is 273.16±0.0001 K
^ Measurement was made in 2002 and has an uncertainty of ±3 kelvins. A 1989 measurement Archived 2010-02-11 at the Wayback Machine produced a value of 5,777.0±2.5 K. Citation: Overview of the Sun (Chapter 1 lecture notes on Solar Physics by Division of Theoretical Physics, Dept. of Physical Sciences, University of Helsinki).
^ The 350 MK value is the maximum peak fusion fuel temperature in a thermonuclear weapon of the Teller–Ulam configuration (commonly known as a hydrogen bomb). Peak temperatures in Gadget-style fission bomb cores (commonly known as an atomic bomb) are in the range of 50 to 100 MK. Citation: Nuclear Weapons Frequently Asked Questions, 3.2.5 Matter At High Temperatures. Link to relevant Web page. Archived 2007-05-03 at the Wayback Machine All referenced data was compiled from publicly available sources.
^ Peak temperature for a bulk quantity of matter was achieved by a pulsed-power machine used in fusion physics experiments. The term bulk quantity draws a distinction from collisions in particle accelerators wherein high temperature applies only to the debris from two subatomic particles or nuclei at any given instant. The >2 GK temperature was achieved over a period of about ten nanoseconds during shot Z1137. In fact, the iron and manganese ions in the plasma averaged 3.58±0.41 GK (309±35 keV) for 3 ns (ns 112 through 115). Ion Viscous Heating in a Magnetohydrodynamically Unstable Z Pinch at Over 2×10 Kelvin, M.G. Haines et al., Physical Review Letters 96 (2006) 075003. Link to Sandia's news release. Archived 2010-05-30 at the Wayback Machine
^ Core temperature of a high–mass (>8–11 solar masses) star after it leaves the main sequence on the Hertzsprung–Russell diagram and begins the alpha process (which lasts one day) of fusing silicon–28 into heavier elements in the following steps: sulfur–32 → argon–36 → calcium–40 → titanium–44 → chromium–48 → iron–52 → nickel–56. Within minutes of finishing the sequence, the star explodes as a Type II supernova. Citation: Holland, Arthur; Williams, Mark. "Stellar Evolution: The Life and Death of Our Luminous Neighbors". GS265. University of Michigan. Archived from the original on 2009-01-16. More informative links can be found here "Chapter 21 Stellar Explosions". Archived from the original on 2013-04-11. Retrieved 2016-02-08., and here "Trans". Archived from the original on 2011-08-14. Retrieved 2016-02-08., and a concise treatise on stars by NASA is here "NASA - Star". Archived from the original on 2010-10-24. Retrieved 2010-10-12..
^ Based on a computer model that predicted a peak internal temperature of 30 MeV (350 GK) during the merger of a binary neutron star system (which produces a gamma–ray burst). The neutron stars in the model were 1.2 and 1.6 solar masses respectively, were roughly 20 km in diameter, and were orbiting around their barycenter (common center of mass) at about 390 Hz during the last several milliseconds before they completely merged. The 350 GK portion was a small volume located at the pair's developing common core and varied from roughly 1 to 7 km across over a time span of around 5 ms. Imagine two city-sized objects of unimaginable density orbiting each other at the same frequency as the G4 musical note (the 28th white key on a piano). It's also noteworthy that at 350 GK, the average neutron has a vibrational speed of 30% the speed of light and a relativistic mass (m) 5% greater than its rest mass (m0). Torus Formation in Neutron Star Mergers and Well-Localized Short Gamma-Ray Bursts Archived 2017-11-22 at the Wayback Machine, R. Oechslin et al. of Max Planck Institute for Astrophysics. Archived 2005-04-03 at the Wayback Machine, arXiv:astro-ph/0507099 v2, 22 Feb. 2006. An html summary Archived 2010-11-09 at the Wayback Machine.
Citations
^ Agency, International Atomic Energy (1974). Thermal discharges at nuclear power stations: their management and environmental impacts: a report prepared by a group of experts as the result of a panel meeting held in Vienna, 23–27 October 1972. International Atomic Energy Agency.
^ Watkinson, John (2001). The Art of Digital Audio. Taylor & Francis. ISBN 978-0-240-51587-8.
^ Middleton, W.E.K. (1966), pp. 89–105.
^ Joanna Thompson (2021-10-14). "Scientists just broke the record for the coldest temperature ever recorded in a lab". livescience.com. Retrieved 2023-04-28.
^ Jaynes, E.T. (1965), pp. 391–398.
^ Cryogenic Society Archived 2020-11-07 at the Wayback Machine (2019).
^ Draft Resolution A "On the revision of the International System of Units (SI)" to be submitted to the CGPM at its 26th meeting (2018) (PDF), archived from the original (PDF) on 2018-04-29, retrieved 2019-10-20
^ Truesdell, C.A. (1980), Sections 11 B, 11H, pp. 306–310, 320–332.
^ Quinn, T. J. (1983).
^ Germer, L.H. (1925). 'The distribution of initial velocities among thermionic electrons', Phys. Rev., 25: 795–807. here
^ Turvey, K. (1990). 'Test of validity of Maxwellian statistics for electrons thermionically emitted from an oxide cathode', European Journal of Physics, 11(1): 51–59. here
^ Zeppenfeld, M., Englert, B.G.U., Glöckner, R., Prehn, A., Mielenz, M., Sommer, C., van Buuren, L.D., Motsch, M., Rempe, G. (2012).
^ Miller, J. (2013).
^ de Podesta, M., Underwood, R., Sutton, G., Morantz, P, Harris, P, Mark, D.F., Stuart, F.M., Vargha, G., Machin, M. (2013). A low-uncertainty measurement of the Boltzmann constant, Metrologia, 50 (4): S213–S216, BIPM & IOP Publishing Ltd
^ Quinn, T.J. (1983), pp. 98–107.
^ Schooley, J.F. (1986), pp. 138–143.
^ Quinn, T.J. (1983), pp. 61–83.
^ Schooley, J.F. (1986), pp. 115–138.
^ Adkins, C.J. (1968/1983), pp. 119–120.
^ Buchdahl, H.A. (1966), pp. 137–138.
^ Tschoegl, N.W. (2000), p. 88.
^ Fermi, E. (1956). Thermodynamics. Dover Publications (still in print). p. 48. eq.(64).
^ Thomson, W. (Lord Kelvin) (1848).
^ Thomson, W. (Lord Kelvin) (1851).
^ Partington, J.R. (1949), pp. 175–177.
^ Roberts, J.K., Miller, A.R. (1928/1960), pp. 321–322.
^ Quinn, T.J. (1983). Temperature, Academic Press, London, ISBN 0-12-569680-9, pp. 160–162.
^ Tisza, L. (1966). Generalized Thermodynamics, M.I.T. Press, Cambridge MA, pp. 47, 57.
^ Münster, A. (1970), Classical Thermodynamics, translated by E.S. Halberstadt, Wiley–Interscience, London, ISBN 0-471-62430-6, pp. 49, 69.
^ Bailyn, M. (1994). A Survey of Thermodynamics, American Institute of Physics Press, New York, ISBN 0-88318-797-3, pp. 14–15, 214.
^ Callen, H.B. (1960/1985), Thermodynamics and an Introduction to Thermostatistics, (first edition 1960), second edition 1985, John Wiley & Sons, New York, ISBN 0-471-86256-8, pp. 146–148.
^ Kondepudi, D., Prigogine, I. (1998). Modern Thermodynamics. From Heat Engines to Dissipative Structures, John Wiley, Chichester, ISBN 0-471-97394-7, pp. 115–116.
^ Tisza, L. (1966). Generalized Thermodynamics, M.I.T. Press, Cambridge MA, p. 58.
^ Milne, E.A. (1929). The effect of collisions on monochromatic radiative equilibrium, Monthly Notices of the Royal Astronomical Society, 88: 493–502.
^ Gyarmati, I. (1970). Non-equilibrium Thermodynamics. Field Theory and Variational Principles, translated by E. Gyarmati and W.F. Heinz, Springer, Berlin, pp. 63–66.
^ Glansdorff, P., Prigogine, I., (1971). Thermodynamic Theory of Structure, Stability and Fluctuations, Wiley, London, ISBN 0-471-30280-5, pp. 14–16.
^ Bailyn, M. (1994). A Survey of Thermodynamics, American Institute of Physics Press, New York, ISBN 0-88318-797-3, pp. 133–135.
^ Callen, H.B. (1960/1985), Thermodynamics and an Introduction to Thermostatistics, (first edition 1960), second edition 1985, John Wiley & Sons, New York, ISBN 0-471-86256-8, pp. 309–310.
^ Bryan, G.H. (1907). Thermodynamics. An Introductory Treatise dealing mainly with First Principles and their Direct Applications, B.G. Teubner, Leipzig, p. 3. "Thermodynamics by George Hartley Bryan". Archived from the original on 2011-11-18. Retrieved 2011-10-02.
^ Pippard, A.B. (1957/1966), p. 18.
^ Adkins,C.J. (1968/1983), p. 20.
^ Bryan, G.H. (1907). Thermodynamics. An Introductory Treatise dealing mainly with First Principles and their Direct Applications, B.G. Teubner, Leipzig, p. 5: "... when a body is spoken of as growing hotter or colder an increase of temperature is always implied, for the hotness and coldness of a body are qualitative terms which can only refer to temperature." "Thermodynamics by George Hartley Bryan". Archived from the original on 2011-11-18. Retrieved 2011-10-02.
^ Mach, E. (1900). Die Principien der Wärmelehre. Historisch-kritisch entwickelt, Johann Ambrosius Barth, Leipzig, section 22, pp. 56–57.
^ Serrin, J. (1986). Chapter 1, 'An Outline of Thermodynamical Structure', pp. 3–32, especially p. 6, in New Perspectives in Thermodynamics, edited by J. Serrin, Springer, Berlin, ISBN 3-540-15931-2.
^ Maxwell, J.C. (1872). Theory of Heat, third edition, Longmans, Green, London, p. 32.
^ Tait, P.G. (1884). Heat, Macmillan, London, Chapter VII, pp. 39–40.
^ Planck, M. (1897/1903). Treatise on Thermodynamics, translated by A. Ogg, Longmans, Green, London, pp. 1–2.
^ Planck, M. (1914), The Theory of Heat Radiation Archived 2011-11-18 at the Wayback Machine, second edition, translated into English by M. Masius, Blakiston's Son & Co., Philadelphia, reprinted by Kessinger.
^ J.S. Dugdale (1996). Entropy and its Physical Interpretation. Taylor & Francis. p. 13. ISBN 978-0-7484-0569-5.
^ F. Reif (1965). Fundamentals of Statistical and Thermal Physics. McGraw-Hill. p. 102. ISBN 9780070518001.
^ M.J. Moran; H.N. Shapiro (2006). "1.6.1". Fundamentals of Engineering Thermodynamics (5 ed.). John Wiley & Sons, Ltd. p. 14. ISBN 978-0-470-03037-0.
^ T.W. Leland, Jr. "Basic Principles of Classical and Statistical Thermodynamics" (PDF). p. 14. Archived (PDF) from the original on 2011-09-28. Consequently we identify temperature as a driving force which causes something called heat to be transferred.
^ Tait, P.G. (1884). Heat, Macmillan, London, Chapter VII, pp. 42, 103–117.
^ Beattie, J.A., Oppenheim, I. (1979). Principles of Thermodynamics, Elsevier Scientific Publishing Company, Amsterdam, ISBN 978-0-444-41806-7, p. 29.
^ Landsberg, P.T. (1961). Thermodynamics with Quantum Statistical Illustrations, Interscience Publishers, New York, p. 17.
^ Thomsen, J.S. (1962). "A restatement of the zeroth law of thermodynamics". Am. J. Phys. 30 (4): 294–296. Bibcode:1962AmJPh..30..294T. doi:10.1119/1.1941991.
^ Maxwell, J.C. (1872). Theory of Heat, third edition, Longman's, Green & Co, London, p. 45.
^ Pitteri, M. (1984). On the axiomatic foundations of temperature, Appendix G6 on pp. 522–544 of Rational Thermodynamics, C. Truesdell, second edition, Springer, New York, ISBN 0-387-90874-9.
^ Truesdell, C., Bharatha, S. (1977). The Concepts and Logic of Classical Thermodynamics as a Theory of Heat Engines, Rigorously Constructed upon the Foundation Laid by S. Carnot and F. Reech, Springer, New York, ISBN 0-387-07971-8, p. 20.
^ Serrin, J. (1978). The concepts of thermodynamics, in Contemporary Developments in Continuum Mechanics and Partial Differential Equations. Proceedings of the International Symposium on Continuum Mechanics and Partial Differential Equations, Rio de Janeiro, August 1977, edited by G.M. de La Penha, L.A.J. Medeiros, North-Holland, Amsterdam, ISBN 0-444-85166-6, pp. 411–451.
^ Maxwell, J.C. (1872). Theory of Heat, third edition, Longmans, Green, London, pp. 155–158.
^ Tait, P.G. (1884). Heat, Macmillan, London, Chapter VII, Section 95, pp. 68–69.
^ Buchdahl, H.A. (1966), p. 73.
^ Kondepudi, D. (2008). Introduction to Modern Thermodynamics, Wiley, Chichester, ISBN 978-0-470-01598-8, Section 32., pp. 106–108.
^ Green, Don; Perry, Robert H. (2008). Perry's Chemical Engineers' Handbook, Eighth Edition (8th ed.). McGraw-Hill Education. p. 660. ISBN 978-0071422949.
^ The kelvin in the SI Brochure Archived 2007-09-26 at the Wayback Machine
^ "Absolute Zero". Calphad.com. Archived from the original on 2011-07-08. Retrieved 2010-09-16.
^ Definition agreed by the 26th General Conference on Weights and Measures (CGPM) Archived 2020-10-09 at the Wayback Machine in November 2018, implemented 20 May 2019
^ Jha, Aditya; Campbell, Douglas; Montelle, Clemency; Wilson, Phillip L. (2023-07-30). "On the Continuum Fallacy: Is Temperature a Continuous Function?". Foundations of Physics. 53 (4): 69. Bibcode:2023FoPh...53...69J. doi:10.1007/s10701-023-00713-x. ISSN 1572-9516.
^ van Strien, Marij (2015-10-01). "Continuity in nature and in mathematics: Boltzmann and Poincaré". Synthese. 192 (10): 3275–3295. doi:10.1007/s11229-015-0701-9. ISSN 1573-0964. S2CID 255075377.
^ Fang, G.; Ward, C. A. (1999-01-01). "Temperature measured close to the interface of an evaporating liquid". Physical Review E. 59 (1): 417–428. Bibcode:1999PhRvE..59..417F. doi:10.1103/PhysRevE.59.417.
^ Newell, Homer E. (1960-02-12). "The Space Environment: As man looks forward to flight into space, he finds the outer regions not completely unknown". Science. 131 (3398): 385–390. doi:10.1126/science.131.3398.385. ISSN 0036-8075. PMID 14426791.
^ Chen, Gang (2022-08-01). "On the molecular picture and interfacial temperature discontinuity during evaporation and condensation". International Journal of Heat and Mass Transfer. 191: 122845. arXiv:2201.07318. doi:10.1016/j.ijheatmasstransfer.2022.122845. ISSN 0017-9310. S2CID 246036409.
^ Cahill, D; et al. (27 Dec 2022). "Nanoscale thermal transport". Journal of Applied Physics. 93 (2): 793–818. doi:10.1063/1.1524305. hdl:2027.42/70161. S2CID 15327316. Retrieved 2023-08-02.
^ Chen, Jie; Xu, Xiangfan; Zhou, Jun; Li, Baowen (2022-04-22). "Interfacial thermal resistance: Past, present, and future". Reviews of Modern Physics. 94 (2): 025002. Bibcode:2022RvMP...94b5002C. doi:10.1103/RevModPhys.94.025002. S2CID 248350864.
^ Aursand, Eskil; Ytrehus, Tor (2019-07-01). "Comparison of kinetic theory evaporation models for liquid thin-films". International Journal of Multiphase Flow. 116: 67–79. doi:10.1016/j.ijmultiphaseflow.2019.04.007. hdl:11250/2594950. ISSN 0301-9322. S2CID 146056093.
^ C. Caratheodory (1909). "Untersuchungen über die Grundlagen der Thermodynamik". Mathematische Annalen. 67 (3): 355–386. doi:10.1007/BF01450409. S2CID 118230148.
^ Swendsen, Robert (March 2006). "Statistical mechanics of colloids and Boltzmann's definition of entropy" (PDF). American Journal of Physics. 74 (3): 187–190. Bibcode:2006AmJPh..74..187S. doi:10.1119/1.2174962. S2CID 59471273. Archived from the original (PDF) on 2020-02-28.
^ Balescu, R. (1975). Equilibrium and Nonequilibrium Statistical Mechanics, Wiley, New York, ISBN 0-471-04600-0, pp. 148–154.
^ Kittel, Charles; Kroemer, Herbert (1980). Thermal Physics (2nd ed.). W.H. Freeman Company. pp. 391–397. ISBN 978-0-7167-1088-2.
^ Kondepudi, D.K. (1987). "Microscopic aspects implied by the second law". Foundations of Physics. 17 (7): 713–722. Bibcode:1987FoPh...17..713K. doi:10.1007/BF01889544. S2CID 120576357.
^ The Feynman Lectures on Physics. 39–5 The ideal gas law
^ "2018 CODATA Value: molar gas constant". The NIST Reference on Constants, Units, and Uncertainty. NIST. 20 May 2019. Retrieved 2019-05-20.
^ "Kinetic Theory". galileo.phys.virginia.edu. Archived from the original on 16 July 2017. Retrieved 27 January 2018.
^ Tolman, R.C. (1938). The Principles of Statistical Mechanics, Oxford University Press, London, pp. 93, 655.
^ Peter Atkins, Julio de Paula (2006). Physical Chemistry (8 ed.). Oxford University Press. p. 9.
^ Maxwell, J.C. (1872). Theory of Heat, third edition, Longman's, Green & Co, London, p. 32.
^ Bailyn, M. (1994). A Survey of Thermodynamics, American Institute of Physics Press, New York, ISBN 0-88318-797-3, p. 23, "..., if a temperature gradient exists, ..., then a flow of heat, ..., must occur to achieve a uniform temperature."
^ Guggenheim, E.A. (1967). Thermodynamics. An Advanced Treatment for Chemists and Physicists, North-Holland Publishing Company., Amsterdam, (1st edition 1949) fifth edition 1965, p. 8, "... will gradually adjust themselves until eventually they do reach mutual equilibrium after which there will of course be no further change."
^ Bailyn, M. (1994). A Survey of Thermodynamics, American Institute of Physics Press, New York, ISBN 0-88318-797-3, p. 22.
^ Guggenheim, E.A. (1967). Thermodynamics. An Advanced Treatment for Chemists and Physicists, North-Holland Publishing Company., Amsterdam, (1st edition 1949) fifth edition 1965, p. 8: "If two systems are both in thermal equilibrium with a third system then they are in thermal equilibrium with each other."
^ Buchdahl, H.A. (1966). The Concepts of Classical Thermodynamics, Cambridge University Press, Cambridge, p. 29: "... if each of two systems is in equilibrium with a third system then they are in equilibrium with each other."
^ Planck, M. (1945). Treatise on Thermodynamics. Dover Publications. p. §90 & §137. eqs.(39), (40), & (65).
^ Prati, E. (2010). "The finite quantum grand canonical ensemble and temperature from single-electron statistics for a mesoscopic device". J. Stat. Mech. 1 (1): P01003. arXiv:1001.2342. Bibcode:2010JSMTE..01..003P. doi:10.1088/1742-5468/2010/01/P01003. S2CID 118339343. arxiv.org Archived 2017-11-22 at the Wayback Machine
^ "Realizing Boltzmann's dream: computer simulations in modern statistical mechanics" (PDF). Archived (PDF) from the original on 2014-04-13. Retrieved 2014-04-11.
^ Prati, E.; et al. (2010). "Measuring the temperature of a mesoscopic electron system by means of single electron statistics". Applied Physics Letters. 96 (11): 113109. arXiv:1002.0037. Bibcode:2010ApPhL..96k3109P. doi:10.1063/1.3365204. S2CID 119209143. Archived from the original on 2016-05-14. Retrieved 2022-03-02. arxiv.org Archived 2017-11-22 at the Wayback Machine
^ Water Science School. "Frozen carbon dioxide (dry ice) sublimates directly into a vapor". USGS.
^ Guggenheim, E.A. (1967) [1949], Thermodynamics. An Advanced Treatment for Chemists and Physicists (fifth ed.), Amsterdam: North-Holland Publishing Company., p. 157: "It is impossible by any procedure, no matter how idealized, to reduce the temperature of any system to zero temperature in a finite number of finite operations."
^ Pippard, A.B. (1957/1966). Elements of Classical Thermodynamics for Advanced Students of Physics, original publication 1957, reprint 1966, Cambridge University Press, Cambridge, page 51: "By no finite series of processes is the absolute zero attainable."
^ Tisza, L. (1966). Generalized Thermodynamics, MIT Press, Cambridge MA, page 96: "It is impossible to reach absolute zero as a result of a finite sequence of operations."
^ Kittel, Charles; Kroemer, Herbert (1980). Thermal Physics (2nd ed.). W.H. Freeman Company. p. Appendix E. ISBN 978-0-7167-1088-2.
^ SVS (2023-08-03). "NASA Scientific Visualization Studio | A Guide to Cosmic Temperatures". SVS. Retrieved 2023-08-06.
^ This the Hawking Radiation for a Schwarzschild black hole of mass M = 4.145×10 M☉. It is too faint to be observed.
^ "World record in low temperatures". Archived from the original on 2009-06-18. Retrieved 2009-05-05.
^ Results of research by Stefan Bathe using the PHENIX Archived 2008-11-20 at the Wayback Machine detector on the Relativistic Heavy Ion Collider Archived 2016-03-03 at the Wayback Machine at Brookhaven National Laboratory Archived 2012-06-24 at the Wayback Machine in Upton, New York. Bathe has studied gold-gold, deuteron-gold, and proton-proton collisions to test the theory of quantum chromodynamics, the theory of the strong force that holds atomic nuclei together. Link to news release. Archived 2009-02-11 at the Wayback Machine
^ How do physicists study particles? Archived 2007-10-11 at the Wayback Machine by CERN Archived 2012-07-07 at the Wayback Machine.
Bibliography of cited references[edit]
Adkins, C.J. (1968/1983). Equilibrium Thermodynamics, (1st edition 1968), third edition 1983, Cambridge University Press, Cambridge UK, ISBN 0-521-25445-0.
Buchdahl, H.A. (1966). The Concepts of Classical Thermodynamics, Cambridge University Press, Cambridge.
Jaynes, E.T. (1965). Gibbs vs Boltzmann entropies, American Journal of Physics, 33(5), 391–398.
Middleton, W.E.K. (1966). A History of the Thermometer and its Use in Metrology, Johns Hopkins Press, Baltimore.
Miller, J (2013). "Cooling molecules the optoelectric way". Physics Today. 66 (1): 12–14. Bibcode:2013PhT....66a..12M. doi:10.1063/pt.3.1840.
Partington, J.R. (1949). An Advanced Treatise on Physical Chemistry, volume 1, Fundamental Principles. The Properties of Gases, Longmans, Green & Co., London, pp. 175–177.
Pippard, A.B. (1957/1966). Elements of Classical Thermodynamics for Advanced Students of Physics, original publication 1957, reprint 1966, Cambridge University Press, Cambridge UK.
Quinn, T.J. (1983). Temperature, Academic Press, London, ISBN 0-12-569680-9.
Schooley, J.F. (1986). Thermometry, CRC Press, Boca Raton, ISBN 0-8493-5833-7.
Roberts, J.K., Miller, A.R. (1928/1960). Heat and Thermodynamics, (first edition 1928), fifth edition, Blackie & Son Limited, Glasgow.
Thomson, W. (Lord Kelvin) (1848). On an absolute thermometric scale founded on Carnot's theory of the motive power of heat, and calculated from Regnault's observations, Proc. Camb. Phil. Soc. (1843/1863) 1, No. 5: 66–71.
Thomson, W. (Lord Kelvin) (March 1851). "On the Dynamical Theory of Heat, with numerical results deduced from Mr Joule's equivalent of a Thermal Unit, and M. Regnault's Observations on Steam". Transactions of the Royal Society of Edinburgh. XX (part II): 261–268, 289–298.
Truesdell, C.A. (1980). The Tragicomical History of Thermodynamics, 1822–1854, Springer, New York, ISBN 0-387-90403-4.
Tschoegl, N.W. (2000). Fundamentals of Equilibrium and Steady-State Thermodynamics, Elsevier, Amsterdam, ISBN 0-444-50426-5.
Zeppenfeld, M.; Englert, B.G.U.; Glöckner, R.; Prehn, A.; Mielenz, M.; Sommer, C.; van Buuren, L.D.; Motsch, M.; Rempe, G. (2012). "Sysiphus cooling of electrically trapped polyatomic molecules". Nature. 491 (7425): 570–573. arXiv:1208.0046. Bibcode:2012Natur.491..570Z. doi:10.1038/nature11595. PMID 23151480. S2CID 4367940.
Further reading[edit]
Chang, Hasok (2004). Inventing Temperature: Measurement and Scientific Progress. Oxford: Oxford University Press. ISBN 978-0-19-517127-3.
Zemansky, Mark Waldo (1964). Temperatures Very Low and Very High. Princeton, NJ: Van Nostrand.
External links[edit]
Temperature at Wikipedia's sister projects
Definitions from WiktionaryMedia from CommonsResources from WikiversityData from Wikidata
Current map of global surface temperatures
vteScales of temperature
Celsius
Delisle
Fahrenheit
Gas mark
Kelvin
Leiden
Newton
Rankine
Réaumur
Rømer
Wedgwood
Conversion formulas and comparison
vteMeteorological data and variablesGeneral
Adiabatic processes
Advection
Buoyancy
Lapse rate
Lightning
Surface solar radiation
Surface weather analysis
Visibility
Vorticity
Wind
Wind shear
Condensation
Cloud
Cloud condensation nuclei (CCN)
Fog
Convective condensation level (CCL)
Lifted condensation level (LCL)
Precipitable water
Precipitation
Water vapor
Convection
Convective available potential energy (CAPE)
Convective inhibition (CIN)
Convective instability
Convective momentum transport
Conditional symmetric instability
Convective temperature (Tc)
Equilibrium level (EL)
Free convective layer (FCL)
Helicity
K Index
Level of free convection (LFC)
Lifted index (LI)
Maximum parcel level (MPL)
Bulk Richardson number (BRN)
Temperature
Dew point (Td)
Dew point depression
Dry-bulb temperature
Equivalent temperature (Te)
Forest fire weather index
Haines Index
Heat index
Humidex
Humidity
Relative humidity (RH)
Mixing ratio
Potential temperature (θ)
Equivalent potential temperature (θe)
Sea surface temperature (SST)
Temperature anomaly
Thermodynamic temperature
Vapor pressure
Virtual temperature
Wet-bulb temperature
Wet-bulb globe temperature
Wet-bulb potential temperature
Wind chill
Pressure
Atmospheric pressure
Baroclinity
Barotropicity
Pressure gradient
Pressure-gradient force (PGF)
Velocity
Maximum potential intensity
vteSI base quantitiesBase quantities
Quantity
SI unit
Name
Symbol
Dimensionsymbol
Unitname
Unitsymbol
time, duration
t
T
second
s
length
l, x, r, etc.
L
metre
m
mass
m
M
kilogram
kg
electric current
I , i
I
ampere
A
thermodynamic temperature
T
Θ
kelvin
K
amount of substance
n
N
mole
mol
luminous intensity
Iv
J
candela
cd
See also
History of the metric system
International System of Quantities
2019 redefinition
Systems of measurement
Category
Outline
Authority control databases: National
Spain
France
BnF data
Germany
Israel
United States
Latvia
Japan
Czech Republic
Portal: Energy | biology | 6143428 | https://sv.wikipedia.org/wiki/Steele%20Glacier | Steele Glacier | Steele Glacier är en glaciär i Kanada. Den ligger i territoriet Yukon, i den västra delen av landet, km väster om huvudstaden Ottawa. Steele Glacier ligger meter över havet.
Terrängen runt Steele Glacier är bergig österut, men västerut är den kuperad. Steele Glacier ligger nere i en dal. Den högsta punkten i närheten är meter över havet, km sydost om Steele Glacier. Trakten runt Steele Glacier är nära nog obefolkad, med mindre än två invånare per kvadratkilometer.. Det finns inga samhällen i närheten.
Trakten runt Steele Glacier är permanent täckt av is och snö. Årsmedeltemperaturen i trakten är °C. Den varmaste månaden är juli, då medeltemperaturen är °C, och den kallaste är januari, med °C.
Kommentarer
Källor
Isformationer i Yukon | swedish | 0.990625 |
temperature_sensed/Sensor.txt | A sensor is a device that produces an output signal for the purpose of detecting a physical phenomenon.
In the broadest definition, a sensor is a device, module, machine, or subsystem that detects events or changes in its environment and sends the information to other electronics, frequently a computer processor.
Sensors are used in everyday objects such as touch-sensitive elevator buttons (tactile sensor) and lamps which dim or brighten by touching the base, and in innumerable applications of which most people are never aware. With advances in micromachinery and easy-to-use microcontroller platforms, the uses of sensors have expanded beyond the traditional fields of temperature, pressure and flow measurement, for example into MARG sensors.
Analog sensors such as potentiometers and force-sensing resistors are still widely used. Their applications include manufacturing and machinery, airplanes and aerospace, cars, medicine, robotics and many other aspects of our day-to-day life. There is a wide range of other sensors that measure chemical and physical properties of materials, including optical sensors for refractive index measurement, vibrational sensors for fluid viscosity measurement, and electro-chemical sensors for monitoring pH of fluids.
A sensor's sensitivity indicates how much its output changes when the input quantity it measures changes. For instance, if the mercury in a thermometer moves 1 cm when the temperature changes by 1 °C, its sensitivity is 1 cm/°C (it is basically the slope dy/dx assuming a linear characteristic). Some sensors can also affect what they measure; for instance, a room temperature thermometer inserted into a hot cup of liquid cools the liquid while the liquid heats the thermometer. Sensors are usually designed to have a small effect on what is measured; making the sensor smaller often improves this and may introduce other advantages.
Technological progress allows more and more sensors to be manufactured on a microscopic scale as microsensors using MEMS technology. In most cases, a microsensor reaches a significantly faster measurement time and higher sensitivity compared with macroscopic approaches. Due to the increasing demand for rapid, affordable and reliable information in today's world, disposable sensors—low-cost and easy‐to‐use devices for short‐term monitoring or single‐shot measurements—have recently gained growing importance. Using this class of sensors, critical analytical information can be obtained by anyone, anywhere and at any time, without the need for recalibration and worrying about contamination.
Classification of measurement errors[edit]
An infrared sensor
A good sensor obeys the following rules:
it is sensitive to the measured property
it is insensitive to any other property likely to be encountered in its application, and
it does not influence the measured property.
Most sensors have a linear transfer function. The sensitivity is then defined as the ratio between the output signal and measured property. For example, if a sensor measures temperature and has a voltage output, the sensitivity is constant with the units [V/K]. The sensitivity is the slope of the transfer function. Converting the sensor's electrical output (for example V) to the measured units (for example K) requires dividing the electrical output by the slope (or multiplying by its reciprocal). In addition, an offset is frequently added or subtracted. For example, −40 must be added to the output if 0 V output corresponds to −40 C input.
For an analog sensor signal to be processed or used in digital equipment, it needs to be converted to a digital signal, using an analog-to-digital converter.
Sensor deviations[edit]
Since sensors cannot replicate an ideal transfer function, several types of deviations can occur which limit sensor accuracy:
Since the range of the output signal is always limited, the output signal will eventually reach a minimum or maximum when the measured property exceeds the limits. The full scale range defines the maximum and minimum values of the measured property.
The sensitivity may in practice differ from the value specified. This is called a sensitivity error. This is an error in the slope of a linear transfer function.
If the output signal differs from the correct value by a constant, the sensor has an offset error or bias. This is an error in the y-intercept of a linear transfer function.
Nonlinearity is deviation of a sensor's transfer function from a straight line transfer function. Usually, this is defined by the amount the output differs from ideal behavior over the full range of the sensor, often noted as a percentage of the full range.
Deviation caused by rapid changes of the measured property over time is a dynamic error. Often, this behavior is described with a bode plot showing sensitivity error and phase shift as a function of the frequency of a periodic input signal.
If the output signal slowly changes independent of the measured property, this is defined as drift. Long term drift over months or years is caused by physical changes in the sensor.
Noise is a random deviation of the signal that varies in time.
A hysteresis error causes the output value to vary depending on the previous input values. If a sensor's output is different depending on whether a specific input value was reached by increasing vs. decreasing the input, then the sensor has a hysteresis error.
If the sensor has a digital output, the output is essentially an approximation of the measured property. This error is also called quantization error.
If the signal is monitored digitally, the sampling frequency can cause a dynamic error, or if the input variable or added noise changes periodically at a frequency near a multiple of the sampling rate, aliasing errors may occur.
The sensor may to some extent be sensitive to properties other than the property being measured. For example, most sensors are influenced by the temperature of their environment.
All these deviations can be classified as systematic errors or random errors. Systematic errors can sometimes be compensated for by means of some kind of calibration strategy. Noise is a random error that can be reduced by signal processing, such as filtering, usually at the expense of the dynamic behavior of the sensor.
Resolution[edit]
The sensor resolution or measurement resolution is the smallest change that can be detected in the quantity that is being measured. The resolution of a sensor with a digital output is usually the numerical resolution of the digital output. The resolution is related to the precision with which the measurement is made, but they are not the same thing. A sensor's accuracy may be considerably worse than its resolution.
For example, the distance resolution is the minimum distance that can be accurately measured by any distance measuring devices. In a time-of-flight camera, the distance resolution is usually equal to the standard deviation (total noise) of the signal expressed in unit of length.
The sensor may to some extent be sensitive to properties other than the property being measured. For example, most sensors are influenced by the temperature of their environment.
Chemical sensor[edit]
A chemical sensor is a self-contained analytical device that can provide information about the chemical composition of its environment, that is, a liquid or a gas phase. The information is provided in the form of a measurable physical signal that is correlated with the concentration of a certain chemical species (termed as analyte). Two main steps are involved in the functioning of a chemical sensor, namely, recognition and transduction. In the recognition step, analyte molecules interact selectively with receptor molecules or sites included in the structure of the recognition element of the sensor. Consequently, a characteristic physical parameter varies and this variation is reported by means of an integrated transducer that generates the output signal.
A chemical sensor based on recognition material of biological nature is a biosensor. However, as synthetic biomimetic materials are going to substitute to some extent recognition biomaterials, a sharp distinction between a biosensor and a standard chemical sensor is superfluous. Typical biomimetic materials used in sensor development are molecularly imprinted polymers and aptamers.
Chemical sensor array[edit]
This section is an excerpt from Chemical sensor array.[edit]
A chemical sensor array is a sensor architecture with multiple sensor components that create a pattern for analyte detection from the additive responses of individual sensor components. There exist several types of chemical sensor arrays including electronic, optical, acoustic wave, and potentiometric devices. These chemical sensor arrays can employ multiple sensor types that are cross-reactive or tuned to sense specific analytes.
Biosensor[edit]
Main article: Biosensor
In biomedicine and biotechnology, sensors which detect analytes thanks to a biological component, such as cells, protein, nucleic acid or biomimetic polymers, are called biosensors.
Whereas a non-biological sensor, even organic (carbon chemistry), for biological analytes is referred to as sensor or nanosensor. This terminology applies for both in-vitro and in vivo applications.
The encapsulation of the biological component in biosensors, presents a slightly different problem that ordinary sensors; this can either be done by means of a semipermeable barrier, such as a dialysis membrane or a hydrogel, or a 3D polymer matrix, which either physically constrains the sensing macromolecule or chemically constrains the macromolecule by bounding it to the scaffold.
Neuromorphic sensors[edit]
Neuromorphic sensors are sensors that physically mimic structures and functions of biological neural entities. One example of this is the event camera.
MOS sensors[edit]
Metal–oxide–semiconductor (MOS) technology originates from the MOSFET (MOS field-effect transistor, or MOS transistor) invented by Mohamed M. Atalla and Dawon Kahng in 1959, and demonstrated in 1960. MOSFET sensors (MOS sensors) were later developed, and they have since been widely used to measure physical, chemical, biological and environmental parameters.
Biochemical sensors[edit]
A number of MOSFET sensors have been developed, for measuring physical, chemical, biological, and environmental parameters. The earliest MOSFET sensors include the open-gate field-effect transistor (OGFET) introduced by Johannessen in 1970, the ion-sensitive field-effect transistor (ISFET) invented by Piet Bergveld in 1970, the adsorption FET (ADFET) patented by P.F. Cox in 1974, and a hydrogen-sensitive MOSFET demonstrated by I. Lundstrom, M.S. Shivaraman, C.S. Svenson and L. Lundkvist in 1975. The ISFET is a special type of MOSFET with a gate at a certain distance, and where the metal gate is replaced by an ion-sensitive membrane, electrolyte solution and reference electrode. The ISFET is widely used in biomedical applications, such as the detection of DNA hybridization, biomarker detection from blood, antibody detection, glucose measurement, pH sensing, and genetic technology.
By the mid-1980s, numerous other MOSFET sensors had been developed, including the gas sensor FET (GASFET), surface accessible FET (SAFET), charge flow transistor (CFT), pressure sensor FET (PRESSFET), chemical field-effect transistor (ChemFET), reference ISFET (REFET), biosensor FET (BioFET), enzyme-modified FET (ENFET) and immunologically modified FET (IMFET). By the early 2000s, BioFET types such as the DNA field-effect transistor (DNAFET), gene-modified FET (GenFET) and cell-potential BioFET (CPFET) had been developed.
Image sensors[edit]
Main articles: Image sensor, Charge-coupled device, and Active-pixel sensor
MOS technology is the basis for modern image sensors, including the charge-coupled device (CCD) and the CMOS active-pixel sensor (CMOS sensor), used in digital imaging and digital cameras. Willard Boyle and George E. Smith developed the CCD in 1969. While researching the MOS process, they realized that an electric charge was the analogy of the magnetic bubble and that it could be stored on a tiny MOS capacitor. As it was fairly straightforward to fabricate a series of MOS capacitors in a row, they connected a suitable voltage to them so that the charge could be stepped along from one to the next. The CCD is a semiconductor circuit that was later used in the first digital video cameras for television broadcasting.
The MOS active-pixel sensor (APS) was developed by Tsutomu Nakamura at Olympus in 1985. The CMOS active-pixel sensor was later developed by Eric Fossum and his team in the early 1990s.
MOS image sensors are widely used in optical mouse technology. The first optical mouse, invented by Richard F. Lyon at Xerox in 1980, used a 5 µm NMOS sensor chip. Since the first commercial optical mouse, the IntelliMouse introduced in 1999, most optical mouse devices use CMOS sensors.
Monitoring sensors[edit]
A LIDAR sensor (bottom, center), as part of the camera system on an iPad Pro.
MOS monitoring sensors are used for house monitoring, office and agriculture monitoring, traffic monitoring (including car speed, traffic jams, and traffic accidents), weather monitoring (such as for rain, wind, lightning and storms), defense monitoring, and monitoring temperature, humidity, air pollution, fire, health, security and lighting. MOS gas detector sensors are used to detect carbon monoxide, sulfur dioxide, hydrogen sulfide, ammonia, and other gas substances. Other MOS sensors include intelligent sensors and wireless sensor network (WSN) technology.
See also[edit]
Actuator
Data acquisition
Data logger
Image sensor
MOSFET
BioFET
Chemical field-effect transistor
ISFET
List of sensors
Machine olfaction
Nanoelectronics
Nanosensor
Sensing floor
Transducer
Wireless sensor network | biology | 1024194 | https://no.wikipedia.org/wiki/Sensor%20%28instrument%29 | Sensor (instrument) | En sensor er et instrument som registrerer en viss påvirkning, f.eks. varme eller kulde, og konverterer registreringen til et signal som kan leses av en observatør eller et annet instrument. Bevegelsedetektor er et eksempel på en sensor som registrerer bevegelse og kan fra det trigge en kontakt eller relé, og dermed utløse en alarm eller et elektronisk varsel.
Dersom en sensor kobles til, eller har en indikasjon på målt verdi, kan den også kalles et måleinstrument.
For at man skal kunne stole på den målte verdien fra en sensor må den kalibreres. Kalibrering er å sammenligne den målte verdien fra sensoren med en måleverdi med kjent usikkerhet, som regel fra et annen sensor med kjent nøyaktighet.
Overføringen av det konverterte signalet er ofte elektrisk som 4-20mA 0-10V og digital, men kan også av være pneumatisk, hydraulisk, mekanisk eller annen art.
Eksempel på sensorer
Temperatursensor - måler temperatur
Trykksensor - måler trykk, finnes blant annet for luft, olje, vann.
Lastcelle - måler vekt.
Fuktighetsensor - måler relativ luftfuktighet.
Avstandssensor - måler avstand til et objekt
Luxmeter - måler lysmengde, finnes også som fotocelle
Bevegelsesensor - detekterer bevegelse. Kalles også nærverssensor
Desibelmåler - måler lydtrykk
CO2-måler - måler innhold av CO2 i luft
Flowsensor - måler massestrømning
Vindmåler - måler vindhastighet
Virtuelle sensorer
Virtuelle sensorer er teknologi der det måles en verdi fra en sensor som ikke fysisk eksisterer. De nødvendige målingene fra prosessen måles og overføres til en matematisk modell som ved hjelp av beregninger kalkulerer ønsket verdi. Grunnen til å bruke en virtuell sensor kan være at en slik sensor ikke finnes eller kan lages, eller at plassen der en ønsker å måle er utilgjengelig eller vanskelig å nå. Eksempel på virtuelle sensorer er duggpunktsensor eller en trykksensor inne i en roterende turbin. Virtuelle sensorer er avhengige av at den matematiske modellen er riktig og får rette data, men krever ikke vedlikehold, kalibrering eller service.
Se også
Transduser
Sensorer
Elektriske komponenter | norwegian_bokmål | 0.404755 |
temperature_sensed/Nerve.txt | A nerve is an enclosed, cable-like bundle of nerve fibers (called axons) in the peripheral nervous system.
Nerves have historically been considered the basic units of the peripheral nervous system. A nerve provides a common pathway for the electrochemical nerve impulses called action potentials that are transmitted along each of the axons to peripheral organs or, in the case of sensory nerves, from the periphery back to the central nervous system. Each axon, within the nerve, is an extension of an individual neuron, along with other supportive cells such as some Schwann cells that coat the axons in myelin.
Within a nerve, each axon is surrounded by a layer of connective tissue called the endoneurium. The axons are bundled together into groups called fascicles, and each fascicle is wrapped in a layer of connective tissue called the perineurium. Finally, the entire nerve is wrapped in a layer of connective tissue called the epineurium. Nerve cells (often called neurons) are further classified as sensory, motor, or mixed nerves.
In the central nervous system, the analogous structures are known as nerve tracts.
Structure[edit]
Cross-section of a nerve
Each nerve is covered on the outside by a dense sheath of connective tissue, the epineurium. Beneath this is a layer of fat cells, the perineurium, which forms a complete sleeve around a bundle of axons. Perineurial septae extend into the nerve and subdivide it into several bundles of fibres. Surrounding each such fibre is the endoneurium. This forms an unbroken tube from the surface of the spinal cord to the level where the axon synapses with its muscle fibres, or ends in sensory receptors. The endoneurium consists of an inner sleeve of material called the glycocalyx and an outer, delicate, meshwork of collagen fibres. Nerves are bundled and often travel along with blood vessels, since the neurons of a nerve have fairly high energy requirements.
Within the endoneurium, the individual nerve fibres are surrounded by a low-protein liquid called endoneurial fluid. This acts in a similar way to the cerebrospinal fluid in the central nervous system and constitutes a blood-nerve barrier similar to the blood–brain barrier. Molecules are thereby prevented from crossing the blood into the endoneurial fluid. During the development of nerve edema from nerve irritation (or injury), the amount of endoneurial fluid may increase at the site of irritation. This increase in fluid can be visualized using magnetic resonance neurography, and thus MR neurography can identify nerve irritation and/or injury.
Categories[edit]
Nerves are categorized into three groups based on the direction that signals are conducted:
Afferent nerves conduct signals from sensory neurons to the central nervous system, for example from the mechanoreceptors in skin.
Efferent nerves conduct signals from the central nervous system along motor neurons to their target muscles and glands.
Mixed nerves contain both afferent and efferent axons, and thus conduct both incoming sensory information and outgoing muscle commands in the same bundle. All spinal nerves are mixed nerves, and some of the cranial nerves are also mixed nerves.
Nerves can be categorized into two groups based on where they connect to the central nervous system:
Spinal nerves innervate (distribute to/stimulate) much of the body, and connect through the vertebral column to the spinal cord and thus to the central nervous system. They are given letter-number designations according to the vertebra through which they connect to the spinal column.
Cranial nerves innervate parts of the head, and connect directly to the brain (especially to the brainstem). They are typically assigned Roman numerals from 1 to 12, although cranial nerve zero is sometimes included. In addition, cranial nerves have descriptive names.
Terminology[edit]
Main article: Anatomical terms of neuroanatomy
Specific terms are used to describe nerves and their actions. A nerve that supplies information to the brain from an area of the body, or controls an action of the body is said to innervate that section of the body or organ. Other terms relate to whether the nerve affects the same side ("ipsilateral") or opposite side ("contralateral") of the body, to the part of the brain that supplies it.
Development[edit]
Nerve growth normally ends in adolescence, but can be re-stimulated with a molecular mechanism known as "Notch signaling".
Regeneration[edit]
If the axons of a neuron are damaged, as long as the cell body of the neuron is not damaged, the axons can regenerate and remake the synaptic connections with neurons with the help of guidepost cells. This is also referred to as neuroregeneration.
The nerve begins the process by destroying the nerve distal to the site of injury allowing Schwann cells, basal lamina, and the neurilemma near the injury to begin producing a regeneration tube. Nerve growth factors are produced causing many nerve sprouts to bud. When one of the growth processes finds the regeneration tube, it begins to grow rapidly towards its original destination guided the entire time by the regeneration tube. Nerve regeneration is very slow and can take up to several months to complete. While this process does repair some nerves, there will still be some functional deficit as the repairs are not perfect.
Function[edit]
A nerve conveys information in the form of electrochemical impulses (as nerve impulses known as action potentials) carried by the individual neurons that make up the nerve. These impulses are extremely fast, with some myelinated neurons conducting at speeds up to 120 m/s. The impulses travel from one neuron to another by crossing a synapse, where the message is converted from electrical to chemical and then back to electrical.
Nerves can be categorized into two groups based on function:
An afferent nerve fiber conducts sensory information from a sensory neuron to the central nervous system, where the information is then processed. Bundles of fibres or axons, in the peripheral nervous system are called nerves, and bundles of afferent fibers are known as sensory nerves.
An efferent nerve fiber conducts signals from a motor neuron in the central nervous system to muscles. Bundles of these fibres are known as efferent nerves.
Nervous system[edit]
Main article: Nervous system
The nervous system is the part of an animal that coordinates its actions by transmitting signals to and from different parts of its body. In vertebrates it consists of two main parts, the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain, brainstem and spinal cord. The PNS consists mainly of nerves, which are enclosed bundles of the long fibers or axons, that connect the CNS to all remaining body parts.
Nerves that transmit signals from the CNS are called motor or efferent nerves, while those nerves that transmit information from the body to the CNS are called sensory or afferent. Spinal nerves serve both functions and are called mixed nerves. The PNS is divided into three separate subsystems, the somatic, autonomic, and enteric nervous systems. Somatic nerves mediate voluntary movement.
The autonomic nervous system is further subdivided into the sympathetic and the parasympathetic nervous systems. The sympathetic nervous system is activated in cases of emergencies to mobilize energy, while the parasympathetic nervous system is activated when organisms are in a relaxed state. The enteric nervous system functions to control the gastrointestinal system. Both autonomic and enteric nervous systems function involuntarily. Nerves that exit from the cranium are called cranial nerves while those exiting from the spinal cord are called spinal nerves.
Clinical significance[edit]
Micrograph demonstrating perineural invasion of prostate cancer. H&E stain.
Cancer can spread by invading the spaces around nerves. This is particularly common in head and neck cancer, prostate cancer and colorectal cancer.
Nerves can be damaged by physical injury as well as conditions like carpal tunnel syndrome (CTS) and repetitive strain injury. Autoimmune diseases such as Guillain–Barré syndrome, neurodegenerative diseases, polyneuropathy, infection, neuritis, diabetes, or failure of the blood vessels surrounding the nerve all cause nerve damage, which can vary in severity.
Multiple sclerosis is a disease associated with extensive nerve damage. It occurs when the macrophages of an individual's own immune system damage the myelin sheaths that insulate the axon of the nerve.
A pinched nerve occurs when pressure is placed on a nerve, usually from swelling due to an injury, or pregnancy and can result in pain, weakness, numbness or paralysis, an example being CTS. Symptoms can be felt in areas far from the actual site of damage, a phenomenon called referred pain. Referred pain can happen when the damage causes altered signalling to other areas.
Neurologists usually diagnose disorders of nerves by a physical examination, including the testing of reflexes, walking and other directed movements, muscle weakness, proprioception, and the sense of touch. This initial exam can be followed with tests such as nerve conduction study, electromyography (EMG), and computed tomography (CT).
Other animals[edit]
A neuron is called identified if it has properties that distinguish it from every other neuron in the same animal—properties such as location, neurotransmitter, gene expression pattern, and connectivity—and if every individual organism belonging to the same species has exactly one neuron with the same set of properties. In vertebrate nervous systems, very few neurons are "identified" in this sense. Researchers believe humans have none—but in simpler nervous systems, some or all neurons may be thus unique.
In vertebrates, the best known identified neurons are the gigantic Mauthner cells of fish. Every fish has two Mauthner cells, located in the bottom part of the brainstem, one on the left side and one on the right. Each Mauthner cell has an axon that crosses over, innervating (stimulating) neurons at the same brain level and then travelling down through the spinal cord, making numerous connections as it goes. The synapses generated by a Mauthner cell are so powerful that a single action potential gives rise to a major behavioral response: within milliseconds the fish curves its body into a C-shape, then straightens, thereby propelling itself rapidly forward. Functionally this is a fast escape response, triggered most easily by a strong sound wave or pressure wave impinging on the lateral line organ of the fish. Mauthner cells are not the only identified neurons in fish—there are about 20 more types, including pairs of "Mauthner cell analogs" in each spinal segmental nucleus. Although a Mauthner cell is capable of bringing about an escape response all by itself, in the context of ordinary behavior other types of cells usually contribute to shaping the amplitude and direction of the response.
Mauthner cells have been described as command neurons. A command neuron is a special type of identified neuron, defined as a neuron that is capable of driving a specific behavior all by itself. Such neurons appear most commonly in the fast escape systems of various species—the squid giant axon and squid giant synapse, used for pioneering experiments in neurophysiology because of their enormous size, both participate in the fast escape circuit of the squid. The concept of a command neuron has, however, become controversial, because of studies showing that some neurons that initially appeared to fit the description were really only capable of evoking a response in a limited set of circumstances.
In organisms of radial symmetry, nerve nets serve for the nervous system. There is no brain or centralised head region, and instead there are interconnected neurons spread out in nerve nets. These are found in Cnidaria, Ctenophora and Echinodermata.
History[edit]
Further information: History of neurology and neurosurgery
Herophilos (335–280 BC) described the functions of the optic nerve in sight and the oculomotor nerve in eye movement. Analysis of the nerves in the cranium enabled him to differentiate between blood vessels and nerves (Ancient Greek: νεῦρον (neûron) "string, plant fiber, nerve").
Modern research has not confirmed William Cullen's 1785 hypothesis associating mental states with physical nerves, although popular or lay medicine may still invoke "nerves" in diagnosing or blaming any sort of psychological worry or hesitancy, as in the common traditional phrases "my poor nerves",
"high-strung", and "nervous breakdown".
See also[edit]
Biology portal
Connective tissue in the peripheral nervous system
Dermatome (anatomy)
List of nerves of the human body
Nerve injury
Neuropathy
Nerve injury classification | biology | 322049 | https://da.wikipedia.org/wiki/Neuron | Neuron | Et neuron (fra græsk: sene, nerve), også kaldet en nervecelle, er en celletype i nervesystemet.
De adskiller sig fra andre celler ved deres mange udløbere, kaldet dendritter og aksoner, og ved at være specialiseret i transmittering af signaler, dels elektrisk intraneuronalt og dels ved hjælp af transmitterstoffer interneuronalt, der virker på andre neuroners eller effektorvævenes receptorer (se for eksempel G-protein-koblede receptorer, GPCR og den motoriske endeplade). Af neurotransmittere kan nævnes serotonin, acetylkolin, dopamin, adrenalin og noradrenalin.
Formålet med hjernens netværk af neuroner er kommunikation og informationsbehandling.
Opbygning
Neuroner er de centrale komponenter i hjernen og rygmarven i centralnervesystemet (CNS) samt ganglier i det perifere nervesystem (PNS). En typisk neuron består af et cellelegeme (soma), dendritter og et axon. Dendritter er tynde strukturer, der udspringer fra cellekroppen og ofte strækker sig flere hundrede mikrometer og forgrener sig flere gange. En Axon, også kaldet en nervefiber når de er myelinerede, udspringer fra cellen ved et sted kaldet axon højen og rejser op til en en meter i mennesker og endnu længere i andre arter. En neurons cellekroppe har ofte flere dendritter, men aldrig til mere end ét Axon. Neuronets lange aksoner kan være omgivet af en isolerende fedtskede kaldet myelinskeden, som har til funktion at øge signaleringshastigheden.
Myelinskeden produceres og vedligeholdes i centralnervesystemet af oligodendrocytter og af Schwannske celler i det perifere nervesystem.
I de fleste tilfælde er neuroner genereret af særlige typer af stamceller. Det menes generelt, at neuroner ikke undergår celledeling, men nyere forskning hos hunde viser, at det i nogle tilfælde sker i nethinden. Astrocytter er stjerneformede gliaceller, der er også blevet observeret at blive til neuroner. Hos mennesker ophører neurogenese stort set i voksenalderen, men i to områder i hjernen, hippocampus og den olfaktoriske pære, er der beviser for generering af nye neuroner.
Kommunikation mellem neuroner
En neuron behandler og sender oplysninger gennem elektriske og kemiske signaler. Nervecellernes signaler bygger på en spændingsforskel mellem nervecellens indre og ydre, kaldet membranpotentialet. Er neuronet i hvile, kaldes spændingsforskellen for hvilemembranpotentialet. Er ændringen i membranpotentialet tilstrækkelig stor, genereres en (alt-eller-intet) elektrokemisk puls kaldet et aktionspotentiale, som bevæger sig hurtigt langs cellens axon og aktiverer synaptiske forbindelser til andre celler.
Neurontyper
Neuroner kan overordnet inddeles i projektionsneuroner Golgi type 1 med et enkelt langt akson, der kan kommunikere med andre regioner af nervesystemet, og interneuroner Golgi type 2 som kun befinder sig i en hjerneregion. Der er dog ikke skarp adskillelse mellem disse to typer.
Specialiserede neuroner omfatter: 1) sensoriske neuroner, der reagerer på berøring, lyd, lys og alle andre stimuli, der påvirker cellerne via vores sanser, som derefter sender signaler til rygmarven og hjernen; 2) motoriske neuroner, der modtager signaler fra hjernen og rygmarven, som bevirker muskelsammentrækninger og påvirker glandulare udgange; 3) interneuroner, der forbinder neuroner i den samme region i hjernen eller rygmarven.
Se også
Dendrit, akson, soma, gliacelle
Kranienerver
Neuropeptid
Eksterne henvisninger | danish | 0.434241 |
temperature_sensed/Room_temperature.txt | Colloquially, room temperature is a range of air temperatures that most people prefer for indoor settings. These temperatures feel comfortable to people wearing typical indoor clothing. But comfortable temperatures can be extended beyond this range depending on humidity, air circulation, and other factors.
In certain fields, like science and engineering, and within a particular context, room temperature can mean different agreed-upon ranges. In contrast, ambient temperature is the actual temperature, as measured by a thermometer, of the air (or other medium and surroundings) in any particular place. The ambient temperature (e.g. an unheated room in winter) may be very different from an ideal room temperature.
Food or beverages may be served at "room temperature", meaning neither heated nor cooled.
Comfort temperatures[edit]
Main article: Thermal comfort
The American Heritage Dictionary of the English Language identifies room temperature as around 20–22 °C (68–72 °F), while the Oxford English Dictionary states that it is "conventionally taken as about 20 °C (68 °F)".
Ideal room temperature varies vastly depending on the surrounding climate. Studies from Indonesia have shown that the range of comfortable temperature falls between 24–29 °C (75–84 °F) for local residents. Studies from Nigeria show a comfortable temperature range of 26–28 °C (79–82 °F), comfortably cool 24–26 °C (75–79 °F) and comfortably warm 28–30 °C (82–86 °F). A field study conducted in Hyderabad, India returned a comfort band of 26–32.45 °C (79–90 °F) with a mean of 29.23 °C (85 °F). A study conducted in Jaipur, India among healthy young men showed that the neutral thermal comfort temperature was analyzed to be 30.15 °C (86 °F), although a range of 25.9–33.8 °C (79–93 °F) was found.
Owing to variations in humidity and (likely) clothing, recommendations for summer and winter may vary; a suggested typical range for summer is 23–25.5 °C (73–78 °F), with that for winter being 20–23.5 °C (68–74 °F). Some studies have suggested that thermal comfort preferences of men and women may differ significantly, with women on average preferring higher ambient temperatures.
In the recent past it was common for house temperatures to be kept below the comfort level; a 1978 UK study found average indoor home temperatures to be 15.8 °C (60.4 °F) while Japan in 1980 had median home temperatures of 13 °C (55 °F) to 15 °C (59 °F).
Rooms may be maintained at an ambient temperature above the comfort temperature in hot weather, or below it in cold weather, if required by cost considerations or practical issues (e.g. lack of air conditioning or relatively high expense of heating.)
In the UK, the Offices, Shops and Railway Premises Act 1963 provides for a minimum temperature in commercial premises, but not for a maximum temperature.
Health effects[edit]
See also: Sick building syndrome
A digital thermometer reading an ambient temperature of 36.4°C (97°F) in an unventilated room during a heat wave; a high indoor temperature can cause heat exhaustion or heat stroke in a person.
The World Health Organization in 1987 found that comfortable indoor temperatures of between 18 and 24 °C (64 and 75 °F) were not associated with health risks for healthy adults with appropriate clothing, humidity, and other factors. For infants, elderly, and those with significant health problems, a minimum 20 °C (68 °F) was recommended. Temperatures lower than 16 °C (61 °F) with humidity above 65% were associated with respiratory hazards including allergies.
The WHO's 2018 guidelines give a strong recommendation that a minimum of 18 °C (64 °F) is a "safe and well-balanced indoor temperature to protect the health of general populations during cold seasons". A higher minimum temperature may be necessary for vulnerable groups including children, the elderly, and people with cardiorespiratory disease and other chronic illnesses. However, the recommendation regarding risk of exposure to high indoor temperatures is only "conditional". Minimal-risk high temperatures range from about 21 to 30 °C (70 to 86 °F) depending on the region, with maximum acceptable temperatures between 25 and 32 °C (77 and 90 °F).
Definitions in science and industry[edit]
Temperature ranges are defined as room temperature for certain products and processes in industry, science, standards, and consumer goods. For instance, for the shipping and storage of pharmaceuticals, the United States Pharmacopeia-National Formulary (USP-NF) defines controlled room temperature as between 20 and 25 °C (68 and 77 °F), with excursions between 15 and 30 °C (59 and 86 °F) allowed, provided the mean kinetic temperature does not exceed 25 °C (77 °F). The European Pharmacopoeia defines it as being simply 15 to 25 °C (59 to 77 °F), and the Japanese Pharmacopeia defines "ordinary temperature" as 15 to 25 °C (59 to 77 °F), with room temperature being 1 to 30 °C (34 to 86 °F). Merriam-Webster gives as a medical definition a range of 15 to 25 °C (59 to 77 °F) as being suitable for human occupancy, and at which laboratory experiments are usually performed.
See also[edit]
Standard conditions for temperature and pressure
ISO 1 – ISO standard temperature, 20°C
Indoor air quality | biology | 15016 | https://sv.wikipedia.org/wiki/Temperatur | Temperatur | Temperatur är en fysikalisk storhet och ett mått på värmetillstånd. Värmeflödet är från en högre temperatur till en lägre temperatur. Vid lika temperatur är föremål i termisk jämvikt, se termodynamikens nollte huvudsats. Vidare kan också olika färgtoner av ljus mätas i så kallad färgtemperatur.
Definition
Temperatur definieras som förändringen i inre energi av entropin
där T är temperaturen i kelvin (K). Partiella derivatan, med volymen V och partikelantalet N fasta, är ofta en monotont växande funktion, vilket innebär att temperaturen alltid är positiv. Ett system har alltså låg temperatur om entropin ändras mycket då det tar upp värme. Värme flödar från ett system med hög temperatur till ett system med låg temperatur till dess att systemet som avger värme inte längre förlorar mer entropi på värmeförflyttningen än systemet som tar emot värme vinner. Då värmeflödet upphör har systemen tillsammans uppnått sin maximala entropi, temperaturen är lika och systemen befinner sig i termisk jämvikt med varandra.
Temperatur kan också beskrivas som den kinetiska energin i en ideal gas, det vill säga rörelsen hos molekylerna/atomerna inom ämnet. Vid högre temperatur rör de sig mer och vid lägre temperatur mindre.
Temperaturskalor
Kelvinskalan – i vetenskapliga sammanhang används idag den absoluta skalan. Den har sin nollpunkt vid den temperatur som ett medium har då det är i sitt grundtillstånd (absoluta nollpunkten) och temperaturen uttrycks i enheten kelvin. Notera att kelvin inte inleds med en versal eftersom det tillhör SI-enheterna. Observera också att "grad Kelvin" (°K) har ersatts med kelvin (K).
Före den moderna uppfattningen om temperatur användes skalor med nollpunkten satt vid andra temperaturer än den absoluta nollpunkten. Av dem används dessa fortfarande i vardagliga sammanhang:
Celsiusskalan – den vanligaste temperaturskalan i Sverige och i många andra europeiska länder. Uppfunnen av svensken Anders Celsius. Temperaturer anges på skalan med enheten grad Celsius (°C), vars steglängd är lika med 1 K. Vatten fryser vid 0 °C och kokar vid 100 °C.
Fahrenheitskalan – används mest i länder utanför Europa, till exempel USA och Jamaica. Temperaturer anges på skalan med enheten grad Fahrenheit (°F). Vatten fryser vid 32 °F och kokar vid 212 °F.
Följande formler kan användas för att konvertera mellan temperaturerna T K, tC °C och tF °F:
T = tC + 273,15
tC = 5/9 · (tF − 32)
Äldre temperaturskalor som inte används längre
Rankine – °R eller °Raa, som har samma steglängd som °F. Utgår från absoluta nollpunkten. Vatten fryser vid 459.67°R och kokar vid 671.64°R
Réaumur – °Ré, °Re eller °R. Steglängden är lika med 1,25 grader Celsius. Vatten fryser vid 0°Ré och kokar vid 80°Ré.
Rømer – °Rø eller °R. Vatten fryser vid 7,05°Rø och kokar vid 60°Rø.
Delisle – °De (stavas ibland de Lisle). Vatten fryser vid 150°De och kokar vid 0°De. Temperatur över kokpunkten räknas som minusgrader.
Omvandlingstabell
Se även
Dolbears lag
Lord Kelvin
Negativ temperatur
Operativ temperatur
Rumstemperatur
Termometer
Väderrekord
Referenser
Externa länkar
Termodynamik
Fysikaliska storheter
Wikipedia:Basartiklar | swedish | 0.40785 |
temperature_sensed/Transient_receptor_potential_channel.txt | Transient receptor potential channels (TRP channels) are a group of ion channels located mostly on the plasma membrane of numerous animal cell types. Most of these are grouped into two broad groups: Group 1 includes TRPC ( "C" for canonical), TRPV ("V" for vanilloid), TRPVL ("VL" for vanilloid-like), TRPM ("M" for melastatin), TRPS ("S" for soromelastatin), TRPN ("N" for mechanoreceptor potential C), and TRPA ("A" for ankyrin). Group 2 consists of TRPP ("P" for polycystic) and TRPML ("ML" for mucolipin). Other less-well categorized TRP channels exist, including yeast channels and a number of Group 1 and Group 2 channels present in non-animals. Many of these channels mediate a variety of sensations such as pain, temperature, different kinds of taste, pressure, and vision. In the body, some TRP channels are thought to behave like microscopic thermometers and used in animals to sense hot or cold. Some TRP channels are activated by molecules found in spices like garlic (allicin), chili pepper (capsaicin), wasabi (allyl isothiocyanate); others are activated by menthol, camphor, peppermint, and cooling agents; yet others are activated by molecules found in cannabis (i.e., THC, CBD and CBN) or stevia. Some act as sensors of osmotic pressure, volume, stretch, and vibration. Most of the channels are activated or inhibited by signaling lipids and contribute to a family of lipid-gated ion channels.
These ion channels have a relatively non-selective permeability to cations, including sodium, calcium and magnesium.
TRP channels were initially discovered in the so-called "transient receptor potential" mutant (trp-mutant) strain of the fruit fly Drosophila, hence their name (see History of Drosophila TRP channels below). Later, TRP channels were found in vertebrates where they are ubiquitously expressed in many cell types and tissues. Most TRP channels are composed of 6 membrane-spanning helices with intracellular N- and C-termini. Mammalian TRP channels are activated and regulated by a wide variety of stimuli and are expressed throughout the body.
Families[edit]
TRP channel groups and families.
In the animal TRP superfamily there are currently 9 proposed families split into two groups, each family containing a number of subfamilies. Group one consists of TRPC, TRPV, TRPVL, TRPA, TRPM, TRPS, and TRPN, while group two contains TRPP and TRPML. There is an additional family labeled TRPY that is not always included in either of these groups. All of these sub-families are similar in that they are molecular sensing, non-selective cation channels that have six transmembrane segments, however, each sub-family is very unique and shares little structural homology with one another. This uniqueness gives rise to the various sensory perception and regulation functions that TRP channels have throughout the body. Group one and group two vary in that both TRPP and TRPML of group two have a much longer extracellular loop between the S1 and S2 transmembrane segments. Another differentiating characteristic is that all the group one sub-families either contain an N-terminal intracellular ankyrin repeat sequence, a C-terminal TRP domain sequence, or both—whereas both group two sub-families have neither. Below are members of the sub-families and a brief description of each:
TRPA[edit]
Family
Sub-Family
Known Taxa
TRPA
TRPA1
Vertebrates, arthropods, and molluscs
TRPA-like
Choanoflagellates, cnidarians, nematodes, arthropods (only crustaceans and myriapods), molluscs, and echinoderms
TRPA5
Arthropods (only crustaceans and insects)
painless
pyrexia
waterwitch
HsTRPA
Specific to hymenopteran insects
TRPA, A for "ankyrin", is named for the large amount of ankyrin repeats found near the N-terminus. TRPA is primarily found in afferent nociceptive nerve fibers and is associated with the amplification of pain signaling as well as cold pain hypersensitivity. These channels have been shown to be both mechanical receptors for pain and chemosensors activated by various chemical species, including isothiocyanates (pungent chemicals in substances such as mustard oil and wasabi), cannabinoids, general and local analgesics, and cinnamaldehyde.
While TRPA1 is expressed in a wide variety of animals, a variety of other TRPA channels exist outside of vertebrates. TRPA5, painless, pyrexia, and waterwitch are distinct phylogenetic branches within the TRPA clade, and are only evidenced to be expressed in crustaceans and insects, while HsTRPA arose as a Hymenoptera-specific duplication of waterwitch. Like TRPA1 and other TRP channels, these function as ion channels in a number of sensory systems. TRPA- or TRPA1-like channels also exists in a variety of species as a phylogenetically distinct clade, but these are less well understood.
TRPC[edit]
Family
Sub-Family
Known Taxa
TRPC
TRPC1
Vertebrates
TRPC2
TRPC3
TRPC4
TRPC5
TRPC6
TRPC7
TRP
Arthropods
TRPgamma
TRPL
Unknown
Choanoflagellates, cnidarians, xenacoelomorphs, lophotrochozoans, and nematodes
TRPC, C for "canonical", is named for being the most closely related to Drosophila TRP, the namesake of TRP channels. The phylogeny of TRPC channels has not been resolved in detail, but they are present across animal taxa. There are actually only six TRPC channels expressed in humans because TRPC2 is found to be expressed solely in mice and is considered a pseudo-gene in humans; this is partly due to the role of TRPC2 in detecting pheromones, which mice have an increased ability compared to humans. Mutations in TRPC channels have been associated with respiratory diseases along with focal segmental glomerulosclerosis in the kidneys. All TRPC channels are activated either by phospholipase C (PLC) or diacyglycerol (DAG).
TRPML[edit]
Family
Sub-Family
Known Taxa
TRPML
Unknown
Cnidarians, basal vertebrates, tunicates, cephalochordates, hemichordates, echinoderms, arthropods, and nematodes
TRPML1
Specific to jawed vertebrates
TRPML2
TRPML3
TRPML, ML for "mucolipin", gets its name from the neurodevelopmental disorder mucolipidosis IV. Mucolipidosis IV was first discovered in 1974 by E.R. Berman who noticed abnormalities in the eyes of an infant. These abnormalities soon became associated with mutations to the MCOLN1 gene which encodes for the TRPML1 ion channel. TRPML is still not highly characterized. The three known vertebrate copies are restricted to jawed vertebrates, with some exceptions (e.g. Xenopus tropicalis).
TRPM[edit]
Family
Sub-Family
Known Taxa
TRPM
Alpha/α (inc. TRPM1, 3, 6, and 7)
Choanoflagellates and animals (except tardigrades)
Beta/β (inc. TRPM2, 4, 5, and 8)
TRPM, M for "melastatin", was found during a comparative genetic analysis between benign nevi and malignant nevi (melanoma). Mutations within TRPM channels have been associated with hypomagnesemia with secondary hypocalcemia. TRPM channels have also become known for their cold-sensing mechanisms, such is the case with TRPM8. Comparative studies have shown that the functional domains and critical amino acids of TRPM channels are highly conserved across species.
Phylogenetics has shown that TRPM channels are split into two major clades, αTRPM and βTRPM. αTRPMs include vertebrate TRPM1, TRPM3, and the "chanzymes" TRPM6 and TRPM7, as well as the only insect TRPM channel, among others. βTRPMs include, but are not limited to, vertebrate TRPM2, TRPM4, TRPM5, and TRPM8 (the cold and menthol sensor). Two additional major clades have been described: TRPMc, which is present only in a variety of arthropods, and a basal clade, which has since been proposed to be a distinct and separate TRP channel family (TRPS).
TRPN[edit]
Family
Sub-Family
Known Taxa
TRPN
TRPN/nompC
Placozoans, cnidarians, nematodes, arthropods, molluscs, annelids, and vertebrates (excluding amniotes)
TRPN was originally described in Drosophila melanogaster and Caenorhabditis elegans as nompC, a mechanically gated ion channel. Only a single TRPN, N for "no mechanoreceptor potential C," or "nompC", is known to be broadly expressed in animals (although some Cnidarians have more), and is notably only a pseudogene in amniote vertebrates. Despite TRPA being named for ankyrin repeats, TRPN channels are thought to have the most of any TRP channel, typically around 28, which are highly conserved across taxa Since its discovery, Drosophila nompC has been implicated in mechanosensation (including mechanical stimulation of the cuticle and sound detection) and cold nociception.
TRPP[edit]
Family
Sub-Family
Known Taxa
TRPP
PKD1-like
Animals (excluding arthropods)
PKD2-like
Animals
Brividos
Insects
TRPP, P for "polycistin", is named for polycystic kidney disease, which is associated with these channels. These channels are also referred to as PKD (polycistic kidney disease) ion channels.
PKD2-like genes (examples include TRPP2, TRPP3, and TRPP5) encode canonical TRP channels. PKD1-like genes encode much larger proteins with 11 transmembrane segments, which do not have all the features of other TRP channels. However, 6 of the transmebrane segments of PKD1-like proteins have substantial sequence homology with TRP channels, indicating they may simply have diversified greatly from other closely related proteins.
Insects have a third sub-family of TRPP, called brividos, which participate in cold sensing.
TRPS[edit]
TRPS, S for Soromelastatin, was named as it forms a sister group to TRPM. TRPS is broadly present in animals, but notably absent in vertebrates and insects (among others). TRPS has not yet been well described functionally, though it is known that the C. elegans TRPS, known as CED-11, is a calcium channel which participates in apoptosis.
TRPV[edit]
Family
Sub-Family
Known Taxa
TRPV
Nanchung
Placozoans, cnidarians, nematodes, annelids, molluscs, and arthropods (possibly excluding arachnids)
Inactive
TRPV1
Specific to vertebrates
TRPV2
TRPV3
TRPV4
TRPV5
TRPV6
TRPV, V for "vanilloid", was originally discovered in Caenorhabditis elegans, and is named for the vanilloid chemicals that activate some of these channels. These channels have been made famous for their association with molecules such as capsaicin (a TRPV1 agonist). In addition to the 6 known vertebrate paralogues, 2 major clades are known outside of the deterostomes: nanchung and Iav. Mechanistic studies of these latter clades have been largely restricted to Drosophila, but phylogenetic analyses has placed a number of other genes from Placozoa, Annelida, Cnidaria, Mollusca, and other arthropods within them. TRPV channels have also been described in protists.
TRPVL[edit]
TRPVL has been proposed to be a sister clade to TRPV, and is limited to the cnidarians Nematostella vectensis and Hydra magnipapillata, and the annelid Capitella teleta. Little is known concerning these channels.
TRPY[edit]
TRPY, Y for "yeast", is highly localized to the yeast vacuole, which is the functional equivalent of a lysosome in a mammalian cell, and acts as a mechanosensor for vacuolar osmotic pressure. Patch clamp techniques and hyperosmotic stimulation have illustrated that TRPY plays a role in intracellular calcium release. Phylogenetic analysis has shown that TRPY1 does not form a part with the other metazoan TRP groups one and two, and is suggested to have evolved after the divergence of metazoans and fungi. Others have indicated that TRPY are more closely related to TRPP.
Structure[edit]
TRP channels are composed of 6 membrane-spanning helices (S1-S6) with intracellular N- and C-termini. Mammalian TRP channels are activated and regulated by a wide variety of stimuli including many post-transcriptional mechanisms like phosphorylation, G-protein receptor coupling, ligand-gating, and ubiquitination. The receptors are found in almost all cell types and are largely localized in cell and organelle membranes, modulating ion entry.
Most TRP channels form homo- or heterotetramers when completely functional. The ion selectivity filter, pore, is formed by the complex combination of p-loops in the tetrameric protein, which are situated in the extracellular domain between the S5 and S6 transmembrane segments. As with most cation channels, TRP channels have negatively charged residues within the pore to attract the positively charged ions.
Group 1 Characteristics[edit]
Each channel in this group is structurally unique, which adds to the diversity of functions that TRP channels possess, however, there are some commonalities that distinguish this group from others. Starting from the intracellular N-terminus there are varying lengths of ankryin repeats (except in TRPM) that aid with membrane anchoring and other protein interactions. Shortly following S6 on the C-terminal end, there is a highly conserved TRP domain (except in TRPA) which is involved with gating modulation and channel multimerization. Other C-terminal modifications such as alpha-kinase domains in TRPM7 and M8 have been seen as well in this group.
Group 2 Characteristics[edit]
Group two most distinguishable trait is the long extracellular span between the S1 and S2 transmembrane segments. Members of group two are also lacking in ankryin repeats and a TRP domain. They have been shown, however, to have endoplasmic reticulum (ER) retention sequences towards on the C-terminal end illustrating possible interactions with the ER.
Function[edit]
TRP channels modulate ion entry driving forces and Ca and Mg transport machinery in the plasma membrane, where most of them are located. TRPs have important interactions with other proteins and often form signaling complexes, the exact pathways of which are unknown. TRP channels were initially discovered in the trp mutant strain of the fruit fly Drosophila which displayed transient elevation of potential in response to light stimuli and were so named transient receptor potential channels. TRPML channels function as intracellular calcium release channels and thus serve an important role in organelle regulation. Importantly, many of these channels mediate a variety of sensations like the sensations of pain, temperature, different kinds of taste, pressure, and vision. In the body, some TRP channels are thought to behave like microscopic thermometers and are used in animals to sense hot or cold. TRPs act as sensors of osmotic pressure, volume, stretch, and vibration. TRPs have been seen to have complex multidimensional roles in sensory signaling. Many TRPs function as intracellular calcium release channels.
Pain and temperature sensation[edit]
TRP ion channels convert energy into action potentials in somatosensory nociceptors. Thermo-TRP channels have a C-terminal domain that is responsible for thermosensation and have a specific interchangeable region that allows them to sense temperature stimuli that is tied to ligand regulatory processes. Although most TRP channels are modulated by changes in temperature, some have a crucial role in temperature sensation. There are at least 6 different Thermo-TRP channels and each plays a different role. For instance, TRPM8 relates to mechanisms of sensing cold, TRPV1 and TRPM3 contribute to heat and inflammation sensations, and TRPA1 facilitates many signaling pathways like sensory transduction, nociception, inflammation and oxidative stress.
Taste[edit]
TRPM5 is involved in taste signaling of sweet, bitter and umami tastes by modulating the signal pathway in type II taste receptor cells. TRPM5 is activated by the sweet glycosides found in the stevia plant.
Several other TRP channels play a significant role in chemosensation through sensory nerve endings in the mouth that are independent from taste buds. TRPA1 responds to mustard oil (allyl isothiocyanate), wasabi, and cinnamon, TRPA1 and TRPV1 responds to garlic (allicin), TRPV1 responds to chilli pepper (capsaicin), TRPM8 is activated by menthol, camphor, peppermint, and cooling agents; TRPV2 is activated by molecules (THC, CBD and CBN) found in marijuana.
TRP-like channels in insect vision[edit]
Figure 1. Light-activated TRPL channels in Periplaneta americana photoreceptors. A, a typical current through TRPL channels was evoked by a 4-s pulse of bright light (horizontal bar). B, a photoreceptor membrane voltage response to the light-induced activation of TRPL channels, data from the same cell are shown
The trp-mutant fruit flies, which lack a functional copy of trp gene, are characterized by a transient response to light, unlike wild-type flies that demonstrate a sustained photoreceptor cell activity in response to light.
A distantly related isoform of TRP channel, TRP-like channel (TRPL), was later identified in Drosophila photoreceptors, where it is expressed at approximately 10- to 20-fold lower levels than TRP protein. A mutant fly, trpl, was subsequently isolated. Apart from structural differences, the TRP and TRPL channels differ in cation permeability and pharmacological properties.
TRP/TRPL channels are solely responsible for depolarization of insect photoreceptor plasma membrane in response to light. When these channels open, they allow sodium and calcium to enter the cell down the concentration gradient, which depolarizes the membrane. Variations in light intensity affect the total number of open TRP/TRPL channels, and, therefore, the degree of membrane depolarization. These graded voltage responses propagate to photoreceptor synapses with second-order retinal neurons and further to the brain.
It is important to note that the mechanism of insect photoreception is dramatically different from that in mammals. Excitation of rhodopsin in mammalian photoreceptors leads to the hyperpolarization of the receptor membrane but not to depolarization as in the insect eye. In Drosophila and, it is presumed, other insects, a phospholipase C (PLC)-mediated signaling cascade links photoexcitation of rhodopsin to the opening of the TRP/TRPL channels. Although numerous activators of these channels such as phosphatidylinositol-4,5-bisphosphate (PIP2) and polyunsaturated fatty acids (PUFAs) were known for years, a key factor mediating chemical coupling between PLC and TRP/TRPL channels remained a mystery until recently. It was found that breakdown of a lipid product of PLC cascade, diacylglycerol (DAG), by the enzyme diacylglycerol lipase, generates PUFAs that can activate TRP channels, thus initiating membrane depolarization in response to light. This mechanism of TRP channel activation may be well-preserved among other cell types where these channels perform various functions.
Clinical significance[edit]
Mutations in TRPs have been linked to neurodegenerative disorders, skeletal dysplasia, kidney disorders, and may play an important role in cancer. TRPs may make important therapeutic targets. There is significant clinical significance to TRPV1, TRPV2, TRPV3 and TRPM8’s role as thermoreceptors, and TRPV4 and TRPA1’s role as mechanoreceptors; reduction of chronic pain may be possible by targeting ion channels involved in thermal, chemical, and mechanical sensation to reduce their sensitivity to stimuli. For instance the use of TRPV1 agonists would potentially inhibit nociception at TRPV1, particularly in pancreatic tissue where TRPV1 is highly expressed. The TRPV1 agonist capsaicin, found in chili peppers, has been indicated to relieve neuropathic pain. TRPV1 agonists inhibit nociception at TRPV1
Role in cancer[edit]
Altered expression of TRP proteins often leads to tumorigenesis, as reported for TRPV1, TRPV6, TRPC1, TRPC6, TRPM4, TRPM5, and TRPM8. TRPV1 and TRPV2 have been implicated in breast cancer. TRPV1 expression in aggregates found at endoplasmic reticulum or Golgi apparatus and/or surrounding these structures in breast cancer patients confer worse survival.
TRPM family of ion channels are particularly associated with prostate cancer where TRPM2 (and its long noncoding RNA TRPM2-AS), TRPM4, and TRPM8 are overexpressed in prostate cancer associated with more aggressive outcomes. TRPM3 has been shown to promote growth and autophagy in clear cell renal cell carcinoma, TRPM4 is overexpressed in diffuse large B-cell lymphoma associated with poorer survival, while TRPM5 has oncogenic properties in melanoma.
TRP channels take center stage in modulating chemotherapy resistance in breast cancer. Some TRP channels such as TRPA1 and TRPC5 are tightly associated with drug resistance during cancer treatment; TRPC5-mediated high Ca influx activates the transcription factor NFATC3 (Nuclear Factor of Activated T Cells, Cytoplasmic 3), which triggers p-glycoprotein (p-gp) transcription. The overexpression of p-gp is widely recognized as a major factor in chemoresistance in cancer cells, as it functions as an active efflux pump that can remove various foreign substances, including chemotherapeutic agents, from within the cell.
TRPC5-mediated chemoresistance: TRPC5 overexpression activates the transcription factor NFATC3 Ca signaling pathway, leading to p-gp overexpression. Moreover, the overexpressed p-gp expels chemotherapeutic drugs such as doxorubicin triggering chemoresistance. Chemoresistant breast cancer cells overexpressing TRPC5 transfer channel units to chemo sensitive recipient cells via extracellular vesicles (EV), leading to the development of TRPC5-mediated chemoresistance in these cells.
Contrarily, other TRP channels, such as TRPV1 and TRPV2, have been demonstrated to potentiate the anti-tumorigenic effects of certain chemotherapeutic agents and TRPV2 is a potential biomarker and therapeutic target in triple negative breast cancer.
Role in inflammatory responses[edit]
In addition to TLR4 mediated pathways, certain members of the family of the transient receptor potential ion channels recognize LPS. LPS-mediated activation of TRPA1 was shown in mice and Drosophila melanogaster flies. At higher concentrations, LPS activates other members of the sensory TRP channel family as well, such as TRPV1, TRPM3 and to some extent TRPM8. LPS is recognized by TRPV4 on epithelial cells. TRPV4 activation by LPS was necessary and sufficient to induce nitric oxide production with a bactericidal effect.
History of Drosophila TRP channels[edit]
The original TRP-mutant in Drosophila was first described by Cosens and Manning in 1969 as "a mutant strain of D. melanogaster which, though behaving phototactically positive in a T-maze under low ambient light, is visually impaired and behaves as though blind". It also showed an abnormal electroretinogram response of photoreceptors to light which was transient rather than sustained as in the "wild type". It was investigated subsequently by Baruch Minke, a post-doc in the group of William Pak, and named TRP according to its behavior in the ERG. The identity of the mutated protein was unknown until it was cloned by Craig Montell, a post-doctoral researcher in Gerald Rubin's research group, in 1989, who noted its predicted structural relationship to channels known at the time and Roger Hardie and Baruch Minke who provided evidence in 1992 that it is an ion channel that opens in response to light stimulation. The TRPL channel was cloned and characterized in 1992 by the research group of Leonard Kelly. In 2013, Montell and his research group found that the TRPL (TRP-like) cation channel was a direct target for tastants in gustatory receptor neurons and could be reversibly down-regulated.
See also[edit]
Endocannabinoid system
Transient receptor potential channel-interacting protein database (2010) | biology | 2565453 | https://sv.wikipedia.org/wiki/Trithemis%20nuptialis | Trithemis nuptialis | Trithemis nuptialis är en trollsländeart som beskrevs av Karsch 1894. Trithemis nuptialis ingår i släktet Trithemis och familjen segeltrollsländor. IUCN kategoriserar arten globalt som livskraftig. Inga underarter finns listade i Catalogue of Life.
Källor
Segeltrollsländor
nuptialis | swedish | 1.018416 |
human_fear_dark/fear_response_dark.txt | Brief Report
Acute Stress Potentiates Anxiety in Humans
Author links open overlay panelChristian Grillon a, Roman Duncko b, Matthew F. Covington, Lori Kopperman a, Mitchel A. Kling c
Show more
Add to Mendeley
Share
Cite
https://doi.org/10.1016/j.biopsych.2007.06.007
Get rights and content
Background
Stress is an important factor in the development and maintenance of anxiety disorders. Stress also potentiates anxiety-like response in animals, but empirical evidence for a similar effect in humans is still lacking.
Methods
To test whether stress increases anxiety in humans, we examined the ability of a social stressor (speech and a counting task) to potentiate the facilitation of startle in the dark. Measures of subjective distress and of hypothalamic–pituitary–adrenal axis and autonomic nervous system activity (e.g., salivary cortisol, α-amylase, blood pressure, and heart rate) were also taken to confirm the effectiveness of the stress manipulation.
Results
Startle was significantly facilitated in the dark. This effect was potentiated by prior exposure to the social stressor. The social stressor induced increases in salivary cortisol and α amylase as well as increases in blood pressure, heart rate, and subjective distress.
Conclusion
The findings indicate that stress potentiates anxiety. Animal studies suggest that such an effect might be mediated by glucocorticoid effects on corticotropin-releasing hormone in limbic structures.
Section snippets
Methods and Materials
Participants were 20 medically and psychiatrically healthy volunteers (9 men) ages 28.1 years (SD = 8.3 years) who gave written informed consent.
The FSD was investigated in two sessions a week apart, one after a social stressor (stress) and the other after no stressor (control) in a between-subject design counterbalanced across subjects. The 10-min social stressor consisted of delivering a speech followed by a backward counting task (see Figure 1 for details). The FSD test started 25 min after
Results
Startle was facilitated by darkness, and this facilitation increased after stress (Figure 2). A Stress Condition (2) × Illumination (2) × Gender (2) ANOVA revealed an Illumination main effect [F(1,18) = 15.9, p < .0009] and a Stress Condition × Illumination interaction [F(1,18) = 6.4, p < .02]. Follow-up tests showed significant FSD in the control [F(1,19) = 6.8, p < .02] and stress condition [F(1,19) = 18.5, p < .0009].
The autonomic, endocrine, cardiovascular, and subjective responses
Discussion
To our knowledge this is the first report showing that unconditioned anxiety is enhanced by prior stress. Anxiety as measured with FSD was sensitized in humans exposed to a social stressor, consistent with animal data (7, 8).
The light-enhanced startle effect in the rat is mediated by CRH in the BNST (18), suggesting that the effect of changes in background illumination on startle (i.e., FSD) is also mediated by CRH acting on receptors in the BNST. Sensitized FSD by stress in humans might | biology | 10282 | https://da.wikipedia.org/wiki/Stress | Stress |
Hvad er stress?
Ordet stress har flere betydninger. I daglig tale kan begrebet stress beskrive at en person er presset på tid, eller har meget at se til. Stress er en betegnelse for en længere belastningstilstand. Den psykologiske betegnelse for stress beskriver en diagnose, som kan føre til udviklingen af fysiske sygdomme. Stress forbindes med negative konsekvenser som, angst og depression.
Virkning af stress
Kortvarig stress kan være nyttig, mens langvarig stress kan medføre alvorlig risiko for udvikling af livstruende sygdomme. Det hænger måske sammen med blandt andet udskillelsen og især effekterne adrenalin og kortisol. Det er dokumenteret at langvarig stress kan skade hjertet og blodkredsløbet (adrenalinet), og der er øget risiko for blodprop, alvorlige infektionssygdomme og kræft (kortisolen). Der er også fundet sammenhæng mellem stress og f.eks. knogleskørhed og type-2-diabetes.
En del stressrelaterede sygdomme er forbundet med aldringsprocessen, som har noget med stoffet interleukon-6 (IL-6) at gøre. Dette stof påvirker immunsystemet.
Symptomer og diagnosticering
De psykiske symptomer på stress er: anspændthed, ængstelse og irritation. De fysiske symptomer kan være hoved- og mavepine samt søvnløshed. Dog vil det være at betragte som en fejl, såfremt en læge (det værende egen læge eller på et hospital) diagnosticerer stress udelukkende ud fra disse symptomer. Stress er en klinisk diagnose og kan ikke objektivt måles med nuværende midler. En blod- eller urinprøve hvori niveauer af stresshormonerne cortisol og adrenalin kan måles, men disse siger intet om det reelle stressniveau individet befinder sig i og derfor er betegnelsen stresshormon egentlig misvisende, da disse kun er forhøjede ved akut og kortvarigt stress.
Det tætteste man kommer på en egentlig stressdiagnose er diagnosen tilpasningsreaktion (belastningsreaktion), som er defineret ved at individet har
Kendt traume eller belastning inden for den seneste måned (ikke af usædvanlig eller katastrofekarakter)
Emotionelle eller adfærdsmæssige symptomer uden at opfylde kriterierne for andre psykiske lidelser
Behandling
Der findes flere råd om stresshåndtering. Nogle af rådene er at
undgå de ting, der har påført en stress
tænke over de ting du stresser om
slappe af
anerkende din stress
sige fra
tale med andre
søg hjælp hos en læge, psykolog eller psykoterapeut.
Lægen kan hjælpe dig med sygemelding så du får ro til at komme ovenpå eller hvis du har brug for medicin.
Psykologen eller psykoterapeuten kan hjælpe dig med at bearbejde årsagerne og give dig redskaber til at håndtere fremtidig stress.
Det første skridt kan være at indse, at man er stresset. Er du i tvivl så tal med din læge eller andre fagpersoner.
Litteratur
Ballegaard, Søren (2020): Sandheden om stress. Gyldendal, 230 sider,
Dræby, Anders (2019): Stress, angst og depression – at fremme og kurere de samme lidelser. I: Glasdam og Boelsbjerg (red.) Folkesundhed – bag om intentioner og strategier. Gad, s. 343-353
Jørgensen, Per Schultz Jørgensen (2005): Familieliv i en stresset kultur. In: “Det svære liv”. Aarhus UniversitetsforlaG
Strøbæk, Pernille S. (2009). “Stress og socialitet”. Psyke & Logos, nr. 30(2), s. 26.
Referencer
Eksterne henvisninger
DR's tema om stresshttps://psykiatrifonden.dk/viden/temaer/stress
Psykiatri
Psykologi | danish | 0.480096 |
human_fear_dark/i-put-a-night-light-in-my-babys-room-so-she-wont-be-afraid-of-the-dark-and-other-well-meant-but-misguided-parenting-mistakes.txt |
About Services Free Consult Sleep & Parenting Hacks Success Videos Life Coaching About Services Free Consult Sleep & Parenting Hacks Success Videos Life Coaching
About Services Free Consult Sleep & Parenting Hacks Success Videos Life Coaching About Services Free Consult Sleep & Parenting Hacks Success Videos Life Coaching
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed. 0 Comments Leave a Reply. Author Abby Wolfson is a pediatric nurse practitioner, certified child sleep consultant and certified life coach for parents. She divides her time between Brooklyn, NY and San Miguel de Allende, Mexico. Archives March 2024 February 2024 January 2024 December 2023 November 2023 October 2023 September 2023 August 2023 July 2023 June 2023 May 2023 April 2023 March 2023 February 2023 January 2023 December 2022 November 2022 October 2022 September 2022 August 2022 July 2022 June 2022 May 2022 April 2022 March 2022 February 2022 January 2022 December 2021 November 2021 October 2021 September 2021 August 2021 July 2021 June 2021 May 2021 April 2021 March 2021 February 2021 January 2021 November 2020 October 2020 September 2020 August 2020 July 2020 June 2020 May 2020 April 2020 March 2020 February 2020 January 2020 December 2019 November 2019 October 2019 Categories All Achieving Your Impossible Goals Adolescents Boundaries Breastfeeding Coaching Developmental Leaps Early Waking Illness Life Coaching Living Internationally Naps Newborns Overtiredness Parent Coaching Positive Reinforcement Postpartum Depression Preschoolers Self Soothing Setting Limits Sibling Challenges Single Parents Sleep Crutches Sleep Deprivation Sleep Essentials Sleep Obstacles Sleep Schedules Sleep Training Methods Sleep Training Success Sleep Transitions Strong Attachment Tantrums Thought Work Time Changes Toddlers Travel Twins Vulnerability Wake Windows RSS Feed
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed. 0 Comments Leave a Reply. Author Abby Wolfson is a pediatric nurse practitioner, certified child sleep consultant and certified life coach for parents. She divides her time between Brooklyn, NY and San Miguel de Allende, Mexico. Archives March 2024 February 2024 January 2024 December 2023 November 2023 October 2023 September 2023 August 2023 July 2023 June 2023 May 2023 April 2023 March 2023 February 2023 January 2023 December 2022 November 2022 October 2022 September 2022 August 2022 July 2022 June 2022 May 2022 April 2022 March 2022 February 2022 January 2022 December 2021 November 2021 October 2021 September 2021 August 2021 July 2021 June 2021 May 2021 April 2021 March 2021 February 2021 January 2021 November 2020 October 2020 September 2020 August 2020 July 2020 June 2020 May 2020 April 2020 March 2020 February 2020 January 2020 December 2019 November 2019 October 2019 Categories All Achieving Your Impossible Goals Adolescents Boundaries Breastfeeding Coaching Developmental Leaps Early Waking Illness Life Coaching Living Internationally Naps Newborns Overtiredness Parent Coaching Positive Reinforcement Postpartum Depression Preschoolers Self Soothing Setting Limits Sibling Challenges Single Parents Sleep Crutches Sleep Deprivation Sleep Essentials Sleep Obstacles Sleep Schedules Sleep Training Methods Sleep Training Success Sleep Transitions Strong Attachment Tantrums Thought Work Time Changes Toddlers Travel Twins Vulnerability Wake Windows RSS Feed
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed. 0 Comments Leave a Reply. Author Abby Wolfson is a pediatric nurse practitioner, certified child sleep consultant and certified life coach for parents. She divides her time between Brooklyn, NY and San Miguel de Allende, Mexico. Archives March 2024 February 2024 January 2024 December 2023 November 2023 October 2023 September 2023 August 2023 July 2023 June 2023 May 2023 April 2023 March 2023 February 2023 January 2023 December 2022 November 2022 October 2022 September 2022 August 2022 July 2022 June 2022 May 2022 April 2022 March 2022 February 2022 January 2022 December 2021 November 2021 October 2021 September 2021 August 2021 July 2021 June 2021 May 2021 April 2021 March 2021 February 2021 January 2021 November 2020 October 2020 September 2020 August 2020 July 2020 June 2020 May 2020 April 2020 March 2020 February 2020 January 2020 December 2019 November 2019 October 2019 Categories All Achieving Your Impossible Goals Adolescents Boundaries Breastfeeding Coaching Developmental Leaps Early Waking Illness Life Coaching Living Internationally Naps Newborns Overtiredness Parent Coaching Positive Reinforcement Postpartum Depression Preschoolers Self Soothing Setting Limits Sibling Challenges Single Parents Sleep Crutches Sleep Deprivation Sleep Essentials Sleep Obstacles Sleep Schedules Sleep Training Methods Sleep Training Success Sleep Transitions Strong Attachment Tantrums Thought Work Time Changes Toddlers Travel Twins Vulnerability Wake Windows RSS Feed
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed. 0 Comments Leave a Reply.
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed. 0 Comments Leave a Reply.
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed. 0 Comments
"I Put a Night Light in My Baby´s Room So She Won´t Be Afraid of the Dark" and Other Well-Meant But Misguided Parenting Mistakes 6/23/2020 0 Comments
As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed.
As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby ' s room is a perfect example of this. Children don't develop a fear of the dark before age 2-3. Before that time, their brains simply don't have the developmental maturity to imagine scary monsters under the bed. And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy. For this reason, putting a night light in your baby 's room is hurting her sleep without any benefit. If she cries at bedtime, it´ s because she doesn ' t want to separate from you, or because she is overtired, not because she 's scared of the dark. Adding a night light will only make things worse in that she ' ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter ' s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning. At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won ' t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom. If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down. Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child 's routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?" Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day. It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn 't you rather have a child that is (nearly) always well-rested and occasionally doesn ' t sleep well because you can 't recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.) As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn 't know better and put her down to nap too late. It was miserable for both of us. If you can 't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed.
As parents, we – with the best of intentions – tell ourselves a lot of stories about our children. And sometimes those stories can seriously undermine our families' needs. Putting a night light in a baby
And light in the bedroom is a bad idea. Light signals the brain that it´s time to be awake. We want to send the opposite message at bedtime and nap time, that it´s time to sleep. Darkness cues the body to release the hormone melatonin, which makes us sleepy.
ll have an even harder time falling asleep. (And by the way, separation anxiety is perfectly normal and is not a reason to keep your tired child up, even if is what she thinks she wants. It´s only making things worse. When you get her caught up on her sleep debt, you will both see this.) The best way to sleep, at all ages, is in total darkness. You should not be able to see your hand in front of your face. You will likely need blackout shades to achieve this level of darkness. Your local hardware store can sell you inexpensive stick-on shades that you can try before you invest in a more permanent solution. You can also do what I did with my oldest, and tape up black garbage bags over the windows. Depressingly ugly but remarkably effective! You may also need to add painter
s tape around the edges of the blackout shades to prevent any light leakage. This may seem like overkill but it´s an easy step to try if your child is waking up too early. Even a tiny bit of morning light can wake a little one in the early morning.
At age 2-3, your child may develop a true fear of the dark. Only at this point should you consider introducing a night light. I use a portable Munchkin night light for my own kids. It turns off on its own after a few minutes, so it won
t disturb their slumber once they drift off, plus the portability is great for those scary midnight trips to the bathroom.
If your child needs a light that stays on all night, pick something that has red light, not blue. Blue light is stimulating to the brain and tells us to wake up. Electronic screens contain blue light and for this reason, you should avoid exposure to screens in the hour before bedtime, too. I recommend turning down all the lights in the house in the hour before bedtime. This is one more cue to the brain to start winding down.
Keep night lights as far from the bed as possible. And keep the number to a mínimum. Your child should not sleep with more than one all-night night light. If your older child (preschool or older) child is sleeping with multiple night lights, you will need to wean him off them. When making big changes to your child
routine, I always suggest having a conversation with your child ahead of time (not at bedtime!), getting his buy-in, and rewarding his cooperation. After all, this is your idea, not his. I might say something like, ¨"I have learned that sleeping with too much light on makes it harder for our brains to relax. I want to work on having fewer night lights in your room. I know that might be hard for you. What prize would you like to earn for working on this?"
Some parents fear that creating a dark sleep environment will create dependence or worse, that their children will wake up cranky and miserable if they sleep in darkness in the middle of the day.
It´s true that if you put your child to sleep in the dark, he may require darkness to sleep. And that dependence on darkness can occasionally be inconvenient. But wouldn
recreate his ideal sleep environment on the go? Versus a child that is so overtired that he will fall asleep wherever he is, but is never well-rested? (A child that always falls asleep in the car is an overtired child.)
As for your child waking up cranky after a nap in the dark? The problem is not the darkness. The problem is the timing of the nap. A correctly-timed nap will not result in crankiness. A correctly-timed nap in the dark will coincide with your child´s natural surge in melatonin, the sleepy hormone, so your child will nap well and wake up well-rested and happy. An incorrectly-timed nap results in that cranky, miserable feeling we adults feel, too, when we sleep at the wrong time. It´s called nap inerti a, and my older daughter had it all the time because I didn
't put her down on time, it's better to keep her up and implement a very early bedtime instead. It can take courage to make big changes in your child's sleep routine, like eliminating night lights. Be patient and encouraging with both of you. If you would like some support along the way, schedule a free chat and get your family the sleep you deserve, guaranteed.
Author Abby Wolfson is a pediatric nurse practitioner, certified child sleep consultant and certified life coach for parents. She divides her time between Brooklyn, NY and San Miguel de Allende, Mexico. Archives March 2024 February 2024 January 2024 December 2023 November 2023 October 2023 September 2023 August 2023 July 2023 June 2023 May 2023 April 2023 March 2023 February 2023 January 2023 December 2022 November 2022 October 2022 September 2022 August 2022 July 2022 June 2022 May 2022 April 2022 March 2022 February 2022 January 2022 December 2021 November 2021 October 2021 September 2021 August 2021 July 2021 June 2021 May 2021 April 2021 March 2021 February 2021 January 2021 November 2020 October 2020 September 2020 August 2020 July 2020 June 2020 May 2020 April 2020 March 2020 February 2020 January 2020 December 2019 November 2019 October 2019 Categories All Achieving Your Impossible Goals Adolescents Boundaries Breastfeeding Coaching Developmental Leaps Early Waking Illness Life Coaching Living Internationally Naps Newborns Overtiredness Parent Coaching Positive Reinforcement Postpartum Depression Preschoolers Self Soothing Setting Limits Sibling Challenges Single Parents Sleep Crutches Sleep Deprivation Sleep Essentials Sleep Obstacles Sleep Schedules Sleep Training Methods Sleep Training Success Sleep Transitions Strong Attachment Tantrums Thought Work Time Changes Toddlers Travel Twins Vulnerability Wake Windows RSS Feed
Author Abby Wolfson is a pediatric nurse practitioner, certified child sleep consultant and certified life coach for parents. She divides her time between Brooklyn, NY and San Miguel de Allende, Mexico. Archives March 2024 February 2024 January 2024 December 2023 November 2023 October 2023 September 2023 August 2023 July 2023 June 2023 May 2023 April 2023 March 2023 February 2023 January 2023 December 2022 November 2022 October 2022 September 2022 August 2022 July 2022 June 2022 May 2022 April 2022 March 2022 February 2022 January 2022 December 2021 November 2021 October 2021 September 2021 August 2021 July 2021 June 2021 May 2021 April 2021 March 2021 February 2021 January 2021 November 2020 October 2020 September 2020 August 2020 July 2020 June 2020 May 2020 April 2020 March 2020 February 2020 January 2020 December 2019 November 2019 October 2019 Categories All Achieving Your Impossible Goals Adolescents Boundaries Breastfeeding Coaching Developmental Leaps Early Waking Illness Life Coaching Living Internationally Naps Newborns Overtiredness Parent Coaching Positive Reinforcement Postpartum Depression Preschoolers Self Soothing Setting Limits Sibling Challenges Single Parents Sleep Crutches Sleep Deprivation Sleep Essentials Sleep Obstacles Sleep Schedules Sleep Training Methods Sleep Training Success Sleep Transitions Strong Attachment Tantrums Thought Work Time Changes Toddlers Travel Twins Vulnerability Wake Windows RSS Feed
Abby Wolfson is a pediatric nurse practitioner, certified child sleep consultant and certified life coach for parents. She divides her time between Brooklyn, NY and San Miguel de Allende, Mexico.
March 2024 February 2024 January 2024 December 2023 November 2023 October 2023 September 2023 August 2023 July 2023 June 2023 May 2023 April 2023 March 2023 February 2023 January 2023 December 2022 November 2022 October 2022 September 2022 August 2022 July 2022 June 2022 May 2022 April 2022 March 2022 February 2022 January 2022 December 2021 November 2021 October 2021 September 2021 August 2021 July 2021 June 2021 May 2021 April 2021 March 2021 February 2021 January 2021 November 2020 October 2020 September 2020 August 2020 July 2020 June 2020 May 2020 April 2020 March 2020 February 2020 January 2020 December 2019 November 2019 October 2019
All Achieving Your Impossible Goals Adolescents Boundaries Breastfeeding Coaching Developmental Leaps Early Waking Illness Life Coaching Living Internationally Naps Newborns Overtiredness Parent Coaching Positive Reinforcement Postpartum Depression Preschoolers Self Soothing Setting Limits Sibling Challenges Single Parents Sleep Crutches Sleep Deprivation Sleep Essentials Sleep Obstacles Sleep Schedules Sleep Training Methods Sleep Training Success Sleep Transitions Strong Attachment Tantrums Thought Work Time Changes Toddlers Travel Twins Vulnerability Wake Windows
SCHEDULE A FREE CONSULT Every child deserves a great night's sleep. © 2019 Peaceful Parent Sleep Coaching. All Rights Reserved. Storybrand Website Design by Red Door Designs
SCHEDULE A FREE CONSULT Every child deserves a great night's sleep. © 2019 Peaceful Parent Sleep Coaching. All Rights Reserved. Storybrand Website Design by Red Door Designs
SCHEDULE A FREE CONSULT Every child deserves a great night's sleep. © 2019 Peaceful Parent Sleep Coaching. All Rights Reserved. Storybrand Website Design by Red Door Designs
© 2019 Peaceful Parent Sleep Coaching. All Rights Reserved. Storybrand Website Design by Red Door Designs | biology | 2890856 | https://sv.wikipedia.org/wiki/Triphaenopsis%20punctisignata | Triphaenopsis punctisignata | Triphaenopsis punctisignata är en fjärilsart som beskrevs av Embrik Strand 1921. Triphaenopsis punctisignata ingår i släktet Triphaenopsis och familjen nattflyn. Inga underarter finns listade i Catalogue of Life.
Källor
Nattflyn
punctisignata | swedish | 1.256814 |
human_fear_dark/dealing-with-fear-of-the-dark.txt |
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog / Helping families get the sleep the need Blog Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog / January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ... Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog / Helping families get the sleep the need Blog Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog / January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ... Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog / Helping families get the sleep the need Blog Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog /
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog / Helping families get the sleep the need Blog Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog /
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog /
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog /
Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog /
Home / About / Approach / Services / Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards Contact / Contact Book a Call Blog /
Virtual Mini Sessions Prenatal/Newborn Sleep Package Pediatric Sleep Packages (4 months-7 years) Parent Coaching My Favourite Sleep Products Gift Cards
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ...
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ...
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ...
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ...
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis Feeling Helpless? 6 Sleep Hacks To ... 8 Tips for Travelling & ...
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis
January 29, 2020 Dealing with Fear of the Dark January 29, 2020 / Lindsay Lewis “Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome. January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
“Mommy, I can’t sleep. I’m scared of the dark.” Sound familiar? Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children. As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off. This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks. When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!” So what’s the best way to handle these fears? As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution. (Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.) Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here. For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety. The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.) Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one. This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night. One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
Fear of the dark usually starts to show up around the 2-year mark. As a toddler’s mind matures, their memory gets longer and their imagination develops. They’ve almost certainly taken a spill on the playground or had some kind of traumatic incident by this point, so they’re aware that there are things out there that can hurt them. They’ve also probably seen a few movies or been read a few books that touch on a couple of spooky or eerie elements, even if they’re geared towards children.
As adults, we’re experienced enough to recognize that the dark isn’t inherently dangerous. But for a toddler, there’s no history to draw on to assure them that they’re safe and secure after the lights go out. So, my first, and most important piece of advice when you’re addressing your little one’s fear of the dark is this; don’t brush it off.
This can be a bit of a tricky landscape to navigate. On the one hand, we absolutely want to show empathy and understanding when something frightens our kids. On the other, we don’t want to add fuel to the fire. This is why I’m not a big fan of “monster spray” or nightly closet checks.
When we tell our kids, “Nope! No monsters in the closet or under your bed! Not that I noticed, anyway, so you’re all good,” it’s not nearly as reassuring as you might think. It’s easy to see how they could interpret that as, “Yeah, there’s absolutely such a thing as monsters, they’re scary as hell, and they do tend to live kids’ closets, but I don’t see one in there at the moment, so... sleep tight!”
As I mentioned, dismissing your little one’s fears as irrational isn’t all that helpful, so ask some questions when they express a fear of the dark. Digging into their concerns is helpful in a couple of ways. It lets them know that you’re taking them seriously, which is very reassuring. It also helps you to assess what it is about the darkness that frightens them and helps you to address it. For example, if they tell you they’re seeing things moving around their room, it might be caused by shadows. Headlights from cars driving by can often shine enough light through curtains or blinds to throw shadows across the room. Coupled with a toddler’s imagination, that can create some seriously intimidating scenes. In that situation, a nightlight or some blackout blinds can prove to be a quick, effective solution.
(Tip: If you’re going to use a nightlight, make sure it’s a warm colour. Blue lights may look soothing, but they stimulate cortisol production, which is the last thing we want at bedtime.)
Now, that’s a bit of an ideal scenario. As you’re likely already aware, getting a clear, concise answer from a toddler about what they are scared of could be tricky. To a toddler, “Paw Patrol” is a reasonable answer to, “What do you want for lunch?” So, you’re likely going to have to work with slightly more obscure information, but we’re showing concern, and that goes a long way here.
For a lot of toddlers, bedtime is the only time of the day that they’re left alone. They’re either playing with friends, hanging close to their parents, or supervised in some way, shape, or form by a grown-up. Bedtime is also the only time they’re exposed to darkness, so you can see how the two things together could easily cause some anxiety.
The obvious (and super fun!) way to ease some of that apprehension is to spend some time together in the dark. Reading books under a blanket with a dim flashlight is a great activity. Some hide and seek with the lights out is tons of fun as well, just as long as you clear any tripping hazards out of the area, you’re going to be playing in. (It doesn’t have to be pitch black. We just want to get some positive associations with low-light situations.)
Shadow puppets are a great too, even though I’m personally terrible at them. Hide-and-Clap is also a classic. A quick Google search will give you dozens of ideas, so pick two or three that you think your child will like, then let them choose one.
This isn’t likely to be an overnight fix, but stay respectful, calm, and consistent. After your little one’s fears have been addressed and they’ve learned that the darkness is more fun than frightening, you’ll start seeing more consolidated sleep and less visits in the middle of the night.
One last little tip, turning down the lights gradually as your little one’s bedtime approaches is a good way to ease them into a dark setting, and also helps to stimulate melatonin production, which will help them get to sleep easier. Two birds, one stone. You’re welcome.
January 29, 2020 / Lindsay Lewis / Infant sleep , healthy sleep , Toddler sleep , sleep tip Sleep Tips Lindsay Lewis
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL Copyright 2024 Lindsay Lewis Sleep Solutions Privacy Policy
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! SCHEDULE A FREE 15 MINUTE DISCOVERY CALL
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit!
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit!
Not sure what package is right for you? Want to tell us more about your sleep situation and learn more about our sleep philosophy before diving in? Book a FREE 15-minute discovery call with us to determine if we are good fit! | biology | 123060 | https://da.wikipedia.org/wiki/The%20Simpsons%20%28s%C3%A6son%209%29 | The Simpsons (sæson 9) | Den niende sæson af tv-serien The Simpsons blev første gang sendt i 1997 og 1998.
Afsnit
The City of New York vs. Homer Simpson
Familien Simpsons skal til New York for at hente Homers bil hjem, efter at Barney Gumble har stjålet den. Men Homer har dårlige minder fra New York.
The Principal and The Pauper
Det er Seymour Skinner´s 20-års jubilæum som inspektør på Springfield Elementary School. De andre lærere forbereder en stor fest sammen med eleverne. Bart Simpson skal lave en kage, Lisa Simpson og Ralph Wiggum skal holde en tale og nogle børn synger en sang. Da arrangementet begynder kører der tilfældigvis en bil forbi skolen. En der sidder i bilen får øje på skiltet hvor der står: "Tillykke Seymour Skinner!". Ham der sidder i bilen går ind på skolen siger at Seymour er en bedrager. Det viser sig at den mystiske person faktisk hedder: Seymour Skinner. Og Seymour Skinner(som han nu ikke hedder) hedder rigtigt: Armin Tamzarian. De var begge med i Vietnamkrigen, hvor de de var kammerater. En dag tror alle tropperne at DEN RIGTIGE Seymour Skinner dør af en bombe, som han jo ikke gjorde. Armin Tamzarian får den opgave at fortælle det til Seymours mor Agnes Skinner. Men han synes det var for synd for hende at hendes egen søn var død, så han begyndte at spille Seymour over for hende. Hun synes også at han ligner Seymour. Så Tamzarian udgør de drømme Skinner havde. Men nu må Tamzarian rejse tilbage til sin hjemby Capitol City. Men kan Springfields befolkning undvære ham, de nu er vant til?
Lisa's Sax
Da Lisas saxofon går i stykker efter et skænderi med Bart, og hun siger hun har haft den så længe hun kan huske det, beslutter Homer sig for at fortælle hende om dengang hun fik den.
Historien kommer nu mest til at omhandle Bart.
Om Barts glæde ved at skulle starte i skole, og hvordan han efter første dag allerede har mistet lysten til at gå i skole.
Til en samtale hos skolens psykolog om Barts manglende glæde ved at gå i skole, påpeger psykologen hvor begavet Lisa er, og foreslår at de sender hende på førskole.
Det har de dog ikke råd til, men beslutter sig for at finde en anden måde at berige Lisas intelligens på.
Alt dette sker samtidig med at der er en massiv hedebølge over Springfield, og Homer har sparet sammen til en aircondition, men på vej ned for at købe den, får Homer et tegn om at han skal købe en Saxofon i stedet.
Tilbage til nutiden har de igen sparet op til en aircondition, men bruger dem på en ny saxofon i stedet.
Treehouse of Horror VIII
- "The Homega Man"
- Fly Vs. Fly
- Easy-Bake coven
. I fly vs. fly , køber Homer en maskine af professor Fink der kan transportere en person fra et sted til et andet. Bart opdager, ved et tilfælde, at man kan ændre DNA ved at putte to organismer ind i maskinen på samme tid. Det opdager han da Hunden og katten løber ind på samme tid , og 2 dyr kommer ud, en med 2 hoveder og en med to røve, som han konkluderer må være Lisas.
Selv går han derind med en flue i håbet om at blive en superflue.
Det går ikke helt som planlagt da han selv bliver rigtig lille med flue krop, og hans krop vandrer rundt med fluens hoved.
Bart forsøger at få processen vendt, men det er fluen dog ikke helt enig i.
Fluen prøver at dræbe Lisa, og det lykkedes Bart at skubbe ham ind i maskinen igen.
Homer og Marge har tilsyneladende ikke lagt mærke til ændringen i huset, før alting er tilbage til normalt, og Homer finder en økse frem for at gøre noget han skulle have gjort for længe siden.
"I'll teach you to play with my machine!"
.Easy-Bake coven er historien om den første Halloween nogensinde.
Springfields hekseafbrændinger har taget overhånd og der skal ikke noget til for at blive anklaget som Heks. Da Marge prøver at hisse pøblen ned, bliver hun selv anklaget for at være heks, og bliver stillet over for et ultimatum. Hun skal hoppe ud over en klippe, og hvis hun ikke er en heks vil hun stige til himmels og få en værdig kristen død, og hvis hun er en heks vil hun flyve væk i sikkerhed. Hvorefter hun selvfølgelig er nødt til at komme tilbage så hun kan blive brændt.
Mod al forventning og til alles overraskelse viser det sig, at Marge Simpson faktisk ER en heks, og hun flyver derfra hen til sine 2 søstre.
De planlægger sammen at kidnappe og spise byens børn, men på deres togt, bliver de overbevist af Maude Flanders om at de hellere skal tage deres kager.
Det synes de er en god ide, og sådan kommer traditionen om Halloween frem.
The Cartridge Family
En fodboldkamp får Homer til at købe en pistol for at beskytte hans familie, men Marge vil have ham til at love at han vil smide den ud, men han gør det ikke i al hemmelighed.
Bart Star
Springfields drenge er blevet for tykke, og det får channel 6 til at opfordre at de skal begynde til Sport.
Bart melder sig til Junior Football holdet The Wildcats, ledet af Ned Flanders, og med Nelson Muntz som deres Quarterback vinder de alle kampene.
Efter mange tilråb fra Homer Simpson om at Ned Flanders er dårlig, overlader Flanders posten som træner til Homer, der er fast besluttet på at elske Bart mere, og derfor lader ham blive den nye Quarterback.
Bart ved godt at han ikke egner sig som Quarterback, og efter flere nederlag og advarsler fra holdet, vælger han at stoppe. Hvilket resulterer i en stor konflikt mellem Bart og Homer. Der blandt andet får Homer til, ved en fejltagelse at sige sit Job op fordi han prøver at vise en pointe om at give op.
The Two Mrs. Nahasapeemapetilons
Efter en aktion på Springfield rådhus, hvor ungkarlene i Springfield har sat dem selv til salg for at skaffe penge til en ny brandbil, og Apu Nahasapeemapetilon viser sig at være meget eftertraktet, beslutter Apu sig om at han nu skal leve livet. Det bliver dog kun for en kort stund, da han modtager et brev fra sin mor om at det er tid til hans arrangerede ægteskab.
Han prøver at undgå det ved at bilde sin mor ind at han er gift med Marge, men efter at Apus mor finder Homer i Marges seng, begynder hun at arrangere.
Apu bliver så gift med Manjula, og de finder und af at det faktisk godt kan lykkes mellem dem.
Lisa the Skeptic
Et skelet er blevet opdaget i Springfield, men det ser ud at have vinger så beboerne begynder at tro det er en engel, hvilket får lisa til at finde en rationel forklaring. Desværre tror ingen på hende.
Realty Bites
Marge er blevet en ejendomsmægler, men hun kan ikke holde ud at lyve for folk, så hun får ikke solgt et eneste, indtil hun begynder at lyve overfor familien Flanders
Miracle on Evergreen Terrace
Efter at have drukket 12 glas vand før sengetid, lykkes det Bart at vågne før resten af familien den 25. december for at gå ned og kigge på sine gaver.
Dette resulterer desværre i at juletræet bliver brændt ned, og han begraver de sølle rester i forhaven under sneen i den tro at det vil skjule det.
Da resten af familien spørger forfærdet til hvad der er sket, bryder Bart grædende sammen og fortæller at der har været en tyv og stjålet alle gaverne.. inklusive træet..
Tragedien spredes over hele Springfield og alle donerer penge til familien i trøst, og de køber sig en ny bil, der kort efter synker til bunds i Springfield Lake.
Senere indrømmer Bart over for familien at det var ham der ved et uheld kom til at brænde træet ned. Familien bliver afbrudt, i færd med at kvæle Bart, af nyhedsfolkene der vil høre til hvordan det går.
Under interviewet finder politiet resterne af træet i forhaven, og familien går til bekendelse om at det var Bart der brændte træet.
Det får hele familien til at blive forhadt i Springfield, indtil de får det afklaret ved at Springfields beboere får deres penge tilbage igennem Simpsons egendele.
All Singing, All Dancing
Homer lejer en Clint Eastwood film for at finde ud at det er en musical. Da Homer syntes at sang er noget svanset noget, begynder Marge og Lisa at rode i fortiden.
Dette udvikler sig til et clip-show fyldt med sang og dans.
Bart Carny
Homer og Bart tager til karneval hvor Bart kommer til at ødelægge Hitlers Bil, og efter slag i hovedet af Nelson Muntz, der spørger, hvad Hitler nogen sinde har gjort ham, bliver Homer og Bart ansat for at betale gælden af.
De bliver på et tidspunkt bedt om at passe ringboden. Og kommer selvfølgelig til at dumme sig så boden bliver beslaglagt af politiet.
Homer tilbyder i stedet ejeren og hans søn at flytte ind hos dem, og der går ikke længe før de to har overtaget Casa del Simpsons. Familien flytter imidlertid op i Barts træhytte, og planlægger hvordan de skal få huset tilbage.
For hvordan fupper man en fupmager?
The Joy of Sect
Homer er blevet hjernevasket ind til en sekt i Springfield. Da Marge får omprogrammeret Homer, begynder han at afsløre sekten hemmeligheder, hvilket giver ham mange problemer.
Das Bus
Springfields folkeskoles bus går i stykker på en øde ø og Lisa forsøger at lave et minisamfund der.
The Last Temptation of Krust
Klovnen Krusty finder ud af at hans materiale ikke dur mere så han finder på nogen nye måder at underholde, men han ender jo selvfølgelig på sine egne måder igen.
Dumbbell Indemnity
Efter lang tids søgen finder Moe Szyslak endelig kærligheden. Renee (Helen Hunt), bliver hans eneste ene. Og han bruger alle sine penge på hende, indtil han en dag ikke har flere, og af frygt for at Renee vil forlade ham, overtaler han Homer til at stjæle hans bil så han kan få forsikrings pengene.
Dette mislykkes da Homer, i stedet for at sætte bilen på jernbanesporerne, tager i drive-in for at se Hail to the Chimp.
Han bliver sat i fængsel efter at have kørt bilen i havet, og Moe søger desperat efter en måde at få Homer ud igen uden at bruge sine penge, eller selv at ende i fængslet.
En plan der indeholder 2 lig, en brændt bar og et nyt liv på Hawaii.
Lisa, the Simpson
Lisa begynder at føle sig dummere og dummere, og Ape Simpson informerer hende om det såkaldte Simpson Gen, der gør, at alle Simpsonerne bliver dummere og dummere med alderen.
Panikken indfinder sig hos Lisa, og hun går i tv for at få alle til at holde mere af deres hjerner.
Det får Homer til at ringe til alle fra familien Simpson i området for at vise hende, at der i hvert fald ikke er noget galt med Simpson-genet.
En plan, der mislykkes, da det viser sig, at alle mændene er dumme.
En skyder fugle i lufthavnen, en bliver kørt ned af kendte og sagsøger dem, og en spiller millionær til fester. Eller det vil han i hvert fald gerne.
Men det viser sig at alle kvinderne i Simpson-klanen er kloge, og at fejlen kun ligger i x-kromosomet.
This Little Wiggy
Bart bliver venner med en af skolens nørder Ralph Wiggum og narre Ralph til at give ham hans fars supernøgle til hele byens huse. På grund af det kommer de til at risikere borgmesteren liv.
Simpson Tide
Homer og hans venner melder sig ind i Flådens Reserve og dermed får Homer på mystisk vis lov til at styre en atomubåd. Det ender selvfølgelig galt og de får både USA og Rusland på nakken af sig.
The Trouble With Trillions
efter Homer for sent finder ud af at man er nødt til at indgi sin selvangivelse HVERT år, bliver han ansat hos IRS til at udspionere sine venner.
Han bliver også sat til at udspionere Montgomery Burns der er mistænkt for at have stjålet en 1 milliard seddel fra USA, der skulle have været givet til Frankrig i sin tid.
I stedet stikker han af sammen med Burns og Smithers til Cuba, og Fidel Castro stjæler sedlen fra ham , og de må padle hjem til Amerika igen.
Girly Edition
Trash of the Titans
AFSNIT NUMMER 200
efter et skænderi med skraldemændene nægter de at hente skrald hos familien Simpson mere.
eftersom skraldet hober sig op i forhaven og Bart får pest, bliver Marge mere og mere irriteret, og det ender med at hun sender en undskyldning til skraldemændene i Homers navn der får dem til at hente skraldet.
Da Homer finder ud af det, bliver han meget vred, og opsøger skraldechefen Ray Patterson, for at få trukket sin undskyldning tilbage, og efter et selvopdigtet skænderi med ham, beslutter Homer at stille op til valget som Skraldechef, og vinde det.
Men Homer har i valgtiden lovet en masse ting, og ting koster penge. Så for at få penge til lønningerne, bliver han betalt af andre byer rundt om i Amerika for at lade dem dumpe skrald i deres gamle miner. Dette resulterer i at Springfield bliver overfyldt med skrald, og de er nødt til at flytte hele Byen 15 km.
King of the Hill
efter en Søndags kirkepicnic hvor både Homer og Flanders deltager i børnenes leg, og Homer bliver ydmyget og Bart flov, lover Homer over for sig selv og Marge at bart aldrig mere skal føle skam over at have Homer til far.
Han begynder at træne i al hemmelighed, og bliver et muskelbundt, undervist af Rainer Wolfcastle.
Han spiser kun proteinbarer fra et firma, der hyrer ham til at bestige bjerget Murderhorn.
Lost Our Lisa
Efter at Bart har limet ting fast til sit ansigt og marge er nødt til at køre ham på hospitalet, bliver lisas tur til den egyptiske udstilling på springfield museeum aflyst.
Hun får narret Homer til at sige hun gerne må tage med bussen selv, og det går galt og hun ender i den forkerte bus på et forkert sted.
Efter at Lenny og Carl bliver forargede over at Homer har ladet sin 8-årige datter tage alene med bussen, bliver han bekymret og tager ud og leder efter hende.
Han finder hende, og de beslutter sig for at tage ind og se udstillingen efter der er lukket, og de opdager en gammel hemlighed.
Afsnittet indeholder Matt Groewnings egen yndlingsreplik.
Da Homer kører i en løbsk kran ude i vandet siger han " I'm not usually a praying man, but if you're out there PLEASE help me Superman"
Natural Born Kissers
Homer og Marge har begge mistet lysten til Sex, men da de en dag strander på en landevej, og er nødt til at tilbringe natten i en lade, genfinder de lysten. de opdager at de tænder på at have sex på steder hvor de kan blive opdaget.
Kort sagt resulterer det i at de er jagtet nøgne af hele byen efter at de blev opdaget på en minigolf bane.
09 | danish | 1.017776 |
human_fear_dark/.txt |
Beauty Fitness Lifestyle Health Food Shopping Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe Search Search Button Lifestyle Healthy Mind Healthy Sleeping Habits Yes, Nickelodeon, I’m a Grown Woman Who Is Afraid of the Dark and Sleeps With the Light On Mary Grace Garis Mary Grace Garis February 26, 2019 Pin It Share on facebook Share on twitter Share on pinterest Share on email I think my nyctophobia turned my partner into Sandra Bullock in Bird Box . At least, that's who he most reminded me of the other night when I saw him wearing the sleep mask he uses as a shield from the light I desperately need to stay on all night. Without it, my anxiety soars sky high and my heart pounds. All of this is because—to answer that famed Nickelodeon-prompted question—I am indeed afraid of the dark. Or, maybe I'm just afraid of Samara from The Ring . I don’t find it a coincidence that I started sleeping with the light on after watching the film in 2002, but many find it puzzling that my fright has extended so many years. I've found that being an adult and simultaneously having a fear of the dark prompts so many questions from the few people who know about it, and the tonal subtext is often, “Why—and why would you ever share that information?” I get the judgment. See, nyctophobia , as Healthline has taught me with a not-so-necessary air of condescension, tends to dissipate with age. However, my fear really isn't that huge of an anomaly. One doc says an estimated 11 percent of the United States population is afraid of the dark , making it a more common phobia than a fear of heights. And unlike with heights, where the visual stimuli is responsible for the palpations, nyctophobia is more rooted in the anxiety of having impaired vision, invoking a feeling of defenselessness. The small lamp on my desk is typically enough to keep me calm and happy, but anything from a bad dream to a before-bed horror movie to even the Sunday Scaries can push me to light up the whole room. The more fearful I get, the more bulbs are necessary, as if illuminating my room is shrouding me from anxiety bogeymen. Related Stories {{ truncate(post.title, 12) }} {{post.sponsorText}} There were, once upon a time, some stipulations. I didn’t force my freshmen-year roommates to keep the light on or alert my random hookups that they needed to save me from the monster under the bed. Historically, having someone with me kept me from freaking, which is a real Freudian take on things. Old Sigmund thought a fear of the dark correlated with separation anxiety, writing in his General Introduction to Psychoanalysis , “I once heard a child, who was afraid of the dark, call into an adjoining room, ‘Auntie, talk to me, I am afraid.’ ‘But what good will that do to you? You cannot see me!’ Whereupon the child answered, ‘If someone speaks, it is brighter.’” Word, little dude. I found myself waking up in a full-force panic attack, terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. And so, for years, having a body in place of a night light made the room brighter until it...didn’t. When my S.O. and I went to the Catskills for his friend’s wedding, I found myself waking up in a full-force panic attack , terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. What’s especially fun and great about this is that it sure as hell cuts into the quality of my sleep. One small Ryerson University study on 93 college-aged men and women found more than half of the 42 who suffered from insomnia reported a fear of the dark . And it’s a real catch-22 because you know what helps you go to sleep? Um, the effing dark. Like, it’s why our iPhones keep us awake every night . According to the American Medical Association , blue or white nighttime light suppresses melatonin release and interrupts circadian biological rhythms. Perhaps, then, the best cure for a fear of the dark is, well, getting used to the dark. So why can’t I face that fear? Truthfully, on my mile-long list of “Things to deal with in therapy,” this doesn’t crack the top 10. It’s concerning and something I’ll need to confront directly...yet it can be bandaged. Maybe I'll switch out my lamp for something more subtle, like an adult-friendly night light—maybe something gemstone-y . I just don’t want to shut off the brightness entirely. Because even if it’s counterproductive, the reality is that I don’t like being left in the dark. Alone. Defenseless. Wondering what kind of person I am, what kind of person I’ll be, and contending with the horrifying stream of consciousness running through my mind, not willing to quiet itself for sleep. I'm just lucky to have a partner who's willing to support me through this—even if doing so opens him up to the risk of the joining the dumbest internet challenge of the year when he wakes up at 4 a.m. to groggily go to the bathroom . Forget the dark— your ex and FOMO might also be culprits for stealing your sleep. Tags: Healthy Mind , Healthy Sleeping Habits , Mental Challenges Loading More Posts... Featured Collection The Beach Is My Happy Place—and Here Are 3 Science-Backed Reasons It Should Be Yours, Too Paid Content Evolve The Beach Is My Happy Place—and Here Are 3 Science-Backed Reasons It Should Be Yours, Too Your official excuse to add "OOD" (ahem, out of doors) to your cal. 4 Mistakes That Are Causing You to Waste Money on Skin-Care Serums, According to an Esthetician Skin-Care Tips 4 Mistakes That Are Causing You to Waste Money on Skin-Care Serums, According to an Esthetician These Are the Best Anti-Chafing Denim Shorts—According to Some Very Happy Reviewers Active Clothing These Are the Best Anti-Chafing Denim Shorts—According to Some Very Happy Reviewers Wellness In Your Inbox Well+Good helps you to pave your own personalized path to living well. Get inspired, on the daily. Subscribe Facebook Pinterest Twitter Youtube Instagram Well+Good About Contact Us Press Advertise Topics Skin-Care Tips Food and Nutrition Holistic Treatment Fitness Tips Relationship Tips For You Well+Good Shop Accessibility Statement California Notice of Collection © 2024 Well+Good LLC. All rights reserved. Read our Privacy Notice , Cookie Notice and Terms and Conditions . Close Close Close {{ customData.cancel_text }} {{ successMessage }} Please wait a moment... Close
Beauty Fitness Lifestyle Health Food Shopping Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe Search Search Button
Beauty Fitness Lifestyle Health Food Shopping Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics Become an Insider Subscribe
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear About Us Well+Good Podcast All Topics
Beauty Skin Care Hair Makeup Nails Fitness Workouts Yoga Running Recovery Interval Training Lifestyle Sex Relationships Career Astrology Travel Health Healthy Body Pregnancy Menstrual Health Gut Health Healthy Mind Food Nutrition Healthy Meals Shopping Sales Footwear Active Clothing Loungewear Fitness Gear
Yes, Nickelodeon, I’m a Grown Woman Who Is Afraid of the Dark and Sleeps With the Light On Mary Grace Garis Mary Grace Garis February 26, 2019 Pin It
Yes, Nickelodeon, I’m a Grown Woman Who Is Afraid of the Dark and Sleeps With the Light On Mary Grace Garis Mary Grace Garis February 26, 2019
Share on facebook Share on twitter Share on pinterest Share on email I think my nyctophobia turned my partner into Sandra Bullock in Bird Box . At least, that's who he most reminded me of the other night when I saw him wearing the sleep mask he uses as a shield from the light I desperately need to stay on all night. Without it, my anxiety soars sky high and my heart pounds. All of this is because—to answer that famed Nickelodeon-prompted question—I am indeed afraid of the dark. Or, maybe I'm just afraid of Samara from The Ring . I don’t find it a coincidence that I started sleeping with the light on after watching the film in 2002, but many find it puzzling that my fright has extended so many years. I've found that being an adult and simultaneously having a fear of the dark prompts so many questions from the few people who know about it, and the tonal subtext is often, “Why—and why would you ever share that information?” I get the judgment. See, nyctophobia , as Healthline has taught me with a not-so-necessary air of condescension, tends to dissipate with age. However, my fear really isn't that huge of an anomaly. One doc says an estimated 11 percent of the United States population is afraid of the dark , making it a more common phobia than a fear of heights. And unlike with heights, where the visual stimuli is responsible for the palpations, nyctophobia is more rooted in the anxiety of having impaired vision, invoking a feeling of defenselessness. The small lamp on my desk is typically enough to keep me calm and happy, but anything from a bad dream to a before-bed horror movie to even the Sunday Scaries can push me to light up the whole room. The more fearful I get, the more bulbs are necessary, as if illuminating my room is shrouding me from anxiety bogeymen. Related Stories {{ truncate(post.title, 12) }} {{post.sponsorText}} There were, once upon a time, some stipulations. I didn’t force my freshmen-year roommates to keep the light on or alert my random hookups that they needed to save me from the monster under the bed. Historically, having someone with me kept me from freaking, which is a real Freudian take on things. Old Sigmund thought a fear of the dark correlated with separation anxiety, writing in his General Introduction to Psychoanalysis , “I once heard a child, who was afraid of the dark, call into an adjoining room, ‘Auntie, talk to me, I am afraid.’ ‘But what good will that do to you? You cannot see me!’ Whereupon the child answered, ‘If someone speaks, it is brighter.’” Word, little dude. I found myself waking up in a full-force panic attack, terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. And so, for years, having a body in place of a night light made the room brighter until it...didn’t. When my S.O. and I went to the Catskills for his friend’s wedding, I found myself waking up in a full-force panic attack , terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. What’s especially fun and great about this is that it sure as hell cuts into the quality of my sleep. One small Ryerson University study on 93 college-aged men and women found more than half of the 42 who suffered from insomnia reported a fear of the dark . And it’s a real catch-22 because you know what helps you go to sleep? Um, the effing dark. Like, it’s why our iPhones keep us awake every night . According to the American Medical Association , blue or white nighttime light suppresses melatonin release and interrupts circadian biological rhythms. Perhaps, then, the best cure for a fear of the dark is, well, getting used to the dark. So why can’t I face that fear? Truthfully, on my mile-long list of “Things to deal with in therapy,” this doesn’t crack the top 10. It’s concerning and something I’ll need to confront directly...yet it can be bandaged. Maybe I'll switch out my lamp for something more subtle, like an adult-friendly night light—maybe something gemstone-y . I just don’t want to shut off the brightness entirely. Because even if it’s counterproductive, the reality is that I don’t like being left in the dark. Alone. Defenseless. Wondering what kind of person I am, what kind of person I’ll be, and contending with the horrifying stream of consciousness running through my mind, not willing to quiet itself for sleep. I'm just lucky to have a partner who's willing to support me through this—even if doing so opens him up to the risk of the joining the dumbest internet challenge of the year when he wakes up at 4 a.m. to groggily go to the bathroom . Forget the dark— your ex and FOMO might also be culprits for stealing your sleep. Tags: Healthy Mind , Healthy Sleeping Habits , Mental Challenges
I think my nyctophobia turned my partner into Sandra Bullock in Bird Box . At least, that's who he most reminded me of the other night when I saw him wearing the sleep mask he uses as a shield from the light I desperately need to stay on all night. Without it, my anxiety soars sky high and my heart pounds. All of this is because—to answer that famed Nickelodeon-prompted question—I am indeed afraid of the dark. Or, maybe I'm just afraid of Samara from The Ring . I don’t find it a coincidence that I started sleeping with the light on after watching the film in 2002, but many find it puzzling that my fright has extended so many years. I've found that being an adult and simultaneously having a fear of the dark prompts so many questions from the few people who know about it, and the tonal subtext is often, “Why—and why would you ever share that information?” I get the judgment. See, nyctophobia , as Healthline has taught me with a not-so-necessary air of condescension, tends to dissipate with age. However, my fear really isn't that huge of an anomaly. One doc says an estimated 11 percent of the United States population is afraid of the dark , making it a more common phobia than a fear of heights. And unlike with heights, where the visual stimuli is responsible for the palpations, nyctophobia is more rooted in the anxiety of having impaired vision, invoking a feeling of defenselessness. The small lamp on my desk is typically enough to keep me calm and happy, but anything from a bad dream to a before-bed horror movie to even the Sunday Scaries can push me to light up the whole room. The more fearful I get, the more bulbs are necessary, as if illuminating my room is shrouding me from anxiety bogeymen. Related Stories {{ truncate(post.title, 12) }} {{post.sponsorText}} There were, once upon a time, some stipulations. I didn’t force my freshmen-year roommates to keep the light on or alert my random hookups that they needed to save me from the monster under the bed. Historically, having someone with me kept me from freaking, which is a real Freudian take on things. Old Sigmund thought a fear of the dark correlated with separation anxiety, writing in his General Introduction to Psychoanalysis , “I once heard a child, who was afraid of the dark, call into an adjoining room, ‘Auntie, talk to me, I am afraid.’ ‘But what good will that do to you? You cannot see me!’ Whereupon the child answered, ‘If someone speaks, it is brighter.’” Word, little dude. I found myself waking up in a full-force panic attack, terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. And so, for years, having a body in place of a night light made the room brighter until it...didn’t. When my S.O. and I went to the Catskills for his friend’s wedding, I found myself waking up in a full-force panic attack , terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. What’s especially fun and great about this is that it sure as hell cuts into the quality of my sleep. One small Ryerson University study on 93 college-aged men and women found more than half of the 42 who suffered from insomnia reported a fear of the dark . And it’s a real catch-22 because you know what helps you go to sleep? Um, the effing dark. Like, it’s why our iPhones keep us awake every night . According to the American Medical Association , blue or white nighttime light suppresses melatonin release and interrupts circadian biological rhythms. Perhaps, then, the best cure for a fear of the dark is, well, getting used to the dark. So why can’t I face that fear? Truthfully, on my mile-long list of “Things to deal with in therapy,” this doesn’t crack the top 10. It’s concerning and something I’ll need to confront directly...yet it can be bandaged. Maybe I'll switch out my lamp for something more subtle, like an adult-friendly night light—maybe something gemstone-y . I just don’t want to shut off the brightness entirely. Because even if it’s counterproductive, the reality is that I don’t like being left in the dark. Alone. Defenseless. Wondering what kind of person I am, what kind of person I’ll be, and contending with the horrifying stream of consciousness running through my mind, not willing to quiet itself for sleep. I'm just lucky to have a partner who's willing to support me through this—even if doing so opens him up to the risk of the joining the dumbest internet challenge of the year when he wakes up at 4 a.m. to groggily go to the bathroom . Forget the dark— your ex and FOMO might also be culprits for stealing your sleep. Tags: Healthy Mind , Healthy Sleeping Habits , Mental Challenges
I think my nyctophobia turned my partner into Sandra Bullock in Bird Box . At least, that's who he most reminded me of the other night when I saw him wearing the sleep mask he uses as a shield from the light I desperately need to stay on all night. Without it, my anxiety soars sky high and my heart pounds. All of this is because—to answer that famed Nickelodeon-prompted question—I am indeed afraid of the dark. Or, maybe I'm just afraid of Samara from The Ring . I don’t find it a coincidence that I started sleeping with the light on after watching the film in 2002, but many find it puzzling that my fright has extended so many years. I've found that being an adult and simultaneously having a fear of the dark prompts so many questions from the few people who know about it, and the tonal subtext is often, “Why—and why would you ever share that information?” I get the judgment. See, nyctophobia , as Healthline has taught me with a not-so-necessary air of condescension, tends to dissipate with age. However, my fear really isn't that huge of an anomaly. One doc says an estimated 11 percent of the United States population is afraid of the dark , making it a more common phobia than a fear of heights. And unlike with heights, where the visual stimuli is responsible for the palpations, nyctophobia is more rooted in the anxiety of having impaired vision, invoking a feeling of defenselessness. The small lamp on my desk is typically enough to keep me calm and happy, but anything from a bad dream to a before-bed horror movie to even the Sunday Scaries can push me to light up the whole room. The more fearful I get, the more bulbs are necessary, as if illuminating my room is shrouding me from anxiety bogeymen. Related Stories {{ truncate(post.title, 12) }} {{post.sponsorText}} There were, once upon a time, some stipulations. I didn’t force my freshmen-year roommates to keep the light on or alert my random hookups that they needed to save me from the monster under the bed. Historically, having someone with me kept me from freaking, which is a real Freudian take on things. Old Sigmund thought a fear of the dark correlated with separation anxiety, writing in his General Introduction to Psychoanalysis , “I once heard a child, who was afraid of the dark, call into an adjoining room, ‘Auntie, talk to me, I am afraid.’ ‘But what good will that do to you? You cannot see me!’ Whereupon the child answered, ‘If someone speaks, it is brighter.’” Word, little dude. I found myself waking up in a full-force panic attack, terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. And so, for years, having a body in place of a night light made the room brighter until it...didn’t. When my S.O. and I went to the Catskills for his friend’s wedding, I found myself waking up in a full-force panic attack , terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare. What’s especially fun and great about this is that it sure as hell cuts into the quality of my sleep. One small Ryerson University study on 93 college-aged men and women found more than half of the 42 who suffered from insomnia reported a fear of the dark . And it’s a real catch-22 because you know what helps you go to sleep? Um, the effing dark. Like, it’s why our iPhones keep us awake every night . According to the American Medical Association , blue or white nighttime light suppresses melatonin release and interrupts circadian biological rhythms. Perhaps, then, the best cure for a fear of the dark is, well, getting used to the dark. So why can’t I face that fear? Truthfully, on my mile-long list of “Things to deal with in therapy,” this doesn’t crack the top 10. It’s concerning and something I’ll need to confront directly...yet it can be bandaged. Maybe I'll switch out my lamp for something more subtle, like an adult-friendly night light—maybe something gemstone-y . I just don’t want to shut off the brightness entirely. Because even if it’s counterproductive, the reality is that I don’t like being left in the dark. Alone. Defenseless. Wondering what kind of person I am, what kind of person I’ll be, and contending with the horrifying stream of consciousness running through my mind, not willing to quiet itself for sleep. I'm just lucky to have a partner who's willing to support me through this—even if doing so opens him up to the risk of the joining the dumbest internet challenge of the year when he wakes up at 4 a.m. to groggily go to the bathroom . Forget the dark— your ex and FOMO might also be culprits for stealing your sleep. Tags: Healthy Mind , Healthy Sleeping Habits , Mental Challenges
I think my nyctophobia turned my partner into Sandra Bullock in Bird Box . At least, that's who he most reminded me of the other night when I saw him wearing the sleep mask he uses as a shield from the light I desperately need to stay on all night. Without it, my anxiety soars sky high and my heart pounds. All of this is because—to answer that famed Nickelodeon-prompted question—I am indeed afraid of the dark.
Or, maybe I'm just afraid of Samara from The Ring . I don’t find it a coincidence that I started sleeping with the light on after watching the film in 2002, but many find it puzzling that my fright has extended so many years. I've found that being an adult and simultaneously having a fear of the dark prompts so many questions from the few people who know about it, and the tonal subtext is often, “Why—and why would you ever share that information?”
I get the judgment. See, nyctophobia , as Healthline has taught me with a not-so-necessary air of condescension, tends to dissipate with age. However, my fear really isn't that huge of an anomaly. One doc says an estimated 11 percent of the United States population is afraid of the dark , making it a more common phobia than a fear of heights.
And unlike with heights, where the visual stimuli is responsible for the palpations, nyctophobia is more rooted in the anxiety of having impaired vision, invoking a feeling of defenselessness. The small lamp on my desk is typically enough to keep me calm and happy, but anything from a bad dream to a before-bed horror movie to even the Sunday Scaries can push me to light up the whole room. The more fearful I get, the more bulbs are necessary, as if illuminating my room is shrouding me from anxiety bogeymen.
There were, once upon a time, some stipulations. I didn’t force my freshmen-year roommates to keep the light on or alert my random hookups that they needed to save me from the monster under the bed. Historically, having someone with me kept me from freaking, which is a real Freudian take on things. Old Sigmund thought a fear of the dark correlated with separation anxiety, writing in his General Introduction to Psychoanalysis , “I once heard a child, who was afraid of the dark, call into an adjoining room, ‘Auntie, talk to me, I am afraid.’ ‘But what good will that do to you? You cannot see me!’ Whereupon the child answered, ‘If someone speaks, it is brighter.’”
I found myself waking up in a full-force panic attack, terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare.
And so, for years, having a body in place of a night light made the room brighter until it...didn’t. When my S.O. and I went to the Catskills for his friend’s wedding, I found myself waking up in a full-force panic attack , terrified by the stillness surrounding me. Forget whatever I was wrestling with in the throes of REM; waking up in darkness is the real nightmare.
What’s especially fun and great about this is that it sure as hell cuts into the quality of my sleep. One small Ryerson University study on 93 college-aged men and women found more than half of the 42 who suffered from insomnia reported a fear of the dark . And it’s a real catch-22 because you know what helps you go to sleep? Um, the effing dark. Like, it’s why our iPhones keep us awake every night . According to the American Medical Association , blue or white nighttime light suppresses melatonin release and interrupts circadian biological rhythms.
Perhaps, then, the best cure for a fear of the dark is, well, getting used to the dark. So why can’t I face that fear? Truthfully, on my mile-long list of “Things to deal with in therapy,” this doesn’t crack the top 10. It’s concerning and something I’ll need to confront directly...yet it can be bandaged. Maybe I'll switch out my lamp for something more subtle, like an adult-friendly night light—maybe something gemstone-y . I just don’t want to shut off the brightness entirely.
Because even if it’s counterproductive, the reality is that I don’t like being left in the dark. Alone. Defenseless. Wondering what kind of person I am, what kind of person I’ll be, and contending with the horrifying stream of consciousness running through my mind, not willing to quiet itself for sleep. I'm just lucky to have a partner who's willing to support me through this—even if doing so opens him up to the risk of the joining the dumbest internet challenge of the year when he wakes up at 4 a.m. to groggily go to the bathroom .
Featured Collection The Beach Is My Happy Place—and Here Are 3 Science-Backed Reasons It Should Be Yours, Too Paid Content Evolve The Beach Is My Happy Place—and Here Are 3 Science-Backed Reasons It Should Be Yours, Too Your official excuse to add "OOD" (ahem, out of doors) to your cal. 4 Mistakes That Are Causing You to Waste Money on Skin-Care Serums, According to an Esthetician Skin-Care Tips 4 Mistakes That Are Causing You to Waste Money on Skin-Care Serums, According to an Esthetician These Are the Best Anti-Chafing Denim Shorts—According to Some Very Happy Reviewers Active Clothing These Are the Best Anti-Chafing Denim Shorts—According to Some Very Happy Reviewers
The Beach Is My Happy Place—and Here Are 3 Science-Backed Reasons It Should Be Yours, Too Paid Content Evolve The Beach Is My Happy Place—and Here Are 3 Science-Backed Reasons It Should Be Yours, Too Your official excuse to add "OOD" (ahem, out of doors) to your cal. 4 Mistakes That Are Causing You to Waste Money on Skin-Care Serums, According to an Esthetician Skin-Care Tips 4 Mistakes That Are Causing You to Waste Money on Skin-Care Serums, According to an Esthetician These Are the Best Anti-Chafing Denim Shorts—According to Some Very Happy Reviewers Active Clothing These Are the Best Anti-Chafing Denim Shorts—According to Some Very Happy Reviewers
Wellness In Your Inbox Well+Good helps you to pave your own personalized path to living well. Get inspired, on the daily. Subscribe Facebook Pinterest Twitter Youtube Instagram Well+Good About Contact Us Press Advertise Topics Skin-Care Tips Food and Nutrition Holistic Treatment Fitness Tips Relationship Tips For You Well+Good Shop
Wellness In Your Inbox Well+Good helps you to pave your own personalized path to living well. Get inspired, on the daily. Subscribe Facebook Pinterest Twitter Youtube Instagram
Wellness In Your Inbox Well+Good helps you to pave your own personalized path to living well. Get inspired, on the daily. Subscribe
Well+Good About Contact Us Press Advertise Topics Skin-Care Tips Food and Nutrition Holistic Treatment Fitness Tips Relationship Tips For You Well+Good Shop
Accessibility Statement California Notice of Collection © 2024 Well+Good LLC. All rights reserved. Read our Privacy Notice , Cookie Notice and Terms and Conditions .
Accessibility Statement California Notice of Collection © 2024 Well+Good LLC. All rights reserved. Read our Privacy Notice , Cookie Notice and Terms and Conditions .
Accessibility Statement California Notice of Collection © 2024 Well+Good LLC. All rights reserved. Read our Privacy Notice , Cookie Notice and Terms and Conditions .
© 2024 Well+Good LLC. All rights reserved. Read our Privacy Notice , Cookie Notice and Terms and Conditions .
© 2024 Well+Good LLC. All rights reserved. Read our Privacy Notice , Cookie Notice and Terms and Conditions . | biology | 207941 | https://no.wikipedia.org/wiki/Foggy%20Dew | Foggy Dew | «Foggy Dew» (eller «The Foggy Dew») er navnet på flere ballader.
Foggy, Foggy Dew
Den eldste versjonen, som også kalles «Foggy, Foggy Dew», er en sorgfull ballade om en ung elsker. Den ble publisert i England omkring 1815, men for de fleste er den kjent gjennom Burl Ives' versjon fra 1940-årene. Ives hevdet at sangen hadde sitt opphav i Amerika i kolonitiden, men det er ikke noen støtte for dette i kildene. Han ble engang arrestert i Mona i Utah for å synge den offentlig, ettersom myndighetene mente den var uanstendig.
Musikken er en versjon fra slutten av det 18. eller begynnelsen av det 19. århundre av «When I First Came to Court» fra 1689.
When I was a bachelor, I liv'd all alone
I worked at the weaver's trade
And the only, only thing that I did that was wrong
Was to woo a fair young maid.
I wooed her in the wintertime
Part of the summer, too
And the only, only thing that I did that was wrong
Was to keep her from the foggy, foggy dew.
One night she knelt close by my side
When I was fast asleep.
She threw her arms around my neck
And she began to weep.
She wept, she cried, she tore her hair
Ah, me! What could I do?
So all night long I held her in my arms
Just to keep her from the foggy foggy dew.
Again I am a bachelor, I live with my son
We work at the weaver's trade.
And every single time I look into his eyes
He reminds me of that fair young maid.
He reminds me of the wintertime
Part of the summer, too,
And the many, many times that I held her in my arms
Just to keep her from the foggy, foggy, dew.
En irsk versjon av sangen begynner med linjene:
When I was a bachelor, airy and young, I followed the roving trade,
And the only harm that ever I did was courting a servant maid.
I courted her all summer long, and part of the winter, too
And many's the time I rode my love all over the foggy dew.'Axel Schiøtz, tenor Herm D. Koppel piano akk. spilte den inn i København 7. mai 1951. Den ble utgitt på 78-platene His Master's Voice X 8009 og på His Master's Voice A.L. 3204. Arrangør var Benjamin Britten.
Irsk sørgevise
«Foggy Dew» er også en sørgevise fra Irland, av ukjent datering. Teksten nedenfor er hentet fra 1931-utgaven av The Home and Community Songbook.
Oh, a wan cloud was drawn o'er the dim weeping dawn
As to Shannon's side I return'd at last,
And the heart in my breast for the girl I lov'd best
Was beating, ah, beating, how loud and fast!
While the doubts and the fears of the long aching years
Seem'd mingling their voices with the moaning flood:
Till full in my path, like a wild water wraith,
My true love's shadow lamenting stood.
But the sudden sun kiss'd the cold, cruel mist
Into dancing show'rs of diamond dew,
And the dark flowing stream laugh'd back to his beam,
And the lark soared aloft in the blue;
While no phantom of night but a form of delight
Ran with arms outspread to her darling boy,
And the girl I love best on my wild throbbing breast
Hid her thousand treasures with cry of joy.
Påskeopprøret
En annen sang ved navn «Foggy Dew», også kjent som «Down the Glen», som kanskje har blitt den best kjente for mange, handler om påskeopprøret i Irland i 1916. Den er attribuert til Charles O'Neill. Det finnes ikke beviser for noen av attribueringene.
Teksten finnes i en rekke versjoner, og fremføres ofte i forkortet versjon. De fleste av de større irske folkemusikkgruppene har spilt den inn, som The Clancy Brothers and Tommy Makem, The Dubliners, The Chieftains (med Sinéad O'Connor som vokalist), Shane MacGowan, The Battering Ram, og Wolfe Tones.
As down the glen one Easter morn to a city fair rode I
There Armed lines of marching men in squadrons passed me by
No pipe did hum nor battle drum did sound its dread tattoo
But the Angelus bell o'er the Liffey's swell rang out through the foggy dew
Right proudly high over Dublin Town they hung out the flag of war Twas better to die 'neath an Irish sky than at Suvla or Sud-El-Bar
And from the plains of Royal Meath strong men came hurrying through
While Britannia's Huns, with their long range guns sailed in through the foggy dew
'T'was England bade our wild geese go, that "small nations might be free";
Their lonely graves are by Suvla's waves or the fringe of the great North Sea.
Oh, had they died by Pearse's side or fought with Cathal Brugha
Their graves we'd keep where the Fenians sleep, 'neath the shroud of the foggy dew.
Oh the night fell black, and the rifles' crack made perfidious Albion reel
In the leaden rain, seven tongues of flame did shine o'er the lines of steel
By each shining blade a prayer was said, that to Ireland her sons be true
But when morning broke, still the war flag shook out its folds the foggy dew
Oh the bravest fell, and the requiem bell rang mournfully and clear
For those who died that Eastertide in the spring time of the year
And the world did gaze, in deep amaze, at those fearless men, but few,
Who bore the fight that freedom's light might shine through the foggy dew
As back through the glen I rode again and my heart with grief was sore
For I parted then with valiant men whom I never shall see more
But to and fro in my dreams I go and I kneel and pray for you,
For slavery fled, O glorious dead, when you fell in the foggy dew.
Referanser
Irske sanger
Britiske sanger | norwegian_bokmål | 1.193555 |
human_fear_dark/fear-of-the-dark-in-children.txt |
Menu What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us View cart
Menu What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us View cart
Menu What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us
Menu What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us
What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us
What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us
What can we help you find? Home About System Features Buy Now News Media Resources FAQ Help with My Snorble Contact Us
Snorble® | Fear of the Dark in Children Resources December 13, 2022 Fear of the Dark in Children Fear of the dark is common in children. However, you can give your little one the tools necessary to confront that fear and help them grow out of it. Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights. These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep. What is fear of the dark called? Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear. What causes us to fear the darkness? Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved. Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off. Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night. When do kids start fearing the dark? Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD. For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious: Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark Can you help your little one overcome their fear of the dark? You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try: Having a conversation about the dark When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it. Offering security in the form of an object For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.) Training for relaxation Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia. Lighting the night with a nightlight Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights. Imagining a better darkness One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep. Making sure bravery doesn’t go unnoticed When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears. Snorble’s not in the dark about nyctophobia Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark. PREORDER NOW Sources: https://www.washingtonpost.com/health/2022/03/26/fear-of-dark-kids-tips/ https://www.whattoexpect.com/toddler/sleep/toddler-afraid-of-the-dark/ https://www.webmd.com/parenting/features/fear-of-the-dark https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/ https://my.clevelandclinic.org/health/diseases/22785-nyctophobia-fear-of-the-dark https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/fear-and-anxiety-children https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know Photo by Caleb Woods on Unsplash Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest $299 Limited quantities available. Buy Now Home Features Buy Now News Resources FAQ Media Careers Contact Us Privacy Terms Help Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad. Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Snorble® | Fear of the Dark in Children Resources December 13, 2022 Fear of the Dark in Children Fear of the dark is common in children. However, you can give your little one the tools necessary to confront that fear and help them grow out of it. Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights. These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep. What is fear of the dark called? Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear. What causes us to fear the darkness? Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved. Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off. Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night. When do kids start fearing the dark? Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD. For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious: Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark Can you help your little one overcome their fear of the dark? You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try: Having a conversation about the dark When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it. Offering security in the form of an object For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.) Training for relaxation Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia. Lighting the night with a nightlight Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights. Imagining a better darkness One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep. Making sure bravery doesn’t go unnoticed When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears. Snorble’s not in the dark about nyctophobia Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark. PREORDER NOW Sources: https://www.washingtonpost.com/health/2022/03/26/fear-of-dark-kids-tips/ https://www.whattoexpect.com/toddler/sleep/toddler-afraid-of-the-dark/ https://www.webmd.com/parenting/features/fear-of-the-dark https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/ https://my.clevelandclinic.org/health/diseases/22785-nyctophobia-fear-of-the-dark https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/fear-and-anxiety-children https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know Photo by Caleb Woods on Unsplash Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest $299 Limited quantities available. Buy Now Home Features Buy Now News Resources FAQ Media Careers Contact Us Privacy Terms Help Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad. Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Snorble® | Fear of the Dark in Children Resources December 13, 2022 Fear of the Dark in Children Fear of the dark is common in children. However, you can give your little one the tools necessary to confront that fear and help them grow out of it. Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights. These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep. What is fear of the dark called? Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear. What causes us to fear the darkness? Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved. Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off. Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night. When do kids start fearing the dark? Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD. For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious: Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark Can you help your little one overcome their fear of the dark? You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try: Having a conversation about the dark When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it. Offering security in the form of an object For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.) Training for relaxation Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia. Lighting the night with a nightlight Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights. Imagining a better darkness One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep. Making sure bravery doesn’t go unnoticed When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears. Snorble’s not in the dark about nyctophobia Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark. PREORDER NOW Sources: https://www.washingtonpost.com/health/2022/03/26/fear-of-dark-kids-tips/ https://www.whattoexpect.com/toddler/sleep/toddler-afraid-of-the-dark/ https://www.webmd.com/parenting/features/fear-of-the-dark https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/ https://my.clevelandclinic.org/health/diseases/22785-nyctophobia-fear-of-the-dark https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/fear-and-anxiety-children https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know Photo by Caleb Woods on Unsplash Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest $299 Limited quantities available. Buy Now
Snorble® | Fear of the Dark in Children Resources December 13, 2022 Fear of the Dark in Children Fear of the dark is common in children. However, you can give your little one the tools necessary to confront that fear and help them grow out of it.
Fear of the dark is common in children. However, you can give your little one the tools necessary to confront that fear and help them grow out of it.
Fear of the dark is common in children. However, you can give your little one the tools necessary to confront that fear and help them grow out of it.
Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights. These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep. What is fear of the dark called? Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear. What causes us to fear the darkness? Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved. Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off. Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night. When do kids start fearing the dark? Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD. For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious: Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark Can you help your little one overcome their fear of the dark? You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try: Having a conversation about the dark When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it. Offering security in the form of an object For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.) Training for relaxation Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia. Lighting the night with a nightlight Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights. Imagining a better darkness One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep. Making sure bravery doesn’t go unnoticed When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears. Snorble’s not in the dark about nyctophobia Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark. PREORDER NOW Sources: https://www.washingtonpost.com/health/2022/03/26/fear-of-dark-kids-tips/ https://www.whattoexpect.com/toddler/sleep/toddler-afraid-of-the-dark/ https://www.webmd.com/parenting/features/fear-of-the-dark https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/ https://my.clevelandclinic.org/health/diseases/22785-nyctophobia-fear-of-the-dark https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/fear-and-anxiety-children https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know Photo by Caleb Woods on Unsplash
Share Share Share on Facebook Tweet Tweet on Twitter Pin it Pin on Pinterest It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights. These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep. What is fear of the dark called? Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear. What causes us to fear the darkness? Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved. Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off. Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night. When do kids start fearing the dark? Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD. For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious: Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark Can you help your little one overcome their fear of the dark? You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try: Having a conversation about the dark When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it. Offering security in the form of an object For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.) Training for relaxation Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia. Lighting the night with a nightlight Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights. Imagining a better darkness One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep. Making sure bravery doesn’t go unnoticed When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears. Snorble’s not in the dark about nyctophobia Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark. PREORDER NOW Sources: https://www.washingtonpost.com/health/2022/03/26/fear-of-dark-kids-tips/ https://www.whattoexpect.com/toddler/sleep/toddler-afraid-of-the-dark/ https://www.webmd.com/parenting/features/fear-of-the-dark https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/ https://my.clevelandclinic.org/health/diseases/22785-nyctophobia-fear-of-the-dark https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/fear-and-anxiety-children https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know Photo by Caleb Woods on Unsplash
It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights. These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep. What is fear of the dark called? Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear. What causes us to fear the darkness? Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved. Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off. Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night. When do kids start fearing the dark? Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD. For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious: Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark Can you help your little one overcome their fear of the dark? You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try: Having a conversation about the dark When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it. Offering security in the form of an object For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.) Training for relaxation Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia. Lighting the night with a nightlight Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights. Imagining a better darkness One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep. Making sure bravery doesn’t go unnoticed When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears. Snorble’s not in the dark about nyctophobia Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark. PREORDER NOW Sources: https://www.washingtonpost.com/health/2022/03/26/fear-of-dark-kids-tips/ https://www.whattoexpect.com/toddler/sleep/toddler-afraid-of-the-dark/ https://www.webmd.com/parenting/features/fear-of-the-dark https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/ https://my.clevelandclinic.org/health/diseases/22785-nyctophobia-fear-of-the-dark https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/fear-and-anxiety-children https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know Photo by Caleb Woods on Unsplash
It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights.
It’s bedtime. You’re about to tuck your little one in when they hit you with this, “There’s a monster in my closet.” You open the closet door to show them that there’s nothing in there except for their clothing. Then there’s a monster under the bed. Once you’ve verified that there’s nothing under the bed, there’s another monster and another. Each monster delays the inevitable shutting off the lights.
These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep.
These monsters aren’t actually monsters and it’s not going to sleep that your kiddo fears, it’s the dark. Fear of the dark is common in children and with your love and help, and a little night support from our smart companion, Snorble®, your youngster can face their fears and get a good night’s sleep.
Fear of the dark is called nyctophobia . It comes from the Greek word for night and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear.
and encompasses the anxiety and symptoms that go along with being scared of being alone in the dark and darkness itself. Nyctophobia is more common in children, however, people of all ages can be plagued by this fear.
Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved.
Back, back, back, back, back, back, back in the day, our ancestors lived and slept out in the open making darkness very dangerous. Predators roamed the night and you never knew who or what threatened your safety. Because our ancestors had to be vigilante even at night, this awareness of the unknown has stayed with us as we evolved.
Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the importance of sleep on their overall well-being and development , they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off.
Moreover, a person who has a troubling or traumatic experience in the dark is more likely to develop a fear of it. Memories of that traumatic event (or any traumatic event for that matter) can often return when we’re ready in bed and know the lights have to turn off so we can get some sleep. As children are learning about bedtime and the
, they’re also dealing with a fear of the unknown. To a child, the world is a big, wondrous place full of things they don’t know, especially when the lights go off.
Fear of the dark in toddlers and preschoolers can be brought on by noises they hear when the lights go out, especially when they have no idea where the noises are coming from or what caused them. In addition, kids are highly influenced by what they see. If they walk in on an older sibling watching a horror movie and catch a glimpse of something unsettling, this could turn into nyctophobia. They could also overhear their parents and caregivers talking about a horrible current event that they don’t understand but can grasp enough to know it’s bad news. Kiddos are creative and have wonderful imaginations but that creativity and imaginative curiosity can lead to phobias at night.
Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD.
Generally, children begin to fear the dark around age two. This fear can last anywhere from a few weeks to a few months to a few years depending on your kiddo’s genetic makeup, if they have experienced a traumatic event, or have an underlying issue that hasn’t been addressed like ADD, PTSD, and OCD.
For anyone of any age, being alone in the dark can feel uncomfortable due to uncertainty. What’s that noise? Who else is in here? However, for children that are nyctophobic there are specific triggers beyond turning the lights off at night that can make them anxious:
Getting ready for bed - the simple act of putting their PJs on and brushing their teeth can make some youngsters uneasy knowing that soon they’ll be in the dark trying to sleep
Walking into a dark place, even during the day - movie theaters, going down to the basement, or any darkened place where a child can’t reach the light switch can be terrifying for them
Watching a movie or TV show with nighttime scenes - even watching a cartoon where the action takes place at night can provoke their fear of the dark
You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try:
You sure can! Besides telling your youngster that you’re not going anywhere while they’re sleeping and even popping your head into their room a few times as they’re dozing off to comfort them, you can try:
When you acknowledge your child’s fear of the dark , you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it.
, you’re telling them that this is a real fear and you understand that they’re having a tough time with it. If you ignore it or tease them, even in a playful manner, this may magnify the fear and make it worse. Instead of saying things like, “Big kids aren’t scared of the dark.” you can instead say something like, “Tell me what’s scaring you so we can talk about it.” This way you’re giving your child a chance to unpack their fear and you can get to the bottom of it.
For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.)
For children trying to cope with the darkness, a security object like a blanket or stuffed animal can help them feel more relaxed and secure during the night. Should they wake up in the middle of the night in a panic because it’s too dark to see, they can hug their security object for comfort. You can also tell your kiddo to talk to their security object for reassurance even if they don’t get a response. Snorble can be used as a security object and your child will get a response when they have a conversation in the middle of the night. (More on that below.)
Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia.
Yoga, meditation, and reflection, are all great ways to train your child to relax before bed. Snorble offers those activities as part of the Bedtime Experience wind-down component. By showing your kiddo how to close their eyes and focus on their breathing, they may eventually learn to do that on their own when faced with nyctophobia.
Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using a nightlight offers security for your child and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights.
Nightlights can help children get over their fear of the dark as long as they’re not exposed to bright blue light while they’re trying to sleep. Using
and allows them to see enough of what’s around them without illuminating the entire room. Snorble’s a nightlight too so you can leave our cuddly smart companion on while your child sleeps to bathe their room in low ambient lights.
One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep.
One of the most precious things about children is their imagination. You can imagine a better darkness with them by playing in the dark with glow-in-the-dark toys. You can also let their imaginations run wild by naming the monsters and boogeymen they think are hiding in the shadows. With a backstory, the scary ogre in their closet can turn into a loving mythical creature that protects them from harm while they sleep.
When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears.
When your little one confronts their fear of the dark, whether it’s sleeping through the night or not asking you to check the closet for monsters before bed, you can give them a little reward. This will not only make them feel good about themselves but give them an incentive to stand up to the darkness and face their fears.
Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark.
Here at Team Snorble, we know first-hand how fear of the dark can keep your little one from getting a good night’s sleep. We were kids once and we now face nyctophobia with our children. But there’s hope! Snorble can act as a nightlight and comes with an entirely customizable Bedtime Experience designed to get your kiddo into bed, fall asleep, and stay asleep. Your youngster can even cuddle Snorble during the night as a security object to protect them from the invisible menaces they think are lurking in the dark.
https://www.parents.com/toddlers-preschoolers/development/fear/how-to-help-your-kid-overcome-their-fear-of-the-dark/
https://www.variationspsychology.com/blogs/why-are-kids-so-afraid-of-the-dark-11-things-parents-should-know
Home Features Buy Now News Resources FAQ Media Careers Contact Us Privacy Terms Help Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad. Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Home Features Buy Now News Resources FAQ Media Careers Contact Us Privacy Terms Help Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad. Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Home Features Buy Now News Resources FAQ Media Careers Contact Us Privacy Terms Help Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad. Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad. Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Sign-up to our newsletter to get exclusive offers and more! Subscribe We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad.
We won’t spam you with calls or texts. When you give us your phone number, we’ll only message you with exciting offers, information about new content and educational games, and the occasional survey. You can opt out at any time! We won’t be mad.
Follow us on social Facebook Twitter Pinterest Instagram YouTube LinkedIn TikTok Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Snorble® and Snorbles® are trademarks of Snorble Incorporated. Patent pending - All rights reserved. Copyright © 2019 - 2024, Snorble Incorporated.
Close Cart Shopping Cart Your cart is currently empty. Enable cookies to use the shopping cart t: e (- ) Remove Item Quantity Discount: - Subtotal $0.00 Taxes and shipping calculated at checkout Checkout
Your cart is currently empty. Enable cookies to use the shopping cart t: e (- ) Remove Item Quantity
Your cart is currently empty. Enable cookies to use the shopping cart t: e (- ) Remove Item Quantity
t: e (- ) Remove Item Quantity
t: e (- ) Remove Item Quantity
t: e (- ) Remove Item Quantity | biology | 695467 | https://sv.wikipedia.org/wiki/Nattskr%C3%A4ck | Nattskräck | Nattskräck är ett fenomen som räknas till parasomnierna. Det förekommer framför allt hos små barn, där det är mycket vanligt. Det förekommer också som symtom vid psykiska störningar hos vuxna. Till skillnad från mardrömmar, minns personen inte några obehagliga drömmar efter uppvaknandet.
Nattskräck utgörs av attacker under sömn med ett beteende som liknar det vid stark ångest. Personen, som fortfarande delvis är i sömn, kan sätta sig upp, skrika, vara våldsam och tröstlös, och uppvisar tecken på autonom aktivering i form av ökad motorik, kallsvettningar, hög puls och blekhet. Ofta har personen glömt episoden morgonen efter, medan andra kan ha diffusa minnesbilder.
Ibland är attackerna förenade med sömngång. Diagnostiskt är detta två skilda fenomen, men i praktiken kan en person ha ett mellanting med drag av båda. Nattskräck uppkommer som regel under sömnens första 2-3 timmar, men alltid under NREM-sömn, den sömnfas som inte är REM-sömn, detta till skillnad från mardrömmar som istället uppkommer under REM.
Hos barn börjar nattskräck vanligen i åldern 3 till 12 år. Prognosen för barn är godartad och växer vanligtvis bort. Ickefarmakologisk behandling går ut på att avlägsna potentiellt skadliga föremål från patientens rum, låsa fönster och dörrar samt undvika sömnbrist. Vid mer svårartade besvär kan läkemedelsbehandling bli aktuell. Kirurgisk behandling, genom att adenoiden opereras bort, kan också ge goda resultat.
Vuxnas nattskräck startar vanligtvis i åldrarna 20 till 30 år. Nattskräck hos vuxna förekommer vid posttraumatiskt stressyndrom (under REM-sömn), hos suicidala personer, hos personer med neuroticism, och vid andra psykiska störningar samt ofta tillsammans med andra sömnstörningar. Hos barn har det ärftliga orsaker och har med den biologiska utvecklingen av hjärnan att göra.
Noter
Sömnstörningar
Barnpsykiatriska störningar | swedish | 0.754397 |
human_fear_dark/help-my-child-is-afraid-of-the-dark.txt |
Search Checkout Contact Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ? 12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business CART Your Cart is currently empty!
Search Checkout Contact Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ? 12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business CART Your Cart is currently empty!
Search Checkout Contact Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
Search Checkout Contact Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
Search Checkout Contact Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
Checkout Contact Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
Account Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
Login Email Password Forgot your password? Sign in Create account Reset your password We will send you an email to reset your password Email Submit Cancel Create account First name Last name Email Password Create Already Have an Account ?
12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business
12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business
12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business
12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business
12 Months Warranty Giving you peace of mind with every purchase Proudly Aussie Family-owned small business
Shop All Dream Guardians Dream Lights Miffy & Friends SALE Personalised Lights NEW Blog FAQ About Contact 0 item
Shop All Dream Guardians Dream Lights Miffy & Friends SALE Personalised Lights NEW Blog FAQ About Contact 0 item
Shop All Dream Guardians Dream Lights Miffy & Friends SALE Personalised Lights NEW Blog FAQ About Contact 0 item
Shop All Dream Guardians Dream Lights Miffy & Friends SALE Personalised Lights NEW Blog FAQ About Contact 0 item
Shop All Dream Guardians Dream Lights Miffy & Friends SALE Personalised Lights NEW Blog FAQ About Contact 0 item
New Product New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Special Product Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per We’ve talked about children’s fear of the dark before, but this is such an ongoing problem for many of us, so we thought we’d revisit it. After all, sleep deprivation is no one’s friend! In a study by Muris et al 2001 children aged 4 -12 and their parents were interviewed about children's night time fears. Results showed that 73.3% of the children reported night time fears, these fears were common among 4 to 6 year olds, became even more frequent in 7 to 9 year olds and then remained relatively stable in 10 to 12 year-olds. My personal experience is in line with the study, my 12 year old son seems to be ok at night now, but my 9 year old daughter is still afraid, although she is getting better. Why do children fear the dark? Kids have the most amazing imaginations and can develop a fear of pretty much anything. (my daughter is currently afraid when the home phone rings?!) A toddler or child in Kindy can be afraid of unfamiliar things or things out of their control. They don’t have the life experience to explain things, so they turn to the land of magic 4 year old logic, where monsters are real and dragons are living in the trees just outside the window. As children get older, they fear what the darkness masks and the threats they are unable to spot. They also start to think about real life dangers, like intruders or being taken. How can I help my child? From my experience, the best way to get over fear or anxiety is to get more information about it and then problem solve. Here are other ways you can help: Their fear is real. Do not dismiss it or tell them to stop being silly. DO NOT make fun of them. Encourage them to talk and really listen, understand exactly what they are afraid of. If a toy or other object is casting a scary shadow, turn the light on, show them and then remove the object before turning the light off. If they are worried about an intruder, show them that the doors and windows are locked. Reassure them that they are safe Their fear could be about something else, like being separated from you or anxious about something. Talk to them honestly. Ask them for suggestions, what would make them feel safe? Remain calm! Practical night time solutions Establish a bedtime routine, predictability helps reduce anxiety. This will help you to start the evening calm and relaxed. This sounds obvious but scary stories, television, social media are a no-go. Remember, children find different things scary, even the news could frighten them. (My 12 year old son finds that the news makes him anxious at times, especially when it is about war.) Put a night light in their room. If they’re older get one that they can control. (this will help to lessen their fear.) Try to look for a light with a remote and a warm light source – no bright white or blue light. We recommend red for overnight use as it is the only colour that doesn’t interfere with the sleepy hormone Melatonin. Be pro-active, look around their room for anything that may cast scary shadows and move it. Help them to choose a special protector to keep them safe at night – a cuddly friend perhaps or even an animal shaped night light . Whatever works for your child. Try playing with glow products or animals in the dark to give them positive experiences in darkness. Relaxation techniques and sounds before sleep really work well Be consistent, do not give up. This could take a while. Don’t forget, you are absolutely not going through this alone. Most of us have been there. And please do remember to applaud your little ones’ bravery as they progress, because they are incredibly brave at times, especially when their fears are so real to them. If things are not getting better and you are worried in anyway, please contact your doctor. *these are the opinions of the My Night Light team and are solutions we have found to work for our children. We are not medical professionals. Leave a comment Name * Email * Comment * Please note, comments need to be approved before they are published.
New Product New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Special Product Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per We’ve talked about children’s fear of the dark before, but this is such an ongoing problem for many of us, so we thought we’d revisit it. After all, sleep deprivation is no one’s friend! In a study by Muris et al 2001 children aged 4 -12 and their parents were interviewed about children's night time fears. Results showed that 73.3% of the children reported night time fears, these fears were common among 4 to 6 year olds, became even more frequent in 7 to 9 year olds and then remained relatively stable in 10 to 12 year-olds. My personal experience is in line with the study, my 12 year old son seems to be ok at night now, but my 9 year old daughter is still afraid, although she is getting better. Why do children fear the dark? Kids have the most amazing imaginations and can develop a fear of pretty much anything. (my daughter is currently afraid when the home phone rings?!) A toddler or child in Kindy can be afraid of unfamiliar things or things out of their control. They don’t have the life experience to explain things, so they turn to the land of magic 4 year old logic, where monsters are real and dragons are living in the trees just outside the window. As children get older, they fear what the darkness masks and the threats they are unable to spot. They also start to think about real life dangers, like intruders or being taken. How can I help my child? From my experience, the best way to get over fear or anxiety is to get more information about it and then problem solve. Here are other ways you can help: Their fear is real. Do not dismiss it or tell them to stop being silly. DO NOT make fun of them. Encourage them to talk and really listen, understand exactly what they are afraid of. If a toy or other object is casting a scary shadow, turn the light on, show them and then remove the object before turning the light off. If they are worried about an intruder, show them that the doors and windows are locked. Reassure them that they are safe Their fear could be about something else, like being separated from you or anxious about something. Talk to them honestly. Ask them for suggestions, what would make them feel safe? Remain calm! Practical night time solutions Establish a bedtime routine, predictability helps reduce anxiety. This will help you to start the evening calm and relaxed. This sounds obvious but scary stories, television, social media are a no-go. Remember, children find different things scary, even the news could frighten them. (My 12 year old son finds that the news makes him anxious at times, especially when it is about war.) Put a night light in their room. If they’re older get one that they can control. (this will help to lessen their fear.) Try to look for a light with a remote and a warm light source – no bright white or blue light. We recommend red for overnight use as it is the only colour that doesn’t interfere with the sleepy hormone Melatonin. Be pro-active, look around their room for anything that may cast scary shadows and move it. Help them to choose a special protector to keep them safe at night – a cuddly friend perhaps or even an animal shaped night light . Whatever works for your child. Try playing with glow products or animals in the dark to give them positive experiences in darkness. Relaxation techniques and sounds before sleep really work well Be consistent, do not give up. This could take a while. Don’t forget, you are absolutely not going through this alone. Most of us have been there. And please do remember to applaud your little ones’ bravery as they progress, because they are incredibly brave at times, especially when their fears are so real to them. If things are not getting better and you are worried in anyway, please contact your doctor. *these are the opinions of the My Night Light team and are solutions we have found to work for our children. We are not medical professionals. Leave a comment Name * Email * Comment * Please note, comments need to be approved before they are published.
New Product New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Special Product Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
New Product New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Special Product Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
New Product New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Special Product Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
New Product New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
New Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Special Product Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Special Product Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
Example product title Example product title Vendor: Vendor Regular price $19.99 Regular price Sale price $19.99 Unit price / per
We’ve talked about children’s fear of the dark before, but this is such an ongoing problem for many of us, so we thought we’d revisit it. After all, sleep deprivation is no one’s friend! In a study by Muris et al 2001 children aged 4 -12 and their parents were interviewed about children's night time fears. Results showed that 73.3% of the children reported night time fears, these fears were common among 4 to 6 year olds, became even more frequent in 7 to 9 year olds and then remained relatively stable in 10 to 12 year-olds. My personal experience is in line with the study, my 12 year old son seems to be ok at night now, but my 9 year old daughter is still afraid, although she is getting better. Why do children fear the dark? Kids have the most amazing imaginations and can develop a fear of pretty much anything. (my daughter is currently afraid when the home phone rings?!) A toddler or child in Kindy can be afraid of unfamiliar things or things out of their control. They don’t have the life experience to explain things, so they turn to the land of magic 4 year old logic, where monsters are real and dragons are living in the trees just outside the window. As children get older, they fear what the darkness masks and the threats they are unable to spot. They also start to think about real life dangers, like intruders or being taken. How can I help my child? From my experience, the best way to get over fear or anxiety is to get more information about it and then problem solve. Here are other ways you can help: Their fear is real. Do not dismiss it or tell them to stop being silly. DO NOT make fun of them. Encourage them to talk and really listen, understand exactly what they are afraid of. If a toy or other object is casting a scary shadow, turn the light on, show them and then remove the object before turning the light off. If they are worried about an intruder, show them that the doors and windows are locked. Reassure them that they are safe Their fear could be about something else, like being separated from you or anxious about something. Talk to them honestly. Ask them for suggestions, what would make them feel safe? Remain calm! Practical night time solutions Establish a bedtime routine, predictability helps reduce anxiety. This will help you to start the evening calm and relaxed. This sounds obvious but scary stories, television, social media are a no-go. Remember, children find different things scary, even the news could frighten them. (My 12 year old son finds that the news makes him anxious at times, especially when it is about war.) Put a night light in their room. If they’re older get one that they can control. (this will help to lessen their fear.) Try to look for a light with a remote and a warm light source – no bright white or blue light. We recommend red for overnight use as it is the only colour that doesn’t interfere with the sleepy hormone Melatonin. Be pro-active, look around their room for anything that may cast scary shadows and move it. Help them to choose a special protector to keep them safe at night – a cuddly friend perhaps or even an animal shaped night light . Whatever works for your child. Try playing with glow products or animals in the dark to give them positive experiences in darkness. Relaxation techniques and sounds before sleep really work well Be consistent, do not give up. This could take a while. Don’t forget, you are absolutely not going through this alone. Most of us have been there. And please do remember to applaud your little ones’ bravery as they progress, because they are incredibly brave at times, especially when their fears are so real to them. If things are not getting better and you are worried in anyway, please contact your doctor. *these are the opinions of the My Night Light team and are solutions we have found to work for our children. We are not medical professionals. Leave a comment Name * Email * Comment * Please note, comments need to be approved before they are published.
We’ve talked about children’s fear of the dark before, but this is such an ongoing problem for many of us, so we thought we’d revisit it. After all, sleep deprivation is no one’s friend! In a study by Muris et al 2001 children aged 4 -12 and their parents were interviewed about children's night time fears. Results showed that 73.3% of the children reported night time fears, these fears were common among 4 to 6 year olds, became even more frequent in 7 to 9 year olds and then remained relatively stable in 10 to 12 year-olds. My personal experience is in line with the study, my 12 year old son seems to be ok at night now, but my 9 year old daughter is still afraid, although she is getting better. Why do children fear the dark? Kids have the most amazing imaginations and can develop a fear of pretty much anything. (my daughter is currently afraid when the home phone rings?!) A toddler or child in Kindy can be afraid of unfamiliar things or things out of their control. They don’t have the life experience to explain things, so they turn to the land of magic 4 year old logic, where monsters are real and dragons are living in the trees just outside the window. As children get older, they fear what the darkness masks and the threats they are unable to spot. They also start to think about real life dangers, like intruders or being taken. How can I help my child? From my experience, the best way to get over fear or anxiety is to get more information about it and then problem solve. Here are other ways you can help: Their fear is real. Do not dismiss it or tell them to stop being silly. DO NOT make fun of them. Encourage them to talk and really listen, understand exactly what they are afraid of. If a toy or other object is casting a scary shadow, turn the light on, show them and then remove the object before turning the light off. If they are worried about an intruder, show them that the doors and windows are locked. Reassure them that they are safe Their fear could be about something else, like being separated from you or anxious about something. Talk to them honestly. Ask them for suggestions, what would make them feel safe? Remain calm! Practical night time solutions Establish a bedtime routine, predictability helps reduce anxiety. This will help you to start the evening calm and relaxed. This sounds obvious but scary stories, television, social media are a no-go. Remember, children find different things scary, even the news could frighten them. (My 12 year old son finds that the news makes him anxious at times, especially when it is about war.) Put a night light in their room. If they’re older get one that they can control. (this will help to lessen their fear.) Try to look for a light with a remote and a warm light source – no bright white or blue light. We recommend red for overnight use as it is the only colour that doesn’t interfere with the sleepy hormone Melatonin. Be pro-active, look around their room for anything that may cast scary shadows and move it. Help them to choose a special protector to keep them safe at night – a cuddly friend perhaps or even an animal shaped night light . Whatever works for your child. Try playing with glow products or animals in the dark to give them positive experiences in darkness. Relaxation techniques and sounds before sleep really work well Be consistent, do not give up. This could take a while. Don’t forget, you are absolutely not going through this alone. Most of us have been there. And please do remember to applaud your little ones’ bravery as they progress, because they are incredibly brave at times, especially when their fears are so real to them. If things are not getting better and you are worried in anyway, please contact your doctor. *these are the opinions of the My Night Light team and are solutions we have found to work for our children. We are not medical professionals.
We’ve talked about children’s fear of the dark before, but this is such an ongoing problem for many of us, so we thought we’d revisit it. After all, sleep deprivation is no one’s friend!
In a study by Muris et al 2001 children aged 4 -12 and their parents were interviewed about children's night time fears. Results showed that 73.3% of the children reported night time fears, these fears were common among 4 to 6 year olds, became even more frequent in 7 to 9 year olds and then remained relatively stable in 10 to 12 year-olds.
My personal experience is in line with the study, my 12 year old son seems to be ok at night now, but my 9 year old daughter is still afraid, although she is getting better.
Kids have the most amazing imaginations and can develop a fear of pretty much anything. (my daughter is currently afraid when the home phone rings?!) A toddler or child in Kindy can be afraid of unfamiliar things or things out of their control. They don’t have the life experience to explain things, so they turn to the land of magic 4 year old logic, where monsters are real and dragons are living in the trees just outside the window.
As children get older, they fear what the darkness masks and the threats they are unable to spot. They also start to think about real life dangers, like intruders or being taken.
From my experience, the best way to get over fear or anxiety is to get more information about it and then problem solve. Here are other ways you can help:
Don’t forget, you are absolutely not going through this alone. Most of us have been there. And please do remember to applaud your little ones’ bravery as they progress, because they are incredibly brave at times, especially when their fears are so real to them.
*these are the opinions of the My Night Light team and are solutions we have found to work for our children. We are not medical professionals.
Leave a comment Name * Email * Comment * Please note, comments need to be approved before they are published.
Leave a comment Name * Email * Comment * Please note, comments need to be approved before they are published.
My Night Light PTY LTD Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents. Quick Links Quick Links Contact Us FAQs Safety Concerns Shipping Information Returns Policy Your Privacy Shop Shop Bedtime Story Books Red Light for Sleep Nursery Night Lights Gift Card Shop All Lights Social Follows Social Follows Facebook Instagram TikTok Contact Information Contact Information 90 Hezlett Rd, Kellyville NSW Australia 1300901590 [email protected] © My Night Light PTY LTD 2018. All Rights Reserved. Powered by Queper Apple Pay Google Pay Visa Mastercard Discover
My Night Light PTY LTD Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents. Quick Links Quick Links Contact Us FAQs Safety Concerns Shipping Information Returns Policy Your Privacy Shop Shop Bedtime Story Books Red Light for Sleep Nursery Night Lights Gift Card Shop All Lights Social Follows Social Follows Facebook Instagram TikTok Contact Information Contact Information 90 Hezlett Rd, Kellyville NSW Australia 1300901590 [email protected]
My Night Light PTY LTD Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents. Quick Links Quick Links Contact Us FAQs Safety Concerns Shipping Information Returns Policy Your Privacy Shop Shop Bedtime Story Books Red Light for Sleep Nursery Night Lights Gift Card Shop All Lights Social Follows Social Follows Facebook Instagram TikTok Contact Information Contact Information 90 Hezlett Rd, Kellyville NSW Australia 1300901590 [email protected]
My Night Light PTY LTD Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents.
My Night Light PTY LTD Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents.
My Night Light PTY LTD Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents.
Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents.
Founded in 2009, My Night Light started out as an idea in the early hours of the morning amidst the cries of a screaming child. Our baby and we, found our peace in these night lights amongst darkness, exhaustion, and fear. We understood the power of a gentle, comforting glow in the darkness and that's when we decided to share that comfort with all parents.
Contact Information Contact Information 90 Hezlett Rd, Kellyville NSW Australia 1300901590 [email protected]
© My Night Light PTY LTD 2018. All Rights Reserved. Powered by Queper Apple Pay Google Pay Visa Mastercard Discover
© My Night Light PTY LTD 2018. All Rights Reserved. Powered by Queper Apple Pay Google Pay Visa Mastercard Discover
© My Night Light PTY LTD 2018. All Rights Reserved. Powered by Queper Apple Pay Google Pay Visa Mastercard Discover | biology | 3482749 | https://sv.wikipedia.org/wiki/Pristomyrmex%20wilsoni | Pristomyrmex wilsoni | Pristomyrmex wilsoni är en myrart som beskrevs av Taylor 1968. Pristomyrmex wilsoni ingår i släktet Pristomyrmex och familjen myror. Inga underarter finns listade i Catalogue of Life.
Källor
Externa länkar
Myror
wilsoni | swedish | 1.383937 |
birches_white_bark/whitebarkpineprotect.txt | [  ](https://biologicaldiversity.org)
For Immediate Release, December 14, 2022
Contact:
|
Noah Greenwald, (503) 484-7495, [email protected]
---|---
Whitebark Pine Protected as Threatened Under Endangered Species Act
_Pine Imperiled in Seven Western States by Disease, Climate Change_
PORTLAND, _Ore._ — The U.S. Fish and Wildlife Service [ announced today
](https://public-inspection.federalregister.gov/2022-27087.pdf) that the
whitebark pine will be listed as threatened under the Endangered Species Act.
The pine is the most widespread tree to receive such protection. It occurs in
high-elevation areas of Washington, Oregon, California, Idaho, Montana,
Wyoming and Nevada.
The tree is primarily threatened by an introduced fungal pathogen known as
white pine blister rust. It is increasingly threatened by climate change,
which is leading to increased fires and mountain pine beetle outbreaks.
Like most imperiled species, the tree is also threatened by habitat
destruction. The whitebark pine is threatened specifically by development for
winter recreation. Despite this additional threat, the Service declined to
designate protected critical habitat.
“It’s just incredibly sad to see so many dead whitebark pines in the high
country,” said Noah Greenwald, endangered species director at the Center for
Biological Diversity. “These exceedingly beautiful trees are an icon of our
western mountains and they need all the help they can get, including
protection from development.”
Efforts are under way to collect seeds from apparently disease-resistant
individuals and outplant seedlings. It’s also important to protect the places
on the landscape where the tree is managing to survive, and critical habitat
protection could have helped with that.
The whitebark pine is a keystone species in high-elevation areas. It provides
food for grizzly bears and many other species. The trees also slow snowmelt,
helping to maintain stream flow into the summer months, which benefits fish
and other aquatic species.
“The whitebark pine is just one of the many species being pushed off the tops
of mountains by climate change and other factors,” said Greenwald. “With
warmer temperatures and earlier snowmelt, we’re also losing wolverines, pikas
and many more. This should be setting off alarm bells that we need to get our
planet-warming pollution under control.”
The loss of whitebark pine, along with the earlier disappearance of the
American chestnut from eastern deciduous forests, should have triggered much
more robust efforts to ensure that pathogens aren’t accidentally introduced
through trade in plants and animals, but this trade continues largely
unhindered.
In the past couple of decades, novel pathogens have killed millions of North
American bats and are killing snakes and rabbits in increasing numbers. It’s
very likely that COVID-19 arose from the wildlife trade, meaning that it’s not
just plants and animals that are being affected by the careless trade of
native species.
_The Center for Biological Diversity is a national, nonprofit conservation
organization with more than 1.7 million members and online activists dedicated
to the protection of endangered species and wild places._
[ 
](https://biologicaldiversity.org)
[ More Press Releases ](/news/breaking/)
#### Programs:
* [ Endangered Species ](/w/news/press-releases/programs/endangered-species/)
[ View for Email ](/w/news/press-releases/whitebark-pine-protected-as-
threatened-under-endangered-species-act-2022-12-14/email_view/)
| biology | 6166658 | https://sv.wikipedia.org/wiki/Wood%20River%2C%20British%20Columbia | Wood River, British Columbia | Wood River är ett vattendrag i Kanada. Det ligger i provinsen British Columbia, i den centrala delen av landet, km väster om huvudstaden Ottawa. Wood River ligger vid sjön Kinbasket Lake.
I omgivningarna runt Wood River växer i huvudsak barrskog. Trakten runt Wood River är nära nog obefolkad, med mindre än två invånare per kvadratkilometer. Trakten ingår i den hemiboreala klimatzonen. Årsmedeltemperaturen i trakten är °C. Den varmaste månaden är augusti, då medeltemperaturen är °C, och den kallaste är december, med °C.
Kommentarer
Källor
Vattendrag i British Columbia | swedish | 1.222467 |
birches_white_bark/Sun_scald.txt | Sun scald is the freezing of bark following high temperatures in the winter season, resulting in permanent visible damage to bark. Fruits may also be damaged. In the northern hemisphere, it is also called southwest injury.
Causes[edit]
The reason the sun can cause so much damage to trees is because of dormancy. When a tree is dormant in the winter it can be reactivated by warm weather. In the northern woods trees are exposed to the most sunlight and heat on the southwest facing side, so this side is heated during warm sunny winter days (in the afternoon, that's why westwards) to the point that it can be awoken from dormancy. The temperature required to wake up a tree depends on plant species and length of day, but it is typically just above freezing. Once active, the cells on the southwest side of the plant are unable to return to dormancy by nightfall, at which time the temperature returns to levels capable of killing active cells. Fluctuating winter temperatures can also cause frost cracks, which result from the expanding and contracting of the tree trunk.
Trees[edit]
Sun scald on ash bark
Slugs sheltering in a sun scald fissure
When sun scald appears on trees it is most frequently a result of reflected light off the snow during winter months. The damage in this case will appear as sunken or dead bark on the trunk of the tree, then later in the tree's life the bark might fall away revealing dead tissue in the tree's cambium layer. This damage will typically be found on the south west facing side of the tree's trunk. It can be found on other sides of the tree if there is light reflection from other sources, like man made structures or reflective rock faces. After a tree is afflicted by sun scald it becomes much more vulnerable to decay organisms. The plant will create walls around the affected area, but sometimes it is not enough to block the infections. The leaves of the tree are also affected by sun scald, particularly on a bright sunny day following a period of warm cloudy humidity. The damage to the leaves will start as bronzing of the epidermis between the veins of the leaf, and if the sunny conditions persist the tissue of the leaf will die.
Fruits[edit]
Sun scald on fruit appears when a fruit is exposed to direct sunlight after cold storage for an extended period of time, typically after harvest. The damage can often lead to the death of the fruit via consumption by insects, animals, bacteria, or fungi. This is the case if the defenses of the fruit are too heavily inflicted, which may occur when the outer skin is damaged to the point that the cell walls/membranes are either gone or weak enough for the plant's enemies to penetrate. Sun scald often presents as more of an internal damage, with the affected tissue gaining a leathery or wrinkled appearance and a pale discoloration. The terms "sun scald" and "sunburn" are frequently used interchangeably, however fruit sunburn is generally distinguished by only occurring pre-harvest and leaving yellow, bronze, or brown discoloration of the skin. Fruits damaged to either extent are usually considered unmarketable due to diminished appearance, taste, and texture, but those of the latter are sometimes sold at a lower grade or utilized as an (often puréed) ingredient. Sun damage cannot be reversed, but the fruit may still be able to fend off attacks and partially recover if properly treated and protected from further damage.
Treatments[edit]
The treatment of sun scalding is fundamentally simple: reduce the intensity of the sun, or block the sun completely.
Wrapping[edit]
The most common method used to prevent sun scalding on the trunks of trees is to wrap the tree up to the first branch with white paper overlapping approximately thirty three percent each time around the tree. The white paper is effective in reflecting the heat of the sun off the tree. The paper should be applied after the tree has gone dormant for the winter and taken off before it becomes active again. If the paper is left on too long it can interfere with the growth of the tree and harbor insects that may damage the tree.
Painting[edit]
Painting the tree white has the same effect as wrapping, although it is a permanent change to the color of the tree. This discoloration can be aesthetically displeasing, so this method is mostly used in orchards and rarely in landscaping.
Shading[edit]
The amount of light a tree receives on its southwest side is correlated with the amount of sun scald the tree endures. Reducing the amount of light the tree is exposed to by planting a shrub or bush strategically to shade the southwest side can be less effective than wrapping or painting, but can have better aesthetic qualities for landscaping.
Fruits[edit]
For fruit the most important part of avoiding sun scald is to be aware of where the fruit has been during its growth. If the fruit grew in the shade then exposing it to the sun will be likely to yield damage to the produce. Because of the importance of keeping shaded fruits out of the sun, leaves of fruit plants should be monitored for wilting and disease. If the leaves that shade a fruit die, the fruit will be exposed and in danger of sun scald. Another measure that can be taken to avoid damage is covering the fruits with straw or screen to block the sun.
See also[edit]
Frost crack | biology | 1649482 | https://sv.wikipedia.org/wiki/Barkbrand | Barkbrand | Barkbrand, eller solbrand, är en skada, som ibland uppkommer hos träd med tunn bark, och vanligen sådana, som föredrar skugga, som bok, avenbok, gran, silvergran m. fl. Den uppträder särskilt om träden plötsligt friställs och utsätts för middags- eller eftermiddagssolen (i sydligt eller västligt läge) ofta speciellt i kombination med frost.
Den orsakas av att bildningsväven (kambium) dödas genom den starka uppvärmningen, varefter den utanför det döda kambiet befintliga barken dör och faller av. Hos fristående träd skyddas stammen från upphettning genom den långt ned gående kronan, men träd, som vuxit i tätt bestånd saknar detta skydd, och när trädet genom friställning utsättes för stark bestrålning av solen kan barkbrand lätt uppstå.
Av ljusälskande träd är somliga, såsom tall, ek m. fl., skyddade genom en tjock bark, och björk genom en ljus bark, som reflekterar solstrålarna. I motsats till andra barkskador övervallas de av barkbrand skadade delarna endast långsamt, men då skadade träd länge kan hålla sig vid liv, finns det ej skäl att avverka i utkanten av ett bestånd stående, skadade träd, eftersom de då innanför de befintliga träden i sin tur lätt skadas när de exponeras för solen.
Värdefulla träd, såsom park- och fruktträd, kan i någon mån skyddas genom bestrykning med kalkmjölk eller lindning med skuggande material.
Källor
Lantmannens uppslagsbok, Stockholm 1923
Skogsbruk
Skogsbrand | swedish | 0.618828 |
birches_white_bark/whitebarkpinereceive.txt | Skip to main content
[  ](/ "FWS Home")
[ U.S. Fish & Wildlife Service ](/ "FWS Home")
Toggle navigation
## Utility (Top) navigation
* [ About Us ](/about) __ Forward
__ Back
* [ About Us ](/about)
* [ Mission & Vision ](/about/mission-and-vision)
* [ Leadership ](/about/leadership)
* [ Our Organization ](/about/programs)
* [ Our Facilities ](/our-facilities)
* [ Regions ](/about/regions)
* [ Laws & Regulations ](/laws) __ Forward
__ Back
* [ Laws & Regulations ](/laws)
* [ Laws, Agreements & Treaties ](/library/categories/laws)
* [ Hunting Regulations ](/refuges/hunting/rules-regulations-and-improved-access)
* [ Library ](/library/collections) __ Forward
__ Back
* [ Library ](/library)
* [ Categories ](/library/categories)
* [ Collections ](/library/collections)
*
* [ Home ](/)
* [ Services ](/services "Access services provided by FWS") __ Forward
__ Back
* [ Services ](/services "Access the landing page for services provided by FWS")
* [ Duck Stamps ](/service/duck-stamps) __ Forward
__ Back
* [ Duck Stamps ](/service/duck-stamps)
* [ Buy a Duck Stamp or E-Stamp ](/service/buy-duck-stamp-or-e-stamp)
* [ Buy a Junior Duck Stamp ](/service/buy-junior-duck-stamp)
* [ Fish Stocking ](/service/fish-stocking)
* [ Importing & Exporting ](/service/importing-and-exporting)
* [ Consultation & Technical Assistance ](/service/consultation-and-technical-assistance) __ Forward
__ Back
* [ Consultation & Technical Assistance ](/service/consultation-and-technical-assistance)
* [ ESA Section 7 Consultation ](/service/section-7-consultations)
* [ Habitat Conservation Planning (HCPs) ](/service/habitat-conservation-plans)
* [ Candidate Conservation Agreements (CCA & CCAA) ](/service/candidate-conservation-agreements-assurances)
* [ Safe Harbour Agreements (SHA) ](/service/safe-harbor-agreements)
* [ Conservation Banking ](/service/conservation-banking)
* [ Coastal Barrier Resources Act Project Consultation ](/service/coastal-barrier-resources-act-project-consultation)
* [ Coastal Barrier Resources System Property Documentation ](/service/coastal-barrier-resources-system-property-documentation)
* [ Financial Assistance ](/service/financial-assistance)
* [ Species Management ](/service/species-management)
* [ Investigational New Animal Drugs (INADS) ](/service/investigational-new-animal-drugs-inads)
* [ Permits ](/service/permits)
* [ Search All Services ](/service/search)
[ 
](/carp/service/duck-stamps) [ Duck Stamps ](/carp/service/duck-stamps) [ One
of the easiest ways that anyone can support bird habitat conservation is by
buying duck stamps. ](/carp/service/duck-stamps)
* [ Species ](/species) __ Forward
__ Back
* [ Species ](/species)
* [ Explore Taxonomic Tree ](/explore-taxonomic-tree)
* [ Find a Species ](/species/search)
[ 
](/story/coolest-mammals-earth) [ Interest story ](/story/coolest-mammals-
earth) [ Bats: “The Coolest Mammals on Earth” ](/story/coolest-mammals-earth)
* [ Visit Us ](/visit-us) __ Forward
__ Back
* [ Visit Us ](/visit-us)
* [ Events ](/events)
* [ Recreation Passes ](/service/federal-recreational-lands-passes)
* [ Outdoor Recreation ](/activities)
* [ Where to Hunt ](/hunting/map)
* [ Where to Fish ](/fishing/map)
* [ Our Locations ](/our-facilities)
[ 
](/activity/auto-tour) [ Scenic Drives ](/activity/auto-tour) [ Tour routes
of great scenic drives on National Wildlife Refuges. ](/activity/auto-tour)
* [ Get Involved ](/get-involved) __ Forward
__ Back
* [ Get Involved ](/get-involved)
* [ Careers and Internships ](/careers)
* [ Volunteering ](/volunteer-opportunity)
* [ Friends Partnerships ](/program/friends-partnerships)
* [ Learning Opportunities ](/training)
* [ Education Programs ](/education-programs)
* [ Events ](/events)
* [ Partnerships ](/partner) __ Forward
__ Back
* [ Partnerships ](/partner)
* [ Partnership Categories ](/partner/categories)
* [ Partner List ](/partner/search)
[ 
](/volunteer-opportunity) [ Volunteer ](/volunteer-opportunity) [ Search for
volunteer opportunities around the country ](/volunteer-opportunity)
* [ Newsroom ](/news) __ Forward
__ Back
* [ Newsroom ](/news)
* [ Press Releases ](/press-release)
* [ Congressional Testimony ](/testimony)
* [ Media Contacts ](/media-contacts)
[ 
](/news/wildlife-wonders) [ Wild Things ](/news/wildlife-wonders) [ News
about wonderful wild things and places ](/news/wildlife-wonders)
* [ Initiatives ](/initiatives) __ Forward
__ Back
* [ Initiatives ](/initiatives)
* [ Combating Wildlife Trafficking ](/initiative/combating-wildlife-trafficking)
* [ Director's Priorities ](/initiative/directors-priorities)
* [ Climate Change ](/initiative/climate-change)
* [ Hunting ](/initiative/hunting)
* [ Fishing ](/initiative/fishing)
* [ Invasive Species ](/initiative/invasive-species)
* [ Pollinators ](/initiative/pollinators)
* [ Protecting Wildlife ](/initiative/protecting-wildlife)
[ 
](/initiative/climate-change) [ Climate Action ](/initiative/climate-change)
[ FWS is taking steps to mitigate climate impacts ](/initiative/climate-
change)
* [ I want to ](/) __ Forward
__ Back
* I Want To
* [ Report Wildlife Crime ](/wildlife-crime-tips "Report wildlife trafficking or other wildlife crime to our tip line")
* [ Do Business with FWS ](/program/contracting "info on contracting with FWS")
* [ Volunteer ](/volunteer-opportunity)
* [ Find a Job or Internship ](/careers)
* [ Visit a Refuge ](/visit-us/refuges)
* [ Buy a Duck Stamp ](/service/buy-duck-stamp-or-e-stamp)
* [ Apply for a Permit ](/service/permits "Apply for a Permit or License")
* [ Find Funding ](/service/financial-assistance)
* [ Find Training ](/training)
[ 
](/jobs) [ Wild work ](/jobs) [ Search employment opportunities with USFWS
](/jobs)
## Search
Search
Search
Enter the terms you wish to search for.
×
# Whitebark pine receives ESA protection as a Threatened species

[ Image Details ](/banner/whitebark-pine-2credit-diana-tombackjpg)
Press Release
Whitebark pine receives Endangered Species Act protection as a Threatened
species
Significant threats continue to challenge this keystone species of the
American West
Dec 14, 2022
Media Contacts
[ Joe Szuszwalak ](/staff-profile/joe-szuszwalak)

[ Image Details ](/media/whitebark-pine-2credit-diana-tombackjpg)
**DENVER —** Today, the U.S. Fish and Wildlife Service (Service) is announcing
a final action to list the whitebark pine ( _Pinus albicaulis_ ) as a
threatened species under the Endangered Species Act (ESA). The Service has
concluded that the whitebark pine is likely to become endangered in the
foreseeable future throughout its range. These protections follow a proposal
to list the species as threatened in [ December 2020 ](/press-
release/2020-12/proposed-protections-whitebark-pine) with a subsequent public
comment period. The proposal and final action to list the whitebark pine were
made based on a rigorous [ Species Status Assessment
](https://ecos.fws.gov/ServCat/DownloadFile/226045) , using the best available
science.
“As a keystone species of the West, extending ESA protections to whitebark
pine is critical to not only the tree itself, but also the numerous plants,
animals, and watersheds that it supports,” **said Service Regional Director
Matt Hogan** . “The Service now looks forward to continuing engagement with
the many whitebark pine conservation partners during the recovery planning
process to ensure this species continues to endure for future generations.”
Whitebark pine trees live in windy, cold, high-elevation or high-latitude
environments across the western United States and southern Canada. As a
keystone species, this five-needled pine influences the health and life cycle
of other native plants and animals. In addition to providing a high-energy
food source for animals, healthy whitebark pine stands also play an essential
role in slowing runoff from snowmelt and reducing soil erosion.
White pine blister rust remains the primary threat to whitebark pine. This
blister rust is a non-native fungal disease that harms whitebark pine trees
across the West. Additional threats impacting the health of the species
include mountain pine beetles, altered wildfire patterns, and climate change
**climate change** __
Climate change includes both global warming driven by human-induced emissions
of greenhouse gases and the resulting large-scale shifts in weather patterns.
Though there have been previous periods of climatic change, since the mid-20th
century humans have had an unprecedented impact on Earth's climate system and
caused change on a global scale.
[ Learn more about climate change ](/glossary/climate-change) . As a result of
these threats, scientists estimate that as of 2016, 51% of all standing
whitebark pine trees are dead.

[ Image Details ](/media/whitebark-pine-3credit-diana-tomback-2jpg)
Providing ESA protections to whitebark pine will boost new and ongoing
research efforts to conserve the species, including future developments in
combatting white pine blister rust. The protections for whitebark pine also
make it illegal to remove, possess, or damage the tree on federal lands, or to
engage in interstate or foreign commerce, including the prohibition of
importing or exporting the tree. Unlike the prohibitions for federal lands, it
remains legal to remove or possess whitebark pine on non-federal lands per the
ESA, as long as those activities are otherwise allowed under state law.
Many partners are already engaged in restoration and conservation efforts for
whitebark pine across the region. The U.S. Forest Service is collaborating
with the Whitebark Pine Ecosystem Foundation (WPEF) and American Forests to
promote strategic range-wide conservation of the species, with the support of
the U.S. Fish and Wildlife Service. This effort will help guide the recovery
process by leveraging the ongoing work of these partnerships to create an
effective strategy to save the species from further decline.
“We have the tools and capability to make populations more resilient to these
threats. Given the scale of this effort, we are approaching restoration both
collaboratively and strategically," **said Diana Tomback, WPEF policy and
outreach coordinator and professor of integrative biology at the University of
Colorado Denver** . “One promising work in progress is the National Whitebark
Pine Restoration Plan, a roadmap for restoration being developed
collaboratively between the WPEF and American Forests, in consultation with
the US Forest Service, the National Park Service, Bureau of Land Management,
and several northwestern tribes.”

[ Image Details ](/media/whitebark-pine-cones)
The Service is not designating critical habitat for this species as part of
the listing because habitat loss is not a threat to the species’ continued
survival; disease from white pine blister rust is the primary threat. A broad
distribution of the species remains across more than 80 million acres in
Washington, Oregon, California, Idaho, Montana, Wyoming, Nevada, and Canada.
In the U.S., 88% of the species range is on federal land managed by the U.S.
Forest Service, National Park Service, and Bureau of Land Management, allowing
for thoughtful management of these lands through the ESA [ consultation
process ](/service/esa-section-7-consultation) .
To allow for further conservation of the species, the Service has included a
4(d) rule with the listing of the whitebark pine. A [ 4(d) rule is one of the
tools in the ESA ](/sites/default/files/documents/section-4d-rules_0.pdf) that
allows the Service to authorize activities that benefit and conserve the
species. In the case of whitebark pine, the Service is allowing research,
forest management, and restoration work on federal lands where it might
otherwise be prohibited. This provision also allows for the collection of
seeds on federal lands for Tribal ceremonial and traditional use.
The final rule to list the whitebark pine as a threatened species and
accompanying 4(d) rule will be published tomorrow in the Federal Register and
is available for public inspection today in the Reading Room: [
https://www.federalregister.gov/d/2022-27087
](https://www.federalregister.gov/d/2022-27087) .
To learn more about the whitebark pine, please visit the [ species profile
page ](/species/whitebark-pine-pinus-albicaulis) , additional questions and
answers about the listing of this species can be found on our website: [ Q&As:
Whitebark pine listing as Threatened ](/project/qas-whitebark-pine-listing-
threatened "Q&As: Whitebark pine listing as Threatened") .
**Additional partner quotes:**
* “The Confederated Salish and Kootenai Tribes’ of Montana commend and honor the USFWS decision listing Whitenbark pine as a threatened species, **said the Confederated Salish and Kootenai Tribes Forestry Department** . “Native Americans continue to be stewards of the land and understand the need for balanced ecosystems. CS&KT does this by utilizing our traditional ecological knowledge taught from story and songs and applying it to western science techniques. Whitebark pine is not only a keystone species for this balance, but it is also part of our first foods and culture. CS&KT supports the listing Whitebark pine to help insure the protection and restoration of the land and of our culture.”
* “We applaud the decision by the USFWS to list whitebark pine as threatened,” **said David Neale, Whitebark Pine Ecosystem Foundation Director and emeritus professor of plant sciences at the University of California, Davis** . “It also brings much-needed attention to the plight of this remarkable tree and builds further public support for the challenging restoration work ahead.”
* “Whitebark pine is the center of an important web of life and provides valuable ecosystem services in western high-elevation forests.” **said Diana Tomback, Whitebark Pine Ecosystem Foundation policy and outreach coordinator and professor of integrative biology at the University of Colorado Denver** . “But it is facing an unprecedented convergence of lethal threats. We have the tools and capability to make populations more resilient to these threats. Given the scale of this effort, we are approaching restoration both collaboratively and strategically. One promising work in progress is the National Whitebark Pine Restoration Plan, a roadmap for restoration being developed collaboratively between the Whitebark Pine Ecosystem Foundation and American Forests, in consultation with the US Forest Service, the National Park Service, Bureau of Land Management, and several northwestern tribes.”
### Story Tags
Endangered and/or Threatened species
Trees
Press Release
### Published
Dec 14, 2022
[ Endangered Species Act ](/news/endangered-species-act)
### Media Contacts
[ Joe Szuszwalak ](/staff-profile/joe-szuszwalak)
### Related Stories


Endangered Species Act
[ Proposed Protections for Whitebark Pine ](/press-release/2020-12/proposed-
protections-whitebark-pine)
Dec 1, 2020
### Latest Press Releases


Energy
[ Seeking public comment for Prosperity Wind Project Habitat Conservation Plan
](/press-release/2024-04/seeking-public-comment-prosperity-wind-project-
habitat-conservation-plan)
Apr 29, 2024


Habitat Restoration
[ $7.5M More in Bipartisan Infrastructure Law Funding to Support Salmon and
Communities ](/press-release/2024-04/75m-more-bipartisan-infrastructure-law-
funding-support-salmon-and-communities)
Apr 25, 2024


Climate Change
[ IRA-Funded Projects in Eastern North Carolina Announced ](/press-
release/2024-04/ira-funded-projects-eastern-north-carolina-announced)
Apr 25, 2024
Pagination
* [ Previous page ]( "Go to previous page")
* [ Current page 1 ](?page=0 "Current page")
* [ Page 2 ](?page=1 "Go to page 2")
* [ Page 3 ](?page=2 "Go to page 3")
* [ Page 4 ](?page=3 "Go to page 4")
* [ Page 5 ](?page=4 "Go to page 5")
* [ Page 6 ](?page=5 "Go to page 6")
* [ Page 7 ](?page=6 "Go to page 7")
* [ Page 8 ](?page=7 "Go to page 8")
* [ Page 9 ](?page=8 "Go to page 9")
* [ Next page ](?page=1 "Go to next page")

Working with others to conserve, protect and enhance fish, wildlife, plants
and their habitats for the continuing benefit of the American people.
## Footer Menu - Employment
* [ Careers & Internships ](/careers)
* [ Contracting ](/program/contracting)
## Footer Menu - Site Links
* [ Leadership ](/about/leadership)
* [ Frequently Asked Questions ](/frequently-asked-questions "Questions from the customer service call center")
* [ Contact FWS ](/contact-us)
## Footer Menu - Legal
* [ Accessibility ](/carp/help/accessibility)
* [ Freedom of Information Act ](/carp/program/fws-freedom-information-act-foia)
* [ Notices ](/carp/notices)
* [ Privacy Policy ](https://www.doi.gov/privacy)
* [ Disclaimers ](/carp/disclaimer)
* [ Information Quality ](/carp/program/information-quality)
* [ Vulnerability Disclosure Policy ](/carp/vulnerability-disclosure-policy)
## Footer Menu - External Links
* [ DOI ](https://www.doi.gov)
* [ USA.GOV ](https://www.usa.gov)
* [ ](https://www.facebook.com/usfws "Follow us on Facebook")
* [ ](https://www.instagram.com/usfws "Follow us on Instagram")
* [ ](https://www.twitter.com/usfws "Follow us on Twitter")
* [ ](https://www.linkedin.com/company/usfws "Find us on LinkedIn")
* [ ](https://www.flickr.com/photos/usfwshq "Find us on Flickr")
* [ ](https://www.youtube.com/usfws "Follow us on YouTube")
### You are exiting the U.S. Fish and Wildlife Service website
You are being directed to [ ]()
We do not guarantee that the websites we link to comply with Section 508
(Accessibility Requirements) of the Rehabilitation Act. Links also do not
constitute endorsement, recommendation, or favoring by the U.S. Fish and
Wildlife Service.
[ I Understand. Take me there. ]() Cancel
| biology | 1980233 | https://sv.wikipedia.org/wiki/Pachyseris%20speciosa | Pachyseris speciosa | Pachyseris speciosa är en korallart som först beskrevs av James Dwight Dana 1846. Pachyseris speciosa ingår i släktet Pachyseris och familjen Agariciidae. IUCN kategoriserar arten globalt som livskraftig. Inga underarter finns listade i Catalogue of Life.
Källor
Externa länkar
Stenkoraller
speciosa | swedish | 1.230508 |
birches_white_bark/fsb.txt | USDA Forest Service R6-NR-FHP-2007-01
Using Verbenone to Protect Whitebark Pine from
Mountain Pine Beetle Attack
Kegley, Sandra J.1
; and Gibson, Kenneth E. 2
1 USDA Forest Service, Forest Health Protection, Coeur d’Alene Field Office, 3815 Schreiber Way,
Coeur d’Alene, ID
2 USDA Forest Service, Forest Health Protection, Missoula Field Office, P.O. Box 7669, Missoula,
MT
Whitebark pine (Pinus albicaulis Engelm.) is currently declining in many areas due to a variety of
factors including white pine blister rust, fire suppression, forest successional processes, and periodic
outbreaks of mountain pine beetle (Dendroctonus ponderosae Hopkins).
In 2005 in northern Idaho and Montana, mountain pine beetle killed nearly 630,000 mature whitebark
pine trees on about 142,000 acres. Many of these stands have also been impacted by white pine
blister rust. Seed is collected from phenotypically blister rust-resistant “plus” trees which are now
under threat of mountain pine beetle attack. Silvicultural methods such as thinning, used to manage
mountain pine beetle in other hosts, may not be applicable in less-dense whitebark pine stands.
Individual, high-value trees can be protected from beetle attack with topical treatments of insecticides
applied to tree boles with high-pressure spray equipment. However, spraying is not practical in
inaccessible areas or on a large scale. Using beetle pheromones to protect high-value trees from
attack has been shown to be a viable management option.
We tested the efficacy of verbenone, an anti-aggregation pheromone for mountain pine beetle, in
protecting individual whitebark pine trees from beetle attack for three years. EPA-registered 5-gram
verbenone pouches, replaced at mid-season, were tested along with new thicker membrane (longer
lasting) pouches, and larger (7.5-gram) pouches. All were compared to untreated controls.
In 2002, we tested verbenone on 150 whitebark pine in northern Idaho. Treatments were: two, 5-
gram pouches per tree; four pouches per tree; and untreated controls. To ensure equal beetle pressure,
each tree was baited with a mountain pine beetle attractant tree bait. Pouches were replaced midseason due to their short elution period. After beetle flight, trees were rated as mass attack, strip
attack, pitch out, or no attack. “Mass attack” means the tree was overcome by beetles and killed.
“Strip attack” is a tree successfully attacked on a portion of its bole, but the tree is not killed. A
“pitch out” is one unsuccessfully attacked. “No attack” is an unattacked tree.
148
Proceedings of the Conference Whitebark Pine: A Pacific Coast Perspective
All untreated trees were killed while over 90% of treated trees survived. There was no statistical
difference between the use of two or four pouches per tree (figure 1).
0%
20%
40%
60%
80%
100%
Control 2 Pouches 4 Pouches
Treatment
% Trees
mass attack
strip attack
pitch out
No attack
Figure 1. A test of whitebark pine trees treated with two or four verbenone pouches per tree in North
Idaho in 2002.
In 2003, we treated 150 whitebark pine in western Montana. Treatments were: two standard 5-gram
pouches per tree; two experimental longer-lasting 5-gram pouches per tree; and untreated controls.
Only standard pouches were replaced mid-season. Attractant tree baits were placed 10-15 feet from
each tree. Results were comparable to 2002. A total of 42% of control trees were killed while over
90% of the treated trees survived (figure 2). There was no significant difference between the two
pouches.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Control Standard
Pouch
(replaced)
New Longerlasting
Pouch
Treatment
% Trees
mass attack
strip attack
pitch out
No attack
Figure 2. Whitebark pine test in 2003 in western Montana using two standard verbenone pouches that
were replaced mid-season, and two experimental longer lasting pouches per tree.
149
USDA Forest Service R6-NR-FHP-2007-01
In 2004, 100 whitebark pine were treated in northern Idaho. Treatments were two, experimental 7.5
gram pouches per tree and untreated controls. The pouches were not replaced mid-season. Attractant
tree baits were placed 5-10 feet from each tree. Treated trees were protected from mass attack (figure
3). While 77% of control trees were killed, 87% of treated trees survived.
0%
20%
40%
60%
80%
100%
Control 7.5 g Pouch
Treatment
Mass Attack
Strip Attack
Pitch Out
No Attack
Figure 3. Whitebark pine test in North Idaho in 2004 using two 7.5 gram verbenone pouches per tree.
In summary, most individual whitebark pine trees, subjected to the presence of a mountain pine beetle
attractant pheromone, were protected from mass beetle attacks. Treating individual whitebark pine
trees with either two registered or two experimental pouches significantly reduced mountain pine
beetle attacks when compared to untreated controls. These tests provide strong evidence that
individual pine trees can be protected from mountain pine beetle attack using verbenone. Standard
(5-gram) verbenone pouches are currently registered and available for use. Used operationally, they
have successfully protected whitebark pine plus trees from mountain pine beetle attack, even in areas
of high beetle populations.
150 | biology | 2530522 | https://sv.wikipedia.org/wiki/Nesoleon%20boschimanus | Nesoleon boschimanus | Nesoleon boschimanus är en insektsart som först beskrevs av Louis Albert Péringuey 1910. Nesoleon boschimanus ingår i släktet Nesoleon och familjen myrlejonsländor. Inga underarter finns listade i Catalogue of Life.
Källor
Myrlejonsländor
boschimanus | swedish | 1.225344 |
birches_white_bark/northendangeredspeci.txt | ## Our stories are fueled by readers like you
Donate $10 Donate $25 Other amount
If you learned something new or were moved by what you read here, **help us do
more of this work with your donation** . We’re only able to bring this news
and analysis to millions of people across the country because of readers like
you. Reader contributions make up 75% of our revenue and are key to the future
of healthy independent media.
Close
* Your Dashboard
Search for: Search
* [ Features ](https://www.hcn.org/topic/features/)
* [ Public Lands ](https://www.hcn.org/topic/public-lands/)
* [ Indigenous Affairs ](https://www.hcn.org/topic/indigenous-affairs/)
* [ Water ](https://www.hcn.org/topic/water/)
* [ Climate Change ](https://www.hcn.org/topic/climate-change/)
* [ Arts & Culture ](https://www.hcn.org/topic/arts-culture/)
* [ Subscribe ](https://www.hcn.org/subscribe/)
* [ Donate Now ](https://www.hcn.org/support/)
* [ The Magazine ](https://www.hcn.org/topic/the-magazine/)
* Jobs & Classifieds Open dropdown menu
* [ Jobs & Classifieds ](https://classifieds.hcn.org/)
* [ Place a Classified Ad ](https://highcountrynews.adperfect.com/channel/C0A801FE1b4c7201BEWYX3FF29A8/publication/C0A801FE1b4c72030CvHv3ED6703)
* [ Display Ad Info ](https://www.hcn.org/advertising/)
Close Close
* Your Dashboard
* [ Features ](https://www.hcn.org/topic/features/)
* [ Public Lands ](https://www.hcn.org/topic/public-lands/)
* [ Indigenous Affairs ](https://www.hcn.org/topic/indigenous-affairs/)
* [ Water ](https://www.hcn.org/topic/water/)
* [ Climate Change ](https://www.hcn.org/topic/climate-change/)
* [ Arts & Culture ](https://www.hcn.org/topic/arts-culture/)
* [ Subscribe ](https://www.hcn.org/subscribe/)
* [ Donate Now ](https://www.hcn.org/support/)
* [ The Magazine ](https://www.hcn.org/topic/the-magazine/)
* Jobs & Classifieds Open dropdown menu
* [ Jobs & Classifieds ](https://classifieds.hcn.org/)
* [ Place a Classified Ad ](https://highcountrynews.adperfect.com/channel/C0A801FE1b4c7201BEWYX3FF29A8/publication/C0A801FE1b4c72030CvHv3ED6703)
* [ Display Ad Info ](https://www.hcn.org/advertising/)
Skip to content
Menu
[ 
](https://www.hcn.org/)
[ High Country News ](https://www.hcn.org/)
A nonprofit independent magazine of unblinking journalism that shines a light
on all of the complexities of the West.
[ Support ](/support/) Menu
Open Search
Search for: Search
Posted in [ Articles ](https://www.hcn.org/category/articles/) [ Endangered
Species ](https://www.hcn.org/topic/endangered-species/)
# How to save the whitebark pine
The tree is getting federal protection. But plenty of people were already
trying to save it.
by [ Kylie Mohr ](https://www.hcn.org/author/kylie-mohr/) January 5, 2023
January 24, 2024
* [ Print ](https://www.hcn.org/articles/north-endangered-species-how-to-save-the-whitebark-pine/#print "Click to print")
* [ Email ](mailto:?subject=%5BShared%20Post%5D%20How%20to%20save%20the%20whitebark%20pine&body=https%3A%2F%2Fwww.hcn.org%2Farticles%2Fnorth-endangered-species-how-to-save-the-whitebark-pine%2F&share=email "Click to email a link to a friend")
* [ Save Article ](https://www.hcn.org/articles/north-endangered-species-how-to-save-the-whitebark-pine/?share=custom-1697321641 "Click to share on Save Article")
* More
*
* [ Twitter ](https://www.hcn.org/articles/north-endangered-species-how-to-save-the-whitebark-pine/?share=twitter "Click to share on Twitter")
* [ Facebook ](https://www.hcn.org/articles/north-endangered-species-how-to-save-the-whitebark-pine/?share=facebook "Click to share on Facebook")
* [ Reddit ](https://www.hcn.org/articles/north-endangered-species-how-to-save-the-whitebark-pine/?share=reddit "Click to share on Reddit")
*
Whitebark pines are unmistakable, with their stout, twisted trunks — shaped
but not dominated by the wind — topped with clumps of needles on upswept
branches. But by 2016, [ over half ](https://www.fs.usda.gov/rmrs/science-
spotlights/landscape-scale-assessments-whitebark-pine) of those still standing
were husks of their former selves, their fate signaled by flaming red needles,
and then, ghostly gray trunks with no branches at all.
The trees are fighting an uphill battle. The invasive blister rust fungus,
mountain pine beetle infestations, changing wildfire patterns and climate
change [ all threaten
](https://www.sciencedirect.com/science/article/pii/S0378112721010203) this
keystone species. It was officially [ listed ](https://www.fws.gov/press-
release/2022-12/whitebark-pine-receives-esa-protection-threatened-species) as
threatened by the U.S. Fish and Wildlife Service in December 2022.
The tree plays important roles in the [ ecosystem
](https://www.americanforests.org/save-the-whitebark-pine/) : Its branches
create shade, which helps retain snowpack, and its roots hold the soil in
place, preventing erosion. Whitebark pine cones nourish animals like grizzlies
and Clark’s nutcrackers and serve as a protein-rich Indigenous food source. At
a conference on whitebark pine conservation in 2019, the late Selis-Qlispe
tribal elder Tony Incashola Sr. remembered snacking on pine nuts when he was
growing up on the Flathead Reservation. “If you come across it in your
travels, it was a treat to have,” the [ _Char-Koosta News_ reported _._
](http://www.charkoosta.com/news/elder-tony-incashola-remembers-whitebark-
pine-from-his-youth/article_4c28c94c-e09f-11e9-9232-274e4ed71e48.html)
 Monique Wynecoop (Pit River/Maidu),
U.S. Forest Service fire ecologist, makes a gift of tobacco and prays at what
remains of a 2,000-year-old whitebark pine tree named ‘Illawia,’ which means
great-great-grandparent in the Salish languge, during a guided hike with
members and guests of the Whitebark Pine Ecosystem Foundation in the mountains
of the Flathead Indian Reservation, Montana, in 2019. The Confederated Salish
and Kootenai Tribes are working to restore whitebark pines on tribal lands.
Credit: Chip Somodevilla/Getty Images
Whitebark pines span an impressive range of 80.5 million subalpine acres in
seven Western states. Across that sprawling region, tribal nations,
conservation groups and federal agencies have already made significant efforts
to protect and restore the tree, even as federal protection has lagged. It’s
been a candidate for protection since 2011 and was listed as endangered in
Canada in 2012, but other higher priority species got the focus in the U.S.
for years.
Listing means new money and formalized safeguards. Fish and Wildlife Service
funding for listed species can be used to boost new and ongoing research into
things like blister rust resilience. The listing allows management and
restoration activities in places where they might otherwise be prohibited,
such as wilderness areas, and makes it illegal to remove or damage the tree on
federal lands, although tribes can still collect seeds on federal lands for
ceremonial and traditional uses. A [ national restoration plan
](https://whitebarkfound.org/our-work/national-whitebark-pine-restoration-
plan/) , created by nonprofits working with the federal government and tribal
nations, is slated to publish in 2023; the listing also triggers the creation
of a recovery plan by Fish and Wildlife, with both plans building off the hard
work already underway on the ground.
### **Growing disease-resistant trees**
White pine blister rust, an invasive fungal disease, is the primary threat to
the pine. Dead red branches and orange-rimmed canker sores are signs of the
disease, which can eventually kill the tree.
Identifying trees that appear to resist it, then growing their offspring in
nurseries and replanting them in the wild, is one way to create tougher
forests. A greenhouse full of yellow tubes in Coeur d’Alene, Idaho, is home to
100,000 1-year-old whitebark pine seedlings. The [ Forest Service nursery
](https://subscriber.politicopro.com/article/eenews/2021/04/26/fleet-fingered-
crew-fights-for-persnickety-pine-002669) there supplies the most rust-
resistant seedlings of all six Forest Service nurseries in the country.
### **Collecting seeds and genetic material**
The Confederated Salish and Kootenai Tribes are [ working hard
](http://www.charkoosta.com/news/whitebark-pine-set-to-make-a-comeback-on-
flathead-reservation/article_15d337da-e0a2-11e9-9ae4-6bbddcf79673.html) to
restore whitebark pines on tribal lands. Starting in 2016, the nations’
forestry and historic preservation departments identified areas with resistant
trees, planted seedlings and caged ripening cones to safeguard them from
hungry critters. Cones are then harvested for nurseries like the one in Idaho.
The tribes also collect genetic material like pollen, to help researchers
investigate drought and blister rust resilience in the trees. Genetic testing
occurs through the [ Whitebark Pine Genome Initiative
](https://whitebarkfound.org/our-work/genome-projects/genome-project/) and
research at the [ Forest Service Dorena Genetic Resource Center.
](https://www.mpgnorth.com/research/dorena-grc-collaboration)
### **Building seed orchards**
Why traipse around in the woods, searching for whitebark pine cones and
climbing trees to harvest them, when you could just head to a designated
orchard? Seed orchards, like the one created in Montana’s Custer Gallatin
National Forest in [ 2013
](https://www.fs.usda.gov/detail/custergallatin/news-
events/?cid=STELPRDB5426775) , are meant to speed up and simplify the seed
sourcing process. While it takes the orchards [ two or more decades
](https://www.sciencedirect.com/science/article/pii/S0378112721010203) to be
operational — trees must be old enough to produce a good cone crop — having
easily accessible seeds is a long-term investment.
### **Protecting trees from mountain pine beetle**
Mountain pine beetles kill whitebark pines [ from within
](https://kids.frontiersin.org/articles/10.3389/frym.2022.678082) . The bugs
swarm the tree, chew a network of paths inside its bark and lay their eggs.
When the eggs hatch, the larvae feed and develop in tissue under the bark —
blocking water and nutrients, and eventually killing the tree. In warmer
years, more beetles [ survive the winter ](https://www.usgs.gov/news/climate-
has-led-beetle-outbreaks-iconic-whitebark-pine-trees) , and also reproduce
faster, and drought-stressed trees are especially susceptible to them.
Targeted approaches can help protect individual trees and their cones from
such onslaughts. Beetle pheromones can trick the bugs at their own game, [
according to
](https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev2_025991.pdf) the
Forest Service: An early 2000s [ study in north Idaho
](https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev2_025991.pdf) found
that an artificially made mountain pine beetle attractant pheromone,
verbenone, protected individual whitebark pines from mass beetle attacks.
Beetles produce verbenone when they’ve attacked a tree and there are too many
beetles, signaling to their colleagues: This tree is occupied. Go somewhere
else. Humans want to mimic this signal to keep beetles away altogether.
### **Designating priority restoration areas**
The [ national whitebark pine restoration plan
](https://whitebarkfound.org/our-work/national-whitebark-pine-restoration-
plan/) , which develops [ priority areas
](https://www.sciencedirect.com/science/article/pii/S0378112722001980) for
restoration, is led by the Whitebark Pine Ecosystem Foundation and American
Forests in consultation with the Forest Service, along with other federal land
management agencies and tribal nations. National parks, Forest Service regions
and Bureau of Land Management field offices all nominated 20% to 30% of their
whitebark pine distribution as potential core areas for the work.
This kind of targeted approach helps deal with the logistical and fiscal
challenges of the trees’ large range. The whitebark pine is the most widely
distributed forest tree protected under the Endangered Species Act.
Eventually, seeds from trees thriving in core areas will disperse into
neighboring areas. The final plan, which will be released this year, will
include nominated core areas, management strategies and estimated
implementation costs for proposed restoration activities.
 In 2019, Nico Matallana, National Park
Service biological science technician, plants a whitebark pine seedling among
trees killed by the 2017 Sprague Creek Fire on Mount Brown in Glacier National
Park, Montana. Credit: Chip Somodevilla/Getty Images
### **Keeping surrounding forests healthy**
Forestry techniques like prescribed fire and thinning can help whitebark
pines, too. Clearing out brush and limiting excess fuel reduces the likelihood
of high-severity fires. Fires [ can benefit
](https://www.mdpi.com/1999-4907/9/10/648) the pines, but they can [ also
](https://whitebarkfound.org/fire-regimes/) harm them.
Whitebark pines can survive low-intensity fires that occur in small portions
of their forests thanks to their thicker bark and deeper roots, while mixed
severity fires can open up habitat for the Clark’s nutcracker, a bird that
scatters the trees’ seeds. Whitebark pine forests in northwestern Montana,
northern Idaho and the Cascades have historically experienced periodic large
fires in which trees are killed but readily regenerate afterward. But today’s
[ more severe, more frequent ](https://www.nrfirescience.org/resource/17562)
wildfires can also kill mature pines, including the rust-resistant ones.
Losing these trees to wildfire would be a big blow to restoration efforts. [
Recent work ](https://www.mdpi.com/1999-4907/9/10/648) from the Forest Service
Rocky Mountain Research Station and Missoula Fire Sciences Laboratory suggests
effective whitebark pine restoration needs to “enhance the benefits and reduce
the losses from wildfires.”
### **Monitoring Clark’s nutcracker**
Whitebark pines need the Clark’s nutcracker to disperse their seeds, sometimes
as far as [ 20 miles ](https://www.fs.usda.gov/pnw/sciencef/scifi130.pdf)
_from their home trees_ , and the Clark’s nutcrackers needs whitebark pines
for calories. The two species’ fates are intertwined. So researchers across
the West are [ monitoring
](https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0227161)
this small bird, worried about its decline as the trees die. Tracking efforts
are underway in areas like North Cascades, Mount Rainier, Yosemite and
Sequoia-Kings Canyon national parks.
“We’re relying on the bird,” said Diana Tomback, a professor at the University
of Colorado Denver who studies whitebark pine. “It will take multiple human
lifetimes for whitebark pines’ range to be re-established, and Clark’s
nutcrackers’ seed dispersal will be the key to this.”
_Kylie Mohr is an editorial fellow for_ High Country News _writing from
Montana. Email her at[ [email protected] ](mailto:[email protected]) or
submit a [ letter to the editor ](https://www.hcn.org/feedback/contact-us) .
See our [ letters to the editor policy ](https://www.hcn.org/policies/lte) . _
[ Follow @thatsmohrlikeit ](https://twitter.com/thatsmohrlikeit)
Republish
Spread the word. News organizations can pick-up quality news, essays and
feature stories for free.
Republish This Story
[  ](http://creativecommons.org/licenses/by-nd/4.0/)
Republish our articles for free, online or in print, under a Creative Commons
license.
Close window X
## Republish this article
[  ](http://creativecommons.org/licenses/by-nd/4.0/)
This work is licensed under a [ Creative Commons Attribution-NoDerivatives 4.0
International License ](http://creativecommons.org/licenses/by-nd/4.0/) .
Our articles are available for republishing, but images are only available on
a case-by-case basis. You can’t republish photographs or illustrations without
written permission from High Country News and/or the photographer. Please
reach out to [ [email protected] ](mailto:[email protected]) to request
illustrations and photography for specific stories. **Republishing
guidelines**
* **Credit the author and High Country News -** We prefer Author Name, High Country News at the byline. At the top of the story, if on the web, please include this text and link: “This story was originally published by High Country News.”
* **Check the image requirements -** Images that are clearly marked CC or from federal agencies are in the creative commons and are available for republishing. Outside of this, you can’t republish photographs or illustrations without written permission from High Country News and/or the photographer. Please reach out to [ [email protected] ](mailto:[email protected]) to request more information.
* **Don’t change anything significant -** Articles must be republished in their entirety. Revisions for house style or references to time (“yesterday” to “today”) are allowed. If larger revisions are necessary, including significant trims or an editor’s note, contact us at [ [email protected] ](mailto:[email protected]) to get approval for the change ahead of republishing.
* **Share on social media -** When sharing on social tag High Country News in your post and note the story is from @highcountrynews
* Twitter: [ @highcountrynews ](https://twitter.com/highcountrynews)
* Instagram: [ @highcountrynews ](https://www.instagram.com/highcountrynews)
* TikTok: [ @highcountrynews ](https://www.tiktok.com/@highcountrynews)
* Facebook: [ @highcountrynews ](https://www.facebook.com/highcountrynews)
* LinkedIn: [ High Country News ](https://www.linkedin.com/company/high-country-news)
* **Stay in touch** \- Let us know you republished the story. Send a link to [ [email protected] ](mailto:[email protected]) once you’ve republished the story and let us know how it’s resonating with your readers.
# How to save the whitebark pine
by Kylie Mohr, High Country News
January 5, 2023
<h1>How to save the whitebark pine</h1> <p class="byline">by Kylie Mohr, High
Country News <br />January 5, 2023</p> <p>Whitebark pines are unmistakable,
with their stout, twisted trunks — shaped but not dominated by the wind —
topped with clumps of needles on upswept branches. But by 2016, <a
href="https://www.fs.usda.gov/rmrs/science-spotlights/landscape-scale-
assessments-whitebark-pine">over half</a> of those still standing were husks
of their former selves, their fate signaled by flaming red needles, and then,
ghostly gray trunks with no branches at all. </p> <p>The trees are
fighting an uphill battle. The invasive blister rust fungus, mountain pine
beetle infestations, changing wildfire patterns and climate change <a
href="https://www.sciencedirect.com/science/article/pii/S0378112721010203">all
threaten</a> this keystone species. It was officially <a
href="https://www.fws.gov/press-release/2022-12/whitebark-pine-receives-esa-
protection-threatened-species">listed</a> as threatened by the U.S. Fish and
Wildlife Service in December 2022. </p> <p>The tree plays important roles
in the <a href="https://www.americanforests.org/save-the-whitebark-
pine/">ecosystem</a>: Its branches create shade, which helps retain snowpack,
and its roots hold the soil in place, preventing erosion. Whitebark pine cones
nourish animals like grizzlies and Clark’s nutcrackers and serve as a protein-
rich Indigenous food source. At a conference on whitebark pine conservation in
2019, the late Selis-Qlispe tribal elder Tony Incashola Sr. remembered
snacking on pine nuts when he was growing up on the Flathead Reservation. “If
you come across it in your travels, it was a treat to have,” the <a
href="http://www.charkoosta.com/news/elder-tony-incashola-remembers-whitebark-
pine-from-his-
youth/article_4c28c94c-e09f-11e9-9232-274e4ed71e48.html"><em>Char-Koosta News
</em>reported<em>.</em></a></p> <figure class="wp-block-image size-full"><img
src="https://www.hcn.org/wp-content/uploads/2026/09/whitebark-
pine-23-2.jpg?fit=1200%2C800&ssl=1" alt="" class="wp-image-70217"
/><figcaption class="wp-element-caption">Monique Wynecoop (Pit River/Maidu),
U.S. Forest Service fire ecologist, makes a gift of tobacco and prays at what
remains of a 2,000-year-old whitebark pine tree named 'Illawia,' which means
great-great-grandparent in the Salish languge, during a guided hike with
members and guests of the Whitebark Pine Ecosystem Foundation in the mountains
of the Flathead Indian Reservation, Montana, in 2019. The Confederated Salish
and Kootenai Tribes are working to restore whitebark pines on tribal
lands.</figcaption></figure> <p>Whitebark pines span an impressive range of
80.5 million subalpine acres in seven Western states. Across that sprawling
region, tribal nations, conservation groups and federal agencies have already
made significant efforts to protect and restore the tree, even as federal
protection has lagged. It’s been a candidate for protection since 2011 and was
listed as endangered in Canada in 2012, but other higher priority species got
the focus in the U.S. for years. </p> <p>Listing means new money and
formalized safeguards. Fish and Wildlife Service funding for listed species
can be used to boost new and ongoing research into things like blister rust
resilience. The listing allows management and restoration activities in places
where they might otherwise be prohibited, such as wilderness areas, and makes
it illegal to remove or damage the tree on federal lands, although tribes can
still collect seeds on federal lands for ceremonial and traditional uses. A <a
href="https://whitebarkfound.org/our-work/national-whitebark-pine-restoration-
plan/">national restoration plan</a>, created by nonprofits working with the
federal government and tribal nations, is slated to publish in 2023; the
listing also triggers the creation of a recovery plan by Fish and Wildlife,
with both plans building off the hard work already underway on the
ground. </p> <h3 class="wp-block-heading"><strong>Growing
disease-resistant trees</strong></h3> <p>White pine blister rust, an invasive
fungal disease, is the primary threat to the pine. Dead red branches and
orange-rimmed canker sores are signs of the disease, which can eventually kill
the tree.</p> <p>Identifying trees that appear to resist it, then growing
their offspring in nurseries and replanting them in the wild, is one way to
create tougher forests. A greenhouse full of yellow tubes in Coeur
d’Alene, Idaho, is home to 100,000 1-year-old whitebark pine seedlings. The <a
href="https://subscriber.politicopro.com/article/eenews/2021/04/26/fleet-
fingered-crew-fights-for-persnickety-pine-002669">Forest Service nursery</a>
there supplies the most rust-resistant seedlings of all six Forest Service
nurseries in the country. </p> <h3 class="wp-block-
heading"><strong>Collecting seeds and genetic material</strong></h3> <p>The
Confederated Salish and Kootenai Tribes are <a
href="http://www.charkoosta.com/news/whitebark-pine-set-to-make-a-comeback-on-
flathead-
reservation/article_15d337da-e0a2-11e9-9ae4-6bbddcf79673.html">working
hard</a> to restore whitebark pines on tribal lands. Starting in 2016, the
nations’ forestry and historic preservation departments identified areas with
resistant trees, planted seedlings and caged ripening cones to safeguard them
from hungry critters. Cones are then harvested for nurseries like the one in
Idaho. The tribes also collect genetic material like pollen, to help
researchers investigate drought and blister rust resilience in the trees.
Genetic testing occurs through the <a href="https://whitebarkfound.org/our-
work/genome-projects/genome-project/">Whitebark Pine Genome Initiative</a> and
research at the <a href="https://www.mpgnorth.com/research/dorena-grc-
collaboration">Forest Service Dorena Genetic Resource
Center. </a></p> <h3 class="wp-block-heading"><strong>Building seed
orchards </strong></h3> <p>Why traipse around in the woods, searching for
whitebark pine cones and climbing trees to harvest them, when you could just
head to a designated orchard? Seed orchards, like the one created in Montana’s
Custer Gallatin National Forest in <a
href="https://www.fs.usda.gov/detail/custergallatin/news-
events/?cid=STELPRDB5426775">2013</a>, are meant to speed up and simplify the
seed sourcing process. While it takes the orchards <a
href="https://www.sciencedirect.com/science/article/pii/S0378112721010203">two
or more decades</a> to be operational — trees must be old enough to produce a
good cone crop — having easily accessible seeds is a long-term
investment. </p> <h3 class="wp-block-heading"><strong>Protecting trees
from mountain pine beetle</strong></h3> <p>Mountain pine beetles kill
whitebark pines <a
href="https://kids.frontiersin.org/articles/10.3389/frym.2022.678082">from
within</a>. The bugs swarm the tree, chew a network of paths inside its bark
and lay their eggs. When the eggs hatch, the larvae feed and develop in tissue
under the bark — blocking water and nutrients, and eventually killing the
tree. In warmer years, more beetles <a
href="https://www.usgs.gov/news/climate-has-led-beetle-outbreaks-iconic-
whitebark-pine-trees">survive the winter</a>, and also reproduce faster, and
drought-stressed trees are especially susceptible to them. </p>
<p>Targeted approaches can help protect individual trees and their cones from
such onslaughts. Beetle pheromones can trick the bugs at their own game, <a
href="https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev2_025991.pdf">according
to</a> the Forest Service: An early 2000s <a
href="https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev2_025991.pdf">study
in north Idaho</a> found that an artificially made mountain pine beetle
attractant pheromone, verbenone, protected individual whitebark pines from
mass beetle attacks. Beetles produce verbenone when they’ve attacked a tree
and there are too many beetles, signaling to their colleagues: This tree is
occupied. Go somewhere else. Humans want to mimic this signal to keep
beetles away altogether.</p> <h3 class="wp-block-heading"><strong>Designating
priority restoration areas</strong></h3> <p>The <a
href="https://whitebarkfound.org/our-work/national-whitebark-pine-restoration-
plan/">national whitebark pine restoration plan</a>, which develops <a
href="https://www.sciencedirect.com/science/article/pii/S0378112722001980">priority
areas</a> for restoration, is led by the Whitebark Pine Ecosystem Foundation
and American Forests in consultation with the Forest Service, along with other
federal land management agencies and tribal nations. National parks, Forest
Service regions and Bureau of Land Management field offices all nominated 20%
to 30% of their whitebark pine distribution as potential core areas for the
work.</p> <p>This kind of targeted approach helps deal with the logistical and
fiscal challenges of the trees’ large range. The whitebark pine is the most
widely distributed forest tree protected under the Endangered Species Act.
Eventually, seeds from trees thriving in core areas will disperse into
neighboring areas. The final plan, which will be released this year, will
include nominated core areas, management strategies and estimated
implementation costs for proposed restoration activities. </p> <figure
class="wp-block-image size-full"><img src="https://www.hcn.org/wp-
content/uploads/2026/09/whitebark-pine-23-1.jpg?fit=1200%2C800&ssl=1"
alt="" class="wp-image-70216" /><figcaption class="wp-element-caption">In
2019, Nico Matallana, National Park Service biological science technician,
plants a whitebark pine seedling among trees killed by the 2017 Sprague Creek
Fire on Mount Brown in Glacier National Park, Montana.</figcaption></figure>
<h3 class="wp-block-heading"><strong>Keeping surrounding forests
healthy</strong></h3> <p>Forestry techniques like prescribed fire and thinning
can help whitebark pines, too. Clearing out brush and limiting excess fuel
reduces the likelihood of high-severity fires. Fires <a
href="https://www.mdpi.com/1999-4907/9/10/648">can benefit</a> the pines, but
they can <a href="https://whitebarkfound.org/fire-regimes/">also</a> harm
them. </p> <p>Whitebark pines can survive low-intensity fires that occur
in small portions of their forests thanks to their thicker bark and deeper
roots, while mixed severity fires can open up habitat for the Clark’s
nutcracker, a bird that scatters the trees’ seeds. Whitebark pine forests in
northwestern Montana, northern Idaho and the Cascades have historically
experienced periodic large fires in which trees are killed but readily
regenerate afterward. But today’s <a
href="https://www.nrfirescience.org/resource/17562">more severe, more
frequent</a> wildfires can also kill mature pines, including the rust-
resistant ones. Losing these trees to wildfire would be a big blow to
restoration efforts. <a href="https://www.mdpi.com/1999-4907/9/10/648">Recent
work</a> from the Forest Service Rocky Mountain Research Station and Missoula
Fire Sciences Laboratory suggests effective whitebark pine restoration needs
to “enhance the benefits and reduce the losses from wildfires.” </p> <h3
class="wp-block-heading"><strong>Monitoring Clark’s
nutcracker </strong></h3> <p>Whitebark pines need the Clark’s nutcracker
to disperse their seeds, sometimes as far as <a
href="https://www.fs.usda.gov/pnw/sciencef/scifi130.pdf">20 miles</a><u> from
their home trees</u>, and the Clark’s nutcrackers needs whitebark pines for
calories. The two species’ fates are intertwined. So researchers across the
West are <a
href="https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0227161">monitoring</a>
this small bird, worried about its decline as the trees die. Tracking efforts
are underway in areas like North Cascades, Mount Rainier, Yosemite and
Sequoia-Kings Canyon national parks.</p> <p>“We’re relying on the bird,” said
Diana Tomback, a professor at the University of Colorado Denver who studies
whitebark pine. “It will take multiple human lifetimes for whitebark pines’
range to be re-established, and Clark’s nutcrackers’ seed dispersal will be
the key to this.” </p> <p><em>Kylie Mohr is an editorial fellow
for </em>High Country News<em> writing from Montana. Email her
at <a href="mailto:[email protected]">[email protected]</a> or
submit a<a href="https://www.hcn.org/feedback/contact-us"> letter to the
editor</a>. See our <a href="https://www.hcn.org/policies/lte">letters to
the editor policy</a>. </em></p> <p><a
href="https://twitter.com/thatsmohrlikeit" class="twitter-follow-button" data-
show-count="false" data-size="large">Follow @thatsmohrlikeit</a></p> <p>//
</p>
This <a target="_blank" href="https://www.hcn.org/articles/north-endangered-
species-how-to-save-the-whitebark-pine/">article</a> first appeared on <a
target="_blank" href="https://www.hcn.org">High Country News</a> and is
republished here under a Creative Commons license.<img
src="https://i0.wp.com/www.hcn.org/wp-content/uploads/2023/05/cropped-
HCN_Logo-Monogram_White_Sq-2.png?fit=150%2C150&ssl=1"
style="width:1em;height:1em;margin-left:10px;"><img id="republication-tracker-
tool-source" src="https://www.hcn.org/?republication-
pixel=true&post=96480&ga4=G-0NS3WVPPTN" style="width:1px;height:1px;">
Copy to Clipboard
1
Tagged: [ Endangered Species ](https://www.hcn.org/topic/endangered-species/)
, [ Forests ](https://www.hcn.org/topic/forests/) , [ Mountain West
](https://www.hcn.org/topic/mountain-west/) , [ Plants
](https://www.hcn.org/topic/plants/) , [ U.S. Fish & Wildlife
](https://www.hcn.org/topic/u-s-fish-wildlife/) , [ U.S. Forest Service
](https://www.hcn.org/topic/u-s-forest-service/)
## [ Kylie Mohr ](https://www.hcn.org/author/kylie-mohr/)
Kylie Mohr is a correspondent for High Country News writing from Montana.
Email her at [ [email protected] ](mailto:[email protected]) or submit a
letter to the editor. See our letters to the editor policy.
[ More by Kylie Mohr ](https://www.hcn.org/author/kylie-mohr/)
Search for: Search
## Support nonprofit news
_High Country News_ relies on donations as well as subscription fees to
produce independent reporting on the West. Help continue the legacy of reader-
supported journalism by making a tax-deductible contribution today.
[ Make a contribution ](?view=donation-select&campaign_key=campaign-
subscriber-1)
_Find out more about how we use your contributions in our[ annual reports and
filings ](https://www.hcn.org/support/annual-reports/) . _
## Subscribe to High Country News
Bypass the paywall, read as much as you’d like and enable _HCN_ to continue
reporting on the West.
Subscribe now
## Get our newsletters
Sign up to receive news and updates from _High Country News_ .
Email address (Required)
Name
This field is for validation purposes and should be left unchanged.
Δ
## Most popular stories
* [ Can ice climbing bring life to an isolated Colorado town in the dead of winter? ](https://www.hcn.org/articles/can-ice-climbing-bring-life-to-an-isolated-colorado-town-in-the-dead-of-winter/)
* [ What’s next for the Owyhee Canyonlands? ](https://www.hcn.org/articles/whats-next-for-the-owyhee-canyonlands/)
* [ Are the Great Salt Lake scientists all right? ](https://www.hcn.org/articles/are-the-great-salt-lake-scientists-alright/)
* [ Wildlife habitat and tribal cultures threatened by Washington’s largest wind farm ](https://www.hcn.org/articles/wildlife-habitat-and-tribal-cultures-threatened-by-washingtons-largest-wind-farm/)
* [ Notes on not sending it ](https://www.hcn.org/issues/notes-on-not-sending-it/)
## Featured Stories
[ 
](https://www.hcn.org/articles/whats-next-for-the-owyhee-canyonlands/)
## [ What’s next for the Owyhee Canyonlands?
](https://www.hcn.org/articles/whats-next-for-the-owyhee-canyonlands/)
[ 
](https://www.hcn.org/articles/wildlife-habitat-and-tribal-cultures-
threatened-by-washingtons-largest-wind-farm/)
## [ Wildlife habitat and tribal cultures threatened by Washington’s largest
wind farm ](https://www.hcn.org/articles/wildlife-habitat-and-tribal-cultures-
threatened-by-washingtons-largest-wind-farm/)
[ 
](https://www.hcn.org/articles/meet-the-tree-sitters-who-occupied-a-ponderosa-
pine/)
## [ Meet the tree-sitters who occupied a ponderosa pine
](https://www.hcn.org/articles/meet-the-tree-sitters-who-occupied-a-ponderosa-
pine/)
## The West in Perspective
## [ Is Biden a public-lands protector? ](https://www.hcn.org/articles/is-
biden-a-public-lands-protector/)
by [ Jonathan Thompson ](https://www.hcn.org/author/jonathan-thompson/)
## [ A border need not be a wall ](https://www.hcn.org/issues/56-4/a-border-
need-not-be-a-wall/)
by [ John Washington ](https://www.hcn.org/author/john-washington/)
## [ Cattle are drinking the Colorado River dry
](https://www.hcn.org/articles/cattle-are-drinking-the-colorado-river-dry/)
by [ Jonathan Thompson ](https://www.hcn.org/author/jonathan-thompson/)
### [ About High Country News ](/about)
* [ Our history ](https://www.hcn.org/about/our-history/timeline/)
* [ How to support _HCN_ ](/support/)
* [ Submissions ](/about/submissions/)
## Know the West.
Get 2 free issues ↓

119 Grand Avenue
PO Box 1090
Paonia, CO 81428
[ (970) 527-4898 ](tel:9705274898)
* [ Facebook ](https://www.facebook.com/highcountrynews)
* [ Twitter ](https://twitter.com/highcountrynews)
* [ Instagram ](https://www.instagram.com/highcountrynews)
* [ TikTok ](https://www.tiktok.com/@highcountrynews)
* [ YouTube ](https://www.youtube.com/user/highcountrynews)
* [ LinkedIn ](https://www.linkedin.com/company/high-country-news)
* [ RSS Feed ](https://www.hcn.org/rss)
* [ Contact Us ](https://hcn.org/contact/)
* [ About us ](https://www.hcn.org/about/)
* [ Careers ](https://www.hcn.org/about/jobs/)
* [ Pitch us a story ](https://www.hcn.org/about/submissions/)
* [ Fellowships ](https://www.hcn.org/about/fellowships/)
* [ Education ](https://www.hcn.org/edu/)
* [ Support our work ](https://www.hcn.org/support/)
* [ Advertise ](https://www.hcn.org/advertising/)
* [ Syndication ](https://www.hcn.org/syndication/)
* [ Subscriber services ](https://www.hcn.org/subscriber-services/)
[ Get 2 free issues ↓
](/subscribe/?view=register&product_id=2071&plan_id=27792&coupon_code=FullTrial2&campaign_key=site_footer_label)

**[ Join now
](/subscribe/?view=register&product_id=2071&plan_id=27792&coupon_code=FullTrial2&campaign_key=site_footer)
** and become a valued member of our community. Your support helps us keep a
watch on the West.
[ Privacy Policy ](https://www.hcn.org/privacy-policy/) | [ Terms of Use
](https://www.hcn.org/terms/)
© 2024 High Country News. All rights reserved. [ Powered by Newspack
](https://newspack.com/)
| biology | 3819770 | https://sv.wikipedia.org/wiki/Holopsis%20carolinae | Holopsis carolinae | Holopsis carolinae är en skalbaggsart som först beskrevs av Thomas Casey 1900. Holopsis carolinae ingår i släktet Holopsis och familjen punktbaggar. Inga underarter finns listade i Catalogue of Life.
Källor
Externa länkar
Punktbaggar
carolinae | swedish | 1.526958 |
wake_up_wrong/Sleep_inertia.txt | Sleep inertia is a physiological state of impaired cognitive and sensory-motor performance that is present immediately after awakening. It persists during the transition of sleep to wakefulness, where an individual will experience feelings of drowsiness, disorientation and a decline in motor dexterity. Impairment from sleep inertia may take several hours to dissipate. In the majority of cases, morning sleep inertia is experienced for 15 to 30 minutes after waking.
Sleep inertia is of concern with decision-making abilities, safety-critical tasks and the ability to operate efficiently soon after awakening. In these situations, it poses an occupational hazard due to the cognitive and motor deficits that may be present.
Symptoms[edit]
"Grogginess", as defined by a drowsy or disoriented state in which there is a dampening of sensory acuity and mental processing.
Impaired motor dexterity and decrease in cognitive ability. These gross impairments may be responsible for the associated increase in reaction time and drop in attentiveness.
Deficits in spatial memory
Reports of heightened subjective fatigue
Reduced vigilance and a desire to go back to sleep
These symptoms are expressed with the greatest intensity immediately after waking, and dissipate following a period of extended wakefulness. The duration of symptoms varies on a conditional basis, with primary expression during the first 15–60 minutes after waking and potentially extending for several hours. Tasks that require more complex cognitive operations will feature greater deficits as compared to a simple motor task; the accuracy of sensory and motor functioning is more impaired by sleep inertia as compared to sheer speed. In order to measure the cognitive and motor deficiencies associated with sleep inertia, a battery of tests may be utilized, including: the psychomotor vigilance task, descending subtraction task (DST), auditory reaction time task, and the finger tapping task.
Causes[edit]
Studies have shown that abrupt awakening during stage 3 sleep, slow-wave sleep (SWS), produces more sleep inertia than awakening during sleep stages 1, 2 or REM sleep.
Prior sleep deprivation increases the percentage of time spent in slow-wave sleep (SWS). Therefore, an individual who was previously sleep deprived will have a greater chance of experiencing sleep inertia.
Adenosine levels in the brain progressively increase with sleep deprivation, and return to normal during sleep. Upon awakening with sleep deprivation, high amounts of adenosine will be bound to receptors in the brain, neural activity slows down, and a feeling of tiredness will result.
Studies show that individuals express a lack of blood flow to the brain upon awakening. Levels of cerebral blood flow (CBF) and cerebral blood flow velocities (CBFV) will take up to 30 minutes to increase and reach daytime levels. Studies using advanced imaging have shown that cerebral blood flow will return to waking levels in the brainstem and thalamus first. Then, after 15 minutes, the brain's anterior cortical regions receive normal daytime blood flow. This 15 minute time period corresponds to the sleep inertia period.
Studies show that drinking alcoholic beverages in the evening causes physiological distress upon wake up. This phenomenon is known colloquially as a hangover.
Treatments and countermeasures[edit]
This section needs more reliable medical references for verification or relies too heavily on primary sources, specifically: This section relies too much on one source, please consider adding additional reliable third party sources, like medical journal review papers or newspaper articles. Please review the contents of the section and add the appropriate references if you can. Unsourced or poorly sourced material may be challenged and removed. Find sources: "Sleep inertia" – news · newspapers · books · scholar · JSTOR (February 2019)
There has been a great deal of research into potential methods to relieve the effects of sleep inertia. The demand for remedies is driven by the occupational hazards of sleep inertia for employees who work extended shifts such as medical professionals, emergency responders, or military personnel. The motor functioning and cognitive ability of these professionals who must immediately respond to a call can pose a safety hazard in the workplace. Below are some of the various methods that have been suggested to combat sleep inertia.
Napping[edit]
When a person is sleep deprived, re-entering sleep may provide a viable route to reduce mental and physical fatigue, but it can also induce sleep inertia. In order to limit sleep inertia, one should avoid waking from the deeper stages of slow-wave sleep. The onset of slow-wave sleep occurs approximately 30 minutes after falling asleep, therefore a nap should be limited to under 30 minutes to prevent waking during slow-wave sleep and enhancing sleep inertia. Furthermore, self-awakening from a short nap was shown to relieve disorientation of sleep inertia as opposed to a forced awakening, but these results may warrant more research into the nature of arousal after sleep periods.
Caffeine[edit]
Caffeine is a xanthine derivative that can cross the blood–brain barrier, as well as the most widely-consumed stimulant compound, present in therapeutic qualities in a variety of food and drink, including tea, coffee, soft drinks and chocolate. The caffeine present in coffee or tea exerts its stimulating action by blocking adenosine receptors in the brain. By antagonizing the adenosine receptors, caffeine limits the effects of adenosine buildup in the brain and increases alertness and attentiveness. Previous research has shown that coupled with a short nap, consuming caffeine prior to the nap can alleviate the effects of sleep inertia. Nonetheless, individual degree of consumption and tolerance to caffeine may be responsible for variation in its efficacy to reduce sleep inertia symptoms.
Light[edit]
The natural light provided by the sunrise may contribute to a reduction in sleep inertia effects. Research simulating increase of light at dawn was shown to potentiate the cortisol awakening response (CAR). The CAR is a spike in blood cortisol levels following awakening, and is associated with the return to an alert cognitive state.
Other[edit]
Some other interventions that could potentially minimize the effects of sleep inertia are sound and temperature. There is moderate evidence that the presence of mild sounds and a sharp decrease in the temperature of the extremities may independently reverse sleep inertia symptoms. Sound, especially music, is thought to increase attentiveness and decrease one's subjective feeling of sleepiness upon awakening. A drop in temperature of the extremities may prevent heat loss upon awakening, facilitating the return of core body temperature to homeostatic daytime levels.
See also[edit]
Circadian rhythm sleep disorder
Delayed sleep phase syndrome
Shift work | biology | 67513 | https://da.wikipedia.org/wiki/S%C3%B8vn | Søvn | Søvn er en regelmæssigt tilbagevendende tilstand af naturlig hvile, som muliggør restitution af krop og sind, samt hjernens lagring af nyligt indlærte ting.
Søvnprocessen er livsnødvendig for alle pattedyr og fugle. Det er stadig uklart, om andre dyr (fisk, reptiler og insekter) sover. Søvn er en fase af legemets døgnrytme, som er baseret på en ca. 24 timers cyklus. Behov for søvn er meget individuelt. Nogle voksne har brug for 8-9 timers søvn, mens andre kan klare sig med 5-6 timer. Børn har generelt brug for mere søvn end voksne.
Søvnen har ikke altid varet 8 timer i træk. Tidligere har man opdelt søvnen i 4 timer to gange dagligt. Der er fx fundet mindst 500 historiske tekster af Roger Ekirch på, at mennesker før den industrielle revolution, har sovet af to omgange i døgnet; fra skumring til et tidspunkt i natten og brugtes tiden til at bede til gud, ryge eller have sex. Og man er herefter gået i seng igen og stået op ved morgengry.
Søvnen deles op i forskellige faser, som afløser hinanden: døs, let søvn, slow-wave søvn (dyb søvn) og deltasøvn (dybeste del). Hvis man sover 8 timer i døgnet, består søvnen typisk af 2 timers dyb søvn, 2 timers REM søvn og 4 timers let søvn. Perioderne afløser hinanden i faser på ca. 1½ times varighed. I REM faserne er der voldsom aktivitet i hjernen, og der drømmes mere end i andre faser.
Der er to former for søvnkvalitet. Den ene er den subjektive del som er følelsen af være udhvilet og har haft en god nattesøvn.
En søvncyklus foregår over 5 stadier, som gentages ca. 4-5 gange i løbet af natten. Ved første stadie sover man meget let, og har let ved at vågne, og hvis man vågner i det her stadie, vil man tro, at man slet ikke har sovet. Herefter kommer man til næste stadie, som gør at man stadig sover let, men alligevel en smule tungere. I dette stadie vågner man sjældent spontant. Efter en halv time kommer man til tredje stadie, hvor man nu sover tungt. Her falder hjerneaktiviteten og hjertefrekvens. I det her stadie har man ingen drømme, og man er svær at vække op fra søvnen. Det er her, at kroppen genopbygger sig. I det næste stadie er det rem-søvnen. Det er her, at man drømmer og ens hjerne er lige så aktiv, som hvis man havde været vågen. Man husker ofte drømme man har lige haft, når man vågner. En cyklus tager 90-110 minutter.
Den objektive søvnkvalitet har man ved at måle de forskellige søvnstadier. Man ser især på hvor meget man befinder sig i dyb søvn og antallet af opvågninger i løbet af natten.
I øjeblikket omkring man falder i søvn kan der forekomme små ryk i kroppen kaldet søvnryk.
Formål
Formålet med søvn er at give hvile, mens kroppen genopbygges og hjernen bearbejder dagens indtryk som gemmes i hukommelsen. Der findes flere hypoteser om, hvorfor søvn er så vigtig. Én af dem er, at hjernen har brug for at være offline, for at være frisk til at lagre information, når vi er vågne. En anden hypotese er, at søvnen fjerner affaldsstoffer fra hjernen. Hvis de ikke fjernes ophobes de, og kan øge risikoen for forskellige sygdomme. Uanset forklaringen er der dog ingen tvivl om, at søvn er vigtigt.
Fysiologi
De mest udtalte fysiologiske ændringer i søvn forekommer i hjernen. Hjernen bruger markant mindre energi under søvn end den gør, når den er vågen, især under ikke-REM-søvn. I områder med nedsat aktivitet gendanner hjernen sin forsyning med adenosintrifosfat (ATP), molekylet, der bruges til kortvarig opbevaring og transport af energi.
REM-søvn
REM-søvnen er den dybeste søvn, som vi mennesker oplever under en normal nattesøvn. Den er en af de fire søvnfaser, man kommer igennem hver nat. Den har en helt afgørende betydning for søvnens kvalitet.
Udtrykket REM er en forkortelse for “rapid eye movement”. Det er opstået fordi under denne søvnfase, sker drømmene og derfor er vores hjerneaktivitet også forhøjet.
Man befinder sig i REM-søvnen flere gange i løbet af en nat, og første gang kommer typisk efter man har sovet i 90 minutter. REM- søvnen varer mellem 10 minutter og en time ad gangen.
Fordi hjernen er meget aktiv, sover man meget let i denne fase - det er også derfor, at man tit vågner midt i en drøm.
Manglende søvn
Konsekvenserne af manglende eller dårlig søvn, kan godt være til at overskue hvis ikke det foregår over en længere periode. Men hvis man kommer i søvnunderskud over længere tid, kan det få alvorlige konsekvenser for den fysiske og mentale sundhed.
Manglende søvn over en længere periode kan nedsætte immunforsvaret og øge risikoen for; infektioner, lavere metabolisme, overvægt, diabetes 2, irritabelt humør, koncentrationsbesvær, psykiske sygdomme(fx depression og angst), hjerteproblemer, fremskynde udviklingen af Alzheimers -og i sin yderste konsekvens forårsage en for tidlig død.
For børn kan manglende søvn have en negativ konsekvens for barnets udvikling og hvis der er tale om søvnbesvær hos barnet, kan det være hensigtsmæssigt at iværksætte visse tiltag for at forbedre søvnen. Det kan være nedgearing før sengetid, mindske stimuli i løbet af dagen, massage og tryk på led og muskler eller introduktion af en tyngdedyne, som dæmper barnets motoriske uro og giver lettere indsovning og færre opvågninger i løbet af natten.
Se også
Biologisk ur
Søvn hos dyr
Søvngængeri
Referencer
Eksterne henvsninger
Videnskab.dks tværvidenskabelige tema om Søvn
Fysiologi
Psykologi
Uløste problemer indenfor neurovidenskab | danish | 0.617238 |
wake_up_wrong/s4010102200317w.txt | Skip to main content
Advertisement
[

](//pubads.g.doubleclick.net/gampad/jump?iu=/270604982/bmc/jphysiolanthropol/articles&sz=728x90,970x90&pos=LB1&doi=10.1186/s40101-022-00317-w&type=article&kwrd=Sleep
loss,Snooze alarm,Awakening,Sleep inertia,Normal nocturnal
sleep,Polysomnograms&pmc=B13004,X12000&)
[   ](https://www.biomedcentral.com)
Search
* [ Explore journals ](//www.biomedcentral.com/journals)
* [ Get published ](//www.biomedcentral.com/getpublished)
* [ About BMC ](//www.biomedcentral.com/about)
* [ My account ](https://www.biomedcentral.com/account)
Search all BMC articles
Search
[ Journal of Physiological Anthropology ](/)
* [ Home ](/)
* [ About ](/about)
* [ Articles ](/articles)
* [ Submission Guidelines ](/submission-guidelines)
* [ Awards ](/awards)
* [ Submit manuscript ](https://www.editorialmanager.com/jpha/)
Effects of using a snooze alarm on sleep inertia after morning awakening
[ Download PDF
](//jphysiolanthropol.biomedcentral.com/counter/pdf/10.1186/s40101-022-00317-w.pdf)
[ Download ePub
](//jphysiolanthropol.biomedcentral.com/counter/epub/10.1186/s40101-022-00317-w.epub)
[ Download PDF
](//jphysiolanthropol.biomedcentral.com/counter/pdf/10.1186/s40101-022-00317-w.pdf)
[ Download ePub
](//jphysiolanthropol.biomedcentral.com/counter/epub/10.1186/s40101-022-00317-w.epub)
* Original article
* [ Open access ](https://www.springernature.com/gp/open-research/about/the-fundamentals-of-open-access-and-open-research)
* Published: 31 December 2022
# Effects of using a snooze alarm on sleep inertia after morning awakening
* Keiko Ogawa [ ORCID: orcid.org/0000-0001-9251-1884 ](http://orcid.org/0000-0001-9251-1884) 1 ,
* Emi Kaizuma-Ueyama 2 &
* Mitsuo Hayashi 1
[ _Journal of Physiological Anthropology_ ](/) ** volume 41 ** , Article
number: 43 ( 2022 ) Cite this article
* 7104 Accesses
* 1 Citations
* 178 Altmetric
* [ Metrics details ](/articles/10.1186/s40101-022-00317-w/metrics)
## Abstract
### Background
Many people use the snooze function of digital alarm clocks for morning
awakening, but the effects of a snooze alarm on waking are unclear. We
examined the effects of a snooze alarm on sleep inertia, which is a
transitional state characterized by reduced arousal and impaired cognitive and
behavioral performance immediately upon awakening.
### Methods
In study 1, healthy Japanese university students responded to a sleep survey
during a psychology class (study 1), and we collected 293 valid responses. In
study 2, we compared a separate sample of university students ( _n_ = 10) for
the effects of using or not using a snooze alarm on sleep inertia immediately
after awakening from normal nocturnal sleep in a sleep laboratory.
### Results
Of 293 valid respondents in study 1, 251 often used a tool to wake up in the
morning (85.7%). Moreover, 70.5% reported often using the snooze function of
their mobile phones, mainly to reduce anxiety about oversleeping. Study 2
indicated no differences in the sleep quality or quantity before awakening
with or without the snooze alarm, except in the last 20 min. However, during
the last 20 min of sleep with snooze alarm, the snooze alarm prolonged waking
and stage N1 sleep. Stage N1 sleep is non-rapid eye movement sleep that is
primarily defined as a drowsy state. Furthermore, Global Vigor values were
enhanced after awakening compared to pre-sleep in the no-snooze condition.
### Conclusions
Using a snooze alarm prolongs sleep inertia compared to a single alarm,
possibly because snooze alarms induce repeated forced awakenings.
## Introduction
Today, the nocturnal sleep duration has become shorter for most people. The
Japan Broadcasting Corporation (NHK) reported in 2020 that the mean sleep time
of Japanese people was 7 h 12 min [ [ 1
](/articles/10.1186/s40101-022-00317-w#ref-CR1 "NHK Broadcasting Culture
Research Institute. National Time Use Survey 2020.
https://www.nhk.or.jp/bunken/yoron-jikan/column/sleep-2020.html
. Accessed 8 May 2022.") ], down from 8 h 13 min in 1960 [ [ 2 ](/articles/10.1186/s40101-022-00317-w#ref-CR2 "NHK Broadcasting Culture Research Institute. National Time use Survey 2015. Tokyo: Japan Broadcast Publishing Co. Ltd; 2016.") ]. Sleep loss (deprivation) has led to increased daytime sleepiness and bad moods on awakening [ [ 3 ](/articles/10.1186/s40101-022-00317-w#ref-CR3 "Carskadon MA, Dement WC. Sleep-related breathing disorders in elderly adults. J Gerontol. 1982;37\(5\):637.") ], with impairments of specific cognitive function [ [ 4 ](/articles/10.1186/s40101-022-00317-w#ref-CR4 "Havekes R, Vecsey CG, Abel T. The impact of sleep deprivation on neuronal and glial signaling pathways important for memory and synaptic plasticity. Cell Signal. 2012;24\(6\):1251–60.
https://doi.org/10.1016/j.cellsig.2012.02.010
.") , [ 5 ](/articles/10.1186/s40101-022-00317-w#ref-CR5 "McCoy JG, Strecker RE. The cognitive cost of sleep lost. Neurobiol Learn Mem. 2011;96\(4\):564–82.
https://doi.org/10.1016/j.nlm.2011.07.004
.") ] and decreased immune, inflammatory and cardiovascular functions [ [ 6 ](/articles/10.1186/s40101-022-00317-w#ref-CR6 "Faraut B, Touchette E, Gamble H, et al. Short sleep duration and increased risk of hypertension: a primary care medicine investigation. J Hypertens. 2012;30\(7\):1354–63.
https://doi.org/10.1097/HJH.0b013e32835465e5
.") ]. Moreover, sleep loss (deprivation) increases sleep inertia [ [ 7 ](/articles/10.1186/s40101-022-00317-w#ref-CR7 "Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG spectral power and cognitive performance during sleep inertia: the effect of normal sleep duration and partial sleep deprivation. Physiol Behav. 2006;87\(1\):177–84.
https://doi.org/10.1016/j.physbeh.2005.09.017
.") , [ 8 ](/articles/10.1186/s40101-022-00317-w#ref-CR8 "Tassi P, Muzet A. Sleep inertia. Sleep Med Rev. 2000;4\(4\):341–53.
https://doi.org/10.1053/smrv.2000.0098
.") ], which is a transitional state of reduced arousal and impaired cognitive and behavioral performance [ 7 , 8 , 9 , 10 , [ 11 ](/articles/10.1186/s40101-022-00317-w#ref-CR11 "Ikeda H, Hayashi M. The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol. 2010;83\(1\):15–9.
https://doi.org/10.1016/j.biopsycho.2009.09.008
.") ] immediately upon awakening.
Sleep inertia appears after normal nocturnal sleep [ [ 8
](/articles/10.1186/s40101-022-00317-w#ref-CR8 "Tassi P, Muzet A. Sleep
inertia. Sleep Med Rev. 2000;4\(4\):341–53.
https://doi.org/10.1053/smrv.2000.0098
.") , [ 11 ](/articles/10.1186/s40101-022-00317-w#ref-CR11 "Ikeda H, Hayashi M. The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol. 2010;83\(1\):15–9.
https://doi.org/10.1016/j.biopsycho.2009.09.008
.") ], and its duration rarely exceeds 30 min [ [ 8 ](/articles/10.1186/s40101-022-00317-w#ref-CR8 "Tassi P, Muzet A. Sleep inertia. Sleep Med Rev. 2000;4\(4\):341–53.
https://doi.org/10.1053/smrv.2000.0098
.") ]. Many factors increase the intensity and duration of sleep inertia. These factors include prior sleep deprivation [ [ 8 ](/articles/10.1186/s40101-022-00317-w#ref-CR8 "Tassi P, Muzet A. Sleep inertia. Sleep Med Rev. 2000;4\(4\):341–53.
https://doi.org/10.1053/smrv.2000.0098
.") ], the length of waking time prior to sleep [ [ 12 ](/articles/10.1186/s40101-022-00317-w#ref-CR12 "Dinges DF, Orne MT, Whitehouse WG, Orne EC. Temporal placement of a nap for alertness: contributions of circadian phase and prior wakefulness. Sleep. 1987;10\(4\):313–29.") ], the sleep stage at awakening [ [ 7 ](/articles/10.1186/s40101-022-00317-w#ref-CR7 "Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG spectral power and cognitive performance during sleep inertia: the effect of normal sleep duration and partial sleep deprivation. Physiol Behav. 2006;87\(1\):177–84.
https://doi.org/10.1016/j.physbeh.2005.09.017
.") , [ 13 ](/articles/10.1186/s40101-022-00317-w#ref-CR13 "Cavallero C, Versace F. Stage at awakening, sleep inertia and performance. Sleep Res Online. 2003;3:89–97.") ], circadian timing of awakening [ [ 14 ](/articles/10.1186/s40101-022-00317-w#ref-CR14 "McHill AW, Hull JT, Cohen DA, Wang W, Czeisler CA, Klerman EB. Chronic sleep restriction greatly magnifies performance decrements immediately after awakening. Sleep. 2019;42\(5\):zsz032.
https://doi.org/10.1093/sleep/zsz032
.") , [ 15 ](/articles/10.1186/s40101-022-00317-w#ref-CR15 "Scheer FA, Shea TJ, Hilton MF, Shea SA. An endogenous circadian rhythm in sleep inertia results in greatest cognitive impairment upon awakening during the biological night. J Biol Rhythm. 2008;23\(4\):353–61.
https://doi.org/10.1177/0748730408318081
.") ], the amount of non-rapid eye movement (NREM) slow-wave sleep (SWS) [ [ 10 ](/articles/10.1186/s40101-022-00317-w#ref-CR10 "Ferrara M, De Gennaro L, Ferlazzo F, Curcio G, Barattucci M, Bertini M. Auditory evoked responses upon awakening from sleep in human subjects. Neurosci Lett. 2001;310\(2-3\):145–8.
https://doi.org/10.1016/s0304-3940\(01\)02107-3
.") ], and the method of waking [ [ 11 ](/articles/10.1186/s40101-022-00317-w#ref-CR11 "Ikeda H, Hayashi M. The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol. 2010;83\(1\):15–9.
https://doi.org/10.1016/j.biopsycho.2009.09.008
.") , [ 16 ](/articles/10.1186/s40101-022-00317-w#ref-CR16 "Kaida K, Nakano E, Nittono H, Hayashi M, Hori T. The effects of self-awakening on heart rate activity in a short afternoon nap. Clin Neurophysiol. 2003;114\(10\):1896–901.
https://doi.org/10.1016/s1388-2457\(03\)00167-6
.") , [ 17 ](/articles/10.1186/s40101-022-00317-w#ref-CR17 "Matsuura N, Hayashi M. Effects of habitual self-awakening on nocturnal sleep, autonomic activity prior to awakening, and subjective condition after awakening. Sleep Biol Rhythms. 2009;7\(3\):172–80.") ]. Sleep deprivation studies on the relationship between sleep loss and sleep inertia have indicated that sleep inertia worsens on partial sleep deprivation (2 h) nights compared to sufficient sleep (8 h) nights [ [ 7 ](/articles/10.1186/s40101-022-00317-w#ref-CR7 "Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG spectral power and cognitive performance during sleep inertia: the effect of normal sleep duration and partial sleep deprivation. Physiol Behav. 2006;87\(1\):177–84.
https://doi.org/10.1016/j.physbeh.2005.09.017
.") ]. Moreover, McHill et al. [ [ 14 ](/articles/10.1186/s40101-022-00317-w#ref-CR14 "McHill AW, Hull JT, Cohen DA, Wang W, Czeisler CA, Klerman EB. Chronic sleep restriction greatly magnifies performance decrements immediately after awakening. Sleep. 2019;42\(5\):zsz032.
https://doi.org/10.1093/sleep/zsz032
.") ] reported that performance was worse after chronic restricted sleep when sleep opportunities for a 24-h day were 5.6 h compared to normal sleep when sleep opportunity for a 24-h day was 8 h. Research on the relationship between sleep stages and sleep inertia has reported that performance decreased upon waking from SWS sleep compared to N1, N2, and REM sleep [ [ 13 ](/articles/10.1186/s40101-022-00317-w#ref-CR13 "Cavallero C, Versace F. Stage at awakening, sleep inertia and performance. Sleep Res Online. 2003;3:89–97.") , [ 18 ](/articles/10.1186/s40101-022-00317-w#ref-CR18 "Bruck D, Pisani DL. The effects of sleep inertia on decision-making performance. J Sleep Res. 1999;8\(2\):95–103.") , [ 19 ](/articles/10.1186/s40101-022-00317-w#ref-CR19 "Stampi C. The effects of polyphasic and ultrashort sleep schedules. In: Stampi C, editor. Why we nap: evolution, chronobiology and functions of Polyphasic and Ultrashort sleep. Boston: Birkhäuser; 1992. p. 137–79.") ]. Studies on the effect of endogenous circadian cycles on sleep inertia indicated that sleep inertia is worse after nights when the core body temperature is lower than the day [ [ 14 ](/articles/10.1186/s40101-022-00317-w#ref-CR14 "McHill AW, Hull JT, Cohen DA, Wang W, Czeisler CA, Klerman EB. Chronic sleep restriction greatly magnifies performance decrements immediately after awakening. Sleep. 2019;42\(5\):zsz032.
https://doi.org/10.1093/sleep/zsz032
.") , [ 15 ](/articles/10.1186/s40101-022-00317-w#ref-CR15 "Scheer FA, Shea TJ, Hilton MF, Shea SA. An endogenous circadian rhythm in sleep inertia results in greatest cognitive impairment upon awakening during the biological night. J Biol Rhythm. 2008;23\(4\):353–61.
https://doi.org/10.1177/0748730408318081
.") ]. Studies have also suggested that the effect of sleep stages and circadian timing on sleep inertia was influenced by sleep loss (deprivation) [ [ 20 ](/articles/10.1186/s40101-022-00317-w#ref-CR20 "Hilditch CJ, McHill AW. Sleep inertia: current insights. Nat Sci Sleep. 2019;11:155–65 Published 2019 Aug 22. 10.2147/NSS.S188911.") ].
Reducing sleep inertia helps people awaken refreshed in the morning. Research
on factors influencing sleep inertia has suggested strategies for reducing
sleep inertia, including sleeping for an optimal duration and waking up from
light sleep. Moreover, self-awakening (SA), a method of waking up at a
predetermined time without an alarm [ [ 11
](/articles/10.1186/s40101-022-00317-w#ref-CR11 "Ikeda H, Hayashi M. The
effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol.
2010;83\(1\):15–9.
https://doi.org/10.1016/j.biopsycho.2009.09.008
.") , [ 16 ](/articles/10.1186/s40101-022-00317-w#ref-CR16 "Kaida K, Nakano E, Nittono H, Hayashi M, Hori T. The effects of self-awakening on heart rate activity in a short afternoon nap. Clin Neurophysiol. 2003;114\(10\):1896–901.
https://doi.org/10.1016/s1388-2457\(03\)00167-6
.") , [ 17 ](/articles/10.1186/s40101-022-00317-w#ref-CR17 "Matsuura N, Hayashi M. Effects of habitual self-awakening on nocturnal sleep, autonomic activity prior to awakening, and subjective condition after awakening. Sleep Biol Rhythms. 2009;7\(3\):172–80.") , [ 21 ](/articles/10.1186/s40101-022-00317-w#ref-CR21 "Moorcroft WH, Kayser KH, Griggs AJ. Subjective and objective confirmation of the ability to self-awaken at a self-predetermined time without using external means. Sleep. 1997;20\(1\):40–5.
https://doi.org/10.1093/sleep/20.1.40
.") ], is an effective strategy for reducing sleep inertia. People who plan to SA show increasing sympathetic nervous activity before waking up, suggesting that the body prepares for waking up before sleep termination [ [ 22 ](/articles/10.1186/s40101-022-00317-w#ref-CR22 "Born J, Hansen K, Marshall L, Mölle M, Fehm HL. Timing the end of nocturnal sleep. Nature. 1999;397\(6714\):29–30.
https://doi.org/10.1038/16166
.") , [ 23 ](/articles/10.1186/s40101-022-00317-w#ref-CR23 "Kräuchi K, Cajochen C, Wirz-Justice A. Waking up properly: is there a role of thermoregulation in sleep inertia? J Sleep Res. 2004;13\(2\):121–7.
https://doi.org/10.1111/j.1365-2869.2004.00398.x
.") ]. For example, Kaida et al. [ [ 16 ](/articles/10.1186/s40101-022-00317-w#ref-CR16 "Kaida K, Nakano E, Nittono H, Hayashi M, Hori T. The effects of self-awakening on heart rate activity in a short afternoon nap. Clin Neurophysiol. 2003;114\(10\):1896–901.
https://doi.org/10.1016/s1388-2457\(03\)00167-6
.") ] observed that the heart rate gradually increased before SA and suggested that the increased heart rate facilitates a smoother transition from sleep to awakening. Allen [ [ 24 ](/articles/10.1186/s40101-022-00317-w#ref-CR24 "Allen RP. Article reviewed: timing the end of nocturnal sleep. Sleep Med. 2001;2\(1\):69–70.
https://doi.org/10.1016/s1389-9457\(00\)00090-3
.") ] indicated that adrenocorticotropin release before awakening, which facilitates sympathetic nervous activity, reduces sleep inertia. In Japan, only 10.3% of university students and 18.9% of workers reported SA [ [ 17 ](/articles/10.1186/s40101-022-00317-w#ref-CR17 "Matsuura N, Hayashi M. Effects of habitual self-awakening on nocturnal sleep, autonomic activity prior to awakening, and subjective condition after awakening. Sleep Biol Rhythms. 2009;7\(3\):172–80.") ]. Mattingly et al. [ [ 25 ](/articles/10.1186/s40101-022-00317-w#ref-CR25 "Mattingly SM, Martinez G, Young J, Cain MK, Striegel A. Snoozing: an examination of a common method of waking. Sleep. 2022;45\(10\):zsac184.
https://doi.org/10.1093/sleep/zsac184
.") ] investigated the effect of sleep duration on the waking method of 385 full-time workers in the USA. They reported that the sleep duration on natural wake days (mean sleep duration: 8.74h ± 3.77 min) was significantly longer than on days when participants used an alarm or a snooze alarm to wake up (mean sleep duration: 7.83h ± 3.54 min for the alarm and 7.95 h ± 3.66 min for the snooze alarm). Studies on Japanese people have suggested that only a few Japanese use SA [ [ 17 ](/articles/10.1186/s40101-022-00317-w#ref-CR17 "Matsuura N, Hayashi M. Effects of habitual self-awakening on nocturnal sleep, autonomic activity prior to awakening, and subjective condition after awakening. Sleep Biol Rhythms. 2009;7\(3\):172–80.") ] because they have a short sleep time [ [ 26 ](/articles/10.1186/s40101-022-00317-w#ref-CR26 "Organization for Economic Co-operation and Development \(OECD\). Social protection and well-being. Time use; 2020.
https://stats.oecd.org/Index.aspx?DataSetCode=TIME_USE#
. Accessed 8 May 2022") , [ 27 ](/articles/10.1186/s40101-022-00317-w#ref-CR27 "Steptoe A, Peacey V, Wardle J. Sleep duration and health in young adults. Arch Intern Med. 2006;166\(16\):1689–92.
https://doi.org/10.1001/archinte.166.16.1689
.") ]. It is also possible that many Japanese people use external tools such as alarms to wake up in the morning because of sleep loss and difficulties in awakening.
The snooze function of alarm clocks helps avoid oversleeping after turning off
the alarm. Some people use the snooze function repeatedly after the first
alarm before they awaken, presumably to awaken gradually and gently. Mattingly
et al. [ [ 25 ](/articles/10.1186/s40101-022-00317-w#ref-CR25 "Mattingly SM,
Martinez G, Young J, Cain MK, Striegel A. Snoozing: an examination of a common
method of waking. Sleep. 2022;45\(10\):zsac184.
https://doi.org/10.1093/sleep/zsac184
.") ] reported that 57% of 450 participants habitually used a snooze alarm, and they (snoozers) had a lighter sleep and a higher resting HR across the whole night and in the last hour of sleep than no-snoozers. Self-awakening and sleep inertia studies have shown that light sleep [ [ 13 ](/articles/10.1186/s40101-022-00317-w#ref-CR13 "Cavallero C, Versace F. Stage at awakening, sleep inertia and performance. Sleep Res Online. 2003;3:89–97.") , [ 18 ](/articles/10.1186/s40101-022-00317-w#ref-CR18 "Bruck D, Pisani DL. The effects of sleep inertia on decision-making performance. J Sleep Res. 1999;8\(2\):95–103.") , [ 19 ](/articles/10.1186/s40101-022-00317-w#ref-CR19 "Stampi C. The effects of polyphasic and ultrashort sleep schedules. In: Stampi C, editor. Why we nap: evolution, chronobiology and functions of Polyphasic and Ultrashort sleep. Boston: Birkhäuser; 1992. p. 137–79.") ] and increased HR before awakening [ [ 24 ](/articles/10.1186/s40101-022-00317-w#ref-CR24 "Allen RP. Article reviewed: timing the end of nocturnal sleep. Sleep Med. 2001;2\(1\):69–70.
https://doi.org/10.1016/s1389-9457\(00\)00090-3
.") ] effectively reduce sleep inertia. On the other hand, Mattingly et al. [ [ 25 ](/articles/10.1186/s40101-022-00317-w#ref-CR25 "Mattingly SM, Martinez G, Young J, Cain MK, Striegel A. Snoozing: an examination of a common method of waking. Sleep. 2022;45\(10\):zsac184.
https://doi.org/10.1093/sleep/zsac184
.") ] expressed concern that lighter sleep across the whole night over the long term might lead to chronic sleep loss. Moreover, an increased resting HR is associated with numerous adverse health effects, including diabetes, heart disease, and mortality. Therefore, Mattingly et al. concluded that research is needed to clarify the effect of the snooze function on human health and sleep.
The present study investigated the efficacy of the snooze function in alarm
clocks on sleep inertia. We conducted a two-part study in which study 1
surveyed participants on the prevalence of using snooze alarms and the daily
settings of snooze alarms. Then, study 2 examined the effects of using a
snooze alarm on sleep inertia after awakening from nocturnal sleep in a
smaller group of participants in a sleep laboratory. We hypothesized that
sleep inertia would decrease in the snooze condition more than in the no-
snooze condition if the physiological effects of snooze alarms were similar to
self-awakening.
## Method: study 1
### Participants
Japanese university students ( _n_ = 296, 169 women; 127 men: age range 18–28
years) attending a psychology class responded to a questionnaire inquiring
about their sleeping and waking habits. The research protocol of this study
was approved by the Research Ethics Committee of Hiroshima University (No.
30-02). The participants were briefed about the study content, and they gave
their informed consent before participating in the study.
### Procedure
Participants responded to a range of questions regarding their sleep habits
and alarm clocks and mobile phone’s snooze functions use on a four-point scale
comprising 1 (often), 2 (sometimes), 3 (few times), or 4 (never). The
questions included “How many minutes before the actual waking time do you set
the alarm?” “How many times do you hit the snooze button between the first and
the last alarm (the wake-up time)?” The participants also gave free responses
to the question, “Why do you use the snooze function?”
## Results: study 1
Tables [ 1 ](/articles/10.1186/s40101-022-00317-w#Tab1) and [ 2
](/articles/10.1186/s40101-022-00317-w#Tab2) show the number of participants
using a tool to wake up in the morning. Data from three participants were
removed from the analysis due to missing values. Of the remaining 293
participants, 251 participants (85.7%) reported they often used a tool to wake
up in the morning, and fewer students reported that they used a tool
“sometimes” (9.6%), “a few times” (3.4%), or “never” (1.4%; _n_ = 4) (Table [
1 ](/articles/10.1186/s40101-022-00317-w#Tab1) ). Moreover, 204 (70.5%) among
289 students, after excluding those who never used a tool to wake up, reported
that they often used the phone’s snooze function, and fewer students reported
that they did so sometimes (9.0%), a few times (6.3%) or never (14.2%; _n_ =
41) (Table [ 1 ](/articles/10.1186/s40101-022-00317-w#Tab1) ). Among the
snooze function users ( _n_ = 248), 47.5% reported using it seven times a
week, 10.0% six times, 26.7% five times, and 15.9% less than four times (Table
[ 2 ](/articles/10.1186/s40101-022-00317-w#Tab2) ). The two main reasons for
using the snooze function were concerns about not awakening on time with a
once-only alarm (50.7%) and the desire to feel secure about waking up before
going to sleep (35.6%).
**Table 1 Participants’ using a tool and the snooze function to wake up**
[ Full size table ](/articles/10.1186/s40101-022-00317-w/tables/1)
**Table 2 Participants’ use of different snooze settings**
[ Full size table ](/articles/10.1186/s40101-022-00317-w/tables/2)
Of the participants that reported using the snooze function, 211 responded to
all the questions about the snooze settings. Among them, 184 participants
(87.2%) set the snooze alarm to ring 30 min before the predetermined waking
time, 11.4% set it to ring 30–60 min before the waking time, and 1.4% set it
to ring over 60 min before the waking time (Table [ 2
](/articles/10.1186/s40101-022-00317-w#Tab2) ). The mean time for setting the
snooze alarm before the predetermined waking time was 20.3 min (SE = 1.3). The
mean frequency of using the snooze alarm was 6.2 (SE = 0.2) times per morning
(the mode = 4 and median = 5). The mean interval between snooze alarms was 7.0
(SE = 0.4) min (mode and median were 5 min).
## Method: study 2
Study 2 compared the effects of using or not using a snooze alarm on sleep
inertia immediately after awakening from normal nocturnal sleep. The
participants in study 2 did not participate in study 1.
### Participants
Study 2 used the same questionnaire as in study 1 to pre-screen ten healthy
Japanese university students (5 women, 5 men: age range 21–26 years) who
reported using an external tool to wake up and used a mobile phone’s snooze
function 5 to 7 times per week ( _M =_ 6.2, SE = 0.3). We conducted a post hoc
power analysis using G*Power 3.1.9.6 [ [ 28
](/articles/10.1186/s40101-022-00317-w#ref-CR28 "Faul F, Erdfelder E, Buchner
A, Lang AG. Statistical power analyses using G*power 3.1: tests for
correlation and regression analyses. Behav Res Methods. 2009;41\(4\):1149–60.
https://doi.org/10.3758/BRM.41.4.1149
.") ]. We assumed the standard criteria for significance (α = .05) and small-to-medium effect size ( _f_ = .23) for a repeated-measures analysis of variance (ANOVA). The power analysis indicated that the 1–β error probability = .47. Study 2’s protocol was approved by the Research Ethics Committee of Hiroshima University (No. 30-03). All participants gave their written informed consent to participate in the study after a briefing about its content.
### Measures
#### Global vigor (GV) and global affect (GA) scores
We assessed the participants’ Global Vigor Scores (GV) to evaluate their
alertness, sleepiness, motivation, loss of effort, and weariness. We also
evaluated their Global Affect Scores (GA) for happiness, sadness, calmness,
and tension [ [ 29 ](/articles/10.1186/s40101-022-00317-w#ref-CR29 "Monk TH. A
visual analogue scale technique to measure global vigor and affect. Psychiatry
Res. 1989;27\(1\):89–99.
https://doi.org/10.1016/0165-1781\(89\)90013-9
.") ]. The participants rated their vigor and affect on the GV and GA before bedtime and after awakening using a Visual Analog Scale ranging from 0 (not at all) to 100 (absolutely) [ [ 30 ](/articles/10.1186/s40101-022-00317-w#ref-CR30 "Monk TH. Subjective ratings of sleepiness--the underlying circadian mechanisms. Sleep. 1987;10\(4\):343–53.
https://doi.org/10.1093/sleep/10.4.343
.") ]. We also used the Japanese version [ [ 31 ](/articles/10.1186/s40101-022-00317-w#ref-CR31 "Shimizu H, Imae K. Development of the Japanese edition of the Spielberger state–trait anxiety inventory \(STAI\) for student use. Japanese J Educ Psychol. 1981;29:348–53 Japanese.") ] of Spielberger’s [ [ 32 ](/articles/10.1186/s40101-022-00317-w#ref-CR32 "Spielberger CD, Gorsuch RL, Lushene E. Manual for the state-trait anxiety inventory \(self-evaluation questionnaire\). Palo Alto: Consulting Psychologists Press; 1970.") ] State Anxiety Scale (STAI) to assess the participants’ anxiety at bedtime. Participants also completed the standardized and revised version of the Oguri-Shirakawa-Azumi (OSA) Sleep Inventory for Middle-Aged Respondents (OSA-MA, [ [ 33 ](/articles/10.1186/s40101-022-00317-w#ref-CR33 "Yamamoto Y, Tanaka H, Takase M, Yamazaki K, Azumi K, Shirakawa S. Chuukounen/koureisya wo taisyoutoshita OSA suiminchousahyou \(MA ban\) no kaihatu to hyoujyunka. \[\(standardization of revised version of OSA sleep inventory for middle age and aged.\) \(in Japanese with English abstract.\)\]. Brain Sci Mental Disord. 1999;10:401–9 Japanese.") ]) immediately after waking up. They rated the condition of five variables on the OSA-MA using a four-point semantic differential (SD) scale. The variables assessed were sleepiness on rising (items included, “I can concentrate,” “I feel a freedom,” “I have a clear head,” and “I can answer quickly.”); initiation and maintenance (items included “I slept well,” “I had a light sleep,” “I dozed a lot,” “I fell asleep quickly,” and “I woke up often.”); frequent dreaming (items included, “I had many nightmares,” and “I dreamed often.”), refreshment (items included, “I am still tired,” “I am tired all over,” “I have a good appetite,” and “I feel bad.”); sleep length (items included, “I had a long sleep.”).
#### Objective performance task
We used this task to measure the participants’ alertness objectively. The task
is a simple auditory reaction time task using only one auditory stimulus (a
computer-generated 66 dB beep tone). Previously studies have used this task to
examine sleep inertia and vigilance [ [ 13
](/articles/10.1186/s40101-022-00317-w#ref-CR13 "Cavallero C, Versace F. Stage
at awakening, sleep inertia and performance. Sleep Res Online. 2003;3:89–97.")
]. The task requires a participant to press a button as quickly as possible
after hearing an auditory stimulus at random intervals of 2–8 s (mean interval
5 s). One trial of this task comprised one auditory stimulus, and 60 trials
comprised a task block that lasted 5 min. Participants performed one task
block in the pre-sleep session and three in the post-sleep session.
#### Polysomnogram
We recorded standard polysomnograms to evaluate the participants’ sleep
quality when using and not using a snooze alarm. We recorded
electroencephalograms (EEGs) at four scalp sites (C3, C4, O1, O2) according to
the international 10% system [ [ 34
](/articles/10.1186/s40101-022-00317-w#ref-CR34 "Sharbrough F. American
Electroencephalographic Society guidelines for standard electrode position
nomenclature. J Clin Neurophysiol. 1991;8\(2\):200–2.") ] with a time constant
of 0.3 s. We recorded horizontal electrooculograms (EOG) from the outer canthi
of both eyes with a time constant of 2 s. The electromyogram (EMG) was
recorded from the mentalis muscles with a time constant of 0.003 seconds. We
conducted the entire recording was conducted using Ag/AgCl electrodes. We used
high-cut filters of 60 Hz for EEG and EOG and 120 Hz for EMG at a sampling
rate of 500 Hz. Electrode impedance was below 10 KΩ. We scored sleep stages in
20-s epochs and calculated sleep variables using EEG, EOG, and EMG
polysomnogram data [ 35 , 36 , [ 37
](/articles/10.1186/s40101-022-00317-w#ref-CR37 "Rechtschaffen A, Kales A. A
manual of standardized terminology, techniques and scoring system for sleep
stages of human subjects. Los Angeles: University of California, Brain
Information Service/BrainResearch Institute; 1968.") ]. We classified
nocturnal sleep into four-sleep stages: stage N1 sleep (shallow non-rapid eye
movement (NREM) sleep, defined as the drowsy state), stage N2 sleep (shallow
NREM sleep), stage N3 sleep (deep NREM sleep), and stage REM sleep (rapid eye
movement (REM) sleep).
### Procedure
Participants slept in their homes during the control night’s week. We
monitored their sleep-wake schedules at home using sleep logs and wrist
actigraphy (Actiwatch AW64, Mini-Mitter Co., Bed, Ore., USA) to confirm that
their sleep-wake habits were constant ( _M_ total sleep time = 373, SE = 21.5
min; _M_ bedtime = 1:29:34, SE = 0:19:22; _M_ awakening time = 7:42:13, SE =
0:13:52). We used the identical sleep-wake schedules as the control nights on
the experimental nights in the sleep laboratory. We also asked the
participants to wake up daily during the control nights by using the snooze
function of an identical mobile phone (Softbank Corp., 831P) to the one to be
used later during experimental nights in the sleep laboratory to familiarize
themselves with the experimental procedure. Based on the results of study 1,
we set the snooze alarm to activate four times at 5-min intervals during the
20-min before a predetermined waking time. The alarm was either stopped by the
participants or continued for 60 s.
After the week of control nights, the participants spent three consecutive
experimental nights in a sleep laboratory. On the experimental days, we
requested the participants to abstain from ingesting substances that affected
sleep and wakefulness, including alcohol, caffeine, and nicotine. The first
experimental night was an adaptation session, and the second and third nights
were data collection nights under the snooze alarm or no-snooze alarm sleep
conditions, which we counterbalanced across the participants. We fixed the
awakening and going to sleep times at the participant’s usual waking and
bedtimes by referring to the control nights ( _M_ total sleep time = 402.0, SE
= 8.0 min _; M_ bedtime = 1:10:00, SE _=_ 0:15:55; _M_ awakening time =
7:52:00 am, SE = 0:13:34).
The participants finished dinner in their homes and arrived at the sleep
laboratory approximately 2 h before bedtime. Then, we attached the
polysomnogram electrodes. In the laboratory, the participants could drink
water freely, and we asked them to go to the toilet before bedtime. Then, the
participants entered a soundproof, air-conditioned isolation unit 20 min
before their predetermined sleep time. We eliminated all time cues, including
natural light, from this unit. Then, the participants completed the GV and GA
Questionnaires [ [ 29 ](/articles/10.1186/s40101-022-00317-w#ref-CR29 "Monk
TH. A visual analogue scale technique to measure global vigor and affect.
Psychiatry Res. 1989;27\(1\):89–99.
https://doi.org/10.1016/0165-1781\(89\)90013-9
.") ] and performed the simple auditory reaction time task for 5 min, 15 min before sleep. Following this, the participants responded to the STAI [ [ 31 ](/articles/10.1186/s40101-022-00317-w#ref-CR31 "Shimizu H, Imae K. Development of the Japanese edition of the Spielberger state–trait anxiety inventory \(STAI\) for student use. Japanese J Educ Psychol. 1981;29:348–53 Japanese.") ]. We instructed them to go to sleep at the usual time. Prior to the awakening time in the snooze alarm condition, a mobile phone’s snooze alarm (50 dB) kept beside the pillow was activated for 1 min, and this alarm recurred at 5-min intervals for 20 min. Participants could stop the alarm before 1 min had elapsed by pressing a button on the mobile phone. We asked the participants to sleep again until the alarm finally woke them up. In the no-snooze condition, the snooze alarm did not activate before waking up. In both conditions, participants were awakened at the predetermined awakening time by an auditory stimulus (a computer-generated 60 dB of beep tone). After waking up, the participants completed the OSA-MA sleep inventory questionnaire [ [ 33 ](/articles/10.1186/s40101-022-00317-w#ref-CR33 "Yamamoto Y, Tanaka H, Takase M, Yamazaki K, Azumi K, Shirakawa S. Chuukounen/koureisya wo taisyoutoshita OSA suiminchousahyou \(MA ban\) no kaihatu to hyoujyunka. \[\(standardization of revised version of OSA sleep inventory for middle age and aged.\) \(in Japanese with English abstract.\)\]. Brain Sci Mental Disord. 1999;10:401–9 Japanese.") ]. Then, they conducted the post-sleep sessions during 30 min after awakening: (1) the 5-min auditory reaction time task (session 1, session 2, session 3); and (2) responding to the GV and GA questionnaire using the visual analog scale for every 10 min.
### Statistical analysis
We used SPSS ver. 28 (IBM) for statistical analyses. We used _t_ -tests to
compare anxiety levels (STAI scores) at bedtime, sleep variables (time in bed,
the total sleep time, sleep efficiency, waking time after sleep onset, sleep
stage times (stages N1, N2, N3, and REM), movement time), and OSA-MA scores on
snooze alarm and no snooze alarm nights. We conducted a two-factor, repeated
measures analysis of variance (ANOVA) with conditions (snooze alarm and no-
snooze alarm) crossed with the experimental sessions to examine task
performance. We used the pre-session performance measurement before sleeping
and the performance measurements of the three post-session after awakening,
including auditory reaction times, correct response rates, and subjective GV
and GA ratings. The ANOVA effect sizes were shown with partial η 2 (ηp 2
). We adjusted the degrees of freedom using Greenhouse-Geisser’s epsilon if
the assumption of sphericity (Mauchly’s sphericity test) was significant. The
probability of significance was adjusted using Bonferroni corrections if the
main effect was significant. We set the statistical significance level at 0.05
for all the analyses.
## Results: study 2
### Sleep variables
The polysomnogram data indicated that the total sleeping time of the whole
night was significantly shorter in the snooze alarm condition ( _M =_ 387.8,
SE = 7.8 min) than in the no-snooze alarm condition ( _M_ = 396.5, SE = 7.8
min; _t_ (9) = 2.780, _p_ = 0.021). Table [ 3
](/articles/10.1186/s40101-022-00317-w#Tab3) shows the sleep data, excluding
the last 20 min before awakening (upper part) and the sleep data of the last
20 min (lower part) on snooze and no-snooze alarm nights. During the sleeping
time excluding the last 20 min, participants slept for an average of 371.9 min
(SE _=_ 7.8) and 376.8 min (SE _=_ 7.9), respectively, on snooze alarm and no-
snooze alarm nights. There were no significant differences in sleep variables
between these two conditions. The total sleep time during the last 20 min was
4 min shorter with the snooze alarm ( _M_ = 15.7, SE = 0.9 min) than without
it ( _M_ = 19.7, SE = 0.2 min; _p_ = 0.002). Moreover, sleep efficiency was
significantly worse ( _p_ = 0.002), and wake time ( _p_ = 0.009), and stage N1
sleep ( _p_ = 0.006) were longer on snooze alarm nights than on no-snooze
alarm nights.
**Table 3 Polysomnogram data for total nights’ sleep (excluding the last 20
min before awakening) and the last 20 min of sleep**
[ Full size table ](/articles/10.1186/s40101-022-00317-w/tables/3)
Six participants woke up in stage N2 sleep, and others had already woken up
before the predetermined waking time in the snooze alarm condition. In
contrast, all the participants except one remained asleep until waking time in
the no-snooze alarm condition. Moreover, participants were aroused in
different sleep stages in the no-snooze alarm condition, such that two
participants were aroused in stage N1 sleep, five were aroused in state N2
sleep, and two were aroused in stage REM sleep, while no participants were
aroused during stage N3 sleep.
The participants were more frequently aroused during the last 20 min of sleep
in the snooze alarm condition ( _M_ = 4.1, SE = 0.77 times) than in the no-
snooze alarm condition ( _M_ = 0.3, SE = 0.21 times; _t_ (9) = 6.042, _p < _
0.001). Also, the number of sleep-stage transitions increased in the snooze
alarm condition ( _M_ = 12.2, SE = 2.0 times) relative to the no-snooze alarm
condition ( _M =_ 3.5, SE = 0.92 times; _t_ (9) = 3.538, _p_ = 0.006). The
sleep stages during the 20 min before awakening indicated that stage N2 and
REM sleep were stable in the no-snooze alarm condition, whereas waking and
stage N1 sleep increased periodically in the snooze alarm condition.
### Objective performance
Figure [ 1 ](/articles/10.1186/s40101-022-00317-w#Fig1) a, b shows auditory
reaction times (RTs) and correct response rates in the snooze and no-snooze
alarm conditions. ANOVAs with the two conditions (snooze and no-snooze)
crossed with the sessions (4 levels: pre-sleep, session 1, session 2, session
3) indicated neither the main effect of the condition ( _F_ (1, 9) = 3.39, _p_
= 0.15, _η_ _p_ _2_ = 0.22) nor session ( _F_ (3, 27) = 0.87, _p_ = 0.47, _η_
_p_ _2_ = 0.09; _W_ (Mauchly’s sphericity test) = 0.327, _p_ = 0.13) on
auditory reaction times, whereas the interaction between condition and session
was marginally significant ( _F_ (3, 27) = 2.57, _p_ = 0.08, _η_ _p_ _2_ =
0.22; _W_ = 0.611, _p_ = 0.58). RTs were significantly longer in session 3 (
_M_ = 238.4, SE = 9.6 ms) in the snooze than in the no-snooze condition ( _M_
= 221.9, SE = 7.9 ms; _p_ = 0.024). Moreover, there was no significant main
effect of the correct response rates on condition ( _F_ (1, 9) = 0.13, _p_ =
0.73, _η_ _p_ _2_ = 0.01) or session ( _F_ (1.98, 17.82) = 0.27, _p_ = 0.76,
_η_ _p_ _2_ = 0.03; _W_ = 0.029, _p_ = 0.00). Furthermore, the interaction
between the condition and the session was not significant ( _F_ (1.55, 14.00)
= 2.02, _p_ = 0.18, _η_ _p_ _2_ = 0.18; _W_ = 0.115, _p_ = 0.01).
**Fig. 1**
[ 
](/articles/10.1186/s40101-022-00317-w/figures/1)
**a** Mean auditory reaction times, **b** mean correct response rates in a
simple auditory reaction time task, **c** mean global vigor (GV) VAS values,
**d** mean global affect (GA) VAS values before and after nocturnal sleep. *
_p_ < 0.05, ** _p_ < 0.01; snooze alarm condition vs no-snooze alarm
condition. The vertical bars reflect standard errors (SE)
[ Full size image ](/articles/10.1186/s40101-022-00317-w/figures/1)
### Subjective reports
We analyzed STAI scores to examine the participants’ pre-sleep anxiety levels
using paired _t tests_ between snooze alarm ( _M_ = 34.6, SE = 1.6) and no-
snooze alarm ( _M_ = 35.9, SE = 2.2) conditions ( _t_ (9) = 1.314, _p_ =
0.221). The results revealed no significant differences in pre-sleep anxiety.
Also, a series of paired _t_ -tests examined post-sleep differences between
snooze alarm and no-snooze alarm conditions on OSA-MA scores assessing
sleepiness ( _M_ = 42.9, SE = 2.8 versus _M_ = 43.6, SE = 2.3; _t_ (9) =
0.368, _p_ = 0.721); sleep-onset and sleep sustainment ( _M_ = 41.6, SE = 2.8
vs. _M_ = 48.1, SE = 3.1; _t_ (9) = 2.401, _p_ = 0.040); dreaming ( _M_ =
46.9, SE = 3.3 vs. _M_ = 44.4, SE = 3.6; _t_ (9) = 0.703, _p_ = 0.500);
recovery from exhaustion ( _M_ = 42.7, SE = 2.0 vs. _M_ = 44.8, SE = 2.0; _t_
(9) = 2.079, _p_ = 0.067); and sleep duration ( _M_ = 42.8, SE = 2.3 vs. _M_ =
45.7, SE = 2.1; _t_ (9) = 2.228, _p_ = 0.053). These _t_ tests indicated
significant differences only for sleep-onset and sleep sustainment, with
participants in the no-snooze alarm condition showing better sleep quality
than those in the snooze alarm condition.
Figure [ 1 ](/articles/10.1186/s40101-022-00317-w#Fig1) c, d shows mean GV and
GA values between snooze and no-snooze alarm conditions. We conducted repeated
measures ANOVAs with two conditions (snooze and no-snooze alarm) crossed with
the sessions (5 levels: pre-sleep and 2, 12, 22, 32 min after from awakening)
to examine differences in GV and GA values between the conditions. The results
indicated that the main effect of condition ( _F_ (1, 9) = 4.05, _p_ = 0.08,
_η_ _p_ _2_ = 0.31) and session ( _F_ (2.23, 20.08) = 1.80, _p_ = 0.19, _η_
_p_ _2_ = 0.17; _W_ = 0.047, _p_ = 0.01) were not significant for GV. However,
the condition by session interaction was significant ( _F_ (4, 36) = 4.35, _p_
= 0.006, _η_ _p_ _2_ = 0.33; _W_ = 0.170, _p_ = 0.17), showing that GV was
significantly enhanced after awakening (2 min: _M_ = 49.9, SE = 3.0 ( _p_ =
0.002), 12 min: _M_ = 51.7, SE = 2.1 ( _p_ < 0.001), 22 min: _M_ = 51.1, SE =
2.3 ( _p_ < 0.001), 32 min: _M_ = 52.6, SE = 3.5 ( _p_ < 0.001)) compared to
pre-sleep ( _M_ = 40.8, SE = 2.2) in the no-snooze condition, which was not
the case in the snooze condition. There was no significant difference among
post four sessions. In contrast, neither the main effect of condition ( _F_
(1, 9) = 0.193, _p_ = 0.67, _η_ _p_ _2_ = 0.19) nor session ( _F_ (2.17,
21.70) = 4.17, _p_ = 0.27, _η_ _p_ _2_ = 0.29; _W_ = 0.057, _p_ = 0.01), nor
the interaction between condition and session was significant for GA ( _F_
(2.00, 19.79) = 0.51, _p_ = 0.61, _η_ _p_ _2_ = 0.05; _W_ = 0.088, _p_ =
0.02).
### Final sleep stage and objective/subjective data
Table [ 4 ](/articles/10.1186/s40101-022-00317-w#Tab4) shows the number of
participants that woke up from stages N1, N2, and REM sleep and those that
were awake, and the number of sleep-stage transitions during the last 20 min
of sleep. The number of participants who woke up from stage N2 sleep was
relatively high in both snooze and no-snooze conditions. Moreover, the sleep-
stage transitions were small for all sleep stages in the no-snooze than the
snooze condition.
**Table 4 Final sleep stage and objective/subjective data**
[ Full size table ](/articles/10.1186/s40101-022-00317-w/tables/4)
Table [ 4 ](/articles/10.1186/s40101-022-00317-w#Tab4) also shows objective
(auditory reaction time) and subjective (Global Vigor score) data after
awakening on the sleep stages just before waking up. The auditory reaction
times significantly differed between the conditions in session 3. Moreover,
the GV score showed a significant difference between pre-sleep and all post-
sleep sessions in the no-snooze condition. Therefore, we calculated the
auditory reaction times by subtracting the pre-sleep session reaction times
from session 3 and GV by subtracting the pre-sleep session scores from the
mean scores in all post-sleep sessions. The results indicated that subjective
reports (Global Vigor) increased after awakening from stage N1 sleep and being
awake in the no-snooze condition. Moreover, the reaction times were better for
all sleep stages in the no-snooze condition than the snooze condition.
## General discussion
This study investigated the efficacy of alarm clocks’ snooze function on sleep
inertia. The results of study 1 indicated that 85.7% of university students in
the survey used an external tool to wake up in the morning. Moreover, 70.5% of
these students often used the snooze function of their mobile phones, mainly
to reduce their anxiety about oversleeping. The most common snooze alarm
setting reported by the students allowed 4 or 5 resets in the last 20 min of
nocturnal sleep, separated by 5-min intervals. Study 2 indicated that sleep
variables were not significantly different between the conditions other than
in the last 20 min before the alarm sounded. However, the wake time and stage
N1 sleep were prolonged after the first snooze alarm in the last 20 min of
nocturnal sleep. In addition, auditory reaction times to the simple auditory
task were slower in session 3, and global vigor deteriorated after awakening
in the snooze alarm compared to the no-snooze alarm condition. These results
suggest that the mobile phones’ snooze function might increase sleep inertia
after waking up. This sleep inertia might be explained by the sleep stage
before awakening and repeated forced awakening during the last 20 min of
sleep.
Sleep length prior to waking [ [ 7 ](/articles/10.1186/s40101-022-00317-w#ref-
CR7 "Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG
spectral power and cognitive performance during sleep inertia: the effect of
normal sleep duration and partial sleep deprivation. Physiol Behav.
2006;87\(1\):177–84.
https://doi.org/10.1016/j.physbeh.2005.09.017
.") ], the circadian timing of awakening [ [ 14 ](/articles/10.1186/s40101-022-00317-w#ref-CR14 "McHill AW, Hull JT, Cohen DA, Wang W, Czeisler CA, Klerman EB. Chronic sleep restriction greatly magnifies performance decrements immediately after awakening. Sleep. 2019;42\(5\):zsz032.
https://doi.org/10.1093/sleep/zsz032
.") , [ 15 ](/articles/10.1186/s40101-022-00317-w#ref-CR15 "Scheer FA, Shea TJ, Hilton MF, Shea SA. An endogenous circadian rhythm in sleep inertia results in greatest cognitive impairment upon awakening during the biological night. J Biol Rhythm. 2008;23\(4\):353–61.
https://doi.org/10.1177/0748730408318081
.") ], the time awake prior to sleep onset [ [ 12 ](/articles/10.1186/s40101-022-00317-w#ref-CR12 "Dinges DF, Orne MT, Whitehouse WG, Orne EC. Temporal placement of a nap for alertness: contributions of circadian phase and prior wakefulness. Sleep. 1987;10\(4\):313–29.") ], the sleep stage at awakening [ [ 7 ](/articles/10.1186/s40101-022-00317-w#ref-CR7 "Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG spectral power and cognitive performance during sleep inertia: the effect of normal sleep duration and partial sleep deprivation. Physiol Behav. 2006;87\(1\):177–84.
https://doi.org/10.1016/j.physbeh.2005.09.017
.") ], and the waking method [ [ 11 ](/articles/10.1186/s40101-022-00317-w#ref-CR11 "Ikeda H, Hayashi M. The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol. 2010;83\(1\):15–9.
https://doi.org/10.1016/j.biopsycho.2009.09.008
.") , [ 16 ](/articles/10.1186/s40101-022-00317-w#ref-CR16 "Kaida K, Nakano E, Nittono H, Hayashi M, Hori T. The effects of self-awakening on heart rate activity in a short afternoon nap. Clin Neurophysiol. 2003;114\(10\):1896–901.
https://doi.org/10.1016/s1388-2457\(03\)00167-6
.") , [ 17 ](/articles/10.1186/s40101-022-00317-w#ref-CR17 "Matsuura N, Hayashi M. Effects of habitual self-awakening on nocturnal sleep, autonomic activity prior to awakening, and subjective condition after awakening. Sleep Biol Rhythms. 2009;7\(3\):172–80.") ] affect sleep inertia. Study 2 controlled for the time awake before sleep onset and the circadian timing of awakening under the two conditions. However, the sleep stage before awakening differed depending on the condition. Previous studies have reported that sleep inertia strongly increases after awakening from deep NREM sleep (stage N3) [ [ 38 ](/articles/10.1186/s40101-022-00317-w#ref-CR38 "Stampi C, Mullington J, Rivers M, Campos JP, Broughton R. Ultrashort sleep schedules: sleep architecture and recuperative value of 80, 50 and 20 min naps. In: Horne JA, editor. Sleep '90. Bochum: Pontenagel Press; 1990. p. 71–4.") ]. However, in this study, stage N3 sleep did not occur in the last 20 min of sleep (Table [ 3 ](/articles/10.1186/s40101-022-00317-w#Tab3) ). We observed that more participants woke up from stage N2 sleep (Table [ 4 ](/articles/10.1186/s40101-022-00317-w#Tab4) ) under both conditions. Six participants woke up from stage N2 sleep; four had already woken up before the final beeping tone in the snooze alarm condition, whereas only one had already woken up in the no-snooze alarm condition. Two participants in the no-snooze alarm condition woke up from stage N1 sleep, five from Stage N2 sleep, and two from Stage REM sleep. In the no-snooze condition, subjective reports (Global Vigor) increased in participants awakening from stage N1 sleep and participants that were awake. Moreover, the reaction times were better for all sleep stages in the no-snooze compared to the snooze condition. The effects of the sleep stages (N1, N2, and REM) before awakening on sleep inertia are especially controversial. Cavallero and Versace [ [ 13 ](/articles/10.1186/s40101-022-00317-w#ref-CR13 "Cavallero C, Versace F. Stage at awakening, sleep inertia and performance. Sleep Res Online. 2003;3:89–97.") ] reported that sleep inertia slowed the reaction time to a simple auditory task after waking up from stage N2 sleep compared to stage REM sleep, suggesting that sleep inertia is more pronounced after awakening from stage N2 sleep. However, Stones [ [ 39 ](/articles/10.1186/s40101-022-00317-w#ref-CR39 "Stones MJ. Memory performance after arousal from different sleep stages. Br J Psychol. 1977;68\(2\):177–81.
https://doi.org/10.1111/j.2044-8295.1977.tb01573.x
.") ] failed to find any effects of sleep stages before awakening on sleep inertia. Koulack and Schultz [ [ 40 ](/articles/10.1186/s40101-022-00317-w#ref-CR40 "Koulack D, Schultz KJ. Task performance after awakenings from different stages of sleep. Percept Mot Skills. 1974;39\(2\):792–4.
https://doi.org/10.2466/pms.1974.39.2.792
.") ] showed that REM sleep increased inertia. Finally, Jewett et al. [ [ 41 ](/articles/10.1186/s40101-022-00317-w#ref-CR41 "Jewett ME, Wyatt JK, Ritz-De Cecco A, Khalsa SB, Dijk DJ, Czeisler CA. Time course of sleep inertia dissipation in human performance and alertness. J Sleep Res. 1999;8\(1\):1–8.
https://doi.org/10.1111/j.1365-2869.1999.00128.x
.") ] did not always observe the effects of sleep stages on sleep inertia. Therefore, it is difficult to make any firm conclusions about this study’s results on sleep stages.
It is also possible that the snooze alarm induces forced awakening whenever it
is activated. In this study, the snooze alarm was activated four times during
the last 20 min of nocturnal sleep at 5-min intervals. Participants tried to
go back to sleep after each forced awakening, suggesting that the snooze alarm
increased their sleep propensity and waking thresholds compared to the no-
snooze alarm condition. Indeed, sleep-stage transitions increased in the
snooze alarm condition ( _M_ = 12.2) than in the no-snooze alarm condition (
_M_ = 3.5). Transitions into and from sleep induce highly predictable
autonomic changes [ [ 42 ](/articles/10.1186/s40101-022-00317-w#ref-CR42
"Ekstedt M, Akerstedt T, Söderström M. Microarousals during sleep are
associated with increased levels of lipids, cortisol, and blood pressure.
Psychosom Med. 2004;66\(6\):925–31.
https://doi.org/10.1097/01.psy.0000145821.25453.f7
.") , [ 43 ](/articles/10.1186/s40101-022-00317-w#ref-CR43 "Trinder J, Padula M, Berlowitz D, et al. Cardiac and respiratory activity at arousal from sleep under controlled ventilation conditions. J Appl Physiol \(1985\). 2001;90\(4\):1455–63.
https://doi.org/10.1152/jappl.2001.90.4.1455
.") ] that might discourage smooth awakening. Moreover, sleep fragmentation is related to daytime sleepiness [ [ 44 ](/articles/10.1186/s40101-022-00317-w#ref-CR44 "Stepanski E, Lamphere J, Badia P, Zorick F, Roth T. Sleep fragmentation and daytime sleepiness. Sleep. 1984;7\(1\):18–26.
https://doi.org/10.1093/sleep/7.1.18
.") ] and detrimental to cogitative and vigilance tasks [ [ 45 ](/articles/10.1186/s40101-022-00317-w#ref-CR45 "Roehrs T, Merlotti L, Petrucelli N, Stepanski E, Roth T. Experimental sleep fragmentation. Sleep. 1994;17\(5\):438–43.
https://doi.org/10.1093/sleep/17.5.438
.") ]. Therefore, sleep inertia might have increased through repeated forced awakening. According to the two-process model of sleep regulation [ [ 46 ](/articles/10.1186/s40101-022-00317-w#ref-CR46 "Borbély AA. A two process model of sleep regulation. Hum Neurobiol. 1982;1\(3\):195–204.") ] consisting of sleep homeostatic (S) and circadian rhythm (C) processes, sleepiness should be minimal when we wake up in the morning. However, there is increased sleepiness (lowered alertness and impaired performance) immediately upon awakening in the morning. Folkard and Akerstedt [ [ 47 ](/articles/10.1186/s40101-022-00317-w#ref-CR47 "Folkard S, Åkerstedt TA. Three-process model of the regulation of alertness-sleepiness. In: Broughton RJ, Ogilvie RD, editors. Sleep, arousal, and performance. Boston: Birkhäuser; 1992. p. 11–26.") ] hypothesized that sleep inertia is a third process of sleep regulation that the S or C processes of the two-process model cannot explain. Moreover, Hilditch and McHill [ [ 20 ](/articles/10.1186/s40101-022-00317-w#ref-CR20 "Hilditch CJ, McHill AW. Sleep inertia: current insights. Nat Sci Sleep. 2019;11:155–65 Published 2019 Aug 22. 10.2147/NSS.S188911.") ] suggested that sleep inertia is an adaptive function of sleep maintenance, particularly in the morning during the last part of nocturnal sleep. Therefore, in the snooze condition, the sleep maintenance function might be enhanced by repeated forced awakening by the snooze alarm due to sleep fragmentation.
This study’s results indicated that sleep inertia only induced a significant
performance difference in session 3. Sleep inertia dissipates asymptotically
within 15 to 30 min after waking [ [ 8
](/articles/10.1186/s40101-022-00317-w#ref-CR8 "Tassi P, Muzet A. Sleep
inertia. Sleep Med Rev. 2000;4\(4\):341–53.
https://doi.org/10.1053/smrv.2000.0098
.") , [ 20 ](/articles/10.1186/s40101-022-00317-w#ref-CR20 "Hilditch CJ, McHill AW. Sleep inertia: current insights. Nat Sci Sleep. 2019;11:155–65 Published 2019 Aug 22. 10.2147/NSS.S188911.") ]. However, we found that sleep inertia in the snooze alarm condition did not follow this dissipation curve as assessed by objective measures because behavioral performance decreased, as indicated by increased reaction times throughout the sessions. Therefore, our findings also support the hypothesis that sleep inertia is a third process that we cannot explain by the two-process sleep regulation model [ [ 47 ](/articles/10.1186/s40101-022-00317-w#ref-CR47 "Folkard S, Åkerstedt TA. Three-process model of the regulation of alertness-sleepiness. In: Broughton RJ, Ogilvie RD, editors. Sleep, arousal, and performance. Boston: Birkhäuser; 1992. p. 11–26.") ]. We might observe sleep inertia in the morning even after we have adequate sleep because sleep inertia maintains sleep in the morning, during the last part of nocturnal sleep [ [ 20 ](/articles/10.1186/s40101-022-00317-w#ref-CR20 "Hilditch CJ, McHill AW. Sleep inertia: current insights. Nat Sci Sleep. 2019;11:155–65 Published 2019 Aug 22. 10.2147/NSS.S188911.") , [ 48 ](/articles/10.1186/s40101-022-00317-w#ref-CR48 "Dijk D-J, Czeisler CA. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci Lett. 1994;166\(1\):63–8.
https://doi.org/10.1016/0304-3940\(94\)90841-9
.") ]. Therefore, we might always experience sleep inertia in the morning regardless of using the snooze function. For that reason, it is also possible that the snooze and no-snooze alarm conditions develop sleep inertia through similar mechanisms in the first half of sleep sessions. Moreover, sleep fragmentation in the snooze alarm condition might induce decreased sleep quality and quantity (increased waking and stage N1 sleep) during the last 20 min of sleep and continue to decrease VG (alertness, sleepiness, motivation, loss of effort, and weariness) immediately upon awakening. We suggest that prolonged sleep inertia, sleepiness, and fatigue might accumulate consistently in the snooze alarm condition, and sleep inertia might not dissipate asymptotically. As a result, we only observed a significant difference in reaction times in the last session (session 3). We suggest that future studies investigate the effect of sleep fragmentation (repeated undesired awakening) on sleep inertia (sleepiness/fatigue and behavioral performance) as a third sleep regulation process [ [ 47 ](/articles/10.1186/s40101-022-00317-w#ref-CR47 "Folkard S, Åkerstedt TA. Three-process model of the regulation of alertness-sleepiness. In: Broughton RJ, Ogilvie RD, editors. Sleep, arousal, and performance. Boston: Birkhäuser; 1992. p. 11–26.") ].
## Limitations and directions for future research
Several limitations constrain the findings of this study. Firstly, the small
sample size might have limited the statistical power of the analyses.
Therefore, future studies should use a larger sample. Secondly, this study
found that university students use tools such as the snooze function because
of their anxiety about waking up in the morning. We suggest that future
studies investigate the effects of the snooze function on other age groups and
professions, including full-time workers. Thirdly, Mattingly et al. [ [ 25
](/articles/10.1186/s40101-022-00317-w#ref-CR25 "Mattingly SM, Martinez G,
Young J, Cain MK, Striegel A. Snoozing: an examination of a common method of
waking. Sleep. 2022;45\(10\):zsac184.
https://doi.org/10.1093/sleep/zsac184
.") ] demonstrated that the waking method depends on the sleep duration; people with a longer sleep duration wake up naturally (self-awakening), and those with a shorter sleep duration wake up using an alarm or a snooze alarm. This study did not investigate individual waking methods based on sleep duration. Therefore, we suggest that future studies investigate this relationship. Finally, the participants knew they would not oversleep because they would be forced to wake up at the predetermined time. Therefore, the anxiety (STAI) scores before sleep were not different between this study’s experimental and control conditions. We suggest that future studies use a scenario in which the participants have to worry whether or not they can wake up in the morning. Moreover, further research is needed to examine sleep outside the laboratory, including the participants’ homes, which can provide helpful information for our daily lives.
## Conclusion
We examined the effect of using a snooze alarm to wake up in the morning on
sleep inertia. Results demonstrated that 70.5% of participants often used
their mobile phones’ snooze function primarily to reduce anxiety about
oversleeping. However, the repeated use of the snooze alarm increases sleep
inertia and fatigue after awakening. People with high anxiety and worry take
longer to fall asleep [ [ 49 ](/articles/10.1186/s40101-022-00317-w#ref-CR49
"Fuller KH, Waters WF, Binks PG, Anderson T. Generalized anxiety and sleep
architecture: a polysomnographic investigation. Sleep. 1997;20:370–6.") ]. In
addition, sleep length and sleep latency are essential for the subjective
feelings of good sleep [ [ 50 ](/articles/10.1186/s40101-022-00317-w#ref-CR50
"Åkerstedt T, Hume K, Minors D, Waterhouse J. The meaning of good sleep: a
longitudinal study of polysomnography and subjective sleep quality. J Sleep
Res. 1994;3:152–8.") ]. Therefore, even though snooze alarms increase sleep
inertia, they might be crucial for reducing anxiety about oversleeping and
maintaining a good night’s sleep, mainly by avoiding prolonging sleep latency.
Van De Werken et al. [ [ 51 ](/articles/10.1186/s40101-022-00317-w#ref-CR51
"Van De Werken M, Giménez MC, De Vries B, Beersma DG, Van Someren EJ, Gordijn
MC. Effects of artificial dawn on sleep inertia, skin temperature, and the
awakening cortisol response. J Sleep Res. 2010;19\(3\):425–35.
https://doi.org/10.1111/j.1365-2869.2010.00828.x
.") ] showed the positive effects of artificial dawn during the last 30 min of nocturnal sleep on reducing sleep inertia. We suggest that further studies investigate appropriate strategies for using the snooze function to reduce sleep inertia and fragmentation with repeated forced awakening, including the alarm’s modality, force, volume, and interval. We must also consider countermeasures [ [ 20 ](/articles/10.1186/s40101-022-00317-w#ref-CR20 "Hilditch CJ, McHill AW. Sleep inertia: current insights. Nat Sci Sleep. 2019;11:155–65 Published 2019 Aug 22. 10.2147/NSS.S188911.") ] for sleep inertia after awakening, including caffeine, light, sounds, music, and temperature.
## Availability of data and materials
The datasets used and/or analyzed during the current study are available from
the corresponding author on reasonable request.
## Abbreviations
ANOVA:
Analysis of variance
EEG:
Electroencephalogram
EMG:
Electromyogram
EOG:
Electrooculograms
GV:
Global vigor
GA:
Global affect
NHK:
The Japan Broadcasting Corporation
NREM:
Non-rapid eye movement
OSA-MA:
Oguri-Shirakawa-Azumi Sleep Inventory for Middle-Aged Respondents
SA:
Self-awakening
SE:
Standard errors
SD:
Semantic differential
Stage N1:
Shallow non-rapid eye movement (NREM) sleep, especially defined as drowsy
state
Stage N2:
Shallow NREM sleep
Stage N3:
Deep NREM sleep
Stage REM:
Rapid eye movement (REM) sleep
STAI:
State anxiety scale
REM:
Rapid eye movement
RT:
Reaction time
## References
1. NHK Broadcasting Culture Research Institute. National Time Use Survey 2020. [ https://www.nhk.or.jp/bunken/yoron-jikan/column/sleep-2020.html ](https://www.nhk.or.jp/bunken/yoron-jikan/column/sleep-2020.html) . Accessed 8 May 2022.
2. NHK Broadcasting Culture Research Institute. National Time use Survey 2015. Tokyo: Japan Broadcast Publishing Co. Ltd; 2016.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=National%20Time%20use%20Survey%202015&publication_year=2016)
3. Carskadon MA, Dement WC. Sleep-related breathing disorders in elderly adults. J Gerontol. 1982;37(5):637.
[ CAS ](/articles/cas-redirect/1:STN:280:DyaL383ktVClsg%3D%3D) [ Google
Scholar ](http://scholar.google.com/scholar_lookup?&title=Sleep-
related%20breathing%20disorders%20in%20elderly%20adults&journal=J%20Gerontol&volume=37&issue=5&publication_year=1982&author=Carskadon%2CMA&author=Dement%2CWC)
4. Havekes R, Vecsey CG, Abel T. The impact of sleep deprivation on neuronal and glial signaling pathways important for memory and synaptic plasticity. Cell Signal. 2012;24(6):1251–60. [ https://doi.org/10.1016/j.cellsig.2012.02.010 ](https://doi.org/10.1016/j.cellsig.2012.02.010) .
[ Article ](https://doi.org/10.1016%2Fj.cellsig.2012.02.010) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BC38XkvF2iu7k%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20impact%20of%20sleep%20deprivation%20on%20neuronal%20and%20glial%20signaling%20pathways%20important%20for%20memory%20and%20synaptic%20plasticity&journal=Cell%20Signal&doi=10.1016%2Fj.cellsig.2012.02.010&volume=24&issue=6&pages=1251-1260&publication_year=2012&author=Havekes%2CR&author=Vecsey%2CCG&author=Abel%2CT)
5. McCoy JG, Strecker RE. The cognitive cost of sleep lost. Neurobiol Learn Mem. 2011;96(4):564–82. [ https://doi.org/10.1016/j.nlm.2011.07.004 ](https://doi.org/10.1016/j.nlm.2011.07.004) .
[ Article ](https://doi.org/10.1016%2Fj.nlm.2011.07.004) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20cognitive%20cost%20of%20sleep%20lost&journal=Neurobiol%20Learn%20Mem&doi=10.1016%2Fj.nlm.2011.07.004&volume=96&issue=4&pages=564-582&publication_year=2011&author=McCoy%2CJG&author=Strecker%2CRE)
6. Faraut B, Touchette E, Gamble H, et al. Short sleep duration and increased risk of hypertension: a primary care medicine investigation. J Hypertens. 2012;30(7):1354–63. [ https://doi.org/10.1097/HJH.0b013e32835465e5 ](https://doi.org/10.1097/HJH.0b013e32835465e5) .
[ Article ](https://doi.org/10.1097%2FHJH.0b013e32835465e5) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BC38Xos1CgtL0%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Short%20sleep%20duration%20and%20increased%20risk%20of%20hypertension%3A%20a%20primary%20care%20medicine%20investigation&journal=J%20Hypertens&doi=10.1097%2FHJH.0b013e32835465e5&volume=30&issue=7&pages=1354-1363&publication_year=2012&author=Faraut%2CB&author=Touchette%2CE&author=Gamble%2CH)
7. Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG spectral power and cognitive performance during sleep inertia: the effect of normal sleep duration and partial sleep deprivation. Physiol Behav. 2006;87(1):177–84. [ https://doi.org/10.1016/j.physbeh.2005.09.017 ](https://doi.org/10.1016/j.physbeh.2005.09.017) .
[ Article ](https://doi.org/10.1016%2Fj.physbeh.2005.09.017) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BD2MXhtlagsL%2FL) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=EEG%20spectral%20power%20and%20cognitive%20performance%20during%20sleep%20inertia%3A%20the%20effect%20of%20normal%20sleep%20duration%20and%20partial%20sleep%20deprivation&journal=Physiol%20Behav&doi=10.1016%2Fj.physbeh.2005.09.017&volume=87&issue=1&pages=177-184&publication_year=2006&author=Tassi%2CP&author=Bonnefond%2CA&author=Engasser%2CO&author=Hoeft%2CA&author=Eschenlauer%2CR&author=Muzet%2CA)
8. Tassi P, Muzet A. Sleep inertia. Sleep Med Rev. 2000;4(4):341–53. [ https://doi.org/10.1053/smrv.2000.0098 ](https://doi.org/10.1053/smrv.2000.0098) .
[ Article ](https://doi.org/10.1053%2Fsmrv.2000.0098) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20inertia&journal=Sleep%20Med%20Rev&doi=10.1053%2Fsmrv.2000.0098&volume=4&issue=4&pages=341-353&publication_year=2000&author=Tassi%2CP&author=Muzet%2CA)
9. Asaoka S, Masaki H, Ogawa K, Murphy TI, Fukuda K, Yamazaki K. Performance monitoring during sleep inertia after a 1-h daytime nap. J Sleep Res. 2010;19(3):436–43. [ https://doi.org/10.1111/j.1365-2869.2009.00811.x ](https://doi.org/10.1111/j.1365-2869.2009.00811.x) .
[ Article ](https://doi.org/10.1111%2Fj.1365-2869.2009.00811.x) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Performance%20monitoring%20during%20sleep%20inertia%20after%20a%201-h%20daytime%20nap&journal=J%20Sleep%20Res&doi=10.1111%2Fj.1365-2869.2009.00811.x&volume=19&issue=3&pages=436-443&publication_year=2010&author=Asaoka%2CS&author=Masaki%2CH&author=Ogawa%2CK&author=Murphy%2CTI&author=Fukuda%2CK&author=Yamazaki%2CK)
10. Ferrara M, De Gennaro L, Ferlazzo F, Curcio G, Barattucci M, Bertini M. Auditory evoked responses upon awakening from sleep in human subjects. Neurosci Lett. 2001;310(2-3):145–8. [ https://doi.org/10.1016/s0304-3940(01)02107-3 ](https://doi.org/10.1016/s0304-3940\(01\)02107-3) .
[ Article ](https://doi.org/10.1016%2Fs0304-3940%2801%2902107-3) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BD3MXmsFelsb0%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Auditory%20evoked%20responses%20upon%20awakening%20from%20sleep%20in%20human%20subjects&journal=Neurosci%20Lett&doi=10.1016%2Fs0304-3940%2801%2902107-3&volume=310&issue=2-3&pages=145-148&publication_year=2001&author=Ferrara%2CM&author=Gennaro%2CL&author=Ferlazzo%2CF&author=Curcio%2CG&author=Barattucci%2CM&author=Bertini%2CM)
11. Ikeda H, Hayashi M. The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol. 2010;83(1):15–9. [ https://doi.org/10.1016/j.biopsycho.2009.09.008 ](https://doi.org/10.1016/j.biopsycho.2009.09.008) .
[ Article ](https://doi.org/10.1016%2Fj.biopsycho.2009.09.008) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effect%20of%20self-
awakening%20from%20nocturnal%20sleep%20on%20sleep%20inertia&journal=Biol%20Psychol&doi=10.1016%2Fj.biopsycho.2009.09.008&volume=83&issue=1&pages=15-19&publication_year=2010&author=Ikeda%2CH&author=Hayashi%2CM)
12. Dinges DF, Orne MT, Whitehouse WG, Orne EC. Temporal placement of a nap for alertness: contributions of circadian phase and prior wakefulness. Sleep. 1987;10(4):313–29.
[ CAS ](/articles/cas-redirect/1:STN:280:DyaL1c%2FhslKluw%3D%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Temporal%20placement%20of%20a%20nap%20for%20alertness%3A%20contributions%20of%20circadian%20phase%20and%20prior%20wakefulness&journal=Sleep.&volume=10&issue=4&pages=313-329&publication_year=1987&author=Dinges%2CDF&author=Orne%2CMT&author=Whitehouse%2CWG&author=Orne%2CEC)
13. Cavallero C, Versace F. Stage at awakening, sleep inertia and performance. Sleep Res Online. 2003;3:89–97.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Stage%20at%20awakening%2C%20sleep%20inertia%20and%20performance&journal=Sleep%20Res%20Online&volume=3&pages=89-97&publication_year=2003&author=Cavallero%2CC&author=Versace%2CF)
14. McHill AW, Hull JT, Cohen DA, Wang W, Czeisler CA, Klerman EB. Chronic sleep restriction greatly magnifies performance decrements immediately after awakening. Sleep. 2019;42(5):zsz032. [ https://doi.org/10.1093/sleep/zsz032 ](https://doi.org/10.1093/sleep/zsz032) .
[ Article ](https://doi.org/10.1093%2Fsleep%2Fzsz032) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Chronic%20sleep%20restriction%20greatly%20magnifies%20performance%20decrements%20immediately%20after%20awakening&journal=Sleep.&doi=10.1093%2Fsleep%2Fzsz032&volume=42&issue=5&publication_year=2019&author=McHill%2CAW&author=Hull%2CJT&author=Cohen%2CDA&author=Wang%2CW&author=Czeisler%2CCA&author=Klerman%2CEB)
15. Scheer FA, Shea TJ, Hilton MF, Shea SA. An endogenous circadian rhythm in sleep inertia results in greatest cognitive impairment upon awakening during the biological night. J Biol Rhythm. 2008;23(4):353–61. [ https://doi.org/10.1177/0748730408318081 ](https://doi.org/10.1177/0748730408318081) .
[ Article ](https://doi.org/10.1177%2F0748730408318081) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=An%20endogenous%20circadian%20rhythm%20in%20sleep%20inertia%20results%20in%20greatest%20cognitive%20impairment%20upon%20awakening%20during%20the%20biological%20night&journal=J%20Biol%20Rhythm&doi=10.1177%2F0748730408318081&volume=23&issue=4&pages=353-361&publication_year=2008&author=Scheer%2CFA&author=Shea%2CTJ&author=Hilton%2CMF&author=Shea%2CSA)
16. Kaida K, Nakano E, Nittono H, Hayashi M, Hori T. The effects of self-awakening on heart rate activity in a short afternoon nap. Clin Neurophysiol. 2003;114(10):1896–901. [ https://doi.org/10.1016/s1388-2457(03)00167-6 ](https://doi.org/10.1016/s1388-2457\(03\)00167-6) .
[ Article ](https://doi.org/10.1016%2Fs1388-2457%2803%2900167-6) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effects%20of%20self-
awakening%20on%20heart%20rate%20activity%20in%20a%20short%20afternoon%20nap&journal=Clin%20Neurophysiol&doi=10.1016%2Fs1388-2457%2803%2900167-6&volume=114&issue=10&pages=1896-1901&publication_year=2003&author=Kaida%2CK&author=Nakano%2CE&author=Nittono%2CH&author=Hayashi%2CM&author=Hori%2CT)
17. Matsuura N, Hayashi M. Effects of habitual self-awakening on nocturnal sleep, autonomic activity prior to awakening, and subjective condition after awakening. Sleep Biol Rhythms. 2009;7(3):172–80.
[ Article ](https://doi.org/10.1111%2Fj.1479-8425.2009.00398.x) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Effects%20of%20habitual%20self-
awakening%20on%20nocturnal%20sleep%2C%20autonomic%20activity%20prior%20to%20awakening%2C%20and%20subjective%20condition%20after%20awakening&journal=Sleep%20Biol%20Rhythms&doi=10.1111%2Fj.1479-8425.2009.00398.x&volume=7&issue=3&pages=172-180&publication_year=2009&author=Matsuura%2CN&author=Hayashi%2CM)
18. Bruck D, Pisani DL. The effects of sleep inertia on decision-making performance. J Sleep Res. 1999;8(2):95–103.
[ Article ](https://doi.org/10.1046%2Fj.1365-2869.1999.00150.x) [ CAS
](/articles/cas-redirect/1:STN:280:DyaK1MzhslehsA%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effects%20of%20sleep%20inertia%20on%20decision-
making%20performance&journal=J%20Sleep%20Res&doi=10.1046%2Fj.1365-2869.1999.00150.x&volume=8&issue=2&pages=95-103&publication_year=1999&author=Bruck%2CD&author=Pisani%2CDL)
19. Stampi C. The effects of polyphasic and ultrashort sleep schedules. In: Stampi C, editor. Why we nap: evolution, chronobiology and functions of Polyphasic and Ultrashort sleep. Boston: Birkhäuser; 1992. p. 137–79.
[ Chapter ](https://link.springer.com/doi/10.1007/978-1-4757-2210-9_10) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effects%20of%20polyphasic%20and%20ultrashort%20sleep%20schedules&doi=10.1007%2F978-1-4757-2210-9_10&pages=137-179&publication_year=1992&author=Stampi%2CC)
20. Hilditch CJ, McHill AW. Sleep inertia: current insights. Nat Sci Sleep. 2019;11:155–65 Published 2019 Aug 22. 10.2147/NSS.S188911.
[ Article ](https://doi.org/10.2147%2FNSS.S188911) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20inertia%3A%20current%20insights&journal=Nat%20Sci%20Sleep&doi=10.2147%2FNSS.S188911&volume=11&pages=155-165&publication_year=2019&author=Hilditch%2CCJ&author=McHill%2CAW)
21. Moorcroft WH, Kayser KH, Griggs AJ. Subjective and objective confirmation of the ability to self-awaken at a self-predetermined time without using external means. Sleep. 1997;20(1):40–5. [ https://doi.org/10.1093/sleep/20.1.40 ](https://doi.org/10.1093/sleep/20.1.40) .
[ Article ](https://doi.org/10.1093%2Fsleep%2F20.1.40) [ CAS ](/articles/cas-
redirect/1:STN:280:DyaK2s3ns1Grsg%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Subjective%20and%20objective%20confirmation%20of%20the%20ability%20to%20self-
awaken%20at%20a%20self-
predetermined%20time%20without%20using%20external%20means&journal=Sleep.&doi=10.1093%2Fsleep%2F20.1.40&volume=20&issue=1&pages=40-45&publication_year=1997&author=Moorcroft%2CWH&author=Kayser%2CKH&author=Griggs%2CAJ)
22. Born J, Hansen K, Marshall L, Mölle M, Fehm HL. Timing the end of nocturnal sleep. Nature. 1999;397(6714):29–30. [ https://doi.org/10.1038/16166 ](https://doi.org/10.1038/16166) .
[ Article ](https://doi.org/10.1038%2F16166) [ CAS ](/articles/cas-
redirect/1:CAS:528:DyaK1MXnsFyqsA%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Timing%20the%20end%20of%20nocturnal%20sleep&journal=Nature.&doi=10.1038%2F16166&volume=397&issue=6714&pages=29-30&publication_year=1999&author=Born%2CJ&author=Hansen%2CK&author=Marshall%2CL&author=M%C3%B6lle%2CM&author=Fehm%2CHL)
23. Kräuchi K, Cajochen C, Wirz-Justice A. Waking up properly: is there a role of thermoregulation in sleep inertia? J Sleep Res. 2004;13(2):121–7. [ https://doi.org/10.1111/j.1365-2869.2004.00398.x ](https://doi.org/10.1111/j.1365-2869.2004.00398.x) .
[ Article ](https://doi.org/10.1111%2Fj.1365-2869.2004.00398.x) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Waking%20up%20properly%3A%20is%20there%20a%20role%20of%20thermoregulation%20in%20sleep%20inertia%3F&journal=J%20Sleep%20Res&doi=10.1111%2Fj.1365-2869.2004.00398.x&volume=13&issue=2&pages=121-127&publication_year=2004&author=Kr%C3%A4uchi%2CK&author=Cajochen%2CC&author=Wirz-
Justice%2CA)
24. Allen RP. Article reviewed: timing the end of nocturnal sleep. Sleep Med. 2001;2(1):69–70. [ https://doi.org/10.1016/s1389-9457(00)00090-3 ](https://doi.org/10.1016/s1389-9457\(00\)00090-3) .
[ Article ](https://doi.org/10.1016%2Fs1389-9457%2800%2900090-3) [ CAS
](/articles/cas-redirect/1:STN:280:DC%2BC2sbjvF2qtA%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Article%20reviewed%3A%20timing%20the%20end%20of%20nocturnal%20sleep&journal=Sleep%20Med&doi=10.1016%2Fs1389-9457%2800%2900090-3&volume=2&issue=1&pages=69-70&publication_year=2001&author=Allen%2CRP)
25. Mattingly SM, Martinez G, Young J, Cain MK, Striegel A. Snoozing: an examination of a common method of waking. Sleep. 2022;45(10):zsac184. [ https://doi.org/10.1093/sleep/zsac184 ](https://doi.org/10.1093/sleep/zsac184) .
[ Article ](https://doi.org/10.1093%2Fsleep%2Fzsac184) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Snoozing%3A%20an%20examination%20of%20a%20common%20method%20of%20waking&journal=Sleep.&doi=10.1093%2Fsleep%2Fzsac184&volume=45&issue=10&publication_year=2022&author=Mattingly%2CSM&author=Martinez%2CG&author=Young%2CJ&author=Cain%2CMK&author=Striegel%2CA)
26. Organization for Economic Co-operation and Development (OECD). Social protection and well-being. Time use; 2020. [ https://stats.oecd.org/Index.aspx?DataSetCode=TIME_USE# ](https://stats.oecd.org/Index.aspx?DataSetCode=TIME_USE#) . Accessed 8 May 2022
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Social%20protection%20and%20well-
being.%20Time%20use&publication_year=2020)
27. Steptoe A, Peacey V, Wardle J. Sleep duration and health in young adults. Arch Intern Med. 2006;166(16):1689–92. [ https://doi.org/10.1001/archinte.166.16.1689 ](https://doi.org/10.1001/archinte.166.16.1689) .
[ Article ](https://doi.org/10.1001%2Farchinte.166.16.1689) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20duration%20and%20health%20in%20young%20adults&journal=Arch%20Intern%20Med&doi=10.1001%2Farchinte.166.16.1689&volume=166&issue=16&pages=1689-1692&publication_year=2006&author=Steptoe%2CA&author=Peacey%2CV&author=Wardle%2CJ)
28. Faul F, Erdfelder E, Buchner A, Lang AG. Statistical power analyses using G*power 3.1: tests for correlation and regression analyses. Behav Res Methods. 2009;41(4):1149–60. [ https://doi.org/10.3758/BRM.41.4.1149 ](https://doi.org/10.3758/BRM.41.4.1149) .
[ Article ](https://doi.org/10.3758%2FBRM.41.4.1149) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Statistical%20power%20analyses%20using%20G%2Apower%203.1%3A%20tests%20for%20correlation%20and%20regression%20analyses&journal=Behav%20Res%20Methods&doi=10.3758%2FBRM.41.4.1149&volume=41&issue=4&pages=1149-1160&publication_year=2009&author=Faul%2CF&author=Erdfelder%2CE&author=Buchner%2CA&author=Lang%2CAG)
29. Monk TH. A visual analogue scale technique to measure global vigor and affect. Psychiatry Res. 1989;27(1):89–99. [ https://doi.org/10.1016/0165-1781(89)90013-9 ](https://doi.org/10.1016/0165-1781\(89\)90013-9) .
[ Article ](https://doi.org/10.1016%2F0165-1781%2889%2990013-9) [ CAS
](/articles/cas-redirect/1:STN:280:DyaL1M7mtFSgtw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=A%20visual%20analogue%20scale%20technique%20to%20measure%20global%20vigor%20and%20affect&journal=Psychiatry%20Res&doi=10.1016%2F0165-1781%2889%2990013-9&volume=27&issue=1&pages=89-99&publication_year=1989&author=Monk%2CTH)
30. Monk TH. Subjective ratings of sleepiness--the underlying circadian mechanisms. Sleep. 1987;10(4):343–53. [ https://doi.org/10.1093/sleep/10.4.343 ](https://doi.org/10.1093/sleep/10.4.343) .
[ Article ](https://doi.org/10.1093%2Fsleep%2F10.4.343) [ CAS ](/articles/cas-
redirect/1:STN:280:DyaL1c%2FhslKksw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Subjective%20ratings%20of%20sleepiness
--
the%20underlying%20circadian%20mechanisms&journal=Sleep.&doi=10.1093%2Fsleep%2F10.4.343&volume=10&issue=4&pages=343-353&publication_year=1987&author=Monk%2CTH)
31. Shimizu H, Imae K. Development of the Japanese edition of the Spielberger state–trait anxiety inventory (STAI) for student use. Japanese J Educ Psychol. 1981;29:348–53 Japanese.
[ Article ](https://doi.org/10.5926%2Fjjep1953.29.4_348) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Development%20of%20the%20Japanese%20edition%20of%20the%20Spielberger%20state%E2%80%93trait%20anxiety%20inventory%20%28STAI%29%20for%20student%20use&journal=Japanese%20J%20Educ%20Psychol&doi=10.5926%2Fjjep1953.29.4_348&volume=29&pages=348-353&publication_year=1981&author=Shimizu%2CH&author=Imae%2CK)
32. Spielberger CD, Gorsuch RL, Lushene E. Manual for the state-trait anxiety inventory (self-evaluation questionnaire). Palo Alto: Consulting Psychologists Press; 1970.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Manual%20for%20the%20state-
trait%20anxiety%20inventory%20%28self-
evaluation%20questionnaire%29&publication_year=1970&author=Spielberger%2CCD&author=Gorsuch%2CRL&author=Lushene%2CE)
33. Yamamoto Y, Tanaka H, Takase M, Yamazaki K, Azumi K, Shirakawa S. Chuukounen/koureisya wo taisyoutoshita OSA suiminchousahyou (MA ban) no kaihatu to hyoujyunka. [(standardization of revised version of OSA sleep inventory for middle age and aged.) (in Japanese with English abstract.)]. Brain Sci Mental Disord. 1999;10:401–9 Japanese.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Chuukounen%2Fkoureisya%20wo%20taisyoutoshita%20OSA%20suiminchousahyou%20%28MA%20ban%29%20no%20kaihatu%20to%20hyoujyunka.%20%5B%28standardization%20of%20revised%20version%20of%20OSA%20sleep%20inventory%20for%20middle%20age%20and%20aged.%29%20%28in%20Japanese%20with%20English%20abstract.%29%5D&journal=Brain%20Sci%20Mental%20Disord&volume=10&pages=401-409&publication_year=1999&author=Yamamoto%2CY&author=Tanaka%2CH&author=Takase%2CM&author=Yamazaki%2CK&author=Azumi%2CK&author=Shirakawa%2CS)
34. Sharbrough F. American Electroencephalographic Society guidelines for standard electrode position nomenclature. J Clin Neurophysiol. 1991;8(2):200–2.
[ Article ](https://doi.org/10.1097%2F00004691-199104000-00007) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=American%20Electroencephalographic%20Society%20guidelines%20for%20standard%20electrode%20position%20nomenclature&journal=J%20Clin%20Neurophysiol.&doi=10.1097%2F00004691-199104000-00007&volume=8&issue=2&pages=200-2&publication_year=1991&author=Sharbrough%2CF)
35. Berry RB, Brooks R, Gamaldo CE, Harding SM, Lloyd RM, Marcus CL, et al. The AASM manual for the scoring of sleep and associated events: rules, terminology and technical specifications. Version 2.5. Darien: American Academy of Sleep Medicine; 2018.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20AASM%20manual%20for%20the%20scoring%20of%20sleep%20and%20associated%20events%3A%20rules%2C%20terminology%20and%20technical%20specifications.%20Version%202.5&publication_year=2018&author=Berry%2CRB&author=Brooks%2CR&author=Gamaldo%2CCE&author=Harding%2CSM&author=Lloyd%2CRM&author=Marcus%2CCL)
36. Hori T, Sugita Y, Koga E, et al. Proposed supplements and amendments to 'A manual of standardized terminology, techniques and scoring system for sleep stages of human Subjects', the Rechtschaffen & Kales (1968) standard. Psychiatry Clin Neurosci. 2001;55(3):305–10. [ https://doi.org/10.1046/j.1440-1819.2001.00810.x ](https://doi.org/10.1046/j.1440-1819.2001.00810.x) .
[ Article ](https://doi.org/10.1046%2Fj.1440-1819.2001.00810.x) [ CAS
](/articles/cas-redirect/1:STN:280:DC%2BD3MzmtlSktQ%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Proposed%20supplements%20and%20amendments%20to%20%27A%20manual%20of%20standardized%20terminology%2C%20techniques%20and%20scoring%20system%20for%20sleep%20stages%20of%20human%20Subjects%27%2C%20the%20Rechtschaffen%20%26%20Kales%20%281968%29%20standard&journal=Psychiatry%20Clin%20Neurosci&doi=10.1046%2Fj.1440-1819.2001.00810.x&volume=55&issue=3&pages=305-310&publication_year=2001&author=Hori%2CT&author=Sugita%2CY&author=Koga%2CE)
37. Rechtschaffen A, Kales A. A manual of standardized terminology, techniques and scoring system for sleep stages of human subjects. Los Angeles: University of California, Brain Information Service/BrainResearch Institute; 1968.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=A%20manual%20of%20standardized%20terminology%2C%20techniques%20and%20scoring%20system%20for%20sleep%20stages%20of%20human%20subjects&publication_year=1968&author=Rechtschaffen%2CA&author=Kales%2CA)
38. Stampi C, Mullington J, Rivers M, Campos JP, Broughton R. Ultrashort sleep schedules: sleep architecture and recuperative value of 80, 50 and 20 min naps. In: Horne JA, editor. Sleep '90. Bochum: Pontenagel Press; 1990. p. 71–4.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Ultrashort%20sleep%20schedules%3A%20sleep%20architecture%20and%20recuperative%20value%20of%2080%2C%2050%20and%2020%20min%20naps&pages=71-74&publication_year=1990&author=Stampi%2CC&author=Mullington%2CJ&author=Rivers%2CM&author=Campos%2CJP&author=Broughton%2CR)
39. Stones MJ. Memory performance after arousal from different sleep stages. Br J Psychol. 1977;68(2):177–81. [ https://doi.org/10.1111/j.2044-8295.1977.tb01573.x ](https://doi.org/10.1111/j.2044-8295.1977.tb01573.x) .
[ Article ](https://doi.org/10.1111%2Fj.2044-8295.1977.tb01573.x) [ CAS
](/articles/cas-redirect/1:STN:280:DyaE2s3gt1ensw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Memory%20performance%20after%20arousal%20from%20different%20sleep%20stages&journal=Br%20J%20Psychol&doi=10.1111%2Fj.2044-8295.1977.tb01573.x&volume=68&issue=2&pages=177-181&publication_year=1977&author=Stones%2CMJ)
40. Koulack D, Schultz KJ. Task performance after awakenings from different stages of sleep. Percept Mot Skills. 1974;39(2):792–4. [ https://doi.org/10.2466/pms.1974.39.2.792 ](https://doi.org/10.2466/pms.1974.39.2.792) .
[ Article ](https://doi.org/10.2466%2Fpms.1974.39.2.792) [ CAS
](/articles/cas-redirect/1:STN:280:DyaE2M7hvFOqtA%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Task%20performance%20after%20awakenings%20from%20different%20stages%20of%20sleep&journal=Percept%20Mot%20Skills&doi=10.2466%2Fpms.1974.39.2.792&volume=39&issue=2&pages=792-794&publication_year=1974&author=Koulack%2CD&author=Schultz%2CKJ)
41. Jewett ME, Wyatt JK, Ritz-De Cecco A, Khalsa SB, Dijk DJ, Czeisler CA. Time course of sleep inertia dissipation in human performance and alertness. J Sleep Res. 1999;8(1):1–8. [ https://doi.org/10.1111/j.1365-2869.1999.00128.x ](https://doi.org/10.1111/j.1365-2869.1999.00128.x) .
[ Article ](https://doi.org/10.1111%2Fj.1365-2869.1999.00128.x) [ CAS
](/articles/cas-redirect/1:STN:280:DyaK1M3gtl2ntw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Time%20course%20of%20sleep%20inertia%20dissipation%20in%20human%20performance%20and%20alertness&journal=J%20Sleep%20Res&doi=10.1111%2Fj.1365-2869.1999.00128.x&volume=8&issue=1&pages=1-8&publication_year=1999&author=Jewett%2CME&author=Wyatt%2CJK&author=Ritz-
De%20Cecco%2CA&author=Khalsa%2CSB&author=Dijk%2CDJ&author=Czeisler%2CCA)
42. Ekstedt M, Akerstedt T, Söderström M. Microarousals during sleep are associated with increased levels of lipids, cortisol, and blood pressure. Psychosom Med. 2004;66(6):925–31. [ https://doi.org/10.1097/01.psy.0000145821.25453.f7 ](https://doi.org/10.1097/01.psy.0000145821.25453.f7) .
[ Article ](https://doi.org/10.1097%2F01.psy.0000145821.25453.f7) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BD2cXpsl2htrY%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Microarousals%20during%20sleep%20are%20associated%20with%20increased%20levels%20of%20lipids%2C%20cortisol%2C%20and%20blood%20pressure&journal=Psychosom%20Med&doi=10.1097%2F01.psy.0000145821.25453.f7&volume=66&issue=6&pages=925-931&publication_year=2004&author=Ekstedt%2CM&author=Akerstedt%2CT&author=S%C3%B6derstr%C3%B6m%2CM)
43. Trinder J, Padula M, Berlowitz D, et al. Cardiac and respiratory activity at arousal from sleep under controlled ventilation conditions. J Appl Physiol (1985). 2001;90(4):1455–63. [ https://doi.org/10.1152/jappl.2001.90.4.1455 ](https://doi.org/10.1152/jappl.2001.90.4.1455) .
[ Article ](https://doi.org/10.1152%2Fjappl.2001.90.4.1455) [ CAS
](/articles/cas-redirect/1:STN:280:DC%2BD3MzgsFChug%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Cardiac%20and%20respiratory%20activity%20at%20arousal%20from%20sleep%20under%20controlled%20ventilation%20conditions&journal=J%20Appl%20Physiol%20%281985%29&doi=10.1152%2Fjappl.2001.90.4.1455&volume=90&issue=4&pages=1455-1463&publication_year=2001&author=Trinder%2CJ&author=Padula%2CM&author=Berlowitz%2CD)
44. Stepanski E, Lamphere J, Badia P, Zorick F, Roth T. Sleep fragmentation and daytime sleepiness. Sleep. 1984;7(1):18–26. [ https://doi.org/10.1093/sleep/7.1.18 ](https://doi.org/10.1093/sleep/7.1.18) .
[ Article ](https://doi.org/10.1093%2Fsleep%2F7.1.18) [ CAS ](/articles/cas-
redirect/1:STN:280:DyaL2c7psFygsw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20fragmentation%20and%20daytime%20sleepiness&journal=Sleep.&doi=10.1093%2Fsleep%2F7.1.18&volume=7&issue=1&pages=18-26&publication_year=1984&author=Stepanski%2CE&author=Lamphere%2CJ&author=Badia%2CP&author=Zorick%2CF&author=Roth%2CT)
45. Roehrs T, Merlotti L, Petrucelli N, Stepanski E, Roth T. Experimental sleep fragmentation. Sleep. 1994;17(5):438–43. [ https://doi.org/10.1093/sleep/17.5.438 ](https://doi.org/10.1093/sleep/17.5.438) .
[ Article ](https://doi.org/10.1093%2Fsleep%2F17.5.438) [ CAS ](/articles/cas-
redirect/1:STN:280:DyaK2M%2FosVSmsQ%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Experimental%20sleep%20fragmentation&journal=Sleep.&doi=10.1093%2Fsleep%2F17.5.438&volume=17&issue=5&pages=438-443&publication_year=1994&author=Roehrs%2CT&author=Merlotti%2CL&author=Petrucelli%2CN&author=Stepanski%2CE&author=Roth%2CT)
46. Borbély AA. A two process model of sleep regulation. Hum Neurobiol. 1982;1(3):195–204.
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=A%20two%20process%20model%20of%20sleep%20regulation&journal=Hum%20Neurobiol&volume=1&issue=3&pages=195-204&publication_year=1982&author=Borb%C3%A9ly%2CAA)
47. Folkard S, Åkerstedt TA. Three-process model of the regulation of alertness-sleepiness. In: Broughton RJ, Ogilvie RD, editors. Sleep, arousal, and performance. Boston: Birkhäuser; 1992. p. 11–26.
[ Google Scholar ](http://scholar.google.com/scholar_lookup?&title=Three-
process%20model%20of%20the%20regulation%20of%20alertness-
sleepiness&pages=11-26&publication_year=1992&author=Folkard%2CS&author=%C3%85kerstedt%2CTA)
48. Dijk D-J, Czeisler CA. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci Lett. 1994;166(1):63–8. [ https://doi.org/10.1016/0304-3940(94)90841-9 ](https://doi.org/10.1016/0304-3940\(94\)90841-9) .
[ Article ](https://doi.org/10.1016%2F0304-3940%2894%2990841-9) [ CAS
](/articles/cas-redirect/1:STN:280:DyaK2c3kslOnsQ%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Paradoxical%20timing%20of%20the%20circadian%20rhythm%20of%20sleep%20propensity%20serves%20to%20consolidate%20sleep%20and%20wakefulness%20in%20humans&journal=Neurosci%20Lett&doi=10.1016%2F0304-3940%2894%2990841-9&volume=166&issue=1&pages=63-68&publication_year=1994&author=Dijk%2CD-J&author=Czeisler%2CCA)
49. Fuller KH, Waters WF, Binks PG, Anderson T. Generalized anxiety and sleep architecture: a polysomnographic investigation. Sleep. 1997;20:370–6.
[ Article ](https://doi.org/10.1093%2Fsleep%2F20.5.370) [ CAS ](/articles/cas-
redirect/1:STN:280:DyaK1c%2FitVKnsw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Generalized%20anxiety%20and%20sleep%20architecture%3A%20a%20polysomnographic%20investigation&journal=Sleep&doi=10.1093%2Fsleep%2F20.5.370&volume=20&pages=370-376&publication_year=1997&author=Fuller%2CKH&author=Waters%2CWF&author=Binks%2CPG&author=Anderson%2CT)
50. Åkerstedt T, Hume K, Minors D, Waterhouse J. The meaning of good sleep: a longitudinal study of polysomnography and subjective sleep quality. J Sleep Res. 1994;3:152–8.
[ Article ](https://doi.org/10.1111%2Fj.1365-2869.1994.tb00122.x) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20meaning%20of%20good%20sleep%3A%20a%20longitudinal%20study%20of%20polysomnography%20and%20subjective%20sleep%20quality&journal=J%20Sleep%20Res&doi=10.1111%2Fj.1365-2869.1994.tb00122.x&volume=3&pages=152-158&publication_year=1994&author=%C3%85kerstedt%2CT&author=Hume%2CK&author=Minors%2CD&author=Waterhouse%2CJ)
51. Van De Werken M, Giménez MC, De Vries B, Beersma DG, Van Someren EJ, Gordijn MC. Effects of artificial dawn on sleep inertia, skin temperature, and the awakening cortisol response. J Sleep Res. 2010;19(3):425–35. [ https://doi.org/10.1111/j.1365-2869.2010.00828.x ](https://doi.org/10.1111/j.1365-2869.2010.00828.x) .
[ Article ](https://doi.org/10.1111%2Fj.1365-2869.2010.00828.x) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Effects%20of%20artificial%20dawn%20on%20sleep%20inertia%2C%20skin%20temperature%2C%20and%20the%20awakening%20cortisol%20response&journal=J%20Sleep%20Res&doi=10.1111%2Fj.1365-2869.2010.00828.x&volume=19&issue=3&pages=425-435&publication_year=2010&author=Werken%2CM&author=Gim%C3%A9nez%2CMC&author=Vries%2CB&author=Beersma%2CDG&author=Someren%2CEJ&author=Gordijn%2CMC)
[ Download references ](https://citation-
needed.springer.com/v2/references/10.1186/s40101-022-00317-w?format=refman&flavour=references)
## Acknowledgements
Not applicable.
## Funding
Not applicable.
## Author information
### Authors and Affiliations
1. Integrated Arts and Human Sciences Program, Graduate School of Humanities and Social Sciences, Hiroshima University, Higashi-Hiroshima, Hiroshima, Japan
Keiko Ogawa & Mitsuo Hayashi
2. School of Integrated Arts and Sciences, Hiroshima University, Higashi-Hiroshima, Hiroshima, Japan
Emi Kaizuma-Ueyama
Authors
1. Keiko Ogawa
[ View author publications
](https://www.biomedcentral.com/search?query=author%23Keiko%20Ogawa)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Keiko%20Ogawa)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Keiko%20Ogawa%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
2. Emi Kaizuma-Ueyama
[ View author publications
](https://www.biomedcentral.com/search?query=author%23Emi%20Kaizuma-Ueyama)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Emi%20Kaizuma-
Ueyama) [ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Emi%20Kaizuma-
Ueyama%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
3. Mitsuo Hayashi
[ View author publications
](https://www.biomedcentral.com/search?query=author%23Mitsuo%20Hayashi)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Mitsuo%20Hayashi)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Mitsuo%20Hayashi%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
### Contributions
EKU and MH conceived and designed research. EKU performed experiments. EKU and
KO analyzed data. EKU, KO and MH interpreted results of experiments. KO
prepared table and figure. KO and EKU drafted the manuscript. KO and MH edited
and revised the manuscript. All authors approved the final version of the
manuscript.
### Corresponding authors
Correspondence to [ Keiko Ogawa ](mailto:[email protected]) or [
Mitsuo Hayashi ](mailto:[email protected]) .
## Ethics declarations
### Ethics approval and consent to participate
All participants were given instructions about the purpose and risks of the
study and were required to give written informed consent. The experimental
procedure was approved by the Research Ethics Committee of Hiroshima
university (No. 30-02, 30-03). This study was performed in accordance with the
Declaration of Helsinki.
### Consent for publication
Not applicable.
### Competing interests
The authors declare that they have no competing interests.
## Additional information
### Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in
published maps and institutional affiliations.
## Rights and permissions
**Open Access** This article is licensed under a Creative Commons Attribution
4.0 International License, which permits use, sharing, adaptation,
distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to
the Creative Commons licence, and indicate if changes were made. The images or
other third party material in this article are included in the article's
Creative Commons licence, unless indicated otherwise in a credit line to the
material. If material is not included in the article's Creative Commons
licence and your intended use is not permitted by statutory regulation or
exceeds the permitted use, you will need to obtain permission directly from
the copyright holder. To view a copy of this licence, visit [
http://creativecommons.org/licenses/by/4.0/
](http://creativecommons.org/licenses/by/4.0/) . The Creative Commons Public
Domain Dedication waiver ( [ http://creativecommons.org/publicdomain/zero/1.0/
](http://creativecommons.org/publicdomain/zero/1.0/) ) applies to the data
made available in this article, unless otherwise stated in a credit line to
the data.
[ Reprints and permissions
](https://s100.copyright.com/AppDispatchServlet?title=Effects%20of%20using%20a%20snooze%20alarm%20on%20sleep%20inertia%20after%20morning%20awakening&author=Keiko%20Ogawa%20et%20al&contentID=10.1186%2Fs40101-022-00317-w©right=The%20Author%28s%29&publication=1880-6805&publicationDate=2022-12-31&publisherName=SpringerNature&orderBeanReset=true&oa=CC%20BY%20%2B%20CC0)
## About this article
[ 
](https://crossmark.crossref.org/dialog/?doi=10.1186/s40101-022-00317-w)
### Cite this article
Ogawa, K., Kaizuma-Ueyama, E. & Hayashi, M. Effects of using a snooze alarm on
sleep inertia after morning awakening. _J Physiol Anthropol_ **41** , 43
(2022). https://doi.org/10.1186/s40101-022-00317-w
[ Download citation ](https://citation-
needed.springer.com/v2/references/10.1186/s40101-022-00317-w?format=refman&flavour=citation)
* Received : 08 August 2022
* Accepted : 09 December 2022
* Published : 31 December 2022
* DOI : https://doi.org/10.1186/s40101-022-00317-w
### Share this article
Anyone you share the following link with will be able to read this content:
Get shareable link
Sorry, a shareable link is not currently available for this article.
Copy to clipboard
Provided by the Springer Nature SharedIt content-sharing initiative
### Keywords
* [ Sleep loss ](/search?query=Sleep%20loss&facet-discipline="Biomedicine")
* [ Snooze alarm ](/search?query=Snooze%20alarm&facet-discipline="Biomedicine")
* [ Awakening ](/search?query=Awakening&facet-discipline="Biomedicine")
* [ Sleep inertia ](/search?query=Sleep%20inertia&facet-discipline="Biomedicine")
* [ Normal nocturnal sleep ](/search?query=Normal%20nocturnal%20sleep&facet-discipline="Biomedicine")
* [ Polysomnograms ](/search?query=Polysomnograms&facet-discipline="Biomedicine")
[ Download PDF
](//jphysiolanthropol.biomedcentral.com/counter/pdf/10.1186/s40101-022-00317-w.pdf)
[ Download ePub
](//jphysiolanthropol.biomedcentral.com/counter/epub/10.1186/s40101-022-00317-w.epub)
Advertisement
[

](//pubads.g.doubleclick.net/gampad/jump?iu=/270604982/bmc/jphysiolanthropol/articles&sz=300x250&pos=MPU1&doi=10.1186/s40101-022-00317-w&type=article&kwrd=Sleep
loss,Snooze alarm,Awakening,Sleep inertia,Normal nocturnal
sleep,Polysomnograms&pmc=B13004,X12000&)
#### Journal of Physiological Anthropology
ISSN: 1880-6805
#### Contact us
* Submission enquiries: [ Access here and click Contact Us ](https://www.editorialmanager.com/jpha/)
* General enquiries: [ [email protected] ](mailto:[email protected])

* [ Read more on our blogs ](https://blogs.biomedcentral.com/)
* [ Receive BMC newsletters ](//www.biomedcentral.com/login)
* [ Manage article alerts ](//www.biomedcentral.com/account)
* [ Language editing for authors ](https://authorservices.springernature.com/go/sn/?utm_source=Website&utm_medium=BMC&utm_campaign=SNAS+Referrals+2022&utm_id=ref2022)
* [ Scientific editing for authors ](https://authorservices.springernature.com/go/sn/?utm_source=Website&utm_medium=BMC&utm_campaign=SNAS+Referrals+2022&utm_id=ref2022)
* [ Policies ](//www.biomedcentral.com/about/policies)
* [ Accessibility ](//www.biomedcentral.com/accessibility)
* [ Press center ](//www.biomedcentral.com/about/press-centre)
* [ Support and Contact ](https://support.biomedcentral.com/support/home)
* [ Leave feedback ](https://biomedcentral.typeform.com/to/VLXboo)
* [ Careers ](//www.biomedcentral.com/about/jobs)
### Follow BMC
* [ BMC Twitter page ](https://twitter.com/biomedcentral)
* [ BMC Facebook page ](https://www.facebook.com/BioMedCentral)
* [ BMC Weibo page ](http://www.weibo.com/biomedcentral)
By using this website, you agree to our [ Terms and Conditions
](//www.biomedcentral.com/terms-and-conditions) , [ Your US state privacy
rights ](https://www.springernature.com/ccpa) , [ Privacy statement
](//www.biomedcentral.com/privacy-statement) and [ Cookies
](//www.biomedcentral.com/cookies) policy. [ Your privacy choices/Manage
cookies ](javascript:void\(0\);) we use in the preference centre.

© 2024 BioMed Central Ltd unless otherwise stated. Part of [ Springer Nature
](https://www.springernature.com) .
*[
DOI
]: Digital Object Identifier
| biology | 498444 | https://no.wikipedia.org/wiki/Asunder | Asunder | Asunder er Heaven Shall Burns første studioalbum. Det ble utgitt 8. april 2000.
Sporliste
«To Inherit the Guilt» (3.05)
«Cold» (4.20)
«Betrayed Again» (3.52)
«Deification» (5.11)
«Pass Away» (3.30)
«Open Arms to the Future» (2.55)
«The Drowned and the Saved» (4.07)
«Where Is the Light» (2.56)
«Asunder» (4.48)
«The Fourth Crusade» (4.52)
«Battlecries» (26.23)
Notater
Låt nr. 10, «The Fourth Crusade», er en coverversjon av Bolt Throwers sang «The IVth Crusade».
Låt nr. 11 er coverversjonen av «Battlecries», originalt av Liar, også 23 minutter stillhet.
Kilder
Asunder fra Encyclopaedia Metallum
Eksterne lenker
Debutalbum fra 2000
Heaven Shall Burn-album | norwegian_bokmål | 0.950343 |
wake_up_wrong/s41467022345032.txt | Skip to main content
Thank you for visiting nature.com. You are using a browser version with
limited support for CSS. To obtain the best experience, we recommend you use a
more up to date browser (or turn off compatibility mode in Internet Explorer).
In the meantime, to ensure continued support, we are displaying the site
without styles and JavaScript.
Advertisement
[

](//pubads.g.doubleclick.net/gampad/jump?iu=/285/nature_communications/article&sz=728x90&c=-1354327232&t=pos%3Dtop%26type%3Darticle%26artid%3Ds41467-022-34503-2%26doi%3D10.1038/s41467-022-34503-2%26techmeta%3D141%26subjmeta%3D1385,208,2811,378,477,631,729%26kwrd%3DCircadian+rhythms+and+sleep,Heritable+quantitative+trait,Human+behaviour)
[ 
](/ncomms)
* [ View all journals ](https://www.nature.com/siteindex)
* Search
* [ Log in ](https://idp.nature.com/auth/personal/springernature?redirect_uri=https://www.nature.com/articles/s41467-022-34503-2)
* Explore content
* About the journal
* Publish with us
* [ Sign up for alerts ](https://idp.nature.com/auth/personal/springernature?redirect_uri=https%3A%2F%2Fwww.nature.com%2Fmy-account%2Falerts%2Fsubscribe-journal%3Flist-id%3D264%26journal-link%3Dhttps%253A%252F%252Fwww.nature.com%252Fncomms%252F)
* [ RSS feed ](https://www.nature.com/ncomms.rss)
1. [ nature ](/)
2. [ nature communications ](/ncomms)
3. [ articles ](/ncomms/articles?type=article)
4. article
How people wake up is associated with previous night’s sleep together with
physical activity and food intake
[ Download PDF ](/articles/s41467-022-34503-2.pdf)
[ Download PDF ](/articles/s41467-022-34503-2.pdf)
* Article
* [ Open access ](https://www.springernature.com/gp/open-research/about/the-fundamentals-of-open-access-and-open-research)
* Published: 19 November 2022
# How people wake up is associated with previous night’s sleep together with
physical activity and food intake
* Raphael Vallat [ ORCID: orcid.org/0000-0003-1779-7653 ](http://orcid.org/0000-0003-1779-7653) 1 ,
* Sarah E. Berry [ ORCID: orcid.org/0000-0002-5819-5109 ](http://orcid.org/0000-0002-5819-5109) 2 ,
* Neli Tsereteli 3 ,
* Joan Capdevila 4 ,
* Haya Al Khatib 2 , 4 ,
* Ana M. Valdes [ ORCID: orcid.org/0000-0003-1141-4471 ](http://orcid.org/0000-0003-1141-4471) 5 , 6 ,
* Linda M. Delahanty [ ORCID: orcid.org/0000-0002-1525-3559 ](http://orcid.org/0000-0002-1525-3559) 7 ,
* David A. Drew [ ORCID: orcid.org/0000-0002-8813-0816 ](http://orcid.org/0000-0002-8813-0816) 8 , 9 ,
* Andrew T. Chan [ ORCID: orcid.org/0000-0001-7284-6767 ](http://orcid.org/0000-0001-7284-6767) 8 , 9 ,
* Jonathan Wolf [ ORCID: orcid.org/0000-0002-0530-2257 ](http://orcid.org/0000-0002-0530-2257) 4 ,
* Paul W. Franks [ ORCID: orcid.org/0000-0002-0520-7604 ](http://orcid.org/0000-0002-0520-7604) 3 , 10 , 11 ,
* Tim D. Spector [ ORCID: orcid.org/0000-0002-9795-0365 ](http://orcid.org/0000-0002-9795-0365) 10 &
* …
* Matthew P. Walker [ ORCID: orcid.org/0000-0002-7839-6389 ](http://orcid.org/0000-0002-7839-6389) 1
Show authors
[ _Nature Communications_ ](/ncomms) ** volume 13 ** , Article number: 7116
( 2022 ) Cite this article
* 76k Accesses
* 5 Citations
* 1521 Altmetric
* [ Metrics details ](/articles/s41467-022-34503-2/metrics)
### Subjects
* [ Circadian rhythms and sleep ](/subjects/circadian-rhythms-and-sleep)
* [ Heritable quantitative trait ](/subjects/heritable-quantitative-trait)
* [ Human behaviour ](/subjects/human-behaviour)
## Abstract
How people wake up and regain alertness in the hours after sleep is related to
how they are sleeping, eating, and exercising. Here, in a prospective
longitudinal study of 833 twins and genetically unrelated adults, we
demonstrate that how effectively an individual awakens in the hours following
sleep is not associated with their genetics, but instead, four independent
factors: sleep quantity/quality the night before, physical activity the day
prior, a breakfast rich in carbohydrate, and a lower blood glucose response
following breakfast. Furthermore, an individual’s set-point of daily alertness
is related to the quality of their sleep, their positive emotional state, and
their age. Together, these findings reveal a set of non-genetic (i.e., not
fixed) factors associated with daily alertness that are modifiable.
### Similar content being viewed by others

### [ Microdosing with psilocybin mushrooms: a double-blind placebo-
controlled study
](https://www.nature.com/articles/s41398-022-02039-0?fromPaywallRec=false)
Article Open access 02 August 2022

### [ A systematic review and multivariate meta-analysis of the physical and
mental health benefits of touch interventions
](https://www.nature.com/articles/s41562-024-01841-8?fromPaywallRec=false)
Article Open access 08 April 2024

### [ Associations of dietary patterns with brain health from behavioral,
neuroimaging, biochemical and genetic analyses
](https://www.nature.com/articles/s44220-024-00226-0?fromPaywallRec=false)
Article Open access 01 April 2024
## Introduction
What factors will influence how you wake up tomorrow morning, predicting
whether or not you will feel alert, and then be able to sustain that level of
attentive waking consciousness across the day? This question is scientifically
elemental but also of societal relevance considering that the failure to
sustain alertness throughout the day is a major causal factor of road traffic
and occupational accidents, accounting for thousands of deaths every year [ 1
](/articles/s41467-022-34503-2#ref-CR1 "Tefft, B. C. & Others. Prevalence of
motor vehicle crashes involving drowsy drivers, United States, 2009-2013.
\(Citeseer, 2014\).") , [ 2 ](/articles/s41467-022-34503-2#ref-CR2 "Rakel, R.
E. Clinical and societal consequences of obstructive sleep apnea and excessive
daytime sleepiness. Postgrad. Med. 121, 86–95 \(2009\).") . Moreover, it is
estimated that insufficient sleep leading to impaired daytime alertness is
responsible for significant work-related loss of productivity, greater
healthcare utilisation and work absenteeism, thereby costing developed nations
about 2% of their gross domestic product each year (i.e., $411 billion dollars
in the United States alone) [ 3 ](/articles/s41467-022-34503-2#ref-CR3
"Hafner, M., Stepanek, M., Taylor, J., Troxel, W. M. & van Stolk, C. Why sleep
matters-the economic costs of insufficient sleep: a cross-country comparative
analysis. Rand Health Q 6, 11 \(2017\).") .
In addition, the inability to transition effectively to a state of functional
cognitive alertness upon awakening from sleep—known as “sleep inertia” [ 4
](/articles/s41467-022-34503-2#ref-CR4 "Vallat, R., Meunier, D., Nicolas, A. &
Ruby, P. Hard to wake up? The cerebral correlates of sleep inertia assessed
using combined behavioral, EEG and fMRI measures. Neuroimage 184, 266–278
\(2019\).") —is a serious safety risk for workers performing hazardous tasks
immediately upon awakening, one that has cost such individuals either their
own lives or the lives of others (e.g. military personnel, healthcare workers,
firefighters, pilots) 5 , 6 , [ 7 ](/articles/s41467-022-34503-2#ref-CR7
"Trotti, L. M. Waking up is the hardest thing I do all day: Sleep inertia and
sleep drunkenness. Sleep. Med. Rev. 35, 76–84 \(2017\).") .
Despite the severity, magnitude, and scope of these consequences, the unique
factors that influence how each of us wakes up and sustain meaningful
alertness throughout a waking day are poorly understood at both the extrinsic
and intrinsic (biological) level.
Addressing these issues, here, we sought to test whether a set of a priori
factors are associated with alertness in the first hours after awakening from
sleep.
Building on previous research, and using the Personalized Responses to Dietary
Composition Trial 1 (“PREDICT1”) [ 8 ](/articles/s41467-022-34503-2#ref-CR8
"Berry, S. et al. Personalised REsponses to DIetary Composition Trial
\(PREDICT\): an intervention study to determine inter-individual differences
in postprandial response to foods. Protocol Exchange
https://doi.org/10.21203/rs.2.20798/v1
\(2020\).") , [ 9 ](/articles/s41467-022-34503-2#ref-CR9 "Berry, S. E. et al. Human postprandial responses to food and potential for precision nutrition. Nat. Med.
https://doi.org/10.1038/s41591-020-0934-0
\(2020\).") , we targeted four interrelated hypotheses, the motivational evidence for which we describe below. First, we tested the hypothesis that an individual’s unique sleep profile the night prior — i.e., their sleep duration, sleep efficiency, and sleep timing — predicts subsequent changes in next-morning alertness. Specifically, a higher-than-typical sleep duration and sleep efficiency for any one individual would uniquely predict superior (higher) next-day alertness for that individual. Second, higher levels of physical activity on the day prior would predict an increase in next-day morning alertness. Third, that the macronutrient composition of breakfast, and independent of that composition, the unique associated blood glucose response, each selectively influences morning alertness. Fourth, and beyond these modifiable lifestyle factors (i.e., sleep, food, physical activity), we additionally sought to test whether daytime alertness is under significant genetic heritability using twin-pair genetic modeling.
## Results
### Modifiable lifestyle factors are associated with day-to-day fluctuations
in morning alertness
In short (and see “Methods”), PREDICT1 is a prospective longitudinal study
including a thousand twins and genetically unrelated adults. During the two
weeks of the study, participants consumed multiple standardised breakfast
meals differing in nutritional composition (Table [ 1
](/articles/s41467-022-34503-2#Tab1) ), while wearing an accelerometer
wristwatch and a continuous glucose monitor (CGM). Participants also recorded
their food intake on a dedicated study app throughout the study, together with
their alertness levels on a 0-100 visual analogue scale at several time points
after the logging of each meal (Fig. [ 1 ](/articles/s41467-022-34503-2#Fig1)
). Demographics and descriptive statistics of the dataset are reported in
Table [ 2 ](/articles/s41467-022-34503-2#Tab2) and Fig. [ 2
](/articles/s41467-022-34503-2#Fig2) .
**Table 1 Nutritional composition of the standardised breakfast meals**
[ Full size table ](/articles/s41467-022-34503-2/tables/1)
**Fig. 1: Experimental design.**
[ 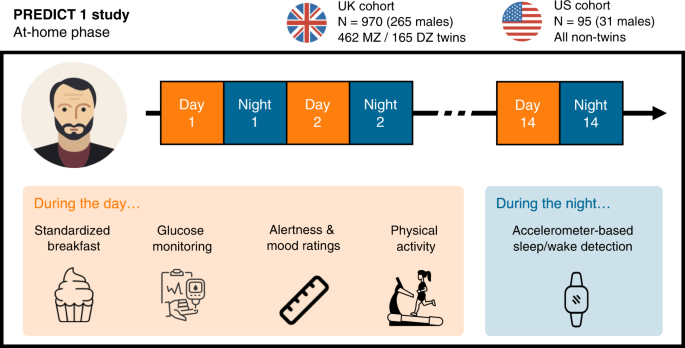
](/articles/s41467-022-34503-2/figures/1)
The Personalized Responses to Dietary Composition Trial (or “PREDICT1”) is a
two-country (UK, US) longitudinal study whose primary goal is to predict
metabolic responses to foods based on the individual’s characteristics,
including molecular biomarkers and lifestyle factors, as well as the
nutritional composition of the food [ 8 ](/articles/s41467-022-34503-2#ref-
CR8 "Berry, S. et al. Personalised REsponses to DIetary Composition Trial
\(PREDICT\): an intervention study to determine inter-individual differences
in postprandial response to foods. Protocol Exchange
https://doi.org/10.21203/rs.2.20798/v1
\(2020\).") . PREDICT1 consists of one clinic baseline visit followed by a two-week home-based study. During the at-home phase, participants consumed multiple standardised test meals differing in macronutrient composition, while wearing an accelerometer wristwatch and a continuous glucose monitor. The former was used to determine sleep/wake activity during the night and physical activity during the day. The continuous glucose monitor was used to measure postprandial glucose response. Participants also recorded their dietary intake, satiety, mood, and exercise on the study app throughout the study. The app also prompted participants to report their alertness levels on a 0–100 visual analogue scale at t = 0 minutes (time of logging of a meal) and regular intervals following the logging of a meal (see “Methods”). Source data are provided as a Source Data file. The cupcake icon, the ruler icon and the smartwatch icon were purchased and downloaded from thenounproject.com. All other icons were purchased and/or downloaded from iconfinder.com. MZ monozygotic, DZ dizygotic.
[ Full size image ](/articles/s41467-022-34503-2/figures/1)
**Table 2 Demographics and descriptive statistics of sleep and alertness**
[ Full size table ](/articles/s41467-022-34503-2/tables/2)
**Fig. 2: Alertness ratings throughout the day.**
[ 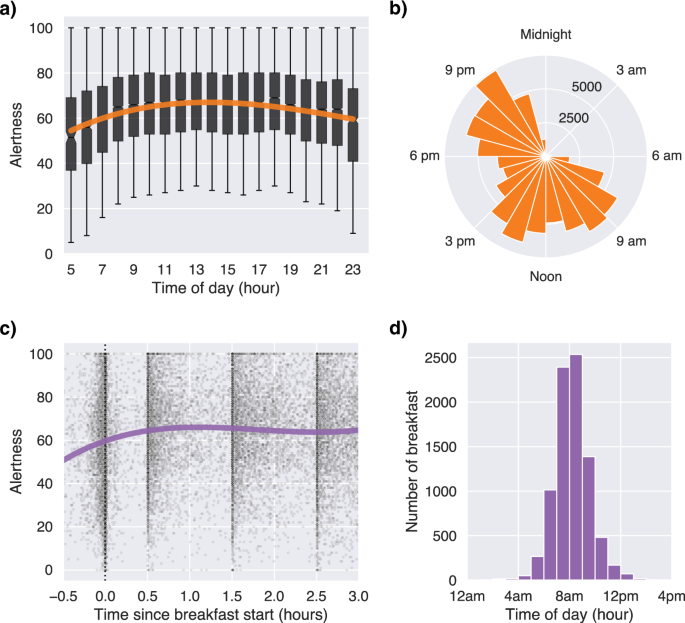
](/articles/s41467-022-34503-2/figures/2)
**a** Alertness as a function of time of day. The orange line shows a cubic
regression of all the alertness ratings logged between 5am and midnight (n =
89,440). Alertness progressively increased in the first hours of the morning,
reached a plateau during midday and progressively decreased in the evening.
Sample size for each unique box is shown in panel B. Box plots show centre
line as median, box limits as upper and lower quartiles. The notches represent
confidence intervals around the median. The whiskers extend from the box
limits by 1x the interquartile range. **b** Polar histogram of the number of
alertness ratings as a function of time of day. **c** Alertness ratings within
the first three hours after breakfast onset. Participants were instructed to
rate their alertness at t = 0 min, t = 30, t = 90 and t = 150 min after
breakfast start. During that ~3 hour period, they were also instructed to fast
and avoid physical activity. Each black dot represents one alertness rating
from one participant. The purple line shows a cubic regression of all the
morning alertness ratings. Alertness immediately increased after breakfast,
and then plateaued for the subsequent 2.5 hours. **d** Distribution of
breakfast start time. By definition in the protocol, the first alertness
rating of the day must coincide with breakfast onset. Source data are provided
as a Source Data file.
[ Full size image ](/articles/s41467-022-34503-2/figures/2)
We first tested the experimental hypothesis that day-to-day fluctuations in
alertness are associated with changes in sleep the night prior, physical
activity across the previous day, as well as first food intake upon awakening
for the analysis day in question (targeting breakfast nutritional composition
and consequential blood sugar response).
The average of all initial morning alertness ratings logged by participants
within the first three hours after the start of the standardised breakfast
meal was used to compute a daily morning alertness score for each participant
(Fig. [ 2c, d ](/articles/s41467-022-34503-2#Fig2) ). This time period was
defined within the experimental design, such that participants were instructed
to first rate their alertness in the morning when they started their
breakfast, and then to rate their alertness at several time points during the
three hours following breakfast start (t = 0, +0.5, +1.5, and +2.5 hours).
Participants were further instructed to avoid any snacking and/or physical
activity during that time window (see “Methods”). This ensured a null or low
impact of potential confounders (snacks, physical activity) of morning
alertness, as well as a high temporal causality between the predictors of
interest — prior sleep, prior physical activity, same-day morning breakfast
composition and consequential glucose response, and the outcome of interest:
alertness. On average, participants started their breakfast at 8h12 am ± 1h24
min (Fig. [ 2d ](/articles/s41467-022-34503-2#Fig2) ). The average latency
between the sleep offset (estimated from the accelerometer) and breakfast
start was 1h08 min ± 1h21 min.
A linear mixed-effects model was built to test the association between each
aforementioned predictor and morning alertness. A detailed description of the
predictors can be found in the “Methods” section. Importantly, all predictors
were included in a single linear mixed model to estimate the contribution of
each predictor, whilst simultaneously adjusting for all the others. All linear
mixed models reported thereafter were adjusted for age, sex, body mass index
(BMI), zygosity, sunrise time, daylight saving time (DST) and weekend. To
account for the natural between-person variability in sleep, the sleep
predictors — duration, efficiency and waketime — were normalised using person-
mean centering, i.e., expressed as a deviation from a person’s average across
the two weeks of the study. Unless otherwise specified, all p-values reported
in the following paragraphs were obtained from two-tailed Wald tests.
Consistent with the hypothesis, analyses demonstrated that (1) prior sleep
parameters, (2) breakfast composition and its associated post-breakfast blood
glucose response, and (3) physical activity of the previous day were each
significantly, and independently, predictive of morning alertness (Fig. [ 3
](/articles/s41467-022-34503-2#Fig3) and Supplementary Table [ 1
](/articles/s41467-022-34503-2#MOESM1) ).
**Fig. 3: Predictors of day-to-day fluctuations in morning alertness.**
[ 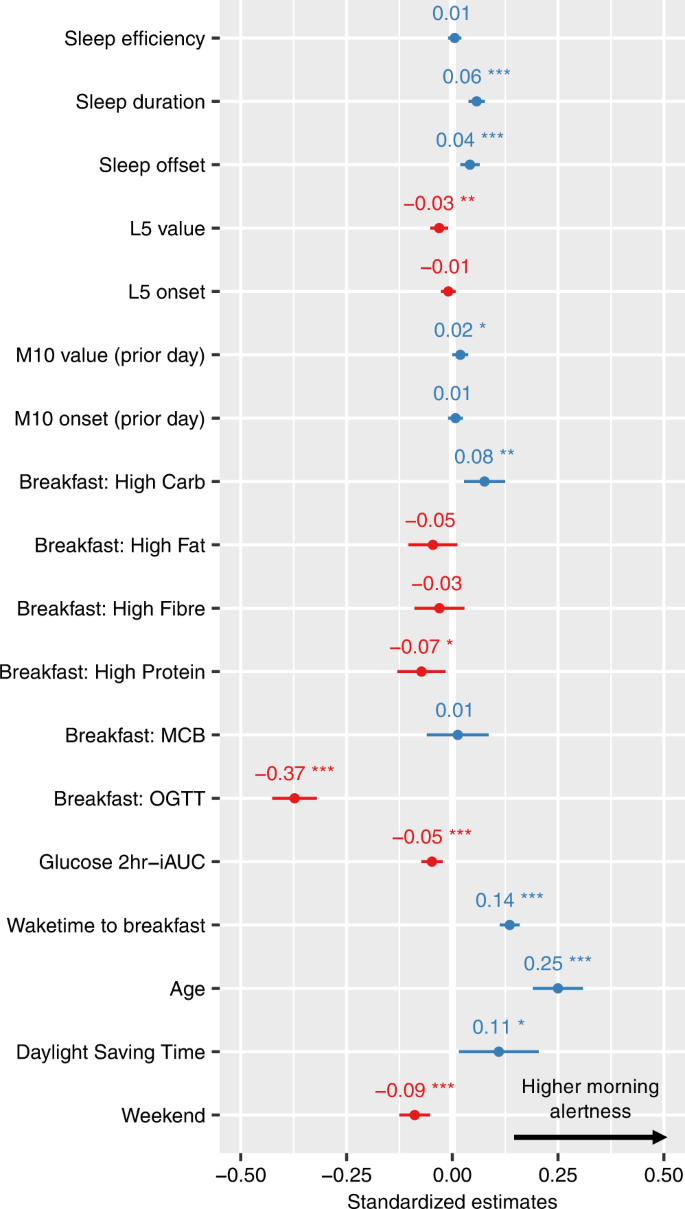
](/articles/s41467-022-34503-2/figures/3)
Standardised regression coefficients and confidence intervals from a linear
mixed effect model. Sex, BMI, zygosity and sunrise time were also included in
the model but are not reported here for conciseness since none of them was a
significant predictor of morning alertness. Unstandardised regression
coefficients and raw p-values can be found in Supplementary Table [ 1
](/articles/s41467-022-34503-2#MOESM1) . Sleep predictors were normalised
using person-mean centering. The dependent variable of the model is morning
alertness, which is calculated by averaging the alertness ratings that were
made within the first three hours after breakfast start (n = 6,744
observations). Family ID and participant ID were defined as nested random
effects of the linear mixed model. Predictors with a positive coefficient (i.e
predicting higher morning alertness) are shown in blue, while those with a
negative coefficient (lower morning alertness) are shown in red. Error bars
represent 95% confidence intervals. Stars indicate significance. P-values are
based on two-tailed Wald tests (degrees of freedom = 6717) and are not
adjusted for multiple comparisons. *p < 0.05, **p < 0.01, ***p < 0.001. L5 =
least active 5 hours of the day, M10 = most active 10 hours of the day, MCB
metabolic challenge breakfast, OGTT Oral Glucose Tolerance Test, iAUC
incremental area under the curve. Source data are provided as a Source Data
file.
[ Full size image ](/articles/s41467-022-34503-2/figures/3)
First, sleeping longer than one’s typical sleep duration was associated with
higher next-morning alertness (p < 0.001). Waking up later than one’s own
typical wake-up time was also associated with higher subsequent alertness,
even when controlling for sleep duration (p < 0.001). A similar effect was
observed for sleep onset: going to bed later than usual for a specific
individual was associated with higher morning alertness (p < 0.001;
Supplementary Table [ 2 ](/articles/s41467-022-34503-2#MOESM1) , note that
sleep offset and sleep onset could not be included in the same model because
of high collinearity with sleep duration). There was no significant
interaction between sleep duration and sleep timing (p = 0.792; Supplementary
Fig. [ 1 ](/articles/s41467-022-34503-2#MOESM1) ). Sleep efficiency did not
significantly predict morning alertness (p = 0.48). Taken together, this first
set of data demonstrates that sleeping longer and/or later than typical is
associated with higher next-morning alertness.
Second, the amount of physical activity that occurred across the prior day
also predicted morning alertness that following day. The average acceleration
value of the most active 10 hours of the previous daytime prior (the so-called
M10, see “Methods”) was associated with higher alertness (p = 0.049).
Conversely, the activity level in the nighttime (so-called L5, see “Methods”)
was related to worse next-day morning alertness (p = 0.004). Therefore, higher
levels of movement activity during the day (indicative of daytime physical
movement activity), yet lower levels of physical movement activity at night,
associated with more continuous and less disrupted sleep [ 10
](/articles/s41467-022-34503-2#ref-CR10 "Gonçalves, B. S. B., Adamowicz, T.,
Louzada, F. M., Moreno, C. R. & Araujo, J. F. A fresh look at the use of
nonparametric analysis in actimetry. Sleep. Med. Rev. 20, 84–91 \(2015\).") ,
each predicted superior morning alertness.
Third, breakfast composition the morning of significantly predicted subsequent
alertness (Fig. [ 3 ](/articles/s41467-022-34503-2#Fig3) and Supplementary
Table [ 1 ](/articles/s41467-022-34503-2#MOESM1) ). Specifically, the High
Carbohydrates breakfast meal was associated with higher morning alertness (p =
0.002), relative to the reference standardised meal consisting of a medium
amount of fat and carbohydrate (the “UK Average”, see Table [ 1
](/articles/s41467-022-34503-2#Tab1) and Supplementary Fig. [ 2
](/articles/s41467-022-34503-2#MOESM1) ). By contrast, the High Protein meal
was associated with lower alertness compared to the reference meal (p =
0.012). The strongest effect, however, was found for days in which
participants consumed a pure glucose liquid for breakfast (i.e., Oral Glucose
Tolerance Test, or OGTT, see Table [ 1 ](/articles/s41467-022-34503-2#Tab1) ).
Here, and compared to the reference standardised meal, consumption of the OGTT
was associated with marked reductions in subsequent alertness (p < 0.001). In
fact, alertness following the OGTT meal was significantly lower than alertness
following all the other standardised breakfast meals (Supplementary Fig. [ 2
](/articles/s41467-022-34503-2#MOESM1) ).
Since the OGTT is a non-standard breakfast meal, and participants were asked
not to consume caffeinated drinks the morning of the OGTT (see “Methods”), the
inclusion of the OGTT as a breakfast meal could potentially bias the
association between alertness and the standardised meals. However, additional
analyses revealed that all the above effects remained similar when removing
the OGTT from the analysis while leaving other standard breakfast meals within
the model (Supplementary Table [ 3 ](/articles/s41467-022-34503-2#MOESM1) ).
Moreover, caffeine intake was largely absent for the majority of observations
(86% null + 12% missing = 98%), suggesting that most breakfast meals were not
accompanied by any caffeinated drinks. Because of this low variance, breakfast
caffeine intake was not included in the model. Nevertheless, including
breakfast caffeine intake did not change the significance of the other
predictors, and breakfast caffeine intake by itself was not a significant
predictor of morning alertness — both with and without the OGTT (p = 0.11 and
p = 0.605 respectively).
The above data describe the association between the nutritional composition of
the breakfast meal consumed and subsequent morning alertness. However, such
data do not address the metabolic downstream consequence of that meal: the
physiological glucose response to the breakfast meal. Addressing this
question, and using a CGM device (see “Methods”), post-breakfast glucose
levels were uniquely associated with subsequent morning alertness, such that a
lower post-breakfast glycemic load (i.e., area under the curve of blood
glucose in the 2 hours following the breakfast) predicted a superior
subsequent alertness (p < 0.001; Fig. [ 3 ](/articles/s41467-022-34503-2#Fig3)
and Supplementary Table [ 1 ](/articles/s41467-022-34503-2#MOESM1) ).
As a collective, analyses of this first component of the study demonstrate
that morning alertness was significantly, and independently so, associated
with the factors of (1) sleep (specifically a longer sleep duration, the
offset timing of a later morning awakening and lower levels of movement during
the night), (2) physical activity (increased activity on the previous day),
(3) breakfast composition (high carbohydrates meal), and (4) post-breakfast
blood glucose response (lower glycemic load).
### Predicting an individual’s baseline levels of alertness
One significant advantage of the current longitudinal design is the ability to
resolve personalised, within-individual unique changes. To analytically
exploit these unique, within-individual dynamics, a hold-out validation was
conducted to determine whether the morning alertness of a specific given
individual on a given morning was accurately predicted by the aforementioned
modifiable lifestyle factors (Fig. [ 4a ](/articles/s41467-022-34503-2#Fig4)
). Indeed, the model was able to explain 59% of the variance in morning
alertness in the testing set (Fig. [ 4b ](/articles/s41467-022-34503-2#Fig4)
). This model was then compared to a dummy model that predicted the average
morning alertness for each individual across all days of the training dataset
(Fig. [ 4c ](/articles/s41467-022-34503-2#Fig4) ). A Bayesian model comparison
revealed that the main experimental model was significantly better than the
dummy model (log[Bayes Factor] =181.3), although the added gain in explained
variance was relatively small (5%). The latter finding suggests a substantive
consistency of morning alertness level among the same individual (i.e., low
within-individual variance). Consistent with this finding, the intraclass
correlation coefficient of the dummy model was 0.56, indicating that between-
individual variability accounted for a larger total amount of variance in
morning alertness.
**Fig. 4: Predictions of morning alertness from the hold-out validation.**
[ 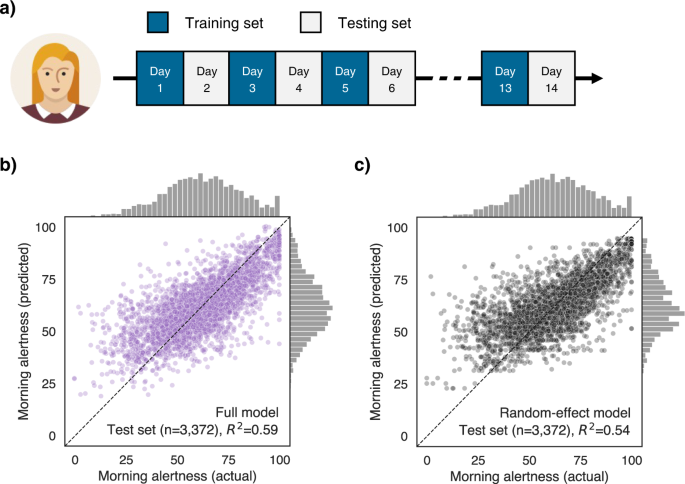
](/articles/s41467-022-34503-2/figures/4)
**a** Hold-out validation strategy. For each participant, the multilevel model
was trained on half the available days and then tested on the remaining half.
Predictors of the multilevel model are shown in Fig. [ 3
](/articles/s41467-022-34503-2#Fig3) . The dependent variable of the model is
morning alertness, which is calculated by averaging the alertness ratings that
were made within the first three hours after breakfast start. Family ID and
participant ID were defined as nested random effects of the linear mixed
model. **b** Scatter plot showing the true and predicted values of morning
alertness from a full model that included all the aforementioned predictors.
Each dot in the scatter plot represents the morning alertness value from one
day from one participant. **c** True and predicted values of morning alertness
from a naive model that only included random effects. Predicted values are
therefore, for each participant, the average of all the morning alertness
values in the training set. Source data are provided as a Source Data file.
The woman icon was downloaded from iconfinder.com.
[ Full size image ](/articles/s41467-022-34503-2/figures/4)
As described above, that nevertheless substantial within-individual variance
can be explained by the a priori collection of preceding factors of 1) prior
sleep, 2) prior physical activity, and 3) morning food composition. However,
these analyses do not address the second main experimental question: what
factor(s) then explain the even larger between-individual variability in
levels of alertness. For example, is this set-point of daytime alertness
across individuals genetically determined, and thus inherently fixed for each
individual. Or rather, is this alertness set-point across the population
influenced by external, modifiable trait factors (e.g. lifestyle, habitual
behaviours, health)? The latter hypothesis is of particular relevance, as it
would suggest that long-term targeted public-health interventions on these
modifiable factors may provide a way to improve (i.e., shift up) an
individual’s set-point in daytime alertness.
To test this hypothesis, the average alertness across all days of the
longitudinal study, termed, “trait daytime alertness”, was calculated for each
participant. A machine-learning approach then evaluated the ranked importance
of trait predictors, including demographics, mental health, mood, habitual
eating behaviours, as well as subjective and objective markers of sleep and
physical activity. The model used a three-fold cross-validation approach
coupled with a gradient boosting estimator to predict the baseline alertness
of all participants based on all above predictors. Gradient boosting
algorithms are optimal for this sort of task as they — unlike standard
regression models — natively handle missing values in predictors and are
robust to highly correlated predictors [ 11
](/articles/s41467-022-34503-2#ref-CR11 "Ke, G. et al. LightGBM: A Highly
Efficient Gradient Boosting Decision Tree. in Advances in Neural Information
Processing Systems 30 \(eds. Guyon, I. et al.\) 3146–3154 \(Curran Associates,
Inc., 2017\).") . The importance rank of each predictor was then calculated on
the full dataset using Shapley (SHAP) values (see “Methods”), which quantify
the exact impact of a given feature — after accounting for all other features
— on the predicted alertness outcome of the model.
Consistent with the hypothesis, the machine-learning model accurately and
significantly predicted what an individual’s typical level of alertness would
be. Indeed, the model explained more than a third of the variance of someone’s
set-point alertness in a previously unseen (cross-validated) dataset (Fig. [
5a ](/articles/s41467-022-34503-2#Fig5) , with no statistical difference
between the means of the predicted and ground-truth alertness values: paired
T-test, T(832)=−0.07, p = 0.94).
**Fig. 5: Predictors of trait alertness.**
[ 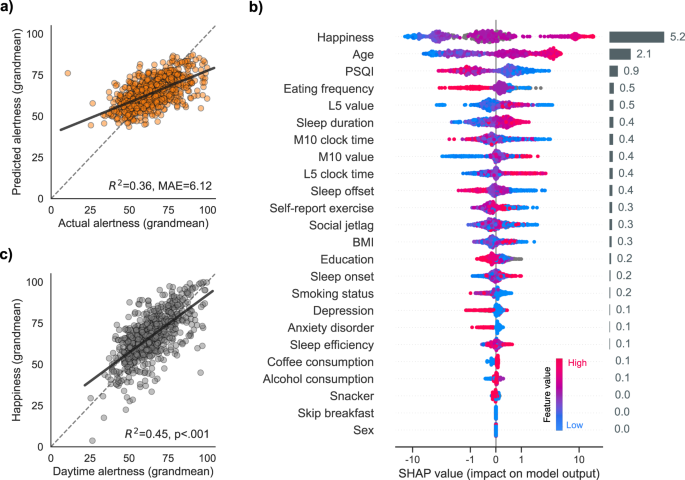
](/articles/s41467-022-34503-2/figures/5)
**a** Cross-validated performance of the gradient boosting algorithm.
Predictions of the trait alertness are plotted against the ground-truth
values. Each dot represents an individual’s predicted and actual trait
alertness. Trait alertness was calculated by averaging, for each individual,
all their alertness rating across the two weeks of the study. **b** Features
of the gradient boosting algorithm, ranked by order of descending importance.
Features importance was calculated using the SHAP method (see “Methods”). Each
dot on the plot is a Shapley value for a given predictor and participant.
Shapley values represent, for each participant, the exact contribution of a
given feature on the output of the model. The colour of the dots represents
the value of a given predictor from low to high (e.g. for age, younger = blue,
older = pink). Global feature importance is shown in the rightmost horizontal
grey bars and was calculated by averaging the absolute Shapley values of a
given predictor across all participants. **c** Trait correlation between
happiness and alertness. Happiness was the most important feature of the
model. Higher trait happiness is associated with higher trait alertness.
Source data are provided as a Source Data file.
[ Full size image ](/articles/s41467-022-34503-2/figures/5)
The importance ranking of the predictors revealed four key top factors that
had the strongest impact on the outcome of the model: (1) mood, (2) age, (3)
sleep, and (4) eating frequency (Fig. [ 5b
](/articles/s41467-022-34503-2#Fig5) ).
Mood, specifically levels of daily happiness, together with the age of the
individual, were the two most significant predictors of trait alertness (Fig.
[ 5b, c ](/articles/s41467-022-34503-2#Fig5) ), such that higher levels of
happiness and increasing chronological age were each positively predictive of
higher inherent levels of alertness (r = 0.67, p < 0.001 and r = 0.345, p <
0.001, respectively; Supplementary Figs. [ 3
](/articles/s41467-022-34503-2#MOESM1) and [ 4
](/articles/s41467-022-34503-2#MOESM1) , with p-values adjusted for multiple
comparisons using the Holm-Bonferroni method).
Interestingly, the former associations between happiness and alertness would
predict that participants who suffer from mood disorders have lower levels of
alertness. Post-hoc analyses afforded a test of this prediction, demonstrating
that this was indeed the case. Specifically, participants with a
current/former medical diagnosis of depression and/or anxiety disorder had
significantly lower baseline alertness compared to the remaining participants
(Welch T-test, depression: T(132)=2.855, p = 0.005; anxiety disorder:
T(165)=3.24, p = 0.001; normal quantile plots (Q–Q plots) were used to check
the assumption of normality; Supplementary Fig. [ 5
](/articles/s41467-022-34503-2#MOESM1) ).
The third key feature predicting baseline alertness was an individual’s
quality of sleep. As assessed with the validated Pittsburgh Sleep Quality
Index (PSQI), the higher an individual’s nightly sleep quality (reflected in
lower PSQI scores), the higher their level of trait alertness (r = −0.14, p =
0.002, Fig. [ 5b ](/articles/s41467-022-34503-2#Fig5) ).
The fourth top feature of predictive importance was the frequency of food
intake. Here, the higher the self-reported frequency of eating (1–2 times vs.
3–4 times vs. 5+ occasions), the lower the levels of trait alertness. Indeed,
participants who reported eating on average 5 times or more a day had a
significantly lower baseline alertness than those who reported eating 3–4
times a day (T(731)=2.94, p = 0.01, adjusted for multiple comparisons using
Tukey’s method; Q-Q plots were used to check the assumption of normality, and
the Levene test was used to verify the assumption of homoscedasticity) or 1–2
times a day (T(731)=2.65, p = 0.02).
### The genetic heritability of alertness
The above findings describe a set of non-genetic, and thus modifiable
lifestyle factors (with the exception of age) that are significantly
associated with alertness, both in terms of day-to-day fluctuations (i.e.,
daily state changes) and trait level of alertness.
However, together, these factors still leave a proportion of the variance in
alertness unexplained, suggesting a role for the non-modifiable factor of an
individual’s genetics to prove influential. To test this last hypothesis, a
standard twin model [ 12 ](/articles/s41467-022-34503-2#ref-CR12 "Polderman,
T. J. C. et al. Meta-analysis of the heritability of human traits based on
fifty years of twin studies. Nat. Genet. 47, 702–709 \(2015\).") was used to
test the extent of the contribution from an individual’s genetics in
predicting their levels of alertness.
Broad-sense heritability was calculated using the prototypical variance
component twin model approach. In short, (but see “Methods” for details), this
approach decomposes the phenotypic variance into a mixture of additive and
non-additive genetic factors (A and D, respectively), shared environmental
factors (C), and individual-specific environmental variance plus measurement
error (E). All models reported thereafter were then adjusted for age and sex.
Focusing first on the baseline levels of across the entire day alertness
described in the previous section, an ACE model yielded a heritability
estimate of 0.25 (95% confidence intervals [CIs] = −0.34 to 0.84), indicating
a modest and non-significant contribution of genetic factors to phenotypic
differences in alertness across individuals (Table [ 3
](/articles/s41467-022-34503-2#Tab3) ). Consistent with our prior hypotheses
and earlier results, the majority of variability in basal alertness was
explained by individual-specific environmental factors (E, 57%). Model-
comparison analyses revealed that trait alertness is significantly influenced
by familial (i.e., A + C) factors, but not by genetic factors alone. That is,
the A and C parameters could be removed individually without significantly
worsening the model fit (p = 0.384 and p = 0.532), but dropping both
parameters resulted in a worse model fit (p < 0.001).
**Table 3 The heritability of alertness and sleep**
[ Full size table ](/articles/s41467-022-34503-2/tables/3)
Second, the same analysis was conducted for trait alertness in the morning
hours following breakfast consumption, rather than across the entire day. This
model yielded a heritability estimate of 0.39 (best-fitting model = AE; Table
[ 3 ](/articles/s41467-022-34503-2#Tab3) ), suggesting that alertness levels
in the first hours after waking up are somewhat more influenced by genetics
than alertness during the rest of the day. Once again, however, the
heritability estimate were not significant (i.e., overlap with zero, −0.26 to
1.03), and individual-specific environmental factors instead explained most of
the variability in trait morning alertness (59%).
Finally, and to facilitate better interpretation of the above effects still
through the lens of genetics, heritability estimates were then calculated for
a subset of predictors including self-report happiness and sleep parameters
(Table [ 3 ](/articles/s41467-022-34503-2#Tab3) ). The heritability estimates
of happiness and sleep offset were roughly in the same range as alertness (
\\({h}^{2}\\) = .32 and \\({h}^{2}\\) = .18 respectively). By contrast,
other sleep parameters such as sleep duration, sleep efficiency, and sleep
onset had higher heritability estimates ( \\({h}^{2}\\) ≥ 0.51), indicating
a stronger contribution of genetic factors for these traits.
Overall, the heritability analyses demonstrate that an individual’s level of
alertness is not strongly dependent on genetics, and reinforce the former
findings that non-genetic (and thus modifiable) extrinsic factors more
strongly predict differences in alertness across this population.
## Discussion
Why is it that we human beings fluctuate in our alertness from one day to the
next? Why do we wake one morning feeling alert, yet another morning, flounder
in that level of alertness upon awakening? The first set of analyses revealed
that three key factors significantly and each independently predict how an
individual awakens with alertness: 1) their prior sleep history, 2) their
levels of prior physical activity, and 3) what they first eat in the morning.
. Importantly, from an interventional perspective, all three of these
categories are largely modifiable, and therefore represent lifestyle-realistic
opportunities, or interventional levers, that may aid how an individual (and
collectively, a society) awake each day, and sustain that waking alertness. We
now discuss each of these factors in detail, starting with sleep.
Nights when an individual slept longer than their own typical sleep amount
(rather than a standardised sleep amount), were associated with a superior
(i.e., higher) degree of next-morning alertness. More than just sleep
duration, however, where that sleep temporally arrived on the 24-hour clock
face was also significantly associated with next-morning alertness.
Specifically, sleeping later into the morning than is typical for a given
individual (which in part, can give rise to longer sleep duration), predicted
higher levels of alertness that following morning.
Such insights emphasise the utility of using a longitudinal study design,
which allows for a definition of a person’s own individualised sleep norms,
and deviations from that person-specific norm, both in sleep amount, and sleep
timing. This is not simply methodological in nature or value, but further
highlights the possibility that adopting a person-centric experimental
approach allows for true individualised recommendations targeting more
effective behavioural change to prevent failures in attentive alertness during
the waking (and working) day.
A next-step challenge will be to determine the underlying mechanism(s) of how
and why sleeping longer and sleeping later, relative to that unique
individual’s typical norm, may transact a benefit upon morning alertness.
We offer three tenable and testable candidates. First, sleeping later in the
morning means that an individual is more likely to wake up farther away from
their circadian nadir, shifting toward the circadian assent. Second, sleeping
later will increase the likelihood of obtaining more (or even awaken from) REM
sleep [ 5 ](/articles/s41467-022-34503-2#ref-CR5 "Hilditch, C. J. & McHill,
A. W. Sleep inertia: current insights. Nat. Sci. Sleep. 11, 155–165
\(2019\).") , [ 13 ](/articles/s41467-022-34503-2#ref-CR13 "Silva, E. J. &
Duffy, J. F. Sleep inertia varies with circadian phase and sleep stage in
older adults. Behav. Neurosci. 122, 928–935 \(2008\).") , [ 14
](/articles/s41467-022-34503-2#ref-CR14 "Skorucak, J., Arbon, E. L., Dijk,
D.-J. & Achermann, P. Response to chronic sleep restriction, extension, and
subsequent total sleep deprivation in humans: adaptation or preserved sleep
homeostasis? Sleep 41, \(2018\).") . Both this first and second possibilities
have been associated with reduced sleep inertia [ 5
](/articles/s41467-022-34503-2#ref-CR5 "Hilditch, C. J. & McHill, A. W. Sleep
inertia: current insights. Nat. Sci. Sleep. 11, 155–165 \(2019\).") , [ 13
](/articles/s41467-022-34503-2#ref-CR13 "Silva, E. J. & Duffy, J. F. Sleep
inertia varies with circadian phase and sleep stage in older adults. Behav.
Neurosci. 122, 928–935 \(2008\).") . The latter benefit upon REM sleep may be
beneficially driven by higher cortical activation, or the increase in body
(and brain) temperature associated with REM sleep [ 13
](/articles/s41467-022-34503-2#ref-CR13 "Silva, E. J. & Duffy, J. F. Sleep
inertia varies with circadian phase and sleep stage in older adults. Behav.
Neurosci. 122, 928–935 \(2008\).") , [ 15 ](/articles/s41467-022-34503-2#ref-
CR15 "Harding, E. C., Franks, N. P. & Wisden, W. Sleep and thermoregulation.
Curr. Opin. Physiol. 15, 7–13 \(2020\).") , [ 16
](/articles/s41467-022-34503-2#ref-CR16 "Wamsley, E. J., Hirota, Y., Tucker,
M. A., Smith, M. R. & Antrobus, J. S. Circadian and ultradian influences on
dreaming: a dual rhythm model. Brain Res. Bull. 71, 347–354 \(2007\).") .
A third (and non-mutually exclusive) explanation for the independent benefit
of sleeping longer concerns sleep pressure. Specifically, the effective
discharging of sleep pressure upon awakening. This dissipation, also known as
discharging of the sleep homeostat (in part, reflecting the clearance of
accumulated adenosine [ 17 ](/articles/s41467-022-34503-2#ref-CR17 "Porkka-
Heiskanen, T. & Kalinchuk, A. V. Adenosine, energy metabolism and sleep
homeostasis. Sleep. Med. Rev. 15, 123–135 \(2011\).") ), is one of the most
reliable predictors of diminished sleep inertia [ 4
](/articles/s41467-022-34503-2#ref-CR4 "Vallat, R., Meunier, D., Nicolas, A. &
Ruby, P. Hard to wake up? The cerebral correlates of sleep inertia assessed
using combined behavioral, EEG and fMRI measures. Neuroimage 184, 266–278
\(2019\).") , [ 5 ](/articles/s41467-022-34503-2#ref-CR5 "Hilditch, C. J. &
McHill, A. W. Sleep inertia: current insights. Nat. Sci. Sleep. 11, 155–165
\(2019\).") , [ 18 ](/articles/s41467-022-34503-2#ref-CR18 "Marzano, C.,
Ferrara, M., Moroni, F. & De Gennaro, L. Electroencephalographic sleep inertia
of the awakening brain. Neuroscience 176, 308–317 \(2011\).") . Indeed, long
sleepers have, on average, less remaining daytime sleep pressure than short
sleepers [ 19 ](/articles/s41467-022-34503-2#ref-CR19 "Aeschbach, D. et al.
Evidence from the waking electroencephalogram that short sleepers live under
higher homeostatic sleep pressure than long sleepers. Neuroscience 102,
493–502 \(2001\).") . Moreover, within an individual, extending sleep duration
leads to a decrease in sleep pressure [ 14
](/articles/s41467-022-34503-2#ref-CR14 "Skorucak, J., Arbon, E. L., Dijk,
D.-J. & Achermann, P. Response to chronic sleep restriction, extension, and
subsequent total sleep deprivation in humans: adaptation or preserved sleep
homeostasis? Sleep 41, \(2018\).") . In the context of our findings, sleeping
longer (relative to one’s own typical sleep amount) may allow a fuller
discharge of sleep pressure during the night, resulting in higher morning
alertness from one day to the next.
Beyond sleep, the second main feature predicting day-to-day changes in morning
alertness was the intensity of physical activity the day prior. In particular,
when the extent of an individual’s physical activity was comparatively greater
the day prior, individuals experienced higher levels of alertness the next
morning. Covariate analyses demonstrated that this effect was not driven by a
simple age-related difference in activity levels. Moreover, even though
physical activity can have modest benefits upon subsequent sleep [ 20
](/articles/s41467-022-34503-2#ref-CR20 "Kredlow, M. A., Capozzoli, M. C.,
Hearon, B. A., Calkins, A. W. & Otto, M. W. The effects of physical activity
on sleep: a meta-analytic review. J. Behav. Med. 38, 427–449 \(2015\).") , the
effects we report between prior physical activity and next-day alertness
remained significant when accounting for the amount of sleep that came in
between. This would suggest the influence of prior physical activity and that
of prior sleep upon alertness are each independent. However, a limitation of
our study is that we did not quantify the electrophysiological quality of
sleep, relevant considering that the effects of physical activity on
subsequent sleep can include changes in the NREM quality of sleep [ 21
](/articles/s41467-022-34503-2#ref-CR21 "Park, I. et al. Exercise improves the
quality of slow-wave sleep by increasing slow-wave stability. Sci. Rep. 11,
4410 \(2021\).") .
The final factor predicting fluctuations in next-day alertness occurred not on
the day prior, or even the night prior, but the morning of. Specifically, it
was the unique macronutrient composition of food that the individual consumed
for their breakfast that offered further explanatory value. Compared to a
reference breakfast meal consisting of a standardised (moderate) amount of
fat, carbohydrates, and protein (approximately 40/50/10% of energy,
respectively), when individuals consumed a higher amount of carbohydrates
(“High Carbs” breakfast), they experienced higher levels of ensuing alertness.
In contrast, the “High Protein” breakfast predicted a diminished, rather than
enhanced, level of alertness following sleep, relative to the reference meal.
Lastly, the consumption of a pure glucose liquid bolus (oral glucose tolerance
test, OGTT) was associated with a marked reduction in alertness levels−the
largest drop relative to all the other standardised breakfast meals.
Importantly, however, all of the above associations for the breakfast food
remained significant when postprandial blood glucose levels were adjusted for
in the statistical model. Similarly, the differences in alertness between the
protein and carbohydrate meals were not driven by differences in total energy
content of each respective meal, since most of the breakfast meals (including
the High Protein, High Carb and standardised reference) were calorically
matched (i.e., isocaloric at ~500 kcal). Instead, these findings suggest that
it is the actual macronutrient composition of the meal itself that contributes
to the statistically independent prediction of subsequent alertness.
That a carbohydrate-rich meal, versus a protein-rich meal, is associated with
a higher level of morning alertness may seem counter to previous reports that
have described either the opposite effect [ 22
](/articles/s41467-022-34503-2#ref-CR22 "Zeng, Y.-C. et al. Influences of
protein to energy ratios in breakfast on mood, alertness and attention in the
healthy undergraduate students. Health 3, 383 \(2011\).") , or no significant
difference relative to protein-rich meals [ 23
](/articles/s41467-022-34503-2#ref-CR23 "Boelsma, E., Brink, E. J., Stafleu,
A. & Hendriks, H. F. J. Measures of postprandial wellness after single intake
of two protein-carbohydrate meals. Appetite 54, 456–464 \(2010\).") . However,
there is evidence of high carbohydrate intake linked to superior alertness
levels, consistent with our findings [ 24 ](/articles/s41467-022-34503-2#ref-
CR24 "Sihvola, N. et al. Breakfast high in whey protein or carbohydrates
improves coping with workload in healthy subjects. Br. J. Nutr. 110, 1712–1721
\(2013\).") . In addition, increasing protein intake in drosophila models was
correlated with decreased postprandial alertness [ 25
](/articles/s41467-022-34503-2#ref-CR25 "Murphy, K. R. et al. Postprandial
sleep mechanics in Drosophila. Elife 5, e19334 \(2016\).") , suggesting that a
meal lower in protein may be optimal for increasing alertness.
Beyond the effects of the high carbohydrate meal itself, one of the strongest
effects revealed in the current study was the marked reduction in alertness on
days when participants consumed a high-sugar amount, here controlled using a
standardised liquid glucose bolus at breakfast (the oral glucose tolerance
test (OGTT)). The OGTT consists of 100% monosaccharide glucose. That a large
dose of glucose predicts a drop in alertness may appear contradictory to the
aforementioned association of the High Carb breakfast linked to an increase in
alertness. However, despite their common high carbohydrate content, there are
three key differences between the High Carb and OGTT meals that aid in
resolving this empirical tension.
First, unlike the OGTT, the High Carb breakfast a contained 23% proportion of
other macronutrients, of which 16% was fat and 7% was protein. The ratio of
carbohydrates-to-protein is known to modulate tryptophan availability in the
brain and thus serotonin synthesis [ 26 ](/articles/s41467-022-34503-2#ref-
CR26 "Wurtman, R. J. et al. Effects of normal meals rich in carbohydrates or
proteins on plasma tryptophan and tyrosine ratios. Am. J. Clin. Nutr. 77,
128–132 \(2003\).") . Specifically, pure carbohydrate (i.e., a ratio of 1:0,
as seen in the OGTT) may drive the strongest sedative effect through a surge
in serotonin synthesis and thus brain-available tryptophan associated with
impaired alertness [ 27 ](/articles/s41467-022-34503-2#ref-CR27 "Mantantzis,
K., Schlaghecken, F., Sünram-Lea, S. I. & Maylor, E. A. Sugar rush or sugar
crash? A meta-analysis of carbohydrate effects on mood. Neurosci. Biobehav.
Rev. 101, 45–67 \(2019\).") . Our finding would therefore suggest that the
combination of other macronutrients (e.g., protein, fat, fibre) paired with
carbohydrate determines the true synergistic impact of the meal on subsequent
alertness.
The second key distinction between the High Carb meal and OGTT meal is the
sugar form. The OGTT consists exclusively of glucose, whereas the High Carb
meal consists of sucrose, which is composed of one molecule of glucose, and
one molecule of fructose. Unlike glucose in the OGTT, the fructose represented
in the High Carb meal results in a more modest effect on blood glucose and
circulating insulin levels [ 28 ](/articles/s41467-022-34503-2#ref-CR28
"Merino, B., Fernández-Díaz, C. M., Cózar-Castellano, I. & Perdomo, G.
Intestinal Fructose and Glucose Metabolism in Health and Disease. Nutrients
12, \(2019\).") . Therefore, the High Carb meal can induce a lower glycemic
response than the OGTT [ 29 ](/articles/s41467-022-34503-2#ref-CR29
"Tsereteli, N. et al. Impact of insufficient sleep on dysregulated blood
glucose control under standardised meal conditions. Diabetologia
https://doi.org/10.1007/s00125-021-05608-y
\(2021\).") . This strong surge in blood glucose levels caused by the OGTT may therefore result in an inhibition of the wake-promoting neurons in the hypothalamus [ 30 ](/articles/s41467-022-34503-2#ref-CR30 "Burdakov, D. K+ channels stimulated by glucose: a new energy-sensing pathway. Pflug. Arch. 454, 19–27 \(2007\).") , and this loss of hypothalamic drive may ultimately lead to reduced alertness.
The third non-mutually exclusive explanation for why the pure glucose meal
significantly impaired alertness concerns the lower total caloric content that
the OGTT represented, relative to the other standardised meals in the study
(300 kcal vs ~500 kcal). Within this mechanistic framework, the caloric
deficit would result in post-consumption hunger [ 31
](/articles/s41467-022-34503-2#ref-CR31 "Wyatt, P. et al. Postprandial
glycaemic dips predict appetite and energy intake in healthy individuals. Nat.
Metab. 3, 523–529 \(2021\).") , the effects of which have been associated with
lower levels of general alertness [ 32 ](/articles/s41467-022-34503-2#ref-
CR32 "Holt, S. H., Delargy, H. J., Lawton, C. L. & Blundell, J. E. The effects
of high-carbohydrate vs high-fat breakfasts on feelings of fullness and
alertness, and subsequent food intake. Int. J. Food Sci. Nutr. 50, 13–28
\(1999\).") .
The macronutrient composition of a meal can significantly dictate the
subsequent change in blood glucose [ 9 ](/articles/s41467-022-34503-2#ref-CR9
"Berry, S. E. et al. Human postprandial responses to food and potential for
precision nutrition. Nat. Med.
https://doi.org/10.1038/s41591-020-0934-0
\(2020\).") . However, different individuals respond with markedly different blood glucose responses to the same macronutrient meal due to a broad collection of reasons [ 9 ](/articles/s41467-022-34503-2#ref-CR9 "Berry, S. E. et al. Human postprandial responses to food and potential for precision nutrition. Nat. Med.
https://doi.org/10.1038/s41591-020-0934-0
\(2020\).") . This suggests that, independent of the nutritional content of the food an individual eats, there is a need to additionally measure, and account for, the inter-individual difference in blood glucose response to that meal.
Guided by this dissociation, the final analyses assessed associations between
alertness and the body’s glycemic food response Here, on days when an
individual experienced a higher blood glucose response to that same
standardised breakfast meal, the lower their levels of alertness that
following morning, and vice versa. Importantly, this effect was not explained
by breakfast macronutrient composition. Therefore, our findings establish that
it is both the macronutrient profile of food and the way in which the body
processes that food — here on the basis of glycemic blood glucose levels. We
show that both independently predict that relationship with morning alertness.
That a higher level of blood glucose is associated with a lower level of
alertness would, after first glance, contravene the popular cultural belief of
a “sugar rush” that boosts vigilance. However, contrary to this common
fiction, and consistent with the current findings, experimental evidence has
established that high glucose consumption results in a “sugar crash” and thus
reduced alertness. Specifically, attentive levels of consciousness and
associated cognition decrease soon after the ingestion of high-glycemic foods,
such as potatoes and sugar-sweetened beverages [ 27
](/articles/s41467-022-34503-2#ref-CR27 "Mantantzis, K., Schlaghecken, F.,
Sünram-Lea, S. I. & Maylor, E. A. Sugar rush or sugar crash? A meta-analysis
of carbohydrate effects on mood. Neurosci. Biobehav. Rev. 101, 45–67
\(2019\).") , [ 33 ](/articles/s41467-022-34503-2#ref-CR33 "Anderson, C. &
Horne, J. A. A high sugar content, low caffeine drink does not alleviate
sleepiness but may worsen it. Hum. Psychopharmacol. 21, 299–303 \(2006\).") .
Adding further support to the notion of a sugar crash demonstrated in the
current study, a recent meta-analysis reported that high subsequent glycemic
responses to morning breakfast meals are associated with impaired, rather than
enhanced, cognitive performance [ 34 ](/articles/s41467-022-34503-2#ref-CR34
"Edefonti, V. et al. The effect of breakfast composition and energy
contribution on cognitive and academic performance: a systematic review. Am.
J. Clin. Nutr. 100, 626–656 \(2014\).") .
How and why alertness levels drop significantly following a high-glycemic meal
has been linked to glucose-sensing neurons in the brain and may accurately
account for our related findings. Indeed, when glucose-sensing neurons are
activated by high(er) levels of blood sugar [ 30
](/articles/s41467-022-34503-2#ref-CR30 "Burdakov, D. K+ channels stimulated
by glucose: a new energy-sensing pathway. Pflug. Arch. 454, 19–27 \(2007\).")
, [ 35 ](/articles/s41467-022-34503-2#ref-CR35 "Afaghi, A., O’Connor, H. &
Chow, C. M. High-glycemic-index carbohydrate meals shorten sleep onset. Am. J.
Clin. Nutr. 85, 426–430 \(2007\).") , [ 36 ](/articles/s41467-022-34503-2#ref-
CR36 "Kim, S. W. & Lee, B. I. Metabolic state, neurohormones, and vagal
stimulation, not increased serotonin, orchestrate postprandial drowsiness.
Biosci. Hypotheses 2, 422–427 \(2009\).") , these sugar-sensing brain cells
inhibit the otherwise wake-promoting orexin system within the lateral
hypothalamus, the blockade of which results in decreased alertness [ 30
](/articles/s41467-022-34503-2#ref-CR30 "Burdakov, D. K+ channels stimulated
by glucose: a new energy-sensing pathway. Pflug. Arch. 454, 19–27 \(2007\).")
.
Beyond empirical and mechanisms insights, the current study may aid more
broadly in the development of behavioural recommendations at a public-health
or government level. Perhaps most applicable, the current results suggest that
avoiding high-glycemic-response breakfast is associated with optimal alertness
throughout the morning. This may be especially germane in the context of
education, where alertness is essential for effective knowledge acquisition in
the classroom. In fact, this issue is particularly noteworthy considering the
rapidly growing trend for teenagers and young adults to consume sugar-
sweetened energy drinks as an alternative to a whole-foods breakfast [ 37
](/articles/s41467-022-34503-2#ref-CR37 "Luneke, A. C. et al. Energy drink
expectancies among college students. J Am. Coll. Health 1–9
https://doi.org/10.1080/07448481.2020.1790569
\(2020\).") —a trend that would only serve to increase sleepiness in morning classes.
Findings from our first experimental question addressed the factors that
account for fluctuations in alertness within an individual, from one day to
the next. However, there was also large and substantial between-person
variability in average levels of alertness. Our second experimental question
sought to explain why this is the case i.e., why do some individuals have a
higher level of overall alertness “set-point” common across all days?
A prominent genetic influence would at first seem logical. However, twin-pair
analyses indicated that the genetic contribution in daytime alertness was
quantifiably small overall, with a heritability estimate of 0.25,
demonstrating that most of the trait variation in alertness comes from non-
genetically shared, individual-specific factors.
Our additional machine-learning analysis further revealed that it is non-
genetic factors which best account for differences in alertness set points
across individuals. Specifically, three independently contributing components
were revealed that most significantly predicted trait levels of daytime
alertness: (1) positive mood (specifically self-report happiness), (2) age,
and (3) an individual’s self-reported sleep quality.
The feature of positive mood, and specifically levels of happiness, was the
strongest predictor of general alertness across individuals, such that the
happier an individual was, the higher their baseline levels of alertness. Two
tenable, non-mutually exclusive, explanations may explain this association.
First, the psychological state of happy mood is accompanied by an aroused
autonomic state during wakefulness [ 38 ](/articles/s41467-022-34503-2#ref-
CR38 "Kreibig, S. D. Autonomic nervous system activity in emotion: a review.
Biol. Psychol. 84, 394–421 \(2010\).") . This increased autonomic activation
may therefore in turn promote cognitive arousal and alertness. Second, there
is a well-established bidirectional association between happiness and
serotonin [ 39 ](/articles/s41467-022-34503-2#ref-CR39 "Young, S. N. How to
increase serotonin in the human brain without drugs. J. Psychiatry Neurosci.
32, 394–399 \(2007\).") ; a neurotransmitter that has a key role in the
modulation of alertness [ 40 ](/articles/s41467-022-34503-2#ref-CR40 "Oken,
B. S., Salinsky, M. C. & Elsas, S. M. Vigilance, alertness, or sustained
attention: physiological basis and measurement. Clin. Neurophysiol. 117,
1885–1901 \(2006\).") . Higher positive mood may thus be associated with
higher levels of brain serotonin, and consequently higher activity in
wakefulness-promoting brain regions.
Interestingly, of the varied sleep measures assessed, it was subjective sleep
quality, rather than any objective sleep metrics, that ranked as a more
important predictor of an individual’s trait alertness. This association with
between-person differences in alertness adds to a rapidly emerging set of data
suggesting that sleep quality, as much if not more than, sleep quantity, most
accurately explains the variability in sleep-dependent brain and body
outcomes, including mental health and well-being [ 41
](/articles/s41467-022-34503-2#ref-CR41 "Jackowska, M., Ronaldson, A., Brown,
J. & Steptoe, A. Biological and psychological correlates of self-reported and
objective sleep measures. J. Psychosom. Res. 84, 52–55 \(2016\).") , [ 42
](/articles/s41467-022-34503-2#ref-CR42 "Ben Simon, E., Vallat, R., Barnes, C.
M. & Walker, M. P. Sleep loss and the socio-emotional brain. Trends Cogn. Sci.
24, 435–450 \(2020\).") , sustained attention [ 43
](/articles/s41467-022-34503-2#ref-CR43 "Gobin, C. M., Banks, J. B., Fins, A.
I. & Tartar, J. L. Poor sleep quality is associated with a negative cognitive
bias and decreased sustained attention. J. Sleep. Res. 24, 535–542 \(2015\).")
, cardiovascular health [ 44 ](/articles/s41467-022-34503-2#ref-CR44
"Hoevenaar-Blom, M. P., Spijkerman, A. M. W., Kromhout, D., van den Berg, J.
F. & Verschuren, W. M. M. Sleep duration and sleep quality in relation to
12-year cardiovascular disease incidence: the MORGEN study. Sleep 34,
1487–1492 \(2011\).") , and all-cause mortality [ 45
](/articles/s41467-022-34503-2#ref-CR45 "Martin, J. L. et al. Poor self-
reported sleep quality predicts mortality within one year of inpatient post-
acute rehabilitation among older adults. Sleep 34, 1715–1721 \(2011\).") .
The current findings must be appreciated within the context of a number of
important limitations. First, alertness was measured subjectively (visual
analogue scale), and should therefore be interpreted in light of the potential
biases associated with self-report methods. That said, subjective alertness
ratings correlate significantly with objective EEG spectral power activity
indexing homeostatic sleep pressure, or sleepiness 46 , 47 , [ 48
](/articles/s41467-022-34503-2#ref-CR48 "Finelli, L. A., Baumann, H., Borbély,
A. A. & Achermann, P. Dual electroencephalogram markers of human sleep
homeostasis: correlation between theta activity in waking and slow-wave
activity in sleep. Neuroscience 101, 523–529 \(2000\).") , suggesting some
level of objective validity. Second, though the population cohort assessed in
this study was composed of healthy individuals, we did not formally screen for
sleep-disordered breathing. Therefore, we cannot exclude the possibility that
a sub-group of participants existed that may have had sleep apnea, which could
have influenced the results. Third, no sleep logs were collected in this
study, with sleep/wake detection relying on accelerometry, which has been
validated against gold-standard polysomnography in both healthy individuals
and patients with sleep disorders (mean concordance statistic of 0.86 and
0.83, respectively) [ 49 ](/articles/s41467-022-34503-2#ref-CR49 "van Hees,
V. T. et al. Estimating sleep parameters using an accelerometer without sleep
diary. Sci. Rep. 8, 12975 \(2018\).") . Fourth, light exposure in the first
hours of the morning was not measured in this study. Bright light exposure has
been shown to increase the cortisol awakening response [ 50
](/articles/s41467-022-34503-2#ref-CR50 "Petrowski, K., Schmalbach, B.,
Niedling, M. & Stalder, T. The effects of post-awakening light exposure on the
cortisol awakening response in healthy male individuals.
Psychoneuroendocrinology 108, 28–34 \(2019\).") and increase daytime alertness
[ 51 ](/articles/s41467-022-34503-2#ref-CR51 "Souman, J. L., Tinga, A. M., Te
Pas, S. F., van Ee, R. & Vlaskamp, B. N. S. Acute alerting effects of light: A
systematic literature review. Behav. Brain Res. 337, 228–239 \(2018\).") , and
may thus have offered additional explanatory insights in predicting morning
alertness. Fifth, that the standardised breakfast meals were associated with
morning alertness differently based on their macronutrient composition should
not be used solely to derive conclusions on recommending absolute quantities
of a single macronutrient (e.g. the simplistic notion that consuming more
grams of carbohydrates will increase alertness). Indeed, every meal in the
study (with the exception of the OGTT) included some amount of each of the
major macronutrients, and the meals were therefore not completely separable in
their macronutrient composition. Last, despite the large sample size of our
study, one cannot exclude a potential selection bias. Specifically,
participants in PREDICT1 study were of robust health (see inclusion criteria),
and therefore may not be representative of a pure random sample of the US and
UK population.
In summary, we demonstrate that fluctuations in morning alertness within an
individual from one day to the next are significantly and independently
predicted by four modifiable factors of (1) sleep the night before, (2) higher
physical activity on the day prior, (3) a breakfast rich in carbohydrates, and
(4) a lower glycemic response in the hours following breakfast consumption.
Beyond an explanation of day-to-day fluctuations in alertness, and counter to
a strong trait determinant model, genetics offered a modest influence upon an
individual’s set-point of alertness. Instead, trait levels of alertness across
individuals were best predicted by their level of positive mood, their age,
and their self-reported sleep quality.
More broadly, our results reveal a set of key factors associated with
alertness that are, for the most part, not fixed. Instead, the majority of
factors associated with alertness are modifiable, and therefore permissive to
behavioural intervention. Such findings may help inform public health
recommendations towards reducing the non-trivial mortality, financial and
societal burden caused by insufficient alertness.
## Methods
### Study design and participants
We sought to test our hypothesis and predictions using the Personalized
Responses to Dietary Composition Trial (or “PREDICT1”). PREDICT1 is a single-
arm, single-blind intervention study, whose overall objective is to understand
glucose, insulin, lipid and other postprandial responses to foods based on the
individual’s characteristics, including molecular biomarkers, lifestyle
factors, in combination with the nutritional composition of the food. The
official start and end dates for the study were 5 June 2018 and 8 May 2019,
the first participant was enrolled on 4 August 2018 and the last clinical
visit was completed on 24 April 2019, with the primary cohort based at King’s
College London in the UK and a second cohort (that underwent the same
profiling as in the UK) assessed at Massachusetts General Hospital in Boston,
MA, USA. In the UK, participants (target enrollment, 1,000 participants) were
recruited from the TwinsUK cohort, a prospective cohort study, and online
advertising. In the USA, participants (target enrollment, 100 participants)
were recruited through online advertising and research participant databases.
The written informed consent and ethical committee approvals covered all
analysis reported in the current study in addition to the key primary outcomes
described in [ 9 ](/articles/s41467-022-34503-2#ref-CR9 "Berry, S. E. et al.
Human postprandial responses to food and potential for precision nutrition.
Nat. Med.
https://doi.org/10.1038/s41591-020-0934-0
\(2020\).") . The trial was registered on ClinicalTrials.gov (registration number: NCT03479866 _,_ first posted on March 27, 2018) as part of the registration for the PREDICT program of research, which also includes two other study protocol cohorts (not analysed in the current study). The trial was run in accordance with the Declaration of Helsinki and Good Clinical Practice. The study was approved in the UK by the Research Ethics Committee and Integrated Application System (IRAS 236407) and in the US by the Institutional Review Board of Partners Healthcare (Protocol # 2018P002078). Participants did not receive financial compensation for taking part in the study.
Study participants were healthy individuals aged 18–65 years, who were able to
provide written informed consent. Exclusion criteria included ongoing
inflammatory disease; cancer in the last three years (excluding skin cancer);
long-term gastrointestinal disorders including irritable bowel disease or
Celiac disease (gluten allergy), but not including irritable bowel syndrome;
taking immunosuppressants or antibiotics as daily medication within the last
three months; capillary glucose level of >12 mmol l–1 (or 216 mg dl–1), or
type 1 diabetes mellitus, or taking medication for type 2 diabetes mellitus;
currently experiencing acute clinically diagnosed depression; heart attack
(myocardial infarction) or stroke in the last 6 months; pregnancy; and vegan
or experiencing an eating disorder or unwilling to consume foods that are part
of the study. Diagnosis or symptoms of any sleep disorders, circadian rhythm
disorders, or neurocognitive disorders were not exclusionary. Similarly, the
use of medication to impact sleep, circadian or brain function was not
exclusionary.
A total of 970 generally healthy adults from the United Kingdom (including
non-twins, monozygotic [MZ] twins and dizygotic [DZ] twins) as well as 95
healthy adults from the United States (all non-twins) were enrolled and
completed baseline clinic measurements, as well as a two-week at-home phase.
For more details on the clinic measurements, we refer the reader to the online
protocol [ 8 ](/articles/s41467-022-34503-2#ref-CR8 "Berry, S. et al.
Personalised REsponses to DIetary Composition Trial \(PREDICT\): an
intervention study to determine inter-individual differences in postprandial
response to foods. Protocol Exchange
https://doi.org/10.21203/rs.2.20798/v1
\(2020\).") .
During the study’s home-phase, participants consumed multiple standardised
test meals differing in macronutrient composition (carbohydrate, fat, protein
and fibre), while wearing a physical activity monitor. Standardised meals were
consumed at breakfast during the first 9-11 days of the home period, and
additionally for lunch on the two first days. Participants recorded their
dietary intake and alertness on the Zoe study app throughout the study.
Following completion of the home-phase, participants returned all study
samples and devices to study staff via standard mail.
### Data collection and analysis
For an exhaustive description of all the outcomes measured in the PREDICT1
study, we refer the reader to the full online protocol [ 8
](/articles/s41467-022-34503-2#ref-CR8 "Berry, S. et al. Personalised
REsponses to DIetary Composition Trial \(PREDICT\): an intervention study to
determine inter-individual differences in postprandial response to foods.
Protocol Exchange
https://doi.org/10.21203/rs.2.20798/v1
\(2020\).") . Briefly, key outcomes included postprandial metabolic responses (blood triglyceride, glucose, and insulin concentrations) to sequential mixed-nutrient dietary challenges administered in a tightly controlled clinical setting on day 1. A second set of outcomes was assessed over the subsequent 13 d at-home period. Primary outcomes include gut microbiome profile, blood lipids and glucose, sleep, physical activity, and hunger and appetite assessment. The main analysis of the primary outcomes has been reported elsewhere [ 8 ](/articles/s41467-022-34503-2#ref-CR8 "Berry, S. et al. Personalised REsponses to DIetary Composition Trial \(PREDICT\): an intervention study to determine inter-individual differences in postprandial response to foods. Protocol Exchange
https://doi.org/10.21203/rs.2.20798/v1
\(2020\).") , [ 52 ](/articles/s41467-022-34503-2#ref-CR52 "Asnicar, F. et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med.
https://doi.org/10.1038/s41591-020-01183-8
\(2021\).") . The current study reports a non-preregistered/exploratory analysis of the association between the secondary outcome of subjective alertness and the primary outcomes of sleep, physical activity, diet, and blood glucose (all measured during the at-home phase of the study). Data from the UK and US sub-cohorts were combined into a single dataset in the current study.
#### Questionnaires
Data on socio-demographic characteristics, medical health, habitual diet, and
lifestyle were collected via a self-administered baseline questionnaire during
the clinic visit. _Education_ . Academic education was measured on a scale
from 0 (no qualifications) to 8 (postgraduate degree). _Subjective sleep_ .
Subjective sleep quality was assessed using the well-validated Pittsburgh
Sleep Quality Index (PSQI) [ 53 ](/articles/s41467-022-34503-2#ref-CR53
"Buysse, D. J., Reynolds, C. F. 3rd, Monk, T. H., Berman, S. R. & Kupfer, D.
J. The Pittsburgh Sleep Quality Index: a new instrument for psychiatric
practice and research. Psychiatry Res. 28, 193–213 \(1989\).") . The PSQI
measures 7 domains of sleep quality over the past month to provide a global
score (0-21) of overall sleep quality, with higher scores indicating poorer
sleep quality. In addition, participants were asked to report their typical
bedtime and waketime, in both weekdays and weekends. The absolute difference
between the midpoint of sleep in weekdays and weekends was then used to
quantify social jetlag [ 54 ](/articles/s41467-022-34503-2#ref-CR54
"Wittmann, M., Dinich, J., Merrow, M. & Roenneberg, T. Social jetlag:
misalignment of biological and social time. Chronobiol. Int. 23, 497–509
\(2006\).") , with higher values indicating a greater mismatch between an
individual’s own biological rhythm and the daily timing determined by social
constraints (assuming that most individuals worked during the weekdays and did
not work during the weekend). _Habitual diet and eating behaviours_ . Habitual
diet was measured using the European Prospective Investigation into Cancer and
Nutrition (EPIC) food frequency questionnaire (FFQ) in the UK cohort and the
Harvard semi-quantitative FFQ in the US cohort. The questionnaire also
included questions related to eating frequency (“how many times do you eat in
a day?”), habitual coffee and alcohol consumption, and whether the
participants usually skip breakfast. _Exercise_ . Self-report exercise
frequency was measured with the following question: “In the past year, how
frequently have you typically engaged in physical exercises that raise your
heart rate and last for 20 minutes at a time?” _Mental health_ .
Current/former clinical diagnosis of depression and anxiety disorder was
measured using the following questions: _“Has a doctor ever told you that you
have/had any of the following conditions? [clinical depression, anxiety or
stress disorder]”_ .
#### Standardised test meals
Upon completion of their baseline visit, participants received a home-phase
meal pack containing test-meals varying in macronutrient composition, which
they consumed according to standardised instructions for breakfast and, on
some days, lunch. Test meals consisted of either an oral glucose tolerance
test (OGTT) or muffins, which were consumed on their own or paired with
chocolate milk, a protein shake or commercial fibre bars. The description and
nutritional composition of the test meals can be found in Table [ 1
](/articles/s41467-022-34503-2#Tab1) . Test meals were consumed in a different
order depending on which protocol group participants were assigned to, as
described in the online protocol [ 8 ](/articles/s41467-022-34503-2#ref-CR8
"Berry, S. et al. Personalised REsponses to DIetary Composition Trial
\(PREDICT\): an intervention study to determine inter-individual differences
in postprandial response to foods. Protocol Exchange
https://doi.org/10.21203/rs.2.20798/v1
\(2020\).") . Test meals were prepared and packaged in the Dietetics Kitchen (Department of Nutritional Sciences, King’s College London). As is common practice for postprandial studies [ 55 ](/articles/s41467-022-34503-2#ref-CR55 "Kolovou, G. D. et al. Assessment and clinical relevance of non-fasting and postprandial triglycerides: an expert panel statement. Curr. Vasc. Pharmacol. 9, 258–270 \(2011\).") , meal sizes were similar across all participants and not normalised by weight or total daily energy expenditure.
Participants were instructed to fast for a minimum of 8 hours prior to
consuming a test breakfast meal (i.e., avoid nighttime snacking), and to fast
for 3 or 4 hours after the test meal consumption. Furthermore, they were
advised to limit exercise and drink only plain, still water during the fasting
periods. When fasting was completed, participants could eat, drink and
exercise as they liked for the rest of the day. Participants were asked to
consume all muffin-based meals within 10 minutes and the OGTT within 5
minutes, and to notify study staff if this was not achieved, in which case the
data were excluded from the analysis.
If the participant chose to accompany their home-phase muffin-based test meals
with a tea or coffee (with up to 40 ml of 0.1% fat cow’s milk, but without any
sugar or sweeteners), they were instructed to consume this drink consistently,
in the same strength and amount, alongside all muffin-based test meals
throughout the study. Participants were instructed to not consume any food or
drink other than water alongside the OGTT. They recorded test meals and any
other dietary intake within fasting periods, including accompanying drinks in
the study app (see next section) with the exact time of consumption and
ingredient quantities so that study staff could monitor compliance. Only test
meals that were completed according to instructions were included in the
analysis.
#### Food and alertness logging via mobile study app
The Zoe study app was developed to support the PREDICT 1 study by serving as
an electronic notebook for study tasks. The app sent participants
notifications and reminders to complete tasks at certain time-points, such as
when their test lunch meals were due. Participants logged their full dietary
intake using the study app over the 14-day study period, including all
standardised test meals and free-living foods, beverages (including water) and
medications.
Data logged into the app was uploaded onto a digital dashboard in real time
and reviewed and assessed for logging accuracy and study guideline compliance
by study staff. The Zoe study app contained a database of generic and branded
food items with nutritional information sourced from generic data sources,
commercial food databases under licence, back-of-pack information from
commercial providers, and publicly available restaurant nutritional data. It
also allowed participants to photograph back of pack labels in cases where
this information was missing from the nutritional database, and where
possible, the photographed information was entered into the database by study
staff.
The app also prompted participants to report their alertness levels on a
visual analogue scale [ 56 ](/articles/s41467-022-34503-2#ref-CR56 "Wright,
K. P. Jr, Hull, J. T. & Czeisler, C. A. Relationship between alertness,
performance, and body temperature in humans. Am. J. Physiol. Regul. Integr.
Comp. Physiol. 283, R1370–7 \(2002\).") by displaying the question “how alert
are you?”. These app notifications appeared at t = 0 (time of logging) and
regular intervals (+0.5, +1.5, +2.5 hours) following the logging of a
breakfast, lunch or dinner meal. However, given the free-living conditions of
the study, participants did occasionally miss one or more ratings, resulting
in a variable number of alertness ratings per day per participant. The app
also prompted participants to report their happiness and anxiousness levels
once per day at ~9 PM local time. Here again, a visual analogue scale was used
with the following question: “How have you been feeling generally over the
whole day: How happy have you felt”?
For both the meals and alertness data, participants with less than 5 days of
valid data were removed (n = 56 [5.3%] and n = 42 [3.49%] participants
excluded, respectively). Second, participants that travelled in a different
timezone during the two weeks of the home-based study were also excluded (n =
65 [5.59%] participants). Data regarding travel to a different time zone in
the weeks before the study was not available.
#### Postprandial glucose
Interstitial glucose was measured every 15 min using Freestyle Libre Pro CGMs
(Abbott). Monitors were fitted by trained nurses on the upper, non-dominant
arm at participants’ baseline visit and were covered with Opsite Flexifix
adhesive film (Smith & Nephew Medical) for improved durability, and were worn
for the entire study duration. The 2 hours incremental area under the curve
(2hr-iAUC) was used for analysis of the postprandial glucose response [ 9
](/articles/s41467-022-34503-2#ref-CR9 "Berry, S. E. et al. Human postprandial
responses to food and potential for precision nutrition. Nat. Med.
https://doi.org/10.1038/s41591-020-0934-0
\(2020\).") . The distribution of glucose 2hr-iAUC was skewed and the data was thus transformed using a Yeo-Johnson power transformation.
#### Sleep/wake
Sleep/wake patterns were measured using a triaxial accelerometer (AX3,
Axivity, Newcastle Upon Tyne, UK). The accelerometer was fitted by clinical
practitioners at the baseline clinic visit on the non-dominant wrist and worn
for the duration of the study (except during water-based activities, including
showers and swimming), after which they were removed on day 15 and mailed back
to study staff. The accelerometer was programmed to measure acceleration at 50
Hz with a dynamic range of ±8 _g_ (where _g_ refers to the standard
acceleration of gravity, i.e., approximately 9.81 m/s 2 ). Non-wear periods
were defined as windows of at least 1 hour with less than 13 m _g_ for at
least 2 out of 3 axes, or where 2 out of 3 axes measured less than 50 m _g_ [
57 ](/articles/s41467-022-34503-2#ref-CR57 "Syed, S., Morseth, B., Hopstock,
L. A. & Horsch, A. Evaluating the performance of raw and epoch non-wear
algorithms using multiple accelerometers and electrocardiogram recordings.
Sci. Rep. 10, 5866 \(2020\).") .
Raw accelerometer data was analyzed using the “GGIR” R package version 1.10-7
[ 58 ](/articles/s41467-022-34503-2#ref-CR58 "van Hees, V. et al. GGIR.
https://doi.org/10.5281/zenodo.3474227
\(2019\).") . Sleep/wake detection was then quantified using the validated algorithm described in ref. [ 49 ](/articles/s41467-022-34503-2#ref-CR49 "van Hees, V. T. et al. Estimating sleep parameters using an accelerometer without sleep diary. Sci. Rep. 8, 12975 \(2018\).") , which uses the variance in the accelerometer z-axis angle together with a set of heuristic rules to determine sleep periods. This algorithm does not require a sleep diary and has been validated against gold-standard polysomnography in both healthy individuals and patients with sleep disorders, with a mean concordance statistic of 0.86 and 0.83, respectively [ 49 ](/articles/s41467-022-34503-2#ref-CR49 "van Hees, V. T. et al. Estimating sleep parameters using an accelerometer without sleep diary. Sci. Rep. 8, 12975 \(2018\).") . For each night and each participant, the following sleep metrics were calculated (see Fig. [ 1 ](/articles/s41467-022-34503-2#Fig1) ): sleep onset, sleep midpoint, sleep offset, sleep duration (defined as the elapsed time from sleep onset through sleep offset, or sleep period time [SPT]), wake after sleep onset (WASO), total sleep time (TST; = SPT - WASO), sleep efficiency (SE, = TST / SPT). SE was calculated using the SPT and not the more common total time in bed as denominator because the absence of sleep diary data precludes the accurate estimation of bedtime prior to sleep. For the same reason, the algorithm is unable to characterise sleep onset latency (the time between going to bed and falling asleep). The GGIR algorithm is not able to detect naps and therefore only nighttime sleep parameters were included in subsequent analyses.
A set of thresholds was then applied to remove invalid nights or participants,
consistent with typical practices [ 59 ](/articles/s41467-022-34503-2#ref-
CR59 "Jones, S. E. et al. Genetic studies of accelerometer-based sleep
measures yield new insights into human sleep behaviour. Nat. Commun. 10, 1585
\(2019\).") . First, any nights with a TST outside the range of 2 to 15 hours,
or a SE below 20%, was excluded (376 nights, 2.5%). Second, nights with more
than 10% classified as invalid were excluded (403 nights, 2.65%). Third,
nights with a sleep onset between 8 AM and 5 PM or a sleep offset after 12 PM
were excluded (45 nights, 0.3%). Finally, participants with less than 5 days
of valid sleep data (n = 60, 5.53%) were removed, consistent with the
preprocessing of the food and alertness data.
#### Physical activity
Physical activity was measured using the accelerometer and features were
calculated, for each day and each participant, using the GGIR software.
Specifically, these features consisted of the M10 and L5 values, and their
associated onset timings. M10 and L5 refer to the most-active 10 hours and the
least-active five hours of each day, respectively, and are commonly studied
measures relating to circadian activity [ 10
](/articles/s41467-022-34503-2#ref-CR10 "Gonçalves, B. S. B., Adamowicz, T.,
Louzada, F. M., Moreno, C. R. & Araujo, J. F. A fresh look at the use of
nonparametric analysis in actimetry. Sleep. Med. Rev. 20, 84–91 \(2015\).") ,
[ 59 ](/articles/s41467-022-34503-2#ref-CR59 "Jones, S. E. et al. Genetic
studies of accelerometer-based sleep measures yield new insights into human
sleep behaviour. Nat. Commun. 10, 1585 \(2019\).") . The M10 was defined as
the 10-h period with the maximum average acceleration, estimated using a 10
hours moving average. The L5 was defined as the 5-h period with the minimum
average acceleration, estimated with a 5-hours moving average. For these two
metrics, the onset timings was also calculated, defined as the number of hours
elapsed from the previous midnight. Once again, participants with less than 5
days of valid physical activity data were removed (n = 51, 4.72%).
#### Data concatenation
Sleep, meal (including postprandial glucose), physical activity and alertness
data were all merged into a single dataframe to facilitate statistical
analyses. Here, a strict inner-merge was performed, meaning that only the
participants and days with valid food, physical activity, alertness and sleep
data were included in the final dataframe. The sleep and physical activity
features were shifted by one day to ensure a valid temporal directionality,
i.e., that sleep/physical activity occurred before, and not after, the
alertness outcome.
A final quality check was applied that consisted of removing, for each day and
each participant, the alertness ratings that were logged one hour or more
before the algorithm’s predicted sleep offset (916 ratings, 1.0%). Concretely,
this removes the alertness ratings that were input before sleep and after
midnight (as these should technically be counted for the previous day).
### Statistical analyses
#### Multilevel modelling
Linear mixed-effects models were used to measure the statistical association
of sleep, food intake and its associated glucose response, and physical
activity with subsequent alertness. Unless specified otherwise, all multilevel
models were adjusted for age, sex, body mass index (BMI), twin status (MZ, DZ
or NT), sunrise time, daylight saving time (DST), and weekend (i.e., whether
the day on which the person wakes up is a Saturday or Sunday); with family
identifier and subject identifier defined as nested random effects.
Income, work schedule (including shift work) and household size were not
available in PREDICT1 and therefore the statistical models could not be
adjusted for those.
To account for the natural between-person variability in sleep, the sleep
predictors — duration, efficiency and waketime — were normalised using person-
mean centering. That is, they were expressed, for each participant separately,
as a deviation from this individual’s average calculated across the two weeks
of the study.
All multilevel analyses were performed in R [ 60
](/articles/s41467-022-34503-2#ref-CR60 "R Core Team. R: A language and
environment for statistical computing. R Foundation for Statistical Computing,
Vienna, Austria.
https://www.R-project.org/
\(2020\).") using the “lme4”, “lmerTest”,“sjPlot” and “emmeans” packages 61 , 62 , 63 , [ 64 ](/articles/s41467-022-34503-2#ref-CR64 "Lenth, R. V. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.5.3.
https://CRAN.R-project.org/package=emmeans
\(2020\).") . Goodness-of-fit was evaluated with the conditional \\({R}^{2}\\) [ 65 ](/articles/s41467-022-34503-2#ref-CR65 "Nakagawa, S. & Schielzeth, H. A general and simple method for obtaining R 2 from generalized linear mixed-effects models. Methods Ecol. Evol. 4, 133–142 \(2013\).") . For all multilevel models, the variance inflation factor (VIF) was used to check for multicollinearity. When multicollinearity was detected (VIF > 5), the correlated predictors were removed from the model and/or split into two separate multilevel models [ 66 ](/articles/s41467-022-34503-2#ref-CR66 "Yu, H., Jiang, S. & Land, K. C. Multicollinearity in hierarchical linear models. Soc. Sci. Res. 53, 118–136 \(2015\).") . Diagnostic plots were used to assess the validity of the fitted models. For each multilevel model, these included scatterplots of standardised residuals by fitted values and observed versus fitted values. Normal quantile plots (Q–Q plots) were used to check the assumption of normality of the residuals and random effects.
The performance of the multilevel model in predicting new, unseen data was
then tested using a hold-out validation. This step is increasingly recommended
to prevent overfitting and improve the interpretability of the findings [ 67
](/articles/s41467-022-34503-2#ref-CR67 "Yarkoni, T. & Westfall, J. Choosing
prediction over explanation in psychology: lessons from machine learning.
Perspect. Psychol. Sci. 12, 1100–1122 \(2017\).") . The dataset was separated
into a training and testing set, based on a split of odd and even days (e.g.
training = days 1, 3, 5, …; testing = days 2, 4, 6, …). Importantly, the
assignment of odd days to the training or testing set was randomly decided for
each participant. Then, a multilevel model was fitted on the training set,
using the same predictors and random effects as the main model. The resulting
regression coefficients were then used to make predictions on the testing set.
Performance was evaluated using the coefficient of determination (r-squared)
between the true and predicted alertness values. All statistical tests
reported in the manuscript are two-tailed.
#### Machine-learning analysis of the trait predictors of alertness
For between-person (i.e., trait) analysis of alertness, a machine-learning
approach was used to evaluate the relative importance of a large number of
trait measures on alertness. These predictors included: the age, sex,
education level, smoking status, BMI, average sleep and physical activity
parameters calculated across the two weeks of the study, self-report measures
of subjective sleep quality (PSQI score) and social jetlag, self-report
happiness, self-report habitual amount of exercise, self-report medical
diagnosis of depression or anxiety disorder, and self-report eating behaviours
— including whether or not the participant usually skip breakfast, habitual
coffee and alcohol consumption, eating frequency and snacking.
Several of these parameters were highly correlated and/or contained missing
values. For these two reasons, the association of these predictors with
alertness could not be evaluated using a standard regression approach, which
would have resulted in a dramatic decrease in sample size as well as invalid
regression coefficients because of multicollinearity. Addressing these issues,
a gradient boosting machine-learning algorithm (LightGBM, [ 11
](/articles/s41467-022-34503-2#ref-CR11 "Ke, G. et al. LightGBM: A Highly
Efficient Gradient Boosting Decision Tree. in Advances in Neural Information
Processing Systems 30 \(eds. Guyon, I. et al.\) 3146–3154 \(Curran Associates,
Inc., 2017\).") ) was used as the primary analytical method for the between-
person analysis. Gradient boosting algorithms are based on decision trees and
are therefore robust to multicollinearity in predictors. In addition, they
natively support missing values, without the need for deletion or imputation.
The LightGBM model was trained with 50 estimators and a random subsampling of
all features and samples (50%) before building each tree. Performed of the
model on unseen data was evaluated using a 3-fold cross-validation of the full
dataset.
Next, Shapley values were used to assess the unique contribution of each
feature in predicting trait alertness. Shapley values have several desirable
properties that make them ideal to evaluate the unbiased feature importance of
the predictors of a statistical model. Specifically, they quantify, for each
observation (i.e., participant), the exact impact of a given feature — after
accounting for all other features — on the outcome of the model. Shapley
values were first computed for each feature and each participant using the
SHAP library [ 68 ](/articles/s41467-022-34503-2#ref-CR68 "Lundberg, S. M. et
al. From local explanations to global understanding with explainable AI for
trees. Nat. Mach. Intell. 2, 56–67 \(2020\).") . Global feature importance was
then calculated by averaging the absolute Shapley values of a given predictor
across all observations.
All machine-learning algorithms were conducted in Python using the “scikit-
learn”, “lightgbm“, “shap” and “pingouin” packages [ 11
](/articles/s41467-022-34503-2#ref-CR11 "Ke, G. et al. LightGBM: A Highly
Efficient Gradient Boosting Decision Tree. in Advances in Neural Information
Processing Systems 30 \(eds. Guyon, I. et al.\) 3146–3154 \(Curran Associates,
Inc., 2017\).") , [ 68 ](/articles/s41467-022-34503-2#ref-CR68 "Lundberg, S.
M. et al. From local explanations to global understanding with explainable AI
for trees. Nat. Mach. Intell. 2, 56–67 \(2020\).") , [ 69
](/articles/s41467-022-34503-2#ref-CR69 "Pedregosa, F. et al. Scikit-learn:
machine learning in python. J. Mach. Learn. Res. 12, 2825–2830 \(2011\).") .
#### Heritability analyses
A large proportion of the main cohort consisted of pairs of identical (MZ) and
fraternal (DZ) twins, which allows a test of genetic influences upon alertness
and sleep inertia. For each dependent variable, we first calculated the intra-
pair correlation separately for MZ and DZ siblings. The former share the vast
majority of their germline DNA sequence [ 70
](/articles/s41467-022-34503-2#ref-CR70 "Jonsson, H. et al. Differences
between germline genomes of monozygotic twins. Nat. Genet. 53, 27–34
\(2021\).") , while the latter are assumed to share on average 50% of their
segregating genetic material. DZ twins are, however, presumed to share their
common environmental influences (e.g. family) to the same extent as MZ twins.
Therefore, the degree to which MZ siblings have a higher correlation for a
specific trait than DZ siblings reflects the extent of genetic influence on
this trait.
Heritability was then calculated using a standard twin model [ 71
](/articles/s41467-022-34503-2#ref-CR71 "Purcell, S. Variance components
models for gene–environment interaction in twin analysis. Twin Res. Hum.
Genet. 5, 554–571 \(2002\).") , which decomposes the observed phenotypic
variation into a combination of additive (A) and non-additive (D) genetic
variance, common environmental variance (C; familial influences that
contribute to twin similarity) and individual-specific environmental variance
plus measurement error (E). The combination of these factors that best matches
the observed data is found with structural equation modelling techniques.
Because the C and D factors are negatively confounded, they cannot be
estimated simultaneously. Therefore, following standard guidelines, an ACE
model was used when the DZ twin correlation was more than half the MZ twin
correlation, and an ADE model otherwise. The broad heritability (
\\({h}^{2}\\) ) was then defined as the percentage of total phenotypic
variance that could be explained by genetic factors (= A in ACE models and A +
D in ADE models).
The significance of genetic factors (A and/or D) was assessed by means of
likelihood ratio tests comparing the full model with a nested model in which
these factors were constrained to be zero. When the fit significantly
worsened, the contribution of genetic factors was considered significant.
Finally, the Akaike Information Criterion (AIC) was used to determine the
best-fitting model, with lower AIC indicating a better fit of the model to the
observed data.
All heritability analyses were conducted using the “mets” R package [ 72
](/articles/s41467-022-34503-2#ref-CR72 "Scheike, T. H., Holst, K. K. &
Hjelmborg, J. B. Estimating heritability for cause specific mortality based on
twin studies. Lifetime Data Anal. 20, 210–233 \(2014\).") . Twin models were
adjusted for age and sex. To account for repeated measurements in the twin
models, analyses focused on the participants’ grand-averaged values [ 73
](/articles/s41467-022-34503-2#ref-CR73 "Wang, C., Roy-Gagnon, M.-H.,
Lefebvre, J.-F., Burkett, K. M. & Dubois, L. Modeling gene-environment
interactions in longitudinal family studies: a comparison of methods and their
application to the association between the IGF pathway and childhood obesity.
BMC Med. Genet. 20, 9 \(2019\).") .
### Reporting summary
Further information on research design is available in the [ Nature Portfolio
Reporting Summary ](/articles/s41467-022-34503-2#MOESM2) linked to this
article.
## Data availability
The data of the at-home phase of the PREDICT1 trial, which supports the
findings of this study, are held by Zoe Ltd. These data were used under
license for the current study and are therefore not publicly available. Data
are however available from the authors upon reasonable request and with
permission of Zoe Ltd. The data of the baseline in-clinic visit of the
PREDICT1 trial are held by the department of Twin Research at King’s College
London. The data can be released to bona fide researchers using normal
procedures overseen by the Wellcome Trust and its guidelines as part of our
core funding. The application can be found at: [
https://twinsuk.ac.uk/resources-for-researchers/access-our-data/
](https://twinsuk.ac.uk/resources-for-researchers/access-our-data/) . Data
must be anonymized and conform to General Data Protection Regulation
standards. [ Source data ](/articles/s41467-022-34503-2#Sec23) are provided
with this paper.
## Code availability
All scripts used to run the analysis are available from the authors upon
request.
## References
1. Tefft, B. C. & Others. _Prevalence of motor vehicle crashes involving drowsy drivers, United States, 2009-2013_ . (Citeseer, 2014).
2. Rakel, R. E. Clinical and societal consequences of obstructive sleep apnea and excessive daytime sleepiness. _Postgrad. Med._ **121** , 86–95 (2009).
[ Article ](https://doi.org/10.3810%2Fpgm.2009.01.1957) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=19179816)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Clinical%20and%20societal%20consequences%20of%20obstructive%20sleep%20apnea%20and%20excessive%20daytime%20sleepiness&journal=Postgrad.%20Med.&doi=10.3810%2Fpgm.2009.01.1957&volume=121&pages=86-95&publication_year=2009&author=Rakel%2CRE)
3. Hafner, M., Stepanek, M., Taylor, J., Troxel, W. M. & van Stolk, C. Why sleep matters-the economic costs of insufficient sleep: a cross-country comparative analysis. _Rand Health Q_ **6** , 11 (2017).
[ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28983434)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5627640) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Why%20sleep%20matters-
the%20economic%20costs%20of%20insufficient%20sleep%3A%20a%20cross-
country%20comparative%20analysis&journal=Rand%20Health%20Q&volume=6&publication_year=2017&author=Hafner%2CM&author=Stepanek%2CM&author=Taylor%2CJ&author=Troxel%2CWM&author=Stolk%2CC)
4. Vallat, R., Meunier, D., Nicolas, A. & Ruby, P. Hard to wake up? The cerebral correlates of sleep inertia assessed using combined behavioral, EEG and fMRI measures. _Neuroimage_ **184** , 266–278 (2019).
[ Article ](https://doi.org/10.1016%2Fj.neuroimage.2018.09.033) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30223060)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Hard%20to%20wake%20up%3F%20The%20cerebral%20correlates%20of%20sleep%20inertia%20assessed%20using%20combined%20behavioral%2C%20EEG%20and%20fMRI%20measures&journal=Neuroimage&doi=10.1016%2Fj.neuroimage.2018.09.033&volume=184&pages=266-278&publication_year=2019&author=Vallat%2CR&author=Meunier%2CD&author=Nicolas%2CA&author=Ruby%2CP)
5. Hilditch, C. J. & McHill, A. W. Sleep inertia: current insights. _Nat. Sci. Sleep._ **11** , 155–165 (2019).
[ Article ](https://doi.org/10.2147%2FNSS.S188911) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31692489)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6710480) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20inertia%3A%20current%20insights&journal=Nat.%20Sci.%20Sleep.&doi=10.2147%2FNSS.S188911&volume=11&pages=155-165&publication_year=2019&author=Hilditch%2CCJ&author=McHill%2CAW)
6. Spaeth, A. M., Goel, N. & Dinges, D. F. Cumulative neurobehavioral and physiological effects of chronic caffeine intake: individual differences and implications for the use of caffeinated energy products. _Nutr. Rev._ **72** , 34–47 (2014).
[ Article ](https://doi.org/10.1111%2Fnure.12151) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25293542)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Cumulative%20neurobehavioral%20and%20physiological%20effects%20of%20chronic%20caffeine%20intake%3A%20individual%20differences%20and%20implications%20for%20the%20use%20of%20caffeinated%20energy%20products&journal=Nutr.%20Rev.&doi=10.1111%2Fnure.12151&volume=72&pages=34-47&publication_year=2014&author=Spaeth%2CAM&author=Goel%2CN&author=Dinges%2CDF)
7. Trotti, L. M. Waking up is the hardest thing I do all day: Sleep inertia and sleep drunkenness. _Sleep. Med. Rev._ **35** , 76–84 (2017).
[ Article ](https://doi.org/10.1016%2Fj.smrv.2016.08.005) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=27692973)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Waking%20up%20is%20the%20hardest%20thing%20I%20do%20all%20day%3A%20Sleep%20inertia%20and%20sleep%20drunkenness&journal=Sleep.%20Med.%20Rev.&doi=10.1016%2Fj.smrv.2016.08.005&volume=35&pages=76-84&publication_year=2017&author=Trotti%2CLM)
8. Berry, S. et al. Personalised REsponses to DIetary Composition Trial (PREDICT): an intervention study to determine inter-individual differences in postprandial response to foods. _Protocol Exchange_ [ https://doi.org/10.21203/rs.2.20798/v1 ](https://doi.org/10.21203/rs.2.20798/v1) (2020).
9. Berry, S. E. et al. Human postprandial responses to food and potential for precision nutrition. _Nat. Med_ . [ https://doi.org/10.1038/s41591-020-0934-0 ](https://doi.org/10.1038/s41591-020-0934-0) (2020).
10. Gonçalves, B. S. B., Adamowicz, T., Louzada, F. M., Moreno, C. R. & Araujo, J. F. A fresh look at the use of nonparametric analysis in actimetry. _Sleep. Med. Rev._ **20** , 84–91 (2015).
[ Article ](https://doi.org/10.1016%2Fj.smrv.2014.06.002) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25065908)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=A%20fresh%20look%20at%20the%20use%20of%20nonparametric%20analysis%20in%20actimetry&journal=Sleep.%20Med.%20Rev.&doi=10.1016%2Fj.smrv.2014.06.002&volume=20&pages=84-91&publication_year=2015&author=Gon%C3%A7alves%2CBSB&author=Adamowicz%2CT&author=Louzada%2CFM&author=Moreno%2CCR&author=Araujo%2CJF)
11. Ke, G. et al. LightGBM: A Highly Efficient Gradient Boosting Decision Tree. in _Advances in Neural Information Processing Systems 30_ (eds. Guyon, I. et al.) 3146–3154 (Curran Associates, Inc., 2017).
12. Polderman, T. J. C. et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. _Nat. Genet._ **47** , 702–709 (2015).
[ Article ](https://doi.org/10.1038%2Fng.3285) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25985137)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC2MXhtFemt7jN) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Meta-
analysis%20of%20the%20heritability%20of%20human%20traits%20based%20on%20fifty%20years%20of%20twin%20studies&journal=Nat.%20Genet.&doi=10.1038%2Fng.3285&volume=47&pages=702-709&publication_year=2015&author=Polderman%2CTJC)
13. Silva, E. J. & Duffy, J. F. Sleep inertia varies with circadian phase and sleep stage in older adults. _Behav. Neurosci._ **122** , 928–935 (2008).
[ Article ](https://doi.org/10.1037%2F0735-7044.122.4.928) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=18729646)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7673910) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20inertia%20varies%20with%20circadian%20phase%20and%20sleep%20stage%20in%20older%20adults&journal=Behav.%20Neurosci.&doi=10.1037%2F0735-7044.122.4.928&volume=122&pages=928-935&publication_year=2008&author=Silva%2CEJ&author=Duffy%2CJF)
14. Skorucak, J., Arbon, E. L., Dijk, D.-J. & Achermann, P. Response to chronic sleep restriction, extension, and subsequent total sleep deprivation in humans: adaptation or preserved sleep homeostasis? _Sleep_ 41, (2018).
15. Harding, E. C., Franks, N. P. & Wisden, W. Sleep and thermoregulation. _Curr. Opin. Physiol._ **15** , 7–13 (2020).
[ Article ](https://doi.org/10.1016%2Fj.cophys.2019.11.008) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32617439)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7323637) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20and%20thermoregulation&journal=Curr.%20Opin.%20Physiol.&doi=10.1016%2Fj.cophys.2019.11.008&volume=15&pages=7-13&publication_year=2020&author=Harding%2CEC&author=Franks%2CNP&author=Wisden%2CW)
16. Wamsley, E. J., Hirota, Y., Tucker, M. A., Smith, M. R. & Antrobus, J. S. Circadian and ultradian influences on dreaming: a dual rhythm model. _Brain Res. Bull._ **71** , 347–354 (2007).
[ Article ](https://doi.org/10.1016%2Fj.brainresbull.2006.09.021) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17208651)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Circadian%20and%20ultradian%20influences%20on%20dreaming%3A%20a%20dual%20rhythm%20model&journal=Brain%20Res.%20Bull.&doi=10.1016%2Fj.brainresbull.2006.09.021&volume=71&pages=347-354&publication_year=2007&author=Wamsley%2CEJ&author=Hirota%2CY&author=Tucker%2CMA&author=Smith%2CMR&author=Antrobus%2CJS)
17. Porkka-Heiskanen, T. & Kalinchuk, A. V. Adenosine, energy metabolism and sleep homeostasis. _Sleep. Med. Rev._ **15** , 123–135 (2011).
[ Article ](https://doi.org/10.1016%2Fj.smrv.2010.06.005) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=20970361)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Adenosine%2C%20energy%20metabolism%20and%20sleep%20homeostasis&journal=Sleep.%20Med.%20Rev.&doi=10.1016%2Fj.smrv.2010.06.005&volume=15&pages=123-135&publication_year=2011&author=Porkka-
Heiskanen%2CT&author=Kalinchuk%2CAV)
18. Marzano, C., Ferrara, M., Moroni, F. & De Gennaro, L. Electroencephalographic sleep inertia of the awakening brain. _Neuroscience_ **176** , 308–317 (2011).
[ Article ](https://doi.org/10.1016%2Fj.neuroscience.2010.12.014) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=21167917)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC3MXhvFKjsL4%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Electroencephalographic%20sleep%20inertia%20of%20the%20awakening%20brain&journal=Neuroscience&doi=10.1016%2Fj.neuroscience.2010.12.014&volume=176&pages=308-317&publication_year=2011&author=Marzano%2CC&author=Ferrara%2CM&author=Moroni%2CF&author=Gennaro%2CL)
19. Aeschbach, D. et al. Evidence from the waking electroencephalogram that short sleepers live under higher homeostatic sleep pressure than long sleepers. _Neuroscience_ **102** , 493–502 (2001).
[ Article ](https://doi.org/10.1016%2FS0306-4522%2800%2900518-2) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=11226688)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BD3MXhsVKmurY%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Evidence%20from%20the%20waking%20electroencephalogram%20that%20short%20sleepers%20live%20under%20higher%20homeostatic%20sleep%20pressure%20than%20long%20sleepers&journal=Neuroscience&doi=10.1016%2FS0306-4522%2800%2900518-2&volume=102&pages=493-502&publication_year=2001&author=Aeschbach%2CD)
20. Kredlow, M. A., Capozzoli, M. C., Hearon, B. A., Calkins, A. W. & Otto, M. W. The effects of physical activity on sleep: a meta-analytic review. _J. Behav. Med._ **38** , 427–449 (2015).
[ Article ](https://link.springer.com/doi/10.1007/s10865-015-9617-6) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25596964)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effects%20of%20physical%20activity%20on%20sleep%3A%20a%20meta-
analytic%20review&journal=J.%20Behav.%20Med.&doi=10.1007%2Fs10865-015-9617-6&volume=38&pages=427-449&publication_year=2015&author=Kredlow%2CMA&author=Capozzoli%2CMC&author=Hearon%2CBA&author=Calkins%2CAW&author=Otto%2CMW)
21. Park, I. et al. Exercise improves the quality of slow-wave sleep by increasing slow-wave stability. _Sci. Rep._ **11** , 4410 (2021).
[ Article ](https://doi.org/10.1038%2Fs41598-021-83817-6) [ ADS
](http://adsabs.harvard.edu/cgi-bin/nph-
data_query?link_type=ABSTRACT&bibcode=2021NatSR..11.4410P) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33627708)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7904822) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BB3MXltl2jt7k%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Exercise%20improves%20the%20quality%20of%20slow-
wave%20sleep%20by%20increasing%20slow-
wave%20stability&journal=Sci.%20Rep.&doi=10.1038%2Fs41598-021-83817-6&volume=11&publication_year=2021&author=Park%2CI)
22. Zeng, Y.-C. et al. Influences of protein to energy ratios in breakfast on mood, alertness and attention in the healthy undergraduate students. _Health_ **3** , 383 (2011).
[ Article ](https://doi.org/10.4236%2Fhealth.2011.36065) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BC3MXhsV2ku77J) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Influences%20of%20protein%20to%20energy%20ratios%20in%20breakfast%20on%20mood%2C%20alertness%20and%20attention%20in%20the%20healthy%20undergraduate%20students&journal=Health&doi=10.4236%2Fhealth.2011.36065&volume=3&publication_year=2011&author=Zeng%2CY-C)
23. Boelsma, E., Brink, E. J., Stafleu, A. & Hendriks, H. F. J. Measures of postprandial wellness after single intake of two protein-carbohydrate meals. _Appetite_ **54** , 456–464 (2010).
[ Article ](https://doi.org/10.1016%2Fj.appet.2009.12.014) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=20060863)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC3cXntlansr0%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Measures%20of%20postprandial%20wellness%20after%20single%20intake%20of%20two%20protein-
carbohydrate%20meals&journal=Appetite&doi=10.1016%2Fj.appet.2009.12.014&volume=54&pages=456-464&publication_year=2010&author=Boelsma%2CE&author=Brink%2CEJ&author=Stafleu%2CA&author=Hendriks%2CHFJ)
24. Sihvola, N. et al. Breakfast high in whey protein or carbohydrates improves coping with workload in healthy subjects. _Br. J. Nutr._ **110** , 1712–1721 (2013).
[ Article ](https://doi.org/10.1017%2FS0007114513000779) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=23591085)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC3sXhslKns77P) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Breakfast%20high%20in%20whey%20protein%20or%20carbohydrates%20improves%20coping%20with%20workload%20in%20healthy%20subjects&journal=Br.%20J.%20Nutr.&doi=10.1017%2FS0007114513000779&volume=110&pages=1712-1721&publication_year=2013&author=Sihvola%2CN)
25. Murphy, K. R. et al. Postprandial sleep mechanics in Drosophila. _Elife_ **5** , e19334 (2016).
26. Wurtman, R. J. et al. Effects of normal meals rich in carbohydrates or proteins on plasma tryptophan and tyrosine ratios. _Am. J. Clin. Nutr._ **77** , 128–132 (2003).
[ Article ](https://doi.org/10.1093%2Fajcn%2F77.1.128) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=12499331)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BD3sXhtFyntg%3D%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Effects%20of%20normal%20meals%20rich%20in%20carbohydrates%20or%20proteins%20on%20plasma%20tryptophan%20and%20tyrosine%20ratios&journal=Am.%20J.%20Clin.%20Nutr.&doi=10.1093%2Fajcn%2F77.1.128&volume=77&pages=128-132&publication_year=2003&author=Wurtman%2CRJ)
27. Mantantzis, K., Schlaghecken, F., Sünram-Lea, S. I. & Maylor, E. A. Sugar rush or sugar crash? A meta-analysis of carbohydrate effects on mood. _Neurosci. Biobehav. Rev._ **101** , 45–67 (2019).
[ Article ](https://doi.org/10.1016%2Fj.neubiorev.2019.03.016) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30951762)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC1MXntFCht7Y%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Sugar%20rush%20or%20sugar%20crash%3F%20A%20meta-
analysis%20of%20carbohydrate%20effects%20on%20mood&journal=Neurosci.%20Biobehav.%20Rev.&doi=10.1016%2Fj.neubiorev.2019.03.016&volume=101&pages=45-67&publication_year=2019&author=Mantantzis%2CK&author=Schlaghecken%2CF&author=S%C3%BCnram-
Lea%2CSI&author=Maylor%2CEA)
28. Merino, B., Fernández-Díaz, C. M., Cózar-Castellano, I. & Perdomo, G. Intestinal Fructose and Glucose Metabolism in Health and Disease. _Nutrients_ 12, (2019).
29. Tsereteli, N. et al. Impact of insufficient sleep on dysregulated blood glucose control under standardised meal conditions. _Diabetologia_ [ https://doi.org/10.1007/s00125-021-05608-y ](https://doi.org/10.1007/s00125-021-05608-y) (2021).
30. Burdakov, D. K+ channels stimulated by glucose: a new energy-sensing pathway. _Pflug. Arch._ **454** , 19–27 (2007).
[ Article ](https://link.springer.com/doi/10.1007/s00424-006-0189-8) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BD2sXitlGjsbw%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=K%2B%20channels%20stimulated%20by%20glucose%3A%20a%20new%20energy-
sensing%20pathway&journal=Pflug.%20Arch.&doi=10.1007%2Fs00424-006-0189-8&volume=454&pages=19-27&publication_year=2007&author=Burdakov%2CD)
31. Wyatt, P. et al. Postprandial glycaemic dips predict appetite and energy intake in healthy individuals. _Nat. Metab._ **3** , 523–529 (2021).
[ Article ](https://doi.org/10.1038%2Fs42255-021-00383-x) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33846643)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7610681) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BB3MXhtlyqsrzF) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Postprandial%20glycaemic%20dips%20predict%20appetite%20and%20energy%20intake%20in%20healthy%20individuals&journal=Nat.%20Metab.&doi=10.1038%2Fs42255-021-00383-x&volume=3&pages=523-529&publication_year=2021&author=Wyatt%2CP)
32. Holt, S. H., Delargy, H. J., Lawton, C. L. & Blundell, J. E. The effects of high-carbohydrate vs high-fat breakfasts on feelings of fullness and alertness, and subsequent food intake. _Int. J. Food Sci. Nutr._ **50** , 13–28 (1999).
[ Article ](https://doi.org/10.1080%2F096374899101382) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=10435117)
[ CAS ](/articles/cas-redirect/1:STN:280:DyaK1Mzmtl2ktw%3D%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effects%20of%20high-
carbohydrate%20vs%20high-
fat%20breakfasts%20on%20feelings%20of%20fullness%20and%20alertness%2C%20and%20subsequent%20food%20intake&journal=Int.%20J.%20Food%20Sci.%20Nutr.&doi=10.1080%2F096374899101382&volume=50&pages=13-28&publication_year=1999&author=Holt%2CSH&author=Delargy%2CHJ&author=Lawton%2CCL&author=Blundell%2CJE)
33. Anderson, C. & Horne, J. A. A high sugar content, low caffeine drink does not alleviate sleepiness but may worsen it. _Hum. Psychopharmacol._ **21** , 299–303 (2006).
[ Article ](https://doi.org/10.1002%2Fhup.769) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=16856218)
[ CAS ](/articles/cas-redirect/1:STN:280:DC%2BD28vksVSitw%3D%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=A%20high%20sugar%20content%2C%20low%20caffeine%20drink%20does%20not%20alleviate%20sleepiness%20but%20may%20worsen%20it&journal=Hum.%20Psychopharmacol.&doi=10.1002%2Fhup.769&volume=21&pages=299-303&publication_year=2006&author=Anderson%2CC&author=Horne%2CJA)
34. Edefonti, V. et al. The effect of breakfast composition and energy contribution on cognitive and academic performance: a systematic review. _Am. J. Clin. Nutr._ **100** , 626–656 (2014).
[ Article ](https://doi.org/10.3945%2Fajcn.114.083683) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=24808492)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC2cXht1ygs7nJ) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effect%20of%20breakfast%20composition%20and%20energy%20contribution%20on%20cognitive%20and%20academic%20performance%3A%20a%20systematic%20review&journal=Am.%20J.%20Clin.%20Nutr.&doi=10.3945%2Fajcn.114.083683&volume=100&pages=626-656&publication_year=2014&author=Edefonti%2CV)
35. Afaghi, A., O’Connor, H. & Chow, C. M. High-glycemic-index carbohydrate meals shorten sleep onset. _Am. J. Clin. Nutr._ **85** , 426–430 (2007).
[ Article ](https://doi.org/10.1093%2Fajcn%2F85.2.426) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17284739)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BD2sXhvFWhu7g%3D) [ Google
Scholar ](http://scholar.google.com/scholar_lookup?&title=High-glycemic-
index%20carbohydrate%20meals%20shorten%20sleep%20onset&journal=Am.%20J.%20Clin.%20Nutr.&doi=10.1093%2Fajcn%2F85.2.426&volume=85&pages=426-430&publication_year=2007&author=Afaghi%2CA&author=O%E2%80%99Connor%2CH&author=Chow%2CCM)
36. Kim, S. W. & Lee, B. I. Metabolic state, neurohormones, and vagal stimulation, not increased serotonin, orchestrate postprandial drowsiness. _Biosci. Hypotheses_ **2** , 422–427 (2009).
[ Article ](https://doi.org/10.1016%2Fj.bihy.2009.07.008) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BD1MXhsFOmsbzI) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Metabolic%20state%2C%20neurohormones%2C%20and%20vagal%20stimulation%2C%20not%20increased%20serotonin%2C%20orchestrate%20postprandial%20drowsiness&journal=Biosci.%20Hypotheses&doi=10.1016%2Fj.bihy.2009.07.008&volume=2&pages=422-427&publication_year=2009&author=Kim%2CSW&author=Lee%2CBI)
37. Luneke, A. C. et al. Energy drink expectancies among college students. _J Am. Coll. Health_ 1–9 [ https://doi.org/10.1080/07448481.2020.1790569 ](https://doi.org/10.1080/07448481.2020.1790569) (2020).
38. Kreibig, S. D. Autonomic nervous system activity in emotion: a review. _Biol. Psychol._ **84** , 394–421 (2010).
[ Article ](https://doi.org/10.1016%2Fj.biopsycho.2010.03.010) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=20371374)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Autonomic%20nervous%20system%20activity%20in%20emotion%3A%20a%20review&journal=Biol.%20Psychol.&doi=10.1016%2Fj.biopsycho.2010.03.010&volume=84&pages=394-421&publication_year=2010&author=Kreibig%2CSD)
39. Young, S. N. How to increase serotonin in the human brain without drugs. _J. Psychiatry Neurosci._ **32** , 394–399 (2007).
[ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=18043762)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2077351) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=How%20to%20increase%20serotonin%20in%20the%20human%20brain%20without%20drugs&journal=J.%20Psychiatry%20Neurosci.&volume=32&pages=394-399&publication_year=2007&author=Young%2CSN)
40. Oken, B. S., Salinsky, M. C. & Elsas, S. M. Vigilance, alertness, or sustained attention: physiological basis and measurement. _Clin. Neurophysiol._ **117** , 1885–1901 (2006).
[ Article ](https://doi.org/10.1016%2Fj.clinph.2006.01.017) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=16581292)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2865224) [ CAS
](/articles/cas-redirect/1:STN:280:DC%2BD28nksVCrtw%3D%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Vigilance%2C%20alertness%2C%20or%20sustained%20attention%3A%20physiological%20basis%20and%20measurement&journal=Clin.%20Neurophysiol.&doi=10.1016%2Fj.clinph.2006.01.017&volume=117&pages=1885-1901&publication_year=2006&author=Oken%2CBS&author=Salinsky%2CMC&author=Elsas%2CSM)
41. Jackowska, M., Ronaldson, A., Brown, J. & Steptoe, A. Biological and psychological correlates of self-reported and objective sleep measures. _J. Psychosom. Res._ **84** , 52–55 (2016).
[ Article ](https://doi.org/10.1016%2Fj.jpsychores.2016.03.017) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=27095159)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Biological%20and%20psychological%20correlates%20of%20self-
reported%20and%20objective%20sleep%20measures&journal=J.%20Psychosom.%20Res.&doi=10.1016%2Fj.jpsychores.2016.03.017&volume=84&pages=52-55&publication_year=2016&author=Jackowska%2CM&author=Ronaldson%2CA&author=Brown%2CJ&author=Steptoe%2CA)
42. Ben Simon, E., Vallat, R., Barnes, C. M. & Walker, M. P. Sleep loss and the socio-emotional brain. _Trends Cogn. Sci_ . **24** , 435–450 (2020).
43. Gobin, C. M., Banks, J. B., Fins, A. I. & Tartar, J. L. Poor sleep quality is associated with a negative cognitive bias and decreased sustained attention. _J. Sleep. Res._ **24** , 535–542 (2015).
[ Article ](https://doi.org/10.1111%2Fjsr.12302) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25913483)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Poor%20sleep%20quality%20is%20associated%20with%20a%20negative%20cognitive%20bias%20and%20decreased%20sustained%20attention&journal=J.%20Sleep.%20Res.&doi=10.1111%2Fjsr.12302&volume=24&pages=535-542&publication_year=2015&author=Gobin%2CCM&author=Banks%2CJB&author=Fins%2CAI&author=Tartar%2CJL)
44. Hoevenaar-Blom, M. P., Spijkerman, A. M. W., Kromhout, D., van den Berg, J. F. & Verschuren, W. M. M. Sleep duration and sleep quality in relation to 12-year cardiovascular disease incidence: the MORGEN study. _Sleep_ **34** , 1487–1492 (2011).
[ Article ](https://doi.org/10.5665%2Fsleep.1382) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=22043119)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3198203) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Sleep%20duration%20and%20sleep%20quality%20in%20relation%20to%2012-year%20cardiovascular%20disease%20incidence%3A%20the%20MORGEN%20study&journal=Sleep&doi=10.5665%2Fsleep.1382&volume=34&pages=1487-1492&publication_year=2011&author=Hoevenaar-
Blom%2CMP&author=Spijkerman%2CAMW&author=Kromhout%2CD&author=Berg%2CJF&author=Verschuren%2CWMM)
45. Martin, J. L. et al. Poor self-reported sleep quality predicts mortality within one year of inpatient post-acute rehabilitation among older adults. _Sleep_ **34** , 1715–1721 (2011).
[ Article ](https://doi.org/10.5665%2Fsleep.1444) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=22131610)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3208850) [
Google Scholar ](http://scholar.google.com/scholar_lookup?&title=Poor%20self-
reported%20sleep%20quality%20predicts%20mortality%20within%20one%20year%20of%20inpatient%20post-
acute%20rehabilitation%20among%20older%20adults&journal=Sleep&doi=10.5665%2Fsleep.1444&volume=34&pages=1715-1721&publication_year=2011&author=Martin%2CJL)
46. Drapeau, C. & Carrier, J. Fluctuation of waking electroencephalogram and subjective alertness during a 25-hour sleep-deprivation episode in young and middle-aged subjects. _Sleep_ **27** , 55–60 (2004).
[ Article ](https://doi.org/10.1093%2Fsleep%2F27.1.55) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=14998238)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Fluctuation%20of%20waking%20electroencephalogram%20and%20subjective%20alertness%20during%20a%2025-hour%20sleep-
deprivation%20episode%20in%20young%20and%20middle-
aged%20subjects&journal=Sleep&doi=10.1093%2Fsleep%2F27.1.55&volume=27&pages=55-60&publication_year=2004&author=Drapeau%2CC&author=Carrier%2CJ)
47. Leproult, R. et al. Individual differences in subjective and objective alertness during sleep deprivation are stable and unrelated. _Am. J. Physiol. Regul. Integr. Comp. Physiol._ **284** , R280–290 (2003).
[ Article ](https://doi.org/10.1152%2Fajpregu.00197.2002) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=12529281)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BD3sXhtlOqsLs%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Individual%20differences%20in%20subjective%20and%20objective%20alertness%20during%20sleep%20deprivation%20are%20stable%20and%20unrelated&journal=Am.%20J.%20Physiol.%20Regul.%20Integr.%20Comp.%20Physiol.&doi=10.1152%2Fajpregu.00197.2002&volume=284&pages=R280-290&publication_year=2003&author=Leproult%2CR)
48. Finelli, L. A., Baumann, H., Borbély, A. A. & Achermann, P. Dual electroencephalogram markers of human sleep homeostasis: correlation between theta activity in waking and slow-wave activity in sleep. _Neuroscience_ **101** , 523–529 (2000).
[ Article ](https://doi.org/10.1016%2FS0306-4522%2800%2900409-7) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=11113301)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BD3cXosFemsbY%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Dual%20electroencephalogram%20markers%20of%20human%20sleep%20homeostasis%3A%20correlation%20between%20theta%20activity%20in%20waking%20and%20slow-
wave%20activity%20in%20sleep&journal=Neuroscience&doi=10.1016%2FS0306-4522%2800%2900409-7&volume=101&pages=523-529&publication_year=2000&author=Finelli%2CLA&author=Baumann%2CH&author=Borb%C3%A9ly%2CAA&author=Achermann%2CP)
49. van Hees, V. T. et al. Estimating sleep parameters using an accelerometer without sleep diary. _Sci. Rep._ **8** , 12975 (2018).
[ Article ](https://doi.org/10.1038%2Fs41598-018-31266-z) [ ADS
](http://adsabs.harvard.edu/cgi-bin/nph-
data_query?link_type=ABSTRACT&bibcode=2018NatSR...812985H) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30154500)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6113241) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Estimating%20sleep%20parameters%20using%20an%20accelerometer%20without%20sleep%20diary&journal=Sci.%20Rep.&doi=10.1038%2Fs41598-018-31266-z&volume=8&publication_year=2018&author=Hees%2CVT)
50. Petrowski, K., Schmalbach, B., Niedling, M. & Stalder, T. The effects of post-awakening light exposure on the cortisol awakening response in healthy male individuals. _Psychoneuroendocrinology_ **108** , 28–34 (2019).
[ Article ](https://doi.org/10.1016%2Fj.psyneuen.2019.05.016) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31220690)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC1MXhtF2nsbnI) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20effects%20of%20post-
awakening%20light%20exposure%20on%20the%20cortisol%20awakening%20response%20in%20healthy%20male%20individuals&journal=Psychoneuroendocrinology&doi=10.1016%2Fj.psyneuen.2019.05.016&volume=108&pages=28-34&publication_year=2019&author=Petrowski%2CK&author=Schmalbach%2CB&author=Niedling%2CM&author=Stalder%2CT)
51. Souman, J. L., Tinga, A. M., Te Pas, S. F., van Ee, R. & Vlaskamp, B. N. S. Acute alerting effects of light: A systematic literature review. _Behav. Brain Res._ **337** , 228–239 (2018).
[ Article ](https://doi.org/10.1016%2Fj.bbr.2017.09.016) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28912014)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Acute%20alerting%20effects%20of%20light%3A%20A%20systematic%20literature%20review&journal=Behav.%20Brain%20Res.&doi=10.1016%2Fj.bbr.2017.09.016&volume=337&pages=228-239&publication_year=2018&author=Souman%2CJL&author=Tinga%2CAM&author=Pas%2CSF&author=Ee%2CR&author=Vlaskamp%2CBNS)
52. Asnicar, F. et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. _Nat. Med_ . [ https://doi.org/10.1038/s41591-020-01183-8 ](https://doi.org/10.1038/s41591-020-01183-8) (2021).
53. Buysse, D. J., Reynolds, C. F. 3rd, Monk, T. H., Berman, S. R. & Kupfer, D. J. The Pittsburgh Sleep Quality Index: a new instrument for psychiatric practice and research. _Psychiatry Res._ **28** , 193–213 (1989).
[ Article ](https://doi.org/10.1016%2F0165-1781%2889%2990047-4) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2748771)
[ CAS ](/articles/cas-redirect/1:STN:280:DyaL1Mzhs1Wmsg%3D%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=The%20Pittsburgh%20Sleep%20Quality%20Index%3A%20a%20new%20instrument%20for%20psychiatric%20practice%20and%20research&journal=Psychiatry%20Res.&doi=10.1016%2F0165-1781%2889%2990047-4&volume=28&pages=193-213&publication_year=1989&author=Buysse%2CDJ&author=Reynolds%2CCF&author=Monk%2CTH&author=Berman%2CSR&author=Kupfer%2CDJ)
54. Wittmann, M., Dinich, J., Merrow, M. & Roenneberg, T. Social jetlag: misalignment of biological and social time. _Chronobiol. Int._ **23** , 497–509 (2006).
[ Article ](https://doi.org/10.1080%2F07420520500545979) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=16687322)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Social%20jetlag%3A%20misalignment%20of%20biological%20and%20social%20time&journal=Chronobiol.%20Int.&doi=10.1080%2F07420520500545979&volume=23&pages=497-509&publication_year=2006&author=Wittmann%2CM&author=Dinich%2CJ&author=Merrow%2CM&author=Roenneberg%2CT)
55. Kolovou, G. D. et al. Assessment and clinical relevance of non-fasting and postprandial triglycerides: an expert panel statement. _Curr. Vasc. Pharmacol._ **9** , 258–270 (2011).
[ Article ](https://doi.org/10.2174%2F157016111795495549) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=21314632)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BC3MXlvFyku78%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Assessment%20and%20clinical%20relevance%20of%20non-
fasting%20and%20postprandial%20triglycerides%3A%20an%20expert%20panel%20statement&journal=Curr.%20Vasc.%20Pharmacol.&doi=10.2174%2F157016111795495549&volume=9&pages=258-270&publication_year=2011&author=Kolovou%2CGD)
56. Wright, K. P. Jr, Hull, J. T. & Czeisler, C. A. Relationship between alertness, performance, and body temperature in humans. _Am. J. Physiol. Regul. Integr. Comp. Physiol._ **283** , R1370–7 (2002).
[ Article ](https://doi.org/10.1152%2Fajpregu.00205.2002) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=12388468)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BD38XpvVWisLg%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Relationship%20between%20alertness%2C%20performance%2C%20and%20body%20temperature%20in%20humans&journal=Am.%20J.%20Physiol.%20Regul.%20Integr.%20Comp.%20Physiol.&doi=10.1152%2Fajpregu.00205.2002&volume=283&pages=R1370-7&publication_year=2002&author=Wright%2CKP&author=Hull%2CJT&author=Czeisler%2CCA)
57. Syed, S., Morseth, B., Hopstock, L. A. & Horsch, A. Evaluating the performance of raw and epoch non-wear algorithms using multiple accelerometers and electrocardiogram recordings. _Sci. Rep._ **10** , 5866 (2020).
[ Article ](https://doi.org/10.1038%2Fs41598-020-62821-2) [ ADS
](http://adsabs.harvard.edu/cgi-bin/nph-
data_query?link_type=ABSTRACT&bibcode=2020NatSR..10.5866S) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32246080)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7125135) [ CAS
](/articles/cas-redirect/1:CAS:528:DC%2BB3cXmsVOrt78%3D) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Evaluating%20the%20performance%20of%20raw%20and%20epoch%20non-
wear%20algorithms%20using%20multiple%20accelerometers%20and%20electrocardiogram%20recordings&journal=Sci.%20Rep.&doi=10.1038%2Fs41598-020-62821-2&volume=10&publication_year=2020&author=Syed%2CS&author=Morseth%2CB&author=Hopstock%2CLA&author=Horsch%2CA)
58. van Hees, V. et al. _GGIR_ . [ https://doi.org/10.5281/zenodo.3474227 ](https://doi.org/10.5281/zenodo.3474227) (2019).
59. Jones, S. E. et al. Genetic studies of accelerometer-based sleep measures yield new insights into human sleep behaviour. _Nat. Commun._ **10** , 1585 (2019).
[ Article ](https://doi.org/10.1038%2Fs41467-019-09576-1) [ ADS
](http://adsabs.harvard.edu/cgi-bin/nph-
data_query?link_type=ABSTRACT&bibcode=2019NatCo..10.1585J) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30952852)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6451011) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Genetic%20studies%20of%20accelerometer-
based%20sleep%20measures%20yield%20new%20insights%20into%20human%20sleep%20behaviour&journal=Nat.%20Commun.&doi=10.1038%2Fs41467-019-09576-1&volume=10&publication_year=2019&author=Jones%2CSE)
60. R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [ https://www.R-project.org/ ](https://www.R-project.org/) (2020).
61. Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. _J. Stat. Softw., Artic._ **67** , 1–48 (2015).
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Fitting%20linear%20mixed-
effects%20models%20using%20lme4&journal=J.%20Stat.%20Softw.%2C%20Artic.&volume=67&pages=1-48&publication_year=2015&author=Bates%2CD&author=M%C3%A4chler%2CM&author=Bolker%2CB&author=Walker%2CS)
62. Kuznetsova, A. et al. lmerTest package: tests in linear mixed effects models. _J. Stat. Softw._ **82** , 1–26 (2017).
[ Article ](https://doi.org/10.18637%2Fjss.v082.i13) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=lmerTest%20package%3A%20tests%20in%20linear%20mixed%20effects%20models&journal=J.%20Stat.%20Softw.&doi=10.18637%2Fjss.v082.i13&volume=82&pages=1-26&publication_year=2017&author=Kuznetsova%2CA)
63. Lüdecke, D. sjPlot: Data visualization for statistics in social science. _R package version_ 2, (2018).
64. Lenth, R. V. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.5.3. [ https://CRAN.R-project.org/package=emmeans ](https://CRAN.R-project.org/package=emmeans) (2020).
65. Nakagawa, S. & Schielzeth, H. A general and simple method for obtaining R 2 from generalized linear mixed-effects models. _Methods Ecol. Evol._ **4** , 133–142 (2013).
[ Article ](https://doi.org/10.1111%2Fj.2041-210x.2012.00261.x) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=A%20general%20and%20simple%20method%20for%20obtaining%20R%202%20from%20generalized%20linear%20mixed-
effects%20models&journal=Methods%20Ecol.%20Evol.&doi=10.1111%2Fj.2041-210x.2012.00261.x&volume=4&pages=133-142&publication_year=2013&author=Nakagawa%2CS&author=Schielzeth%2CH)
66. Yu, H., Jiang, S. & Land, K. C. Multicollinearity in hierarchical linear models. _Soc. Sci. Res._ **53** , 118–136 (2015).
[ Article ](https://doi.org/10.1016%2Fj.ssresearch.2015.04.008) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=26188442)
[ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Multicollinearity%20in%20hierarchical%20linear%20models&journal=Soc.%20Sci.%20Res.&doi=10.1016%2Fj.ssresearch.2015.04.008&volume=53&pages=118-136&publication_year=2015&author=Yu%2CH&author=Jiang%2CS&author=Land%2CKC)
67. Yarkoni, T. & Westfall, J. Choosing prediction over explanation in psychology: lessons from machine learning. _Perspect. Psychol. Sci._ **12** , 1100–1122 (2017).
[ Article ](https://doi.org/10.1177%2F1745691617693393) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28841086)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6603289) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Choosing%20prediction%20over%20explanation%20in%20psychology%3A%20lessons%20from%20machine%20learning&journal=Perspect.%20Psychol.%20Sci.&doi=10.1177%2F1745691617693393&volume=12&pages=1100-1122&publication_year=2017&author=Yarkoni%2CT&author=Westfall%2CJ)
68. Lundberg, S. M. et al. From local explanations to global understanding with explainable AI for trees. _Nat. Mach. Intell._ **2** , 56–67 (2020).
[ Article ](https://doi.org/10.1038%2Fs42256-019-0138-9) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32607472)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7326367) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=From%20local%20explanations%20to%20global%20understanding%20with%20explainable%20AI%20for%20trees&journal=Nat.%20Mach.%20Intell.&doi=10.1038%2Fs42256-019-0138-9&volume=2&pages=56-67&publication_year=2020&author=Lundberg%2CSM)
69. Pedregosa, F. et al. Scikit-learn: machine learning in python. _J. Mach. Learn. Res._ **12** , 2825–2830 (2011).
[ MathSciNet ](http://www.ams.org/mathscinet-getitem?mr=2854348) [ MATH
](http://www.emis.de/MATH-item?1280.68189) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Scikit-
learn%3A%20machine%20learning%20in%20python&journal=J.%20Mach.%20Learn.%20Res.&volume=12&pages=2825-2830&publication_year=2011&author=Pedregosa%2CF)
70. Jonsson, H. et al. Differences between germline genomes of monozygotic twins. _Nat. Genet._ **53** , 27–34 (2021).
[ Article ](https://doi.org/10.1038%2Fs41588-020-00755-1) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33414551)
[ CAS ](/articles/cas-redirect/1:CAS:528:DC%2BB3MXosVOksw%3D%3D) [ Google
Scholar
](http://scholar.google.com/scholar_lookup?&title=Differences%20between%20germline%20genomes%20of%20monozygotic%20twins&journal=Nat.%20Genet.&doi=10.1038%2Fs41588-020-00755-1&volume=53&pages=27-34&publication_year=2021&author=Jonsson%2CH)
71. Purcell, S. Variance components models for gene–environment interaction in twin analysis. _Twin Res. Hum. Genet._ **5** , 554–571 (2002).
[ Article ](https://doi.org/10.1375%2F136905202762342026) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Variance%20components%20models%20for%20gene%E2%80%93environment%20interaction%20in%20twin%20analysis&journal=Twin%20Res.%20Hum.%20Genet.&doi=10.1375%2F136905202762342026&volume=5&pages=554-571&publication_year=2002&author=Purcell%2CS)
72. Scheike, T. H., Holst, K. K. & Hjelmborg, J. B. Estimating heritability for cause specific mortality based on twin studies. _Lifetime Data Anal._ **20** , 210–233 (2014).
[ Article ](https://link.springer.com/doi/10.1007/s10985-013-9244-x) [
MathSciNet ](http://www.ams.org/mathscinet-getitem?mr=3181012) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=23378036)
[ MATH ](http://www.emis.de/MATH-item?1322.62036) [ Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Estimating%20heritability%20for%20cause%20specific%20mortality%20based%20on%20twin%20studies&journal=Lifetime%20Data%20Anal.&doi=10.1007%2Fs10985-013-9244-x&volume=20&pages=210-233&publication_year=2014&author=Scheike%2CTH&author=Holst%2CKK&author=Hjelmborg%2CJB)
73. Wang, C., Roy-Gagnon, M.-H., Lefebvre, J.-F., Burkett, K. M. & Dubois, L. Modeling gene-environment interactions in longitudinal family studies: a comparison of methods and their application to the association between the IGF pathway and childhood obesity. _BMC Med. Genet._ **20** , 9 (2019).
[ Article ](https://link.springer.com/doi/10.1186/s12881-018-0739-x) [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30634949)
[ PubMed Central ](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6329142) [
Google Scholar
](http://scholar.google.com/scholar_lookup?&title=Modeling%20gene-
environment%20interactions%20in%20longitudinal%20family%20studies%3A%20a%20comparison%20of%20methods%20and%20their%20application%20to%20the%20association%20between%20the%20IGF%20pathway%20and%20childhood%20obesity&journal=BMC%20Med.%20Genet.&doi=10.1186%2Fs12881-018-0739-x&volume=20&publication_year=2019&author=Wang%2CC&author=Roy-
Gagnon%2CM-H&author=Lefebvre%2CJ-F&author=Burkett%2CKM&author=Dubois%2CL)
[ Download references ](https://citation-
needed.springer.com/v2/references/10.1038/s41467-022-34503-2?format=refman&flavour=references)
## Acknowledgements
We express our sincere thanks to the participants of the PREDICT1 study. We
thank the staff of Zoe Ltd, the Department of Twin Research and Genetic
Epidemiology, and Massachusetts General Hospital and the Translational and
Clinical Research Centre for their tireless work in contributing to the
running of the study and data collection. We thank Abbott for their support
with using their CGMs. This work was supported by Zoe Ltd. TwinsUK is funded
by the Wellcome Trust, Medical Research Council, European Union, Chronic
Disease Research Foundation (CDRF), Zoe Ltd and the National Institute for
Health Research (NIHR)-funded BioResource, Clinical Research Facility and
Biomedical Research Centre based at Guy’s and St Thomas’ NHS Foundation Trust
in partnership with King’s College London.
## Author information
### Authors and Affiliations
1. Center for Human Sleep Science, Department of Psychology, University of California, Berkeley, CA, USA
Raphael Vallat & Matthew P. Walker
2. Department of Nutritional Sciences, King’s College London, London, UK
Sarah E. Berry & Haya Al Khatib
3. Department of Clinical Sciences, Lund University, Malmö, Sweden
Neli Tsereteli & Paul W. Franks
4. Zoe Ltd, London, UK
Joan Capdevila, Haya Al Khatib & Jonathan Wolf
5. School of Medicine, University of Nottingham, Nottingham, UK
Ana M. Valdes
6. Nottingham NIHR Biomedical Research Centre, Nottingham, UK
Ana M. Valdes
7. Diabetes Unit, Massachusetts General Hospital and Harvard Medical School, Boston, MA, USA
Linda M. Delahanty
8. Clinical & Translational Epidemiology Unit, Massachusetts General Hospital and Harvard Medical School, Boston, MA, USA
David A. Drew & Andrew T. Chan
9. Division of Gastroenterology, Massachusetts General Hospital and Harvard Medical School, Boston, MA, USA
David A. Drew & Andrew T. Chan
10. Department of Twin Research and Genetic Epidemiology, King’s College London, London, UK
Paul W. Franks & Tim D. Spector
11. Department of Nutrition, Harvard Chan School of Public Health, Boston, MA, USA
Paul W. Franks
Authors
1. Raphael Vallat
[ View author publications ](/search?author=Raphael%20Vallat)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Raphael%20Vallat)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Raphael%20Vallat%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
2. Sarah E. Berry
[ View author publications ](/search?author=Sarah%20E.%20Berry)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Sarah%20E.%20Berry)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Sarah%20E.%20Berry%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
3. Neli Tsereteli
[ View author publications ](/search?author=Neli%20Tsereteli)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Neli%20Tsereteli)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Neli%20Tsereteli%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
4. Joan Capdevila
[ View author publications ](/search?author=Joan%20Capdevila)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Joan%20Capdevila)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Joan%20Capdevila%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
5. Haya Al Khatib
[ View author publications ](/search?author=Haya%20Al%20Khatib)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Haya%20Al%20Khatib)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Haya%20Al%20Khatib%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
6. Ana M. Valdes
[ View author publications ](/search?author=Ana%20M.%20Valdes)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Ana%20M.%20Valdes)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Ana%20M.%20Valdes%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
7. Linda M. Delahanty
[ View author publications ](/search?author=Linda%20M.%20Delahanty)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Linda%20M.%20Delahanty)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Linda%20M.%20Delahanty%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
8. David A. Drew
[ View author publications ](/search?author=David%20A.%20Drew)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=David%20A.%20Drew)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22David%20A.%20Drew%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
9. Andrew T. Chan
[ View author publications ](/search?author=Andrew%20T.%20Chan)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Andrew%20T.%20Chan)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Andrew%20T.%20Chan%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
10. Jonathan Wolf
[ View author publications ](/search?author=Jonathan%20Wolf)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Jonathan%20Wolf)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Jonathan%20Wolf%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
11. Paul W. Franks
[ View author publications ](/search?author=Paul%20W.%20Franks)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Paul%20W.%20Franks)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Paul%20W.%20Franks%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
12. Tim D. Spector
[ View author publications ](/search?author=Tim%20D.%20Spector)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Tim%20D.%20Spector)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Tim%20D.%20Spector%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
13. Matthew P. Walker
[ View author publications ](/search?author=Matthew%20P.%20Walker)
You can also search for this author in [ PubMed
](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=Matthew%20P.%20Walker)
[ Google Scholar
](http://scholar.google.co.uk/scholar?as_q=&num=10&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=%22Matthew%20P.%20Walker%22&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en)
### Contributions
Conceptualization: R.V., S.E.B., A.M.V., D.A.D., A.T.C., J.W., P.W.F., T.D.S.,
M.P.W. Investigation: S.E.B., J.C., D.A.D, A.T.C, J.W, T.D.S. Formal analysis:
R.V, S.E.B., N.T., J.C., H.A.K., M.P.W. Data curation: R.V., N.T., J.C.,
H.A.K. Writing - original draft: R.V. and M.P.W. Writing: review & editing:
R.V., S.E.B., N.T., J.C., H.A.K., A.M.V., L.M.D., D.A.D., A.T.C., J.W.,
P.W.F., T.D.S., M.P.W.
### Corresponding authors
Correspondence to [ Raphael Vallat ](mailto:[email protected]) or [
Matthew P. Walker ](mailto:[email protected]) .
## Ethics declarations
### Competing interests
T.D.S., S.E.B., L.M.D, A.M.V. and P.W.F. are consultants to Zoe Ltd (“Zoe”).
J.W. and J.C. are or have been employees of Zoe and TDS is a co-founder with
an equity interest. M.P.W. serves as a consultant for and has equity interest
in Oura, Bryte, Shuni and StimScience. Other authors have no Competing
Interests to declare.
## Peer review
### Peer review information
_Nature Communications_ thanks the anonymous reviewers for their contribution
to the peer review of this work.
## Additional information
**Publisher’s note** Springer Nature remains neutral with regard to
jurisdictional claims in published maps and institutional affiliations.
## Supplementary information
### [ Supplementary Information ](https://static-
content.springer.com/esm/art%3A10.1038%2Fs41467-022-34503-2/MediaObjects/41467_2022_34503_MOESM1_ESM.pdf)
### [ Reporting Summary ](https://static-
content.springer.com/esm/art%3A10.1038%2Fs41467-022-34503-2/MediaObjects/41467_2022_34503_MOESM2_ESM.pdf)
## Source data
### [ Source data ](https://static-
content.springer.com/esm/art%3A10.1038%2Fs41467-022-34503-2/MediaObjects/41467_2022_34503_MOESM3_ESM.xlsx)
## Rights and permissions
**Open Access** This article is licensed under a Creative Commons Attribution
4.0 International License, which permits use, sharing, adaptation,
distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to
the Creative Commons license, and indicate if changes were made. The images or
other third party material in this article are included in the article’s
Creative Commons license, unless indicated otherwise in a credit line to the
material. If material is not included in the article’s Creative Commons
license and your intended use is not permitted by statutory regulation or
exceeds the permitted use, you will need to obtain permission directly from
the copyright holder. To view a copy of this license, visit [
http://creativecommons.org/licenses/by/4.0/
](http://creativecommons.org/licenses/by/4.0/) .
[ Reprints and permissions
](https://s100.copyright.com/AppDispatchServlet?title=How%20people%20wake%20up%20is%20associated%20with%20previous%20night%E2%80%99s%20sleep%20together%20with%20physical%20activity%20and%20food%20intake&author=Raphael%20Vallat%20et%20al&contentID=10.1038%2Fs41467-022-34503-2©right=The%20Author%28s%29&publication=2041-1723&publicationDate=2022-11-19&publisherName=SpringerNature&orderBeanReset=true&oa=CC%20BY)
## About this article
[ 
](https://crossmark.crossref.org/dialog/?doi=10.1038/s41467-022-34503-2)
### Cite this article
Vallat, R., Berry, S.E., Tsereteli, N. _et al._ How people wake up is
associated with previous night’s sleep together with physical activity and
food intake. _Nat Commun_ **13** , 7116 (2022).
https://doi.org/10.1038/s41467-022-34503-2
[ Download citation ](https://citation-
needed.springer.com/v2/references/10.1038/s41467-022-34503-2?format=refman&flavour=citation)
* Received : 26 May 2021
* Accepted : 27 October 2022
* Published : 19 November 2022
* DOI : https://doi.org/10.1038/s41467-022-34503-2
### Share this article
Anyone you share the following link with will be able to read this content:
Get shareable link
Sorry, a shareable link is not currently available for this article.
Copy to clipboard
Provided by the Springer Nature SharedIt content-sharing initiative
## This article is cited by
* ### [ Shorter self-reported sleep duration is associated with worse virtual spatial navigation performance in men ](https://doi.org/10.1038/s41598-024-52662-8)
* Emre Yavuz
* Christoffer J. Gahnstrom
* Hugo J. Spiers
_Scientific Reports_ (2024)
* ### [ Individual sleep need is flexible and dynamically related to cognitive function ](https://doi.org/10.1038/s41562-024-01827-6)
* Anders M. Fjell
* Kristine B. Walhovd
_Nature Human Behaviour_ (2024)
* ### [ Sleep Apnea, Obesity, and Diabetes — an Intertwined Trio ](https://doi.org/10.1007/s11892-023-01510-6)
* Soumya Kurnool
* Karen C. McCowen
* Atul Malhotra
_Current Diabetes Reports_ (2023)
## Comments
By submitting a comment you agree to abide by our [ Terms ](/info/tandc.html)
and [ Community Guidelines ](/info/community-guidelines.html) . If you find
something abusive or that does not comply with our terms or guidelines please
flag it as inappropriate.
[ Download PDF ](/articles/s41467-022-34503-2.pdf)
## Associated content
Collection
### [ Top 25 Health Sciences Articles of 2022
](https://www.nature.com/collections/hefjafigfj)
Advertisement
[

](//pubads.g.doubleclick.net/gampad/jump?iu=/285/nature_communications/article&sz=300x250&c=539351078&t=pos%3Dright%26type%3Darticle%26artid%3Ds41467-022-34503-2%26doi%3D10.1038/s41467-022-34503-2%26techmeta%3D141%26subjmeta%3D1385,208,2811,378,477,631,729%26kwrd%3DCircadian+rhythms+and+sleep,Heritable+quantitative+trait,Human+behaviour)
## Explore content
* [ Research articles ](/ncomms/research-articles)
* [ Reviews & Analysis ](/ncomms/reviews-and-analysis)
* [ News & Comment ](/ncomms/news-and-comment)
* [ Videos ](/ncomms/video)
* [ Collections ](/ncomms/collections)
* [ Subjects ](/ncomms/browse-subjects)
* [ Follow us on Facebook ](https://www.facebook.com/NatureCommunications)
* [ Follow us on Twitter ](https://twitter.com/NatureComms)
* [ Sign up for alerts ](https://www.nature.com/my-account/alerts/subscribe-journal?list-id=264)
* [ RSS feed ](https://www.nature.com/ncomms.rss)
## About the journal
* [ Aims & Scope ](/ncomms/aims)
* [ Editors ](/ncomms/editors)
* [ Journal Information ](/ncomms/journal-information)
* [ Open Access Fees and Funding ](/ncomms/open-access)
* [ Calls for Papers ](/ncomms/calls-for-papers)
* [ Editorial Values Statement ](/ncomms/editorial-values-statement)
* [ Journal Metrics ](/ncomms/journal-impact)
* [ Editors' Highlights ](/ncomms/editorshighlights)
* [ Contact ](/ncomms/contact)
* [ Editorial policies ](/ncomms/editorial-policies)
* [ Top Articles ](/ncomms/top-articles)
## Publish with us
* [ For authors ](/ncomms/submit)
* [ For Reviewers ](/ncomms/for-reviewers)
* [ Language editing services ](https://authorservices.springernature.com/go/sn/?utm_source=For+Authors&utm_medium=Website_Nature&utm_campaign=Platform+Experimentation+2022&utm_id=PE2022)
* [ Submit manuscript ](https://mts-ncomms.nature.com/)
## Search
Search articles by subject, keyword or author
Show results from All journals This journal
Search
[ Advanced search ](/search/advanced)
### Quick links
* [ Explore articles by subject ](/subjects)
* [ Find a job ](/naturecareers)
* [ Guide to authors ](/authors/index.html)
* [ Editorial policies ](/authors/editorial_policies/)
Nature Communications ( _Nat Commun_ ) ISSN 2041-1723 (online)
## nature.com sitemap
### About Nature Portfolio
* [ About us ](https://www.nature.com/npg_/company_info/index.html)
* [ Press releases ](https://www.nature.com/npg_/press_room/press_releases.html)
* [ Press office ](https://press.nature.com/)
* [ Contact us ](https://support.nature.com/support/home)
### Discover content
* [ Journals A-Z ](https://www.nature.com/siteindex)
* [ Articles by subject ](https://www.nature.com/subjects)
* [ protocols.io ](https://www.protocols.io/)
* [ Nature Index ](https://www.natureindex.com/)
### Publishing policies
* [ Nature portfolio policies ](https://www.nature.com/authors/editorial_policies)
* [ Open access ](https://www.nature.com/nature-research/open-access)
### Author & Researcher services
* [ Reprints & permissions ](https://www.nature.com/reprints)
* [ Research data ](https://www.springernature.com/gp/authors/research-data)
* [ Language editing ](https://authorservices.springernature.com/language-editing/)
* [ Scientific editing ](https://authorservices.springernature.com/scientific-editing/)
* [ Nature Masterclasses ](https://masterclasses.nature.com/)
* [ Research Solutions ](https://solutions.springernature.com/)
### Libraries & institutions
* [ Librarian service & tools ](https://www.springernature.com/gp/librarians/tools-services)
* [ Librarian portal ](https://www.springernature.com/gp/librarians/manage-your-account/librarianportal)
* [ Open research ](https://www.nature.com/openresearch/about-open-access/information-for-institutions)
* [ Recommend to library ](https://www.springernature.com/gp/librarians/recommend-to-your-library)
### Advertising & partnerships
* [ Advertising ](https://partnerships.nature.com/product/digital-advertising/)
* [ Partnerships & Services ](https://partnerships.nature.com/)
* [ Media kits ](https://partnerships.nature.com/media-kits/)
* [ Branded content ](https://partnerships.nature.com/product/branded-content-native-advertising/)
### Professional development
* [ Nature Careers ](https://www.nature.com/naturecareers/)
* [ Nature Conferences ](https://conferences.nature.com)
### Regional websites
* [ Nature Africa ](https://www.nature.com/natafrica)
* [ Nature China ](http://www.naturechina.com)
* [ Nature India ](https://www.nature.com/nindia)
* [ Nature Italy ](https://www.nature.com/natitaly)
* [ Nature Japan ](https://www.natureasia.com/ja-jp)
* [ Nature Middle East ](https://www.nature.com/nmiddleeast)
* [ Privacy Policy ](https://www.nature.com/info/privacy)
* [ Use of cookies ](https://www.nature.com/info/cookies)
* Your privacy choices/Manage cookies
* [ Legal notice ](https://www.nature.com/info/legal-notice)
* [ Accessibility statement ](https://www.nature.com/info/accessibility-statement)
* [ Terms & Conditions ](https://www.nature.com/info/terms-and-conditions)
* [ Your US state privacy rights ](https://www.springernature.com/ccpa)
[ 
](https://www.springernature.com/)
© 2024 Springer Nature Limited
Close banner Close

Sign up for the _Nature Briefing_ newsletter — what matters in science, free
to your inbox daily.
Email address
Sign up
I agree my information will be processed in accordance with the _Nature_ and
Springer Nature Limited [ Privacy Policy
](https://www.nature.com/info/privacy) .
Close banner Close
Get the most important science stories of the day, free in your inbox. [ Sign
up for Nature Briefing
](https://www.nature.com/briefing/signup/?brieferEntryPoint=MainBriefingBanner)


*[
DOI
]: Digital Object Identifier
*[
ISSN
]: International Standard Serial Number
| biology | 1139746 | https://no.wikipedia.org/wiki/Kongevann | Kongevann | Kongevann eller aqua regia (latin) er en blanding av salpetersyre og saltsyre, optimalt i et molforhold på 1: 3. Aqua regia er en gul-oransje (noen ganger rød) røykende væske, kalt av alkymister fordi den kan oppløse edle metaller, gull og platina, men ikke alle metaller.
Produksjon og dekomponering
Ved blanding av konsentrert saltsyre og konsentrert salpetersyre oppstår kjemiske reaksjoner. Disse reaksjonene resulterer i de flyktige produktene, nitrosylklorid og klorgass:
HNO_{3 (aq)} {+} 3HCl_{(aq)} ->NOCl_{(g)} {+} Cl_{2 (g)} {+} 2H2O_{(l)}
Nitrosylklorid kan videre spaltes til nitrogenoksid og klor:
2NOCl_{(g)}<=>2NO_{(g)} {+}Cl_{2 (g)}
Denne dissosiasjonen er likevektsbegrenset. I tillegg til nitrosylklorid og klor inneholder gassen over vannregion nitrogenoksid. Fordi nitrogenoksid reagerer lett med atmosfærisk oksygen, inneholder gassene som produseres også nitrogendioksid, NO2:
2NO_{(g)} {+ }O_{2 (g)}-> 2NO_{2 (g)}
Kjemien
Oppløse gull
Aqua regia løser opp gull, selv om ingen av bestanddelene av syre vil gjøre det alene, fordi hver syre i kombinasjon utfører en annen oppgave. Salpetersyre er en kraftig oksidasjonsmiddel, som faktisk vil oppløse en nesten umulig mengde gull og danne gullioner (Au3 +). Saltsyren gir en klar tilførsel av kloridioner (Cl−), som reagerer med gullionene for å produsere tetrakloroaurat (III) -ioner, i løsning. Reaksjonen med saltsyre er en likevektsreaksjon som favoriserer dannelse av klorauratanioner (AuCl4−). Dette resulterer i en fjerning av gullioner fra løsningen og tillater videre oksidasjon av gull å finne sted. Gullet oppløses for å bli kloroaursyre. I tillegg kan gull løses opp av klor som er tilstede. Egnede ligninger er:
Au + 3HNO3 +4HCl <=>> [AuCl4]- +3[NO2] +[H3O]+ +2H2O
eller
Au + HNO3 +4HCl <=>> [AuCl4]- +[NO] +[H3O]+ +H2O
Hvis aqua regia-oppløsningen bare inneholder gull, kan fast tetrakloraurinsyre fremstilles ved å koke av overflødig aqua regia, og fjerne gjenværende salpetersyre ved gjentatt oppvarming med saltsyre. Dette trinnet reduserer salpetersyre. Hvis elementært gull er ønsket, kan det reduseres selektivt med svoveldioksid, hydrazin, oksalsyre, etc. Ligningen for reduksjon av gull med svoveldioksid er:
2AuCl-_{4 (aq)} +3SO_{2 (g)} {+}6H2O_{(l)} ->2Au_{(s)} {+}12H+_{(aq)} {+3SO^{2-}_{4 (aq)} {+}8Cl^{-}_{(aq)}}
Oppløse platina
Lignende ligninger kan skrives for platina. Som med gull kan oksidasjonsreaksjonen skrives med nitrogenoksid eller nitrogendioksid som nitrogenoksidprodukt:
Pt_{(s)} {+}4NO^{-}_{3 (aq)} {+}8H^{+}_{(aq)} -> Pt^{4+}_{(aq)} {+}4NO_{ (g)} {+}4H2O_{(l)}
3Pt_{(s)} {+}4NO^{-}_{3 (aq)} {+}16H^{+}_{(aq)} -> 3Pt^{4+}_{(aq)} {+}4NO_{ (g)} {+}8H2O_{(l)}
Det oksiderte platina-ionet reagerer deretter med kloridioner, noe som resulterer i kloroplatination:
Pt^{4+}_{(aq)} {+}6Cl^{-}_{(aq)} ->PtCl^{2-}_{6 (aq)}Eksperimentelle bevis viser at reaksjonen av platina med aqua regia er betydelig mer kompleks. De første reaksjonene produserer en blanding av kloroplatinsyre (H2PtCl4) og nitrosoplatinisk klorid ((NO) 2PtCl4). Nitrosoplatinisk klorid er et fast produkt. Hvis full oppløsning av platina er ønsket, må gjentatte ekstraksjoner av gjenværende faste stoffer med konsentrert saltsyre utføres:
2Pt_{(s)} {+} 2HNO3_{(aq)} {+} 8 HCl_{(aq)} ->(NO)2PtCl4_{(s)} {+}
H2PtCl4_{(aq)} {+} 4 H2O_{(l)}
og
(NO)2PtCl4_{(s)} {+} 2 HCl_{(aq)}<=> H2PtCl4 _{(aq)} {+} 2 NOCl_{(g)}
Klorplatinsyren kan oksideres til klorplatinsyre ved å mette løsningen med klor under oppvarming:
H2PtCl4_{(aq)} {+} Cl2_{(g)} -> H2PtCl6_{(aq)}
Å oppløse platina (fast form) i aqua regia var oppdagelsesmåten for de tetteste metallene iridium og osmium, som begge finnes i platina malm og ikke vil bli oppløst av syren, men samler seg i bunnen av karet.
Utfelling av oppløst platina
Når platinagruppemateller renses med aqua regia uftelles gull ved behandling med jern (II) klorid. Platina i løsningen omdannes som heksaklorplatinat (IV) til ammoniumheksaklorplatinat ved tilsetning av ammoniumklorid. Dette ammoniumsaltet er ekstremt uoppløselig, og det kan filtreres av. Med sterk oppvarming konverterer det til platina metall:
3(NH4)2PtCl6 -> 3 Pt + 2N2 + 2NH4Cl + 16HCl
Uutfelt heksaklorplatinat (IV) reduseres med elementært sink, og en lignende metode er egnet for utvinning av platina fra laboratorierester i liten skala.
Reaksjon med tinn
Aqua regia reagerer med tinn for å danne tinn (IV) klorid, som inneholder tinn i sin høyeste oksidasjonstilstand:
4HCl + 2HNO3 +Sn ->SnCl4 +NO2 +NO +3H2O
Referanser
Syrer | norwegian_bokmål | 1.281224 |
wake_up_wrong/sleepinertiacurrenti.txt | Javascript is currently disabled in your browser. Several features of this
site will not function whilst javascript is disabled.
* [ Journals ](browse_journals.php)
* [ Why Publish With Us? ](about-us)
* [ Editorial Policies ](editorial-policies)
* [ Author Guidelines ](author-guidelines)
* [ Peer Review Guidelines ](peer-review-guidelines)
* [ Open Outlook ](open_outlook.php)
* [ Hot topics ](browse_hot_topics.php)
* [ Podcasts ](podcasts)
* [ Blog ](blog)
* [ Reprints ](bulk_reprints.php)
* [ Submit New Manuscript ](submit_step_1.php)
 Menu
* [ About ](about_dovepress.php)
* [ Contact ](contact)
* [ Sustainability ](sustainability.php)
* [ Awards ](author-awards)
* [ Press Center ](press-releases)
* [ Testimonials ](why-publish-with-dove-press/testimonials)
* [ Blog ](blog)
* [ Favored Author Program ](why-publish-with-dove-press/favored-authors)
* [ Permissions ](permissions.php)
* [ Pre-Submission ](presubmission_check.php)
* [ Reprints ](bulk_reprints.php)
* [ **Login** ](login.php)
* [  ](http://www.dovepress.com.cn)
[  ](index.php)
open access to scientific and medical research
[ Advanced search ](search_advance.php)
* [ Journals ](browse_journals.php)
* [ Why publish with us? ](about-us)
* [ Editorial Policies ](editorial_policies.php)
* [ Author Information ](author-guidelines)
* [ Peer Review Guidelines ](peer-review-guidelines)
* [ Open Outlook ](open_outlook.php)
* [ Hot topics ](browse_hot_topics.php)
* [ Podcasts ](podcasts)
#### 9 0 8 6 5
Papers Published
[ Submit New Manuscript ](submit_step_1.php)
[ Login to view existing manuscript status ](script_status.php)
[ Sign Up for Journal alerts ](register.php)
#### About Dove Press
Open access peer-reviewed scientific and medical journals.
[ Learn more ](https://www.dovepress.com/about_dovepress.php)
#### Open Access

Dove Medical Press is a member of the OAI.
[ Learn more ](https://www.dovepress.com/why-publish-with-dove-press/open-
access/)
#### Reprints
Bulk reprints for the pharmaceutical industry.
[ Learn more ](bulk_reprints.php)
#### Favored Authors
We offer real benefits to our authors, including fast-track processing of
papers.
[ Learn more ](fav_author.php)
#### Promotional Article Monitoring
Register your specific details and specific drugs of interest and we will
match the information you provide to articles from our extensive database and
email PDF copies to you promptly.
[ Learn more ](https://www.dovepress.com/pam_reprint_form.php)
##### Social Media
[ ](https://www.facebook.com/DoveMedicalPress) [
](https://twitter.com/DovePress) [ ](https://www.linkedin.com/company/dove-
medical-press) [ ](https://www.youtube.com/user/dovepress)
[ Live Support Chat
](http://messenger.providesupport.com/messenger/0u5bkmd8ytkci1n86hdd6hcgj4.html)
Back to [ Journals ](browse_journals.php) » [ Nature and Science of Sleep
](nature-and-science-of-sleep-journal) » [ Volume 11 ](nature-and-science-of-
sleep-archive73-v1408)
Listen
Review

#
Sleep inertia: current insights
[
](https://twitter.com/intent/tweet?text=Sleep+inertia%3A+current+insights+%7C+NSS+%40DovePress&related=DovePress&url=https://www.dovepress.com/sleep-
inertia-current-insights-peer-reviewed-fulltext-article-NSS "share this page
on Twitter") [
](https://www.facebook.com/sharer.php?u=https://www.dovepress.com/sleep-
inertia-current-insights-peer-reviewed-fulltext-article-NSS&t=Sleep inertia:
current insights | NSS "share this page on Facebook") [
](qr.php?p=https://www.dovepress.com/sleep-inertia-current-insights-peer-
reviewed-fulltext-article-NSS "share this page on WeChat") [
](https://www.linkedin.com/shareArticle?mini=true&url=https://www.dovepress.com/sleep-
inertia-current-insights-peer-reviewed-fulltext-article-NSS&title=Sleep
inertia: current insights | NSS&source=Dove Press "share this page on
Linkedin") [
](https://www.reddit.com/submit?url=https://www.dovepress.com/sleep-inertia-
current-insights-peer-reviewed-fulltext-article-NSS "share this page on
Reddit") [ ](/cdn-cgi/l/email-
protection#340b4741565e515740097b44515a14755757514747145546405d575851145b5a14705b4251147951505d57555814644651474712565b504d09775c51575f145b414014405c5d47145546405d5758510e3e145c404044470e1b1b4343431a505b425144465147471a575b591b4758515144195d5a5146405d551957414646515a40195d5a475d535c40471944515146194651425d51435150195241585840514c40195546405d575851197a6767
"share this page via Email")
[
](https://twitter.com/intent/tweet?text=Sleep+inertia%3A+current+insights+%7C+NSS+%40DovePress&related=DovePress&url=https://www.dovepress.com/sleep-
inertia-current-insights-peer-reviewed-fulltext-article-NSS "share this page
on Twitter") [
](https://www.facebook.com/sharer.php?u=https://www.dovepress.com/sleep-
inertia-current-insights-peer-reviewed-fulltext-article-NSS&t=Sleep inertia:
current insights | NSS "share this page on Facebook") [
](qr.php?p=https://www.dovepress.com/sleep-inertia-current-insights-peer-
reviewed-fulltext-article-NSS "share this page on WeChat") [
](https://www.linkedin.com/shareArticle?mini=true&url=https://www.dovepress.com/sleep-
inertia-current-insights-peer-reviewed-fulltext-article-NSS&title=Sleep
inertia: current insights | NSS&source=Dove Press "share this page on
Linkedin") [
](https://www.reddit.com/submit?url=https://www.dovepress.com/sleep-inertia-
current-insights-peer-reviewed-fulltext-article-NSS "share this page on
Reddit") [ ](/cdn-cgi/l/email-
protection#a699d5d3c4ccc3c5d29be9d6c3c886e7c5c5c3d5d586c7d4d2cfc5cac386c9c886e2c9d0c386ebc3c2cfc5c7ca86f6d4c3d5d580c4c9c2df9be5cec3c5cd86c9d3d286d2cecfd586c7d4d2cfc5cac39cac86ced2d2d6d59c8989d1d1d188c2c9d0c3d6d4c3d5d588c5c9cb89d5cac3c3d68bcfc8c3d4d2cfc78bc5d3d4d4c3c8d28bcfc8d5cfc1ced2d58bd6c3c3d48bd4c3d0cfc3d1c3c28bc0d3cacad2c3ded28bc7d4d2cfc5cac38be8f5f5
"share this page via Email")
* Fulltext
* [ Metrics ](article_metric.php?article_id=48029)
* [ Get Permission ](get_permissions.php?a_id=48029)
* Cite this article
**Authors** [ Hilditch CJ ](author_profile.php?id=1020966) , [ McHill AW
](author_profile.php?id=905583) [ ](https://orcid.org/0000-0002-9428-6884
"https://orcid.org/0000-0002-9428-6884")
**Received** 2 April 2019
**Accepted for publication** 10 August 2019
**Published** 22 August 2019 Volume 2019:11 Pages 155—165
**DOI** [ https://doi.org/10.2147/NSS.S188911
](https://doi.org/10.2147/NSS.S188911)
**Checked for plagiarism** Yes
**Review by** [ Single anonymous peer review ](peer-review-guidelines)
**Peer reviewer comments** 3
**Editor who approved publication:** [ Dr Sutapa Mukherjee
](public_profile.php?id=1019855)
[ Download Article [PDF] ](getfile.php?fileID=52180)
Cassie J Hilditch, 1 Andrew W McHill 2
1 Fatigue Countermeasures Laboratory, San Jose State University Research
Foundation, Moffett Field, San Jose, CA 94035, USA; 2 Oregon Institute of
Occupational Health Sciences, Oregon Health and Science University, Portland,
OR 97239, USA
Correspondence: Andrew W McHill
Oregon Institute of Occupational Health Sciences, Oregon Health and Science
University, 3181 SW Sam Jackson Park Road - L606
Portland, OR 97239 USA
Tel +1 503 494 2594
Fax +1 503 494 4278
Email [ [email protected] ](/cdn-cgi/l/email-protection)
**Abstract:** Sleep inertia, or the grogginess felt upon awakening, is
associated with significant cognitive performance decrements that dissipate as
time awake increases. This impairment in cognitive performance has been
observed in both tightly controlled in-laboratory studies and in real-world
scenarios. Further, these decrements in performance are exaggerated by prior
sleep loss and the time of day in which a person awakens. This review will
examine current insights into the causes of sleep inertia, factors that may
positively or negatively influence the degree of sleep inertia, the
consequences of sleep inertia both in the laboratory and in real-world
settings, and lastly discuss potential countermeasures to lessen the impact of
sleep inertia.
**Keywords:** cognitive performance, countermeasures, grogginess
## Introduction
“ … immediately after getting up, irrespective of the hour, one is not at
one’s best,” Kleitman 1 observed, in perhaps the earliest known description
of sleep inertia in the sleep literature. Sleep inertia is the term used to
refer to the temporary time of sleepiness, disorientation and impaired
cognitive performance experienced upon awakening. 2 Early investigations 3
– 8 of the paradoxical phenomenon of “waking up tired”, although somewhat
limited by low participant numbers and methodological design, have had their
findings of impaired performance upon awakening supported by a wealth of
subsequent, more rigorous studies. Understanding the underlying causes,
consequences, and countermeasures to sleep inertia is important for managing
this period of impaired performance for those working on-call or napping on-
shift who are then required to perform safety-critical tasks soon after
waking. The aim of this review is to summarize decades of research that have
led to current insights into the following areas of sleep inertia:
characterization of neurophysiological correlates; contributing factors
influencing severity; neurobehavioral and real-world consequences; and
finally, proactive and reactive countermeasures. The review will first define
sleep inertia as part of the three-process model of sleep regulation 9 and
then address each of the topics listed above.
## Sleep inertia: the third process of sleep regulation
The propensity for sleep and sleepiness is governed by two main processes: a
homeostatic drive or pressure for sleep (Process S), and a circadian rhythm of
arousal (Process C). 10 , 11 Process S promotes sleepiness as hours of
wakefulness accumulate and is dissipated with time spent asleep. Process C is
driven by a circadian pacemaker, the suprachiasmatic nucleus, to promote sleep
at night and alertness during the day. 12 Thus, the well-accepted two-
process model of sleep regulation would theoretically predict higher alertness
and performance following depletion of the homeostatic drive, which often
coincides with a rise in the circadian drive for alertness following habitual
morning awakenings. What is observed at this time, however, is a short, yet
salient, decrease in alertness and performance immediately after waking. 4 –
6 Therefore, a third-process (Process W; or Process I, in some descriptions)
9 is necessary to better describe these fluctuations in sleepiness. In a
three-process model ( Figure 1 ), sleep inertia (green line) is immediately
present at sleep offset, briefly counteracting the low homeostatic drive for
sleep (red line) and the rising circadian drive for wakefulness (blue line).
[

](getfile_article_fulltext.php?filename=article_fulltext%2Fs188000%2F188911/img/NSS_A_188911_O_F0001g.jpg)
|
**Figure 1** A schematic of the three-process model of sleep regulation.
---|---
The exact function of sleep inertia remains largely unknown. 13 ** , ** 14
From an evolutionary perspective, one might posit that the ability to rapidly
awaken from sleep would be advantageous, for example when awakening in
response to a potential threat. 13 ** , ** 14 A more gradual awakening,
however, may also be protective given the complexity of neural circuitry in
transitioning from one state to another, 14 as is discussed in the
neurophysiology section below. Sleep inertia may, therefore, be an adaptive
mechanism to promote sleep upon awakening so that sleep is maintained when the
awakening is undesired. For example, as with the timing of the circadian
nadir, 15 sleep inertia may help to maintain sleep in the later part of a
nocturnal sleep episode when homeostatic sleep pressure has largely
dissipated. It is only when the restoration of wakefulness needs to be rapid
that the process of sleep inertia appears maladaptive. Vyazovskiy et al 14
also propose that the gradual awakening following rapid eye movement (REM)
sleep may be a “reset” function to minimize hypnopompic intrusions into
wakefulness. These theories as to the function of sleep inertia require
further research. Animal studies investigating the evolutionary progression of
sleep inertia across species may be useful in addressing this gap in the
literature. Currently, neurophysiological studies have provided perhaps the
best insights into this area, as summarized in the next section.
## Neurophysiological basis of sleep inertia
While the transition from sleep to wakefulness has received comparatively
little attention relative to the transition from wakefulness to sleep, there
is growing evidence for the underlying neurophysiological causes of sleep
inertia. For example, spectral analyses of electroencephalography (EEG) after
waking have offered insight into the encephalographic substratum of sleep
inertia. Compared to pre-sleep wakefulness, post-sleep EEG typically contains
higher delta power (associated with deep sleep) and lower beta power
(associated with wakefulness). 16 – 19 Reported changes in theta power
(associated with drowsiness or REM sleep) have been less consistent. 16 ,
17 , 19 Regional differences in power have also been discovered, with a
greater increase in delta waves observed in the posterior regions, which may
suggest slower reactivation of these areas after waking. 16 – 18 Further,
a recent study by Vallat et al 19 assessed the functional connectivity of
different brain regions post-awakening and observed that, compared to those
waking from lighter N2 sleep (non-REM stage 2 by Rechtschaffen & Kale
scoring), 20 participants waking from deeper N3 sleep (non-REM Stage 3 and
4) had higher connectivity between the default mode network and brain regions
responsible for sensory-motor control and attention. The authors posited that
this reduction of functional brain network segregation from the default mode
network, which is also observed during sleep and periods of elevated
sleepiness, 21 may be responsible for the physical and cognitive effects
observed during sleep inertia.
Studies of brain activity upon awakening have observed that cerebral blood
flow velocity is lower than pre-sleep levels for up to half an hour after
waking, reflecting the typical dissipation of cognitive effects. 22 – 24
Additional investigation of these changes discovered brain-region dependent
changes, with the prefrontal cortical regions (responsible for executive
function) taking longer to return to baseline levels. 23 , 25 Another
theory for the neurological basis of sleep inertia came from Van Dongen et
al's observation that sleep inertia is suppressed by caffeine consumed just
prior to the sleep opportunity. 26 Given caffeine acts as an adenosine
receptor antagonist, sleep inertia observed following awakenings in sleep
restriction scenarios may be partly due to left over adenosine stores that
have not been removed during sleep.
While there are few animal studies investigating the transition from sleep to
wake, Vyazovskiy et al 14 looked at neuronal activity in rodents following
spontaneous arousals from sleep. Their study found lower neuronal activity
after waking at a population level, but that individual neurons varied, with a
subset of neurons being silent for 1 min after waking. Further, the authors
observed that neuronal silence was higher following REM sleep compared to
waking from non-REM sleep.
Thus, the neurobehavioral impairments observed after waking appear to be due
to a delay in transitioning several independent neurophysiological processes
and that this transitional lag is most evident in regions related to higher
order processes necessary for cognitive tasks. 14 , 16 – 19 , 21 – 26
These transitions could be targeted in the development of novel interventions
to enhance the transition from sleep to a fully alert state.
## Contributing factors: the influence of sleep and circadian rhythms on
sleep inertia
There are several factors influencing the severity and duration of sleep
inertia. However, as is discussed at the end of this section, sleep inertia
can still occur in the absence of these exacerbating factors.
### Waking after prior sleep loss
Sleep inertia is worse under conditions of prior sleep loss. In studies
comparing sleep inertia following an 8-h sleep opportunity to partial sleep
deprivation, performance upon waking was significantly worse after the partial
sleep deprivation night. 27 , 28 Extended wakefulness prior to a recovery
sleep episode can also exacerbate the sleep inertia observed following
recovery sleep. 29 , 30 In a between-subjects design, Dinges et al 30
allocated a 2-h nap to participants after varying durations of prior
wakefulness (6, 18, 30, 42 and 54 hrs). Reaction times slowed and the number
of correct subtractions decreased as time awake prior to the nap increased. In
a within-subjects design, Rosa et al 29 also measured performance after a
2-h nap opportunity following either 16 hrs of wakefulness, or up to 64 hrs of
wakefulness, with worse performance observed after waking from the nap
following 64 hrs. Sleep inertia is also worsened by cumulative sleep loss.
Balkin and Badia’s 31 observation of increased sleep inertia effects across
four nights of disrupted sleep was recently supported by an in-laboratory
study in which participants were studied under conditions of chronic sleep
restriction (equivalent to sleep opportunities of 5.6 hrs per 24-h day).
Notably, compared to a control condition (equivalent to sleep opportunities of
8 hrs per 24-h day), participants undergoing chronic sleep restriction
experienced a 10% worsening of performance immediately upon awakening, with
average levels of performance failing to reach baseline levels at 70 mins post
awakening. 32 Together, these studies suggest that sleep loss, in the form
of restricted sleep, extended wakefulness, or cumulative sleep loss,
contributes to increased sleep inertia effects.
### Waking during the night
Sleep inertia effects are greatest during the biological night, near the
circadian low in core body temperature. 30 , 33 – 35 Using a protocol
designed to spread behaviors evenly across all hours of the 24-h day (ie,
forced desynchrony protocol), Scheer et al 33 found that circadian rhythms
significantly influenced the number of correct responses on an addition task
performed within two minutes of waking. In this study, amplitude in circadian
variation observed immediately after waking was greater than that observed at
later testing points. This finding suggests that circadian rhythms exert an
effect on sleep inertia, independent to its effect on background performance.
This effect has also been observed in older participants (>55 y). 34
While Scheer et al 33 independently assessed the effects of circadian
rhythms and sleep pressure on sleep inertia, Dinges et al 30 investigated
the interaction between the two processes. In their study, participants took a
2-h nap near the peak (approximately 15:00) or trough (approximately 03:00) of
the circadian cycle, following varying amounts of prior sleep loss (6–54 hrs
of prior wakefulness). This study found that during the peak in alertness, the
effect of prior sleep deprivation on sleep inertia appeared to be attenuated.
Meanwhile, during the trough, these effects were exacerbated. This interaction
between sleep loss, circadian timing, and performance during sleep inertia has
also been found under conditions of chronic sleep restriction. 32 The
results of these studies suggest that circadian rhythms have a direct effect
on sleep inertia, and also moderate the effects of sleep deprivation. This
interaction creates a non-linear trend in performance as sleep deprivation
increases.
Studies designed to systematically assess the impact of circadian rhythms on
sleep inertia show clear support for a time-of-day influence, with worse
performance immediately after waking during the circadian low. This effect has
also been demonstrated to be in addition to circadian effects on background
performance and to modulate the influence of homeostatic pressure.
### Waking from deep sleep
Mixed observations have been reported on whether the depth of sleep, or the
stage of sleep, at awakening has a significant effect on sleep inertia. The
increased amount of, and greater propensity to wake from, slow wave sleep
(SWS) under conditions of sleep pressure may be associated with the observed
increase in sleep inertia following sleep loss. Similarly, the observation
that sleep inertia is less likely to occur after short naps (≤30 mins) 36 –
38 may be due to the typical delay in SWS onset of 30 mins. 39 This section
explores the evidence for and against the influence of SWS on sleep inertia.
Perhaps the most frequently cited evidence for an effect of sleep depth on
sleep inertia is Dinges et al's 30 study of 2-h naps during 54 hrs of sleep
deprivation. Their study measured reaction time by the speed with which
participants answered a wake-up phone call and showed that increasing sleep
depth was associated with slower response speed. This was particularly evident
for awakenings from slow wave Stage 4 sleep (now classified as N3 under the
AASM scoring guidelines). 40 Furthermore, the amount of SWS during the
preceding nap was associated with worse performance on a descending
subtraction task performed immediately after answering the phone. Several
other studies have also observed sleep stage at awakening as a key predictor
of performance impairment upon waking. Stampi 41 reported that participants
waking from SWS showed a 41% reduction in performance upon awakening compared
to performance pre-nap, whereas participants waking from Stage 2 (N2) sleep
showed similar performance to those who were already awake. Sleep depth as
assessed by power spectral analysis showed that the amount of delta activity
(a marker of SWS) in the 10 mins prior to waking from a restricted sleep
episode was negatively correlated with performance immediately after waking.
27 Further, performance on a decision-making task was significantly worse
upon waking from SWS compared to REM sleep. 42 Together, the results from
the above studies suggest a positive relationship between greater sleep depth
and greater sleep inertia.
Many more-recent studies have also found an association between sleep depth
and sleep inertia. However, these studies were not necessarily designed to
systematically investigate this relationship and therefore rarely support
their observations with statistical analysis. 36 , 37 , 43 In
particular, studies of afternoon naps showing that the amount of SWS and the
severity of sleep inertia increase with nap length have not confirmed the role
of SWS with further analysis. 36 , 37 Regardless of the mechanisms
underlying the relationship between greater nap length and greater sleep
inertia, these observations lend support to the use of short naps in an effort
to minimize sleep inertia.
In contrast, some studies have reported no association between sleep depth 33
, 44 , 45 or sleep stages at awakening 33 , 45 , 46 and post-sleep
performance. This lack of association has been demonstrated at all times of
day under controlled prior sleep–wake conditions. 33 and following extended
wakefulness during the night and day. 45 This lack of association is
particularly prevalent in the napping literature (for review see Hilditch et
al, 2017 47 ). 45 , 48 – 50 Perhaps the most striking observation
comes from Lovato et al 48 who, despite 91% of participants waking from SWS
and over 50% of the prior sleep period containing SWS, did not observe any
performance impairment following a nap taken during a simulated night shift.
The debate in the literature as to whether sleep depth influences sleep
inertia may be due to variations in methodologies and definitions of sleep
depth. A direct comparison of the studies presented here, however, reveals
some common factors that may help to explain the apparent discrepancies. For
example, the effects of sleep depth on sleep inertia tend to be observed
following longer naps (2 hrs) 27 , 30 compared to shorter naps (20–60
mins), 45 and under higher homeostatic pressure 27 , 30 but rarely under
reduced homeostatic pressure. 27 , 33 , 51 , 52 These studies suggest
that the influence of sleep depth on sleep inertia may be mediated by the
lengths of both prior wakefulness and prior sleep. From the limited studies
reviewed, the effect of time of day does not appear to interact with this
relationship; 33 , 44 , 45 however, its direct influence on sleep
inertia 33 might still obscure measurement of the direct influence of sleep
depth. Thus, interactions with prior sleep–wake factors appear to change the
relative influence of sleep depth on subsequent performance, although a direct
investigation of these interactions has not been conducted.
Overall, it is difficult to synthesize a clear conclusion as to the role of
SWS in sleep inertia. There is long-standing evidence supporting the
association between greater sleep depth and greater sleep inertia. 27 , 30
, 41 This traditional view, however, is now being challenged by more recent
literature which suggests that this relationship may not be as robust as
initially thought. 33 , 45 Different study designs and measures of sleep
depth make it difficult to compare between studies. However, the current
literature suggests that the lengths of prior wakefulness and prior sleep may
influence the association between sleep depth and sleep inertia.
### Impact of contributing factors on sleep inertia duration
The majority of studies examining sleep inertia were not designed to directly
assess the duration of sleep inertia and therefore include too few data points
to make firm conclusions about the impact of contributing factors on the
duration of sleep inertia. In addition, most studies directly observing the
time course of sleep inertia have not directly compared contributing factors.
One study, however, did directly observe the duration of sleep inertia under
two different conditions. Achermann et al's study 44 observed that the time
course of sleep inertia following an 8-h nocturnal sleep episode and a 2-h
evening nap was the same, suggesting that circadian timing and sleep duration
under these conditions did not impact duration. Brooks and Lack 37 compared
four different short, afternoon nap lengths and found that while a 10-min nap
resulted in immediate performance improvements, a 30-min nap did not provide
improvements until 35 mins or up to 95 mins after waking, depending on the
task. This suggests that the duration of sleep inertia is dependent on both
length of nap and type of task. Comparing across studies, Hilditch et al 38
found that both a 10-min and 30-min nap terminated at 04:00 following acute
sleep loss provided no improvements to performance throughout the sleep
inertia testing period (up to 60 mins) nor across the remainder of the night
(up to 2.5 hrs). 53 Taken together, these studies suggest that circadian
timing and prior sleep-wake history influence sleep inertia duration as well
as severity, although the relative influence of these factors cannot be
determined from these observations.
### Sleep inertia in the absence of high homeostatic and circadian pressures
While the factors discussed above have been shown to exacerbate sleep inertia
effects, it is important to note that sleep inertia has been observed even in
the absence of these factors. For example, Wertz et al's 54 striking
demonstration of the impact of sleep inertia relative to 24 hrs of wakefulness
was observed in healthy participants, following an 8-h, habitually-timed sleep
opportunity. Similarly, studies have demonstrated sleep inertia effects
lasting at least one hour under unremarkable, well-rested conditions. 44 ,
51 Results from studies measuring performance and alertness after waking from
daytime naps also show signs of sleep inertia, even when the prior night of
sleep is unrestricted. 44 , 55 Scheer et al 33 demonstrated the
circadian influence on sleep inertia magnitude in non-sleep restricted
conditions, but also showed that sleep inertia is present, albeit less severe,
during times outside of the circadian low. Finally, Hilditch et al 50
observed performance impairment immediately following a 10-min nap taken at
07:00 following extended wakefulness, demonstrating that a short nap,
depending on the context, does not always avoid sleep inertia. Together, the
findings from these studies suggest that sleep inertia is a ubiquitous
phenomenon that, while exacerbated by certain factors, can potentially occur
at any time.
## Consequences: effects of sleep inertia on neurobehavioral outcomes
Despite its relatively short-acting effects, sleep inertia is a notable cause
of performance impairment and has been associated with severe, real-world
consequences.
### Time course of sleep inertia effects
Studies comparing sleep inertia to pre-sleep values have typically shown a
return to these levels within 30 mins of awakening 36 , 37 , 46 , 48 ,
56 and sometimes as soon as 15 mins after awakening. 45 , 52 , 57
Studies that have systematically measured alertness and performance across the
period after waking, however, report an asymptotic dissipation of sleep
inertia. 9 , 44 , 51 While the initial dissipation of impairment is
rapid, full recovery does not appear to be complete until at least an hour
after awakening. Jewett et al 51 investigated the time course of sleep
inertia in a constant routine protocol in which measures of subjective
alertness and cognitive throughput were taken regularly from one minute to
four hours after scheduled awakening. Under these conditions, Jewett et al 51
found, as in the findings of Folkard and Åkerstedt, 9 that subjective
alertness continued to improve for up to two hours after awakening.
Performance impairment on an addition task, however, took up to 3.5 hrs to
dissipate. 51 These tests were performed following a habitual morning
awakening, so the influence of the rise in circadian alertness across this
period cannot be extricated from this observation.
Interestingly, subjective alertness recovered faster than objective
performance in Jewett et al's 51 study, but was slower in Achermann et al's
44 study. Achermann et al 44 also reported that there was no correlation
between objective performance and subjective sleepiness. The difference in
time course in performance measures between the two studies may be explained
by differences in the tasks performed. However, the desynchronization of time
course between subjective and objective measures in both studies highlights a
concern when using self-assessment after waking, especially if alertness
recovers faster than cognitive performance. Subjective ratings of alertness
and performance have been shown to be inconsistent predictors of objective
performance under conditions of partial 58 and chronic 59 sleep loss.
Achermann et al's 44 study suggests that subjective ratings might also be a
poor indicator of performance across the dissipation of sleep inertia.
Hilditch et al 38 reported a self-rating scale of performance (as opposed to
alertness) across the dissipation of sleep inertia and found that despite
worse objective performance after waking from a 30-min night-time nap compared
to pre-nap, participants rated their performance as significantly better
during this period. These findings highlight the need to measure both
subjective and objective outcomes when investigating sleep inertia effects.
Furthermore, a recent study by Ritchie et al 60 suggests that a
participant’s morning or evening preference (chronotype) should also be
measured when estimating the time course of sleep inertia, with the
observation that later chronotypes took longer to recover from sleep inertia
than early types.
### Comparison of the effects of sleep inertia with the effects of sleep loss
Despite a relatively rapid recovery from sleep inertia in the first 15–30 mins
after waking, impairment during this initial period can be equivalent to, or
worse than, the effects of sleep loss. For example, Wertz et al 54 showed
that performance on an addition test immediately after waking was
significantly more impaired than after one night of sleep deprivation.
Similarly, Miccoli et al 28 found no difference in mean response times on a
10-min psychomotor vigilance test (PVT; a simple response time task) for the
first hour following a night of sleep deprivation compared to waking from a
night of restricted sleep. This indicates that any benefits of the three hours
of sleep in the partial sleep restriction condition were masked by sleep
inertia for at least an hour after waking. Remarkably, Rosa et al 29 found
that waking from two hours of recovery sleep following 64 hrs of sleep
deprivation was associated with worse performance on both a memory task and
auditory reaction time task compared to before the recovery sleep. Performance
after waking was also worse than at the same time of day during the prior
sleep deprivation period (ie after approximately 41 hrs of wakefulness).
In another study involving 64 hrs of sleep deprivation, a group given 20-min
naps every six hours across this period performed worse after waking than the
total sleep deprivation group. Furthermore, the sleep inertia experienced
after short naps was so intolerable that six participants in the nap condition
withdrew from the study, whereas all participants in the sleep deprivation
group completed the study. 61
Together, the findings from these studies highlight the potential severity of
sleep inertia performance deficits relative to extreme sleep loss.
Furthermore, these results suggest that, at least in the short term,
performance impairment after waking from recovery sleep can actually be worse
than the impairment caused by the prior sleep loss itself. Therefore,
following prior sleep loss, observable benefits from recovery sleep may be
delayed by sleep inertia for up to an hour after waking.
### Impact of sleep inertia on different measures of cognitive performance
As the sleep inertia literature expands, a debate has begun as to whether all
cognitive tasks are equally affected immediately after waking. Some studies
have found that, in contrast to the impairment observed after sleep
deprivation, only the reaction time or “speed” component of tasks is
negatively affected during sleep inertia. 28 , 46 , 52 , 55 , 61 ,
62 However, several other studies have found equal effects on both speed and
accuracy 27 , 31 , 45 or greater effects on accuracy. 63 Variations
between studies such as task type, time of testing, instructions to the
participants (eg instructed to perform as fast and/or as accurately as
possible), and the length and timing of sleep may account for these
discrepancies. Two studies have also claimed that, while overall average
response speed may slow as a result of sleep inertia, lapses, which represent
a substantial delay in response speed, are not a neurobehavioral feature of
sleep inertia, but rather are only associated with sleep loss induced
microsleeps. 28 , 52
It has also been argued that higher cognitive tasks that require greater
attentional load are more susceptible to the effects of sleep inertia than
simple tasks. 64 , 65 Studies have reported the effects of sleep inertia
on complex cognitive performance tasks such as memory, 44 calculations, 30
, 54 , 65 decision making, 42 , 66 and a spatial-configuration visual
search task. 65 However, sleep inertia has also been observed using simple
reaction time tasks. 26 , 45 , 62 Conflicting results have also been
obtained with regard to effect sizes across different tasks. Santhi et al 62
showed the largest effect sizes for simpler tasks such as PVT and a 1-back
working memory task compared to a 3-back working memory task. In another
study, medium effect sizes were found for moderately complex tasks such as
addition and digit-symbol substitution, but the largest effect size was
reported for cognitive throughput on a higher-order spatial-configuration
task. 65 Given the range of methodologies used across these studies, a clear
hypothesis for the differential effects observed across tasks has yet to be
put forward.
### Real-world impacts of sleep inertia
Sleep inertia is a challenge to workers who need to perform safety-critical
tasks, make important decisions, or operate a vehicle soon after waking. To
this end, several reviews of alertness management in operational settings have
highlighted the need to manage sleep inertia in order to maintain safety. 64
, 67 – 69
While some facets of cognition may be more affected by sleep inertia than
others, 62 , 65 real-world tasks often involve a combination of multiple
cognitive domains. For example, operating a vehicle safely requires
situational awareness, information processing, decision making, memory and, in
some instances, rapid response times. Studies of complex psychomotor tasks
that better emulate tasks in the real world, for example, a fire management
task, 42 a military enemy attack exercise, 66 or putting on a space suit,
8 have all been shown to be susceptible to the effects of sleep inertia.
Further, interviews with nurses 70 and surveys of emergency service pilots
71 have identified sleep inertia as a prevalent issue amongst on-call and
night shift workers. In the military, a retrospective analysis of more than
400 US Air Force accidents showed that accidents associated with pilot error
were most common during the first hour after waking, suggesting a potential
sleep inertia effect. 72 Sleep inertia has also been cited as a contributing
factor in several commercial incidents across multiple industries which have
resulted in damage, 73 , 74 injuries, 75 and deaths. 76
Taken together, these findings suggest that the effects of sleep inertia on
simple, complex, and operational tasks has the potential to negatively impact
upon safety-critical activities in the real world.
## Countermeasures to sleep inertia
Our knowledge of the factors influencing sleep inertia can help to develop
proactive strategies for managing sleep inertia, such as optimal sleep length
and timing of awakening. For example, based on the literature summarized
above, a planned awakening should take into account as many of the following
criteria as possible: limit the duration of the period of wakefulness before
the sleep episode; minimize the amount of sleep loss prior to the sleep
episode – both acute and chronic; avoid waking during the circadian low in
alertness (biological night); and, if waking from a nap, limit sleep duration
to less than 30 mins. While following these guidelines can reduce the risk of
sleep inertia, as discussed earlier, they do not guarantee a sleep inertia-
free wake up. Thus, the most effective proactive countermeasure appears to be
caffeine. When taken before a short nap (eg 20 mins), caffeine has been shown
to alleviate the symptoms of sleep inertia following the nap. 26 , 77 The
limitation of these proactive strategies is that they often require a planned
sleep opportunity, a designated wake time, and control over prior sleep-wake
history. For many on-call shift workers it is not always feasible to plan the
length and timing of a sleep opportunity, and in many cases prior sleep-wake
history may be poor. Thus, there is a need for countermeasures that can be
implemented upon waking (“reactive” countermeasures).
Hilditch et al 78 recently reviewed the literature investigating reactive
countermeasures and concluded that there is currently no clear empirical
evidence to fully support the use of any reactive countermeasure to provide
immediate and objective effects. As mentioned above, when administered before
sleep, caffeine has been shown to eliminate the effects of sleep inertia. 26
However, there are several limitations to the effectiveness and application of
this countermeasure in a reactive scenario. When administered after sleep,
even in a rapidly-absorbed chewing gum format, the effects of caffeine are
delayed such that while the duration of sleep inertia may be truncated, the
initial, most severe period of effects are unaffected by caffeine. 79
Furthermore, while caffeine is indeed a field-deployable and operationally
viable countermeasure in many cases, the relatively long-lasting stimulant
effects may be unwanted in situations in which it is preferable for the worker
to fall back asleep within a few hours of waking. 80
Other countermeasures such as light, sound, and temperature have been
investigated. The outcomes of the trials, however, have been either
unsuccessful, or uninterpretable due to methodological limitations such as
lack of a control group, not enough testing points, or no evidence of sleep
inertia in the control condition. 81 – 86 We below summarize findings of
the most commonly investigated countermeasures.
To date, two studies have investigated the use of brief 81 and sustained 82
light exposure after waking to reduce sleep inertia. Bright light exposure has
been shown to directly improve alertness and cognitive performance during the
day, night and following sleep restriction. 87 Therefore, there is potential
for bright light to improve alertness and performance during the sleep inertia
period. One study reported a significant improvement to subjective alertness,
however, neither study observed a significant improvement on objective
performance measures. While these results suggest that both brief and
sustained light exposure after waking is of limited effectiveness in reducing
sleep inertia effects, it is worth noting that the exposures in these studies
were during the day (~07:00 and 13:00). The use of light during nocturnal
awakenings may, therefore, have a different effect.
Noise can promote arousal and has previously been shown to attenuate hypo-
vigilance during sleep deprivation. 88 , 89 Early investigations on the
use of sound to reduce sleep inertia effects have been promising. Tassi et al
83 exposed participants to pink noise after a 1-h nap at 01:00 and observed
that pink noise eliminated the sleep inertia effect observed in the no-noise
group. This effect was less obvious when tested at 04:00. The sleep stage at
waking was not controlled in this study and may have contributed to the mixed
results at different test times. Hayashi et al 84 took a different approach,
playing music after waking from a short afternoon nap. While playing music has
not been shown to have a long-term alerting effect, 90 its short-term
effects may be useful in the context of sleep inertia. Indeed, the researchers
reported that music reduced subjective sleepiness, and that music preferred by
the participants led to improved cognitive performance for up to 20 mins after
waking. Sound may be an operationally viable (ie delivered through headphones)
and relatively brief and immediate alerting strategy for use in the field.
While the relationship of body temperature to sleep onset has been extensively
investigated, its relationship to sleep offset has received less attention.
Some studies have shown, however, that changes in the distal-proximal
temperature gradient (DPG) after waking correlate with subjective sleepiness.
85 This relationship has been demonstrated across different circadian phases
in a multi-nap protocol but has yet to be tested with objective performance
measures. 86 It has been proposed that cooling the extremities immediately
after waking may accelerate the DPG changes and, in turn, accelerate recovery
from sleep inertia effects. This theory has yet to be tested with an
intervention study. Manipulating body temperature may also be a useful
deployable strategy, the effects of which may be reversible in situations
where a sleep opportunity is presented later in the night.
While there are no studies to our knowledge that have investigated the
effectiveness of exercise to reduce sleep inertia effects, anecdotally this is
an avenue that may be worthy of research. Sleep inertia is associated with a
slow return to waking levels of cerebral blood flow in the anterior cortical
regions of the brain; 23 therefore, moderate cardiac activity may accelerate
this process. Implementation of this strategy would also be relatively
convenient in most work place settings.
Given the potential for catastrophic events associated with sleep inertia, the
need to develop preventative strategies and reactive countermeasures to
minimize both the severity and duration of sleep inertia effects in safety-
critical scenarios is crucial.
## Summary
As discussed in the previous sections, sleep inertia is characterized by
impaired performance and reduced alertness immediately after waking. These
effects dissipate asymptotically with the most significant effects occurring
within 30 mins of waking. Sleep inertia interacts with the homeostatic and
circadian processes to influence performance immediately after waking.
Evidence suggests that waking after acute or chronic prior sleep loss, during
the circadian low, or from deeper stages of sleep can exacerbate sleep
inertia. Sleep inertia is of great importance as the associated performance
impairment can be equivalent to, or greater than, that observed after up to 40
hrs of sleep deprivation. Sleep inertia effects have been observed on a range
of tasks from simple reaction time tests to complex cognitive tasks. While
field studies of sleep inertia are lacking, evidence of sleep inertia in real-
world scenarios has been demonstrated in several operational incidents, often
with catastrophic consequences. While there is evidence that caffeine can help
to reduce the impact of sleep inertia, there is a need for further research
into reactive countermeasures that can be deployed in operational scenarios.
## Acknowledgment
Dr. Hilditch is supported by a NASA System Wide Safety grant. Dr. McHill is
supported by NIH K01HL146992 and the Oregon Institute of Occupational Health
Sciences at Oregon Health & Science University. The funders had no role in
study design, data collection and analysis, decision to publish, or
preparation of the manuscript.
## Disclosure
Dr. McHill reports speaker honorarium or travel reimbursement fees from the
Utah Sleep Research Society and the California Precast Concrete Association.
The authors report no other conflicts of interest in this work.
## References
1\. Kleitman N. _Sleep and Wakefulness_ . Chicago: University of Chicago
Press; 1939.
2\. Lubin A, Hord DJ, Tracy ML, Johnson LC. Effects of exercise, bedrest and
napping on performance decrement during 40 hours. _Psychophysiology_ .
1976;13(4):334–339.
3\. Broughton RJ. Sleep disorders: disorders of arousal? Enuresis,
somnambulism, and nightmares occur in confusional states of arousal, not in
“dreaming sleep”. _Science_ . 1968;159(3819):1070–1078.
doi:10.1126/science.159.3819.1070
4\. Langdon DE, Hartman B _Performance upon sudden awakening. Technical
Report SAM-TDR_ . Brooks AFB, TX: USAF School of Aerospace Medicine; November
1961.
5\. Hartman BO, Langdon DE _A Second Study on Performance Upon Sudden
Awakening. Technical Report SAM-TR-65-61_ . Brooks AFB, TX: USAF School of
Aerospace Medicine; August 1965.
6\. Hartman BO, Langdon DE, McKenzie RE _A Third Study on Performance Upon
Sudden Awakening. Technical Report SAM-TR-65-63_ . Brooks AFB, TX: USAF School
of Aerospace Medicine; August 1965.
7\. Webb WB, Agnew H. Reaction time and serial response efficiency on arousal
from sleep. _Percept Mot Skills_ . 1964;18(3):783–784.
doi:10.2466/pms.1964.18.3.783
8\. Seminara JL, Shavelson RJ. Effectiveness of space crew performance
subsequent to sudden sleep arousal. _Aerosp Med_ . 1969;40(7):723–727.
9\. Folkard S, Åkerstedt T. A three-process model of the regulation of
alertness-sleepiness. In: Broughton RJ, Ogilvie RD, editors. _Sleep, Arousal,
and Performance_ . Boston: Birkhäuser; 1992:11–26.
10\. Borbély AA. A two process model of sleep regulation. _Hum Neurobiol_ .
1982;1(3):195–204.
11\. Achermann P. The two-process model of sleep regulation revisited. _Aviat
Space Environ Med_ . 2004;75(Supplement 1):A37–A43.
12\. Turek F. Introduction: master circadian clock and master circadian
rhythm. In: Kryger MH, Roth T, Dement WC, editors. _Principles and Practice of
Sleep Medicine_ . 5th ed. St Louis, MO: Elsevier Saunders; 2011:360–362.
13\. Peter-Derex L, Magnin M, Bastuji HJN. Heterogeneity of arousals in human
sleep: a stereo-electroencephalographic study. _NeuroImage_ .
2015;123:229–244. doi:10.1016/j.neuroimage.2015.07.057
14\. Vyazovskiy VV, Cui N, Rodriguez AV, Funk C, Cirelli C, Tononi G. The
dynamics of cortical neuronal activity in the first minutes after spontaneous
awakening in rats and mice. _Sleep_ . 2014;37(8):1337–1347.
doi:10.5665/sleep.3926
15\. Dijk D-J, Czeisler CA. Paradoxical timing of the circadian rhythm of
sleep propensity serves to consolidate sleep and wakefulness in humans.
_Neurosci Lett_ . 1994;166(1):63–68. doi:10.1016/0304-3940(94)90841-9
16\. Ferrara M, Curcio G, Fratello F, et al. The electroencephalographic
substratum of the awakening. _Behav Brain Res_ . 2006;167(2):237–244.
doi:10.1016/j.bbr.2005.09.012
17\. Marzano C, Ferrara M, Moroni F, De Gennaro L. Electroencephalographic
sleep inertia of the awakening brain. _Neuroscience_ . 2011;176:308–317.
doi:10.1016/j.neuroscience.2010.12.014
18\. Gorgoni M, Ferrara M, D’Atri A, et al. EEG topography during sleep
inertia upon awakening after a period of increased homeostatic sleep pressure.
_Sleep Med_ . 2015;16(7):883–890. doi:10.1016/j.sleep.2015.03.009
19\. Vallat R, Meunier D, Nicolas A, Ruby P. Hard to wake up? The cerebral
correlates of sleep inertia assessed using combined behavioral, EEG and fMRI
measures. _NeuroImage_ . 2019;184:266–278.
doi:10.1016/j.neuroimage.2018.09.033
20\. Rechtschaffen A, Kales A. _A Manual of Standardized Terminology,
Techniques and Scoring System for Sleep Stages of Human Subjects_ . Los
Angeles: Brain Information Service/Brain Research Institute, University of
California; 1968.
21\. Picchioni D, Duyn JH, Horovitz SG. Sleep and the functional connectome.
_NeuroImage_ . 2013;80:387–396. doi:10.1016/j.neuroimage.2013.05.067
22\. Hajak G, Klingelhofer J, Schulz-Varszegi M, et al. Relationship between
cerebral blood flow velocities and cerebral electrical activity in sleep.
_Sleep_ . 1994;17(1):11–19. doi:10.1093/sleep/17.1.11
23\. Balkin TJ, Braun AR, Wesensten NJ, et al. The process of awakening: a
PET study of regional brain activity patterns mediating the re‐establishment
of alertness and consciousness. _Brain_ . 2002;125(10):2308–2319.
doi:10.1093/brain/awf228
24\. Kuboyama T, Hori A, Sato T, Mikami T, Yamaki T, Ueda S. Changes in
cerebral blood flow velocity in healthy young men during overnight sleep and
while awake. _Electroencephalogr Clin Neurophysiol_ . 1997;102(2):125–131.
doi:10.1016/s0921-884x(96)95054-7
25\. Sakai F, Meyer JS, Karacan I, Derman S, Yamamoto M. Normal human sleep:
Regional cerebral hemodynamics. _Ann Neurol_ . 1980;7(5):471–478.
doi:10.1002/ana.410070514
26\. Van Dongen HPA, Price NJ, Mullington JM, Szuba MP, Kapoor SC, Dinges DF.
Caffeine eliminates psychomotor vigilance deficits from sleep inertia. _Sleep_
. 2001;24(7):813–819. doi:10.1093/sleep/24.7.813
27\. Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A. EEG
spectral power and cognitive performance during sleep inertia: the effect of
normal sleep duration and partial sleep deprivation. _Physiol Behav_ .
2006;87(1):177–184. doi:10.1016/j.physbeh.2005.09.017
28\. Miccoli L, Versace F, Koterle S, Cavallero C. Comparing sleep-loss
sleepiness and sleep inertia: lapses make the difference. _Chronobiol Int_ .
2008;25(5):725–744. doi:10.1080/07420520802397228
29\. Rosa RR, Bonnet MH, Warm JS. Recovery of performance during sleep
following sleep deprivation. _Psychophysiology_ . 1983;20(2):152–159.
30\. Dinges D, Orne M, Orne E. Assessing performance upon abrupt awakening
from naps during quasi-continuous operations. _Behav Res Methods_ .
1985;17(1):37–45. doi:10.3758/BF03200895
31\. Balkin TJ, Badia P. Relationship between sleep inertia and sleepiness:
cumulative effects of four nights of sleep disruption/restriction on
performance following abrupt nocturnal awakenings. _Biol Psychol_ .
1988;27(3):245–258. doi:10.1016/0301-0511(88)90034-8
32\. McHill AW, Hull JT, Cohen DA, Wang W, Czeisler CA, Klerman EB. Chronic
sleep restriction greatly magnifies performance decrements immediately after
awakening. _Sleep_ . 2019;42. doi:10.1093/sleep/zsz032
33\. Scheer FA, Shea TJ, Hilton MF, Shea SA. An endogenous circadian rhythm
in sleep inertia results in greatest cognitive impairment upon awakening
during the biological night. _J Biol Rhythms_ . 2008;23(4):353–361.
doi:10.1177/0748730408318081
34\. Silva EJ, Duffy JF. Sleep inertia varies with circadian phase and sleep
stage in older adults. _Behav Neurosci_ . 2008;122(4):928–935.
doi:10.1037/0735-7044.122.4.928
35\. Hilditch CJ, Short M, Van Dongen HP, et al. Sleep inertia during a
simulated 6-h on/6-h off fixed split duty schedule. _Chronobiol Int_ .
2016;33(6):685–696. doi:10.3109/07420528.2016.1167724
36\. Tietzel AJ, Lack LC. The short-term benefits of brief and long naps
following nocturnal sleep restriction. _Sleep_ . 2001;24(3):293–300.
doi:10.1093/sleep/24.3.293
37\. Brooks A, Lack L. A brief afternoon nap following nocturnal sleep
restriction: which nap duration is most recuperative? _Sleep_ .
2006;29(6):831–840. doi:10.1093/sleep/29.6.831
38\. Hilditch CJ, Centofanti SA, Dorrian J, Banks S. A 30-minute, but not a
10-minute nighttime nap is associated with sleep inertia. _Sleep_ .
2016;39(3):675–685. (in press). doi:10.5665/sleep.5550
39\. Gillberg M, Åkerstedt T. The dynamics of the first sleep cycle. _Sleep_
. 1991;14(2):147–154.
40\. Iber C, Ancoli-Israel S, Chesson AJ, Quan S; for the American Academy of
Sleep Medicine. _The AASM Manual for the Scoring of Sleep and Associated
Events: Rules, Terminology and Technical Specifications_ . Westchester, IL:
American Academy of Sleep Medicine (AASM); 2007.
41\. Stampi C. The effects of polyphasic and ultrashort sleep schedules. In:
Stampi C, editor. _Why We Nap: Evolution, Chronobiology and Functions of
Polyphasic and Ultrashort Sleep_ . Boston: Birkhäuser; 1992:137–179.
42\. Bruck D, Pisani DL. The effects of sleep inertia on decision-making
performance. _J Sleep Res_ . 1999;8(2):95–103.
43\. Ferrara M, De Gennaro L, Bertini M. The effects of slow-wave sleep (SWS)
deprivation and time of night on behavioral performance upon awakening.
_Physiol Behav_ . 1999;68(1–2):55–61. doi:10.1016/s0031-9384(99)00150-x
44\. Achermann P, Werth E, Dijk D-J, Borbély AA. Time course of sleep inertia
after nighttime and daytime sleep episodes. _Arch Ital Biol_ .
1995;134(1):109–119.
45\. Signal TL, van den Berg MJ, Mulrine HM, Gander PH. Duration of sleep
inertia after napping during simulated night work and in extended operations.
_Chronobiol Int_ . 2012;29(6):769–779. doi:10.3109/07420528.2012.686547
46\. Salame P, Otzenberger H, Ehrhart J, et al. Effects of sleep inertia on
cognitive performance following a 1-hour nap. _Work Stress_ .
1995;9(4):528–539. doi:10.1080/02678379508256898
47\. Hilditch CJ, Dorrian J, Banks S. A review of short naps and sleep
inertia: do naps of 30 min or less really avoid sleep inertia and slow-wave
sleep? _Sleep Med Rev_ . 2017;32:176–190. doi:10.1016/j.sleep.2016.12.016
48\. Lovato N, Lack L, Ferguson S, Tremaine R. The effects of a 30-min nap
during night shift following a prophylactic sleep in the afternoon. _Sleep
Biol Rhythms_ . 2009;7(1):34–42. doi:10.1111/j.1479-8425.2009.00382.x
49\. Kubo T, Takahashi M, Takeyama H, et al. How do the timing and length of
a night-shift nap affect sleep inertia? _Chronobiol Int_ .
2010;27(5):1031–1044. doi:10.3109/07420528.2010.489502
50\. Hilditch CJ, Dorrian J, Centofanti SA, Van Dongen HP, Banks S. Sleep
inertia associated with a 10-min nap before the commute home following a night
shift: a laboratory simulation study. _Accid Anal Prev_ . 2017;99:411–415.
doi:10.1016/j.aap.2015.11.010
51\. Jewett ME, Wyatt JK, Ritz-De Cecco A, Khalsa SB, Dijk D-J, Czeisler CA.
Time course of sleep inertia dissipation in human performance and alertness.
_J Sleep Res_ . 1999;8(1):1–8.
52\. Ikeda H, Hayashi M. The effect of self-awakening from nocturnal sleep on
sleep inertia. _Biol Psychol_ . 2010;83(1):15–19.
doi:10.1016/j.biopsycho.2009.09.008
53\. Centofanti SA, Hilditch CJ, Dorrian J, Banks S. The impact of short
night-time naps on performance, sleepiness and mood during a simulated night
shift. _Chronobiol Int_ . 2016;33(6):706–715.
doi:10.3109/07420528.2016.1167722
54\. Wertz AT, Ronda JM, Czeisler CA, Wright KP Jr. Effects of sleep
inertia on cognition. _JAMA_ . 2006;295(2):163–164. doi:10.1001/jama.295.2.163
55\. Hofer-Tinguely G, Achermann P, Landolt HP, et al. Sleep inertia:
performance changes after sleep, rest and active waking. _Brain Res Cogn Brain
Res_ . 2005;22(3):323–331. doi:10.1016/j.cogbrainres.2004.09.013
56\. Tremaine R, Dorrian J, Lack L, et al. The relationship between
subjective and objective sleepiness and performance during a simulated night-
shift with a nap countermeasure. _Appl Ergon_ . 2010;42(1):52–61.
doi:10.1016/j.apergo.2010.04.005
57\. Kaida K, Nittono H, Hayashi M, Hori T. Effects of self-awakening on
sleep structure of a daytime short nap and on subsequent arousal levels.
_Percept Mot Skills_ . 2003;97(3 Pt 2):1073–1084.
doi:10.2466/pms.2003.97.3f.1073
58\. Biggs SN, Smith A, Dorrian J, et al. Perception of simulated driving
performance after sleep restriction and caffeine. _J Psychosom Res_ .
2007;63(6):573–577. doi:10.1016/j.jpsychores.2007.06.017
59\. Van Dongen HPA, Maislin G, Mullington JM, Dinges DF. The cumulative cost
of additional wakefulness: dose-response effects on neurobehavioral functions
and sleep physiology from chronic sleep restriction and total sleep
deprivation. _Sleep_ . 2003;26(2):117–126. doi:10.1093/sleep/26.2.117
60\. Ritchie HK, Burke TM, Dear TB, McHill AW, Axelsson J, Wright KP Jr .
Impact of sleep inertia on visual selective attention for rare targets and the
influence of chronotype. _J Sleep Res_ . 2017;26(5):551–558.
doi:10.1111/jsr.12525
61\. Naitoh P, Kelly T, Babkoff H. Sleep inertia: best time not to wake up?
_Chronobiol Int_ . 1993;10(2):109–118.
62\. Santhi N, Groeger JA, Archer SN, Giménez M, Schlangen LJM, Dijk D-J.
Morning sleep inertia in alertness and performance: effect of cognitive domain
and white light conditions. _PloS One_ . 2013;8(11):e79688.
doi:10.1371/journal.pone.0079688
63\. Ferrara M, De Gennaro L, Casagrande M, Bertini M. Selective slow-wave
sleep deprivation and time-of-night effects on cognitive performance upon
awakening. _Psychophysiology_ . 2000;37(4):440–446.
64\. Muzet A, Nicolas A, Tassi P, Dewasmes G, Bonneau A. Implementation of
napping in industry and the problem of sleep inertia. _J Sleep Res_ .
1995;4(S2):67–69.
65\. Burke TM, Scheer FA, Ronda JM, Czeisler CA, Wright KP. Sleep inertia,
sleep homeostatic and circadian influences on higher-order cognitive
functions. _J Sleep Res_ . 2015;24(4):364–371. doi:10.1111/jsr.12291
66\. Horne J, Moseley R. Sudden early-morning awakening impairs immediate
tactical planning in a changing ‘emergency’ scenario. _J Sleep Res_ .
2011;20(2):275–278. doi:10.1111/j.1365-2869.2010.00904.x
67\. Rosekind MR, Smith RM, Miller DL, et al. Alertness management: strategic
naps in operational settings. _J Sleep Res_ . 1995;4(S2):62–66.
68\. Ferrara M, De Gennaro L. The sleep inertia phenomenon during the sleep-
wake transition: theoretical and operational issues. _Aviat Space Environ Med_
. 2000;71(8):843–848.
69\. Caldwell JA, Prazinko BF, Caldwell JL _Fatigue in Aviation Sustained
Operations, the Utility of Napping, and the Problem of Sleep Inertia. Report_
No. _RTO-EN-016_ . Neuilly-sur-Seine Cedex: North Atlantic Treaty Organisation
(NATO); 2002.
70\. Fallis WM, McMillan DE, Edwards MP. Napping during night shift:
practices, preferences, and perceptions of critical care and emergency
department nurses. _Crit Care Nurse_ . 2011;31(2):e1–e11.
doi:10.4037/ccn2011710
71\. Gregory KB, Winn W, Johnson K, Rosekind MR. Pilot fatigue survey:
exploring fatigue factors in air medical operations. _Air Med J_ .
2010;29(6):309–319. doi:10.1016/j.amj.2010.07.002
72\. Ribak J, Ashkenazi IE, Klepfish A, et al. Diurnal rhythmicity and air
force flight accidents due to pilot error. _Aviat Space Environ Med_ .
1983;54(12 Pt 1):1096–1099.
73\. Marine Accident Investigation Branch. _Heavy Contact by Skandi Foula
with OMS Resolution, Aberdeen Harbour 29 May 2010. Accident Report No.
15/2011_ . London: UK Department for Transport: UK Department for Transport;
2011.
74\. Armentrout JJ, Holland DA, O’Toole KJ, Ercoline WR. Fatigue and related
human factors in the near crash of a large military aircraft. _Aviat Space
Environ Med_ . 2006;77(9):963–970.
75\. Transportation Safety Board of Canada. _Pitch Excursion: Air Canada,
Boeing 767-333, C-GHLQ, North Atlantic Ocean, 55°00ʹN 029°00ʹW, 14 January
2011. Aviation Investigation Report No. A11F0012_ . Gatineau: Government of
Canada; 2011:A11F0012.
76\. Government of India Ministry of Civil Aviation. _Report on Accident to
Air India Express Boeing 737-800 Aircraft VT-AXV on 22nd May 2010 at
Mangalore_ . New Delhi: Government of India; 2010.
77\. Gray DM, Banks S, Dorrian J, Munro E, Nielsen J, Centofanti S. Combining
caffeine and a nap to improve alertness during a simulated nightshift. _Sleep_
. 2019;42(Supplement 1):A78–A78. doi:10.1093/sleep/zsz067.190
78\. Hilditch CJ, Dorrian J, Banks S. Time to wake up: reactive
countermeasures to sleep inertia. _Ind Health_ . 2016;54:2015–2236.
79\. Newman RA, Kamimori GH, Wesensten NJ, Picchioni D, Balkin TJ. Caffeine
gum minimizes sleep inertia. _Percept Mot Skills_ . 2013;116(1):280–293.
doi:10.2466/29.22.25.PMS.116.1.280-293
80\. McHill AW, Smith BJ, Wright KP. Effects of caffeine on skin and core
temperatures, alertness, and recovery sleep during circadian misalignment. _J
Biol Rhythms_ . 2014;29(2):131–143. doi:10.1177/0748730414523078
81\. Hayashi M, Masuda A, Hori T. The alerting effects of caffeine, bright
light and face washing after a short daytime nap. _Clin Neurophysiol_ .
2003;114(12):2268–2278.
82\. Santhi N, Groeger JA, Archer SN, Giménez M, Schlangen LJ, Dijk DJ.
Morning sleep inertia in alertness and performance: effect of cognitive domain
and white light conditions. _PloS One_ . 2013;8(11):e79688.
doi:10.1371/journal.pone.0079688
83\. Tassi P, Nicolas A, Dewasmes G, et al. Effects of noise on sleep inertia
as a function of circadian placement of a one-hour nap. _Percept Mot Skills_ .
1992;75(1):291–302. doi:10.2466/pms.1992.75.1.291
84\. Hayashi M, Uchida C, Shoji T, Hori T. The effects of the preference for
music on sleep inertia after a short daytime nap. _Sleep Biol Rhythms_ .
2004;2(3):184–191. doi:10.1111/j.1479-8425.2004.00142.x
85\. Kräuchi K, Cajochen C, Wirz-Justice A. Waking up properly: is there a
role of thermoregulation in sleep inertia? _J Sleep Res_ . 2004;13(2):121–127.
doi:10.1111/j.1365-2869.2004.00398.x
86\. Kräuchi K, Knoblauch V, Wirz-Justice A, Cajochen C. Challenging the
sleep homeostat does not influence the thermoregulatory system in men:
evidence from a nap vs. sleep-deprivation study. _Am J Physiol Regul Integr
Comp Physiol_ . 2006;290(4):R1052–R1061. doi:10.1152/ajpregu.00381.2005
87\. Cajochen C. Alerting effects of light. _Sleep Med Rev_ .
2007;11(6):453–464. doi:10.1016/j.smrv.2007.07.009
88\. Wilkinson RT. Interaction of noise with knowledge of results and sleep
deprivation. _J Exp Psychol_ . 1963;66(4):332–337.
89\. Poulton EC. Arousing environmental stresses can improve performance,
whatever people say. _Aviat Space Environ Med_ . 1976;47(11):1193–1204.
90\. Reyner LA, Horne JA. Evaluation of “in-car” countermeasures to
sleepiness: cold air and radio. _Sleep_ . 1998;21(1):46–50.

© 2019 The Author(s). This work is published and licensed by Dove Medical
Press Limited. The full terms of this license are available at [
https://www.dovepress.com/terms.php ](https://www.dovepress.com/terms.php) and
incorporate the [ Creative Commons Attribution - Non Commercial (unported,
v3.0) License ](//creativecommons.org/licenses/by-nc/3.0/) . By accessing the
work you hereby accept the Terms. Non-commercial uses of the work are
permitted without any further permission from Dove Medical Press Limited,
provided the work is properly attributed. For permission for commercial use of
this work, please see paragraphs 4.2 and 5 of [ our Terms
](https://www.dovepress.com/terms.php) .
[ Download Article [PDF] ](getfile.php?fileID=52180)
[ Contact Us ](contact) • [ Privacy Policy ](https://www.informa.com/privacy-
policy/) • [ Associations & Partners ](partners.php) • [ Testimonials ](why-
publish-with-dove-press/testimonials) • [ Terms & Conditions ](terms.php) •
Recommend this site • [ Cookies ](cookies.php) • Top
[ Contact Us ](contact) • [ Privacy Policy ](privacy.php)
**© Copyright 2024** • **Dove Press Ltd** • software development by
maffey.com • Web Design by Adhesion
The opinions expressed in all articles published here are those of the
specific author(s), and do not necessarily reflect the views of Dove Medical
Press Ltd or any of its employees.
Dove Medical Press is part of Taylor & Francis Group, the Academic Publishing
Division of Informa PLC
Copyright 2017 Informa PLC. All rights reserved. This site is owned and
operated by Informa PLC ( “Informa”) whose registered office is 5 Howick
Place, London SW1P 1WG. Registered in England and Wales. Number 3099067. UK
VAT Group: GB 365 4626 36
*[
2 April 2019
]: These are actual dates the paper was submitted to, accepted for, and published in the journal.
| biology | 12917 | https://no.wikipedia.org/wiki/S%C3%B8vn | Søvn | Søvn er tilstanden som mennesker og dyr periodisk inntar for å hvile; en tilstand med sterkt nedsatt bevissthet.
Under søvn skjer det flere fysiologiske forandringer i kroppen:
puls, stoffskifte og kroppstemperatur reduseres
skjelettmusklene slapper av
blodårene utvides
øynene lukkes
Søvn reguleres av døgnrytmen, av homeostase (tiden som er gått siden den siste adekvate søvnen) og, hos mennesker, av villet atferd.
Det er fremdeles usikkert hva som skjer under søvnen som gjør den til en livsnødvendighet, men alle dyr som er blitt forsket på, dør når de holdes fra å sove. Rotter, for eksempel, lever ikke lenger enn ca. 14 dager uten søvn.
Det er blitt antatt at hjernen under søvn kvitter seg med skadelige rester av stoffskiftet, at skadde/svekkede hjerneceller blir reparert og at dagens minner blir bearbeidet og enten lagret i hukommelsen eller «forkastet». Men dette er kun fremlagte ideer.
Søvnen opptrer i to hovedstadier, NREM og REM, som inntreffer med regelmessige intervaller. REM står for rapid eye movement, og NREM betyr ikke-REM. Man drømmer som oftest i REM-stadiet, men forskning ved bruk av EEG-målinger viser at man også kan drømme i NREM-stadiene. NREM-søvn deles opp i fire stadier, to lette (1 og 2) og to dype (3 og 4), som blir etterfulgt av en periode med REM-søvn. Rekkefølgen er 1 – 2 – 3 – 4 – 3 – 2 – REM for en syklus på 90 til 110 minutter. Syklusen gjentas utover natten, som oftest 4 – 5 ganger, med større porsjoner NREM den første halvdelen av natten og mer REM fram mot morgenen.
Stadium-1 er lett søvn (theta-bølger) og tildels likner våken tilstand. Stadium-2 er noe dypere. Stadium-3 (delta-bølger) og -4 er dyp søvn, og det er i dette stadiet svært vanskelig å vekke individet. Tendensen er å slå sammen stadium-3 og -4. Stadium-3 har 20 – 50 % delta-bølger mens stadium 4 inneholder mer enn 50 % delta-bølger.
I REM-søvn økes hjernens aktivitet drastisk, og den jobber faktisk hardere enn i våken tilstand. Skjelettmuskulaturen blir paralysert, og øynene kan bevege seg rundt som om de ser etter noe i mørket under øyelokkene.
Som hos alle arter der nyfødte ikke er fullt utviklet, har småbarn mye REM-søvn. Nyfødte sover opptil 18 timer i døgnet, ca. 50 % REM. Antall timer voksne mennesker sover er helt individuelt; omfattende studier viser at det selvrapporterte gjennomsnittet er 7,5 timer pr. døgn. Eldre mennesker trenger like mye søvn som da de var yngre, men den er ofte mer oppdelt.
Søvnmangel
Søvnmangel over kortere perioder —som ett døgn— har ved forsøk vist tendenser til redusert kortidshukommelse, trøtthet, og redusert oppmerksomhet. Uteblir søvn over flere døgn, svekkes individet ytterligere og kan få store konsentrasjonsproblemer, hallusinasjoner og paranoia. Virkningen kan sammenlignes med beruselse.
Barn bør sove minst 9 timer hver dag. Voksne derimot, bør sove 7 eller 8 timer. Disse påstandene støttes av The National Sleep Foundation (NSF) i USA; en uavhengig, frivillig nullprofittorganisasjon som har som mål å forbedre offentlig helse og sikkerhet ved å oppnå forståelse av søvn og søvnforstyrrelser, og å støtte søvnrelatert utdanning, forskning og forståelse. Mange søvneksperter er uenig i at åtte-timers-behovet gjelder generelt. Isteden er den beste søvnlengden mellom 6,5 og 7,5 timer. Kanskje har timetallet åtte oppstått som et retorisk eller pedagogisk grep ved en tredeling av døgnet i «åtte timer arbeid, åtte timer (religiøse) studier og åtte timer søvn» fra jødenes forskrifter i talmud og blitt forsterket av arbeiderbevegelsens slagord «åtte timer arbeid, åtte timer fritid og åtte timer hvile».
Randy Gardner holder verdensrekorden (Guinness rekordbok) for å holde seg våken lengst. I 1965 holdt syttenåringen seg våken i vel ti døgn i sammenheng med et skoleprosjekt. Han ble alvorlig mentalt svekket under forsøket, men var normal igjen etter få dager. Ved slutten av forsøket klarte likevel Gardener å slå forsøkslederen i actionspillet pinball. Det blir ikke nye rekordholdere; Guinness vil ikke godkjenne flere, på grunn av helsefaren. På grunn av dette finnes det ikke lenger noen offisiell rekordholder, bortsett fra Randy Gardner, men briten Tony Wright holder den uoffisielle rekorden etter å ha holdt seg våken i mer enn 264 timer (11 døgn) i mai 2007.
Søvndeprivasjon blir også benyttet som en torturmetode.
Søvnforstyrrelser
Det er mange grunner til inadekvat søvn, både blant barn og voksne, både akutte og kroniske. Pasienter kan oppleve å få for lite søvn, søvn til feil tid (døgnrytmeforstyrrelser), eller søvn av for dårlig kvalitet. Leger vil ofte først foreslå søvnhygieniske tiltak. Sykdommer, både mentale og fysiske, kan påvirke søvnen. De vanligste søvnforstyrrelser er søvnløshet og søvnapné. Andre søvnforstyrrelser er narkolepsi, marerittlidelse, søvnparalyse, søvngjengeri, skrekkanfall og søvnrelatert spiseforstyrrelse.
Fysisk aktivitet
Regelmessig fysisk aktivitet har en positiv effekt på søvnkvaliteten. Den kan også ha en positiv virkning på søvnproblemer, men det er ingen garanti for at fysisk aktivitet løser dette. Det er viktig å være tålmodig og ikke forvente resultater med en gang. Fysisk aktivitet tidlig på dagen er det beste, om man er i stor fysisk aktivitet om kvelden vil det ha en negativ effekt.
Søvn og vekt
Søvn er også med å regulere vekta. Når du sover reduserer kroppen blant annet mengden av stresshormonet kortisol, slik at kroppen kan produsere andre hormoner, som ghrelin og leptin. Disse er på hver sin måte med på å styre sult- og metthetsfølelse din. En god natts søvn regulerer nivåene av leptin og ghrelin.
Ghrelin er et hormon som ofte kalles sulthormonet. Når du sover, reduseres nivåene av dette hormonet og du ønsker mindre mat. Hvis du sover for lite lagrer kroppen mer ghrelin som gir signaler til hjernen om at du bør spise mer.
Leptin kalles gjerne for appetitthormonet, og er hormonet som gjør at du føler deg mett. Når du sover stimuleres kroppen til å produsere mer leptin. Dette hormonet styres også av fett. Jo mer fettmasse, jo mer leptin.
En studie med 1000 deltakere fant at de som hadde underskudd på søvn hadde 14,9% høyere ghrelin-nivåer og 15,5% lavere leptin-nivåer enn de som fikk tilstrekkelig søvn.
De med søvnunderskudd hadde også høyere BMI.
Se også
Drøm
Søvn hos dyr
Høneblund
Bevisstløs
Referanser
Eksterne lenker
Artikler om Søvn i Tidsskrift for Den norske legeforening
«Slik virker søvn», fra nettstedet forskning.no
Søvnsykdommer for enhver smak - artikkel fra forskning.no 25.3.03 | norwegian_bokmål | 0.680322 |
wake_up_wrong/PMC5136610.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List Ind Health v.54(6); 2016 Nov PMC5136610
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of indhealth
Ind Health. 2016 Nov; 54(6): 528–541.
Published online 2016 May 18. doi: 10.2486/indhealth.2015-0236
PMCID: PMC5136610
PMID: 27193071
Time to wake up: reactive countermeasures to sleep inertia
Cassie J. HILDITCH,1,* Jillian DORRIAN,1 and Siobhan BANKS1
Author information Article notes Copyright and License information PMC Disclaimer
Go to:
Abstract
Sleep inertia is the period of impaired performance and grogginess experienced after waking. This period of impairment is of concern to workers who are on-call, or nap during work hours, and need to perform safety-critical tasks soon after waking. While several studies have investigated the best sleep timing and length to minimise sleep inertia effects, few have focused on countermeasures -especially those that can be implemented after waking (i.e. reactive countermeasures). This structured review summarises current literature on reactive countermeasures to sleep inertia such as caffeine, light, and temperature and discusses evidence for the effectiveness and operational viability of each approach. Current literature does not provide a convincing evidence-base for a reactive countermeasure. Caffeine is perhaps the best option, although it is most effective when administered prior to sleep and is therefore not strictly reactive. Investigations into light and temperature have found promising results for improving subjective alertness; further research is needed to determine whether these countermeasures can also attenuate performance impairment. Future research in this area would benefit from study design features highlighted in this review. In the meantime, it is recommended that proactive sleep inertia countermeasures are used, and that safety-critical tasks are avoided immediately after waking.
Keywords: Body temperature, Caffeine, Countermeasures, Light, Napping, Self-awakening, Shift work, Sleep inertia
Impaired performance and alertness upon waking is known as “sleep inertia”1, 2). Impairment is most severe immediately upon waking and then dissipates, generally returning to baseline levels within 15–60 min3, 4, 5, 6). Sleep inertia is a concern for industries in which workers perform safety-critical tasks soon after waking. Motorists are also at risk when driving too soon after waking, for example when following government recommendations to nap if tired during long drives7, 8). Sleep inertia has been a contributing factor in several major accidents and incidents9, 10, 11, 12). For example, an air crash involving 158 fatalities resulted from poor decisions made by the Captain who had just woken from an in-flight nap10). In another example, heavy contact between supply vessels occurred after the Chief Officer over-slept and consequently arrived on the bridge within minutes of waking11).
Go to:
Factors that Influence Sleep Inertia
Sleep inertia is typically measured in laboratory settings using a combination of sleepiness scales and cognitive tests. However, the sensitivity of these cognitive tasks to sleep inertia varies13, 14). Consequently, the severity of sleep inertia observed can depend on the task used to measure it. Test batteries are often administered immediately after waking and repeated intermittently for up to one hour. Sleep inertia has been measured under a variety of conditions ranging from a full night’s sleep, to short naps at different times of day. Much of this research has focussed on establishing the best length and timing of sleep to minimise sleep inertia. Studies suggest that avoiding sleep periods ending during the circadian low5, 15), and keeping naps to less than 30 min4, 16, 17, 18) to avoid waking from deep sleep15) can minimise sleep inertia magnitude. Prior sleep loss can also exacerbate sleep inertia15, 19), which is particularly important in shiftwork where workers often experience extended wakefulness and/or sleep loss20, 21). Indeed, naps have been suggested as a potentially effective countermeasure to sleepiness following sleep loss and during the circadian low22, 23, 24, 25, 26). Generally the long-term benefits provided by the nap outweigh the short-term detriments associated with sleep inertia4, 16, 25). The challenge is to maximise the benefits of a nap while minimising and/or managing sleep inertia. In addition, there may also be individual factors that contribute to overall sleep inertia severity. For example, some sleep medications27) or sleep disorders may exacerbate sleep inertia symptoms, although there is currently very little research in this area.
Go to:
Taking a Reactive Approach
Research investigating proactive strategies for optimal sleep length and timing to minimise sleep inertia and maximise alertness is important for informing industry guidelines on rest breaks and shift scheduling. In operational environments, however, it is not always feasible to plan the length and timing of a sleep period. For example, when workers are on-call (such as in the emergency services or the military), or for workers such as healthcare professionals who take naps during extended-hours or night shifts. For these workers, the need to process crucial and complex information or to engage in safety critical activities almost immediately after waking could occur at any time. Given this, it is surprising that few studies have directly sought to reduce the effects of sleep inertia through reactive countermeasures. That is, strategies implemented upon wake-up, as opposed to proactive strategies such as planning sleep timing and duration. This review examines the literature on potential reactive countermeasures to sleep inertia including caffeine, light, and temperature, and discusses possible avenues for future research.
Go to:
Search Methods
Three electronic databases (PubMed, Science Direct, and Scopus) were searched on July 8, 2015 for the following search terms paired with “sleep inertia”: adrenaline, caffeine, countermeasures, epinephrine, light, nap, noise, on-call, self-awakening, shift work, sound, and temperature. This resulted in 256 articles. Two articles were manually retrieved from reference lists. Seventeen articles remained after filtering for duplicates (n=150) and studies not meeting the inclusion criteria outlined below (n=91). Figure 1 illustrates the search and selection process. The large number of duplicate articles was a result of using multiple search terms and multiple databases which each returned similar results. The selected articles are summarised in Table 1. Note that one study is included three times as it investigated three countermeasures28).
An external file that holds a picture, illustration, etc.
Object name is indhealth-54-528-g001.jpg
Fig. 1.
Flowchart illustrating the structured narrative review selection process for articles populating Table 1.
Table 1.
A summary of papers investigating countermeasures to sleep inertia
Counter-measure Authors Prior sleep/wake protocol Sleep length Wake-up timing Inertia testing points post-sleep Subjective alertness improved?^ Objective performance improved?^ Reactive?
Caffeine Reyner & Horne, 199730) Sleep restriction (5 h TIB) 15 min 14:40 Drive: 0–180 min; 30-min bins
KSS: every 200 s No obvious signs of inertia, but no nap-only comparison group. No obvious signs of inertia, but no nap-only comparison group. No
Van Dongen et al., 200131) 88 h extended wakefulness with 2-h naps every 12 h 2 h 04:45 & 16:45 5, 75 min Not reported Yes (5 min) No
Hayashi et al., 200328) Habitual sleep 20 min 13:00 0, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 min Yes (15 min) Yes (15 min) No
Newman et al., 201332) Habitual sleep 1 h & ~6 h 01:00 & 06:00 0, 6, 12, 18 min Not reported Yes (18 min) Yes
Light
(post-wake) Hayashi et al., 200328) Habitual sleep 20 min 13:00 0, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 min Yes (15 min) No improvement Yes
Santhi et al., 201313) Sleep restriction (6.5 h TIB) 6.5 h 06:42–07:27, depending on habitual wake-up 3, 33, 63, 93 min No improvement No improvement Yes
Light
(pre-wake) Van de Werken et al., 201035) Habitual sleep ~8 h ~07:00 1, 15, 30, 45, 60, 90 min Yes (15 min) No improvement No
Giménez et al., 201033) Habitual sleep ~8 h ~07:00 N/A Yes (25 min reduction time needed to feel fully awake) Not reported No
Harrison et al., 201134) Habitual sleep 90 min 15:30 3 min No No No
Thompson et al., 201436) Habitual workday sleep 8 h Habitual workday timing (mean not reported) 5, 35, 75 min Yes (better on average over whole testing period) Yes (better on average over whole testing period) No
Sound Tassi et al., 199237) Habitual sleep 1 h 01:00 & 04:00 0, 40 min.
30-min test divided into 3 min bins. Not reported Yes (3 min) Yes
Hayashi et al., 200438) Habitual sleep 20 min 14:20 1, 6, 11, 16 min Yes (1 min) No control group.
High preference music better than low preference (1 min) Yes
Temperature Krauchi et al., 200439) Unknown, assume habitual 8 h (nocturnal sleep) & 2 h (afternoon nap) 07:00 & 18:00 KSS at 0, 30, 60, 90, 120 min Correlation with distal-proximal skin temperature gradient Not reported Yes
Krauchi et al., 200640) Habitual sleep 75 min Multiple wake-ups across the circadian cycle KSS at 0, 30, 60, 90, 120 min Correlation with distal skin temperature Not reported Yes
Self-awakening Kaida et al., 200341) Habitual sleep 15–20 min 14:20 5, 10, 15, 20, 25, 30 min Yes (5 min) No
Greater P300 amplitude (15 min) No
Kaida et al., 200342) Habitual sleep 15–20 min 14:20 5, 10, 15, 20, 25, 30 min Yes (10 min) No
Heart rate increased (3 min before waking) No
Ikeda & Hayashi 20106) Habitual sleep Habitual (mean 7.3 h) Habitual (mean 08:06) 1, 16, 31, 46 min Yes (fatigue - better on average over whole testing period)
No (sleepiness not improved) Yes (in first 15 min test session) No
Ikeda et al., 201443) Habitual sleep Partial restriction (5 h) Habitual (range: 05:30–08:30) Immediately after waking (home study) No Yes (only one test point) No
Face-washing Hayashi et al., 200328) Habitual sleep 20 min 13:00 0, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 min Yes (1 min) No improvement Yes
Open in a separate window
Table notes: ^first recorded improvement; KSS: Karolinska sleepiness scale; N/A: not applicable; TIB: time in bed.
Given the diversity of methods and the exploratory nature of the many approaches to managing sleep inertia, a traditional systematic review style was not implemented. Rather than limiting the scope of the studies included in the review, a structured narrative approach was chosen to allow collation across a broad range of research in this area29). Many of the studies included in this review have methodological limitations that may have rendered them ineligible for traditional systematic review. Nevertheless, these studies highlight important research ideas, and point the way for future studies. In this way, this inclusive review is intended as a “call to arms” for future research initiatives.
Studies in Table 1 met the following inclusion criteria: original article; published in peer-reviewed journal; written in English; used healthy, human, adult participants; and directly trialled a countermeasure to sleep inertia or provided evidence for a potential countermeasure. The majority of screened articles were excluded due to not investigating a countermeasure to sleep inertia or not reporting original research (e.g. review papers) (Fig. 1). All studies in the table were laboratory-based; 15 were experimental and two were observational. Sample size ranged from n=8 to n=23 in 14 within-participant studies, and n=16 to n=44 in three between-participants studies. Both male and female participants were included in all but three studies, which only included male participants. Outcome measures, testing points, timing of awakening, and prior sleep/wake history, and length of sleep periods varied, and as such, direct comparisons between studies were not possible. Six countermeasure categories were identified: caffeine28, 30, 31, 32); light13, 28, 33, 34, 35, 36); sound37, 38); temperature39, 40); self-awakening6, 41, 42, 43); and face-washing28).
Go to:
Caffeine
Caffeine is a readily available and cost effective stimulant used strategically, socially and often habitually44, 45). Caffeine promotes alertness by blocking adenosine receptors. Adenosine, a by-product of cellular energy expenditure, is a neurotransmitter inhibitor which increases with time awake and causes drowsiness46). Caffeine has been shown to improve performance and alertness under conditions of high sleep pressure such as sleep deprivation and during the night47, 48). The search yielded four studies that examined caffeine as a countermeasure to sleep inertia (Table 1). Van Dongen et al.31) found that sustained, low dose caffeine counteracted sleep inertia immediately following 2-h naps across 88 h of sleep deprivation. Notably, caffeine was administered hourly for a plasma concentration approximately 3.7 mg/L, equivalent to one-quarter cup of coffee every hour for 66 h. That is, caffeine was not administered directly after the nap, rather, at regular intervals before and after each nap. In this study, performance on a 10-min psychomotor vigilance task (PVT; a measure of behavioural alertness and vigilance, in which the participant is required to respond to stimuli presented at random intervals as quickly as possible by pressing a button) under placebo conditions deteriorated immediately post-nap, but there was no change in performance pre- to post-nap in the caffeine condition. Testing points over an hour after the nap showed no differences between the two groups. From these results the authors concluded that caffeine had eliminated the effect of sleep inertia on this task. There was evidence to suggest that caffeine affected the sleep architecture of the naps in this study. For example, it took longer for participants to fall asleep, and the amount of time spent in deep sleep was significantly reduced in some of the nap opportunities. However, this effect was not consistent across all naps, yet the reduction of sleep inertia was consistent following all naps. Therefore, the authors concluded that the action of caffeine on sleep inertia was unlikely to be due to the alteration of prior sleep bout.
More recently, Newman et al.32) trialled a caffeinated chewing gum (100 mg) administered immediately upon waking and observed improved performance on a 5-min PVT relative to placebo at 12–18 min post-nap. Caffeine administered via gum reaches peak plasma levels more quickly than via a pill49). However, there were no differences observed between conditions until at least 12–18 min post-nap, suggesting that the caffeine gum, as in the pill form46), has a delayed effect, and is therefore unable to attenuate sleep inertia during the initial, most critical period of wakefulness. The caffeine gum may, however, limit the duration of sleep inertia and therefore provide faster recovery post-nap. It is worth noting that data from the placebo condition did not consistently show clear sleep inertia effects (i.e. performance was not always worse immediately after waking). Therefore, it is difficult to determine whether caffeine had an effect on sleep inertia per se, or improved general performance under conditions of mild sleep restriction and high circadian pressure (awoken at 01:00 and 06:00 after a 1-h and 6-h sleep opportunity, respectively). As an alternative mechanism, studies have shown that chewing non-caffeinated gum can improve alertness and cognitive performance50, 51). Therefore, chewing may have acted as a countermeasure in Newman and colleagues’ study32), thus reducing sleep inertia effects in both the placebo and caffeine groups. If so, the caffeine effect would be acting on top of the chewing effect, leading to delayed improvements on background performance. Overall, the use of caffeinated gum offers promising results for truncating sleep inertia duration. This study demonstrates the importance of using a placebo gum, and ideally another placebo or control condition, so that it is possible to isolate the effects of chewing and caffeine on sleep inertia. Further study is warranted to determine whether different caffeine dosages may be more effective, or different performance tasks may be more sensitive to detect changes in sleep inertia following reactive caffeine gum administration.
Other studies have administered caffeine by adding it to decaffeinated coffee which was consumed prior to a short daytime nap28, 30). Hayashi and colleagues28) administered 200 mg of caffeine (equivalent to 2–3 cups of instant coffee52, 53)) prior to a 20-min nap. Despite pre-nap administration, significant improvements in alertness and measured performance in the caffeine condition were not observed until 15 min after the nap. However, during this 15 min period, there were no differences in alertness or performance between the nap-only and no-nap group. This indicated a lack of observable sleep inertia, which may have been due to the experimental design. Mean total sleep time during the 20-min nap opportunity ending at 13:00 was 14.8 min, with no slow wave sleep (SWS, or “deep sleep”). These conditions are not particularly conducive to sleep inertia4, 16). This study demonstrates that, when investigating sleep inertia countermeasures, it is important to use experimental circumstances that are most likely to produce sleep inertia. For example, following: scheduled awakenings at night5, 15); extended wakefulness or prior sleep loss15, 19); or longer naps4, 16, 18).
Reyner and Horne30), using 150 mg of caffeine in decaffeinated coffee (equivalent to approximately two cups of coffee52, 53)), found that the combination of caffeine and a 15-min nap was associated with better performance across a 2-h drive than following either countermeasure in isolation. The driving task commenced 5 min after the nap, within the sleep inertia period. However, as investigation of sleep inertia was not a research aim of the study, the data were reported in 30-min bins which do not allow investigation of performance changes across this peak sleep inertia period. The first 30-min bin, however, did not show any obvious signs of sleep inertia (i.e. worse performance) relative to subsequent bins. Interpretation of these results with regards to sleep inertia is further limited as performance in the caffeine-plus-nap group was not directly compared to a nap-only group, only to results from a group in a previous study54). In the previous study54), participants drove for 2 h before receiving caffeine and/or nap, whereas in the follow-up study30) they drove for 1 h. Therefore the post-treatment results of these conditions should be compared with this in mind.
There is a government-led road safety campaign, based on the above study, to promote the use of a caffeine-nap combination as a drowsy driving countermeasure55). Further, despite a lack of studies investigating sleep inertia following short naps at night, other government websites promote the use of short naps (≤30 min) with no mention of potential sleep inertia7, 8). Current policies and recommendations to counteract sleepiness while driving require a stronger evidence base with regards to sleep inertia. The studies reviewed highlight the need for experimental and observational testing which allows for the direct assessment of the impact of caffeine in combination with a nap on driving performance immediately after waking at different times of day.
There are several considerations when applying these results in the workplace. In most studies28, 30, 31), caffeine was administered before the nap. Each of these studies yielded improvements post-nap in caffeine compared to placebo groups. In unpredictable scenarios, however, such as working on-call, there may not be sufficient warning or opportunity to use caffeine proactively. Proactive use may only be effective prior to short naps. There is the potential for sleep to be disrupted if caffeine is taken beforehand, resulting in a trade-off between reduced sleep inertia and reduced total sleep or sleep quality28, 31). However, when caffeine was administered post-nap, even in a rapid acting chewing gum format, caffeine effects were not seen until 12–18 min post-nap, which, while useful in some scenarios, may not be fast enough in time-critical situations32). Therefore, when used proactively, caffeine has the potential to eliminate sleep inertia effects almost immediately after short naps. Further research into the use of caffeinated gum is needed to explore the potential benefits of this reactive countermeasure beyond the demonstrated reduction in sleep inertia duration.
Another consideration in the application of these results to operational settings is habitual use and individual caffeine sensitivity. Van Dongen and colleagues31) employed a two-week caffeine-free wash-out before the study to minimise the effects of individual differences in caffeine tolerance. This study still observed large between-subjects variability in plasma caffeine concentrations. The other studies in Table 1 did not report habitual caffeine use30), or allowed participants to drink up to four cups of coffee per day until the experimental period28, 32). This may have influenced the effects of the caffeine, with low-caffeine users more sensitive to the effects of the experimental dosage compared to high-caffeine users46). Alternatively, high-caffeine users may have experienced withdrawals in the placebo condition56). Not controlling for habitual caffeine use, however, is perhaps more ecologically valid, as participants had a range of habitual caffeine intakes, just as a work force would. Studies of caffeine use in shift workers report high levels of caffeine intake especially for: older workers, those on night and morning shifts, and following reduced sleep45, 57). Although it is important to understand caffeine effects under controlled conditions, it is also important to understand individual differences in caffeine response under natural conditions. Translating results from controlled studies into real-world scenarios is a critical step before recommending caffeine as a countermeasure to sleep inertia. Further, the long-lasting effects of caffeine may disturb subsequent sleep opportunities58), so it may not be suitable for use on some occasions, such as towards the end of a night shift.
Go to:
Light
Melatonin is a hormone which helps to regulate sleep-wake cycles and is suppressed under conditions of bright light exposure59). Light also serves as a zeitgeber (time-giver) to entrain our internal body (circadian) rhythms to a 24-h cycle60, 61). Light signals are sent from the eye to a cluster of cells in the brain responsible for our circadian rhythms, the suprachiasmatic nucleus. In this way, light can be used at key times to manipulate circadian timing, leading to changes in sleep patterns62, 63). In addition to its circadian entrainment effects, bright light exposure has also been shown to directly improve alertness and cognitive performance during the day, night, and following sleep restriction64, 65, 66, 67, 68). Investigation into optimising the intensity, wavelength and duration of light exposure to manipulate both sleep and performance are ongoing. There is potential for bright light exposure to directly improve alertness and performance during the sleep inertia period.
Few studies have investigated the effect of light immediately following waking on performance and alertness during this period. The first study of this type exposed participants to bright light (2,000 lux) for 1 min after waking from a 20-min nap28). This brief exposure to bright light did not reduce sleep inertia within the first 15 min after waking. It did, however, modestly improve subjective ratings for the following 45 min. More recently, Santhi and co-authors13) investigated the effect of four light conditions (dim, blue-intermediate, blue-enhanced, and bright blue-enhanced) on a range of cognitive tasks during the sleep inertia period. Participants were woken from a 6.5-h nocturnal sleep and were exposed to light for 4 h. There were no differences between the four conditions in subjective alertness or performance measured every half-hour for the first two hours and then every hour up to 4 hours after waking. The relatively infrequent testing points during the initial sleep inertia period may have limited observations in this protocol. Participants in this study were selected based on a self-reported need for at least 60 min to feel fully alert after waking. Indeed, subjective ratings of sleepiness showed a clear sleep inertia pattern, with highest sleepiness ratings immediately after waking which dissipated across subsequent testing points. However, the performance tasks varied in their sensitivity to sleep inertia. This demonstrates the important methodological consideration of choosing an appropriate task to measure sleep inertia. Regardless of the outcome measured, however, there was no significant effect of light on performance or alertness within 4 h of waking. The improvement in working memory observed 4 h after waking was likely due to light affecting general performance rather than sleep inertia per se. These studies demonstrated that both brief and sustained light exposure after waking did not improve performance during the sleep inertia period. These results suggest that light exposure may be of limited effectiveness during the sleep inertia period. However, it may be worth investigating different light intensities and qualities.
As well as investigating light exposure after waking, some studies have looked at light exposure before waking. This approach assumes a different action pathway of light. Instead of direct alerting properties of bright light exposure during wake, exposure to light before waking applies the theory that light will act to lighten sleep before waking, therefore reducing sleep inertia35). However, Harrison and colleagues34) found no difference in measured performance or alertness following a 90-min afternoon nap during which participants were exposed to varying light intensities (0, 1, 80, 6,400 lux). Similarly, Van de Werken et al.35), despite observing improvements in subjective alertness, found no differences in post-sleep performance on a simple reaction time and addition task between a dawn simulation condition and control. In the dawn simulation, light was increased to 300 lux in the 30 min prior to wake-up; there was no light before wake-up in the control condition. There were no sleep stage differences between conditions in either the 90-min nap34) or during the 30 min of artificial dawn35).
In contrast, modest performance benefits were observed both on cognitive and physical (cycling) tasks following dawn simulation, with no differences in sleep quality as measured by actigraphy36). Giménez et al.33) found that participants reported needing less time to feel fully alert after stronger artificial dawn light levels (250 lux vs 0 lux and 50 lux). Together these studies suggest that dawn light may improve subjective alertness. Further studies of the effect of dawn light on objective performance during the immediate wake-up period are necessary to determine the overall efficacy of dawn lighting to reduce subjective and objective sleep inertia.
Light may have an additional action pathway during the sleep inertia period via the cortisol awakening response (CAR). Cortisol is a hormone typically associated with stress response and follows a diurnal pattern with higher levels during the day and lower levels at night69). The CAR refers to the sharp increase in cortisol upon waking in the morning70). This response is greater in the presence of light presented immediately after waking (800 lux for 1 h)71). Less intense light presented before waking (dawn simulation: light gradually increased to 250–300 lux over 30 min before waking) has had mixed effect35, 72). Furthermore, a greater CAR has been associated with increased levels of self-reported arousal72, 73) and reduced sleepiness74).
The role of cortisol and the CAR in sleep inertia is unknown. A clue to its role comes from the discovery that SWS is associated with lower cortisol75). Deep sleep has also been associated with greater sleep inertia15, 76), although this relationship has not been consistently demonstrated18, 77). The relationship between deep sleep and sleep inertia may be mediated by cortisol, although this has not been directly investigated. If the relationship between cortisol and sleep inertia can be better established, manipulating cortisol through light interventions or exogenous administration may offer an alternative sleep inertia countermeasure. The inclusion of measures of cortisol, alertness, and performance across the sleep inertia period would be beneficial to systematically track inter-relationships between these factors during this time.
Using light after waking would be relatively straightforward to implement in the workplace. For example, strategic exposure to light boxes at work has successfully helped shift workers adjust their circadian rhythm to better match their shifts78, 79). However, a trial of bright light in a chemical plant uncovered several limitations including poor compliance with the interventions and variations in light exposure amongst workers due to different operational tasks80). The effectiveness of such a countermeasure may vary by workplace. However, a light box, or light-emitting glasses81) used immediately after waking, could feasibly be implemented as a reactive countermeasure. While using light before waking would be unsuitable for implementation in unpredictable sleep/wake environments, it may have an application for scheduled rest periods. For example, a dawn simulation light box could be used in a workplace napping room. An additional caution to be added to the use of bright light as a countermeasure to sleep inertia is the potentially unwanted effect of entrainment. For example, bright light exposure, particularly near the circadian nadir, could delay or advance circadian phase62, 63). This could lead to changes in sleep timing that are counterproductive to the work schedule and ultimately lead to sleep loss, sleepiness and poor performance.
Go to:
Sound
Sound such as pink noise (random noise with more low frequency components than white noise) can be used as a sleeping aid by providing a constant, ambient auditory background thereby minimising the impact of random noises on sleep initiation and maintenance82, 83). Conversely, noise has been used to promote alertness under sleep deprivation conditions84) or as a positive stressor to improve performance85). This inconsistency in the literature may be due to variation in noise type (e.g. continuous versus intermittent). The effect of noise may also be task dependent, with significant effects more likely observed on simple compared to complex tasks86, 87, 88). The search identified two studies which used different sounds in an attempt to reduce sleep inertia.
Tassi and colleagues37) exposed participants to continuous noise during a spatial memory test performed after waking. In this study37), spatial memory after waking from a 1-h nap at 01:00 during the no noise condition was worse for up to 15 min compared to the total sleep deprivation condition (no nap, no noise). Performance in the group exposed to 75 dB of pink noise (random noise with more low frequency components than white noise) after waking, did not differ from total sleep deprivation. The authors suggest that noise had an arousing effect which reduced sleep inertia. Noise delivered after a nap ending at 04:00, however, was ineffective and may have exacerbated the effects of partial sleep deprivation under these conditions. Given the conflicting results from this study and the literature generally, more research is required to establish the effect of pink noise exposure under different sleep conditions, and on different tasks. The background noise level of the work place may also impact on the effectiveness of this countermeasure. For example, high levels of ambient noise are typically associated with elevated stress and fatigue in noisy work environments such as factories89, 90). The results of Tassi and colleagues’ study37) suggest, however, that under certain conditions, exposure to pink noise after waking could minimise sleep inertia.
Hayashi and colleagues38) took a different approach, playing excitatory music at 60 dB during a 20-min testing period after waking. Music, especially music that the participant preferred, reduced subjective sleepiness post-nap compared to a no-music control. Similarly, performance on a visual oddball task was improved with high preference music compared to low preference music. Future research in this area would benefit from the inclusion of a no nap control group to confirm that the effects observed target sleep inertia relative to general performance. Reyner and Horne91), under conditions of sleep restriction, found that playing the radio only had a brief, positive effect on alertness and simulated driving performance, and was therefore not suitable to long-distance driving. Playing music may, however, be effective in short-term sleep inertia scenarios.
From the limited literature it appears that sound (noise or music) has the potential to improve performance, at least briefly, under certain conditions. Further research in this area would help to determine the most effective sound to target sleep inertia symptoms. From an operational perspective, implementation in a work setting should be relatively easy. Indeed, background music has been used successfully in both industrial and office settings to improve vigilance and performance at work92). However, rather than background music, sound delivery would likely need to be via headphones, especially in shared sleeping environments. For example, a personal music player with a collection of songs pre-selected by shift workers (high preference music38)) could be kept in a napping room for use immediately after waking.
Go to:
Temperature
Many studies have investigated the role of thermoregulation in moderating sleep onset and maintenance (for review, see Lack et al.93)). At sleep onset there is a drop in core body temperature (CBT) and an associated increase vasodilation and blood flow to the extremities in order to lose heat. This change in body temperature is thought to promote sleep onset and maintain sleep throughout the nocturnal sleep period93). Furthermore, the association between the rate of distal skin temperature increase (measured at the extremities) and sleep onset can be observed at different phases of circadian rhythm in CBT94). Manipulation of skin temperature and CBT have been found to improve sleep quality95), performance, and maintenance of wakefulness96). For example, strategies to warm the extremities (e.g. a hot water bottle at the feet) can help to promote sleep onset97). The role of thermoregulation at sleep offset, however, has received less attention98).
A tantalising preliminary study in this area has demonstrated a relationship between subjective measures of sleep inertia and changes in the distal-proximal skin temperature gradient (DPG)39). In this study, the reduction in subjective sleepiness correlated with a decrease in the DPG. Furthermore, the relationship between distal skin temperature and subjective sleepiness remained in a multiple nap protocol in which sleep and wake opportunities were rapidly alternated (75 min sleep / 150 min wake)40). The authors suggest that actively manipulating the temperature of extremities through simple cooling strategies that evoke distal vasoconstriction and reduce heat loss from the periphery may result in faster sleep inertia dissipation. Van De Werken35) also found that dawn light accelerated the DPG change after waking and that this was associated with a decrease in subjective sleepiness, but not objective performance.
From an operational perspective, if changing body temperature only changed subjective feelings, it may lead to a false sense of improved objective alertness and performance after waking99, 100, 101, 102, 103, 104). Studies further investigating the relationship between DPG and objective measures are well-warranted, however, as an intervention would be relatively easy to implement in most work places. For example, workers could place their hands and feet in a receptacle filled with cold or iced water to rapidly lose heat from their extremities immediately after waking. Such a countermeasure would also have the advantage of not negatively influencing future sleep opportunities (c.f. caffeine, light).
Go to:
Self-awakening
Some studies have observed that waking from deeper stages of sleep (i.e. SWS) is associated with greater sleep inertia15, 76). This has led to studies of self-awakening to minimise the chances of waking from deep sleep. Self-awakening refers to spontaneously waking after a set period of time, without the use of an external stimulus such as an alarm. Kaida and colleagues41, 42) compared self-awakening to being woken by an experimenter after a 15-min afternoon nap. They reported that self-awakening led to lighter sleep, as scored using hypnagogic scoring105), a method which looks at the first two lighter stages of non-rapid eye movement sleep (Stage 1 and Stage 2)106) in greater resolution41). There were no differences observed across stages according to Rechtschaffen and Kales106) criteria41, 42). This finding suggests that traditional sleep scoring methods may not be sensitive enough to detect electroencephalographic changes induced by self-awakening techniques. Spectral analysis of these sleep episodes may be another useful method for understanding the mechanisms underlying self-awakening.
Kaida and colleagues41) measured physiological arousal levels by evaluating the amplitude of a specific brain response to an audible tone presented immediately after waking and repeated every 5 min for 30 min. There was a greater response after self- compared to forced-awakening at 15 min post-wake, but not at earlier or later testing points. The authors argue that this greater response following self-awakening is indicative of greater physiological arousal41). In a second study42), subjective sleepiness was improved at 10 min post-wake, but improvements in performance on an auditory oddball task were not observed until later. These results should be interpreted with caution as the experimental design (i.e. a short afternoon nap) may not have generated sleep inertia. Indeed, there was no difference in performance between the nap and no-nap group, and no immediate subjective ratings of sleep inertia. This study is another example highlighting the importance, when investigating sleep inertia countermeasures, of setting experimental conditions that are likely to produce sleep inertia. For example, Ikeda and colleagues investigated self-awakening following partial sleep restriction43, 107). In contrast to Kaida and colleagues’ studies41, 42), improvement was observed on cognitive tasks following self-awakening, but there were no significant differences in subjective sleepiness ratings.
While there is some evidence to support the use of self-awakening to reduce sleep inertia, a potential limitation in applying this technique is the inconsistent success rate of self-awakening. In Kaida and colleagues’ studies41, 42), there was a 71–82% success rate for self-awakening within ± 5 min of the 15-min target, much higher than previous studies cited by the authors which range from 18–42%41). The study, however, selected participants who were well-rested and claimed they were able to self-awaken from nocturnal sleep. Therefore this approach may be most beneficial for those who find it easier to self-awaken. It would be interesting to further this research under conditions better representing shift work scenarios, e.g. following chronic sleep restriction or extended periods of wakefulness21). In Ikeda and Hayashi’s study6), for example, participants habitually woke from an alarm, and subsequently only nine of the 30 attempted self-awakenings in the study (30%) were within 30 min of target wake time. As workers are generally required to return to shift at a specific time, it is unlikely that self-awakening would be a reliable strategy. Furthermore, this method is not a viable countermeasure for unscheduled wake-ups. For naps taken at home without time pressures, however, the self-awakening technique may offer relief from subjective feelings of sleep inertia.
Go to:
Face Washing and Common Countermeasures
Anecdotally, people have wake-up routines which reduce sleep inertia. However, while commonly used countermeasures to driver fatigue such as cold air or playing the radio have been investigated91), few studies have examined their use for sleep inertia. Hayashi and colleagues’28) trialled face-washing as a practical, and commonly used, sleep inertia countermeasure following a 20-min afternoon nap. The authors describe an immediate, but short-lived, reduction in subjective sleepiness relative to nap-only, with no differences in performance on a memory search task. These results suggest that face-washing may improve subjective alertness after waking. Further research using different performance measures would be useful to establish whether the null findings for performance were specific to the memory search task used. It is possible that other tasks may be more sensitive to inertia.
In a study examining the time course of sleep inertia, Jewett et al.3) found no differences in performance on an addition task between a constant routine condition (lying in semi-recumbent position) and an ambulatory condition in which participants were able to get up, eat breakfast, and shower. Both groups were kept under the same semi-recumbent, fasting conditions until at least 35 min after waking, therefore it is not clear whether these activities would have made a difference if undertaken sooner after waking. Another study reported a positive effect of breakfast on morning mood and memory, but did not report when participants woke, so it is unclear whether it was strictly a study of sleep inertia108). More studies on common countermeasures such as showers, meals, or physical exercise are worthy of investigation. For example, a shower may act to change overall body temperature, and therefore might be more effective than face washing. Empirical research is needed to confirm effective common practices, and to de-bunk ineffective strategies.
Go to:
Adrenaline and Operational Scenarios
Finally, when considering sleep inertia in real-world scenarios, adrenaline is often proposed as a natural countermeasure. Adrenaline is a stress hormone released from the adrenal glands109). Adrenaline may play a role in mitigating impairment during the sleep inertia period110). Laboratory studies of sleep inertia are typically conducted in low-stress environments. It is unknown whether a change in stress levels may affect sleep inertia, or the effectiveness of countermeasures. Further, there are no studies of adrenaline and its effect on performance and alertness during the sleep inertia period. This is likely due to constraints associated with collecting blood samples during the sleep inertia period and the short half-life of adrenaline111). Anecdotal and experimental evidence suggest that adrenaline may not be a reliable countermeasure to sleep inertia. Emergency service workers often report habituating to their high pressure roles, no longer feeling the “rush of adrenaline”. Reports are supported by a study that described changes to hormone levels following stressful field exercises and military training112). Special Forces soldiers had a different response than non-Special Forces soldiers, suggesting a degree of adaptation to stress. However, this adaptation may not be common for shift workers in less stressful occupations. Whether the hormonal response immediately upon waking in high stress scenarios changes with experience, and/or varies between individuals, is unknown. Nor do we know whether increases in hormones such as adrenaline can restore cognitive function during the sleep inertia period, or whether they may in fact further compromise clear thinking and decision-making12). Until there are studies directly investigating the role of adrenaline on cognitive performance during sleep inertia, reliance on this endogenous countermeasure is not recommended.
This area would also benefit from study in the field, which to date, are few110). A survey of emergency medical service pilots revealed that sleep inertia had occasionally compromised flight safety113) and some critical care nurses report avoiding napping on night shift due to concerns about performance impairment upon waking114). Taken together, these studies suggest that there is potential for sleep inertia-related safety risks, or at least, that the perception of sleep inertia as a safety issue exists in the field. Several incident investigation reports have cited sleep inertia as a contributing factor9, 10, 11, 12). Systematic studies of the effect of sleep inertia in real-world situations are essential for progressing our understanding of sleep inertia, and the development of effective countermeasures for practical implementation.
Go to:
Conclusions
Examination of the current literature reveals a gap in the evidence-base for the implementation of a reactive countermeasure to sleep inertia which is effective within 15 min of waking. Caffeine is perhaps the closest option, although to target immediate performance and alertness, it needs to be administered prior to a short sleep bout, and in this way, is not a reactive strategy. This countermeasure may be useful, therefore, in situations where napping can be planned. Given that current investigations into light and temperature are in their infancy, further study is warranted to understand the physiological mechanisms underlying these interventions and whether performance can be improved after waking, in line with some encouraging results from subjective measures.
A challenge in identifying an effective sleep inertia countermeasure is that effectiveness may depend on sleep inertia severity. The majority of the studies reviewed assessed sleep inertia following afternoon naps, or upon waking from habitual night time sleep. There were no clear differences in the efficacy of countermeasures between these two scenarios. However, strategies that work under these conditions may not be sufficient in other scenarios. In shift work conditions, for example, the magnitude of sleep inertia may be exacerbated due to factors such as prior sleep loss, time of day, and sleep length. The impact of these factors on the effectiveness of countermeasures has yet to be investigated. Studies investigating sleep inertia countermeasures should consider directly comparing the effect of these factors, or testing countermeasures in scenarios more common to shift work (e.g. a night shift nap).
Identifying a viable countermeasure also involves assessing operational viability. Many of the countermeasures discussed could be implemented in a workplace. Developing a countermeasure that can attenuate the magnitude and/or duration of sleep inertia in both scheduled and unpredictable shift work environments warrants further investigation. In the meantime, it is recommended that proactive sleep inertia countermeasures are used, and that safety-critical tasks are avoided immediately after waking.
Go to:
Acknowledgments
The authors have no conflicts of interest to declare.
Go to:
References
1. Lubin A, Hord DJ, Tracy ML, Johnson LC (1976) Effects of exercise, bedrest and napping on performance decrement during 40 hours. Psychophysiology 13, 334–9. [PubMed] [Google Scholar]
2. Tassi P, Muzet A (2000) Sleep inertia. Sleep Med Rev 4, 341–53. [PubMed] [Google Scholar]
3. Jewett ME, Wyatt JK, Ritz-De Cecco A, Khalsa SB, Dijk DJ, Czeisler CA (1999) Time course of sleep inertia dissipation in human performance and alertness. J Sleep Res 8, 1–8. [PubMed] [Google Scholar]
4. Tietzel AJ, Lack LC (2001) The short-term benefits of brief and long naps following nocturnal sleep restriction. Sleep 24, 293–300. [PubMed] [Google Scholar]
5. Scheer FA, Shea TJ, Hilton MF, Shea SA (2008) An endogenous circadian rhythm in sleep inertia results in greatest cognitive impairment upon awakening during the biological night. J Biol Rhythms 23, 353–61. [PMC free article] [PubMed] [Google Scholar]
6. Ikeda H, Hayashi M (2010) The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol 83, 15–9. [PubMed] [Google Scholar]
7. Transport Accident Commission http://www.tac.vic.gov.au/road-safety/tac-campaigns/fatigue/big-hit. Accessed May 6, 2015.
8. Transport for NSW http://roadsafety.transport.nsw.gov.au/stayingsafe/fatigue/stoprevivesurvive.html. Accessed May 6, 2015.
9. Armentrout JJ, Holland DA, O’Toole KJ, Ercoline WR (2006) Fatigue and related human factors in the near crash of a large military aircraft. Aviat Space Environ Med 77, 963–70. [PubMed] [Google Scholar]
10. Government of India Ministry of Civil Aviation (2010) Report on Accident to Air India Express Boeing 737–800 Aircraft VT-AXV on 22nd May 2010 at Mangalore.
11. Marine Accident Investigation Branch (2011) Heavy Contact by Skandi Foula with OMS Resolution, Aberdeen Harbour 29 May 2010. UK Department for Transport: London, UK. [Google Scholar]
12. Transportation Safety Board of Canada (2011) Aviation Investigation Report - A11F0012.
13. Santhi N, Groeger JA, Archer SN, Gimenez M, Schlangen LJM, Dijk DJ (2013) Morning sleep inertia in alertness and performance: effect of cognitive domain and white light conditions. PLoS One 8, e79688. [PMC free article] [PubMed] [Google Scholar]
14. Burke TM, Scheer FA, Ronda JM, Czeisler CA, Wright KP Jr (2015) Sleep inertia, sleep homeostatic and circadian influences on higher-order cognitive functions. J Sleep Res 24, 364–71. [PMC free article] [PubMed] [Google Scholar]
15. Dinges DF, Orne MT, Orne EC (1985) Assessing performance upon abrupt awakening from naps during quasi-continuous operations. Behav Res Methods 17, 37–45. [Google Scholar]
16. Brooks A, Lack L (2006) A brief afternoon nap following nocturnal sleep restriction: which nap duration is most recuperative? Sleep 29, 831–40. [PubMed] [Google Scholar]
17. Lovato N, Lack L, Ferguson S, Tremaine R (2009) The effects of a 30-min nap during night shift following a prophylactic sleep in the afternoon. Sleep Biol Rhythms 7, 34–42. [Google Scholar]
18. Signal TL, van den Berg MJ, Mulrine HM, Gander PH (2012) Duration of sleep inertia after napping during simulated night work and in extended operations. Chronobiol Int 29, 769–79. [PubMed] [Google Scholar]
19. Miccoli L, Versace F, Koterle S, Cavallero C (2008) Comparing sleep-loss sleepiness and sleep inertia: lapses make the difference. Chronobiol Int 25, 725–44. [PubMed] [Google Scholar]
20. Åkerstedt T. (1988) Sleepiness as a consequence of shift work. Sleep 11, 17–34. [PubMed] [Google Scholar]
21. Rajaratnam SM, Arendt J (2001) Health in a 24-h society. Lancet 358, 999–1005. [PubMed] [Google Scholar]
22. Lumley M, Roehrs T, Zorick F, Lamphere J, Roth T (1986) The alerting effects of naps in sleep-deprived subjects. Psychophysiology 23, 403–8. [PubMed] [Google Scholar]
23. Dinges DF, Orne MT, Whitehouse WG, Orne EC (1987) Temporal placement of a nap for alertness: contributions of circadian phase and prior wakefulness. Sleep 10, 313–29. [PubMed] [Google Scholar]
24. Kubo T, Takeyama H, Matsumoto S, Ebara T, Murata K, Tachi N, Itani T (2007) Impact of nap length, nap timing and sleep quality on sustaining early morning performance. Ind Health 45, 552–63. [PubMed] [Google Scholar]
25. Mulrine HM, Signal TL, van den Berg MJ, Gander PH (2012) Post-sleep inertia performance benefits of longer naps in simulated nightwork and extended operations. Chronobiol Int 29, 1249–57. [PubMed] [Google Scholar]
26. Ruggiero JS, Redeker NS (2014) Effects of napping on sleepiness and sleep-related performance deficits in night-shift workers: a systematic review. Biol Res Nurs 16, 134–42. [PMC free article] [PubMed] [Google Scholar]
27. Frey DJ, Ortega JD, Wiseman C, Farley CT, Wright KP Jr (2011) Influence of zolpidem and sleep inertia on balance and cognition during nighttime awakening: a randomized placebo-controlled trial. J Am Geriatr Soc 59, 73–81. [PubMed] [Google Scholar]
28. Hayashi M, Masuda A, Hori T (2003) The alerting effects of caffeine, bright light and face washing after a short daytime nap. Clin Neurophysiol 114, 2268–78. [PubMed] [Google Scholar]
29. Collins JA, Fauser BC (2005) Balancing the strengths of systematic and narrative reviews. Hum Reprod Update 11, 103–4. [PubMed] [Google Scholar]
30. Reyner LA, Horne JA (1997) Suppression of sleepiness in drivers: combination of caffeine with a short nap. Psychophysiology 34, 721–5. [PubMed] [Google Scholar]
31. Van Dongen HPA, Price NJ, Mullington JM, Szuba MP, Kapoor SC, Dinges DF (2001) Caffeine eliminates psychomotor vigilance deficits from sleep inertia. Sleep 24, 813–9. [PubMed] [Google Scholar]
32. Newman RA, Kamimori GH, Wesensten NJ, Picchioni D, Balkin TJ (2013) Caffeine gum minimizes sleep inertia. Percept Mot Skills 116, 280–93. [PubMed] [Google Scholar]
33. Giménez MC, Hessels M, van de Werken M, de Vries B, Beersma DG, Gordijn MC (2010) Effects of artificial dawn on subjective ratings of sleep inertia and dim light melatonin onset. Chronobiol Int 27, 1219–41. [PubMed] [Google Scholar]
34. Harrison EM, Gorman MR, Mednick SC (2011) The effect of narrowband 500 nm light on daytime sleep in humans. Physiol Behav 103, 197–202. [PubMed] [Google Scholar]
35. Van De Werken M, Giménez MC, De Vries B, Beersma DG, Van Someren EJ, Gordijn MC (2010) Effects of artificial dawn on sleep inertia, skin temperature, and the awakening cortisol response. J Sleep Res 19, 425–35. [PubMed] [Google Scholar]
36. Thompson A, Jones H, Gregson W, Atkinson G (2014) Effects of dawn simulation on markers of sleep inertia and post-waking performance in humans. Eur J Appl Physiol 114, 1049–56. [PubMed] [Google Scholar]
37. Tassi P, Nicolas A, Dewasmes G, Eschenlauer R, Ehrhart J, Salame P, Muzet A, Libert JP (1992) Effects of noise on sleep inertia as a function of circadian placement of a one-hour nap. Percept Mot Skills 75, 291–302. [PubMed] [Google Scholar]
38. Hayashi M, Uchida C, Shoji T, Hori T (2004) The effects of the preference for music on sleep inertia after a short daytime nap. Sleep Biol Rhythms 2, 184–91. [Google Scholar]
39. Kräuchi K, Cajochen C, Wirz-Justice A (2004) Waking up properly: is there a role of thermoregulation in sleep inertia? J Sleep Res 13, 121–7. [PubMed] [Google Scholar]
40. Kräuchi K, Knoblauch V, Wirz-Justice A, Cajochen C (2006) Challenging the sleep homeostat does not influence the thermoregulatory system in men: evidence from a nap vs. sleep-deprivation study. Am J Physiol Regul Integr Comp Physiol 290, R1052–61. [PubMed] [Google Scholar]
41. Kaida K, Nittono H, Hayashi M, Hori T (2003) Effects of self-awakening on sleep structure of a daytime short nap and on subsequent arousal levels. Percept Mot Skills 97, 1073–84. [PubMed] [Google Scholar]
42. Kaida K, Nakano E, Nittono H, Hayashi M, Hori T (2003) The effects of self-awakening on heart rate activity in a short afternoon nap. Clin Neurophysiol 114, 1896–901. [PubMed] [Google Scholar]
43. Ikeda H, Kubo T, Kuriyama K, Takahashi M (2014) Self-awakening improves alertness in the morning and during the day after partial sleep deprivation. J Sleep Res 23, 673–80. [PubMed] [Google Scholar]
44. Nehlig A. (1999) Are we dependent upon coffee and caffeine? A review on human and animal data. Neurosci Biobehav Rev 23, 563–76. [PubMed] [Google Scholar]
45. Dorrian J, Paterson J, Dawson D, Pincombe J, Grech C, Rogers AE (2011) Sleep, stress and compensatory behaviors in Australian nurses and midwives. Rev Saude Publica 45, 922–30. [PubMed] [Google Scholar]
46. Fredholm BB, Bättig K, Holmén J, Nehlig A, Zvartau EE (1999) Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev 51, 83–133. [PubMed] [Google Scholar]
47. Wesensten NJ, Belenky G, Kautz MA, Thorne DR, Reichardt RM, Balkin TJ (2002) Maintaining alertness and performance during sleep deprivation: modafinil versus caffeine. Psychopharmacology (Berl) 159, 238–47. [PubMed] [Google Scholar]
48. Wyatt JK, Cajochen C, Ritz-De Cecco A, Czeisler CA, Dijk DJ (2004) Low-dose repeated caffeine administration for circadian-phase-dependent performance degradation during extended wakefulness. Sleep 27, 374–81. [PubMed] [Google Scholar]
49. Kamimori GH, Karyekar CS, Otterstetter R, Cox DS, Balkin TJ, Belenky GL, Eddington ND (2002) The rate of absorption and relative bioavailability of caffeine administered in chewing gum versus capsules to normal healthy volunteers. Int J Pharm 234, 159–67. [PubMed] [Google Scholar]
50. Allen AP, Jacob TJ, Smith AP (2014) Effects and after-effects of chewing gum on vigilance, heart rate, EEG and mood. Physiol Behav 133, 244–51. [PubMed] [Google Scholar]
51. Morgan K, Johnson AJ, Miles C (2014) Chewing gum moderates the vigilance decrement. Br J Psychol 105, 214–25. [PubMed] [Google Scholar]
52. Brice CF, Smith AP (2002) Factors associated with caffeine consumption. Int J Food Sci Nutr 53, 55–64. [PubMed] [Google Scholar]
53. Mandel HG. (2002) Update on caffeine consumption, disposition and action. Food Chem Toxicol 40, 1231–4. [PubMed] [Google Scholar]
54. Horne JA, Reyner LA (1996) Counteracting driver sleepiness: effects of napping, caffeine, and placebo. Psychophysiology 33, 306–9. [PubMed] [Google Scholar]
55. Department for Transport http://think.direct.gov.uk/fatigue.html. Accessed May 6, 2015.
56. Juliano LM, Griffiths RR (2004) A critical review of caffeine withdrawal: empirical validation of symptoms and signs, incidence, severity, and associated features. Psychopharmacology (Berl) 176, 1–29. [PubMed] [Google Scholar]
57. Dorrian J, Baulk SD, Dawson D (2011) Work hours, workload, sleep and fatigue in Australian Rail Industry employees. Appl Ergon 42, 202–9. [PubMed] [Google Scholar]
58. McHill AW, Smith BJ, Wright KP Jr (. 2014) Effects of caffeine on skin and core temperatures, alertness, and recovery sleep during circadian misalignment. J Biol Rhythms 29, 131–43. [PubMed] [Google Scholar]
59. Cagnacci A, Elliott JA, Yen SS (1992) Melatonin: a major regulator of the circadian rhythm of core temperature in humans. J Clin Endocrinol Metab 75, 447–52. [PubMed] [Google Scholar]
60. Czeisler CA, Allan JS, Strogatz SH, Ronda JM, Sánchez R, Ríos CD, Freitag WO, Richardson GS, Kronauer RE (1986) Bright light resets the human circadian pacemaker independent of the timing of the sleep-wake cycle. Science 233, 667–71. [PubMed] [Google Scholar]
61. Wever RA. (1989) Light effects on human circadian rhythms: a review of recent Andechs experiments. J Biol Rhythms 4, 161–85. [PubMed] [Google Scholar]
62. Rosenthal NE, Joseph-Vanderpool JR, Levendosky AA, Johnston SH, Allen R, Kelly KA, Souetre E, Schultz PM, Starz KE (1990) Phase-shifting effects of bright morning light as treatment for delayed sleep phase syndrome. Sleep 13, 354–61. [PubMed] [Google Scholar]
63. Eastman CI, Martin SK (1999) How to use light and dark to produce circadian adaptation to night shift work. Ann Med 31, 87–98. [PubMed] [Google Scholar]
64. Campbell SS, Dawson D (1990) Enhancement of nighttime alertness and performance with bright ambient light. Physiol Behav 48, 317–20. [PubMed] [Google Scholar]
65. French J, Hannon P, Brainard GC (1990) Effects of bright illuminance on body temperature and human performance. Ann Rev Chronopharmacol 7, 37–40. [Google Scholar]
66. Phipps-Nelson J, Redman JR, Dijk DJ, Rajaratnam SM (2003) Daytime exposure to bright light, as compared to dim light, decreases sleepiness and improves psychomotor vigilance performance. Sleep 26, 695–700. [PubMed] [Google Scholar]
67. Vandewalle G, Balteau E, Phillips C, Degueldre C, Moreau V, Sterpenich V, Albouy G, Darsaud A, Desseilles M, Dang-Vu TT, Peigneux P, Luxen A, Dijk DJ, Maquet P (2006) Daytime light exposure dynamically enhances brain responses. Curr Biol 16, 1616–21. [PubMed] [Google Scholar]
68. Cajochen C. (2007) Alerting effects of light. Sleep Med Rev 11, 453–64. [PubMed] [Google Scholar]
69. Sherman B, Wysham C, Pfohl B (1985) Age-related changes in the circadian rhythm of plasma cortisol in man. J Clin Endocrinol Metab 61, 439–43. [PubMed] [Google Scholar]
70. Clow A, Hucklebridge F, Stalder T, Evans P, Thorn L (2010) The cortisol awakening response: more than a measure of HPA axis function. Neurosci Biobehav Rev 35, 97–103. [PubMed] [Google Scholar]
71. Scheer FA, Buijs RM (1999) Light affects morning salivary cortisol in humans. J Clin Endocrinol Metab 84, 3395–8. [PubMed] [Google Scholar]
72. Thorn L, Hucklebridge F, Esgate A, Evans P, Clow A (2004) The effect of dawn simulation on the cortisol response to awakening in healthy participants. Psychoneuroendocrinology 29, 925–30. [PubMed] [Google Scholar]
73. Thorn L, Hucklebridge F, Evans P, Clow A (2009) The cortisol awakening response, seasonality, stress and arousal: a study of trait and state influences. Psychoneuroendocrinology 34, 299–306. [PubMed] [Google Scholar]
74. Dahlgren A, Kecklund G, Theorell T, Åkerstedt T (2009) Day-to-day variation in saliva cortisol--relation with sleep, stress and self-rated health. Biol Psychol 82, 149–55. [PubMed] [Google Scholar]
75. Vgontzas AN, Mastorakos G, Bixler EO, Kales A, Gold PW, Chrousos GP (1999) Sleep deprivation effects on the activity of the hypothalamic-pituitary-adrenal and growth axes: potential clinical implications. Clin Endocrinol (Oxf) 51, 205–15. [PubMed] [Google Scholar]
76. Tassi P, Bonnefond A, Engasser O, Hoeft A, Eschenlauer R, Muzet A (2006) EEG spectral power and cognitive performance during sleep inertia: the effect of normal sleep duration and partial sleep deprivation. Physiol Behav 87, 177–84. [PubMed] [Google Scholar]
77. Achermann P, Werth E, Dijk DJ, Borbely AA (1995) Time course of sleep inertia after nighttime and daytime sleep episodes. Arch Ital Biol 134, 109–19. [PubMed] [Google Scholar]
78. Stewart KT, Hayes BC, Eastman CI (1995) Light treatment for NASA shiftworkers. Chronobiol Int 12, 141–51. [PubMed] [Google Scholar]
79. Boivin DB, James FO (2002) Circadian adaptation to night-shift work by judicious light and darkness exposure. J Biol Rhythms 17, 556–67. [PubMed] [Google Scholar]
80. Budnick LD, Lerman SE, Nicolich MJ (1995) An evaluation of scheduled bright light and darkness on rotating shiftworkers: trial and limitations. Am J Ind Med 27, 771–82. [PubMed] [Google Scholar]
81. Wright HR, Lack LC, Partridge KJ (2001) Light emitting diodes can be used to phase delay the melatonin rhythm. J Pineal Res 31, 350–5. [PubMed] [Google Scholar]
82. Kawada T, Suzuki S (1993) Sleep induction effects of steady 60 dB (A) pink noise. Ind Health 31, 35–8. [PubMed] [Google Scholar]
83. Zhou J, Liu D, Li X, Ma J, Zhang J, Fang J (2012) Pink noise: effect on complexity synchronization of brain activity and sleep consolidation. J Theor Biol 306, 68–72. [PubMed] [Google Scholar]
84. Wilkinson RT. (1963) Interaction of noise with knowledge of results and sleep deprivation. J Exp Psychol 66, 332–7. [PubMed] [Google Scholar]
85. Poulton EC. (1976) Arousing environmental stresses can improve performance, whatever people say. Aviat Space Environ Med 47, 1193–204. [Google Scholar]
86. Koelega HS, Brinkman JA (1986) Noise and vigilance: an evaluative review. Hum Factors 28, 465–81. [PubMed] [Google Scholar]
87. Hockey GRJ. (1970) Effect of loud noise on attentional selectivity. Q J Exp Psychol 22, 28–36. [Google Scholar]
88. Hancock PA. (1984) Environmental stressors. In: Sustained attention in human performance, Warm JS (Ed.), Wiley, New York. [Google Scholar]
89. Melamed S, Bruhis S (1996) The effects of chronic industrial noise exposure on urinary cortisol, fatigue and irritability: a controlled field experiment. J Occup Environ Med 38, 252–6. [PubMed] [Google Scholar]
90. Kjellberg A, Muhr P, Sköldström B (1998) Fatigue after work in noise—an epidemiological survey study and three quasi-experimental field studies. Noise Health 1, 47–55. [PubMed] [Google Scholar]
91. Reyner LA, Horne JA (1998) Evaluation “in-car” countermeasures to sleepiness: cold air and radio. Sleep 21, 46–50. [PubMed] [Google Scholar]
92. Penn PE, Bootzin RR (1990) Behavioural techniques for enhancing alertness and performance in shift work. Work Stress 4, 213–26. [Google Scholar]
93. Lack LC, Gradisar M, Van Someren EJ, Wright HR, Lushington K (2008) The relationship between insomnia and body temperatures. Sleep Med Rev 12, 307–17. [PubMed] [Google Scholar]
94. Lack L, Gradisar M (2002) Acute finger temperature changes preceding sleep onsets over a 45-h period. J Sleep Res 11, 275–82. [PubMed] [Google Scholar]
95. Raymann RJ, Swaab DF, Van Someren EJ (2008) Skin deep: enhanced sleep depth by cutaneous temperature manipulation. Brain 131, 500–13. [PubMed] [Google Scholar]
96. Fronczek R, Raymann RJ, Romeijn N, Overeem S, Fischer M, van Dijk JG, Lammers GJ, Van Someren EJ (2008) Manipulation of core body and skin temperature improves vigilance and maintenance of wakefulness in narcolepsy. Sleep 31, 233–40. [PMC free article] [PubMed] [Google Scholar]
97. Kräuchi K, Cajochen C, Werth E, Wirz-Justice A (1999) Warm feet promote the rapid onset of sleep. Nature 401, 36–7. [PubMed] [Google Scholar]
98. Kräuchi K, Cajochen C, Wirz-Justice A (2005) Thermophysiologic aspects of the three-process-model of sleepiness regulation. Clin Sports Med 24, 287–300, ix. [PubMed] [Google Scholar]
99. Bruck D, Pisani DL (1999) The effects of sleep inertia on decision-making performance. J Sleep Res 8, 95–103. [PubMed] [Google Scholar]
100. Dorrian J, Lamond N, Dawson D (2000) The ability to self-monitor performance when fatigued. J Sleep Res 9, 137–44. [PubMed] [Google Scholar]
101. Dorrian J, Lamond N, Holmes AL, Burgess HJ, Roach GD, Fletcher A, Dawson D (2003) The ability to self-monitor performance during a week of simulated night shifts. Sleep 26, 871–7. [PubMed] [Google Scholar]
102. Biggs SN, Smith A, Dorrian J, Reid K, Dawson D, van den Heuvel C, Baulk S (2007) Perception of simulated driving performance after sleep restriction and caffeine. J Psychosom Res 63, 573–7. [PubMed] [Google Scholar]
103. Tremaine R, Dorrian J, Lack L, Lovato N, Ferguson S, Zhou X, Roach G (2010) The relationship between subjective and objective sleepiness and performance during a simulated night-shift with a nap countermeasure. Appl Ergon 42, 52–61. [PubMed] [Google Scholar]
104. Hilditch CJ, Dorrian J, Centofanti SA, Van Dongen HP, Banks S (2015) Sleep inertia associated with a 10-min nap before the commute home following a night shift: A laboratory simulation study. Accid Anal Prev S0001-4575(15)30126-3; Epub ahead of print (In press). [PubMed] [Google Scholar]
105. Hori T, Hayashi M, Morikawa T (1994) Topographical EEG changes and the hypnagogic experience. In: Sleep onset: Normal and abnormal processes, Ogilvie R and Harsh JR (Eds.), 237–53, American Psychological Association, Washington, DC. [Google Scholar]
106. Rechtschaffen A, Kales A (1968) A manual of standardized terminology, techniques and scoring system for sleep stages of human subjects. Los Angeles: Brain Information Service/Brain Research Institute, University of California. [PubMed] [Google Scholar]
107. Ikeda H, Hayashi M (2010) The effect of self-awakening from nocturnal sleep on sleep inertia. Biol Psychol 83, 15–9. [PubMed] [Google Scholar]
108. Smith AP, Clark R, Gallagher J (1999) Breakfast cereal and caffeinated coffee: effects on working memory, attention, mood, and cardiovascular function. Physiol Behav 67, 9–17. [PubMed] [Google Scholar]
109. Matter R, Carroll J, Dyer C (2000) Neuroendocrine responses to stress. In: The biology of animal stress: basic principles and implications for animal welfare, Moberg GP and Mench JA (Eds.), 43–76, CABI Publishing, New York. [Google Scholar]
110. Rosekind MR, Solutions A (2008) Managing fatigue in EMS flight operations: challenges and opportunities. Alertness Solutions White Paper.
111. Ferreira SH, Vane JR (1967) Half-lives of peptides and amines in the circulation. Nature 215, 1237–40. [PubMed] [Google Scholar]
112. Weeks SR, McAuliffe CL, Durussel D, Pasquina PF (2010) Physiological and psychological fatigue in extreme conditions: the military example. PM R 2, 438–41. [PubMed] [Google Scholar]
113. Gregory KB, Winn W, Johnson K, Rosekind MR (2010) Pilot fatigue survey: exploring fatigue factors in air medical operations. Air Med J 29, 309–19. [PubMed] [Google Scholar]
114. Fallis WM, McMillan DE, Edwards MP (2011) Napping during night shift: practices, preferences, and perceptions of critical care and emergency department nurses. Crit Care Nurse 31, e1–11. [PubMed] [Google Scholar]
Articles from Industrial Health are provided here courtesy of National Institute of Occupational Safety and Health, Japan
OTHER FORMATS
PDF (1.6M)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 401166 | https://no.wikipedia.org/wiki/D%C3%B8gnrytmeforstyrrelse | Døgnrytmeforstyrrelse | Døgnrytmeforstyrrelser eller døgnrytmelidelser er oftest sett på som en type søvnforstyrrelse. Søvn-våkenhetsproblemer er de mest merkbare av problemkomplekset, men døgnrytmene omfatter mange flere fysiologiske og atferdsmessige faktorer enn som så, bl.a. tidspunkt for utskillelse av ulike hormoner og appetitt. Mennesker med døgnrytmeforstyrrelser kan ikke sovne og våkne på tidspunkt som samfunnet krever for skole, arbeid og sosiale gjøremål. Med mindre de har en annen søvnforstyrrelse i tillegg, har de søvn av normal kvalitet.
Hadde mennesker blitt mer og mer søvnig for hver time som går siden de stod opp, hadde de ikke kunnet være våkne i de vanlige 16 timene pr. dag. Mot den tendensen arbeider individets døgnrytme, som normalt holder det våkent til godt utpå kvelden. Både A- og B-mennesker er normale, og det store flertallet er innimellom. Noen er rettnok mer fleksible enn andre. Mennesker med døgnrytmeforstyrrelser er lite fleksible i så måte, og døgnrytmen ligger utenfor det normale.
De seks døgnrytmeforstyrrelser
Det finnes seks typer døgnrytmeforstyrrelse, der de to førstnevnte, jetlag og skiftarbeid-syndrom, er midlertidige og påført utenfra.
De fire siste er kroniske, og ICD (International Classification of Diseases), så vel som MeSH, registrerer dem som Sykdommer i nervesystemet. Grunnen til søvnfaseforskyvningene er ikke helt klarlagt. De går i en viss grad igjen i slekten og ser ut til å være genetisk betinget (hPer2- og hPer3-genene). Noen tilfelle er oppstått etter hodeskade.
Døgnrytmens syklus, eller periode, kan være betydelig kortere eller, oftest, lengre enn 24 timer. Effekten av lys til øyets netthinne, som justerer døgnrytmen, kan være redusert eller i feil fase. Pasientene har mye større vanskeligheter med å justere døgnrytmen enn friske mennesker; friske som har spesielt vanskelig for å bli vant til skiftarbeid, kan ha til dels lignende plager. I ICSD's definisjoner av forsinket søvnfasesyndrom (s. 128–133), fremskutt søvnfasesyndrom (s. 134) og ikke-24-time søvn–våken-syndrom (s. 138) er betegnelse «intractable and chronic» brukt; altså kronisk og uregjerlig.
En annen lidelse som kan ha visse likhetstrekk med døgnrytmeforstyrrelser, også når det gjelder behandling, er sesongavhengig depresjon, men denne er fremdeles klassifisert som en psykisk lidelse, ikke en sykdom i nervesystemet.
Jetlag
Jetlag er et uttrykk for den fysiologiske reaksjonen som inntreffer når man endrer kroppens døgnrytme ved å reise gjennom flere tidssoner ved fly.
Skiftarbeidsyndrom
Skiftarbeidsyndrom omfatter problem som skiftarbeidere får med døgnrytmen og søvnen. Skiftarbeid inkluderer fast nattskift så vel som roterende skiftarbeid.
Forsinket søvnfasesyndrom
Kronisk type
Forsinket søvnfasesyndrom er en tilstand hvor kroppens klokke er stilt inn til å fungere noen timer senere enn hos normale mennesker. Syndromet oppstår enten tidlig i barndommen eller i puberteten – svært sjelden etter 20-årsalderen. Ca. 0,15 % av voksne har syndromet, likelig fordelt mellom kvinner og menn. Alle markørene for døgnrytme er affiserte: kroppstemperatur, melatonin, kortisol, appetitt m.m. så vel som søvnfase. Søvnen er normal, og søvnrytmen er regelmessig, bare forsinket; dvs. at rytmen justeres til naturens døgn på 24 timer, bare til «feil» tid. Søvnen kommer ikke før kl. 02–06, og det er svært vanskelig å komme seg opp av sengen før 8 timer senere. Diagnosen forsinket søvnfasesyndrom gis ikke hvis man på egen hånd kan normalisere døgnrytmen.
Forsinket søvnfasesyndrom er kronisk for voksne. Det blir ikke kurert, men kan bli forsøkt behandlet (se nedenfor). Det er ofte feildiagnostisert, da det fremdeles er mange leger som ikke er klar over problem med døgnrytmen.
Opptil 50 % av dem som er beskrevet i litteraturen, har eller har hatt depresjon eller en annen psykisk lidelse, omtrent tilsvarende forekomsten av psykiske lidelser blant pasienter med kronisk insomnia. Da vel 50 % ikke har psykiske problemer, er det klart at slike lidelser ikke kan være den eneste årsaken til forsinket søvnfasesyndrom og ikke nødvendigvis er en årsak i det hele tatt. Å prøve å fungere i 08–16-samfunnet fører som oftest til søvnmangel for disse pasientene, og nyere forskning viser at nettopp søvnmangel kan forårsake psykiske lidelser.
Bruk av søvnmedisiner over lengre tid kan tvinge søvnmønstret på plass. Dette fungerer ikke for alle og er ikke en god løsning pga. bivirkninger som avhengighet, depresjon, angst, kvalme og varig søvnmangel. Medisiner som modafinil (markedsført som blant annet Provigil, Vigil og Modiodal) kan brukes i tillegg for bedre våkenhet på dagtid; modafinil er godkjent i USA for skiftarbeidere i arbeidstiden.
Syndromet ble først beskrevet i 1981 av Dr. Elliot D. Weitzman mfl. Norge er et av de landene som bruker forholdsvis mye midler på forskning på området, blant annet på Universitetet i Bergen.
Voksne med forsinket søvnfasesyndrom kalles gjerne B- eller også C-mennesker.
Ungdomstype
Mange flere ungdommer enn voksne har forsinket søvnfasesyndrom, opp mot 7 %, flere gutter enn jenter. I tillegg kommer de som velger B-menneske-livsstilen. De fleste «vokser det av seg» i 20-årsalderen.
Søvnmangelen går fort utover konsentrasjon, sosiale og atletiske prestasjoner, skolegang, jobb, og livet generelt. Barn og ungdom er spesielt utsatt for misdiagnoser av mental karakter, for eksempel skolevegring.
Fremskutt søvnfasesyndrom
Fremskutt søvnfasesyndrom kan sies å være det motsatte av forsinket søvnfasesyndrom. Det er mye sjeldnere. Mennesker med dette syndromet er nesten uimotståelig søvnige tidlig på kvelden og legger seg rundt kl. 19–20. De har normal søvn og våkner etter de vanlige 8 timene. Dette syndromet går svært tydelig igjen i familier. To ulike mutasjoner i hPer2-genet er avslørt av Louis Ptáčeks forskningsteam ved University of California; ingen av de to mutasjonene er funnet i mennesker som ikke har syndromet.
Ikke-24-time søvn–våken-syndrom
Også kalt hypernychthemeral syndrom, men ofte kalt bare «Non-24», dette syndromet ligner på forsinket søvnfasesyndrom. Det er dog vanskeligere å leve med, da pasientene ikke kan justere kroppsrytmene til naturens 24 timer. De våkner opptil 1–2 timer senere hver dag og sovner tilsvarende senere hver dag. Etter noen uker kommer de tilbake til utgangspunktet. Behandlingen er den samme som for forsinket søvnfasesyndrom, men er enda sjeldnere vellykket.
De fleste tilfeller, men ikke alle, er blant blinde.
Irregulært søvn–våken-mønster
Søvnen varierer fra dag til dag, vanligvis i minst tre søvnepisoder pr. døgn, som dog tilsvarer den normale mengden søvn for alderen. Grunnen kan være feil med døgnrytmen, ofte som følge av demens.
Diagnostiske verktøy
Polysomnografi
Polysomnografi er en søvntest; resultatet kalles en polysomnogram (PSG). Mange kroppsfunksjoner er målt mens pasienten sover: hjerneaktivitet, øyeaktivitet, muskulær aktivitet, pustefunksjon og oksygennivået i blodet, m.m. Når testen er brukt i tilfeller av døgnrytmeforstyrrelser, er det helst for å utelukke andre søvnforstyrrelser så som narkolepsi eller søvnapné.
MSLT (Multiple Sleep Latency Test)
MSLT er en enkel test for søvnighet, som regel gjort dagen etter polysomnografi. Pasienten må legge seg kl. 10, kl. 12, kl. 14 og kl. 16 i 20 minutter hver gang. De samme målingene som ved polysomnografi blir gjort. Av spesiell interesse er hvor raskt søvn er oppnådd og hvilke søvnstadier pasienten er innom.
Søvndagbok eller -logg
Sammen med klinisk intervju, er søvndagboken det viktigste verktøyet i diagnostikk av døgnrytmeforstyrrelser. Den ser ofte ut som en kalenderside med ruter for hver dag der pasienten skal skrive tidspunkt for å legge seg, sovne, ev. våkenepisoder i løpet av natten, våkne, stå opp, om man våknet spontant eller av vekkerklokke o.l., medikamenter m.m. Søvnspesialister og klinikker har gjerne utviklet hvert sitt skjema.
Aktigrafi
Aktigrafi for søvn bruker et lite elektronisk måleinstrument som ligner på et armbandsur. Pasienten har det på den ikke-dominante armen i en eller flere uker. Det har et minne, og det måler i det minste armbevegelser. Noen av dem måler også lysnivået. Det har gjerne en knapp som pasienten trykker på for å indikere spesifikke hendelser, som medisininntak og leggetid.
Aktigrafimålinger viser søvnmønstre og er brukt i diagnostikk – ofte i samband med søvndagboka, i vurdering av effekten av visse terapier og i forskning. Aktigrafi kan bli brukt i tillegg til eller i stedet for polysomnografi i ulike situasjoner.
Klinisk intervju
Søvnspesialisten, som ofte er en nevrolog, barnelege eller psykiater, spør detaljert om pasientens vaner og historikk.
Behandling
Innen pasienter søker medisinsk hjelp, har de fleste prøvd mange ganger og på mange måter å endre søvnmønsteret. Dette gjelder spesielt de som sover på dagtid, da dette er noe som er dårlig akseptert av samfunnet. Pasientene har gjerne blitt fortalt så mange ganger at de er late og mangler viljestyrke, at de kan ha begynt å tro at dette er sant.
For mange pasienter er det å få en diagnose i seg selv noe stort.
Søvnspesialisten forklarer hvilke terapier som kan forsøkes om pasienten vil prøve å endre døgnrytmen, og understreker at ingen av dem er garantert å virke. Rehabilitering til et annet yrke som bedre passer til søvnmønstret, drøftes. Hvis pasienten studerer, er kveldsskole eller studier online en mulighet.
Flere metoder kan brukes når man vil justere døgnrytmen. For de som vil få sove tidligere kan kronoterapi brukes, helst i samarbeid med en søvnspesialist. De andre metodene som kan brukes hver for seg eller i kombinasjon er sterk lys til øynene, svært lite lys ev. med bruk av oransje laserbriller, og små doser av hormonet melatonin—alle til riktig tid for det resultatet man er ute etter.
Kronoterapi
Ved kronoterapi forsøker man å flytte døgnrytmen «rundt klokken», litt etter litt. Dette krever disiplin og helst en samarbeidsvillig familie. Resultatet er sjelden varig, da sykdom, sykt barn, festligheter e.l. vil tvinge mange tilbake til den gamle naturlige rytmen, som gjerne vil feste seg på nytt.
For forsinket søvnfasesyndrom, for eksempel, er det to måter å gjøre dette på.
Senere hver dag
Pasienten går til sengs 2–3 timer senere hver dag inntil ønsket leggetid nås. Man kan f.eks. legge seg kl. 06 en dag og kl. 09, 12, 15, 18 og 21 de påfølgende dagene. Man legger seg deretter kl. 21 i flere dager for å feste den nye rutinen. Man kan så la leggetiden «krype» langsomt til ønsket tid, f.eks. kl. 22.30. Det er tøft å holde seg til 27-timers "døgn", men man bør ikke bruke mer enn en uke på denne behandlingen da man kan risikere å utvikle "ikke-24-time søvn–våken-syndrom" (se nedenfor) som er enda vanskeligere å leve med. Man bør snakke med en søvnspesialist på forehand.
Tidligere en gang i uken
Pasienten sover så regelmessig som mulig i kroppens naturlige rytmen i en uke. Den 6. dagen er man oppe en hel dag, en hel natt og hele dagen deretter. Den natten går man til sengs 90 minutter tidligere enn vanlig og beholder denne leggetiden i 6 netter. Dette repeteres i så mange uker som er nødvendig for å nå ønsket leggetid.
Lysterapi og begrenset belysning, og/eller melatonin
Både sterkt lys til øynene og tilførsel av hormonet melatonin, flytende eller i tablettform, og begrenset lys påvirker døgnrytmen, alt etter tidspunktet på døgnet. Presise tidspunkt er såpass individuelle at en del eksperimentering må som regel til. Behandlingene må brukes daglig for å ha noe særlig effekt.
Illustrasjonen viser fase-respons kurvene (PRC, phase response curves) for menneske for sterkt og svakt lys (de mørke og lyse lilla fargene) og for melatonin (grøn).
Søvnperioden refererer til ikke bare ett tilfeldig døgn, men til den regelmessige, daglige perioden uten bruk av vekkerklokke e.l. De positive og negative tallene til venstre viser hvor mange timer fremskyndelse eller forsinkelse man kan regne med ved påførsel av den aktuelle terapien til det aktuelle tidspunktet.
Lys til øynene like etter spontanoppvåkning fører til at man kan sovne tidligere om kveldene enn vanlig, spesielt hvis man begrenser lys i timene før leggetid. Omvendt, lys om kvelden fører til oppvåkning senere om morgenene. Som man ser av kurven, vil sterkt lys med en gang etter oppvåkning gi opptil to timer fremskyndelse av søvnfasen, mens bare en time senere er virkningen nesten halvert.
For de som ønsker å få sove tidligere kan inntak av hormonet melatonin like før leggetid hjelpe for innsovningen (svak hypnotisk effekt), men har da liten effekt på døgnrytmen. Derimot kan melatonin flere timer før leggetid ha en lignende effekt som lys ved oppvåkning, bare noe mindre. Tilsvarende vil melatonin like etter oppvakning føre til at leggetid kan utsettes litt om kvelden. Effekten av de to behandlingsformene er additiv, dvs. at en person som greier å finne de optimale tidspunktene for behandlingene og er normalt sensitiv til fase-skifte ved lys og melatonin, vil kunne fremskynde søvnfasen med opptil tre timer. Behandlingen må som regel brukes daglig.
Doser på 3 mg melatonin var lenge standard, men forskning de siste årene har vist at 1 mg eller enda mindre kan være like (eller mer) effektiv. Tilsvarende er lysterapi med rundt 10 000 lux standard, mens styrke på det halve og enda mindre kan være effektiv. (Vanlig innendørs belysning er vanligvis godt under 500 lux.)
Melatonin er reseptbelagt eller ikke tilgjengelig i Europa, reseptbelagt på registreringsfritak i Norge. I USA og Canada kalles hormonet for et kostholdssupplement, og det kan der kjøpes fritt.
Da blått og til dels grønt lys har størst effekt på melatonin, kan de som vil få sove tidlegere bruke oransje laserbriller om kveldene for ikke å ødelegge hormonet i sirkulasjon i kroppen.
Referanser
Eksterne lenker
– Opplysninger, på engelsk, ved og for pasienter med Delayed Sleep-Phase Syndrome og pårørende
– Informasjon om døgnrytmelidelser, på engelsk
Søvnforstyrrelser
Nevrologi
Sykdommer i nervesystemet
Nevrofysiologi | norwegian_bokmål | 0.66941 |
neuron_onto/31theneuronisthebuil.txt | Skip to content
[ 
](https://opentextbc.ca/)
Toggle Menu
Primary Navigation
* [ Home ](https://opentextbc.ca/introductiontopsychology)
* [ Read ](https://opentextbc.ca/introductiontopsychology/front-matter/about-bccampus/)
* [ Sign in ](https://opentextbc.ca/introductiontopsychology/wp-login.php?redirect_to=https%3A%2F%2Fopentextbc.ca%2Fintroductiontopsychology%2Fchapter%2F3-1-the-neuron-is-the-building-block-of-the-nervous-system%2F)
* Search in book: Search
Want to create or adapt books like this? [ Learn more
](https://pressbooks.com/adapt-open-
textbooks?utm_source=book&utm_medium=banner&utm_campaign=bbc) about how
Pressbooks supports open publishing practices. [ ](javascript:void\(\))
Book Contents Navigation
Contents
1. [ About BCcampus Open Education ](https://opentextbc.ca/introductiontopsychology/front-matter/about-bccampus/)
2. [ Approach and Pedagogy ](https://opentextbc.ca/introductiontopsychology/front-matter/approach-and-pedagogy/)
Charles Stangor
3. [ Acknowledgments ](https://opentextbc.ca/introductiontopsychology/front-matter/acknowledgments/)
4. [ Dedication ](https://opentextbc.ca/introductiontopsychology/front-matter/dedication/)
Charles Stangor
5. Chapter 1. Introducing Psychology
1. [ 1\. Introducing Psychology ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-1-introducing-psychology/)
2. [ 1.1 Psychology as a Science ](https://opentextbc.ca/introductiontopsychology/chapter/1-1-psychology-as-a-science/)
3. [ 1.2 The Evolution of Psychology: History, Approaches, and Questions ](https://opentextbc.ca/introductiontopsychology/chapter/1-2-the-evolution-of-psychology-history-approaches-and-questions/)
4. [ 1.3 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/1-3-chapter-summary/)
6. Chapter 2. Introduction to Major Perspectives
1. [ 2\. Introduction to Major Perspectives ](https://opentextbc.ca/introductiontopsychology/chapter/2-introduction-to-major-perspectives/)
Jennifer Walinga
2. [ 2.1 Biological Psychology ](https://opentextbc.ca/introductiontopsychology/chapter/2-1-biological-psychology-structuralism-and-functionalism/)
Jennifer Walinga
3. [ 2.2 Psychodynamic Psychology ](https://opentextbc.ca/introductiontopsychology/chapter/2-2-psychodynamic-and-behavioural-psychology/)
Jennifer Walinga
4. [ 2.3 Behaviourist Psychology ](https://opentextbc.ca/introductiontopsychology/chapter/2-3-behaviourist-psychology/)
Jennifer Walinga
5. [ 2.4 Humanist, Cognitive, and Evolutionary Psychology ](https://opentextbc.ca/introductiontopsychology/chapter/2-4-humanist-cognitive-and-evolutionary-psychology/)
Jennifer Walinga
6. [ 2.5 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/2-5-chapter-summary/)
Jennifer Walinga
7. Chapter 3. Psychological Science
1. [ 3\. Psychological Science ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-2-psychological-science/)
2. [ 3.1 Psychologists Use the Scientific Method to Guide Their Research ](https://opentextbc.ca/introductiontopsychology/chapter/2-1-psychologists-use-the-scientific-method-to-guide-their-research/)
3. [ 3.2 Psychologists Use Descriptive, Correlational, and Experimental Research Designs to Understand Behaviour ](https://opentextbc.ca/introductiontopsychology/chapter/2-2-psychologists-use-descriptive-correlational-and-experimental-research-designs-to-understand-behavior/)
4. [ 3.3 You Can Be an Informed Consumer of Psychological Research ](https://opentextbc.ca/introductiontopsychology/chapter/2-3-you-can-be-an-informed-consumer-of-psychological-research/)
5. [ 3.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/2-4-chapter-summary/)
8. Chapter 4. Brains, Bodies, and Behaviour
1. [ 4\. Brains, Bodies, and Behaviour ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-3-brains-bodies-and-behavior/)
2. [ 4.1 The Neuron Is the Building Block of the Nervous System ](https://opentextbc.ca/introductiontopsychology/chapter/3-1-the-neuron-is-the-building-block-of-the-nervous-system/)
3. [ 4.2 Our Brains Control Our Thoughts, Feelings, and Behaviour ](https://opentextbc.ca/introductiontopsychology/chapter/3-2-our-brains-control-our-thoughts-feelings-and-behavior/)
4. [ 4.3 Psychologists Study the Brain Using Many Different Methods ](https://opentextbc.ca/introductiontopsychology/chapter/3-3-psychologists-study-the-brain-using-many-different-methods/)
5. [ 4.4 Putting It All Together: The Nervous System and the Endocrine System ](https://opentextbc.ca/introductiontopsychology/chapter/3-4-putting-it-all-together-the-nervous-system-and-the-endocrine-system/)
6. [ 4.5 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/3-5-chapter-summary/)
9. Chapter 5. Sensing and Perceiving
1. [ 5\. Sensing and Perceiving ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-4-sensing-and-perceiving/)
2. [ 5.1 We Experience Our World through Sensation ](https://opentextbc.ca/introductiontopsychology/chapter/4-1-we-experience-our-world-through-sensation/)
3. [ 5.2 Seeing ](https://opentextbc.ca/introductiontopsychology/chapter/4-2-seeing/)
4. [ 5.3 Hearing ](https://opentextbc.ca/introductiontopsychology/chapter/4-3-hearing/)
5. [ 5.4 Tasting, Smelling, and Touching ](https://opentextbc.ca/introductiontopsychology/chapter/4-4-tasting-smelling-and-touching/)
6. [ 5.5 Accuracy and Inaccuracy in Perception ](https://opentextbc.ca/introductiontopsychology/chapter/4-5-accuracy-and-inaccuracy-in-perception/)
7. [ 5.6 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/4-6-chapter-summary/)
10. Chapter 6. States of Consciousness
1. [ 6\. States of Consciousness ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-5-states-of-consciousness/)
2. [ 6.1 Sleeping and Dreaming Revitalize Us for Action ](https://opentextbc.ca/introductiontopsychology/chapter/5-1-sleeping-and-dreaming-revitalize-us-for-action/)
3. [ 6.2 Altering Consciousness with Psychoactive Drugs ](https://opentextbc.ca/introductiontopsychology/chapter/5-2-altering-consciousness-with-psychoactive-drugs/)
4. [ 6.3 Altering Consciousness without Drugs ](https://opentextbc.ca/introductiontopsychology/chapter/5-3-altering-consciousness-without-drugs/)
5. [ 6.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/5-4-chapter-summary/)
11. Chapter 7. Growing and Developing
1. [ 7\. Growing and Developing ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-6-growing-and-developing/)
2. [ 7.1 Conception and Prenatal Development ](https://opentextbc.ca/introductiontopsychology/chapter/6-1-conception-and-prenatal-development/)
3. [ 7.2 Infancy and Childhood: Exploring and Learning ](https://opentextbc.ca/introductiontopsychology/chapter/6-2-infancy-and-childhood-exploring-and-learning/)
4. [ 7.3 Adolescence: Developing Independence and Identity ](https://opentextbc.ca/introductiontopsychology/chapter/6-3-adolescence-developing-independence-and-identity/)
5. [ 7.4 Early and Middle Adulthood: Building Effective Lives ](https://opentextbc.ca/introductiontopsychology/chapter/6-4-early-and-middle-adulthood-building-effective-lives/)
6. [ 7.5 Late Adulthood: Aging, Retiring, and Bereavement ](https://opentextbc.ca/introductiontopsychology/chapter/6-5-late-adulthood-aging-retiring-and-bereavement/)
7. [ 7.6 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/6-6-chapter-summary/)
12. Chapter 8. Learning
1. [ 8\. Learning ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-7-learning/)
2. [ 8.1 Learning by Association: Classical Conditioning ](https://opentextbc.ca/introductiontopsychology/chapter/7-1-learning-by-association-classical-conditioning/)
3. [ 8.2 Changing Behaviour through Reinforcement and Punishment: Operant Conditioning ](https://opentextbc.ca/introductiontopsychology/chapter/7-2-changing-behavior-through-reinforcement-and-punishment-operant-conditioning/)
4. [ 8.3 Learning by Insight and Observation ](https://opentextbc.ca/introductiontopsychology/chapter/7-3-learning-by-insight-and-observation/)
5. [ 8.4 Using the Principles of Learning to Understand Everyday Behaviour ](https://opentextbc.ca/introductiontopsychology/chapter/7-4-using-the-principles-of-learning-to-understand-everyday-behavior/)
6. [ 8.5 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/7-5-chapter-summary/)
13. Chapter 9. Remembering and Judging
1. [ 9\. Remembering and Judging ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-8-remembering-and-judging/)
2. [ 9.1 Memories as Types and Stages ](https://opentextbc.ca/introductiontopsychology/chapter/8-1-memories-as-types-and-stages/)
3. [ 9.2 How We Remember: Cues to Improving Memory ](https://opentextbc.ca/introductiontopsychology/chapter/8-2-how-we-remember-cues-to-improving-memory/)
4. [ 9.3 Accuracy and Inaccuracy in Memory and Cognition ](https://opentextbc.ca/introductiontopsychology/chapter/8-3-accuracy-and-inaccuracy-in-memory-and-cognition/)
5. [ 9.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/8-4-chapter-summary/)
14. Chapter 10. Intelligence and Language
1. [ 10\. Intelligence and Language ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-9-intelligence-and-language/)
2. [ 10.1 Defining and Measuring Intelligence ](https://opentextbc.ca/introductiontopsychology/chapter/9-1-defining-and-measuring-intelligence/)
3. [ 10.2 The Social, Cultural, and Political Aspects of Intelligence ](https://opentextbc.ca/introductiontopsychology/chapter/9-2-the-social-cultural-and-political-aspects-of-intelligence/)
4. [ 10.3 Communicating with Others: The Development and Use of Language ](https://opentextbc.ca/introductiontopsychology/chapter/9-3-communicating-with-others-the-development-and-use-of-language/)
5. [ 10.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/9-4-chapter-summary/)
15. Chapter 11. Emotions and Motivations
1. [ 11\. Emotions and Motivations ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-10-emotions-and-motivations/)
2. [ 11.1 The Experience of Emotion ](https://opentextbc.ca/introductiontopsychology/chapter/10-1-the-experience-of-emotion/)
3. [ 11.2 Stress: The Unseen Killer ](https://opentextbc.ca/introductiontopsychology/chapter/10-2-stress-the-unseen-killer/)
4. [ 11.3 Positive Emotions: The Power of Happiness ](https://opentextbc.ca/introductiontopsychology/chapter/10-3-positive-emotions-the-power-of-happiness/)
5. [ 11.4 Two Fundamental Human Motivations: Eating and Mating ](https://opentextbc.ca/introductiontopsychology/chapter/10-4-two-fundamental-human-motivations-eating-and-mating/)
6. [ 11.5 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/10-5-chapter-summary/)
16. Chapter 12. Personality
1. [ 12\. Personality ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-11-personality/)
2. [ 12.1 Personality and Behaviour: Approaches and Measurement ](https://opentextbc.ca/introductiontopsychology/chapter/11-1-personality-and-behavior-approaches-and-measurement/)
3. [ 12.2 The Origins of Personality ](https://opentextbc.ca/introductiontopsychology/chapter/11-2-the-origins-of-personality/)
4. [ 12.3 Is Personality More Nature or More Nurture? Behavioural and Molecular Genetics ](https://opentextbc.ca/introductiontopsychology/chapter/11-3-is-personality-more-nature-or-more-nurture-behavioral-and-molecular-genetics/)
5. [ 12.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/11-4-chapter-summary/)
17. Chapter 13. Defining Psychological Disorders
1. [ 13\. Defining Psychological Disorders ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-12-defining-psychological-disorders/)
2. [ 13.1 Psychological Disorder: What Makes a Behaviour Abnormal? ](https://opentextbc.ca/introductiontopsychology/chapter/12-1-psychological-disorder-what-makes-a-behavior-abnormal/)
3. [ 13.2 Anxiety and Dissociative Disorders: Fearing the World Around Us ](https://opentextbc.ca/introductiontopsychology/chapter/12-2-anxiety-and-dissociative-disorders-fearing-the-world-around-us/)
4. [ 13.3 Mood Disorders: Emotions as Illness ](https://opentextbc.ca/introductiontopsychology/chapter/12-3-mood-disorders-emotions-as-illness/)
5. [ 13.4 Schizophrenia: The Edge of Reality and Consciousness ](https://opentextbc.ca/introductiontopsychology/chapter/12-4-schizophrenia-the-edge-of-reality-and-consciousness/)
6. [ 13.5 Personality Disorders ](https://opentextbc.ca/introductiontopsychology/chapter/12-5-personality-disorders/)
7. [ 13.6 Somatoform, Factitious, and Sexual Disorders ](https://opentextbc.ca/introductiontopsychology/chapter/12-6-somatoform-factitious-and-sexual-disorders/)
8. [ 13.7 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/12-7-chapter-summary/)
18. Chapter 14. Treating Psychological Disorders
1. [ 14\. Treating Psychological Disorders ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-13-treating-psychological-disorders/)
2. [ 14.1 Reducing Disorder by Confronting It: Psychotherapy ](https://opentextbc.ca/introductiontopsychology/chapter/13-1-reducing-disorder-by-confronting-it-psychotherapy/)
3. [ 14.2 Reducing Disorder Biologically: Drug and Brain Therapy ](https://opentextbc.ca/introductiontopsychology/chapter/13-2-reducing-disorder-biologically-drug-and-brain-therapy/)
4. [ 14.3 Reducing Disorder by Changing the Social Situation ](https://opentextbc.ca/introductiontopsychology/chapter/13-3-reducing-disorder-by-changing-the-social-situation/)
5. [ 14.4 Evaluating Treatment and Prevention: What Works? ](https://opentextbc.ca/introductiontopsychology/chapter/13-4-evaluating-treatment-and-prevention-what-works/)
6. [ 14.5 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/13-5-chapter-summary/)
19. Chapter 15. Psychology in Our Social Lives
1. [ 15\. Psychology in Our Social Lives ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-14-psychology-in-our-social-lives/)
2. [ 15.1 Social Cognition: Making Sense of Ourselves and Others ](https://opentextbc.ca/introductiontopsychology/chapter/14-1-social-cognition-making-sense-of-ourselves-and-others/)
3. [ 15.2 Interacting With Others: Helping, Hurting, and Conforming ](https://opentextbc.ca/introductiontopsychology/chapter/14-2-interacting-with-others-helping-hurting-and-conforming/)
4. [ 15.3 Working With Others: The Costs and Benefits of Social Groups ](https://opentextbc.ca/introductiontopsychology/chapter/14-3-working-with-others-the-costs-and-benefits-of-social-groups/)
5. [ 15.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/14-4-chapter-summary/)
20. Chapter 16. Stress, Health, and Coping
1. [ 16\. Health, Stress, and Coping ](https://opentextbc.ca/introductiontopsychology/chapter/chapter-15-health-stress-and-coping/)
Jennifer Walinga
2. [ 16.1 Health and Stress ](https://opentextbc.ca/introductiontopsychology/chapter/15-1-health-and-stress/)
Jennifer Walinga
3. [ 16.2 Stress and Coping ](https://opentextbc.ca/introductiontopsychology/chapter/15-2-stress-and-coping/)
Jennifer Walinga
4. [ 16.3 Stress, Health, and Coping in the Workplace ](https://opentextbc.ca/introductiontopsychology/chapter/15-3-stress-health-and-coping-in-the-workplace/)
Jennifer Walinga
5. [ 16.4 Chapter Summary ](https://opentextbc.ca/introductiontopsychology/chapter/15-4-chapter-summary/)
Jennifer Walinga
21. [ About the Authors ](https://opentextbc.ca/introductiontopsychology/back-matter/about-the-authors/)
22. [ Versioning History ](https://opentextbc.ca/introductiontopsychology/back-matter/versioning-history/)
# [ Introduction to Psychology – 1st Canadian Edition
](https://opentextbc.ca/introductiontopsychology/)
Chapter 4. Brains, Bodies, and Behaviour
# 4.1 The Neuron Is the Building Block of the Nervous System
### Learning Objectives
1. Describe the structure and functions of the neuron.
2. Draw a diagram of the pathways of communication within and between neurons.
3. List three of the major neurotransmitters and describe their functions.
The nervous system is composed of more than 100 billion cells known as
_neurons_ . A **neuron** is _a cell in the nervous system whose function it is
to receive and transmit information_ . As you can see in Figure 4.1,
“Components of the Neuron,” neurons are made up of three major parts: a cell
body, or **soma** , which _contains the nucleus of the cell and keeps the cell
alive_ ; a branching treelike fibre known as the **dendrite** , which
_collects information from other cells and sends the information to the soma_
; and a long, segmented fibre known as the **axon** , which _transmits
information away from the cell body toward other neurons or to the muscles and
glands_ . Figure 4.2 shows a photograph of neurons taken using confocal
microscopy.
[ 
](http://opentextbc.ca/introductiontopsychology/wp-
content/uploads/sites/9/2013/11/6a3f0732c22683476ea201ffc5e428ad.jpg) Figure
4.1 Components of the Neuron.
[ 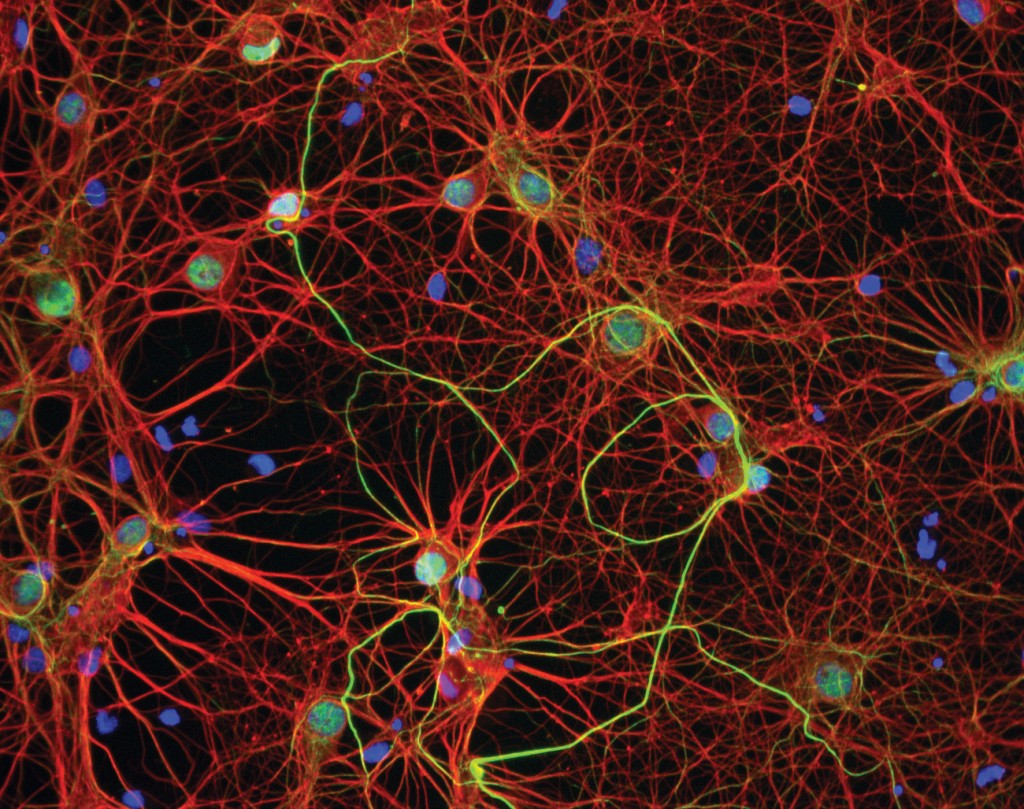
](http://opentextbc.ca/introductiontopsychology/wp-
content/uploads/sites/9/2013/11/fce5078fbe8ec74e95c8a435e607ad11.jpg) Figure
4.2 The nervous system, including the brain, is made up of billions of
interlinked neurons. This vast interconnected web is responsible for all human
thinking, feeling, and behaviour.
Some neurons have hundreds or even thousands of dendrites, and these dendrites
may themselves be branched to allow the cell to receive information from
thousands of other cells. The axons are also specialized, and some, such as
those that send messages from the spinal cord to the muscles in the hands or
feet, may be very long — even up to several feet in length. To improve the
speed of their communication, and to keep their electrical charges from
shorting out with other neurons, axons are often surrounded by a _myelin
sheath_ . The **myelin sheath** is _a layer of fatty tissue surrounding the
axon of a neuron that both acts as an insulator and allows faster transmission
of the electrical signal_ . Axons branch out toward their ends, and at the tip
of each branch is a _terminal button_ .
## Neurons Communicate Using Electricity and Chemicals
The nervous system operates using an _electrochemical_ process. An electrical
charge moves through the neuron itself, and chemicals are used to transmit
information between neurons. Within the neuron, when a signal is received by
the dendrites, it is transmitted to the soma in the form of an electrical
signal, and, if the signal is strong enough, it may then be passed on to the
axon and then to the terminal buttons. If the signal reaches the terminal
buttons, they are signalled to emit chemicals known as _neurotransmitters_ ,
which communicate with other neurons across the spaces between the cells,
known as _synapses_ .
[  ](http://opentextbc.ca/introductiontopsychology/wp-
content/uploads/sites/9/2014/09/electrochemical-action-of-the-neuron.png) _The
following video clip shows a model of the electrochemical action of the neuron
and neurotransmitters:_
**[ The Electrochemical Action of the Neuron [YouTube]
](http://www.youtube.com/watch?v=TKG0MtH5crc) : **
http://www.youtube.com/watch?v=TKG0MtH5crc
The electrical signal moves through the neuron as a result of changes in the
electrical charge of the axon. Normally, the axon remains in the **resting
potential** , _a state in which the interior of the neuron contains a greater
number of negatively charged ions than does the area outside the cell_ . When
the segment of the axon that is closest to the cell body is stimulated by an
electrical signal from the dendrites, and if this electrical signal is strong
enough that it passes a certain level or _threshold_ , the cell membrane in
this first segment opens its gates, allowing positively charged sodium ions
that were previously kept out to enter. This _change in electrical charge that
occurs in a neuron when a nerve impulse is transmitted_ is known as the
**action potential** . Once the action potential occurs, the number of
positive ions exceeds the number of negative ions in this segment, and the
segment temporarily becomes positively charged.
As you can see in Figure 4.3, “The Myelin Sheath and the Nodes of Ranvier,”
the axon is segmented by a series of _breaks between the sausage-like segments
of the myelin sheath_ . Each of these gaps is a **node of Ranvier** . [1]
The electrical charge moves down the axon from segment to segment, in a set of
small jumps, moving from node to node. When the action potential occurs in the
first segment of the axon, it quickly creates a similar change in the next
segment, which then stimulates the next segment, and so forth as the positive
electrical impulse continues all the way down to the end of the axon. As each
new segment becomes positive, the membrane in the prior segment closes up
again, and the segment returns to its negative resting potential. In this way
the action potential is transmitted along the axon, toward the terminal
buttons. The entire response along the length of the axon is very fast — it
can happen up to 1,000 times each second.
[ 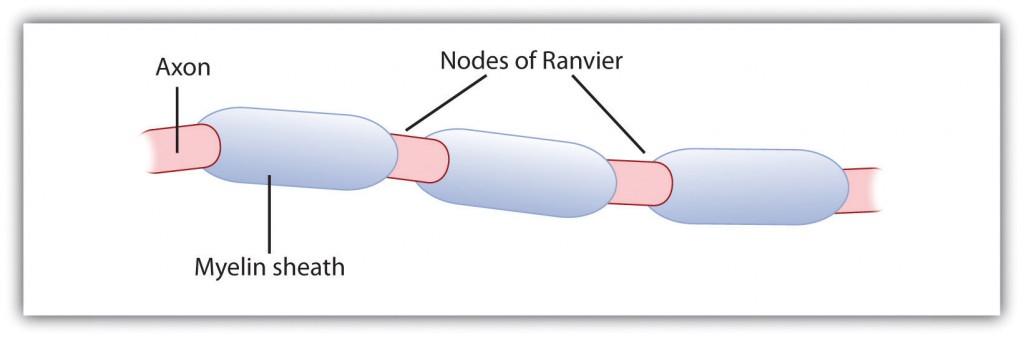
](http://opentextbc.ca/introductiontopsychology/wp-
content/uploads/sites/9/2013/11/496bba516a0f96bc27d754827642e444.jpg) Figure
4.3 The Myelin Sheath and the Nodes of Ranvier. The myelin sheath wraps around
the axon but also leaves small gaps called the nodes of Ranvier. The action
potential jumps from node to node as it travels down the axon.
An important aspect of the action potential is that it operates in an _all or
nothing_ manner. What this means is that the neuron either fires completely,
such that the action potential moves all the way down the axon, or it does not
fire at all. Thus neurons can provide more energy to the neurons down the line
by firing faster but not by firing more strongly. Furthermore, the neuron is
prevented from repeated firing by the presence of a **refractory period** — _a
brief time after the firing of the axon in which the axon cannot fire again
because the neuron has not yet returned to its resting potential._
## Neurotransmitters: The Body’s Chemical Messengers
Not only do the neural signals travel via electrical charges _within_ the
neuron, but they also travel via chemical transmission _between_ the neurons.
Neurons are separated by junction areas known as **synapses** , [2] _areas
where the terminal buttons at the end of the axon of one neuron nearly, but
don’t quite, touch the dendrites of another_ . The synapses provide a
remarkable function because they allow each axon to communicate with many
dendrites in neighbouring cells. Because a neuron may have synaptic
connections with thousands of other neurons, the communication links among the
neurons in the nervous system allow for a highly sophisticated communication
system.
When the electrical impulse from the action potential reaches the end of the
axon, it signals the terminal buttons to release _neurotransmitters_ into the
synapse. A **neurotransmitter** is _a chemical that relays signals across the
synapses between neurons_ . Neurotransmitters travel across the synaptic space
between the terminal button of one neuron and the dendrites of other neurons,
where they bind to the dendrites in the neighbouring neurons. Furthermore,
different terminal buttons release different neurotransmitters, and different
dendrites are particularly sensitive to different neurotransmitters. The
dendrites will admit the neurotransmitters only if they are the right shape to
fit in the receptor sites on the receiving neuron. For this reason, the
receptor sites and neurotransmitters are often compared to a lock and key
(Figure 4.4, “The Synapse”).
[ 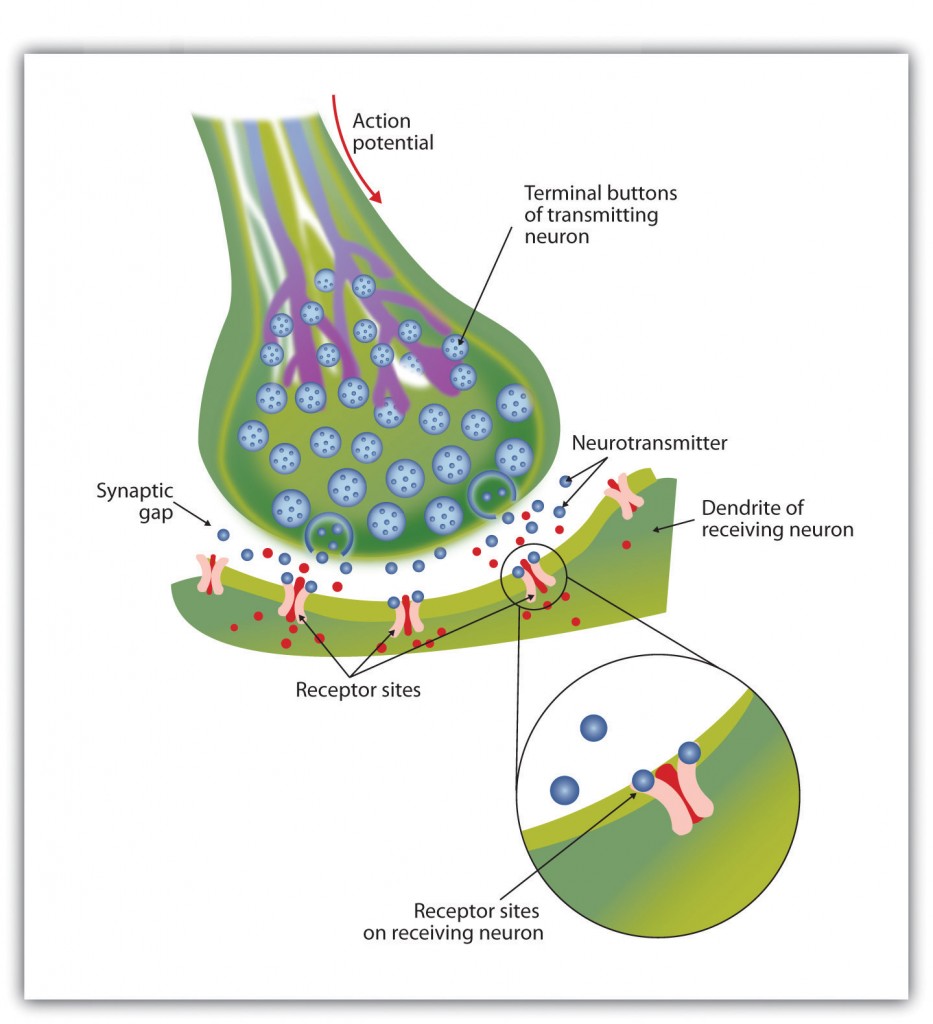
](http://opentextbc.ca/introductiontopsychology/wp-
content/uploads/sites/9/2013/11/6ede49aada9a9dd71c44517dc814ca3a.jpg) Figure
4.4 The Synapse. When the nerve impulse reaches the terminal button, it
triggers the release of neurotransmitters into the synapse. The
neurotransmitters fit into receptors on the receiving dendrites in the manner
of a lock and key.
When neurotransmitters are accepted by the receptors on the receiving neurons,
their effect may be either **excitatory** (i.e., _they make the cell more
likely to fire_ ) or **inhibitory** (i.e., _they make the cell less likely to
fire)_ . Furthermore, if the receiving neuron is able to accept more than one
neurotransmitter, it will be influenced by the excitatory and inhibitory
processes of each. If the excitatory effects of the neurotransmitters are
greater than the inhibitory influences of the neurotransmitters, the neuron
moves closer to its firing threshold; if it reaches the threshold, the action
potential and the process of transferring information through the neuron
begins.
Neurotransmitters that are not accepted by the receptor sites must be removed
from the synapse in order for the next potential stimulation of the neuron to
happen. This process occurs in part through the breaking down of the
neurotransmitters by enzymes, and in part through **reuptake** , _a process in
which neurotransmitters that are in the synapse are reabsorbed into the
transmitting terminal buttons, ready to again be released after the neuron
fires_ .
More than 100 chemical substances produced in the body have been identified as
neurotransmitters, and these substances have a wide and profound effect on
emotion, cognition, and behaviour. Neurotransmitters regulate our appetite,
our memory, our emotions, as well as our muscle action and movement. And as
you can see in Table 4.1, “The Major Neurotransmitters and Their Functions,”
some neurotransmitters are also associated with psychological and physical
diseases.
Drugs that we might ingest — either for medical reasons or recreationally —
can act like neurotransmitters to influence our thoughts, feelings, and
behaviour. An **agonist** is _a drug that has chemical properties similar to a
particular neurotransmitter and thus mimics the effects of the
neurotransmitter_ . When an agonist is ingested, it binds to the receptor
sites in the dendrites to excite the neuron, acting as if more of the
neurotransmitter had been present. As an example, cocaine is an agonist for
the neurotransmitter dopamine. Because dopamine produces feelings of pleasure
when it is released by neurons, cocaine creates similar feelings when it is
ingested. An antagonist is _a drug that reduces or stops the normal effects of
a neurotransmitter_ . When an **antagonist** is ingested, it binds to the
receptor sites in the dendrite, thereby blocking the neurotransmitter. As an
example, the poison curare is an antagonist for the neurotransmitter
acetylcholine. When the poison enters the brain, it binds to the dendrites,
stops communication among the neurons, and usually causes death. Still other
drugs work by blocking the reuptake of the neurotransmitter itself — when
reuptake is reduced by the drug, more neurotransmitter remains in the synapse,
increasing its action.
Table 4.1 The Major Neurotransmitters and Their Functions [Skip Table]
---
Neurotransmitter | Description and function | Notes
Acetylcholine (ACh) | A common neurotransmitter used in the spinal cord and
motor neurons to stimulate muscle contractions. It’s also used in the brain to
regulate memory, sleeping, and dreaming. | Alzheimer’s disease is associated
with an undersupply of acetylcholine. Nicotine is an agonist that acts like
acetylcholine.
Dopamine | Involved in movement, motivation, and emotion, Dopamine produces
feelings of pleasure when released by the brain’s reward system, and it’s also
involved in learning. | Schizophrenia is linked to increases in dopamine,
whereas Parkinson’s disease is linked to reductions in dopamine (and dopamine
agonists may be used to treat it).
Endorphins | Released in response to behaviours such as vigorous exercise,
orgasm, and eating spicy foods. | Endorphins are natural pain relievers.
They are related to the compounds found in drugs such as opium, morphine, and
heroin. The release of endorphins creates the runner’s high that is
experienced after intense physical exertion.
GABA (gamma-aminobutyric acid) | The major inhibitory neurotransmitter in
the brain. | A lack of GABA can lead to involuntary motor actions, including
tremors and seizures. Alcohol stimulates the release of GABA, which inhibits
the nervous system and makes us feel drunk. Low levels of GABA can produce
anxiety, and GABA agonists (tranquilizers) are used to reduce anxiety.
Glutamate | The most common neurotransmitter, it’s released in more than 90%
of the brain’s synapses. Glutamate is found in the food additive MSG
(monosodium glutamate). | Excess glutamate can cause overstimulation,
migraines, and seizures.
Serotonin | Involved in many functions, including mood, appetite, sleep, and
aggression. | Low levels of serotonin are associated with depression, and
some drugs designed to treat depression (known as selective serotonin reuptake
inhibitors, or SSRIs) serve to prevent their reuptake.
### Key Takeaways
* The central nervous system (CNS) is the collection of neurons that make up the brain and the spinal cord.
* The peripheral nervous system (PNS) is the collection of neurons that link the CNS to our skin, muscles, and glands.
* Neurons are specialized cells, found in the nervous system, which transmit information. Neurons contain a dendrite, a soma, and an axon.
* Some axons are covered with a fatty substance known as the myelin sheath, which surrounds the axon, acting as an insulator and allowing faster transmission of the electrical signal.
* The dendrite is a treelike extension that receives information from other neurons and transmits electrical stimulation to the soma.
* The axon is an elongated fibre that transfers information from the soma to the terminal buttons.
* Neurotransmitters relay information chemically from the terminal buttons and across the synapses to the receiving dendrites using a lock and key type of system.
* The many different neurotransmitters work together to influence cognition, memory, and behaviour.
* Agonists are drugs that mimic the actions of neurotransmitters, whereas antagonists are drugs that block the actions of neurotransmitters.
### Exercises and Critical Thinking
1. Draw a picture of a neuron and label its main parts.
2. Imagine an action that you engage in every day and explain how neurons and neurotransmitters might work together to help you engage in that action.
## Image Attributions
**Figure 4.2:** “ [ Confocal microscopy of mouse brain, cortex
](http://www.flickr.com/photos/zeissmicro/10799674936/in/photostream/) ” by
ZEISS Microscopy
(http://www.flickr.com/photos/zeissmicro/10799674936/in/photostream/) used
under [ CC BY-NC-ND 2.0 ](http://creativecommons.org/licenses/by-nc-
nd/2.0/deed.en_CA) (http://creativecommons.org/licenses/by-nc-
nd/2.0/deed.en_CA) license.
* * *
1. The break in the myelin sheath of a nerve fibre. ↵
2. The small gap between neurons across which nerve impulses are transmitted. ↵
Previous/next navigation
[ Previous: 4. Brains, Bodies, and Behaviour
](https://opentextbc.ca/introductiontopsychology/chapter/chapter-3-brains-
bodies-and-behavior/ "Previous: 4. Brains, Bodies, and Behaviour")
[ Next: 4.2 Our Brains Control Our Thoughts, Feelings, and Behaviour
](https://opentextbc.ca/introductiontopsychology/chapter/3-2-our-brains-
control-our-thoughts-feelings-and-behavior/ "Next: 4.2 Our Brains Control Our
Thoughts, Feelings, and Behaviour")
Back to top
## License

[ Introduction to Psychology - 1st Canadian Edition
](https://opentextbc.ca/introductiontopsychology) Copyright © 2014 by
Jennifer Walinga and Charles Stangor is licensed under a [ Creative Commons
Attribution-NonCommercial-ShareAlike 4.0 International License
](https://creativecommons.org/licenses/by-nc-sa/4.0/) , except where otherwise
noted.
## Share This Book
Share on Twitter
[ ](https://pressbooks.com "Pressbooks")
[ Powered by Pressbooks ](https://pressbooks.com)
* [ Guides and Tutorials ](https://pressbooks.com/support/)
* | [ Pressbooks Directory ](https://pressbooks.directory)
* | [ Contact ](https://collection.bccampus.ca/contact)
[  Pressbooks
on YouTube ](https://www.youtube.com/user/pressbooks) [ Pressbooks on Twitter
](https://twitter.com/intent/follow?screen_name=pressbooks)

| biology | 893782 | https://da.wikipedia.org/wiki/Musculus%20semispinalis | Musculus semispinalis | Semispinalis er en gruppe af tre muskler der hører til transversospinales. Disse er semispinalis capitis, semispinalis cervicis og semispinalis thoracis; navngivet efter deres lokation på i ryggen. De udspringer fra spidsentværtapperne af rygsøjlen, begyndende omkring thorax, og insererer så på tværtappen af en tværtap 4-7 segmenter over deres udspring.
Undtagelsen er musculus semispinalis capitis, som ender på kraniebunden; samt musculus semispinalis cervicis, som ender på torntappen af C2. Begge disse er strøj der kan separeres fra den gængse musculus semispinalis thoracis.
Referencer
Muskelsystemet
Dybe rygmuskler | danish | 1.204484 |
neuron_onto/Autapse.txt | An autapse is a chemical or electrical synapse from a neuron onto itself. It can also be described as a synapse formed by the axon of a neuron on its own dendrites, in vivo or in vitro.
History[edit]
The term "autapse" was first coined in 1972 by Van der Loos and Glaser, who observed them in Golgi preparations of the rabbit occipital cortex while originally conducting a quantitative analysis of neocortex circuitry. Also in the 1970s, autapses have been described in dog and rat cerebral cortex, monkey neostriatum, and cat spinal cord.
In 2000, they were first modeled as supporting persistence in recurrent neural networks. In 2004, they were modeled as demonstrating oscillatory behavior, which was absent in the same model neuron without autapse. More specifically, the neuron oscillated between high firing rates and firing suppression, reflecting the spike bursting behavior typically found in cerebral neurons. In 2009, autapses were, for the first time, associated with sustained activation. This proposed a possible function for excitatory autapses within a neural circuit. In 2014, electrical autapses were shown to generate stable target and spiral waves in a neural model network. This indicated that they played a significant role in stimulating and regulating the collective behavior of neurons in the network. In 2016, a model of resonance was offered.
Autapses have been used to simulate "same cell" conditions to help researchers make quantitative comparisons, such as studying how N-methyl-D-aspartate receptor (NMDAR) antagonists affect synaptic versus extrasynaptic NMDARs.
Formation[edit]
Recently, it has been proposed that autapses could possibly form as a result of neuronal signal transmission blockage, such as in cases of axonal injury induced by poisoning or impeding ion channels. Dendrites from the soma in addition to an auxiliary axon may develop to form an autapse to help remediate the neuron's signal transmission.
Structure and function[edit]
Autapses can be either glutamate-releasing (excitatory) or GABA-releasing (inhibitory), just like their traditional synapse counterparts. Similarly, autapses can be electrical or chemical by nature.
Broadly speaking, negative feedback in autapses tends to inhibit excitable neurons whereas positive feedback can stimulate quiescent neurons.
Although the stimulation of inhibitory autapses did not induce hyperpolarizing inhibitory post-synaptic potentials in interneurons of layer V of neocortical slices, they have been shown to impact excitability. Upon using a GABA-antagonist to block autapses, the likelihood of an immediate subsequent second depolarization step increased following a first depolarization step. This suggests that autapses act by suppressing the second of two closely timed depolarization steps and therefore, they may provide feedback inhibition onto these cells. This mechanism may also potentially explain shunting inhibition.
In cell culture, autapses have been shown to contribute to the prolonged activation of B31/B32 neurons, which significantly contribute food-response behavior in Aplysia. This suggests that autapses may play a role in mediating positive feedback. The B31/B32 autapse was unable to play a role in initiating the neuron's activity, although it is believed to have helped sustain the neuron's depolarized state. The extent to which autapses maintain depolarization remains unclear, particularly since other components of the neural circuit (i.e. B63 neurons) are also capable of providing strong synaptic input throughout the depolarization. Additionally, it has been suggested that autapses provide B31/B32 neurons with the ability to quickly repolarize. Bekkers (2009) has proposed that specifically blocking the contribution of autapses and then assessing the differences with or without blocked autapses could better illuminate the function of autapses.
Hindmarsh–Rose (HR) model neurons have demonstrated chaotic, regular spiking, quiescent, and periodic patterns of burst firing without autapses. Upon the introduction of an electrical autapse, the periodic state switches to the chaotic state and displays an alternating behavior that increases in frequency with a greater autaptic intensity and time delay. On the other hand, excitatory chemical autapses enhanced the overall chaotic state. The chaotic state was reduced and suppressed in the neurons with inhibitory chemical autapses. In HR model neurons without autapses, the pattern of firing altered from quiescent to periodic and then to chaotic as DC current was increased. Generally, HR model neurons with autapses have the ability to swap into any firing pattern, regardless of the prior firing pattern.
Location[edit]
Neurons from several brain regions, such as the neocortex, substantia nigra, and hippocampus have been found to contain autapses.
Autapses have been observed to be relatively more abundant in GABAergic basket and dendrite-targeting cells of the cat visual cortex compared to spiny stellate, double bouquet, and pyramidal cells, suggesting that the degree of neuron self-innervation is cell-specific. Additionally, dendrite-targeting cell autapses were, on average, further from the soma compared to basket cell autapses.
80% of layer V pyramidal neurons in developing rat neocortices contained autaptic connections, which were located more so on basal dendrites and apical oblique dendrites rather than main apical dendrites. The dendritic positions of synaptic connections of the same cell type were similar to those of autapses, suggesting that autaptic and synaptic networks share a common mechanism of formation.
Disease implications[edit]
In the 1990s, paroxysmal depolarizing shift-type interictal epileptiform discharges has been suggested to be primarily dependent on autaptic activity for solitary excitatory hippocampal rat neurons grown in microculture.
More recently, in human neocortical tissues of patients with intractable epilepsy, the GABAergic output autapses of fast-spiking (FS) neurons have been shown to have stronger asynchronous release (AR) compared to both non-epileptic tissue and other types of synapses involving FS neurons. The study found similar results using a rat model as well. An increase in residual Ca2+ concentration in addition to the action potential amplitude in FS neurons was suggested to cause this increase in AR of epileptic tissue. Anti-epileptic drugs could potentially target this AR of GABA that seems to rampantly occur at FS neuron autapses.
Effects of drugs[edit]
Using a glia-conditioned medium to treat glia-free purified rat retinal ganglion microcultures has been shown to significantly increase the number of autapses per neuron compared to a control. This suggests that glia-derived soluble, proteinase K-sensitive factors induce autapse formation in rat retinal ganglion cells. | biology | 322049 | https://da.wikipedia.org/wiki/Neuron | Neuron | Et neuron (fra græsk: sene, nerve), også kaldet en nervecelle, er en celletype i nervesystemet.
De adskiller sig fra andre celler ved deres mange udløbere, kaldet dendritter og aksoner, og ved at være specialiseret i transmittering af signaler, dels elektrisk intraneuronalt og dels ved hjælp af transmitterstoffer interneuronalt, der virker på andre neuroners eller effektorvævenes receptorer (se for eksempel G-protein-koblede receptorer, GPCR og den motoriske endeplade). Af neurotransmittere kan nævnes serotonin, acetylkolin, dopamin, adrenalin og noradrenalin.
Formålet med hjernens netværk af neuroner er kommunikation og informationsbehandling.
Opbygning
Neuroner er de centrale komponenter i hjernen og rygmarven i centralnervesystemet (CNS) samt ganglier i det perifere nervesystem (PNS). En typisk neuron består af et cellelegeme (soma), dendritter og et axon. Dendritter er tynde strukturer, der udspringer fra cellekroppen og ofte strækker sig flere hundrede mikrometer og forgrener sig flere gange. En Axon, også kaldet en nervefiber når de er myelinerede, udspringer fra cellen ved et sted kaldet axon højen og rejser op til en en meter i mennesker og endnu længere i andre arter. En neurons cellekroppe har ofte flere dendritter, men aldrig til mere end ét Axon. Neuronets lange aksoner kan være omgivet af en isolerende fedtskede kaldet myelinskeden, som har til funktion at øge signaleringshastigheden.
Myelinskeden produceres og vedligeholdes i centralnervesystemet af oligodendrocytter og af Schwannske celler i det perifere nervesystem.
I de fleste tilfælde er neuroner genereret af særlige typer af stamceller. Det menes generelt, at neuroner ikke undergår celledeling, men nyere forskning hos hunde viser, at det i nogle tilfælde sker i nethinden. Astrocytter er stjerneformede gliaceller, der er også blevet observeret at blive til neuroner. Hos mennesker ophører neurogenese stort set i voksenalderen, men i to områder i hjernen, hippocampus og den olfaktoriske pære, er der beviser for generering af nye neuroner.
Kommunikation mellem neuroner
En neuron behandler og sender oplysninger gennem elektriske og kemiske signaler. Nervecellernes signaler bygger på en spændingsforskel mellem nervecellens indre og ydre, kaldet membranpotentialet. Er neuronet i hvile, kaldes spændingsforskellen for hvilemembranpotentialet. Er ændringen i membranpotentialet tilstrækkelig stor, genereres en (alt-eller-intet) elektrokemisk puls kaldet et aktionspotentiale, som bevæger sig hurtigt langs cellens axon og aktiverer synaptiske forbindelser til andre celler.
Neurontyper
Neuroner kan overordnet inddeles i projektionsneuroner Golgi type 1 med et enkelt langt akson, der kan kommunikere med andre regioner af nervesystemet, og interneuroner Golgi type 2 som kun befinder sig i en hjerneregion. Der er dog ikke skarp adskillelse mellem disse to typer.
Specialiserede neuroner omfatter: 1) sensoriske neuroner, der reagerer på berøring, lyd, lys og alle andre stimuli, der påvirker cellerne via vores sanser, som derefter sender signaler til rygmarven og hjernen; 2) motoriske neuroner, der modtager signaler fra hjernen og rygmarven, som bevirker muskelsammentrækninger og påvirker glandulare udgange; 3) interneuroner, der forbinder neuroner i den samme region i hjernen eller rygmarven.
Se også
Dendrit, akson, soma, gliacelle
Kranienerver
Neuropeptid
Eksterne henvisninger | danish | 0.434241 |
neuron_onto/NBK526047.txt | **Warning:** The NCBI web site requires JavaScript to function. [ more...
](/guide/browsers/#enablejs "Learn how to enable JavaScript")

An official website of the United States government
Here's how you know

**The .gov means it's official.**
Federal government websites often end in .gov or .mil. Before sharing
sensitive information, make sure you're on a federal government site.

**The site is secure.**
The **https://** ensures that you are connecting to the official website and
that any information you provide is encrypted and transmitted securely.
[ 
](/)
[ Log in ](https://account.ncbi.nlm.nih.gov) Show account info
Close
#### Account
Logged in as:
** username **
* [ Dashboard ](/myncbi/)
* [ Publications ](/myncbi/collections/bibliography/)
* [ Account settings ](/account/settings/)
* [ Log out ](/account/signout/)
[ Access keys ](https://www.ncbi.nlm.nih.gov/guide/browsers/#ncbi_accesskeys)
[ NCBI Homepage ](https://www.ncbi.nlm.nih.gov) [ MyNCBI Homepage ](/myncbi/)
Main Content Main Navigation
# [ Bookshelf ](/books/ "Bookshelf home")
##
Search database Books All Databases Assembly Biocollections BioProject
BioSample Books ClinVar Conserved Domains dbGaP dbVar Gene Genome GEO
DataSets GEO Profiles GTR Identical Protein Groups MedGen MeSH NLM
Catalog Nucleotide OMIM PMC PopSet Protein Protein Clusters Protein
Family Models PubChem BioAssay PubChem Compound PubChem Substance PubMed
SNP SRA Structure Taxonomy ToolKit ToolKitAll ToolKitBookgh
Search term
Search
* [ Browse Titles ](/books/browse/)
* [ Advanced ](/books/advanced/)
* [ Help ](/books/NBK3833/)
* [ Disclaimer ](https://www.ncbi.nlm.nih.gov/books/about/disclaimer/)
NCBI Bookshelf. A service of the National Library of Medicine, National
Institutes of Health.
StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-.
[  ](/books/n/statpearls/ "Table of Contents Page")
## StatPearls [Internet].
Show details
Treasure Island (FL): [ StatPearls Publishing ](https://www.statpearls.com/) ;
2024 Jan-.
Search term
# Physiology, Synapse
Michael J. Caire ; Vamsi Reddy ; Matthew Varacallo .
Author Information and Affiliations
#### Authors
Michael J. Caire 1 ; Vamsi Reddy 2 ; Matthew Varacallo 3 .
#### Affiliations
1 LSU School of Medicine - Shreveport
2 McKinsey & Company
3 Penn Highlands Healthcare System
Last Update: March 27, 2023 .
## Introduction
The human brain is made up of approximately 86 billion neurons that “talk” to
each other using a combination of electrical and chemical (electrochemical)
signals.
The places where neurons connect and communicate with each other are called
synapses. Each neuron has anywhere between a few to hundreds of thousands of
synaptic connections, and these connections can be with itself, neighboring
neurons, or neurons in other regions of the brain. A synapse is made up of a
presynaptic and postsynaptic terminal.
The presynaptic terminal is at the end of an axon and is the place where the
electrical signal (the action potential) is converted into a chemical signal
(neurotransmitter release). The postsynaptic terminal membrane is less than 50
nanometers away and contains specialized receptors. The neurotransmitter
rapidly (in microseconds) diffuses across the synaptic cleft and binds to
specific receptors.
The type of neurotransmitter released from the presynaptic terminal and the
specific receptors present on the corresponding postsynaptic terminal is
critical in determining the quality and intensity of information transmitted
by neurons. The postsynaptic neuron integrates all the signals it receives to
determine what it does next, for example, to fire an action potential of its
own or not. [1] [2]
## Cellular Level
**Neurons**
In the simplest sense, the neuron consists of a cell body, axons, and
dendrites.
**Cell Body**
The cell body contains the nucleus and is the site of metabolic activity. Most
of the neurotransmitters that will eventually be released at the synapse are
synthesized here.
**Dendrites**
These are small projections from the cell body that serves a receptive role in
the physiology of the neuron. They receive incoming signals from other neurons
and relay them to the cell body, where the signals are integrated, and a
response will be initiated.
**Axons**
Generally, the outflow tract of the neuron. It is a cylindrical tube that is
covered by the axolemma and is supported by neurofilaments and microtubules.
The microtubules will help to transport the neurotransmitters from the cell
body down to the pre-synaptic terminal, where they will be released.
**Synapses**
The synapse itself is the site of transmission from the pre-synaptic neuron to
the post-synaptic neuron. The structures found on either side of the synapse
vary depending on the type of synapse:
**Axodendritic**
A connection formed between the axon of one neuron and the dendrite of
another. These tend to be excitatory synapses.
**Axosomatic**
A direct connection between the axon of one neuron to the cell body of another
neuron. These tend to be inhibitory synapses.
**Axoaxonic**
A connection between the terminal of one axon and another axon. These synapses
generate serve a regulatory role; the afferent axon will modulate the release
of neurotransmitters from the efferent axon.
The above discussion focuses on chemical synapses, which involve the release
of a chemical neurotransmitter between the 2 neurons. This is the most common
type of synapse in the mammalian central nervous system (CNS). However, it is
important to note that there are electrical synapses, where electrical current
(or signals) will pass directly from one neuron to another through gap
junctions. The differences between the two will be expanded on in the
mechanism section. [3] [4]
## Development
Two neurons form the neurological synapse, or in some instances, a neuron and
an anatomical structure. This review will focus on 2 neurons composing the
synapse. Neurons initially develop from the embryonic neural tube, which has 3
layers:
* The _ventricular zone_ , which surrounds the central canal of the tube. This tube will eventually become the ependyma.
* The _intermediate zone_ , which is formed by dividing cells of the ventricular zone. This zone stretches from the outermost portion of the ventricular zone to the outermost layer of the neural tube, known as the pial layer.
* The _marginal zone_ , which is formed by extensions of the nerve cells of the intermediate zone.
The intermediate zone will go on to form the gray matter, while the nerve
processes that make up the marginal zone will become white matter once
myelinated.
The neurons must then differentiate from their precursors. The order in which
they do this is based upon their size, with the largest neurons (motor
neurons) differentiating first. Around the time of birth, the smaller neurons
(sensory neurons) will develop, along with glial cells. Glial cells are cells
that will aid in the differentiation of the neurons and will facilitate their
growth in the direction of their target locations. Later, glial cells will
participate in the reuptake of excess neurotransmitters in the synaptic cleft.
## Mechanism
**Synapses**
As previously mentioned, there are 2 major types of synapses: electrical and
chemical.
In mammals, the majority of synapses are chemical. Chemical synapses can be
differentiated from electrical synapses by a few distinguishing criteria: they
use neurotransmitters to relay the signal and vesicles are used to store and
transport the neurotransmitter from the cell body to the terminal;
furthermore, the pre-synaptic terminal will have a very active membrane and
the post-synaptic membrane consists of a thick cell membrane made up of many
receptors. In between these 2 membranes is a very distinct cleft (easily
visualized with electron microscopy) and the chemical neurotransmitter
released must diffuse across this cleft to elicit a response in the receptive
neuron. Because of this, the synaptic delay, defined as the time it takes for
current in the pre-synaptic neuron to be transmitted to the post-synaptic
neuron, is approximately 0.5 to 1.0 ms.
This is different from the electrical synapse, which will typically consist of
2 membranes located much closer to each other than in a chemical synapse.
These membranes possess channels formed by proteins known as connexins, which
allow the direct passage of current from one neuron to the next and do not
rely on neurotransmitters. The synaptic delay is significantly shorter in
electrical synapses versus chemical synapses.
The rest of the discussion will focus on chemical synapses, which have a lot
of variation and diversity of their own. They vary not only between shape and
structure, but also the chemical that is transmitted. Synapses can be
excitatory or inhibitory, and use a variety of chemical molecules and proteins
that will be discussed shortly.
Multiple types of neurotransmitters used in synaptic communication including,
but not limited to:
* Acetylcholine (ACh): One of the most important neurotransmitters found in multiple synapses in the body, including, but not limited to, the neuromuscular junction, autonomic ganglia, caudate nucleus, and the limbic system. Generally, ACh is an excitatory neurotransmitter at the neuromuscular junction and in the autonomic ganglia. In the brain, Ach is synthesized in the basal nucleus of Meynert.
* Norepinephrine (NE): The most important molecule in sympathetic nervous system signaling, except for the sweat glands. In the brain, NE is mainly found in the locus coeruleus and lateral tegmental nuclei.
* Dopamine (DA): Dopamine signaling is generally inhibitory. There are three major dopaminergic pathways in the brain, the nigrostriatal, mesolimbic, and mesocortical; each of which serve different roles. One of the most well-known disease states involving dopamine is Parkinson's disease, where there is degeneration of dopaminergic neurons in the substantia nigra.
* Serotonin (5-HT): Produced from tryptophan using tryptophan hydroxylase, which is mostly found in the brain (raphe nucleus) and the gastrointestinal (GI) tract. Serotonin is mostly known for its role as a regulatory neurotransmitter and is therefore implicated in various mood states and diseases.
* Other common neurotransmitters include other catecholamines, gamma-aminobutyric acid (GABA), glycine, and glutamic acid.
The easiest approach to understanding synaptic transmission is to think of it
as a stepwise process beginning with the synthesis of the neurotransmitter and
ending with its release.
1. Synthesis: **** The neurotransmitter is synthesized in the cell body, where it will then be transmitted down the microtubules of the axon to the pre-synaptic terminal, or it is synthesized directly in the pre-synaptic terminal from recycled neurotransmitters. The neurotransmitter is then stored in presynaptic vesicles until its release.
2. Release: **** The neurotransmitter is released in a regulated fashion from the pre-synaptic neuron into the synaptic cleft.
3. Receptor activation: **** The neurotransmitter binds to post-synaptic receptors and produces a response in the post-synaptic neuron.
4. Signal termination: **** The signal must be terminated by some mechanism, normally by the elimination of excess neurotransmitters from the synaptic cleft.
**Synthesis**
Neurotransmitters are synthesized differently depending on which type they
are. They can be a small molecule chemical, such as dopamine and serotonin, or
they can be small neuropeptides, such as enkephalin.
* Neuropeptides are synthesized in the cell body using the typical protein synthesis and translation pathways (rough endoplasmic reticulum and Golgi apparatus), then will be packaged into large, dense-core vesicles along with a protease. These vesicles are rapidly transported down the axon using microtubular proteins such as kinesin. When they arrive at the pre-synaptic terminal, they are ready to be released.
* Small molecule neurotransmitters are synthesized in the cell body, where they are then transported down the axon in small, clear core vesicles. Upon arriving at the pre-synaptic terminal, enzymes will modify the small molecule neurotransmitter, and they can then be released from the vesicles into the cleft.
**Release**
Now that the neurotransmitters are stored in the vesicles in the pre-synaptic
terminal, they must be released into the cleft. Along the membrane of the
vesicle and the presynaptic membrane are proteins known as SNARE proteins;
these proteins are essential in the binding of the vesicles to the membrane
and the release of their contents. As the action potential propagates down the
pre-synaptic neuron, the membrane will depolarize. Once the action potential
arrives at the pre-synaptic terminal, the depolarization of the membrane will
allow the voltage-dependent calcium channels to open, allowing the rapid
influx of calcium into the pre-synaptic terminal. The influx of calcium causes
the SNARE proteins to activate and change conformation, allowing the fusion of
vesicles to the membrane and the release of their contents. The
neurotransmitter will spill into the synaptic cleft, and the vesicle membrane
is recovered via endocytosis.
**Receptor Activation**
Once the neurotransmitter binds to the post-synaptic neuron, it can generally
cause one of 2 types of receptors to be activated. It will either activate a
ligand-gated ion channel or a G-protein receptor.
* Ligand-Gated Ion Channel: When the neurotransmitter binds to this receptor, there is a direct opening or closing of the attached ion channel. In other words, the neurotransmitter acts directly on the target ion channel. This type of receptor is described as “fast” because it generally only takes a few milliseconds to produce a response and is terminated very quickly. Depending on which neurotransmitter is binding to the receptor, these types of receptors can be excitatory or inhibitory.
* G-Protein Coupled Receptors: These types of receptors are will produce a response (open or close an ion channel) by activating a signaling cascade involving secondary messengers. The most common secondary messengers are cyclic adenosine monophosphate (cAMP), inositol triphosphate (IP3), and diacylglycerol (DAG). When the neurotransmitter binds to the receptor, it activates the G-protein, which binds to guanosine triphosphate (GTP), and is activated. This will activate the secondary messenger cascade, which will eventually lead to the phosphorylation of ion channels. Due to multiple steps having to take place to generate the final response, this pathway is generally described as “slow,” and generally, the effects last longer (seconds to minutes).
**Signal Termination**
Inactivation of the signal must involve clearing the neurotransmitter from the
synapse in at least 1 of 3 ways:
* Re-uptake: Re-uptake can either be pre-synaptic or by glial cells. One important point to remember involving reuptake is that only small molecule chemical neurotransmitters can be taken back up, neuropeptides cannot participate in re-uptake; they must be eliminated by other means, such as degradation.
* In pre-synaptic reuptake, the pre-synaptic neuron will use either endocytosis or specific transporters to remove the neurotransmitter from the synapse. The advantage of this mechanism is that the neurotransmitter can be recycled, which will prevent the neuron from having to re-synthesize the neurotransmitter every cycle of release.
* In some cases, such as with glutamate, a glial cell will be involved in the re-uptake. Glutamate is toxic to the cell, so it is stored inside the neuron as glutamine. When glutamate is released into the synapse, it will be taken up by the glial cell using a specific transporter, converted into glutamine via glutaminase, then returned to the neuron to be recycled.
* Enzymatic Destruction: The neurotransmitter can be destroyed directly either in the cleft or in the pre-synaptic terminal using certain enzymes. Two major enzymes are involved in the destruction of the neurotransmitter:
* Monoamine Oxidases (MAO): These enzymes are responsible for oxidizing, and therefore inactivating, the monoamines. They do this by using oxygen to remove the amine group. These are split into MAO-A and MAO-B based on substrates. MAO-A is mostly responsible for breaking down serotonin, melatonin, norepinephrine, and epinephrine. Both forms break down dopamine, tyramine, and tryptamine equally. MAO-B also breaks down phenethylamine and benzylamine.
* Catechol-O-Methyltransferase (COMT): Generally, COMT is responsible for degrading catecholamines, including dopamine, epinephrine, and norepinephrine, as well as most substances with a catechol structure.
It is important to note that both of the above enzymes are very frequent
targets of therapeutic medications. By eliminating these enzymes, the
neurotransmitter will remain in the synapse for longer, which can be
beneficial in eliminating the symptoms of many disease processes.
* Diffusion: **** In the simplest form of termination, the neurotransmitter can simply diffuse out of the synaptic cleft and away from the receptors and into nearby blood vessels. This will decrease the concentration of the neurotransmitter in the synapse, gradually reducing the effect the neurotransmitter has on the post-synaptic neuron. [5] [6]
## Clinical Significance
The synapse is the fundamental functional unit of neuronal communication.
Because of this, diseases that target the synapse can present with severe
clinical consequences. A few examples are listed below:
**Myasthenia Gravis**
Myasthenia gravis is an auto-immune disease process that causes muscle
weakness that usually presents in a descending fashion. It can cause ptosis,
diminished facial expression, respiratory depression, and other signs/symptoms
of weakness. In general, it is worse after activity and better with rest. The
pathogenesis of myasthenia gravis involves diminished communication between
the neuron and the muscle at the neuromuscular junction (NMJ). The reason for
this is that antibodies will either block or destroy the acetylcholine
receptors at the NMJ, preventing the ACh from binding and depolarizing the
muscle, therefore, inhibiting contraction. These antibodies block step three
(receptor activation) of the synaptic communication pathway.
**Lambert-Eaton Syndrome**
Lambert-Eaton syndrome is also an auto-immune condition producing dysfunction
at the neuromuscular junction; however, it involves the pre-synaptic neuron.
Instead of antibodies directed against the ACh receptors as in myasthenia
gravis, the antibodies here are directed against the calcium channels on the
pre-synaptic neuron. This prevents calcium influx from occurring, which
prevents the fusion of vesicles with the pre-synaptic membrane and the release
of the neurotransmitters into the synapse. These antibodies prevent step two
(neurotransmitter release) of the synaptic communication pathway.
**Botulism/Tetanus**
In both of these disease processes, the causative agent is a toxin produced by
a bacteria that acts as a protease that cleaves the SNARE proteins. This
prevents the release of neurotransmitters at the junction by inhibiting
vesicular fusion.
* _Botulism:_ The botulinum toxin, produced by _Clostridium botulinum_ , prevents the release of acetylcholine, which is a stimulatory neurotransmitter. This inhibits stimulatory effects, which prevents muscle contraction and causes flaccid paralysis.
* _Tetanus:_ The tetanus toxin, produced by _Clostridium tetani_ , prevents the release of GABA and glycine, both of which are inhibitory neurotransmitters. Specifically, their release is inhibited in the Renshaw cells in the spinal cord. This produces symptoms resembling an upper motor neuron lesion: spastic paralysis, lockjaw, and opisthotonus.
## Review Questions
* [ Access free multiple choice questions on this topic. ](https://www.statpearls.com/account/trialuserreg/?articleid=29802&utm_source=pubmed&utm_campaign=reviews&utm_content=29802)
* [ Comment on this article. ](https://www.statpearls.com/articlelibrary/commentarticle/29802/?utm_source=pubmed&utm_campaign=comments&utm_content=29802)
[ 
](/books/NBK526047/figure/article-29802.image.f1/?report=objectonly "Figure")
#### [ Figure
](/books/NBK526047/figure/article-29802.image.f1/?report=objectonly)
Anatomy of Neurons. A. Two connected neurons. Neurons have a soma that
contains a nucleus, an axon, and a dendritic tree. A single synapse (red
circle) is formed at the point where an axon's neuron (black) connects to
another neuron's [ (more...)
](/books/NBK526047/figure/article-29802.image.f1/?report=objectonly)
## References
1\.
Jones RA, Harrison C, Eaton SL, Llavero Hurtado M, Graham LC, Alkhammash L,
Oladiran OA, Gale A, Lamont DJ, Simpson H, Simmen MW, Soeller C, Wishart TM,
Gillingwater TH. Cellular and Molecular Anatomy of the Human Neuromuscular
Junction. Cell Rep. 2017 Nov 28; 21 (9):2348-2356. [ [ PMC free article
: PMC5723673 ](/pmc/articles/PMC5723673/) ] [ [ PubMed : 29186674
](https://pubmed.ncbi.nlm.nih.gov/29186674) ]
2\.
Napper RM, Harvey RJ. Number of parallel fiber synapses on an individual
Purkinje cell in the cerebellum of the rat. J Comp Neurol. 1988 Aug 08; 274
(2):168-77. [ [ PubMed : 3209740 ](https://pubmed.ncbi.nlm.nih.gov/3209740)
]
3\.
Harris AL. Electrical coupling and its channels. J Gen Physiol. 2018 Dec 03;
150 (12):1606-1639. [ [ PMC free article : PMC6279368
](/pmc/articles/PMC6279368/) ] [ [ PubMed : 30389716
](https://pubmed.ncbi.nlm.nih.gov/30389716) ]
4\.
Südhof TC. Towards an Understanding of Synapse Formation. Neuron. 2018 Oct
24; 100 (2):276-293. [ [ PMC free article : PMC6226307
](/pmc/articles/PMC6226307/) ] [ [ PubMed : 30359597
](https://pubmed.ncbi.nlm.nih.gov/30359597) ]
5\.
Südhof TC. The presynaptic active zone. Neuron. 2012 Jul 12; 75 (1):11-25.
[ [ PMC free article : PMC3743085 ](/pmc/articles/PMC3743085/) ] [ [ PubMed
: 22794257 ](https://pubmed.ncbi.nlm.nih.gov/22794257) ]
6\.
Lisman JE, Raghavachari S, Tsien RW. The sequence of events that underlie
quantal transmission at central glutamatergic synapses. Nat Rev Neurosci.
2007 Aug; 8 (8):597-609. [ [ PubMed : 17637801
](https://pubmed.ncbi.nlm.nih.gov/17637801) ]
**Disclosure:** Michael Caire declares no relevant financial relationships
with ineligible companies.
**Disclosure:** Vamsi Reddy declares no relevant financial relationships with
ineligible companies.
**Disclosure:** Matthew Varacallo declares no relevant financial relationships
with ineligible companies.
[ Copyright ](/books/about/copyright/) © 2024, StatPearls Publishing LLC.
This book is distributed under the terms of the Creative Commons Attribution-
NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0) ( [
http://creativecommons.org/licenses/by-nc-nd/4.0/
](https://creativecommons.org/licenses/by-nc-nd/4.0/) ), which permits others
to distribute the work, provided that the article is not altered or used
commercially. You are not required to obtain permission to distribute this
article, provided that you credit the author and journal.
Bookshelf ID: NBK526047 PMID: [ 30252303
](https://pubmed.ncbi.nlm.nih.gov/30252303 "PubMed record of this page")
Share
### Views
* [ PubReader ](/books/NBK526047/?report=reader)
* [ Print View ](/books/NBK526047/?report=printable)
* Cite this Page
Caire MJ, Reddy V, Varacallo M. Physiology, Synapse. [Updated 2023 Mar 27].
In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024
Jan-.
### In this Page
* Introduction
* Cellular Level
* Development
* Mechanism
* Clinical Significance
* Review Questions
* References
### Bulk Download
* [ Bulk download StatPearls data from FTP ](https://ftp.ncbi.nlm.nih.gov/pub/litarch/3d/12/)
### Related information
* [ PMC ](/books/?Db=pmc&DbFrom=books&Cmd=Link&LinkName=books_pmc_refs&IdsFromResult=4616406)
PubMed Central citations
* [ PubMed ](/books/?Db=pubmed&DbFrom=books&Cmd=Link&LinkName=books_pubmed_refs&IdsFromResult=4616406)
Links to PubMed
### Similar articles in PubMed
* [ Review Molecules involved in the formation of synaptic connections in muscle and brain. ](/pubmed/11245999) [Matrix Biol. 2001]
Review Molecules involved in the formation of synaptic connections in muscle
and brain.
_Ruegg MA._ _Matrix Biol. 2001 Feb; 20(1):3-12._
* [ Delivery of different genes into presynaptic and postsynaptic neocortical neurons connected by a BDNF-TrkB synapse. ](/pubmed/30710509) [Brain Res. 2019]
Delivery of different genes into presynaptic and postsynaptic neocortical
neurons connected by a BDNF-TrkB synapse.
_Nagayach A, Singh A, Geller AI._ _Brain Res. 2019 Jun 1; 1712:16-24. Epub
2019 Jan 30._
* [ Review The versatile synapse. ](/pubmed/6150966) [J Exp Biol. 1984]
Review The versatile synapse.
_Pitman RM._ _J Exp Biol. 1984 Sep; 112:199-224._
* [ Review Thematic Minireview Series: Molecular Mechanisms of Synaptic Plasticity. ](/pubmed/26453312) [J Biol Chem. 2015]
Review Thematic Minireview Series: Molecular Mechanisms of Synaptic
Plasticity.
_Colbran RJ._ _J Biol Chem. 2015 Nov 27; 290(48):28594-5. Epub 2015 Oct 9._
* [ Review In vivo measurements of neurotransmitters by microdialysis sampling. ](/pubmed/16570388) [Anal Chem. 2006]
Review In vivo measurements of neurotransmitters by microdialysis sampling.
_Watson CJ, Venton BJ, Kennedy RT._ _Anal Chem. 2006 Mar 1; 78(5):1391-9._
[ See reviews...
](/sites/entrez?db=pubmed&cmd=link&linkname=pubmed_pubmed_reviews&uid=30252303)
[ See all...
](/sites/entrez?db=pubmed&cmd=link&linkname=pubmed_pubmed&uid=30252303)
### Recent Activity
[ Clear ](javascript:historyDisplayState\('ClearHT'\)) [ Turn Off
](javascript:historyDisplayState\('HTOff'\)) [ Turn On
](javascript:historyDisplayState\('HTOn'\))
* [ Physiology, Synapse - StatPearls ](/portal/utils/pageresolver.fcgi?recordid=6630022d12fea5609aaaa64e)
Physiology, Synapse - StatPearls
Your browsing activity is empty.
Activity recording is turned off.
[ Turn recording back on ](javascript:historyDisplayState\('HTOn'\))
[ See more... ](/sites/myncbi/recentactivity)
Follow NCBI
[ Twitter ](https://twitter.com/ncbi) [ Facebook
](https://www.facebook.com/ncbi.nlm) [ LinkedIn
](https://www.linkedin.com/company/ncbinlm) [ GitHub
](https://github.com/ncbi) [ NCBI Insights Blog
](https://ncbiinsights.ncbi.nlm.nih.gov/)
[ Connect with NLM ](https://www.nlm.nih.gov/socialmedia/index.html)
* [ Twitter ](https://twitter.com/NLM_NIH)
* [ Facebook ](https://www.facebook.com/nationallibraryofmedicine)
* [ Youtube ](https://www.youtube.com/user/NLMNIH)
National Library of Medicine
[ 8600 Rockville Pike
Bethesda, MD 20894
](https://www.google.com/maps/place/8600+Rockville+Pike,+Bethesda,+MD+20894/@38.9959508,-77.101021,17z/data=!3m1!4b1!4m5!3m4!1s0x89b7c95e25765ddb:0x19156f88b27635b8!8m2!3d38.9959508!4d-77.0988323)
[ Web Policies ](https://www.nlm.nih.gov/web_policies.html)
[ FOIA ](https://www.nih.gov/institutes-nih/nih-office-director/office-
communications-public-liaison/freedom-information-act-office)
[ HHS Vulnerability Disclosure ](https://www.hhs.gov/vulnerability-disclosure-
policy/index.html)
[ Help ](https://support.nlm.nih.gov/)
[ Accessibility ](https://www.nlm.nih.gov/accessibility.html)
[ Careers ](https://www.nlm.nih.gov/careers/careers.html)
* [ NLM ](//www.nlm.nih.gov/)
* [ NIH ](https://www.nih.gov/)
* [ HHS ](https://www.hhs.gov/)
* [ USA.gov ](https://www.usa.gov/)

| biology | 4660065 | https://sv.wikipedia.org/wiki/Syzygium%20neurophyllum | Syzygium neurophyllum | Syzygium neurophyllum är en myrtenväxtart som beskrevs av Neil Snow. Syzygium neurophyllum ingår i släktet Syzygium och familjen myrtenväxter. Inga underarter finns listade i Catalogue of Life.
Källor
Myrtenväxter
neurophyllum | swedish | 0.957925 |
neuron_onto/thesynapse.txt | If you're seeing this message, it means we're having trouble loading external
resources on our website.
If you're behind a web filter, please make sure that the domains
***.kastatic.org** and ***.kasandbox.org** are unblocked.
__ To log in and use all the features of Khan Academy, please enable
JavaScript in your browser.
Courses
[ Search ](/search)
[ ](/)
[ Donate
](https://donate.khanacademy.org/give/419869/#!/donation/checkout?c_src2=Webapp.header)
[ Log in ](/login?continue=%2Fscience%2Fbiology%2Fhuman-biology%2Fneuron-
nervous-system%2Fa%2Fthe-synapse) [ Sign up
](/signup?continue=%2Fscience%2Fbiology%2Fhuman-biology%2Fneuron-nervous-
system%2Fa%2Fthe-synapse)
[ Search for courses, skills, and videos ](/search)
Main content
##
Biology library
### [ Course: Biology library ](/science/biology) > [ Unit 33
](/science/biology/human-biology)
Lesson 2: The neuron and nervous system
* [ Anatomy of a neuron ](/science/biology/human-biology/neuron-nervous-system/v/anatomy-of-a-neuron)
* [ Overview of neuron structure and function ](/science/biology/human-biology/neuron-nervous-system/a/overview-of-neuron-structure-and-function)
* [ The membrane potential ](/science/biology/human-biology/neuron-nervous-system/a/the-membrane-potential)
* [ Electrotonic and action potentials ](/science/biology/human-biology/neuron-nervous-system/v/electrotonic-action-potential)
* [ Saltatory conduction in neurons ](/science/biology/human-biology/neuron-nervous-system/v/saltatory-conduction-neurons)
* [ Neuronal synapses (chemical) ](/science/biology/human-biology/neuron-nervous-system/v/neuronal-synapses-chemical)
* [ The synapse ](/science/biology/human-biology/neuron-nervous-system/a/the-synapse)
* [ Neurotransmitters and receptors ](/science/biology/human-biology/neuron-nervous-system/a/neurotransmitters-their-receptors)
* [ Q & A: Neuron depolarization, hyperpolarization, and action potentials ](/science/biology/human-biology/neuron-nervous-system/a/depolarization-hyperpolarization-and-action-potentials)
* [ Overview of the functions of the cerebral cortex ](/science/biology/human-biology/neuron-nervous-system/v/overview-of-the-functions-of-the-cerebral-cortex)
[ Science ](/science) >
[ Biology library ](/science/biology) >
[ Human biology ](/science/biology/human-biology) >
[ The neuron and nervous system ](/science/biology/human-biology/neuron-
nervous-system)
© 2024 Khan Academy
[ Terms of use ](/about/tos) [ Privacy Policy ](/about/privacy-policy) [
Cookie Notice ](/about/cookie-policy)
# The synapse
[

Google Classroom ]()
How neurons communicate with each other at synapses. Chemical vs. electrical
synapses.
## Key points
* Neurons communicate with one another at junctions called **synapses** . At a synapse, one neuron sends a message to a target neuron—another cell.
* Most synapses are **chemical** ; these synapses communicate using chemical messengers. Other synapses are **electrical** ; in these synapses, ions flow directly between cells.
* At a chemical synapse, an action potential triggers the presynaptic neuron to release **neurotransmitters** . These molecules bind to receptors on the postsynaptic cell and make it more or less likely to fire an action potential.
## Introduction
A single neuron, or nerve cell, can do a lot! It can maintain a [ resting
potential ](/science/biology/human-biology/neuron-nervous-system/a/the-
membrane-potential) —voltage across the membrane. It can fire nerve impulses,
or [ action potentials ](/science/biology/human-biology/neuron-nervous-
system/v/electrotonic-action-potential) . And it can carry out the metabolic
processes required to stay alive.
A neuron’s signaling, however, is much more exciting—no pun intended!—when we
consider its interactions with other neurons. Individual neurons make
connections to target neurons and stimulate or inhibit their activity, forming
circuits that can process incoming information and carry out a response.
How do neurons "talk" to one another? The action happens at the **synapse** ,
the point of communication between two neurons or between a neuron and a
target cell, like a muscle or a gland. At the synapse, the firing of an action
potential in one neuron—the **presynaptic** , or sending, neuron—causes the
transmission of a signal to another neuron—the **postsynaptic** , or
receiving, neuron—making the postsynaptic neuron either more or less likely to
fire its own action potential.
Schematic of synaptic transmission. An action potential travels down the axon
of the pre-synaptic—sending—cell and arrives at the axon terminal. The axon
terminal is adjacent to the dendrite of the post-synaptic—receiving—cell. This
spot of close connection between axon and dendrite is the synapse.
In this article, we'll take a closer look at the synapse and the mechanisms
neurons use to send signals across it. To get the most out of this article,
you may want to learn about [ neuron structure ](/science/biology/human-
biology/neuron-nervous-system/v/anatomy-of-a-neuron) and [ action potentials
](/science/biology/human-biology/neuron-nervous-system/v/electrotonic-action-
potential) first.
## Electrical or chemical transmission?
At the end of the 19th and beginning of the 20th century, there was a lot of
controversy about whether synaptic transmission was electrical or chemical.
* Some people thought that signaling across a synapse involved the flow of ions directly from one neuron into another—electrical transmission.
* Other people thought it depended on the release of a chemical from one neuron, causing a response in the receiving neuron—chemical transmission.
We now know that synaptic transmission can be either electrical or chemical—in
some cases, both at the same synapse!
Chemical transmission is more common, and more complicated, than electrical
transmission. So, let's take a look at chemical transmission first.
## Overview of transmission at chemical synapses
Chemical transmission involves release of chemical messengers known as
**neurotransmitters** . Neurotransmitters carry information from the pre-
synaptic—sending—neuron to the post-synaptic—receiving—cell.
As you may remember from the article on [ neuron structure and function
](/science/biology/human-biology/neuron-nervous-system/a/overview-of-neuron-
structure-and-function) , synapses are usually formed between nerve
terminals—axon terminals—on the sending neuron and the cell body or dendrites
of the receiving neuron.
Schematic of synaptic transmission. An action potential travels down the axon
of the presynaptic—sending—cell and arrives at multiple axon terminals
branching off from the axon. The axon terminal is adjacent to the dendrite of
the postsynaptic—receiving—cell. This spot of close connection between axon
and dendrite is the synapse.
A single axon can have multiple branches, allowing it to make synapses on
various postsynaptic cells. Similarly, a single neuron can receive thousands
of synaptic inputs from many different presynaptic—sending—neurons.
Inside the axon terminal of a sending cell are many **synaptic vesicles** .
These are membrane-bound spheres filled with neurotransmitter molecules. There
is a small gap between the axon terminal of the presynaptic neuron and the
membrane of the postsynaptic cell, and this gap is called the **synaptic
cleft** .
Image showing pre-synaptic cell's axon terminal containing synaptic vesicles
with neurotransmitters. Voltage-gated calcium channels are on the outside
surface of the axon terminal. Across the synaptic cleft, there is the post-
synaptic cell surface covered in receptors (ligand-gated ion channels) for the
neurotransmitter.
When an action potential, or nerve impulse, arrives at the axon terminal, it
activates voltage-gated calcium channels in the cell membrane. Ca 2 \+
, which is present at a much higher concentration outside the neuron than
inside, rushes into the cell. The Ca 2 \+ allows synaptic vesicles to
fuse with the axon terminal membrane, releasing neurotransmitter into the
synaptic cleft.
Image showing what happens when action potential arrives at axon terminal,
causing ion flow and depolarization of target cell. Step by step: 1\. Action
potential reaches axon terminal and depolarizes membrane. 2\. Voltage-gated
calcium channels open and calcium ions flow in. 3\. Calcium ion influx
triggers synaptic vesicles to release neurotransmitter. 4\. Neurotransmitter
binds to receptors on target cell (in this case, causing positive ions to flow
in).
The molecules of neurotransmitter diffuse across the synaptic cleft and bind
to receptor proteins on the postsynaptic cell. Activation of postsynaptic
receptors leads to the opening or closing of ion channels in the cell
membrane. This may be **depolarizing** —make the inside of the cell more
positive—or **hyperpolarizing** —make the inside of the cell more
negative—depending on the ions involved.
In some cases, these effects on channel behavior are direct: the receptor is a
ligand-gated ion channel, as in the diagram above. In other cases, the
receptor is not an ion channel itself but activates ion channels through a
signaling pathway. See the article on [ neurotransmitters and receptors
](/science/biology/human-biology/neuron-nervous-system/a/neurotransmitters-
their-receptors) for more info.
## Excitatory and inhibitory postsynaptic potentials
When a neurotransmitter binds to its receptor on a receiving cell, it causes
ion channels to open or close. This can produce a localized change in the
membrane potential—voltage across the membrane—of the receiving cell.
* In some cases, the change makes the target cell _more_ likely to fire its own action potential. In this case, the shift in membrane potential is called an **excitatory postsynaptic potential** , or **EPSP** .
* In other cases, the change makes the target cell _less_ likely to fire an action potential and is called an **inhibitory post-synaptic potential** , or **IPSP** .
An EPSP is depolarizing: it makes the inside of the cell more positive,
bringing the membrane potential closer to its threshold for firing an action
potential. Sometimes, a single EPSP isn't large enough bring the neuron to
threshold, but it can sum together with other EPSPs to trigger an action
potential.
IPSPs have the opposite effect. That is, they tend to keep the membrane
potential of the postsynaptic neuron below threshold for firing an action
potential. IPSPs are important because they can counteract, or cancel out, the
excitatory effect of EPSPs.
## Spatial and temporal summation
How do EPSPs and IPSPs interact? Basically, a postsynaptic neuron adds
together, or integrates, all of the excitatory and inhibitory inputs it
receives and “decides” whether to fire an action potential.
* The integration of postsynaptic potentials that occur in different locations—but at about the same time—is known as **spatial summation** .
* The integration of postsynaptic potentials that occur in the same place—but at slightly different times—is called **temporal summation** .
For instance, let’s suppose that excitatory synapses are made on two different
dendrites of the same postsynaptic neuron, as shown below. Neither synapse can
produce an EPSP quite large enough to bring the membrane potential to
threshold at the axon hillock—the place where the action potential is
triggered, boxed below. If both subthreshold EPSPs occurred at the same time,
however, they could sum, or add up, to bring the membrane potential to
threshold.
Illustration of spatial summation. A neuron has two synapses onto two
different dendrites, both of which are excitatory. Neither synapse produces a
large enough excitatory postsynaptic potential, EPSP, when it signals to
generate an action potential at the hillock— the place where the axon joins
the cell body and where the action potential is initiated. However, when the
synapses fire at nearly the same time, the EPSPs add up to produce an above-
threshold depolarization, triggering an action potential.
This process is shown on a graph of voltage in millivolts vs. time in
milliseconds. The graph monitors the membrane potential—voltage—at the axon
hillock. Initially, it is at –70 mV, the resting potential. Then, one synapse
fires, resulting in a small depolarization to roughly –60 mV. This is not
sufficient to reach the threshold of –55 mV. However, just a tiny bit later,
the other synapse fires, and it "adds on" to the first depolarization,
resulting in a total depolarization that reaches –55 mV and triggers an action
potential—depolarization to +40 mV, followed by a repolarization and
hyperpolarization below –90 mV, and then a gradual recovery to –70 mV, the
resting membrane potential.
Image credit: modified from _[ Communication between neurons: Figure 2
](http://cnx.org/contents/[email protected]:p74vr6PZ@3/Communication-Between-
Neurons) _ by OpenStax College, Anatomy & Physiology, [ CC BY 3.0
](http://creativecommons.org/licenses/by/3.0/) and _[ Action potential
](https://commons.wikimedia.org/wiki/File:Action_potential.svg) _ by tiZom, [
CC BY-SA 3.0 ](https://creativecommons.org/licenses/by-sa/3.0/deed.en) ; the
modified image is licensed under a [ CC BY-SA 3.0
](https://creativecommons.org/licenses/by-sa/3.0/deed.en) license
On the other hand, if an IPSP occurred together with the two EPSPs, it might
prevent the membrane potential from reaching threshold and keep the neuron
from firing an action potential. These are examples of spatial summation.
What about temporal summation? A key point is that postsynaptic potentials
aren’t instantaneous: instead, they last for a little while before they
dissipate. If a presynaptic neuron fires quickly twice in row, causing two
EPSPs, the second EPSP may arrive before the first one has dissipated, bumping
the membrane potential above threshold. This is an example of temporal
summation.
## Signal termination
A synapse can only function effectively if there is some way to "turn off" the
signal once it's been sent. Termination of the signal lets the postsynaptic
cell return to its normal resting potential, ready for new signals to arrive.
For the signal to end, the synaptic cleft must be cleared of neurotransmitter.
There are a few different ways to get this done. The neurotransmitter may be
broken down by an enzyme, it may be sucked back up into the presynaptic
neuron, or it may simply diffuse away. In some cases, neurotransmitter can
also be "mopped up" by nearby glial cells—not shown in the diagram below.
Reuptake by the presynaptic neuron, enzymatic degradation, and diffusion away
from the synapse reduce neurotransmitter levels, terminating the signal.
Image credit: modified from _[ Nervous system: Figure 9
](http://cnx.org/contents/pMqJxKsZ@7/Nervous-System) _ by OpenStax College,
Biology, adapted by Robert Bear and David Rintoul, [ CC BY 4.0
](http://cnx.org/contents/pMqJxKsZ@7/Nervous-System)
Anything that interferes with the processes that terminate the synaptic signal
can have significant physiological effects. For instance, some insecticides
kill insects by inhibiting an enzyme that breaks down the neurotransmitter
acetylcholine. On a more positive note, drugs that interfere with reuptake of
the neurotransmitter serotonin in the human brain are used as antidepressants,
for example, Prozac. 1
## Chemical synapses are flexible
If you've learned about action potentials, you may remember that the action
potential is an all-or-none response. That is, it either happens at its full
strength, or it doesn't happen at all.
Synaptic signaling, on the other hand, is much more flexible. For instance, a
sending neuron can "dial up" or "dial down" the amount of neurotransmitter it
releases in response to the arrival of an action potential. Similarly, a
receiving cell can alter the number of receptors it puts on its membrane and
how readily it responds to activation of those receptors. These changes can
strengthen or weaken communication at a particular synapse.
Presynaptic and postsynaptic cells can dynamically change their signaling
behavior based on their internal state or the cues they receive from other
cells. This type of plasticity, or capacity for change, makes the synapse a
key site for altering neural circuit strength and plays a role in learning and
memory. Synaptic plasticity is also involved in addiction.
In addition, different presynaptic and postsynaptic cells produce different
neurotransmitters and neurotransmitter receptors, with different interactions
and different effects on the postsynaptic cell. For more information, take a
look at the article on [ neurotransmitters and receptors
](/science/biology/human-biology/neuron-nervous-system/a/neurotransmitters-
their-receptors) .
## Electrical synapses
At **electrical synapses** , unlike chemical synapses, there is a direct
physical connection between the presynaptic neuron and the postsynaptic
neuron. This connection takes the form of a channel called a **gap junction**
, which allows current—ions—to flow directly from one cell into another.
Electrical synapse showing presynaptic cell, gap junction, post-synaptic cell,
and movement of positive ions from pre-synaptic cell to post-synaptic cell.
Image credit: based on similar image in Pereda 2 , Figure 1
Electrical synapses transmit signals more rapidly than chemical synapses do.
Some synapses are both electrical and chemical. At these synapses, the
electrical response occurs earlier than the chemical response.
What are the benefits of electrical synapses? For one thing, they're
fast—which could be important, say, in a circuit that helps an organism escape
from a predator. Also, electrical synapses allow for the synchronized activity
of groups of cells. In many cases, they can carry current in both directions
so that depolarization of a postsynaptic neuron will lead to depolarization of
a presynaptic neuron. This kind of bends the definitions of presynaptic and
postsynaptic!
What are the downsides of electrical synapses? Unlike chemical synapses,
electrical synapses cannot turn an excitatory signal in one neuron into an
inhibitory signal in another. More broadly, they lack the versatility,
flexibility, and capacity for signal modulation that we see in chemical
synapses.
References
This article is licensed under a [ CC BY-NC-SA 4.0
](https://creativecommons.org/licenses/by-nc-sa/4.0/) license.
## Works cited
2. David E. Sadava, David M. Hillis, H. Craig Heller, and May Berenbaum, "How Do Neurons Communicate with Other Cells?" In _Life: The Science of Biology_ , 9th ed. (Sunderland: Sinauer Associates, 2009), 961.
3. Alberto E. Pereda, "Electrical Synapses and Their Functional Interactions with Chemical Synapses," _Nature Reviews Neuroscience_ 15 (2014): 250-263, [ http://dx.doi.org/10.1038/nrn3708 ](http://dx.doi.org/10.1038/nrn3708) .
## References
Kandel, E.R., J. H. Schwartz, and T. M. Jessell. "An Introduction to Synaptic
Transmission." In _Essentials of Neuroscience and Behavior_ , 179-195.
Norwalk: Appleton & Lange, 1995.
Loewi, Otto. "Nobel Lecture: The Chemical Transmission of Nerve Action."
NobelPrize.org. Accessed March 22, 2016. [
http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/loewi-
lecture.html
](http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/loewi-
lecture.html) .
Nicholls, J.G., A. R. Martin, B. G. Wallace, and P. A. Fuchs. "Principles of
Direct Synaptic Transmission." In _From Neuron to Brain_ , 155-176. 4th ed.
Sunderland: Sinauer Associates, 2001.
Openstax College, Biology. "How Neurons Communicate." OpenStax CNX. Last
modified February 29, 2016. [ http://cnx.org/contents/[email protected]:cs_Pb-
GW@5/How-Neurons-Communicate ](http://cnx.org/contents/[email protected]:cs_Pb-
GW@5/How-Neurons-Communicate) .
Pereda, Alberto E. "Electrical Synapses and Their Functional Interactions with
Chemical Synapses." _Nature Reviews Neuroscience_ 15 (2014): 250-263. [
http://dx.doi.org/10.1038/nrn3708 ](http://dx.doi.org/10.1038/nrn3708) .
Purves, D., G. J. Augustine, D. Fitzpatrick, L. C. Katz, A.-S. LaMantia, and
J. O. McNamara. "Synaptic Transmission." In _Neuroscience_ , 85-98.
Sunderland: Sinauer Associates, 1997.
Reece, Jane B., Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V.
Minorsky, and Robert B. Jackson. "Neurons Communicate with Other Cells at
Synapses." In _Campbell Biology_ , 1076. 10th ed. San Francisco: Pearson,
2011.
Sadava, David E., David M. Hillis, H. Craig Heller, and May Berenbaum. "How Do
Neurons Communicate with Other Cells?" In _Life: The Science of Biology_ ,
956-962. 9th ed. Sunderland: Sinauer Associates, 2009.
## Suggestions for further reading
Dale, Henry. "Nobel Lecture: Some Recent Extensions of the Chemical
Transmission of the Effects of Nerve Impulses." NobelPrize.org. Accessed March
22, 2016. [
http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/dale-
lecture.html
](http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/dale-
lecture.html) .
Loewi, Otto. "Nobel Lecture: The Chemical Transmission of Nerve Action."
NobelPrize.org. Accessed March 22, 2016. [
http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/loewi-
lecture.html
](http://www.nobelprize.org/nobel_prizes/medicine/laureates/1936/loewi-
lecture.html) .
Sakmann, Bert. "Sir Bernard Katz. 26 March 1911 - 20 April 2003." _Biogr. Mems
Fell. R. Soc._ 53 (2007): 185-202. [ http://dx.doi.org/10.1098/rsbm.2007.0013
](http://dx.doi.org/10.1098/rsbm.2007.0013) .
Questions Tips & Thanks
## Want to join the conversation?
[ Log in ](/login?continue=%2Fscience%2Fbiology%2Fhuman-biology%2Fneuron-
nervous-system%2Fa%2Fthe-synapse%3FloggedInViaDiscussion%3Dtrue)
Sort by:
Top Voted
* [  ](/profile/kaid_322754331915428616537245/discussion)
[ anshuman28dubey ](/profile/kaid_322754331915428616537245/discussion)
[ 8 years ago Posted 8 years ago. Direct link to anshuman28dubey's post “is
there any thing betwee...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfMzIyNzU0MzMxOTE1NDI4NjE2NTM3MjQ1DAsSCEZlZWRiYWNrGICAgIDAnIIKDA&qa_expand_type=question)
more
is there any thing between the synaps any fluid or anyting else?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 17 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_323215414219590867697280/discussion)
[ Ilka Plesse ](/profile/kaid_323215414219590867697280/discussion)
[ 8 years ago Posted 8 years ago. Direct link to Ilka Plesse's post “Yes, the
synaptic cleft i...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfMzIzMjE1NDE0MjE5NTkwODY3Njk3MjgwDAsSCEZlZWRiYWNrGICAgIDg2o0KDA&qa_expand_type=answer)


more
Yes, the synaptic cleft is filled with extracellular fluid.
Comment Button navigates to signup page
( 28 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Show more...
* [  ](/profile/kaid_244726536382598737957354/discussion)
[ Anthony Thomas ](/profile/kaid_244726536382598737957354/discussion)
[ 7 years ago Posted 7 years ago. Direct link to Anthony Thomas's post “I
know this article talke...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfMjQ0NzI2NTM2MzgyNTk4NzM3OTU3MzU0DAsSCEZlZWRiYWNrGICAgIDg2o0KDA&qa_expand_type=question)
more
I know this article talked about the flexibility of synapses, but I still
don't understand how different kinds of signals can be transmitted. Does a
certain neuron only send one kind of signal only (different signals would be
sent by different neurons) or does a certain neuron send multiple kinds of
signals by sending different kinds of neurotransmitters? If the former, how
are specific cells targeted? If the latter, how are specific neurotransmitters
released?
Answer Button navigates to signup page • 1 comment Comment on Anthony
Thomas's post “I know this article talke...”
( 13 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “Each neuron may be connec...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3sPWz9EKDA&qa_expand_type=answer)

more
Each neuron may be connected to up to 10,000 other neurons, passing signals to
each other via as many as 1,000 trillion synaptic connections.
Meaning that a **certain neuron** sends multiple kinds of signals by sending
different kinds of neurotransmitters.
Functionally related neurons connect to form neural networks (also known as
neural nets or assemblies). The connections between neurons are not static,
though, they change over time. The more signals sent between two neurons, the
stronger the connection grows.
When stimulated by an electrical pulse, neurotransmitters of various types are
released, and they cross the cell membrane into the synaptic gap between
neurons. These chemicals then bind to chemical receptors in the dendrites of
the receiving (post-synaptic) neuron. In the process, they cause changes in
the permeability of the cell membrane to specific ions, opening up special
gates or channels which let in a flood of charged particles (ions of calcium,
sodium, potassium, and chloride).
Meaning that upon stimulation, **many** neurotransmitters are being released
into the synaptic cleft.
[ https://human-memory.net/brain-neurons-synapses/ ](https://human-
memory.net/brain-neurons-synapses/)
But there a thing called membrane capacitance. Another direct measure of
exocytosis is the increase in membrane area due to the incorporation of the
secretory granule or vesicle membrane into the plasma membrane. This can be
measured by increases in membrane capacitance (Cm).
The specific capacitance is mainly determined by the thickness and dielectric
constant of the phospholipid bilayer membrane and is similar for intracellular
organelles and the plasma membrane.
[ https://www.ncbi.nlm.nih.gov/books/NBK27911/
](https://www.ncbi.nlm.nih.gov/books/NBK27911/)
There are only differences between fast secreting neurotransmitters (Acth,
dopamine) and slow releasing neuropeptides from neuroendocrine cells.
Comment Button navigates to signup page
( 12 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_580948424022679867231947/discussion)
[ Mark Young ](/profile/kaid_580948424022679867231947/discussion)
[ 6 years ago Posted 6 years ago. Direct link to Mark Young's post “The
'Synaptic Cleft' has ...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNTgwOTQ4NDI0MDIyNjc5ODY3MjMxOTQ3DAsSCEZlZWRiYWNrGICAgICA5JEKDA&qa_expand_type=question)
more
The 'Synaptic Cleft' has an approximately 20 nm separation, How do the
axion/dendrite pre/post synaptic terminals maintain their correct separation,
that is, how do they stop themselves from touching or stop themselves from
separating too far?
And if there is indeed- Failures in this gap separation, then what would be
the diseases associated with both the touching condition and the over
separation condition of the terminals?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 9 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_244565626671905857566821/discussion)
[ Gopu Kapoor ](/profile/kaid_244565626671905857566821/discussion)
[ 5 years ago Posted 5 years ago. Direct link to Gopu Kapoor's post “In the
Synaptic Cleft, th...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfMjQ0NTY1NjI2NjcxOTA1ODU3NTY2ODIxDAsSCEZlZWRiYWNrGICAgICeyZcKDA&qa_expand_type=answer)
more
In the Synaptic Cleft, there are neurotransmitters that are diffusing from one
neuron to the next neuron, and then undergoing receptor-mediated endocytosis
with the receptors in the "receiving" neuron (which should require space for
receiving them). These neurotransmitters would diffuse away from the synaptic
cleft or an enzyme would help clear the rest of the neurotransmitters. I
believe that for these enzymes to be able to "sweep away" those
neurotransmitters, some space is needed. Furthermore, as only milliseconds
pass between some action potentials, the synaptic cleft maintains its
distance.
Comment Button navigates to signup page
( 2 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_578486592868103296365042/discussion)
[ Julia Pudar ](/profile/kaid_578486592868103296365042/discussion)
[ 6 years ago Posted 6 years ago. Direct link to Julia Pudar's post “in
"Overview of transmiss...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNTc4NDg2NTkyODY4MTAzMjk2MzY1MDQyDAsSCEZlZWRiYWNrGICAgICAuYQKDA&qa_expand_type=question)
more
in "Overview of transmission at chemical synapses," it was stated that a
depolarization of the membrane causes an influx of Ca2+ ions into the cell.
However, doesn't this influx on positive charge cause depolarization of the
cell?
I don't understand why depolarizing the membrane would stimulate further
depolarization. Wouldn't hyper-polarization of the membrane cause this?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 6 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “You got confused because ...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3qPahPoLDA&qa_expand_type=answer)
more
You got confused because both carry a positive charge. I will tell you that
speaking of the number of ions, this is almost minor.
Yes, we say an influx of Ca+ ions, but there are not many Ca+ ions. Sometimes,
only one is enough to bind to SNARE complex of one vesicle to release
neurotransmitters-
Comment Button navigates to signup page
( 6 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_987453463799803618775127/discussion)
[ Adithya Sharanya ](/profile/kaid_987453463799803618775127/discussion)
[ 6 years ago Posted 6 years ago. Direct link to Adithya Sharanya's post
“what makes an EPSP or IPS...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfOTg3NDUzNDYzNzk5ODAzNjE4Nzc1MTI3DAsSCEZlZWRiYWNrGICAgICg55gKDA&qa_expand_type=question)
more
what makes an EPSP or IPSP, how are they determined to be excitatory or
inhibitory?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 5 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “EPSPs are graded potentia...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3qPLrLQJDA&qa_expand_type=answer)
more
EPSPs are graded potentials that can initiate an AP in the axon, whereas IPSPs
produce a graded potential that lessens the chance of an AP in an axon.
I found one paper where it was investigated, but again - Cell-autonomous
molecular mechanisms that control the balance of excitatory and inhibitory
synapse function remain poorly understood; no proteins that regulate
excitatory and inhibitory synapse strength in a coordinated reciprocal manner
have been identified.
The knockdown of cadherin-10 reduces excitatory but increases inhibitory
synapse size and strength.
[ https://www.ncbi.nlm.nih.gov/pubmed/29030434
](https://www.ncbi.nlm.nih.gov/pubmed/29030434)
Comment Button navigates to signup page
( 2 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Show more...
* [  ](/profile/kaid_1071742132013690487345956/discussion)
[ Sunny Yu ](/profile/kaid_1071742132013690487345956/discussion)
[ 5 years ago Posted 5 years ago. Direct link to Sunny Yu's post “Where is
the ACTH broken ...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJBCxIIVXNlckRhdGEiHmthaWRfMTA3MTc0MjEzMjAxMzY5MDQ4NzM0NTk1NgwLEghGZWVkYmFjaxiAgN6I1pj3CAw&qa_expand_type=question)
more
Where is the ACTH broken down into ethanoic acid and choline by the
acetylcholinesterase, in the cleft, or postsynaptic neuron? It is definitely
broken down after it enters the postsynaptic neuron, right? Why ACTH can not
go back to the presynaptic neuron directly, but has to be broken down and
brought back?
Answer Button navigates to signup page • 1 comment Comment on Sunny Yu's
post “Where is the ACTH broken ...”
( 4 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “Acetylcholine interacts w...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3oPI7YUIDA&qa_expand_type=answer)
more
Acetylcholine interacts with postsynaptic receptors a few milliseconds before
it is being degraded down by acetylcholinesterase. Then both taken up by
presynaptic nerve terminal and recycled.
Meaning that degradation happens in the synaptic cleft, actually on the
postsynaptic neurons, but right after it already finished its role.
[ https://www.us.elsevierhealth.com/vst-nurse-anesthesia-e-
book-9780323444378.html?dmnum ](https://www.us.elsevierhealth.com/vst-nurse-
anesthesia-e-book-9780323444378.html?dmnum) =12449
Comment Button navigates to signup page
( 2 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_363573332064498997399332/discussion)
[ Hieu Le ](/profile/kaid_363573332064498997399332/discussion)
[ 5 years ago Posted 5 years ago. Direct link to Hieu Le's post “How did cell
membrane evo...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfMzYzNTczMzMyMDY0NDk4OTk3Mzk5MzMyDAsSCEZlZWRiYWNrGICA5t6_ndUKDA&qa_expand_type=question)
more
How did cell membrane evolve in synaptic clefts?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 3 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “A primordial neurosecreto...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3qP6losIDA&qa_expand_type=answer)
more
A primordial neurosecretory apparatus in choanoflagellates was identified and
it was found that the mechanism, by which presynaptic proteins required for
secretion of neurotransmitters interact, is conserved in choanoflagellates and
metazoans. Moreover, studies on the postsynaptic protein homolog Homer
revealed unexpected localization patterns in choanoflagellates and new binding
partners, both of which are conserved in metazoans.
I think this paper will satisfy you, everyzhing int he one place:
[ https://jeb.biologists.org/content/218/4/506
](https://jeb.biologists.org/content/218/4/506)
Comment Button navigates to signup page
( 3 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_173770543983285893762140/discussion)
[ natascha.b2000 ](/profile/kaid_173770543983285893762140/discussion)
[ 6 years ago Posted 6 years ago. Direct link to natascha.b2000's post “Why
are the neurotransmit...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfMTczNzcwNTQzOTgzMjg1ODkzNzYyMTQwDAsSCEZlZWRiYWNrGICAgICA5JEKDA&qa_expand_type=question)
more
Why are the neurotransmitter molecules cleaved so fast in the synaptic cleft (
50molecules/ms)?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 3 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “Because it could cause po...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3qO5t_gKDA&qa_expand_type=answer)
more
Because it could cause potential disadvantages:
1\. further delaying of response
2\. overcrowding of neurotransmitters and too enhanced answer
3\. what if new stimulation happens in a short time and neurotransmitters are
not recycled? Who would respond to it then?
Comment Button navigates to signup page
( 2 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_814250287910645312371081/discussion)
[ somto luck ](/profile/kaid_814250287910645312371081/discussion)
[ 6 years ago Posted 6 years ago. Direct link to somto luck's post “how do
synapses affect yo...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfODE0MjUwMjg3OTEwNjQ1MzEyMzcxMDgxDAsSCEZlZWRiYWNrGICAgICA5JEKDA&qa_expand_type=question)
more
how do synapses affect your reaction time?
Answer Button navigates to signup page • 1 comment Comment on somto luck's
post “how do synapses affect yo...”
( 3 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
* [  ](/profile/kaid_747203976436023184790018/discussion)
[ Ivana - Science trainee
](/profile/kaid_747203976436023184790018/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Ivana - Science trainee's
post “They little delay the rea...” ](/science/biology/human-biology/neuron-
nervous-system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJACxIIVXNlckRhdGEiHWthaWRfNzQ3MjAzOTc2NDM2MDIzMTg0NzkwMDE4DAsSCEZlZWRiYWNrGICA3sP0oMgKDA&qa_expand_type=answer)
more
They little delay the reaction.
The overall synaptic delay and estimated number of synapses (ENOS) of simple
tactile reaction neuronal circuits of normal subjects did not significantly
vary with site of tactile stimulation or effector organ.
The overall synaptic delay in the tactile reaction neuronal circuits between
SOS and the left and right big toes were significantly lower in sniffers than
in control subjects. This may be due to a decrease in either the average
synaptic delay, the number of synapses, or both in the tactile reaction
neuronal circuits between sites of stimulation and big toes (but not index
fingers) in sniffers.
[ https://www.ncbi.nlm.nih.gov/pubmed/3393601
](https://www.ncbi.nlm.nih.gov/pubmed/3393601)
Comment Button navigates to signup page
( 2 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
* [  ](/profile/kaid_1144457361975706908217650/discussion)
[ Sana Awan ](/profile/kaid_1144457361975706908217650/discussion)
[ 4 years ago Posted 4 years ago. Direct link to Sana Awan's post “Can we see
electrical syn...” ](/science/biology/human-biology/neuron-nervous-
system/a/the-
synapse?qa_expand_key=ag5zfmtoYW4tYWNhZGVteXJBCxIIVXNlckRhdGEiHmthaWRfMTE0NDQ1NzM2MTk3NTcwNjkwODIxNzY1MAwLEghGZWVkYmFjaxiAgLWt7_7ICQw&qa_expand_type=question)
more
Can we see electrical synapses in FMRI just like its name electrical? Do the
both look like the same in FMRI?
Answer Button navigates to signup page • Comment Button navigates to
signup page
( 3 votes )
* Upvote
Button navigates to signup page
* Downvote
Button navigates to signup page
* Flag
Button navigates to signup page
more
Answer

Show preview Show formatting options
Post answer
Loading...
| biology | 4662406 | https://sv.wikipedia.org/wiki/Syzygium%20neurocalyx | Syzygium neurocalyx | Syzygium neurocalyx är en myrtenväxtart som först beskrevs av Asa Gray, och fick sitt nu gällande namn av Erling Christophersen. Syzygium neurocalyx ingår i släktet Syzygium och familjen myrtenväxter. Inga underarter finns listade i Catalogue of Life.
Källor
Myrtenväxter
neurocalyx | swedish | 1.05762 |
neuron_onto/PMC7203343.txt | U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Bookshelf
Search database
Books
Search term
Search
Browse Titles AdvancedHelpDisclaimer
Cover of StatPearls
StatPearls [Internet].
Show details
Search term
Physiology, Synapse
Michael J. Caire; Vamsi Reddy; Matthew Varacallo.
Author Information and Affiliations
Last Update: March 27, 2023.
Go to:
Introduction
The human brain is made up of approximately 86 billion neurons that “talk” to each other using a combination of electrical and chemical (electrochemical) signals.
The places where neurons connect and communicate with each other are called synapses. Each neuron has anywhere between a few to hundreds of thousands of synaptic connections, and these connections can be with itself, neighboring neurons, or neurons in other regions of the brain. A synapse is made up of a presynaptic and postsynaptic terminal.
The presynaptic terminal is at the end of an axon and is the place where the electrical signal (the action potential) is converted into a chemical signal (neurotransmitter release). The postsynaptic terminal membrane is less than 50 nanometers away and contains specialized receptors. The neurotransmitter rapidly (in microseconds) diffuses across the synaptic cleft and binds to specific receptors.
The type of neurotransmitter released from the presynaptic terminal and the specific receptors present on the corresponding postsynaptic terminal is critical in determining the quality and intensity of information transmitted by neurons. The postsynaptic neuron integrates all the signals it receives to determine what it does next, for example, to fire an action potential of its own or not. [1][2]
Go to:
Cellular Level
Neurons
In the simplest sense, the neuron consists of a cell body, axons, and dendrites.
Cell Body
The cell body contains the nucleus and is the site of metabolic activity. Most of the neurotransmitters that will eventually be released at the synapse are synthesized here.
Dendrites
These are small projections from the cell body that serves a receptive role in the physiology of the neuron. They receive incoming signals from other neurons and relay them to the cell body, where the signals are integrated, and a response will be initiated.
Axons
Generally, the outflow tract of the neuron. It is a cylindrical tube that is covered by the axolemma and is supported by neurofilaments and microtubules. The microtubules will help to transport the neurotransmitters from the cell body down to the pre-synaptic terminal, where they will be released.
Synapses
The synapse itself is the site of transmission from the pre-synaptic neuron to the post-synaptic neuron. The structures found on either side of the synapse vary depending on the type of synapse:
Axodendritic
A connection formed between the axon of one neuron and the dendrite of another. These tend to be excitatory synapses.
Axosomatic
A direct connection between the axon of one neuron to the cell body of another neuron. These tend to be inhibitory synapses.
Axoaxonic
A connection between the terminal of one axon and another axon. These synapses generate serve a regulatory role; the afferent axon will modulate the release of neurotransmitters from the efferent axon.
The above discussion focuses on chemical synapses, which involve the release of a chemical neurotransmitter between the 2 neurons. This is the most common type of synapse in the mammalian central nervous system (CNS). However, it is important to note that there are electrical synapses, where electrical current (or signals) will pass directly from one neuron to another through gap junctions. The differences between the two will be expanded on in the mechanism section. [3][4]
Go to:
Development
Two neurons form the neurological synapse, or in some instances, a neuron and an anatomical structure. This review will focus on 2 neurons composing the synapse. Neurons initially develop from the embryonic neural tube, which has 3 layers:
The ventricular zone, which surrounds the central canal of the tube. This tube will eventually become the ependyma.
The intermediate zone, which is formed by dividing cells of the ventricular zone. This zone stretches from the outermost portion of the ventricular zone to the outermost layer of the neural tube, known as the pial layer.
The marginal zone, which is formed by extensions of the nerve cells of the intermediate zone.
The intermediate zone will go on to form the gray matter, while the nerve processes that make up the marginal zone will become white matter once myelinated.
The neurons must then differentiate from their precursors. The order in which they do this is based upon their size, with the largest neurons (motor neurons) differentiating first. Around the time of birth, the smaller neurons (sensory neurons) will develop, along with glial cells. Glial cells are cells that will aid in the differentiation of the neurons and will facilitate their growth in the direction of their target locations. Later, glial cells will participate in the reuptake of excess neurotransmitters in the synaptic cleft.
Go to:
Mechanism
Synapses
As previously mentioned, there are 2 major types of synapses: electrical and chemical.
In mammals, the majority of synapses are chemical. Chemical synapses can be differentiated from electrical synapses by a few distinguishing criteria: they use neurotransmitters to relay the signal and vesicles are used to store and transport the neurotransmitter from the cell body to the terminal; furthermore, the pre-synaptic terminal will have a very active membrane and the post-synaptic membrane consists of a thick cell membrane made up of many receptors. In between these 2 membranes is a very distinct cleft (easily visualized with electron microscopy) and the chemical neurotransmitter released must diffuse across this cleft to elicit a response in the receptive neuron. Because of this, the synaptic delay, defined as the time it takes for current in the pre-synaptic neuron to be transmitted to the post-synaptic neuron, is approximately 0.5 to 1.0 ms.
This is different from the electrical synapse, which will typically consist of 2 membranes located much closer to each other than in a chemical synapse. These membranes possess channels formed by proteins known as connexins, which allow the direct passage of current from one neuron to the next and do not rely on neurotransmitters. The synaptic delay is significantly shorter in electrical synapses versus chemical synapses.
The rest of the discussion will focus on chemical synapses, which have a lot of variation and diversity of their own. They vary not only between shape and structure, but also the chemical that is transmitted. Synapses can be excitatory or inhibitory, and use a variety of chemical molecules and proteins that will be discussed shortly.
Multiple types of neurotransmitters used in synaptic communication including, but not limited to:
Acetylcholine (ACh): One of the most important neurotransmitters found in multiple synapses in the body, including, but not limited to, the neuromuscular junction, autonomic ganglia, caudate nucleus, and the limbic system. Generally, ACh is an excitatory neurotransmitter at the neuromuscular junction and in the autonomic ganglia. In the brain, Ach is synthesized in the basal nucleus of Meynert.
Norepinephrine (NE): The most important molecule in sympathetic nervous system signaling, except for the sweat glands. In the brain, NE is mainly found in the locus coeruleus and lateral tegmental nuclei.
Dopamine (DA): Dopamine signaling is generally inhibitory. There are three major dopaminergic pathways in the brain, the nigrostriatal, mesolimbic, and mesocortical; each of which serve different roles. One of the most well-known disease states involving dopamine is Parkinson's disease, where there is degeneration of dopaminergic neurons in the substantia nigra.
Serotonin (5-HT): Produced from tryptophan using tryptophan hydroxylase, which is mostly found in the brain (raphe nucleus) and the gastrointestinal (GI) tract. Serotonin is mostly known for its role as a regulatory neurotransmitter and is therefore implicated in various mood states and diseases.
Other common neurotransmitters include other catecholamines, gamma-aminobutyric acid (GABA), glycine, and glutamic acid.
The easiest approach to understanding synaptic transmission is to think of it as a stepwise process beginning with the synthesis of the neurotransmitter and ending with its release.
Synthesis: The neurotransmitter is synthesized in the cell body, where it will then be transmitted down the microtubules of the axon to the pre-synaptic terminal, or it is synthesized directly in the pre-synaptic terminal from recycled neurotransmitters. The neurotransmitter is then stored in presynaptic vesicles until its release.
Release: The neurotransmitter is released in a regulated fashion from the pre-synaptic neuron into the synaptic cleft.
Receptor activation: The neurotransmitter binds to post-synaptic receptors and produces a response in the post-synaptic neuron.
Signal termination: The signal must be terminated by some mechanism, normally by the elimination of excess neurotransmitters from the synaptic cleft.
Synthesis
Neurotransmitters are synthesized differently depending on which type they are. They can be a small molecule chemical, such as dopamine and serotonin, or they can be small neuropeptides, such as enkephalin.
Neuropeptides are synthesized in the cell body using the typical protein synthesis and translation pathways (rough endoplasmic reticulum and Golgi apparatus), then will be packaged into large, dense-core vesicles along with a protease. These vesicles are rapidly transported down the axon using microtubular proteins such as kinesin. When they arrive at the pre-synaptic terminal, they are ready to be released.
Small molecule neurotransmitters are synthesized in the cell body, where they are then transported down the axon in small, clear core vesicles. Upon arriving at the pre-synaptic terminal, enzymes will modify the small molecule neurotransmitter, and they can then be released from the vesicles into the cleft.
Release
Now that the neurotransmitters are stored in the vesicles in the pre-synaptic terminal, they must be released into the cleft. Along the membrane of the vesicle and the presynaptic membrane are proteins known as SNARE proteins; these proteins are essential in the binding of the vesicles to the membrane and the release of their contents. As the action potential propagates down the pre-synaptic neuron, the membrane will depolarize. Once the action potential arrives at the pre-synaptic terminal, the depolarization of the membrane will allow the voltage-dependent calcium channels to open, allowing the rapid influx of calcium into the pre-synaptic terminal. The influx of calcium causes the SNARE proteins to activate and change conformation, allowing the fusion of vesicles to the membrane and the release of their contents. The neurotransmitter will spill into the synaptic cleft, and the vesicle membrane is recovered via endocytosis.
Receptor Activation
Once the neurotransmitter binds to the post-synaptic neuron, it can generally cause one of 2 types of receptors to be activated. It will either activate a ligand-gated ion channel or a G-protein receptor.
Ligand-Gated Ion Channel: When the neurotransmitter binds to this receptor, there is a direct opening or closing of the attached ion channel. In other words, the neurotransmitter acts directly on the target ion channel. This type of receptor is described as “fast” because it generally only takes a few milliseconds to produce a response and is terminated very quickly. Depending on which neurotransmitter is binding to the receptor, these types of receptors can be excitatory or inhibitory.
G-Protein Coupled Receptors: These types of receptors are will produce a response (open or close an ion channel) by activating a signaling cascade involving secondary messengers. The most common secondary messengers are cyclic adenosine monophosphate (cAMP), inositol triphosphate (IP3), and diacylglycerol (DAG). When the neurotransmitter binds to the receptor, it activates the G-protein, which binds to guanosine triphosphate (GTP), and is activated. This will activate the secondary messenger cascade, which will eventually lead to the phosphorylation of ion channels. Due to multiple steps having to take place to generate the final response, this pathway is generally described as “slow,” and generally, the effects last longer (seconds to minutes).
Signal Termination
Inactivation of the signal must involve clearing the neurotransmitter from the synapse in at least 1 of 3 ways:
Re-uptake: Re-uptake can either be pre-synaptic or by glial cells. One important point to remember involving reuptake is that only small molecule chemical neurotransmitters can be taken back up, neuropeptides cannot participate in re-uptake; they must be eliminated by other means, such as degradation.
In pre-synaptic reuptake, the pre-synaptic neuron will use either endocytosis or specific transporters to remove the neurotransmitter from the synapse. The advantage of this mechanism is that the neurotransmitter can be recycled, which will prevent the neuron from having to re-synthesize the neurotransmitter every cycle of release.
In some cases, such as with glutamate, a glial cell will be involved in the re-uptake. Glutamate is toxic to the cell, so it is stored inside the neuron as glutamine. When glutamate is released into the synapse, it will be taken up by the glial cell using a specific transporter, converted into glutamine via glutaminase, then returned to the neuron to be recycled.
Enzymatic Destruction: The neurotransmitter can be destroyed directly either in the cleft or in the pre-synaptic terminal using certain enzymes. Two major enzymes are involved in the destruction of the neurotransmitter:
Monoamine Oxidases (MAO): These enzymes are responsible for oxidizing, and therefore inactivating, the monoamines. They do this by using oxygen to remove the amine group. These are split into MAO-A and MAO-B based on substrates. MAO-A is mostly responsible for breaking down serotonin, melatonin, norepinephrine, and epinephrine. Both forms break down dopamine, tyramine, and tryptamine equally. MAO-B also breaks down phenethylamine and benzylamine.
Catechol-O-Methyltransferase (COMT): Generally, COMT is responsible for degrading catecholamines, including dopamine, epinephrine, and norepinephrine, as well as most substances with a catechol structure.
It is important to note that both of the above enzymes are very frequent targets of therapeutic medications. By eliminating these enzymes, the neurotransmitter will remain in the synapse for longer, which can be beneficial in eliminating the symptoms of many disease processes.
Diffusion: In the simplest form of termination, the neurotransmitter can simply diffuse out of the synaptic cleft and away from the receptors and into nearby blood vessels. This will decrease the concentration of the neurotransmitter in the synapse, gradually reducing the effect the neurotransmitter has on the post-synaptic neuron. [5][6]
Go to:
Clinical Significance
The synapse is the fundamental functional unit of neuronal communication. Because of this, diseases that target the synapse can present with severe clinical consequences. A few examples are listed below:
Myasthenia Gravis
Myasthenia gravis is an auto-immune disease process that causes muscle weakness that usually presents in a descending fashion. It can cause ptosis, diminished facial expression, respiratory depression, and other signs/symptoms of weakness. In general, it is worse after activity and better with rest. The pathogenesis of myasthenia gravis involves diminished communication between the neuron and the muscle at the neuromuscular junction (NMJ). The reason for this is that antibodies will either block or destroy the acetylcholine receptors at the NMJ, preventing the ACh from binding and depolarizing the muscle, therefore, inhibiting contraction. These antibodies block step three (receptor activation) of the synaptic communication pathway.
Lambert-Eaton Syndrome
Lambert-Eaton syndrome is also an auto-immune condition producing dysfunction at the neuromuscular junction; however, it involves the pre-synaptic neuron. Instead of antibodies directed against the ACh receptors as in myasthenia gravis, the antibodies here are directed against the calcium channels on the pre-synaptic neuron. This prevents calcium influx from occurring, which prevents the fusion of vesicles with the pre-synaptic membrane and the release of the neurotransmitters into the synapse. These antibodies prevent step two (neurotransmitter release) of the synaptic communication pathway.
Botulism/Tetanus
In both of these disease processes, the causative agent is a toxin produced by a bacteria that acts as a protease that cleaves the SNARE proteins. This prevents the release of neurotransmitters at the junction by inhibiting vesicular fusion.
Botulism: The botulinum toxin, produced by Clostridium botulinum, prevents the release of acetylcholine, which is a stimulatory neurotransmitter. This inhibits stimulatory effects, which prevents muscle contraction and causes flaccid paralysis.
Tetanus: The tetanus toxin, produced by Clostridium tetani, prevents the release of GABA and glycine, both of which are inhibitory neurotransmitters. Specifically, their release is inhibited in the Renshaw cells in the spinal cord. This produces symptoms resembling an upper motor neuron lesion: spastic paralysis, lockjaw, and opisthotonus.
Go to:
Review Questions
Access free multiple choice questions on this topic.
Comment on this article.
Anatomy of Neurons
Figure
Anatomy of Neurons. A. Two connected neurons. Neurons have a soma that contains a nucleus, an axon, and a dendritic tree. A single synapse (red circle) is formed at the point where an axon's neuron (black) connects to another neuron's (more...)
Go to:
References
1.
Jones RA, Harrison C, Eaton SL, Llavero Hurtado M, Graham LC, Alkhammash L, Oladiran OA, Gale A, Lamont DJ, Simpson H, Simmen MW, Soeller C, Wishart TM, Gillingwater TH. Cellular and Molecular Anatomy of the Human Neuromuscular Junction. Cell Rep. 2017 Nov 28;21(9):2348-2356. [PMC free article] [PubMed]
2.
Napper RM, Harvey RJ. Number of parallel fiber synapses on an individual Purkinje cell in the cerebellum of the rat. J Comp Neurol. 1988 Aug 08;274(2):168-77. [PubMed]
3.
Harris AL. Electrical coupling and its channels. J Gen Physiol. 2018 Dec 03;150(12):1606-1639. [PMC free article] [PubMed]
4.
Südhof TC. Towards an Understanding of Synapse Formation. Neuron. 2018 Oct 24;100(2):276-293. [PMC free article] [PubMed]
5.
Südhof TC. The presynaptic active zone. Neuron. 2012 Jul 12;75(1):11-25. [PMC free article] [PubMed]
6.
Lisman JE, Raghavachari S, Tsien RW. The sequence of events that underlie quantal transmission at central glutamatergic synapses. Nat Rev Neurosci. 2007 Aug;8(8):597-609. [PubMed]
Disclosure: Michael Caire declares no relevant financial relationships with ineligible companies.
Disclosure: Vamsi Reddy declares no relevant financial relationships with ineligible companies.
Disclosure: Matthew Varacallo declares no relevant financial relationships with ineligible companies.
Copyright © 2024, StatPearls Publishing LLC.
This book is distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0) ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ), which permits others to distribute the work, provided that the article is not altered or used commercially. You are not required to obtain permission to distribute this article, provided that you credit the author and journal.
Bookshelf ID: NBK526047PMID: 30252303
Share on FacebookShare on Twitter
Views
PubReader
Print View
Cite this Page
In this Page
Introduction
Cellular Level
Development
Mechanism
Clinical Significance
Review Questions
References
Bulk Download
Bulk download StatPearls data from FTP
Related information
PMC
PubMed
Similar articles in PubMed
Review Molecules involved in the formation of synaptic connections in muscle and brain.
[Matrix Biol. 2001]
Delivery of different genes into presynaptic and postsynaptic neocortical neurons connected by a BDNF-TrkB synapse.
[Brain Res. 2019]
Review The versatile synapse.
[J Exp Biol. 1984]
Review Thematic Minireview Series: Molecular Mechanisms of Synaptic Plasticity.
[J Biol Chem. 2015]
Review In vivo measurements of neurotransmitters by microdialysis sampling.
[Anal Chem. 2006]
See reviews...
See all...
Recent Activity
ClearTurn Off
Physiology, Synapse - StatPearls
Types of Eye Movements and Their Functions - Neuroscience
Physiology, Baroreceptors - StatPearls
Anatomy, Head and Neck: Eye Orbicularis Oculi Muscle - StatPearls
See more...
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov | biology | 12496 | https://sv.wikipedia.org/wiki/Signalsubstans | Signalsubstans | En signalsubstans, även neurotransmittor och transmittorsubstans, är en molekyl som förmedlar en nervsignal på kemisk väg från en nervcell till en annan i nervsystemet. Neurotransmittorer existerar både i det centrala nervsystemet (CNS) och i det perifera nervsystemet (PNS). Neurotransmittorer lagras i vesiklar inom specialiserade ansvällningar på axonen som utgör den presynaptiska delen av synapsen, och som svar på en aktionspotential frisläpps de till den synaptiska klyftan. I mottagarcellen binder de då till specifika receptorer och två saker kan inträffa:
Jonkanaler öppnas och den postsynaptiska cellens membranpotential förändras.
En kemisk reaktion kan startas.
Vanliga neurotransmittorer i centrala nervsystemet är serotonin, GABA, dopamin, histamin, adrenalin, noradrenalin och acetylkolin. Acetylkolin är också en vanlig transmittor i motoriska nerver (perifera nervsystemet). Totalt finns det drygt femtio identifierade transmittorsubstanser som delas upp i tre huvudklasser.
Små transmittormolekyler
Dessa transmittorsubstanser är snabbverkande, fylls på snabbt och framställs från mat. Dessutom binder de ofta till jonkanaler (se synaps).
Exempel på små transmittormolekyler är:
Acetylkolin
Adrenalin
Anandamid
Dopamin
GABA
Glycin
Glutaminsyra (glutamat)
Histamin
IGF
Noradrenalin
Serotonin
Neuropeptider
Framställningen av neuropeptider styrs av DNA. Påfyllningen är långsam och de är för stora för att kunna utvinnas från mat. Dessutom binder de inte till jonkanaler.
Exempel på neuropeptider är oxytocin, insulin, endorfin och gastrin.
Transmittorgaser
Dessa transmittorsubstanser är ovanliga. De produceras i synapsen. De som har identifierats är kväveoxid (NO) och kolmonoxid (CO).
Se även
Hormon
Neurovetenskap
Källor
Externa länkar
Från nerv till piller
Neurologi
Biokemi
Cellbiologi | swedish | 0.34637 |
breathe_out_of_one_nostril/Respiratory_system.txt |
The respiratory system (also respiratory apparatus, ventilatory system) is a biological system consisting of specific organs and structures used for gas exchange in animals and plants. The anatomy and physiology that make this happen varies greatly, depending on the size of the organism, the environment in which it lives and its evolutionary history. In land animals, the respiratory surface is internalized as linings of the lungs. Gas exchange in the lungs occurs in millions of small air sacs; in mammals and reptiles, these are called alveoli, and in birds, they are known as atria. These microscopic air sacs have a very rich blood supply, thus bringing the air into close contact with the blood. These air sacs communicate with the external environment via a system of airways, or hollow tubes, of which the largest is the trachea, which branches in the middle of the chest into the two main bronchi. These enter the lungs where they branch into progressively narrower secondary and tertiary bronchi that branch into numerous smaller tubes, the bronchioles. In birds, the bronchioles are termed parabronchi. It is the bronchioles, or parabronchi that generally open into the microscopic alveoli in mammals and atria in birds. Air has to be pumped from the environment into the alveoli or atria by the process of breathing which involves the muscles of respiration.
In most fish, and a number of other aquatic animals (both vertebrates and invertebrates), the respiratory system consists of gills, which are either partially or completely external organs, bathed in the watery environment. This water flows over the gills by a variety of active or passive means. Gas exchange takes place in the gills which consist of thin or very flat filaments and lammellae which expose a very large surface area of highly vascularized tissue to the water.
Other animals, such as insects, have respiratory systems with very simple anatomical features, and in amphibians, even the skin plays a vital role in gas exchange. Plants also have respiratory systems but the directionality of gas exchange can be opposite to that in animals. The respiratory system in plants includes anatomical features such as stomata, that are found in various parts of the plant.
Mammals
Anatomy
Main articles: Lung and Respiratory tract
Fig. 1. Respiratory system
Fig. 2. The lower respiratory tract, or "Respiratory Tree"TracheaMainstem bronchusLobar bronchusSegmental bronchusBronchioleAlveolar ductAlveolus
In humans and other mammals, the anatomy of a typical respiratory system is the respiratory tract. The tract is divided into an upper and a lower respiratory tract. The upper tract includes the nose, nasal cavities, sinuses, pharynx and the part of the larynx above the vocal folds. The lower tract (Fig. 2.) includes the lower part of the larynx, the trachea, bronchi, bronchioles and the alveoli.
The branching airways of the lower tract are often described as the respiratory tree or tracheobronchial tree (Fig. 2). The intervals between successive branch points along the various branches of "tree" are often referred to as branching "generations", of which there are, in the adult human, about 23. The earlier generations (approximately generations 0–16), consisting of the trachea and the bronchi, as well as the larger bronchioles which simply act as air conduits, bringing air to the respiratory bronchioles, alveolar ducts and alveoli (approximately generations 17–23), where gas exchange takes place. Bronchioles are defined as the small airways lacking any cartilaginous support.
The first bronchi to branch from the trachea are the right and left main bronchi. Second, only in diameter to the trachea (1.8 cm), these bronchi (1 -1.4 cm in diameter) enter the lungs at each hilum, where they branch into narrower secondary bronchi known as lobar bronchi, and these branch into narrower tertiary bronchi known as segmental bronchi. Further divisions of the segmental bronchi (1 to 6 mm in diameter) are known as 4th order, 5th order, and 6th order segmental bronchi, or grouped together as subsegmental bronchi.
Compared to the 23 number (on average) of branchings of the respiratory tree in the adult human, the mouse has only about 13 such branchings.
The alveoli are the dead end terminals of the "tree", meaning that any air that enters them has to exit via the same route. A system such as this creates dead space, a volume of air (about 150 ml in the adult human) that fills the airways after exhalation and is breathed back into the alveoli before environmental air reaches them. At the end of inhalation, the airways are filled with environmental air, which is exhaled without coming in contact with the gas exchanger.
Ventilatory volumes
Fig. 3 Output of a 'spirometer'. Upward movement of the graph, read from the left, indicates the intake of air; downward movements represent exhalation.
Main articles: Breathing and Lung volumes
The lungs expand and contract during the breathing cycle, drawing air in and out of the lungs. The volume of air moved in or out of the lungs under normal resting circumstances (the resting tidal volume of about 500 ml), and volumes moved during maximally forced inhalation and maximally forced exhalation are measured in humans by spirometry. A typical adult human spirogram with the names given to the various excursions in volume the lungs can undergo is illustrated below (Fig. 3):
Not all the air in the lungs can be expelled during maximally forced exhalation(ERV). This is the residual volume(volume of air remaining even after a forced exhalation) of about 1.0-1.5 liters which cannot be measured by spirometry. Volumes that include the residual volume (i.e. functional residual capacity of about 2.5-3.0 liters, and total lung capacity of about 6 liters) can therefore also not be measured by spirometry. Their measurement requires special techniques.
The rates at which air is breathed in or out, either through the mouth or nose or into or out of the alveoli are tabulated below, together with how they are calculated. The number of breath cycles per minute is known as the respiratory rate. An average healthy human breathes 12-16 times a minute.
Measurement
Equation
Description
Minute ventilation
tidal volume * respiratory rate
the total volume of air entering, or leaving, the nose or mouth per minute or normal respiration.
Alveolar ventilation
(tidal volume – dead space) * respiratory rate
the volume of air entering or leaving the alveoli per minute.
Dead space ventilation
dead space * respiratory rate
the volume of air that does not reach the alveoli during inhalation, but instead remains in the airways, per minute.
Mechanics of breathing
Fig. 6 Real-time magnetic resonance imaging (MRI) of the chest movements of human thorax during breathing
Main article: Breathing § Mechanics
The "pump handle" and "bucket handle movements" of the ribsFig. 4 The effect of the muscles of inhalation in expanding the rib cage. The particular action illustrated here is called the pump handle movement of the rib cage.Fig. 5 In this view of the rib cage the downward slope of the lower ribs from the midline outwards can be clearly seen. This allows a movement similar to the "pump handle effect", but in this case, it is called the bucket handle movement. The color of the ribs refers to their classification, and is not relevant here.
BreathingFig. 7 The muscles of breathing at rest: inhalation on the left, exhalation on the right. Contracting muscles are shown in red; relaxed muscles in blue. Contraction of the diaphragm generally contributes the most to the expansion of the chest cavity (light blue). However, at the same time, the intercostal muscles pull the ribs upwards (their effect is indicated by arrows) also causing the rib cage to expand during inhalation (see diagram on other side of the page). The relaxation of all these muscles during exhalation causes the rib cage and abdomen (light green) to elastically return to their resting positions. Compare with Fig. 6, the MRI video of the chest movements during the breathing cycle.Fig. 8 The muscles of forceful breathing (inhalation and exhalation). The color code is the same as on the left. In addition to a more forceful and extensive contraction of the diaphragm, the intercostal muscles are aided by the accessory muscles of inhalation to exaggerate the movement of the ribs upwards, causing a greater expansion of the rib cage. During exhalation, apart from the relaxation of the muscles of inhalation, the abdominal muscles actively contract to pull the lower edges of the rib cage downwards decreasing the volume of the rib cage, while at the same time pushing the diaphragm upwards deep into the thorax.
In mammals, inhalation at rest is primarily due to the contraction of the diaphragm. This is an upwardly domed sheet of muscle that separates the thoracic cavity from the abdominal cavity. When it contracts, the sheet flattens, (i.e. moves downwards as shown in Fig. 7) increasing the volume of the thoracic cavity in the antero-posterior axis. The contracting diaphragm pushes the abdominal organs downwards. But because the pelvic floor prevents the lowermost abdominal organs from moving in that direction, the pliable abdominal contents cause the belly to bulge outwards to the front and sides, because the relaxed abdominal muscles do not resist this movement (Fig. 7). This entirely passive bulging (and shrinking during exhalation) of the abdomen during normal breathing is sometimes referred to as "abdominal breathing", although it is, in fact, "diaphragmatic breathing", which is not visible on the outside of the body. Mammals only use their abdominal muscles during forceful exhalation (see Fig. 8, and discussion below). Never during any form of inhalation.
As the diaphragm contracts, the rib cage is simultaneously enlarged by the ribs being pulled upwards by the intercostal muscles as shown in Fig. 4. All the ribs slant downwards from the rear to the front (as shown in Fig. 4); but the lowermost ribs also slant downwards from the midline outwards (Fig. 5). Thus the rib cage's transverse diameter can be increased in the same way as the antero-posterior diameter is increased by the so-called pump handle movement shown in Fig. 4.
The enlargement of the thoracic cavity's vertical dimension by the contraction of the diaphragm, and its two horizontal dimensions by the lifting of the front and sides of the ribs, causes the intrathoracic pressure to fall. The lungs' interiors are open to the outside air and being elastic, therefore expand to fill the increased space, pleura fluid between double-layered pleura covering of lungs helps in reducing friction while lungs expansion and contraction. The inflow of air into the lungs occurs via the respiratory airways (Fig. 2). In a healthy person, these airways begin with the nose. (It is possible to begin with the mouth, which is the backup breathing system. However, chronic mouth breathing leads to, or is a sign of, illness.) It ends in the microscopic dead-end sacs called alveoli, which are always open, though the diameters of the various sections can be changed by the sympathetic and parasympathetic nervous systems. The alveolar air pressure is therefore always close to atmospheric air pressure (about 100 kPa at sea level) at rest, with the pressure gradients because of lungs contraction and expansion cause air to move in and out of the lungs during breathing rarely exceeding 2–3 kPa.
During exhalation, the diaphragm and intercostal muscles relax. This returns the chest and abdomen to a position determined by their anatomical elasticity. This is the "resting mid-position" of the thorax and abdomen (Fig. 7) when the lungs contain their functional residual capacity of air (the light blue area in the right hand illustration of Fig. 7), which in the adult human has a volume of about 2.5–3.0 liters (Fig. 3). Resting exhalation lasts about twice as long as inhalation because the diaphragm relaxes passively more gently than it contracts actively during inhalation.
Fig. 9 The changes in the composition of the alveolar air during a normal breathing cycle at rest. The scale on the left, and the blue line, indicate the partial pressures of carbon dioxide in kPa, while that on the right and the red line, indicate the partial pressures of oxygen, also in kPa (to convert kPa into mm Hg, multiply by 7.5).
The volume of air that moves in or out (at the nose or mouth) during a single breathing cycle is called the tidal volume. In a resting adult human, it is about 500 ml per breath. At the end of exhalation, the airways contain about 150 ml of alveolar air which is the first air that is breathed back into the alveoli during inhalation. This volume air that is breathed out of the alveoli and back in again is known as dead space ventilation, which has the consequence that of the 500 ml breathed into the alveoli with each breath only 350 ml (500 ml - 150 ml = 350 ml) is fresh warm and moistened air. Since this 350 ml of fresh air is thoroughly mixed and diluted by the air that remains in the alveoli after a normal exhalation (i.e. the functional residual capacity of about 2.5–3.0 liters), it is clear that the composition of the alveolar air changes very little during the breathing cycle (see Fig. 9). The oxygen tension (or partial pressure) remains close to 13-14 kPa (about 100 mm Hg), and that of carbon dioxide very close to 5.3 kPa (or 40 mm Hg). This contrasts with composition of the dry outside air at sea level, where the partial pressure of oxygen is 21 kPa (or 160 mm Hg) and that of carbon dioxide 0.04 kPa (or 0.3 mmHg).
During heavy breathing (hyperpnea), as, for instance, during exercise, inhalation is brought about by a more powerful and greater excursion of the contracting diaphragm than at rest (Fig. 8). In addition, the "accessory muscles of inhalation" exaggerate the actions of the intercostal muscles (Fig. 8). These accessory muscles of inhalation are muscles that extend from the cervical vertebrae and base of the skull to the upper ribs and sternum, sometimes through an intermediary attachment to the clavicles. When they contract, the rib cage's internal volume is increased to a far greater extent than can be achieved by contraction of the intercostal muscles alone. Seen from outside the body, the lifting of the clavicles during strenuous or labored inhalation is sometimes called clavicular breathing, seen especially during asthma attacks and in people with chronic obstructive pulmonary disease.
During heavy breathing, exhalation is caused by relaxation of all the muscles of inhalation. But now, the abdominal muscles, instead of remaining relaxed (as they do at rest), contract forcibly pulling the lower edges of the rib cage downwards (front and sides) (Fig. 8). This not only drastically decreases the size of the rib cage, but also pushes the abdominal organs upwards against the diaphragm which consequently bulges deeply into the thorax (Fig. 8). The end-exhalatory lung volume is now well below the resting mid-position and contains far less air than the resting "functional residual capacity". However, in a normal mammal, the lungs cannot be emptied completely. In an adult human, there is always still at least 1 liter of residual air left in the lungs after maximum exhalation.
The automatic rhythmical breathing in and out, can be interrupted by coughing, sneezing (forms of very forceful exhalation), by the expression of a wide range of emotions (laughing, sighing, crying out in pain, exasperated intakes of breath) and by such voluntary acts as speech, singing, whistling and the playing of wind instruments. All of these actions rely on the muscles described above, and their effects on the movement of air in and out of the lungs.
Although not a form of breathing, the Valsalva maneuver involves the respiratory muscles. It is, in fact, a very forceful exhalatory effort against a tightly closed glottis, so that no air can escape from the lungs. Instead, abdominal contents are evacuated in the opposite direction, through orifices in the pelvic floor. The abdominal muscles contract very powerfully, causing the pressure inside the abdomen and thorax to rise to extremely high levels. The Valsalva maneuver can be carried out voluntarily but is more generally a reflex elicited when attempting to empty the abdomen during, for instance, difficult defecation, or during childbirth. Breathing ceases during this maneuver.
Gas exchange
Main article: Gas exchange
Mechanism of gas exchangeFig. 11 A highly diagrammatic illustration of the process of gas exchange in the mammalian lungs, emphasizing the differences between the gas compositions of the ambient air, the alveolar air (light blue) with which the pulmonary capillary blood equilibrates, and the blood gas tensions in the pulmonary arterial (blue blood entering the lung on the left) and venous blood (red blood leaving the lung on the right). All the gas tensions are in kPa. To convert to mm Hg, multiply by 7.5.Fig. 12 A diagrammatic histological cross-section through a portion of lung tissue showing a normally inflated alveolus (at the end of a normal exhalation), and its walls containing the pulmonary capillaries (shown in cross-section). This illustrates how the pulmonary capillary blood is completely surrounded by alveolar air. In a normal human lung, all the alveoli together contain about 3 liters of alveolar air. All the pulmonary capillaries contain about 100 ml of blood.
Fig. 10 A histological cross-section through an alveolar wall showing the layers through which the gases have to move between the blood plasma and the alveolar air. The dark blue objects are the nuclei of the capillary endothelial and alveolar type I epithelial cells (or type 1 pneumocytes). The two red objects labeled "RBC" are red blood cells in the pulmonary capillary blood.
The primary purpose of the respiratory system is the equalizing of the partial pressures of the respiratory gases in the alveolar air with those in the pulmonary capillary blood (Fig. 11). This process occurs by simple diffusion, across a very thin membrane (known as the blood–air barrier), which forms the walls of the pulmonary alveoli (Fig. 10). It consists of the alveolar epithelial cells, their basement membranes and the endothelial cells of the alveolar capillaries (Fig. 10). This blood gas barrier is extremely thin (in humans, on average, 2.2 μm thick). It is folded into about 300 million small air sacs called alveoli (each between 75 and 300 µm in diameter) branching off from the respiratory bronchioles in the lungs, thus providing an extremely large surface area (approximately 145 m) for gas exchange to occur.
The air contained within the alveoli has a semi-permanent volume of about 2.5-3.0 liters which completely surrounds the alveolar capillary blood (Fig. 12). This ensures that equilibration of the partial pressures of the gases in the two compartments is very efficient and occurs very quickly. The blood leaving the alveolar capillaries and is eventually distributed throughout the body therefore has a partial pressure of oxygen of 13-14 kPa (100 mmHg), and a partial pressure of carbon dioxide of 5.3 kPa (40 mmHg) (i.e. the same as the oxygen and carbon dioxide gas tensions as in the alveoli). As mentioned in the section above, the corresponding partial pressures of oxygen and carbon dioxide in the ambient (dry) air at sea level are 21 kPa (160 mmHg) and 0.04 kPa (0.3 mmHg) respectively.
This marked difference between the composition of the alveolar air and that of the ambient air can be maintained because the functional residual capacity is contained in dead-end sacs connected to the outside air by fairly narrow and relatively long tubes (the airways: nose, pharynx, larynx, trachea, bronchi and their branches down to the bronchioles), through which the air has to be breathed both in and out (i.e. there is no unidirectional through-flow as there is in the bird lung). This typical mammalian anatomy combined with the fact that the lungs are not emptied and re-inflated with each breath (leaving a substantial volume of air, of about 2.5-3.0 liters, in the alveoli after exhalation), ensures that the composition of the alveolar air is only minimally disturbed when the 350 ml of fresh air is mixed into it with each inhalation. Thus the animal is provided with a very special "portable atmosphere", whose composition differs significantly from the present-day ambient air. It is this portable atmosphere (the functional residual capacity) to which the blood and therefore the body tissues are exposed – not to the outside air.
The resulting arterial partial pressures of oxygen and carbon dioxide are homeostatically controlled. A rise in the arterial partial pressure of CO2 and, to a lesser extent, a fall in the arterial partial pressure of O2, will reflexly cause deeper and faster breathing until the blood gas tensions in the lungs, and therefore the arterial blood, return to normal. The converse happens when the carbon dioxide tension falls, or, again to a lesser extent, the oxygen tension rises: the rate and depth of breathing are reduced until blood gas normality is restored.
Since the blood arriving in the alveolar capillaries has a partial pressure of O2 of, on average, 6 kPa (45 mmHg), while the pressure in the alveolar air is 13-14 kPa (100 mmHg), there will be a net diffusion of oxygen into the capillary blood, changing the composition of the 3 liters of alveolar air slightly. Similarly, since the blood arriving in the alveolar capillaries has a partial pressure of CO2 of also about 6 kPa (45 mmHg), whereas that of the alveolar air is 5.3 kPa (40 mmHg), there is a net movement of carbon dioxide out of the capillaries into the alveoli. The changes brought about by these net flows of individual gases into and out of the alveolar air necessitate the replacement of about 15% of the alveolar air with ambient air every 5 seconds or so. This is very tightly controlled by the monitoring of the arterial blood gases (which accurately reflect composition of the alveolar air) by the aortic and carotid bodies, as well as by the blood gas and pH sensor on the anterior surface of the medulla oblongata in the brain. There are also oxygen and carbon dioxide sensors in the lungs, but they primarily determine the diameters of the bronchioles and pulmonary capillaries, and are therefore responsible for directing the flow of air and blood to different parts of the lungs.
It is only as a result of accurately maintaining the composition of the 3 liters of alveolar air that with each breath some carbon dioxide is discharged into the atmosphere and some oxygen is taken up from the outside air. If more carbon dioxide than usual has been lost by a short period of hyperventilation, respiration will be slowed down or halted until the alveolar partial pressure of carbon dioxide has returned to 5.3 kPa (40 mmHg). It is therefore strictly speaking untrue that the primary function of the respiratory system is to rid the body of carbon dioxide “waste”. The carbon dioxide that is breathed out with each breath could probably be more correctly be seen as a byproduct of the body's extracellular fluid carbon dioxide and pH homeostats
If these homeostats are compromised, then a respiratory acidosis, or a respiratory alkalosis will occur. In the long run these can be compensated by renal adjustments to the H and HCO3 concentrations in the plasma; but since this takes time, the hyperventilation syndrome can, for instance, occur when agitation or anxiety cause a person to breathe fast and deeply thus causing a distressing respiratory alkalosis through the blowing off of too much CO2 from the blood into the outside air.
Oxygen has a very low solubility in water, and is therefore carried in the blood loosely combined with hemoglobin. The oxygen is held on the hemoglobin by four ferrous iron-containing heme groups per hemoglobin molecule. When all the heme groups carry one O2 molecule each the blood is said to be “saturated” with oxygen, and no further increase in the partial pressure of oxygen will meaningfully increase the oxygen concentration of the blood. Most of the carbon dioxide in the blood is carried as bicarbonate ions (HCO3) in the plasma. However the conversion of dissolved CO2 into HCO3 (through the addition of water) is too slow for the rate at which the blood circulates through the tissues on the one hand, and through alveolar capillaries on the other. The reaction is therefore catalyzed by carbonic anhydrase, an enzyme inside the red blood cells. The reaction can go in both directions depending on the prevailing partial pressure of CO2. A small amount of carbon dioxide is carried on the protein portion of the hemoglobin molecules as carbamino groups. The total concentration of carbon dioxide (in the form of bicarbonate ions, dissolved CO2, and carbamino groups) in arterial blood (i.e. after it has equilibrated with the alveolar air) is about 26 mM (or 58 ml/100 ml), compared to the concentration of oxygen in saturated arterial blood of about 9 mM (or 20 ml/100 ml blood).
Control of ventilation
Main article: Control of ventilation
Ventilation of the lungs in mammals occurs via the respiratory centers in the medulla oblongata and the pons of the brainstem. These areas form a series of neural pathways which receive information about the partial pressures of oxygen and carbon dioxide in the arterial blood. This information determines the average rate of ventilation of the alveoli of the lungs, to keep these pressures constant. The respiratory center does so via motor nerves which activate the diaphragm and other muscles of respiration.
The breathing rate increases when the partial pressure of carbon dioxide in the blood increases. This is detected by central blood gas chemoreceptors on the anterior surface of the medulla oblongata. The aortic and carotid bodies, are the peripheral blood gas chemoreceptors which are particularly sensitive to the arterial partial pressure of O2 though they also respond, but less strongly, to the partial pressure of CO2. At sea level, under normal circumstances, the breathing rate and depth, is determined primarily by the arterial partial pressure of carbon dioxide rather than by the arterial partial pressure of oxygen, which is allowed to vary within a fairly wide range before the respiratory centers in the medulla oblongata and pons respond to it to change the rate and depth of breathing.
Exercise increases the breathing rate due to the extra carbon dioxide produced by the enhanced metabolism of the exercising muscles. In addition, passive movements of the limbs also reflexively produce an increase in the breathing rate.
Information received from stretch receptors in the lungs' limits tidal volume (the depth of inhalation and exhalation).
Responses to low atmospheric pressures
The alveoli are open (via the airways) to the atmosphere, with the result that alveolar air pressure is exactly the same as the ambient air pressure at sea level, at altitude, or in any artificial atmosphere (e.g. a diving chamber, or decompression chamber) in which the individual is breathing freely. With expansion of the lungs the alveolar air occupies a larger volume, and its pressure falls proportionally, causing air to flow in through the airways, until the pressure in the alveoli is again at the ambient air pressure. The reverse happens during exhalation. This process (of inhalation and exhalation) is exactly the same at sea level, as on top of Mt. Everest, or in a diving chamber or decompression chamber.
Fig. 14 A graph showing the relationship between total atmospheric pressure and altitude above sea level.
However, as one rises above sea level the density of the air decreases exponentially (see Fig. 14), halving approximately with every 5500 m rise in altitude. Since the composition of the atmospheric air is almost constant below 80 km, as a result of the continuous mixing effect of the weather, the concentration of oxygen in the air (mmols O2 per liter of ambient air) decreases at the same rate as the fall in air pressure with altitude. Therefore, in order to breathe in the same amount of oxygen per minute, the person has to inhale a proportionately greater volume of air per minute at altitude than at sea level. This is achieved by breathing deeper and faster (i.e. hyperpnea) than at sea level (see below).
Fig. 13 Aerial photo of Mount Everest from the south, behind Nuptse and Lhotse.
There is, however, a complication that increases the volume of air that needs to be inhaled per minute (respiratory minute volume) to provide the same amount of oxygen to the lungs at altitude as at sea level. During inhalation, the air is warmed and saturated with water vapor during its passage through the nose passages and pharynx. Saturated water vapor pressure is dependent only on temperature. At a body core temperature of 37 °C it is 6.3 kPa (47.0 mmHg), irrespective of any other influences, including altitude. Thus at sea level, where the ambient atmospheric pressure is about 100 kPa, the moistened air that flows into the lungs from the trachea consists of water vapor (6.3 kPa), nitrogen (74.0 kPa), oxygen (19.7 kPa) and trace amounts of carbon dioxide and other gases (a total of 100 kPa). In dry air the partial pressure of O2 at sea level is 21.0 kPa (i.e. 21% of 100 kPa), compared to the 19.7 kPa of oxygen entering the alveolar air. (The tracheal partial pressure of oxygen is 21% of [100 kPa – 6.3 kPa] = 19.7 kPa). At the summit of Mt. Everest (at an altitude of 8,848 m or 29,029 ft), the total atmospheric pressure is 33.7 kPa, of which 7.1 kPa (or 21%) is oxygen. The air entering the lungs also has a total pressure of 33.7 kPa, of which 6.3 kPa is, unavoidably, water vapor (as it is at sea level). This reduces the partial pressure of oxygen entering the alveoli to 5.8 kPa (or 21% of [33.7 kPa – 6.3 kPa] = 5.8 kPa). The reduction in the partial pressure of oxygen in the inhaled air is therefore substantially greater than the reduction of the total atmospheric pressure at altitude would suggest (on Mt Everest: 5.8 kPa vs. 7.1 kPa).
A further minor complication exists at altitude. If the volume of the lungs were to be instantaneously doubled at the beginning of inhalation, the air pressure inside the lungs would be halved. This happens regardless of altitude. Thus, halving of the sea level air pressure (100 kPa) results in an intrapulmonary air pressure of 50 kPa. Doing the same at 5500 m, where the atmospheric pressure is only 50 kPa, the intrapulmonary air pressure falls to 25 kPa. Therefore, the same change in lung volume at sea level results in a 50 kPa difference in pressure between the ambient air and the intrapulmonary air, whereas it result in a difference of only 25 kPa at 5500 m. The driving pressure forcing air into the lungs during inhalation is therefore halved at this altitude. The rate of inflow of air into the lungs during inhalation at sea level is therefore twice that which occurs at 5500 m. However, in reality, inhalation and exhalation occur far more gently and less abruptly than in the example given. The differences between the atmospheric and intrapulmonary pressures, driving air in and out of the lungs during the breathing cycle, are in the region of only 2–3 kPa. A doubling or more of these small pressure differences could be achieved only by very major changes in the breathing effort at high altitudes.
All of the above influences of low atmospheric pressures on breathing are accommodated primarily by breathing deeper and faster (hyperpnea). The exact degree of hyperpnea is determined by the blood gas homeostat, which regulates the partial pressures of oxygen and carbon dioxide in the arterial blood. This homeostat prioritizes the regulation of the arterial partial pressure of carbon dioxide over that of oxygen at sea level. That is to say, at sea level the arterial partial pressure of CO2 is maintained at very close to 5.3 kPa (or 40 mmHg) under a wide range of circumstances, at the expense of the arterial partial pressure of O2, which is allowed to vary within a very wide range of values, before eliciting a corrective ventilatory response. However, when the atmospheric pressure (and therefore the partial pressure of O2 in the ambient air) falls to below 50-75% of its value at sea level, oxygen homeostasis is given priority over carbon dioxide homeostasis. This switch-over occurs at an elevation of about 2500 m (or about 8000 ft). If this switch occurs relatively abruptly, the hyperpnea at high altitude will cause a severe fall in the arterial partial pressure of carbon dioxide, with a consequent rise in the pH of the arterial plasma. This is one contributor to high altitude sickness. On the other hand, if the switch to oxygen homeostasis is incomplete, then hypoxia may complicate the clinical picture with potentially fatal results.
There are oxygen sensors in the smaller bronchi and bronchioles. In response to low partial pressures of oxygen in the inhaled air these sensors reflexively cause the pulmonary arterioles to constrict. (This is the exact opposite of the corresponding reflex in the tissues, where low arterial partial pressures of O2 cause arteriolar vasodilation.) At altitude this causes the pulmonary arterial pressure to rise resulting in a much more even distribution of blood flow to the lungs than occurs at sea level. At sea level, the pulmonary arterial pressure is very low, with the result that the tops of the lungs receive far less blood than the bases, which are relatively over-perfused with blood. It is only in the middle of the lungs that the blood and air flow to the alveoli are ideally matched. At altitude, this variation in the ventilation/perfusion ratio of alveoli from the tops of the lungs to the bottoms is eliminated, with all the alveoli perfused and ventilated in more or less the physiologically ideal manner. This is a further important contributor to the acclimatatization to high altitudes and low oxygen pressures.
The kidneys measure the oxygen content (mmol O2/liter blood, rather than the partial pressure of O2) of the arterial blood. When the oxygen content of the blood is chronically low, as at high altitude, the oxygen-sensitive kidney cells secrete erythropoietin (EPO) into the blood. This hormone stimulates the red bone marrow to increase its rate of red cell production, which leads to an increase in the hematocrit of the blood, and a consequent increase in its oxygen carrying capacity (due to the now high hemoglobin content of the blood). In other words, at the same arterial partial pressure of O2, a person with a high hematocrit carries more oxygen per liter of blood than a person with a lower hematocrit does. High altitude dwellers therefore have higher hematocrits than sea-level residents.
Other functions of the lungs
Local defenses
Irritation of nerve endings within the nasal passages or airways, can induce a cough reflex and sneezing. These responses cause air to be expelled forcefully from the trachea or nose, respectively. In this manner, irritants caught in the mucus which lines the respiratory tract are expelled or moved to the mouth where they can be swallowed. During coughing, contraction of the smooth muscle in the airway walls narrows the trachea by pulling the ends of the cartilage plates together and by pushing soft tissue into the lumen. This increases the expired airflow rate to dislodge and remove any irritant particle or mucus.
Respiratory epithelium can secrete a variety of molecules that aid in the defense of the lungs. These include secretory immunoglobulins (IgA), collectins, defensins and other peptides and proteases, reactive oxygen species, and reactive nitrogen species. These secretions can act directly as antimicrobials to help keep the airway free of infection. A variety of chemokines and cytokines are also secreted that recruit the traditional immune cells and others to the site of infections.
Surfactant immune function is primarily attributed to two proteins: SP-A and SP-D. These proteins can bind to sugars on the surface of pathogens and thereby opsonize them for uptake by phagocytes. It also regulates inflammatory responses and interacts with the adaptive immune response. Surfactant degradation or inactivation may contribute to enhanced susceptibility to lung inflammation and infection.
Most of the respiratory system is lined with mucous membranes that contain mucosa-associated lymphoid tissue, which produces white blood cells such as lymphocytes.
Prevention of alveolar collapse
Main article: Pulmonary surfactant
The lungs make a surfactant, a surface-active lipoprotein complex (phospholipoprotein) formed by type II alveolar cells. It floats on the surface of the thin watery layer which lines the insides of the alveoli, reducing the water's surface tension.
The surface tension of a watery surface (the water-air interface) tends to make that surface shrink. When that surface is curved as it is in the alveoli of the lungs, the shrinkage of the surface decreases the diameter of the alveoli. The more acute the curvature of the water-air interface the greater the tendency for the alveolus to collapse. This has three effects. Firstly, the surface tension inside the alveoli resists expansion of the alveoli during inhalation (i.e. it makes the lung stiff, or non-compliant). Surfactant reduces the surface tension and therefore makes the lungs more compliant, or less stiff, than if it were not there. Secondly, the diameters of the alveoli increase and decrease during the breathing cycle. This means that the alveoli have a greater tendency to collapse (i.e. cause atelectasis) at the end of exhalation than at the end of inhalation. Since surfactant floats on the watery surface, its molecules are more tightly packed together when the alveoli shrink during exhalation. This causes them to have a greater surface tension-lowering effect when the alveoli are small than when they are large (as at the end of inhalation, when the surfactant molecules are more widely spaced). The tendency for the alveoli to collapse is therefore almost the same at the end of exhalation as at the end of inhalation. Thirdly, the surface tension of the curved watery layer lining the alveoli tends to draw water from the lung tissues into the alveoli. Surfactant reduces this danger to negligible levels, and keeps the alveoli dry.
Pre-term babies who are unable to manufacture surfactant have lungs that tend to collapse each time they breathe out. Unless treated, this condition, called respiratory distress syndrome, is fatal. Basic scientific experiments, carried out using cells from chicken lungs, support the potential for using steroids as a means of furthering the development of type II alveolar cells. In fact, once a premature birth is threatened, every effort is made to delay the birth, and a series of steroid injections is frequently administered to the mother during this delay in an effort to promote lung maturation.
Contributions to whole body functions
The lung vessels contain a fibrinolytic system that dissolves clots that may have arrived in the pulmonary circulation by embolism, often from the deep veins in the legs. They also release a variety of substances that enter the systemic arterial blood, and they remove other substances from the systemic venous blood that reach them via the pulmonary artery. Some prostaglandins are removed from the circulation, while others are synthesized in the lungs and released into the blood when lung tissue is stretched.
The lungs activate one hormone. The physiologically inactive decapeptide angiotensin I is converted to the aldosterone-releasing octapeptide, angiotensin II, in the pulmonary circulation. The reaction occurs in other tissues as well, but it is particularly prominent in the lungs. Angiotensin II also has a direct effect on arteriolar walls, causing arteriolar vasoconstriction, and consequently a rise in arterial blood pressure. Large amounts of the angiotensin-converting enzyme responsible for this activation are located on the surfaces of the endothelial cells of the alveolar capillaries. The converting enzyme also inactivates bradykinin. Circulation time through the alveolar capillaries is less than one second, yet 70% of the angiotensin I reaching the lungs is converted to angiotensin II in a single trip through the capillaries. Four other peptidases have been identified on the surface of the pulmonary endothelial cells.
Vocalization
The movement of gas through the larynx, pharynx and mouth allows humans to speak, or phonate. Vocalization, or singing, in birds occurs via the syrinx, an organ located at the base of the trachea. The vibration of air flowing across the larynx (vocal cords), in humans, and the syrinx, in birds, results in sound. Because of this, gas movement is vital for communication purposes.
Temperature control
Panting in dogs, cats, birds and some other animals provides a means of reducing body temperature, by evaporating saliva in the mouth (instead of evaporating sweat on the skin).
Clinical significance
Disorders of the respiratory system can be classified into several general groups:
Airway obstructive conditions (e.g., emphysema, bronchitis, asthma)
Pulmonary restrictive conditions (e.g., fibrosis, sarcoidosis, alveolar damage, pleural effusion)
Vascular diseases (e.g., pulmonary edema, pulmonary embolism, pulmonary hypertension)
Infectious, environmental and other "diseases" (e.g., pneumonia, tuberculosis, asbestosis, particulate pollutants)
Primary cancers (e.g. bronchial carcinoma, mesothelioma)
Secondary cancers (e.g. cancers that originated elsewhere in the body, but have seeded themselves in the lungs)
Insufficient surfactant (e.g. respiratory distress syndrome in pre-term babies) .
Disorders of the respiratory system are usually treated by a pulmonologist and respiratory therapist.
Where there is an inability to breathe or insufficiency in breathing, a medical ventilator may be used.
Exceptional mammals
Cetaceans
This section is an excerpt from Cetacea § Respiration.[edit]
Cetaceans have lungs, meaning they breathe air. An individual can last without a breath from a few minutes to over two hours depending on the species. Cetacea are deliberate breathers who must be awake to inhale and exhale. When stale air, warmed from the lungs, is exhaled, it condenses as it meets colder external air. As with a terrestrial mammal breathing out on a cold day, a small cloud of 'steam' appears. This is called the 'spout' and varies across species in shape, angle and height. Species can be identified at a distance using this characteristic.
The structure of the respiratory and circulatory systems is of particular importance for the life of marine mammals. The oxygen balance is effective. Each breath can replace up to 90% of the total lung volume. For land mammals, in comparison, this value is usually about 15%. During inhalation, about twice as much oxygen is absorbed by the lung tissue as in a land mammal. As with all mammals, the oxygen is stored in the blood and the lungs, but in cetaceans, it is also stored in various tissues, mainly in the muscles. The muscle pigment, myoglobin, provides an effective bond. This additional oxygen storage is vital for deep diving, since beyond a depth around 100 m (330 ft), the lung tissue is almost completely compressed by the water pressure.
Horses
Main article: Respiratory system of the horse
Horses are obligate nasal breathers which means that they are different from many other mammals because they do not have the option of breathing through their mouths and must take in air through their noses. A flap of tissue called the soft palate blocks off the pharynx from the mouth (oral cavity) of the horse, except when swallowing. This helps to prevent the horse from inhaling food, but does not allow use of the mouth to breathe when in respiratory distress, a horse can only breathe through its nostrils.
Elephants
The elephant is the only mammal known to have no pleural space. Instead, the parietal and visceral pleura are both composed of dense connective tissue and joined to each other via loose connective tissue. This lack of a pleural space, along with an unusually thick diaphragm, are thought to be evolutionary adaptations allowing the elephant to remain underwater for long periods while breathing through its trunk which emerges as a snorkel.
In the elephant the lungs are attached to the diaphragm and breathing relies mainly on the diaphragm rather than the expansion of the ribcage.
Birds
See also: Bird anatomy § Respiratory system
Fig. 15 The arrangement of the air sacs, and lungs in birds
Fig. 16 The anatomy of bird's respiratory system, showing the relationships of the trachea, primary and intra-pulmonary bronchi, the dorso- and ventro-bronchi, with the parabronchi running between the two. The posterior and anterior air sacs are also indicated, but not to scale.
Fig. 17 A dove skeleton, showing the movement of the chest during inhalation. Arrow 1 indicates the movement of the vertebral ribs. Arrow 2 shows the consequent movement of the sternum (and its keel). The two movements increase the vertical and transverse diameters of the chest portion of the trunk of the bird. Key: 1. skull; 2. cervical vertebrae; 3. furcula; 4. coracoid; 5. vertebral ribs; 6. sternum and its keel; 7. patella; 8. tarsus; 9. digits; 10. tibia (tibiotarsus); 11. fibula (tibiotarsus); 12. femur; 13. ischium (innominate); 14. pubis (innominate); 15. ilium (innominate); 16. caudal vertebrae; 17. pygostyle; 18. synsacrum; 19. scapula; 20. dorsal vertebrae; 21. humerus; 22. ulna; 23. radius; 24. carpus (carpometacarpus); 25. metacarpus (carpometacarpus); 26. digits; 27. alula
The respiratory system of birds differs significantly from that found in mammals. Firstly, they have rigid lungs which do not expand and contract during the breathing cycle. Instead an extensive system of air sacs (Fig. 15) distributed throughout their bodies act as the bellows drawing environmental air into the sacs, and expelling the spent air after it has passed through the lungs (Fig. 18). Birds also do not have diaphragms or pleural cavities.
Bird lungs are smaller than those in mammals of comparable size, but the air sacs account for 15% of the total body volume, compared to the 7% devoted to the alveoli which act as the bellows in mammals.
Inhalation and exhalation are brought about by alternately increasing and decreasing the volume of the entire thoraco-abdominal cavity (or coelom) using both their abdominal and costal muscles. During inhalation the muscles attached to the vertebral ribs (Fig. 17) contract angling them forwards and outwards. This pushes the sternal ribs, to which they are attached at almost right angles, downwards and forwards, taking the sternum (with its prominent keel) in the same direction (Fig. 17). This increases both the vertical and transverse diameters of thoracic portion of the trunk. The forward and downward movement of, particularly, the posterior end of the sternum pulls the abdominal wall downwards, increasing the volume of that region of the trunk as well. The increase in volume of the entire trunk cavity reduces the air pressure in all the thoraco-abdominal air sacs, causing them to fill with air as described below.
During exhalation the external oblique muscle which is attached to the sternum and vertebral ribs anteriorly, and to the pelvis (pubis and ilium in Fig. 17) posteriorly (forming part of the abdominal wall) reverses the inhalatory movement, while compressing the abdominal contents, thus increasing the pressure in all the air sacs. Air is therefore expelled from the respiratory system in the act of exhalation.
Fig. 19 The cross-current respiratory gas exchanger in the lungs of birds. Air is forced from the air sacs unidirectionally (from right to left in the diagram) through the parabronchi. The pulmonary capillaries surround the parabronchi in the manner shown (blood flowing from below the parabronchus to above it in the diagram). Blood or air with a high oxygen content is shown in red; oxygen-poor air or blood is shown in various shades of purple-blue.
During inhalation air enters the trachea via the nostrils and mouth, and continues to just beyond the syrinx at which point the trachea branches into two primary bronchi, going to the two lungs (Fig. 16). The primary bronchi enter the lungs to become the intrapulmonary bronchi, which give off a set of parallel branches called ventrobronchi and, a little further on, an equivalent set of dorsobronchi (Fig. 16). The ends of the intrapulmonary bronchi discharge air into the posterior air sacs at the caudal end of the bird. Each pair of dorso-ventrobronchi is connected by a large number of parallel microscopic air capillaries (or parabronchi) where gas exchange occurs (Fig. 16). As the bird inhales, tracheal air flows through the intrapulmonary bronchi into the posterior air sacs, as well as into the dorsobronchi, but not into the ventrobronchi (Fig. 18). This is due to the bronchial architecture which directs the inhaled air away from the openings of the ventrobronchi, into the continuation of the intrapulmonary bronchus towards the dorsobronchi and posterior air sacs. From the dorsobronchi the inhaled air flows through the parabronchi (and therefore the gas exchanger) to the ventrobronchi from where the air can only escape into the expanding anterior air sacs. So, during inhalation, both the posterior and anterior air sacs expand, the posterior air sacs filling with fresh inhaled air, while the anterior air sacs fill with "spent" (oxygen-poor) air that has just passed through the lungs.
Fig. 18 Inhalation-exhalation cycle in birds.
During exhalation the pressure in the posterior air sacs (which were filled with fresh air during inhalation) increases due to the contraction of the oblique muscle described above. The aerodynamics of the interconnecting openings from the posterior air sacs to the dorsobronchi and intrapulmonary bronchi ensures that the air leaves these sacs in the direction of the lungs (via the dorsobronchi), rather than returning down the intrapulmonary bronchi (Fig. 18). From the dorsobronchi the fresh air from the posterior air sacs flows through the parabronchi (in the same direction as occurred during inhalation) into ventrobronchi. The air passages connecting the ventrobronchi and anterior air sacs to the intrapulmonary bronchi direct the "spent", oxygen poor air from these two organs to the trachea from where it escapes to the exterior. Oxygenated air therefore flows constantly (during the entire breathing cycle) in a single direction through the parabronchi.
The blood flow through the bird lung is at right angles to the flow of air through the parabronchi, forming a cross-current flow exchange system (Fig. 19). The partial pressure of oxygen in the parabronchi declines along their lengths as O2 diffuses into the blood. The blood capillaries leaving the exchanger near the entrance of airflow take up more O2 than do the capillaries leaving near the exit end of the parabronchi. When the contents of all capillaries mix, the final partial pressure of oxygen of the mixed pulmonary venous blood is higher than that of the exhaled air, but is nevertheless less than half that of the inhaled air, thus achieving roughly the same systemic arterial blood partial pressure of oxygen as mammals do with their bellows-type lungs.
The trachea is an area of dead space: the oxygen-poor air it contains at the end of exhalation is the first air to re-enter the posterior air sacs and lungs. In comparison to the mammalian respiratory tract, the dead space volume in a bird is, on average, 4.5 times greater than it is in mammals of the same size. Birds with long necks will inevitably have long tracheae, and must therefore take deeper breaths than mammals do to make allowances for their greater dead space volumes. In some birds (e.g. the whooper swan, Cygnus cygnus, the white spoonbill, Platalea leucorodia, the whooping crane, Grus americana, and the helmeted curassow, Pauxi pauxi) the trachea, which some cranes can be 1.5 m long, is coiled back and forth within the body, drastically increasing the dead space ventilation. The purpose of this extraordinary feature is unknown.
Reptiles
Main article: Reptile § Respiratory system
Fig. 20 X-ray video of a female American alligator while breathing.
The anatomical structure of the lungs is less complex in reptiles than in mammals, with reptiles lacking the very extensive airway tree structure found in mammalian lungs. Gas exchange in reptiles still occurs in alveoli however. Reptiles do not possess a diaphragm. Thus, breathing occurs via a change in the volume of the body cavity which is controlled by contraction of intercostal muscles in all reptiles except turtles. In turtles, contraction of specific pairs of flank muscles governs inhalation and exhalation.
Amphibians
Main article: Amphibian § Respiratory system
Both the lungs and the skin serve as respiratory organs in amphibians. The ventilation of the lungs in amphibians relies on positive pressure ventilation. Muscles lower the floor of the oral cavity, enlarging it and drawing in air through the nostrils into the oral cavity. With the nostrils and mouth closed, the floor of the oral cavity is then pushed up, which forces air down the trachea into the lungs. The skin of these animals is highly vascularized and moist, with moisture maintained via secretion of mucus from specialised cells, and is involved in cutaneous respiration. While the lungs are of primary organs for gas exchange between the blood and the environmental air (when out of the water), the skin's unique properties aid rapid gas exchange when amphibians are submerged in oxygen-rich water.
Some amphibians have gills, either in the early stages of their development (e.g. tadpoles of frogs), while others retain them into adulthood (e.g. some salamanders).
Fish
Main article: Fish physiology § Respiration
Fig. 21. The operculum or gill cover of a pike has been pulled open to expose the gill arches bearing filaments.
Fig. 22. A comparison between the operations and effects of a cocurrent and a countercurrent flow exchange system is depicted by the upper and lower diagrams respectively. In both, it is assumed that red has a higher value (e.g. of temperature or the partial pressure of a gas) than blue and that the property being transported in the channels, therefore, flows from red to blue. In fish a countercurrent flow (lower diagram) of blood and water in the gills is used to extract oxygen from the environment.
Fig. 23 The respiratory mechanism in bony fish. The inhalatory process is on the left, the exhalatory process on the right. The movement of water is indicated by the blue arrows.
Oxygen is poorly soluble in water. Fully aerated fresh water therefore contains only 8–10 ml O2/liter compared to the O2 concentration of 210 ml/liter in the air at sea level. Furthermore, the coefficient of diffusion (i.e. the rate at which a substances diffuses from a region of high concentration to one of low concentration, under standard conditions) of the respiratory gases is typically 10,000 faster in air than in water. Thus oxygen, for instance, has a diffusion coefficient of 17.6 mm/s in air, but only 0.0021 mm/s in water. The corresponding values for carbon dioxide are 16 mm/s in air and 0.0016 mm/s in water. This means that when oxygen is taken up from the water in contact with a gas exchanger, it is replaced considerably more slowly by the oxygen from the oxygen-rich regions small distances away from the exchanger than would have occurred in air. Fish have developed gills deal with these problems. Gills are specialized organs containing filaments, which further divide into lamellae. The lamellae contain a dense thin walled capillary network that exposes a large gas exchange surface area to the very large volumes of water passing over them.
Gills use a countercurrent exchange system that increases the efficiency of oxygen-uptake from the water. Fresh oxygenated water taken in through the mouth is uninterruptedly "pumped" through the gills in one direction, while the blood in the lamellae flows in the opposite direction, creating the countercurrent blood and water flow (Fig. 22), on which the fish's survival depends.
Water is drawn in through the mouth by closing the operculum (gill cover), and enlarging the mouth cavity (Fig. 23). Simultaneously the gill chambers enlarge, producing a lower pressure there than in the mouth causing water to flow over the gills. The mouth cavity then contracts, inducing the closure of the passive oral valves, thereby preventing the back-flow of water from the mouth (Fig. 23). The water in the mouth is, instead, forced over the gills, while the gill chambers contract emptying the water they contain through the opercular openings (Fig. 23). Back-flow into the gill chamber during the inhalatory phase is prevented by a membrane along the ventroposterior border of the operculum (diagram on the left in Fig. 23). Thus the mouth cavity and gill chambers act alternately as suction pump and pressure pump to maintain a steady flow of water over the gills in one direction. Since the blood in the lamellar capillaries flows in the opposite direction to that of the water, the consequent countercurrent flow of blood and water maintains steep concentration gradients for oxygen and carbon dioxide along the entire length of each capillary (lower diagram in Fig. 22). Oxygen is, therefore, able to continually diffuse down its gradient into the blood, and the carbon dioxide down its gradient into the water. Although countercurrent exchange systems theoretically allow an almost complete transfer of a respiratory gas from one side of the exchanger to the other, in fish less than 80% of the oxygen in the water flowing over the gills is generally transferred to the blood.
In certain active pelagic sharks, water passes through the mouth and over the gills while they are moving, in a process known as "ram ventilation". While at rest, most sharks pump water over their gills, as most bony fish do, to ensure that oxygenated water continues to flow over their gills. But a small number of species have lost the ability to pump water through their gills and must swim without rest. These species are obligate ram ventilators and would presumably asphyxiate if unable to move. Obligate ram ventilation is also true of some pelagic bony fish species.
There are a few fish that can obtain oxygen for brief periods of time from air swallowed from above the surface of the water. Thus lungfish possess one or two lungs, and the labyrinth fish have developed a special "labyrinth organ", which characterizes this suborder of fish. The labyrinth organ is a much-folded suprabranchial accessory breathing organ. It is formed by a vascularized expansion of the epibranchial bone of the first gill arch, and is used for respiration in air. This organ allows labyrinth fish to take in oxygen directly from the air, instead of taking it from the water in which they reside through the use of gills. The labyrinth organ helps the oxygen in the inhaled air to be absorbed into the bloodstream. As a result, labyrinth fish can survive for a short period of time out of water, as they can inhale the air around them, provided they stay moist. Labyrinth fish are not born with functional labyrinth organs. The development of the organ is gradual and most juvenile labyrinth fish breathe entirely with their gills and develop the labyrinth organs when they grow older.
Invertebrates
Arthropods
See also: Spiracle (arthropods)
Some species of crab use a respiratory organ called a branchiostegal lung. Its gill-like structure increases the surface area for gas exchange which is more suited to taking oxygen from the air than from water. Some of the smallest spiders and mites can breathe simply by exchanging gas through the surface of the body. Larger spiders, scorpions and other arthropods use a primitive book lung.
Insects
Main article: Respiratory system of insects
Most insects breath passively through their spiracles (special openings in the exoskeleton) and the air reaches every part of the body by means of a series of smaller and smaller tubes called 'trachaea' when their diameters are relatively large, and 'tracheoles' when their diameters are very small. The tracheoles make contact with individual cells throughout the body. They are partially filled with fluid, which can be withdrawn from the individual tracheoles when the tissues, such as muscles, are active and have a high demand for oxygen, bringing the air closer to the active cells. This is probably brought about by the buildup of lactic acid in the active muscles causing an osmotic gradient, moving the water out of the tracheoles and into the active cells. Diffusion of gases is effective over small distances but not over larger ones, this is one of the reasons insects are all relatively small. Insects which do not have spiracles and trachaea, such as some Collembola, breathe directly through their skins, also by diffusion of gases.
The number of spiracles an insect has is variable between species, however, they always come in pairs, one on each side of the body, and usually one pair per segment. Some of the Diplura have eleven, with four pairs on the thorax, but in most of the ancient forms of insects, such as Dragonflies and Grasshoppers there are two thoracic and eight abdominal spiracles. However, in most of the remaining insects, there are fewer. It is at the level of the tracheoles that oxygen is delivered to the cells for respiration.
Insects were once believed to exchange gases with the environment continuously by the simple diffusion of gases into the tracheal system. More recently, however, large variation in insect ventilatory patterns has been documented and insect respiration appears to be highly variable. Some small insects do not demonstrate continuous respiratory movements and may lack muscular control of the spiracles. Others, however, utilize muscular contraction of the abdomen along with coordinated spiracle contraction and relaxation to generate cyclical gas exchange patterns and to reduce water loss into the atmosphere. The most extreme form of these patterns is termed discontinuous gas exchange cycles.
Molluscs
Main article: Respiratory system of gastropods
Molluscs generally possess gills that allow gas exchange between the aqueous environment and their circulatory systems. These animals also possess a heart that pumps blood containing hemocyanin as its oxygen-capturing molecule. Hence, this respiratory system is similar to that of vertebrate fish. The respiratory system of gastropods can include either gills or a lung.
Plants
Main article: Photosynthesis
Plants use carbon dioxide gas in the process of photosynthesis, and exhale oxygen gas as waste. The chemical equation of photosynthesis is 6 CO2 (carbon dioxide) and 6 H2O (water), which in the presence of sunlight makes C6H12O6 (glucose) and 6 O2 (oxygen). Photosynthesis uses electrons on the carbon atoms as the repository for the energy obtained from sunlight. Respiration is the opposite of photosynthesis. It reclaims the energy to power chemical reactions in cells. In so doing the carbon atoms and their electrons are combined with oxygen forming CO2 which is easily removed from both the cells and the organism. Plants use both processes, photosynthesis to capture the energy and oxidative metabolism to use it.
Plant respiration is limited by the process of diffusion. Plants take in carbon dioxide through holes, known as stomata, that can open and close on the undersides of their leaves and sometimes other parts of their anatomy. Most plants require some oxygen for catabolic processes (break-down reactions that release energy). But the quantity of O2 used per hour is small as they are not involved in activities that require high rates of aerobic metabolism. Their requirement for air, however, is very high as they need CO2 for photosynthesis, which constitutes only 0.04% of the environmental air. Thus, to make 1 g of glucose requires the removal of all the CO2 from at least 18.7 liters of air at sea level. But inefficiencies in the photosynthetic process cause considerably greater volumes of air to be used.
See also
Great Oxidation Event – Paleoproterozoic surge in atmospheric oxygen
Respiratory adaptation – Breathing chages caused by exertion
Spirometry – Pulmonary function test
Pulmonary function testing (PFT) | biology | 1987979 | https://no.wikipedia.org/wiki/Luftb%C3%A5ren%20smitte | Luftbåren smitte | Luftbåren smitte er overføring av sykdom ved små svevede partikler som kan overføres luftveien, over varierende tid og avstand. Vektor er dråper, også så små dråper at de omtales som aerosoler, mikrodråper.
Sykdommer som kan smitteoverføres luftbårent kan være knyttet til virus, bakteria eller fungi, og de kan spres ved pust, rop, sang, tale, host, nys, støv i bevegelse, spraying eller til og med spylig av toalett.
Referanser
Luftbårne sykdommer | norwegian_bokmål | 0.916957 |
breathe_out_of_one_nostril/Breathing.txt |
Breathing (spiration or ventilation) is the process of moving air into and from the lungs to facilitate gas exchange with the internal environment, mostly to flush out carbon dioxide and bring in oxygen.
All aerobic creatures need oxygen for cellular respiration, which extracts energy from the reaction of oxygen with molecules derived from food and produces carbon dioxide as a waste product. Breathing, or external respiration, brings air into the lungs where gas exchange takes place in the alveoli through diffusion. The body's circulatory system transports these gases to and from the cells, where cellular respiration takes place.
The breathing of all vertebrates with lungs consists of repetitive cycles of inhalation and exhalation through a highly branched system of tubes or airways which lead from the nose to the alveoli. The number of respiratory cycles per minute is the breathing or respiratory rate, and is one of the four primary vital signs of life. Under normal conditions the breathing depth and rate is automatically, and unconsciously, controlled by several homeostatic mechanisms which keep the partial pressures of carbon dioxide and oxygen in the arterial blood constant. Keeping the partial pressure of carbon dioxide in the arterial blood unchanged under a wide variety of physiological circumstances, contributes significantly to tight control of the pH of the extracellular fluids (ECF). Over-breathing (hyperventilation) and under-breathing (hypoventilation), which decrease and increase the arterial partial pressure of carbon dioxide respectively, cause a rise in the pH of ECF in the first case, and a lowering of the pH in the second. Both cause distressing symptoms.
Breathing has other important functions. It provides a mechanism for speech, laughter and similar expressions of the emotions. It is also used for reflexes such as yawning, coughing and sneezing. Animals that cannot thermoregulate by perspiration, because they lack sufficient sweat glands, may lose heat by evaporation through panting.
Mechanics[edit]
Further information: Muscles of respiration
The "pump handle" and "bucket handle movements" of the ribsThe effect of the muscles of inhalation in expanding the rib cage. The particular action illustrated here is called the pump handle movement of the rib cage.In this view of the rib cage the downward slope of the lower ribs from the midline outwards can be clearly seen. This allows a movement similar to the "pump handle effect", but in this case, it is called the bucket handle movement.
BreathingThe muscles of breathing at rest: inhalation on the left, exhalation on the right. Contracting muscles are shown in red; relaxed muscles in blue. Contraction of the diaphragm generally contributes the most to the expansion of the chest cavity (light blue). However, at the same time, the intercostal muscles pull the ribs upwards (their effect is indicated by arrows) also causing the rib cage to expand during inhalation (see diagram on another side of the page). The relaxation of all these muscles during exhalation causes the rib cage and abdomen (light green) to elastically return to their resting positions. Compare these diagrams with the MRI video at the top of the page.The muscles of forceful breathing (inhalation and exhalation). The color code is the same as on the left. In addition to a more forceful and extensive contraction of the diaphragm, the intercostal muscles are aided by the accessory muscles of inhalation to exaggerate the movement of the ribs upwards, causing a greater expansion of the rib cage. During exhalation, apart from the relaxation of the muscles of inhalation, the abdominal muscles actively contract to pull the lower edges of the rib cage downwards decreasing the volume of the rib cage, while at the same time pushing the diaphragm upwards deep into the thorax.
The lungs are not capable of inflating themselves, and will expand only when there is an increase in the volume of the thoracic cavity. In humans, as in the other mammals, this is achieved primarily through the contraction of the diaphragm, but also by the contraction of the intercostal muscles which pull the rib cage upwards and outwards as shown in the diagrams on the right. During forceful inhalation (Figure on the right) the accessory muscles of inhalation, which connect the ribs and sternum to the cervical vertebrae and base of the skull, in many cases through an intermediary attachment to the clavicles, exaggerate the pump handle and bucket handle movements (see illustrations on the left), bringing about a greater change in the volume of the chest cavity. During exhalation (breathing out), at rest, all the muscles of inhalation relax, returning the chest and abdomen to a position called the "resting position", which is determined by their anatomical elasticity. At this point the lungs contain the functional residual capacity of air, which, in the adult human, has a volume of about 2.5–3.0 liters.
During heavy breathing (hyperpnea) as, for instance, during exercise, exhalation is brought about by relaxation of all the muscles of inhalation, (in the same way as at rest), but, in addition, the abdominal muscles, instead of being passive, now contract strongly causing the rib cage to be pulled downwards (front and sides). This not only decreases the size of the rib cage but also pushes the abdominal organs upwards against the diaphragm which consequently bulges deeply into the thorax. The end-exhalatory lung volume is now less air than the resting "functional residual capacity". However, in a normal mammal, the lungs cannot be emptied completely. In an adult human, there is always still at least one liter of residual air left in the lungs after maximum exhalation.
Diaphragmatic breathing causes the abdomen to rhythmically bulge out and fall back. It is, therefore, often referred to as "abdominal breathing". These terms are often used interchangeably because they describe the same action.
When the accessory muscles of inhalation are activated, especially during labored breathing, the clavicles are pulled upwards, as explained above. This external manifestation of the use of the accessory muscles of inhalation is sometimes referred to as clavicular breathing, seen especially during asthma attacks and in people with chronic obstructive pulmonary disease.
Passage of air[edit]
Main article: Respiratory tract
This is a diagram showing how inhalation and exhalation is controlled by a variety of muscles, and what that looks like from a general overall view.
Upper airways[edit]
The lower airways.TracheaMainstem bronchusLobar bronchusSegmental bronchusBronchioleAlveolar ductAlveolus
Inhaled air is warmed and moistened by the wet, warm nasal mucosa, which consequently cools and dries. When warm, wet air from the lungs is breathed out through the nose, the cold hygroscopic mucus in the cool and dry nose re-captures some of the warmth and moisture from that exhaled air. In very cold weather the re-captured water may cause a "dripping nose".
Ideally, air is breathed first out and secondly in through the nose. The nasal cavities (between the nostrils and the pharynx) are quite narrow, firstly by being divided in two by the nasal septum, and secondly by lateral walls that have several longitudinal folds, or shelves, called
nasal conchae, thus exposing a large area of nasal mucous membrane to the air as it is inhaled (and exhaled). This causes the inhaled air to take up moisture from the wet mucus, and warmth from the underlying blood vessels, so that the air is very nearly saturated with water vapor and is at almost body temperature by the time it reaches the larynx. Part of this moisture and heat is recaptured as the exhaled air moves out over the partially dried-out, cooled mucus in the nasal passages, during exhalation. The sticky mucus also traps much of the particulate matter that is breathed in, preventing it from reaching the lungs.
Lower airways[edit]
The anatomy of a typical mammalian respiratory system, below the structures normally listed among the "upper airways" (the nasal cavities, the pharynx, and larynx), is often described as a respiratory tree or tracheobronchial tree (figure on the left). Larger airways give rise to branches that are slightly narrower, but more numerous than the "trunk" airway that gives rise to the branches. The human respiratory tree may consist of, on average, 23 such branchings into progressively smaller airways, while the respiratory tree of the mouse has up to 13 such branchings. Proximal divisions (those closest to the top of the tree, such as the trachea and bronchi) function mainly to transmit air to the lower airways. Later divisions such as the respiratory bronchioles, alveolar ducts and alveoli are specialized for gas exchange.
The trachea and the first portions of the main bronchi are outside the lungs. The rest of the "tree" branches within the lungs, and ultimately extends to every part of the lungs.
The alveoli are the blind-ended terminals of the "tree", meaning that any air that enters them has to exit the same way it came. A system such as this creates dead space, a term for the volume of air that fills the airways at the end of inhalation, and is breathed out, unchanged, during the next exhalation, never having reached the alveoli. Similarly, the dead space is filled with alveolar air at the end of exhalation, which is the first air to breathed back into the alveoli during inhalation, before any fresh air which follows after it. The dead space volume of a typical adult human is about 150 ml.
Gas exchange[edit]
Main article: Gas exchange
The primary purpose of breathing is to refresh air in the alveoli so that gas exchange can take place in the blood. The equilibration of the partial pressures of the gases in the alveolar blood and the alveolar air occurs by diffusion. After exhaling, adult human lungs still contain 2.5–3 L of air, their functional residual capacity or FRC. On inhalation, only about 350 mL of new, warm, moistened atmospheric air is brought in and is well mixed with the FRC. Consequently, the gas composition of the FRC changes very little during the breathing cycle. This means that the pulmonary, capillary blood always equilibrates with a relatively constant air composition in the lungs and the diffusion rate with arterial blood gases remains equally constant with each breath. Body tissues are therefore not exposed to large swings in oxygen and carbon dioxide tensions in the blood caused by the breathing cycle, and the peripheral and central chemoreceptors measure only gradual changes in dissolved gases. Thus the homeostatic control of the breathing rate depends only on the partial pressures of oxygen and carbon dioxide in the arterial blood, which then also maintains a constant pH of the blood.
Control[edit]
Main article: Control of ventilation
The rate and depth of breathing is automatically controlled by the respiratory centers that receive information from the peripheral and central chemoreceptors. These chemoreceptors continuously monitor the partial pressures of carbon dioxide and oxygen in the arterial blood. The first of these sensors are the central chemoreceptors on the surface of the medulla oblongata of the brain stem which are particularly sensitive to pH as well as the partial pressure of carbon dioxide in the blood and cerebrospinal fluid. The second group of sensors measure the partial pressure of oxygen in the arterial blood. Together the latter are known as the peripheral chemoreceptors, and are situated in the aortic and carotid bodies. Information from all of these chemoreceptors is conveyed to the respiratory centers in the pons and medulla oblongata, which responds to fluctuations in the partial pressures of carbon dioxide and oxygen in the arterial blood by adjusting the rate and depth of breathing, in such a way as to restore the partial pressure of carbon dioxide to 5.3 kPa (40 mm Hg), the pH to 7.4 and, to a lesser extent, the partial pressure of oxygen to 13 kPa (100 mm Hg). For example, exercise increases the production of carbon dioxide by the active muscles. This carbon dioxide diffuses into the venous blood and ultimately raises the partial pressure of carbon dioxide in the arterial blood. This is immediately sensed by the carbon dioxide chemoreceptors on the brain stem. The respiratory centers respond to this information by causing the rate and depth of breathing to increase to such an extent that the partial pressures of carbon dioxide and oxygen in the arterial blood return almost immediately to the same levels as at rest. The respiratory centers communicate with the muscles of breathing via motor nerves, of which the phrenic nerves, which innervate the diaphragm, are probably the most important.
Automatic breathing can be overridden to a limited extent by simple choice, or to facilitate swimming, speech, singing or other vocal training. It is impossible to suppress the urge to breathe to the point of hypoxia but training can increase the ability to hold one's breath. Conscious breathing practices have been shown to promote relaxation and stress relief but have not been proven to have any other health benefits.
Other automatic breathing control reflexes also exist. Submersion, particularly of the face, in cold water, triggers a response called the diving reflex. This has the initial result of shutting down the airways against the influx of water. The metabolic rate slows down. This is coupled with intense vasoconstriction of the arteries to the limbs and abdominal viscera, reserving the oxygen that is in blood and lungs at the beginning of the dive almost exclusively for the heart and the brain. The diving reflex is an often-used response in animals that routinely need to dive, such as penguins, seals and whales. It is also more effective in very young infants and children than in adults.
Composition[edit]
Further information: Atmospheric chemistry
Following on from the above diagram, if the exhaled air is breathed out through the mouth on a cold and humid conditions, the water vapor will condense into a visible cloud or mist.
Inhaled air is by volume 78% nitrogen, 20.95% oxygen and small amounts of other gases including argon, carbon dioxide, neon, helium, and hydrogen.
The gas exhaled is 4% to 5% by volume of carbon dioxide, about a 100 fold increase over the inhaled amount. The volume of oxygen is reduced by about a quarter, 4% to 5%, of total air volume. The typical composition is:
5.0–6.3% water vapor
79% nitrogen
13.6–16.0% oxygen
4.0–5.3% carbon dioxide
1% argon
parts per million (ppm) of hydrogen, from the metabolic activity of microorganisms in the large intestine.
ppm of carbon monoxide from degradation of heme proteins.
1 ppm of ammonia.
Trace many hundreds of volatile organic compounds, especially isoprene and acetone. The presence of certain organic compounds indicates disease.
In addition to air, underwater divers practicing technical diving may breathe oxygen-rich, oxygen-depleted or helium-rich breathing gas mixtures. Oxygen and analgesic gases are sometimes given to patients under medical care. The atmosphere in space suits is pure oxygen. However, this is kept at around 20% of Earthbound atmospheric pressure to regulate the rate of inspiration.
Effects of ambient air pressure[edit]
Breathing at altitude[edit]
See also: Effects of high altitude on humans
Fig. 4 Atmospheric pressure
Atmospheric pressure decreases with the height above sea level (altitude) and since the alveoli are open to the outside air through the open airways, the pressure in the lungs also decreases at the same rate with altitude. At altitude, a pressure differential is still required to drive air into and out of the lungs as it is at sea level. The mechanism for breathing at altitude is essentially identical to breathing at sea level but with the following differences:
The atmospheric pressure decreases exponentially with altitude, roughly halving with every 5,500 metres (18,000 ft) rise in altitude. The composition of atmospheric air is, however, almost constant below 80 km, as a result of the continuous mixing effect of the weather. The concentration of oxygen in the air (mmols O2 per liter of air) therefore decreases at the same rate as the atmospheric pressure. At sea level, where the ambient pressure is about 100 kPa, oxygen constitutes 21% of the atmosphere and the partial pressure of oxygen (PO2) is 21 kPa (i.e. 21% of 100 kPa). At the summit of Mount Everest, 8,848 metres (29,029 ft), where the total atmospheric pressure is 33.7 kPa, oxygen still constitutes 21% of the atmosphere but its partial pressure is only 7.1 kPa (i.e. 21% of 33.7 kPa = 7.1 kPa). Therefore, a greater volume of air must be inhaled at altitude than at sea level in order to breathe in the same amount of oxygen in a given period.
During inhalation, air is warmed and saturated with water vapor as it passes through the nose and pharynx before it enters the alveoli. The saturated vapor pressure of water is dependent only on temperature; at a body core temperature of 37 °C it is 6.3 kPa (47.0 mmHg), regardless of any other influences, including altitude. Consequently, at sea level, the tracheal air (immediately before the inhaled air enters the alveoli) consists of: water vapor (PH2O = 6.3 kPa), nitrogen (PN2 = 74.0 kPa), oxygen (PO2 = 19.7 kPa) and trace amounts of carbon dioxide and other gases, a total of 100 kPa. In dry air, the PO2 at sea level is 21.0 kPa, compared to a PO2 of 19.7 kPa in the tracheal air (21% of [100 – 6.3] = 19.7 kPa). At the summit of Mount Everest tracheal air has a total pressure of 33.7 kPa, of which 6.3 kPa is water vapor, reducing the PO2 in the tracheal air to 5.8 kPa (21% of [33.7 – 6.3] = 5.8 kPa), beyond what is accounted for by a reduction of atmospheric pressure alone (7.1 kPa).
The pressure gradient forcing air into the lungs during inhalation is also reduced by altitude. Doubling the volume of the lungs halves the pressure in the lungs at any altitude. Having the sea level air pressure (100 kPa) results in a pressure gradient of 50 kPa but doing the same at 5500 m, where the atmospheric pressure is 50 kPa, a doubling of the volume of the lungs results in a pressure gradient of the only 25 kPa. In practice, because we breathe in a gentle, cyclical manner that generates pressure gradients of only 2–3 kPa, this has little effect on the actual rate of inflow into the lungs and is easily compensated for by breathing slightly deeper. The lower viscosity of air at altitude allows air to flow more easily and this also helps compensate for any loss of pressure gradient.
All of the above effects of low atmospheric pressure on breathing are normally accommodated by increasing the respiratory minute volume (the volume of air breathed in — or out — per minute), and the mechanism for doing this is automatic. The exact increase required is determined by the respiratory gases homeostatic mechanism, which regulates the arterial PO2 and PCO2. This homeostatic mechanism prioritizes the regulation of the arterial PCO2 over that of oxygen at sea level. That is to say, at sea level the arterial PCO2 is maintained at very close to 5.3 kPa (or 40 mmHg) under a wide range of circumstances, at the expense of the arterial PO2, which is allowed to vary within a very wide range of values, before eliciting a corrective ventilatory response. However, when the atmospheric pressure (and therefore the atmospheric PO2) falls to below 75% of its value at sea level, oxygen homeostasis is given priority over carbon dioxide homeostasis. This switch-over occurs at an elevation of about 2,500 metres (8,200 ft). If this switch occurs relatively abruptly, the hyperventilation at high altitude will cause a severe fall in the arterial PCO2 with a consequent rise in the pH of the arterial plasma leading to respiratory alkalosis. This is one contributor to high altitude sickness. On the other hand, if the switch to oxygen homeostasis is incomplete, then hypoxia may complicate the clinical picture with potentially fatal results.
Breathing at depth[edit]
Typical breathing effort when breathing through a diving regulator
Pressure increases with the depth of water at the rate of about one atmosphere – slightly more than 100 kPa, or one bar, for every 10 meters. Air breathed underwater by divers is at the ambient pressure of the surrounding water and this has a complex range of physiological and biochemical implications. If not properly managed, breathing compressed gasses underwater may lead to several diving disorders which include pulmonary barotrauma, decompression sickness, nitrogen narcosis, and oxygen toxicity. The effects of breathing gasses under pressure are further complicated by the use of one or more special gas mixtures.
Air is provided by a diving regulator, which reduces the high pressure in a diving cylinder to the ambient pressure. The breathing performance of regulators is a factor when choosing a suitable regulator for the type of diving to be undertaken. It is desirable that breathing from a regulator requires low effort even when supplying large amounts of air. It is also recommended that it supplies air smoothly without any sudden changes in resistance while inhaling or exhaling. In the graph, right, note the initial spike in pressure on exhaling to open the exhaust valve and that the initial drop in pressure on inhaling is soon overcome as the Venturi effect designed into the regulator to allow an easy draw of air. Many regulators have an adjustment to change the ease of inhaling so that breathing is effortless.
Respiratory disorders[edit]
Medical condition
Breathing patternsGraph showing normal as well as different kinds of pathological breathing patterns.
Abnormal breathing patterns include Kussmaul breathing, Biot's respiration and Cheyne–Stokes respiration.
Other breathing disorders include shortness of breath (dyspnea), stridor, apnea, sleep apnea (most commonly obstructive sleep apnea), mouth breathing, and snoring. Many conditions are associated with obstructed airways. Chronic mouth breathing may be associated with illness. Hypopnea refers to overly shallow breathing; hyperpnea refers to fast and deep breathing brought on by a demand for more oxygen, as for example by exercise. The terms hypoventilation and hyperventilation also refer to shallow breathing and fast and deep breathing respectively, but under inappropriate circumstances or disease. However, this distinction (between, for instance, hyperpnea and hyperventilation) is not always adhered to, so that these terms are frequently used interchangeably.
A range of breath tests can be used to diagnose diseases such as dietary intolerances.
A rhinomanometer uses acoustic technology to examine the air flow through the nasal passages.
Society and culture[edit]
The word "spirit" comes from the Latin spiritus, meaning breath. Historically, breath has often been considered in terms of the concept of life force. The Hebrew Bible refers to God breathing the breath of life into clay to make Adam a living soul (nephesh). It also refers to the breath as returning to God when a mortal dies. The terms spirit, prana, the Polynesian mana, the Hebrew ruach and the psyche in psychology are related to the concept of breath.
In tai chi, aerobic exercise is combined with breathing exercises to strengthen the diaphragm muscles, improve posture and make better use of the body's qi. Different forms of meditation, and yoga advocate various breathing methods. A form of Buddhist meditation called anapanasati meaning mindfulness of breath was first introduced by Buddha. Breathing disciplines are incorporated into meditation, certain forms of yoga such as pranayama, and the Buteyko method as a treatment for asthma and other conditions.
In music, some wind instrument players use a technique called circular breathing. Singers also rely on breath control.
Common cultural expressions related to breathing include: "to catch my breath", "took my breath away", "inspiration", "to expire", "get my breath back".
Breathing and mood[edit]
A young gymnast breathes deeply before performing his exercise.
Certain breathing patterns have a tendency to occur with certain moods. Due to this relationship, practitioners of various disciplines consider that they can encourage the occurrence of a particular mood by adopting the breathing pattern that it most commonly occurs in conjunction with. For instance, and perhaps the most common recommendation is that deeper breathing which utilizes the diaphragm and abdomen more can encourage relaxation. Practitioners of different disciplines often interpret the importance of breathing regulation and its perceived influence on mood in different ways. Buddhists may consider that it helps precipitate a sense of inner-peace, holistic healers that it encourages an overall state of health and business advisers that it provides relief from work-based stress.
Breathing and physical exercise[edit]
During physical exercise, a deeper breathing pattern is adapted to facilitate greater oxygen absorption. An additional reason for the adoption of a deeper breathing pattern is to strengthen the body's core. During the process of deep breathing, the thoracic diaphragm adopts a lower position in the core and this helps to generate intra-abdominal pressure which strengthens the lumbar spine. Typically, this allows for more powerful physical movements to be performed. As such, it is frequently recommended when lifting heavy weights to take a deep breath or adopt a deeper breathing pattern.
See also[edit]
Agonal respiration – Abnormal pattern of breathing (not related to death rattle)
Ataxic respiration – Abnormal pattern of breathing
Bad breath – Presence of unpleasant odors in exhaled breath
Breath gas analysis – monitoring volatile organic compounds present in the exhaled breathPages displaying wikidata descriptions as a fallback
Breathing gas – Gas used for human respiration
Carbon cycle – Natural processes of carbon exchange
Central sleep apnea – Sleep-related disorder in which the effort to breathe is diminished
Eupnea – Natural, comfortable form of breathing in mammals
Liquid breathing – Respiration of oxygen-rich liquid by a normally air-breathing organism
Mouth breathing – Breathing method in humans
Nasal cycle – Unconscious alternation of the nasal cavities
Nitrogen washout – Test for measuring anatomic dead space in the lung during a respiratory cycle
Obligate nasal breathing – physiological necessity to breathe through the nose rather than the mouthPages displaying wikidata descriptions as a fallback
Respiratory adaptation – Breathing chages caused by exertion | biology | 1989624 | https://sv.wikipedia.org/wiki/Halopteris%20diaphragmatica | Halopteris diaphragmatica | Halopteris diaphragmatica är en nässeldjursart som först beskrevs av Chantal Billard 1911. Halopteris diaphragmatica ingår i släktet Halopteris och familjen Halopterididae. Inga underarter finns listade i Catalogue of Life.
Källor
Hydrozoer
diaphragmatica | swedish | 1.229522 |
breathe_out_of_one_nostril/Nasal_cycle.txt | The nasal cycle is the unconscious alternating partial congestion and decongestion of the nasal cavities in humans and other animals. This results in greater airflow through one nostril with periodic alternation between the nostrils. It is a physiological congestion of the nasal conchae, also called the nasal turbinates (curled bony projections within the nasal cavities), due to selective activation of one half of the autonomic nervous system by the hypothalamus. It should not be confused with pathological nasal congestion.
Description[edit]
The nasal cycle was studied and discussed in the ancient Indian yoga of literature of pranayama. In the modern western literature, it was first described by the German physician Richard Kayser in 1895.
In 1927, Heetderks described the alternating turgescence of the inferior turbinates in 80% of a normal population. According to Heetderks, the cycle is the result of alternating congestion and decongestion of the nasal conchae or turbinates, predominantly the inferior turbinates, which are by far the largest of the turbinates in each nasal fossa. Turbinates consist of bony projections covered by erectile tissue, much like the tissues of the penis and clitoris. The turbinates in one fossa fill up with blood while the opposite turbinates decongest by shunting blood away. This cycle, which is controlled by the autonomic nervous system, has a mean duration of two and a half hours but varies widely with age, body-posture, and other conditions. He further observed and documented that the turbinates in the dependent nasal fossa fill when the patient is lying down. The nasal cycle is an alternation in both time and between left and right sides, with the total resistance in the nose remaining constant. The asymmetric airflow may have some benefit to overall olfactory sensitivity. In patients with a fixed septal deviation and intermittent nasal obstruction, the interplay of the nasal cycle becomes evident; the sensation of obstruction frequently mirrors the congestion phase.
It is possible that the nasal cycle may exacerbate the nasal congestion caused by the common cold, as the lack of motility of the cilia in one half of the nose may lead to an uncomfortable sensation of not being able to shift mucus by blowing the nose.
Benefits for breathing[edit]
It has been shown that the cilia of the congested side suspend their motility until that side decongests. Thus the cycle ensures that one side of the nose is always moist, to facilitate humidification, which is one of the three functions of the nose, the other two being filtration and warming of inspired air prior to its entering the lungs.
Benefits for olfaction[edit]
Some odor chemicals bind with olfactory receptors easily, even under conditions of high airflow, and other odors need more time, under low airflow conditions, to bind with receptors. With high airflow on one side and low airflow on the other side, the olfactory center detects a greater range of smells.
Distinction[edit]
The nasal cycle should not be confused with pathological nasal congestion: individuals with normal nasal breathing usually do not realize their breathing is asymmetric unless there is underlying nasal obstruction. In pathological conditions, however, the nasal cycle may influence the symptoms.
Research on the effects[edit]
A 1994 study suggested that breathing through alternate nostrils can affect brain hemisphere symmetry on EEG topography. A later study in 2007 showed that this cycle (as well as manipulation through forced nostril breathing on one side) has an effect on endogenous ultradian rhythms of the autonomic and central nervous system.
However, more recent research has shown no statistically significant association between spontaneously (i.e., not forced) dominant nostril and active brain hemisphere. | biology | 2892787 | https://sv.wikipedia.org/wiki/Condica%20palpalis | Condica palpalis | Condica palpalis är en fjärilsart som beskrevs av Walker 1856. Condica palpalis ingår i släktet Condica och familjen nattflyn. Inga underarter finns listade i Catalogue of Life.
Källor
Nattflyn
palpalis | swedish | 1.358981 |
breathe_out_of_one_nostril/Nose.txt |
A nose is a protuberance in vertebrates that houses the nostrils, or nares, which receive and expel air for respiration alongside the mouth. Behind the nose are the olfactory mucosa and the sinuses. Behind the nasal cavity, air next passes through the pharynx, shared with the digestive system, and then into the rest of the respiratory system. In humans, the nose is located centrally on the face and serves as an alternative respiratory passage especially during suckling for infants.
The protruding nose that is completely separate from the mouth part is a characteristic found only in therian mammals. It has been theorized that this unique mammalian nose evolved from the anterior part of the upper jaw of the reptilian-like ancestors (synapsids).
Air treatment[edit]
3D medical animation still shot depicting a human nose
Acting as the first interface between the external environment and an animal's delicate internal lungs, a nose conditions incoming air, both as a function of thermal regulation and filtration during respiration, as well as enabling the sensory perception of smell.
Hair inside nostrils filter incoming air, as a first line of defense against dust particles, smoke, and other potential obstructions that would otherwise inhibit respiration, and as a kind of filter against airborne illness. In addition to acting as a filter, mucus produced within the nose supplements the body's effort to maintain temperature, as well as contributes moisture to integral components of the respiratory system. Capillary structures of the nose warm and humidify air entering the body; later, this role in retaining moisture enables conditions for alveoli to properly exchange O2 for CO2 (i.e., respiration) within the lungs. During exhalation, the capillaries then aid recovery of some moisture, mostly as a function of thermal regulation, again.
Sense of direction[edit]
The wet nose of dogs is useful for the perception of direction. The sensitive cold receptors in the skin detect the place where the nose is cooled the most and this is the direction a particular smell that the animal just picked up comes from.
Structure in air-breathing forms[edit]
The nose of a tapir
In amphibians and lungfish, the nostrils open into small sacs that, in turn, open into the forward roof of the mouth through the choanae. These sacs contain a small amount of olfactory epithelium, which, in the case of caecilians, also lines a number of neighbouring tentacles. Despite the general similarity in structure to those of amphibians, the nostrils of lungfish are not used in respiration, since these animals breathe through their mouths. Amphibians also have a vomeronasal organ, lined by olfactory epithelium, but, unlike those of amniotes, this is generally a simple sac that, except in salamanders, has little connection with the rest of the nasal system.
In reptiles, the nasal chamber is generally larger, with the choanae located much further back in the roof of the mouth. In crocodilians, the chamber is exceptionally long, helping the animal to breathe while partially submerged. The reptilian nasal chamber is divided into three parts: an anterior vestibule, the main olfactory chamber, and a posterior nasopharynx. The olfactory chamber is lined by olfactory epithelium on its upper surface and possesses a number of turbinates to increase the sensory area. The vomeronasal organ is well-developed in lizards and snakes, in which it no longer connects with the nasal cavity, opening directly into the roof of the mouth. It is smaller in turtles, in which it retains its original nasal connection, and is absent in adult crocodilians.
Birds have a similar nose to reptiles, with the nostrils located at the upper rear part of the beak. Since they generally have a poor sense of smell, the olfactory chamber is small, although it does contain three turbinates, which sometimes have a complex structure similar to that of mammals. In many birds, including doves and fowls, the nostrils are covered by a horny protective shield. The vomeronasal organ of birds is either under-developed or altogether absent, depending on the species.
Elephants have prehensile noses.
The nasal cavities in mammals are both fused into one. Among most species they are exceptionally large, typically occupying up to half the length of the skull. In some groups, however, including primates, bats, and cetaceans, the nose has been secondarily reduced, and these animals consequently have a relatively poor sense of smell. The nasal cavity of mammals has been enlarged, in part, by the development of a palate cutting off the entire upper surface of the original oral cavity, which consequently becomes part of the nose, leaving the palate as the new roof of the mouth. The enlarged nasal cavity contains complex turbinates forming coiled scroll-like shapes that help to warm the air before it reaches the lungs. The cavity also extends into neighbouring skull bones, forming additional air cavities known as paranasal sinuses.
In cetaceans, the nose has been reduced to one or two blowholes, which are the nostrils that have migrated to the top of the head. This adaptation gave cetaceans a more streamlined body shape and the ability to breathe while mostly submerged. Conversely, the elephant's nose has elaborated into a long, muscular, manipulative organ called the trunk.
The vomeronasal organ of mammals is generally similar to that of reptiles. In most species, it is located in the floor of the nasal cavity, and opens into the mouth via two nasopalatine ducts running through the palate, but it opens directly into the nose in many rodents. It is, however, lost in bats, and in many primates, including humans.
In fish[edit]
Fish have a relatively good sense of smell. Unlike that of tetrapods, the nose has no connection with the mouth, nor any role in respiration. Instead, it generally consists of a pair of small pouches located behind the nostrils at the front or sides of the head. In many cases, each of the nostrils is divided into two by a fold of skin, allowing water to flow into the nose through one side and out through the other.
The pouches are lined by olfactory epithelium, and commonly include a series of internal folds to increase the surface area, often forming an elaborate "olfactory rosette". In some teleosts, the pouches branch off into additional sinus-like cavities, while in coelacanths, they form a series of tubes.
In the earliest vertebrates, there was only one nostril and olfactory pouch, and the nasal passage was connected to the hypophysis. The same anatomy is observed in the most primitive living vertebrates, the lampreys and hagfish. In gnathostome ancestors, the olfactory apparatus gradually became paired (presumably to allow sense of direction of smells), and freeing the midline from the nasal passage allowed evolution of jaws.
See also[edit]
Nasal bridge
Obligate nasal breathing
Rhinarium, the wet, naked surface around the nostrils in most mammals, absent in haplorrhine primates such as humans | biology | 657435 | https://da.wikipedia.org/wiki/Lunge%20%28bredere%20betydning%29 | Lunge (bredere betydning) | [[Fil:heart-and-lungs.jpg|thumb|Menneskets lunger flankeret af hjertet og de store blodårer i brystkassen<ref name = "GA">Gray's Anatomy of the Human Body, 20th ed. 1918.</ref>]]
Lungerne (latin pulmones) er den livsnødvendige vejrtrækning (åndedrættet, respirationen) foregår i mange luftåndende dyr, inklusiv de fleste tetrapoder, nogle få fisk og nogle få snegle. I pattedyr og de mere komplekse livsformer, er de to lunger lokaliseret nær rygraden på hver side af hjertet. Lungernes primære funktion er at transportere ilt fra Jordens atmosfære ind i blodstrømmen, og frigive carbondioxid fra blodstrømmen ud i atmosfæren. Denne gasudveksling lader sig gøre i en mosaik af specialiserede celler som danner millioner af små exceptionelt tyndvæggede luftsække kaldet alveoli.
Fugle
Fugles lunger er relativt små, men er forbundet til 8 eller 9 luftsække, som findes igennem det meste af kroppen, og bliver efter tur forbundet til luftrum i knoglerne. Ved inhalation, strømmer luften gennem fuglens luftrør og i luftsækkene. Luft strømmer kontinuert fra luftsækkene bagerst, gennem lungerne, som har en stort set konstant rumfang, til luftsækkene forrest. Fra her, ekshaleres luften. Disse lunger med stort set konstant rumfang kaldes "kredsløbslunger", i kontrast til "lunger af bælgtypen", som findes i de fleste andre dyr.
Fuglelunger indeholder millioner af små parallelle passager kaldet parabronchi.
Dinosaur
Forskere har fremvist bevis og argumenter for luftsække i sauropoder, "prosauropoder", coelurosaurer, ceratosaurer, og theropoderne Aerosteon og Coelophysis.
I "avancerede" sauropoder ("neosauropoder") har de bagerste og hofternes vertebraer tegn på luftsække.
Bevis på luftsække er også fundet i theropoder. Forskning indikerer at coelurosaur fossiler, ceratosaurs,
og theropoderne Coelophysis og Aerosteon udviser bevis på luftsække. Coelophysis, fra den sene kridttid, er en af de tidligste dinosaurer vis fossiler viser bevis på kanaler til luftsække. Aerosteon'', en sen kridttid allosaur, havde de mest fugle-lignende luftsække fundet indtil videre.
Krokodiller, varaner og flyveøgler
Både krokodiller og varaner har udviklet fuglelignende lunger, hvilket giver en envejs luftstrøm og endda luftsække.
De nu uddøde flyveøgler havde tilsyneladende yderligere udviklet denne lungetype, forlænget luftsækkene ud i vingemembranerne - og i tilfældene med lonchodectidae, tupuxuara og azhdarchoidea, i bagbenene.
Kilder/referencer
Se også
Menneskelunge
Asfyksi
Gælle
blad (organ) | danish | 0.760972 |
breathe_out_of_one_nostril/Nostril.txt | A nostril (or naris /ˈnɛərɪs/, pl.: nares /ˈnɛəriːz/) is either of the two orifices of the nose. They enable the entry and exit of air and other gasses through the nasal cavities. In birds and mammals, they contain branched bones or cartilages called turbinates, whose function is to warm air on inhalation and remove moisture on exhalation. Fish do not breathe through noses, but they do have two small holes used for smelling, which can also be referred to as nostrils (with the exception of Cyclostomi, which have just one nostril).
In humans, the nasal cycle is the normal ultradian cycle of each nostril's blood vessels becoming engorged in swelling, then shrinking.
The nostrils are separated by the septum. The septum can sometimes be deviated, causing one nostril to appear larger than the other. With extreme damage to the septum and columella, the two nostrils are no longer separated and form a single larger external opening.
Like other tetrapods, humans have two external nostrils (anterior nares) and two additional nostrils at the back of the nasal cavity, inside the head (posterior nares, posterior nasal apertures or choanae). They also connect the nose to the throat (the nasopharynx), aiding in respiration. Though all four nostrils were on the outside the head of the aquatic ancestors of modern tetrapods, the nostrils for outgoing water (excurrent nostrils) migrated to the inside of the mouth, as evidenced by the discovery of Kenichthys campbelli, a 395-million-year-old fossilized fish which shows this migration in progress. It has two nostrils between its front teeth, similar to human embryos at an early stage. If these fail to join up, the result is a cleft palate.
Each external nostril contains approximately 1,000 strands of nasal hair, which function to filter foreign particles such as pollen and dust.
It is possible for humans to smell different olfactory inputs in the two nostrils and experience a perceptual rivalry akin to that of binocular rivalry when there are two different inputs to the two eyes. Furthermore, scent information from the two nostrils leads to two types of neural activity with the first cycle corresponding to the ipsilateral and the second cycle corresponding to the contralateral odor representations.
The Procellariiformes are distinguished from other birds by having tubular extensions of their nostrils.
Widely-spaced nostrils, like those of the hammerhead shark, may be useful in determining the direction of an odour's source.
See also[edit]
Dilator naris muscle
Nasal cycle | biology | 873360 | https://da.wikipedia.org/wiki/Nasal%20konsonant | Nasal konsonant | En nasal lukkelyd fremkommer når velum sænkes, således at luften kan strømme frit gennem næsen. Mundhulen fungerer fortsat som et resonanskammer for lyden, men luften slipper ikke ud gennem munden, da den bliver blokeret af tungen eller læberne. Dermed er det ikke næsen selv som udgør forskellen mellem de forskellige nasale lukkelyde, men derimod mundens artikulation, som i plosiver.
Liste over nasale lukkelyder:
[] er en stemt bilabial nasal
[] er en stemt labiodental nasal (SAMPA: [F])
[] er en alveolar eller dental nasal: se alveolar nasal
[] stemt retrofleks nasal (SAMPA: [n`])
[] stemt palatal nasal (SAMPA: [J]); er en udbredt lyd in europæiske sprog som i spansk ñ; fransk og italiensk gn; katalansk og ungarsk ny; eller portugisisk nh.
[] stemt velar nasal (SAMPA: [N]), som i ting.
[] stemt uvular nasal (SAMPA: [N\])
Fonetik | danish | 0.760683 |
vision_of_lights/Phosphene.txt | A phosphene is the phenomenon of seeing light without light entering the eye. The word phosphene comes from the Greek words phos (light) and phainein (to show). Phosphenes that are induced by movement or sound may be associated with optic neuritis.
Phosphenes can be induced by mechanical, electrical, or magnetic stimulation of the retina or visual cortex, or by random firing of cells in the visual system. Phosphenes have also been reported by meditators (called nimitta), people who endure long periods without visual stimulation (the prisoner's cinema), or those who ingest psychedelic drugs.
Causes[edit]
Mechanical stimulation[edit]
The most common phosphenes are pressure phosphenes, caused by rubbing or applying pressure on or near the closed eyes. They have been known since antiquity, and described by the Greeks. The pressure mechanically stimulates the cells of the retina. Experiences include a darkening of the visual field that moves against the rubbing, a diffuse colored patch that also moves against the rubbing, well defined shapes such as bright circles that exist near or opposite to where pressure is being applied, a scintillating and ever-changing and deforming light grid with occasional dark spots (like a crumpling fly-spotted flyscreen), and a sparse field of intense blue points of light. Pressure phosphenes can persist briefly after the rubbing stops and the eyes are opened, allowing the phosphenes to be seen on the visual scene. Hermann von Helmholtz and others have published drawings of their pressure phosphenes. One example of a pressure phosphene is demonstrated by gently pressing the side of one's eye and observing a colored ring of light on the opposite side, as detailed by Isaac Newton.
Another common phosphene is "seeing stars" from a sneeze, laughter, a heavy and deep cough, blowing of the nose, a blow on the head or low blood pressure (such as on standing up too quickly or prior to fainting). It is possible these involve some mechanical stimulation of the retina, but they may also involve mechanical and metabolic (such as from low oxygenation or lack of glucose) stimulation of neurons of the visual cortex or of other parts of the visual system.
Less commonly, phosphenes can also be caused by some diseases of the retina and nerves, such as multiple sclerosis. The British National Formulary lists phosphenes as an occasional side effect of at least one anti-anginal medication.
The name "phosphene" was coined by J. B. H. Savigny, better known as the ship's surgeon of the wrecked French frigate Méduse. It was first employed by Serre d'Uzes to test retinal function prior to cataract surgery.
Electrical stimulation[edit]
Phosphenes have been created by electrical stimulation of the brain, reported by neurologist Otfrid Foerster as early as 1929. Brindley and Lewin (1968) inserted a matrix of stimulating electrodes directly into the visual cortex of a 52-year-old blind female, using small pulses of electricity to create phosphenes. These phosphenes were points, spots, and bars of colorless or colored light. Brindley and Rushton (1974) used the phosphenes to create a visual prosthesis, in this case by using the phosphenes to depict Braille spots.
In recent years, researchers have successfully developed experimental brain–computer interfaces or neuroprostheses that stimulate phosphenes to restore vision to people blinded through accidents. Notable successes include the human experiments by William H. Dobelle and Mark Humayun and animal research by Dick Normann.
A noninvasive technique that uses electrodes on the scalp, transcranial magnetic stimulation, has also been shown to produce phosphenes.
Experiments with humans have shown that when the visual cortex is stimulated above the calcarine fissure, phosphenes are produced in the lower part of the visual field, and vice versa.
Others[edit]
Phosphenes have been produced by intense, changing magnetic fields, such as with transcranial magnetic stimulation (TMS). These fields can be positioned on different parts of the head to stimulate cells in different parts of the visual system. They also can be induced by alternating currents that entrain neural oscillation as with transcranial alternating current stimulation. In this case they appear in the peripheral visual field. This claim has been disputed. The alternative hypothesis is that current spread from the occipital electrode evokes phosphenes in the retina. Phosphenes created by magnetic fields are known as magnetophosphenes.
Astronauts exposed to radiation in space have reported seeing phosphenes. Patients undergoing radiotherapy have reported seeing blue flashes of light during treatment; the underlying phenomenon has been shown to resemble Cherenkov radiation.
Phosphenes can be caused by some medications, such as Ivabradine.
Mechanism[edit]
Most vision researchers believe that phosphenes result from the normal activity of the visual system after stimulation of one of its parts from some stimulus other than light. For example, Grüsser et al. showed that pressure on the eye results in activation of retinal ganglion cells in a similar way to activation by light. An ancient, discredited theory is that light is generated in the eye. A version of this theory has been revived, except, according to its author, that "phosphene lights are [supposed to be] due to the intrinsic perception of induced or spontaneous increased biophoton emission of cells in various parts of the visual system (from retina to cortex)"
Anthropological research[edit]
In 1988, David Lewis-Williams and T. A. Dowson published an article about phosphenes and other entoptic phenomena. They argued that non-figurative art of the Upper Paleolithic depicts visions of phosphenes and neurological "form constants", probably enhanced by hallucinogenic drugs.
Research[edit]
Research has looked into visual prosthesis for the blind, which involves use of arrays of electrodes implanted in the skull over the occipital lobe to produce phosphenes. There have been long term implants of this type. Risks, such as infections and seizures, have been an impediment to their development.
A possible use of phosphenes as part of a brain to brain communication system has been reported. The system called BrainNet, produces phosphenes using transcranial magnetic stimulation (TMS). The goal of the research is to connect thoughts brain to brain using a system where signals are detected using electroencephalography (EEG) and delivered using transcranial magnetic stimulation (TMS). An experiment was conducted with 5 different groups, each contained three people. The subjects were split into two groups. Two subjects functioned as the senders, and were connected to EEG electrodes, and a third person functioned as the receiver, who wore the TMS helmet. Each person was stationed in front of a television screen with a Tetris-style game. The senders had to determine if there was a need to rotate the falling blocks, but without the ability to rotate them – only the receiver was able to perform this operation. At the edges of each screen, were two icons with two flashing lights in two different frequencies, (one at 15 Hz and the other at 17 Hz). The sender focused on one icon, or the other to signal that the block should be rotated to the right or the left. The EEG produced a unique signal, which was transmitted to the TMS helmet of the receiver, who perceived phosphenes which differed for the 15 Hz and 17 Hz signal, and rotated the block in the relevant direction. The experiment achieved 81% success.
See also[edit]
Closed-eye hallucination – Class of hallucination
Dark retreat – Tibetan Buddhism advanced practice
Isolation tank – Pitch-black, light-proof, soundproof environment heated to the same temperature as the skin
Prisoner's cinema – Visual phenomenon involving seeing animated lights in the darkness
Scintillating scotoma – Visual aura associated with migraine
Photopsia – Presence of perceived flashes of light in one's field of vision
Visual snow – Class of hallucinationPages displaying short descriptions of redirect targets
HPPD – Medical condition | biology | 24270 | https://da.wikipedia.org/wiki/Fotonisk%20krystal | Fotonisk krystal | Fotoniske krystaller er metamaterialer, der udgøres af periodiske dielektriske eller metal-dielektriske (nano)strukturer som er designet til at påvirke elektromagnetiske bølgers (EM) udbredelse på den samme måde som det periodiske potential i et halvledende krystal påvirker elektronernes bevægelse ved at definere tilladte og forbudte elektriske energibånd.
Fraværet af tilladte udbredelsesenergibånd indeni strukturen for et interval af bølgelængder kaldes et fotonisk båndgab, som forårsager bemærkelsesværdige optiske fænomener, som bl.a. resulterer i; spontan udsendelse, generering af alle regnbuens farver ud fra infrarødt-lys, højreflekterende spejle som virker i alle retninger og lysleder med lavt tab, og materialer med negativt brydningsindeks.
Fotonisk krystal er grundlæggende set baseret på det fysiske fænomen diffraktion.
Kilder/referencer
Se også
Kvantemekanik
Optik
Nanoteknologi
Kvanteø
Opal
Eksterne henvisninger
Webarchive backup: Ingeniøren nr. -3/1999: Dansk gennembrud i fiberoptik Danske og engelske forskere har sammen bevist, at optisk fiber med huller i kan transportere lys over lange afstande.
Vejviser: Photonic Crystal and Photonic Band Gap Links
Cnet, August 31, 2000, Why photonics? Citat: "...Demand for photonic equipment is skyrocketing. Internet traffic on the backbone networks has been doubling every three months and shows no sign of abating. Companies are laying fiber in just about every cross-country right-of-way they can find. First it was along the railways, and now it's the gas pipelines, sewers and just about any conduit you can think of... "
November 3, 2000, Sandia LabNews: Cheesecloth-like photonics device bends light with little loss Citat: "...the cheesecloth-like structure can be considered essentially a wire for light...Because of the very small light loss, the technique offers the potential of ultimately replacing electronic chips with faster, cooler photonic chips...two-dimensional crystals are cheaper and far easier to build..."
Number 646 #1, July 16, 2003, AIP: Photonic Crystal Shifts Energy Citat: "...Shawn Lin and his Sandia colleagues, in the course of their studies of photonic crystals, have seemed to challenge the venerable formulation, made by Max Planck a hundred years ago, of what kind of emission spectrum a body should have..."
CERN Courier: Photonic crystal makes flat lens Citat: "...The key to creating the flat lens lies with the recent advent of materials – photonic crystals – that effectively have a negative index of refraction...the principle could herald a revolution in optics..."
BBC News: 3 January, 2001, Sea mouse promises bright future Citat: "...The sea mouse, or Aphrodita, has spines that normally appear deep red in colour. But when light falls on a spine perpendicular to its axis, stripes of different colours appear – strong blues and greens..."The simple structure responsible for this effect is a remarkable example of photonic engineering by a living organism."..."These structures may have application in photonic communications, where there is much interest in fabricating photonic crystal fibres with similar morphology."..."
Metamaterialer
Fotonik | danish | 0.653631 |
vision_of_lights/Vision_and_Light.txt | Skip to main content
Table of Contents menu
search Search build_circle Toolbar fact_check Homework cancel Exit
Reader Mode
* school Campus Bookshelves
* menu_book Bookshelves
* perm_media Learning Objects
* login Login
* how_to_reg Request Instructor Account
* hub Instructor Commons
## Search
Search this book
Submit Search
* Downloads expand_more
* Download Page (PDF)
* Download Full Book (PDF)
* Resources expand_more
* Periodic Table
* Physics Constants
* Scientific Calculator
* Reference expand_more
* Reference & Cite
* Tools expand_more
* Help expand_more
* Get Help
* Feedback
* Readability
##
x
selected template will load here
## Error
This action is not available.
chrome_reader_mode Enter Reader Mode
Photoreceptors
Supplemental Modules (Biological Chemistry)
{ }
{ Chemistry_of_Vision : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Photoreceptor_Excitation : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Photoreceptor_Proteins : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Vision_and_Light : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()"
}
{ Carbohydrates : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Enzymes : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Lipids : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Medicinal_Chemistry : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Metabolism : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Nucleic_Acids : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Pharmaceuticals : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Photoreceptors : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Photosynthesis : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Proteins : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()",
Vitamins_Cofactors_and_Coenzymes : "property get [Map
MindTouch.Deki.Logic.ExtensionProcessorQueryProvider+<>c__DisplayClass230_0.<PageSubPageProperty>b__1]()"
}
Sat, 17 Feb 2024 02:43:13 GMT
Vision and Light
480
480
Joshua Halpern
{ }
Anonymous
Anonymous User
2
false
false
[ "article:topic", "showtoc:no", "license:ccbyncsa", "licenseversion:40" ]
[ "article:topic", "showtoc:no", "license:ccbyncsa", "licenseversion:40" ]
https://chem.libretexts.org/@app/auth/3/login?returnto=https%3A%2F%2Fchem.libretexts.org%2FBookshelves%2FBiological_Chemistry%2FSupplemental_Modules_(Biological_Chemistry)%2FPhotoreceptors%2FVision_and_Light
1. Search site
Search Search
Go back to previous article
2. 1. Username
Password
Sign in
2. Sign in
* Sign in
* Forgot password
Expand/collapse global hierarchy
1. Home
2. Bookshelves
3. Biological Chemistry
4. Supplemental Modules (Biological Chemistry)
5. Photoreceptors
6. Vision and Light
Expand/collapse global location
# Vision and Light
1. Last updated
2. Save as PDF
3.
* Page ID
480
\\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup}
{\mathbf{#1}} } \\) \\(
\newcommand{\vecd}[1]{\overset{-\\!-\\!\rightharpoonup}{\vphantom{a}\smash
{#1}}} \\)\\(\newcommand{\id}{\mathrm{id}}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\) \\(
\newcommand{\kernel}{\mathrm{null}\,}\\) \\(
\newcommand{\range}{\mathrm{range}\,}\\) \\(
\newcommand{\RealPart}{\mathrm{Re}}\\) \\(
\newcommand{\ImaginaryPart}{\mathrm{Im}}\\) \\(
\newcommand{\Argument}{\mathrm{Arg}}\\) \\( \newcommand{\norm}[1]{\| #1 \|}\\)
\\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\) \\(\newcommand{\id}{\mathrm{id}}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\) \\(
\newcommand{\kernel}{\mathrm{null}\,}\\) \\(
\newcommand{\range}{\mathrm{range}\,}\\) \\(
\newcommand{\RealPart}{\mathrm{Re}}\\) \\(
\newcommand{\ImaginaryPart}{\mathrm{Im}}\\) \\(
\newcommand{\Argument}{\mathrm{Arg}}\\) \\( \newcommand{\norm}[1]{\| #1 \|}\\)
\\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\\) \\(
\newcommand{\Span}{\mathrm{span}}\\)\\(\newcommand{\AA}{\unicode[.8,0]{x212B}}\\)
1. Introduction
2. Mechanism of Vision
3. Signal Transduction Pathway
4. References
5. Contributors
Vision is such an everyday occurrence that we seldom stop to think and wonder
how we are able to see the objects that surround us. Yet the vision process is
a fascinating example of how light can produce molecular changes. The retina
contain the molecules that undergo a chemical change upon absorbing light, but
it is the brain that actually makes sense of the visual information to create
an image.
## Introduction
Light is one of the most important resources for civilization, it provides
energy as it pass along by the sun. Light influence our everyday live. Living
organisms sense light from the environment by photoreceptors. Light, as waves
carry energy, contains energy by different wavelength. In vision, light is the
stimulus input. Light energy goes into eyes stimulate photoreceptor in eyes.
However, as an energy wave, energy is passed on through light at different
wavelength.
Light, as waves carry energy, contains energy by different wavelength. From
long wavelength to short wavelength, energy increase. 400 nm to 700 nm is
visible spectrum.
Light energy can convert chemical to other forms. Vitamin A , also known as
retinol, anti-dry eye vitamins, is a required nutrition for human health. The
predecessor of vitamin A is present in the variety of plant carotene. Vitamin
A is critical for vision because it is needed by the retina of eye. Retinol
can be convert to retinal, and retinal is a chemical necessary for rhodopsin.
As light enters the eye, the 11- cis -retinal is isomerized to the
all-"trans" form.
## Mechanism of Vision
The molecule cis-retinal can absorb light at a specific wavelength. When
visible light hits the cis-retinal, the cis-retinal undergoes an
isomerization , or change in molecular arrangement, to all-trans-retinal. The
new form of trans-retinal does not fit as well into the protein, and so a
series of geometry changes in the protein begins. The resulting complex is
referred to a bathrhodopsin (there are other intermediates in this process,
but we'll ignore them for now).
The reaction above shows Lysine side-chain from the opsin react with 11-cis-
retinal when stimulated. By removing the oxygen atom from the retinal and two
hydrogen atom from the free amino group of the lysine, the linkage show on the
picture above is formed, and it is called Schiff base.
## Signal Transduction Pathway
As the protein changes its geometry, it initiates a cascade of biochemical
reactions that results in changes in charge so that a large potential
difference builds up across the plasma membrane. This potential difference is
passed along to an adjoining nerve cell as an electrical impulse. The nerve
cell carries this impulse to the brain, where the visual information is
interpreted.
The light image is mapped on the surface of the retina by activating a series
of light-sensitive cells known as rods and cones or photoreceptors. The rods
and cones convert the light into electrical impulses which are transmitted to
the brain via nerve fibers. The brain then determines, which nerve fibers
carried the electrical impulse activate by light at certain photoreceptors,
and then creates an image.
The retina is lined with many millions of photoreceptor cells that consist of
two types: 7 million cones provide color information and sharpness of images,
and 120 million rods are extremely sensitive detectors of white light to
provide night vision. The tops of the rods and cones contain a region filled
with membrane-bound discs, which contain the molecule cis-retinal bound to a
protein called opsin. The resulting complex is called rhodopsin or "visual
purple".
Figure 3: A fundus photograph of the back of the retina. The white area is
the beginning of the optical nerve (optic disc). The image in this photo is
the right eye of eric anthamatten. (CC-BY-SA-4.0; TheGoose aPrisoner) .
In human eyes, rod and cones react to light stimulation, and a series of
chemical reactions happen in cells. These cells receive light, and pass on
signals to other receiver cells. This chain of process is class signal
transduction pathway. Signal transduction pathway is a mechanism that describe
the ways cells react and response to stimulation.
## References
1. Biochemistry, L. Stryer (W.H. Freeman and Co, San Francisco, 1975).
2. The Cambridge Guide to the Material World, Rodney Cotterill (Cambridge University Press, Cambridge, 1985)
## Contributors
* {{template.Contrib Ophardt()}}
* * *
Vision and Light is shared under a CC BY-NC-SA 4.0 license and was
authored, remixed, and/or curated by LibreTexts.
1. Back to top
2. * Photoreceptor Proteins
* Photosynthesis
* Was this article helpful?
* Yes
* No
*
## Recommended articles
1. Article type
Section or Page
License
CC BY-NC-SA
License Version
4.0
Show Page TOC
no on page
2. Tags
This page has no tags.
1. © Copyright 2024 Chemistry LibreTexts
2. Powered by CXone Expert ®
The LibreTexts libraries are Powered by NICE CXone Expert and are supported
by the Department of Education Open Textbook Pilot Project, the UC Davis
Office of the Provost, the UC Davis Library, the California State University
Affordable Learning Solutions Program, and Merlot. We also acknowledge
previous National Science Foundation support under grant numbers 1246120,
1525057, and 1413739. Legal . Accessibility Statement For more information
contact us at [email protected] .
| biology | 34062 | https://sv.wikipedia.org/wiki/Maple | Maple | Maple är ett datoralgebrasystem, det vill säga ett datorprogram för symbolisk lösning av matematiska problem och tekniska beräkningar, från företaget Maplesoft. Maple utvecklades 1981 vid Symbolic Computation Group - University of Waterloo i Ontario, Kanada. Maple täcker aspekter av teknisk databehandling, inklusive visualisering, dataanalys, matrisberäkning och anslutning. En verktygslåda, MapleSim, lägger till funktionalitet för multidomain fysisk modellering och kodgenerering.
Användningsområden
Matematiska beräkningar: symbolisk och numerisk lösning av matematiska problem t. ex. ekvationer, integraler och "pretty-printing" av matematiska formler
Differentialekvation, Ordinär differentialekvation, Partiell differentialekvation
Elementär och speciell funktionsbibliotek
Finansiell modellering, analys och applikationsutveckling
Flervariabelanalys
Grafisk framställning
Komplexa tal och komplex analys
Linjär algebra: Singulärvärdesuppdelning
Matrisoperationer och matrisalgebra
Optimeringslära, Linjärprogrammering
Statistik och dataanalys: Regressionsanalys, Hypotesprövning, ANOVA, Principalkomponentanalys
Test och uppmätning: Hårdvarukoppling och dataanalys för test och mätapplikationer
Teknisk beräkning: analys, signalprocesser, visualisering och algoritmutveckling
Tidsserieanalys
Anslutningar
Matlab anslutningar
Excel add-in
Kodgenerering för C, C#, Fortran, Java, Javascript, Julia, Matlab, Perl, Python, R, VB
Gränssnitt till CAD, HTTP, SQL, .NET
Exempel
Procedur
myfac := proc(n::nonnegint)
local out, i;
out := 1;
for i from 2 to n do
out := out * i
end do;
out
end proc;
Funktion
myfac := n -> product( i, i=1..n );
Integration
.
int(cos(x/a), x);
Resultat:
Determinant
M:= Matrix([[1,2,3], [a,b,c], [x,y,z]]);
LinearAlgebra:-Determinant(M);
Resultat:
Serie expansion
series(tanh(x),x=0,15)
Resultat:
Lös ekvation numeriskt
f := x^53-88*x^5-3*x-5 = 0
fsolve(f)
Resultat:
-1.097486315, -.5226535640, 1.099074017
Visualisering av en enkel variabel funktion
plot(x*sin(x),x=-10..10);
Visualisering av en funktion av två variabler
plot3d(2-x-(y^2-x^2)^0.5), x=0..1, y=0..1);
Animation
plots:-animate(subs(k = .5, f), x=-30..30, t=-10..10, numpoints=200, frames=50, color=red, thickness=3);
plots:-animate3d(cos(t*x)*sin(3*t*y), x=-Pi..Pi, y=-Pi..Pi, t=1..2);
M := Matrix([[400,400,200], [100,100,-400], [1,1,1]], datatype=float[8]):
plot3d(1, x=0..2*Pi, y=0..Pi, axes=none, coords=spherical, viewpoint=[path=M]);
Laplace transform
f := (1+A*t+B*t^2)*exp(c*t);
inttrans:-laplace(f, t, s);
Resultat:
inttrans:-invlaplace(1/(s-a),s,x)
Resultat:
Fourier transform
inttrans:-fourier(sin(x),x,w)
Resultat:
Se även
Mathcad
Mathematica
MATLAB
GNU Maxima
Referenser
Noter
Externa länkar
Maplesoft
Produktdokumentation
MapleSim
Maple til danske gymnasier
Datoralgebrasystem
Matematikprogram
Programspråk
Statistikprogram
Programvaror 1982 | swedish | 0.853418 |
vision_of_lights/content.aspx.txt | Skip to main content
Coronavirus (COVID-19): Latest Updates | Visitation Policies Visitation Policies Visitation Policies Visitation Policies Visitation Policies | COVID-19 Testing | Vaccine Information Vaccine Information Vaccine Information
menu
* Patient Care
* Education
* Medical Education
* Residency & Fellowship
* Graduate Education
* Postdoctoral Affairs
* Dental Education
* Nursing Education
* Libraries
* Alumni
More information on Education
* URMC Research
* URMC Research Labs
* UR Health Research
* Education & Training at the University of Rochester
* Shared Resource Labs and Facilities
* Technology Transfer
More information on Research
* Community
* Center for Community Health & Prevention
* Government & Community Relations
* Monroe County Community Health Improvement Plan
* Pediatric Community Resources
* Health Equity
* Community Dentistry
* Mental Health Community Resources
* Health Research
More information on Community
* About URMC
* For Referring Physicians at UR Medicine
* Get Care Now
* MyChart Login
## Health Encyclopedia
###
* Tests & Procedures
* Interactive Encyclopedia Tools
* Healthy Living
* Your Family
* Drug Reference
* Herbs, Vitamins & Supplements
* Prevention Planner
URMC / Encyclopedia / Content
Search Encyclopedia
# Normal Vision
## What is normal vision?
To understand how certain problems can affect your child's vision, it’s
important to know how normal vision happens. For children with normal vision,
the following things happen in this order:
1. Light enters the eye through the cornea. This is the clear, dome-shaped surface that covers the front of the eye.
2. From the cornea, the light passes through the pupil. The iris, or the colored part of your eye, controls the amount of light passing through.
3. From there, it then hits the lens. This is the clear structure inside the eye that focuses light rays onto the retina.
4. Next, light passes through the vitreous humor. This is the clear, jelly-like substance that fills the center of the eye. It helps to keep the eye round in shape.
5. Finally, the light reaches the retina. This is the light-sensitive nerve layer that lines the back of the eye.
6. The optic nerve is then responsible for carrying the signals to the visual cortex of the brain. The visual cortex turns the signals into images.
Medical Reviewers:
* Chris Haupert MD
* Tara Novick BSN MSN
* Whitney Seltman MD
Ask a Medical Librarian Make an Appointment
## Physicians & Services
* Physicians who treat the Eyes
* Pediatric Ophthalmology at Golisano Children's Hospital
* Flaum Eye Institute
## Interactive Tools
* Eyesight Quiz
* Vision Quiz
* URMC Information
* About URMC
* Our Hospitals
* Directions & Parking
* Newsroom
* Job Opportunities
* Closings & Cancellations
* Contact Information
* Tools
* Get Care Now
* MyChart Patient Portal
* Find a Provider
* Online Bill Pay
* Price Transparency
* Health Encyclopedia
* Event Calendar
* Faculty Lists
* General Information
* University of Rochester
* Policies
* Faculty & Staff Directory
* Emergency Information
©2024 University of Rochester Medical Center Rochester, NY
* Intranet
* Email
* HRMS
* Remote Access (VPN)
* Privacy Topics
Viewing: K1
*[
NY
]: New York
*[
HRMS
]: Human Resources Management System
*[
(VPN)
]: Virtual Private Network
| biology | 117199 | https://nn.wikipedia.org/wiki/1505%20Koranna | 1505 Koranna | 1505 Koranna er ein hovudbelteasteroide. Han blei oppdaga den 21. april 1939 av Cyril V. Jackson ifrå Union Observatory i Johannesburg.
Kjelder
1505 Koranna - baneparametrar hjå JPL Solar System Dynamics
Oppdagarar og oppdagingsplassar i Engelsk Wikipedia sine asteroidelister. Listene baserer seg på Minor Planet Center - «Discovery Circumstances: Numbered Minor Planets»
Koranna
Koranna | norwegian_nynorsk | 1.196159 |
vision_of_lights/vision-it-all-starts-with-light.txt | * Thinking, Sensing & Behaving
* Aging
* Brain Development
* Childhood & Adolescence
* Diet & Lifestyle
* Emotions, Stress & Anxiety
* Hearing
* Language
* Learning & Memory
* Movement
* Pain
* Sleep
* Smell
* Taste
* Thinking & Awareness
* Touch
* Vision
* See All
* Diseases & Disorders
* Addiction
* Alzheimer's & Dementia
* Cancer
* Childhood Disorders
* Epilepsy
* Immune System Disorders
* Injury
* Mental Health
* Neurodegenerative Disorders
* Infectious Disease
* COVID-19
* Neurological Disorders A-Z
* Therapies
* See All
* Brain Anatomy & Function
* Anatomy
* Body Systems
* Cells & Circuits
* Evolution
* Genes & Molecules
* See All
* Neuroscience in Society
* The Arts & the Brain
* Law, Economics & Ethics
* Neuroscience in the News
* Supporting Research
* Tech & the Brain
* See All
* In the Lab
* Animals in Research
* BRAIN Initiative
* Meet the Researcher
* Neuro-technologies
* Tools & Techniques
* See All
* Explore
* 3D Brain
* Core Concepts
* For Educators
* Ask an Expert
* Glossary
* The Brain Facts Book
* About Us
* MyAccount
For Educators
Log in
* Thinking, Sensing & Behaving
* Aging
* Brain Development
* Childhood & Adolescence
* Diet & Lifestyle
* Emotions, Stress & Anxiety
* Hearing
* Language
* Learning & Memory
* Movement
* Pain
* Sleep
* Smell
* Taste
* Thinking & Awareness
* Touch
* Vision
* See All
* Diseases & Disorders
* Addiction
* Alzheimer's & Dementia
* Cancer
* Childhood Disorders
* Epilepsy
* Immune System Disorders
* Injury
* Mental Health
* Neurodegenerative Disorders
* Infectious Disease
* COVID-19
* Neurological Disorders A-Z
* Therapies
* See All
* Brain Anatomy & Function
* Anatomy
* Body Systems
* Cells & Circuits
* Evolution
* Genes & Molecules
* See All
* Neuroscience in Society
* The Arts & the Brain
* Law, Economics & Ethics
* Neuroscience in the News
* Supporting Research
* Tech & the Brain
* See All
* In the Lab
* Animals in Research
* BRAIN Initiative
* Meet the Researcher
* Neuro-technologies
* Tools & Techniques
* See All
# Vision: It all Starts with Light
* Published 1 Apr 2012
* Reviewed 1 Apr 2012
* Source BrainFacts/SfN
To be able to see anything, eyes first need to process light. Vision begins
with light passing through the cornea, which does about three-quarters of the
focusing, and then the lens, which adjusts the focus. Both combine to produce
a clear image of the visual world on a sheet of photoreceptors called the
retina, which is part of the central nervous system but located at the back of
the eye.
Photoreceptors gather visual information by absorbing light and sending
electrical signals to other retinal neurons for initial processing and
integration. The signals are then sent via the optic nerve to other parts of
brain, which ultimately processes the image and allows us to see.
As in a camera, the image on the retina is reversed: Objects to the right of
center project images to the left part of the retina and vice versa; objects
above the center project to the lower part and vice versa.
The size of the pupil, which regulates how much light enters the eye, is
controlled by the iris. The shape of the lens is altered by the muscles just
behind the iris so that near or far objects can be brought into focus on the
retina.
Primates, including humans, have well-developed vision using two eyes, called
binocular vision. Visual signals pass from each eye along the million or so
fibers of the optic nerve to the optic chiasm, where some nerve fibers cross
over. This crossover allows both sides of the brain to receive signals from
both eyes.
When you look at a scene with both eyes, the objects to your left register on
the right side of the retina. This visual information then maps to the right
side of the cortex. The result is that the left half of the scene you are
watching registers in the cerebrum’s right hemisphere. Conversely, the right
half of the scene registers in the cerebrum’s left hemisphere. A similar
arrangement applies to movement and touch: Each half of the cerebrum is
responsible for processing information received from the opposite half of the
body.
Scientists know much about the way cells encode visual information in the
retina, but relatively less about the lateral geniculate nucleus — an
intermediate way station between the retina and visual cortex — and the visual
cortex. Studies about the inner workings of the retina give us the best
knowledge we have to date about how the brain analyzes and processes sensory
information.
Photoreceptors, about 125 million in each human eye, are neurons specialized
to turn light into electrical signals. Two major types of photoreceptors are
rods and cones. Rods are extremely sensitive to light and allow us to see in
dim light, but they do not convey color. Rods constitute 95 percent of all
photoreceptors in humans. Most of our vision, however, comes from cones that
work under most light conditions and are responsible for acute detail and
color vision.
The human eye contains three types of cones (red, green and blue), each
sensitive to a different range of colors. Because their sensitivities overlap,
cones work in combination to convey information about all visible colors. You
might be surprised to know that we can see thousands of colors using only
three types of cones, but computer monitors use a similar process to generate
a spectrum of colors. The central part of the human retina, where light is
focused, is called the fovea, which contains only red and green cones. The
area around the fovea, called the macula, is critical for reading and driving.
Death of photoreceptors in the macula, called macular degeneration, is a
leading cause of blindness among the elderly population in developed
countries, including the United States.
The retina contains three organized layers of neurons. The rod and cone
photoreceptors in the first layer send signals to the middle layer
(interneurons), which then relays signals to the third layer, consisting of
multiple different types of ganglion cells, specialized neurons near the inner
surface of the retina. The axons of the ganglion cells form the optic nerve.
Each neuron in the middle and third layer typically receives input from many
cells in the previous layer, and the number of inputs varies widely across the
retina.
Near the center of the gaze, where visual acuity is highest, each ganglion
cell receives inputs — via the middle layer — from one cone or, at most, a
few, allowing us to resolve very fine details. Near the margins of the retina,
each ganglion cell receives signals from many rods and cones, explaining why
we cannot see fine details on either side. Whether large or small, the region
of visual space providing input to a visual neuron is called its receptive
field.
### CONTENT PROVIDED BY
BrainFacts/SfN
Related Topics Vision Cells & Circuits Anatomy Thinking, Sensing &
Behaving Brain Anatomy & Function
### Also In Vision
Perceptual Disorders: Alice in Wonderland Syndrome and Prosopagnosia
Vision Word Search
Beyond the Rainbow: The Incredible Visual World of Fishes
How ‘One Little Pink Dot’ Marked the Spot of the Brain’s Face-Detecting Center
Baby Mantis Shrimp Perceive UV Light Like Their Parents
Restoring Vision in Older Adults Can Reduce Risk of Dementia
### Trending
Popular articles on BrainFacts.org
1. Depression’s Impact on Memory
2. ICYMI: COVID-19 Linked to Blood Clots and Strokes
3. The Neuron
4. What Part of the Brain Deals With Anxiety? What Can Brains Affected by Anxiety Tell us?
5. Proteins That Balance Our Moods
#### BrainFacts Book
Download a copy of the newest edition of the book, Brain Facts: A Primer on
the Brain and Nervous System.
Download
#### Brain Awareness Video Contest
Submit a short video about any neuroscience topic for a chance to win $4,000
and a trip to SfN's Annual Meeting!
Learn More
#### Best of BrainFacts Newsletter
Our editors' picks from this month's articles.
SIGN UP
* Like
* Subscribe
* Follow
* Follow
* Subscribe
* About
* Contact
* For Educators
* Donate
######
###### SUPPORTING PARTNERS
* *
* Disclaimer
* Privacy Policy
* Accessibility Policy
* Terms and Conditions
* Manage Cookies
Some pages on this website provide links that require Adobe Reader to view.
Copyright © Society for Neuroscience
| biology | 84004 | https://sv.wikipedia.org/wiki/Syn | Syn | Syn är den del av nervsystemet som gör det möjligt för en organism att se. Den räknas även till människans sinnen. Olika djur har olika synsystem och kan uppfatta ljus av olika våglängder. Människan kan uppfatta ljus mellan 380 och 750 nm vilket brukar kallas synligt ljus. Synen bygger på samverkan mellan en rad organ i kroppen.
För att ett skarpt seende skall komma till stånd fordras förutom bildens inträffande på näthinnan, ett klart medium, näthinnans, synnervens och hjärnans integritet, tillräcklig belysning, samt att avståndet till föremålet är tillräckligt stort. Det finns ca 130 miljoner synsinnesceller i ögonen.
Historia
1802 postulerade Thomas Young existensen av tre typer av fotoreceptorer i ögat som var känsliga för var och speciella våglängde i det synliga ljusspektrat. Hermann von Helmholtz utvecklade teorin ytterligare 1850 och menade att de tre typerna kunde klassificeras efter de tre färgerna RGB och att ljusstyrkan tolkades av hjärnan som synliga kulörer. Teorin bevisades över ett århundrade senare år 1964 när mätningar på en enda tappcell kunde göras med hjälp av mikrospektrofotopi.
Fysikern John Strutt, 3:e baron Rayleigh ställde upp kriterier för vilken vinkel som behövs för optisk upplösning vilken är beroende på ljusets våglängd.
Ögat
Ögat registrerar elektromagnetisk strålning som bryts av hornhinna och lins och som projiceras på näthinnan (retina) längst bak i ögat. Cellerna där är specialiserade fotoreceptorer och brukar delas in i stavar som registrerar ljusstyrka och tre typer av tappar som registrerar färgerna rött, blått och grönt. Att man inte ser färg i mörker beror på att de färgkänsliga tapparna kräver mycket ljus för att aktiveras. I mörker är det endast stavarna som skickar information till hjärnan och de kan inte "se" färg. Nerver som är fästa på de celler som är sammankopplade med fotoreceptorerna kommer samman till synnerven vid blinda fläcken som är den punkt i ögat där nerverna passerar ut till hjärnan och här finns inga sinnesceller. Den gula fläcken på näthinnan består av enbart tappar och det är på detta område som ljuset ska brytas för att man ska se skarpt med det direkta seendet, fovea. Resten av näthinnan står för det indirekta seendet, och har inte lika stark skärpa då det består av mestadels stavar. Stavarna är kopplade med flera celler vilket gör att upplösningen försämras, tapparna däremot kopplar endast med en cell och kan därför skicka högupplöst information till hjärnan. Ögat kontrolleras av en rad viljestyrda och icke viljestyrda muskler vilket gör att man kan vrida på ögonen och av en ringmuskel som drar ut linsen vilket justerar skärpan. Att man har två ögon ger ett djupseende som består i att hjärnan kan tolka de skillnader som finns i bilderna de olika ögonen förmedlar och gör att man lättare kan bedöma avstånd, speciellt till närliggande föremål.
Nervsystemet
Synnerven transporterar signaler från ögat till synnervskorsningen (chiasm), som är en korsning där synnerverna korsas. De passerar sedan genom optiska gångar till laterala knäkroppen (lateral geniculate nucleus), en mellanstation på väg mot syncentrum i bakre delen av hjärnbarken. Väl där tolkas synsignalerna.
Synfel
Brytningsfel
Närsynthet
Översynthet
Astigmatism
Ögonsjukdomar
Presbyopi (nedsatt ackommodationsförmåga).
Grå starr
Grön starr
Färgblindhet beror ofta på att någon av de tre olika typerna av tappar saknas och är ofta medfött.
Blindhet
Referenser
Noter
Externa länkar
På engelska Wikipedia finns två artiklar visual system (synsystemet) och visual perception(synuppfattningen) som behandlar synen från olika perspektiv.
Sinnesorgan | swedish | 0.545248 |
vision_of_lights/LIGHT,_EYE_AND_VISION.txt | WikiLectures WikiLectures
### Search
Log in ↓
Personal tools
* Not logged in
* Talk
* Contributions
* Log in
## Navigation menu
Navigation
* Help
* Create article
* Recent changes
* Contacts
* Support
Print/export
* Download as PDF
* Printable version
Namespaces
Page Discussion
Actions
Views
* Read
* Edit
* Edit source
* View history
Tools
Tools
* What links here
* Related changes
* Special pages
* Printable version
* Permanent link
* Page information
* Cite this page
# LIGHT, EYE AND VISION
From WikiLectures
×
Send
Thank you for your comments.
Thank you for reviewing this article.
Your review hasn't been inserted (one review per article per day allowed)!
Visible light is a kind of electromagnetic radiation, it is a narrow part of
the electromagnetic spectrum that is visible to the human eye. Light, like
other types of electromagnetic radiation, exhibits characteristics of both
waves and particles - this is referred to as the dual nature of light. Light
sometimes acts like particles, e.g. when comes into contact with matter. At
the same time light acts like electromagnetic waves - it undergoes diffraction
and interference just like a wave. Light propagation is affected by the
phenomena like refraction, reflection, diffraction and interference. As an
electromagnetic wave, light has both electric and magnetic field components,
which oscillate in a fixed relationship to one another, perpendicular to each
other and perpendicular to the direction of wave propagation. The
electromagnetic wave can be characterized by its frequency and wavelength.
Photon as the particle of light has no charge and always is moving at the
speed of light (has zero mass and rest energy). Its energy is related to the
frequency of the electromagnetic wave by the formula E = h∙f therefore, a
photon can be assumed as a discrete quantum of energy. Photons can be
destroyed (or created) when radiation is absorbed or emitted.
## Contents
* 1 Frequency, wavelength, energy and colour
* 2 Photometry and brightness
* 3 Ultraviolet and infrared light
* 4 Eye
* 5 Vision defects
* 6 Astigmatism
* 7 Cataract
* 8 Glaucoma
* 9 Colour-blindness
### Frequency, wavelength, energy and colour [ edit | edit source ]
The frequency, wavelength and energy of photons are elementary parameters of
the wave and they are related to each other. A typical human eye will respond
to wavelengths from 390 to 700 nm (in vacuum or air). In terms of frequency
this corresponds to a band in the vicinity of 430–790 THz. The energy ( E )
carried by a photon is given by the frequency: E = h . f, where h is the
Planck constant. The unit used for quantification of the amount of energy is
typically electron volt (eV), a unit of energy equal to approximately 1.6×10
−19 J.
Our perception of colour is based on perception of the light wavelength (or
combination of light wavelengths). A light-adapted eye generally has its
maximum sensitivity at around 555 nm (540 THz), in the green region of the
optical spectrum. The spectrum does not contain all the colours that the human
eye and brain can distinguish. Unsaturated colours such as variations of pink
or purple are absent because they can only be made by a mix of multiple
wavelengths. Colours containing only one wavelength are called pure colours.
### Photometry and brightness [ edit | edit source ]
Brightness is an attribute of visual perception, which appears when a source
radiates or reflects light. In other words, brightness is the perception
elicited by the luminance of a visual target. This is a subjective
attribute/property of an object being observed.
The human eye is not equally sensitive to all wavelengths of visible light.
The standardized model of the eyes response to light as a function of
wavelength is given by the luminosity function. Note that the eye has
different responses as a function of wavelength when it is adapted to light
conditions (photopic vision) and dark conditions (scotopic vision).
The luminosity describes the average spectral sensitivity of human visual
perception of brightness. It is based on a subjective judgement, in which a
pair of different coloured lights are brighter to describe relative
sensitivity to light of different wavelengths. It should not be considered
perfectly accurate in every case, but it is a very good representation of
visual sensitivity of the human eye and it is valuable as a baseline for
experimental purposes.
### Ultraviolet and infrared light [ edit | edit source ]
Ultraviolet (UV) light is electromagnetic radiation with a wavelength
shorter than visible light, but longer than X-rays. It is in the range between
400- 100 nm, corresponding to photon energies from 3 eV to 124 eV. It is named
ultraviolet because the spectrum consists of electromagnetic waves with
frequencies higher than those humans identify as the color violet. These
frequencies are invisible to humans, but wavelengths near UV are visible to
several insects and birds.
Of the ultraviolet radiation that reaches the Earth's surface up to 95% is
UVA. Ordinary window glass passes about 90% of the light above 350 nm, but
blocks over 90% of the light below 300 nm.
The shorter bands of UVC, as well as more energetic radiation produced by the
Sun, generate the ozone in the ozone layer when single oxygen atoms produced
by UV photolysis of dioxygen react with more dioxygen. The ozone layer is
important for blocking UVB and part of UVC. Although the shortest wavelengths
of UVC (and those even shorter) are blocked by ordinary air.
Infrared (IR) light is electromagnetic radiation with longer wavelengths
than those of visible light, extending from the nominal red edge of the
visible spectrum at 700 nanometres (nm) to 1 mm. This range of wavelengths
corresponds to a frequency range of approximately 430 THz down to 300 GHz.
Most of the thermal radiation emitted by objects near room temperature is
infrared.
Infrared radiation can be used to remotely determine the temperature of
objects (if the emissivity is known). This is termed thermography.
Thermographic cameras detect radiation in the infrared range of the
electromagnetic spectrum (roughly 900–14,000 nanometers or 0.9–14 μm) and
produce images of that radiation. Infrared radiation is emitted by all
objects, based on their temperature. The black body radiation law states that;
thermography makes it possible to "see" ones environment with or without
visible illumination. The amount of radiation emitted by an object increases
with temperature, therefore thermography allows us to see variations in
temperature.
### Eye [ edit | edit source ]
Eye is the organ of vision - it converts light into electrical signals
transmitted by neurons. The shape of the eye is a slightly asymmetrical globe,
whose diameter is about 25mm. The visible front part of the eye consists of
iris, cornea, pupil, sclera (the white part) and conjunctiva (a clear layer of
tissue covering the front of the eye, except the cornea).
The cornea is the front part of the eye. It is a transparent layer (does not
have blood vessels) with a thickness of about 0.6 mm. Cornea represents the
strongest part of the refracting power of the eye. It provides about 70% of
the power of the eyes optical system. The refractive power of the cornea is
about 40 dioptres. The index of refraction of the cornea is about 1.376 and
most of the refraction is at the cornea-air interface. Due to a constant
refractive power its focus is fixed.
The iris is a thin, circular structure in the eye responsible for
controlling the diameter and size of the pupil and the amount of light
reaching the retina. The color of the iris is often referred to as "eye
color." The iris is a contractile structure, consisting mainly of smooth
muscle surrounding the pupil. Light enters the eye through the pupil, and the
iris regulates the amount of light by controlling the size of the pupil
(similarly to the shutter in camera).
Just behind the iris and pupil lies the lens. The lens focuses light at the
back of the eye. About 9mm in diameter and 4 mm thick, the crystalline lens
provides perhaps 20% of the refracting power of the eye. The index ranges from
about 1.406 at the center to about 1.386 in outer layers making it a gradient
index lens. It is pliable, and changes shape for close focusing. This is known
as accommodation.
Most of the eye is filled with a clear gel called the vitreous. Light projects
through the pupil and the lens to the back of the eye. The inside lining of
the eye is covered by special light-sensing cells that are collectively called
the retina. The retina covers about 65 percent of the eyes interior surface.
Photosensitive cells called rods and cones in the retina convert light energy
into electrical signals, that are carried to the brain by the optic nerve. In
the middle of the retina is a small dimple called the fovea or fovea
centralis. This is the center of the eyes sharpest vision and the location of
most color perception.
The optic nerve is a bundle of nerve fibers that carries the electrical signal
from the retina to the brain for processing. The point where the optic nerve
exits through the retina does not have any rods or cones, and thus produces a
" blind spot ".
Though the eye receives data from a field of about 200 degrees, the acuity
over most of that range is poor. To form high resolution images, the light
must fall on the fovea, and that limits the acute vision angle to about 15
degrees. In low light, this fovea constitutes a second blind spot since it is
exclusively cones which have low light sensitivity. At night, to get most
acute vision one must shift the vision slightly to one side, say 4 to 12
degrees so that the light falls on some rods.
### Vision defects [ edit | edit source ]
Common eye defects like myopia (nearsightedness), hyperopia (farsightedness)
and astigmatism are often called "refractive errors". They affect how the eyes
bend or "refract" light and they can usually be corrected by relatively simple
compensating lenses or by surgery changing the shape of the cornea in order to
improve the refractive state of the eye. Reshaping of the cornea is frequently
performed by photoablation using laser.
Myopia (Nearsightedness)
Myopia, or nearsightedness, is a vision condition in which close objects are
seen clearly, but objects farther away appear blurred. Nearsightedness occurs
if the eyeball is too long (axial myopia) or the cornea has too much curvature
(refractive myopia). As a result, the light entering the eye is not focused
correctly and distant objects look blurred.
Nearsightedness is a very common vision condition affecting nearly 30 percent
of the US and EU population. Some research supports the theory that
nearsightedness is hereditary. There is also growing evidence that it is
influenced by the visual stress of too much close work.
[[|framed|center|
http://www.wikilectures.eu/index.php/File:4.2.5lightineye.png ]]
Generally, nearsightedness first occurs in school-age children. Because the
eye continues to grow during childhood, it typically progresses until about
the age of 20. However, nearsightedness may also develop in adults due to
visual stress or health conditions such as diabetes.
An optometrist can prescribe diverging eyeglasses or contact lenses. This will
correct the nearsightedness by bending the visual images that enter the eyes
and by focusing the images correctly at the back of the eye. Depending on the
severity of nearsightedness, the subject may only need to wear glasses or
contact lenses for certain activities, such as watching a movie or driving a
car. If the person is severely nearsighted, lenses and glasses may need to be
worn all the time. Laser procedures are also a possible treatment for
nearsightedness in adults. They involve reshaping the cornea by removing a
small amount of eye tissue. This is accomplished by using a highly focused
laser beam on the surface of the eye.
Hyperopia (Farsightedness)
Farsightedness or hyperopia, is a vision condition in which distant objects
are seen clearly, but close objects does not come into proper focus.
Farsightedness occurs if the eyeball is too short or the cornea has too little
curvature. Light entering the eye is therefore not focused correctly. Common
signs of farsightedness include difficulty in concentrating and maintaining a
clear focus on near objects. Also eye strain, fatigue and/or headaches after
close work. Aching or burning of the eyes and irritability or nervousness
after sustained concentration can also occur.
In case of farsightedness, vision can be corrected by converging eyeglasses or
contact lenses - to optically correct farsightedness by altering the way the
light enters eyes. Laser procedure is also a possible treatment similar to
that of nearsightedness.
Presbyopia is a vision condition in which the crystalline lens of your eye
loses its flexibility. This makes it difficult for you to focus on close
objects. Presbyopia may seem to occur suddenly, but the actual loss of
flexibility takes place over a number of years. It usually becomes noticeable
in the early to mid-40s and is a natural part of the aging process of the eye.
It is not a disease, and cannot be prevented. Some signs of presbyopia include
the tendency to hold reading materials at arm's length, blurred vision at
normal reading distance and eye fatigue, along with headaches when doing close
work. A comprehensive optometric examination will include testing for
presbyopia.
### Astigmatism [ edit | edit source ]
Astigmatism is a vision condition that causes blurred vision due to the
irregular shape of the cornea or the curvature of the lens inside the eye. An
irregular shaped cornea or lens prevents light from focusing properly on the
retina. As a result, vision becomes blurred at any distance.
Astigmatism is a very common vision condition and most people have some degree
of it. Slight amounts of astigmatism usually don't affect vision and don't
require treatment. However, larger amounts causes distorted or blurred vision,
eye discomfort and headaches.
Astigmatism frequently occurs with other vision conditions like near-
sightedness (myopia) and farsightedness (hyperopia). Compensation for
astigmatism can be made by eyeglasses with cylindrical lens (i.e. a lens that
has different radii of curvature in different planes), contact lenses or
refractive surgery.
### Cataract [ edit | edit source ]
Cataract is a condition that occurs when the inner lens of the eye becomes
darkened or opaque. The lens may be surgically replaced with a plastic lens.
This can have dramatic results in restoring vision to the eye. The implanted
lens is of fixed focal length, meaning it is not capable of accommodation like
the natural lens. This is usually not a major concern, because persons that
develop cataracts after the age of 60 do not have much accommodation remaining
anyways. This is due to the inner lens turning less pliable with age.
### Glaucoma [ edit | edit source ]
Glaucoma is a disease which damage the eyes optic nerve and can result in
vision loss and blindness. People with glaucoma will slowly lose their
peripheral (side) vision. If glaucoma remains untreated, people may not notice
objects to the side and out of the corner of their eye. They seem to be
looking through a tunnel. Over time, straight-ahead (central) vision may
decrease until no vision remains. The vision lost from the disease cannot be
restored.
### Colour-blindness [ edit | edit source ]
Colour blindness is the inability to see colour, or the decreased ability to
perceive colour differences under normal lighting conditions. Colour blindness
affects a significant percentage of the population. There is no actual
blindness but there is a deficiency of colour vision. The most usual cause, is
a fault in the development of one or more sets of retinal cones that perceive
colour in light and transmit that information to the optic nerve.
Colour blindness can also be produced by physical or chemical damage to the
eye, the optic nerve, or parts of the brain.
Retrieved from "
https://www.wikilectures.eu/index.php?title=LIGHT,_EYE_AND_VISION&oldid=92190
"
Categories :
* Biophysics
* Ophthalmology
* *
* This page was last edited on 26 April 2024, at 21:46.
* WikiLectures, project of the First Faculty of Medicine, Charles University , as the contribution to the MEFANET project. • ISSN 1804-9885 • Content is available under Creative Commons Attribution-ShareAlike 4.0 license, additional terms may apply. See Author Law , Terms of use and General Disclaimer for details. Created with the support of OP VVV project CZ.02.2.69/0.0/0.0/16_015/0002362.
* Privacy policy
* About WikiLectures
* Disclaimers
* Developers
* Statistics
* Cookie statement
| biology | 66706 | https://sv.wikipedia.org/wiki/Optik | Optik | Optik är läran om ljusets utbredning och brytning (från grek. optikē) och är en gren av fysiken inom vilken förklaringar ges till olika optiska fenomen. Under det tidiga 1000-talet, skrev Alhazen (Ibn al-Haytham) Boken om optik (Kitab al-manazir) i vilken han undersökte reflexion och refraktion och föreslog ett nytt system för att förklara synlighet och ljus grundat på observationer och experiment.
Optiken brukar vanligen beskrivas som det synliga, det infraröda och det ultravioletta ljusets uppträdande. Men eftersom ljuset är en elektromagnetisk vågrörelse, uppträder liknande fenomen med röntgenstrålning, mikrovågor, radiovågor och andra former av elektromagnetisk strålning. Optiken kan således betraktas som en underavdelning till elektromagnetism. Vissa optiska fenomen kan bara förklaras genom ljusets kvantegenskaper (till exempel den fotoelektriska effekten). Optiken spänner därför över både klassisk fysik och kvantfysik.
Optiken är emellertid ett fält som ofta anses till största delen skilt från fysikens område. Optiken har sin egen identitet, sina egna föreningar och konferenser. De rent vetenskapliga aspekterna av området kallas optisk vetenskap eller optisk fysik. Tillämpad optisk vetenskap benämns optisk ingenjörskonst. Tillämpningar av optisk ingenjörskonst som hör ihop speciellt med belysning kallas för belysningsteknik. Vart och ett av dessa områden tenderar att vara mycket olika i sina tillämpningar, teknisk skicklighet, inriktning och yrkestillhörighet.
Tack vare den breda praktiska tillämpningen av ljusvetenskapen, har den optiska vetenskapen och den optiska ingenjörskunskapen kommit att bli mycket tvärvetenskapliga områden. Optisk vetenskap ingår nu som en del i många närliggande discipliner vilka innefattar elektrisk ingenjörskonst, fysik, psykologi, medicin och andra.
Klassisk optik
Klassisk eller geometrisk optik, ibland kallad för strålgångsoptik beskriver ljusets utbredning i form av strålar. Strålar böjs vid interferens mellan två olika media och kan böjas i ett medium där refraktionsindex är beroende av det fysiska läget. Strålen i geometrisk optik är vinkelrät mot vågfronten i fysisk optik.
Reflexion
Refraktion
Diffraktion
Dispersion
Polarisering
Koherens
Spridning
Fermats princip
Geometrisk optik:
Speglar
Prismaoptik
Modern optik
Modern optik är en term som används för att beskriva de områden av optisk vetenskap och ingenjörskonst som blev populära under 1900-talet. Dessa områden av optisk vetenskap beskriver oftast ljusets elektromagnetiska egenskaper eller ljusets kvantumegenskaper men innehåller också andra ämnesområden.
Laser
Holografi
Radiometri
Fotometri
Optometri
Optometri är läran om synfel, korrektionsglas och synhjälpmedel. Enligt svensk myndighetsdefinition (se t.ex. Karolinska Institutets huvudområdesdefinition av optometri utgörs optometri av ”kunskaper om människans ögon, syn, synsystem och synprocesser/perception, samt hur inre och yttre faktorer påverkar synen.
Optiker, ortoptister & optometritekniker är tre yrken inom ämnet optometri.
Andra optiska områden
Bildbehandling
Fotografi
Informationsteori
Se även
Lins
Foton
Spektroskopi
Geometrisk optik
Våglängd
Aberration
Partikel-våg dualism
Referenser
Noter
Externa länkar | swedish | 0.519966 |
eye_capture_shape/Peripheral_vision.txt | Peripheral vision, or indirect vision, is vision as it occurs outside the point of fixation, i.e. away from the center of gaze or, when viewed at large angles, in (or out of) the "corner of one's eye". The vast majority of the area in the visual field is included in the notion of peripheral vision. "Far peripheral" vision refers to the area at the edges of the visual field, "mid-peripheral" vision refers to medium eccentricities, and "near-peripheral", sometimes referred to as "para-central" vision, exists adjacent to the center of gaze.
Boundaries[edit]
Inner boundaries[edit]
The inner boundaries of peripheral vision can be defined in any of several ways depending on the context. In everyday language the term "peripheral vision" is often used to refer to what in technical usage would be called "far peripheral vision." This is vision outside of the range of stereoscopic vision. It can be conceived as bounded at the center by a circle 60° in radius or 120° in diameter, centered around the fixation point, i.e., the point at which one's gaze is directed. However, in common usage, peripheral vision may also refer to the area outside a circle 30° in radius or 60° in diameter. In vision-related fields such as physiology, ophthalmology, optometry, or vision science in general, the inner boundaries of peripheral vision are defined more narrowly in terms of one of several anatomical regions of the central retina, in particular the fovea and the macula.
The fovea is a cone-shaped depression in the central retina measuring 1.5 mm in diameter, corresponding to 5° of the visual field. The outer boundaries of the fovea are visible under a microscope, or with microscopic imaging technology such as OCT or microscopic MRI. When viewed through the pupil, as in an eye exam (using an ophthalmoscope or retinal photography), only the central portion of the fovea may be visible. Anatomists refer to this as the clinical fovea, and say that it corresponds to the anatomical foveola, a structure with a diameter of 0.35 mm corresponding to 1 degree of the visual field. In clinical usage the central part of the fovea is typically referred to simply as the fovea.
In terms of visual acuity, "foveal vision" may be defined as vision using the part of the retina in which a visual acuity of at least 20/20 (6/6 metric or 0.0 LogMAR; internationally 1.0) is attained. This corresponds to using the foveal avascular zone (FAZ) with a diameter of 0.5 mm representing 1.5° of the visual field. Although often idealized as perfect circles, the central structures of the retina tend to be irregular ovals. Thus, foveal vision may also be defined as the central 1.5–2° of the visual field. Vision within the fovea is generally called central vision, while vision outside of the fovea, or even outside the foveola, is called peripheral, or indirect vision.
A ring-shaped region surrounding the fovea, known as the parafovea, is sometimes taken to represent an intermediate form of vision called paracentral vision. The parafovea has an outer diameter of 2.5 mm representing 8° of the visual field.
The macula, the next larger region of the retina, is defined as having at least two layers of ganglia (bundles of nerves and neurons) and is sometimes taken as defining the boundaries of central vs. peripheral vision (but this is controversial). Estimates of the macula’s size differ, its diameter estimated at 6° – 10° (corresponding to 1.7 – 2.9 mm), up to 17° of the visual field (5.5 mm). The term is familiar in the general public through the widespread macular degeneration (AMD) at older age, where central vision is lost.
When viewed from the pupil, as in an eye exam, only the central portion of the macula may be visible. Known to anatomists as the clinical macula (and in clinical setting as simply the macula) this inner region is thought to correspond to the anatomical fovea.
A dividing line between near and mid peripheral vision at 30° radius can be based on several features of visual performance.
Visual acuity declines systematically up to 30° eccentricity: At 2°, acuity is half the foveal value, at 4° one-third, at 6° one-fourth etc. At 30°, it is one-sixteenth the foveal value. From thereon the decline is steeper. (Note that it would be wrong to say, the value were halved every 2°, as said in some textbooks or in previous versions of this article.)
Color perception is strong at 20° but weak at 40°. In dark-adapted vision, light sensitivity corresponds to rod density, which peaks just at 18°. From 18° towards the center, rod density declines rapidly. From 18° away from the center, rod density declines more gradually, in a curve with distinct inflection points resulting in two humps. The outer edge of the second hump is at about 30°, and corresponds to the outer edge of good night vision.
Outer boundaries[edit]
Classical image of the shape and size of the visual field
The outer boundaries of peripheral vision correspond to the boundaries of the visual field as a whole. For a single eye, the extent of the visual field can be (roughly) defined in terms of four angles, each measured from the fixation point, i.e., the point at which one's gaze is directed. These angles, representing four cardinal directions, are 60° upwards, 60° nasally (towards the nose), 70–75° downwards, and 100–110° temporally (away from the nose and towards the temple). For both eyes the combined visual field is 130–135° vertically and 200–220° horizontally.
Characteristics[edit]
The loss of peripheral vision while retaining central vision is known as tunnel vision, and the loss of central vision while retaining peripheral vision is known as central scotoma.
Peripheral vision is weak in humans, especially at distinguishing detail, color, and shape. This is because the density of receptor and ganglion cells in the retina is greater at the center and lowest at the edges, and, moreover, the representation in the visual cortex is much smaller than that of the fovea (see visual system for an explanation of these concepts). The distribution of receptor cells across the retina is different between the two main types, rod cells and cone cells. Rod cells are unable to distinguish color and peak in density in the near periphery (at 18° eccentricity), while cone cell density is highest in the very center, the fovea. Note that this does not mean that there is a lack of cones representing in the periphery; colors can be distinguished in peripheral vision.
Flicker fusion thresholds decline towards the periphery, but do that at a lower rate than other visual functions; so the periphery has a relative advantage at noticing flicker. Peripheral vision is also relatively good at detecting motion (a feature of Magno cells).
Central vision is relatively weak in the dark (scotopic vision) since cone cells lack sensitivity at low light levels. Rod cells, which are concentrated further away from the fovea, operate better than cone cells in low light. This makes peripheral vision useful for detecting faint light sources at night (like faint stars). Because of this, pilots are taught to use peripheral vision to scan for aircraft at night.
Ovals A, B and C show which portions of the chess situation chess masters can reproduce correctly with their peripheral vision. Lines show path of foveal fixation during 5 seconds when the task is to memorize the situation as correctly as possible. Image from based on data by
The distinctions between foveal (sometimes also called central) and peripheral vision are reflected in subtle physiological and anatomical differences in the visual cortex. Different visual areas contribute to the processing of visual information coming from different parts of the visual field, and a complex of visual areas located along the banks of the interhemispheric fissure (a deep groove that separates the two brain hemispheres) has been linked to peripheral vision. It has been suggested that these areas are important for fast reactions to visual stimuli in the periphery, and monitoring body position relative to gravity.
Functions[edit]
The main functions of peripheral vision are:
recognition of well-known structures and forms with no need to focus by the foveal line of sight
identification of similar forms and movements (Gestalt psychology laws)
delivery of sensations which form the background of detailed visual perception
Extreme peripheral vision[edit]
Side-view of the human eye, viewed approximately 90° temporal, illustrating how the iris and pupil appear rotated towards the viewer due to the optical properties of the cornea and the aqueous humor.
When viewed at large angles, the iris and pupil appear to be rotated toward the viewer due to the optical refraction in the cornea. As a result, the pupil may still be visible at angles greater than 90°.
Cone-rich rim of the retina[edit]
The rim of the retina contains a large concentration of cone cells. The retina extends farthest in the superior-nasal 45° quadrant (in the direction from the pupil to the bridge of the nose) with the greatest extent of the visual field in the opposite direction, the inferior temporal 45° quadrant (from the pupil of either eye towards the bottom of the nearest ear). Vision at this extreme part of the visual field is thought to be possibly concerned with threat detection, measuring optical flow, color constancy, or circadian rhythm.
See also[edit]
Averted vision
Bitemporal hemianopsia
Depth perception
Eye movement
Eye movement in music reading
Fovea
Perimetry
Visual field
Visual perception
Tunnel vision
Binocular vision
Macular degeneration
Glaucoma | biology | 1461639 | https://sv.wikipedia.org/wiki/Stereopsis | Stereopsis | Stereopsis (från stereo- "fast kropp" eller "tredimensionell", och opsis som betyder vy eller syn) är den process inom visual perception som skapar ett djupseende utifrån de två olika vyer som projiceras på näthinnan i ett mänskligt ögonpar och som skiljer sig lite från varandra. Skillnaden mellan de två näthinnebilderna kallas ”horisontell skillnad”, ”retinal disparitet”, eller binokulär disparitet. Skillnaderna uppstår på grund av ögonens olika placering i huvudet. Stereopsis brukar allmänt refereras till som djupseende. Detta är inte helt korrekt, eftersom djupseende beror på många fler monokulära faktorer än stereoskopiska, och individer med bara ett fungerande öga har en hyfsat bra, främst erfarenhetsmässig, förmåga till uppfattning om djup utom i konstruerade fall (som stereoskopiska bilder), där förmågan till stereoseende gör att föremålet kan urskiljas ur de båda tvådimensionella bilderna.
Historia
Stereopsis beskrev första gången av Charles Wheatstone 1838. ”… medvetandet uppfattar ett objekt i tre dimensioner via två olika bilder som projiceras på de två näthinnorna ...”. Han insåg att eftersom varje öga ser den visuella världen från en lite annorlunda horisontell position, så avviker bilderna i de två ögonen lite från varandra. Föremål på olika avstånd från ögonen ger bilder i de två ögonen som skiljer sig åt i den horisontella positionen, vilket ger en ”horisontell skillnad”, också känd som "retinal disparitet" och binokulär disparitet. Wheatstone visade att detta var ett effektivt sätt att ge djupverkan genom att skapa illusionen om djup i platta bilder som skilde sig åt endast i den horisontella positionen. För att visa bilderna för de två ögonen separat, uppfann Wheatstone stereoskopet.
Leonardo da Vinci hade också insett att föremål som befann sig på olika avstånd från ögonen gav bilder som avvek i sin horisontella position, men han hade bara dragit slutsatsen att detta gjorde det omöjligt för en målare att ge en realistisk avbildning av djupet i ett landskap i en enda kanvasduk. Leonardo valde som sitt närliggande objekt en pelare med cirkulärt tvärsnitt och för det avlägsna föremålet en platt vägg. Hade han valt något annat närliggande objekt, hade han kunnat upptäcka den horisontella dispariteten. Hans pelare var ett av de få föremål som skapar en identisk bild av sig självt i de båda ögonen.
Stereopsis blev populärt under Viktoriansk tid genom uppfinningen av prismastereoskopet av David Brewster. Detta, i kombination med fotografin, betydde att tiotusentals stereogram kom att produceras.
Se även
Ortoptist
Binokulär syn
Bibliografi
Julesz, B. (1971). Foundations of cyclopean perception. Chicago: University of Chicago Press
Scott B. Steinman, Barbara A. Steinman and Ralph Philip Garzia. (2000). Foundations of Binocular Vision: A Clinical perspective. McGraw-Hill Medical. .
Cabani, I. (2007). Segmentation et mise en correspondance couleur - Application: étude et conception d'un système de stéréovision couleur pour l'aide à la conduite automobile.
Referenser
Noter
Externa länkar
Middlebury Stereo Vision Page
VIP Laparoscopic / Endoscopic Video Dataset (Stereo Medical Images)
What is Stereo Vision?
Learn about Stereograms then make your own Magic Eye
International Orthoptic Association
Stereoskopi
Syn
es:Visión estereoscópica | swedish | 0.586146 |
eye_capture_shape/Determine-Eye-Shape.txt | Skip to Content
Quizzes
* PRO
Courses Guides New Tech Help Pro Expert Videos About wikiHow Pro Upgrade
Sign In
* QUIZZES
* EDIT
Edit this Article
* EXPLORE
Tech Help Pro About Us Random Article Quizzes
Request a New Article Community Dashboard This Or That Game
Popular Categories
Arts and Entertainment Artwork Books Movies
Computers and Electronics Computers Phone Skills Technology Hacks
Health Men's Health Mental Health Women's Health
Relationships Dating Love Relationship Issues
Hobbies and Crafts Crafts Drawing Games
Education & Communication Communication Skills Personal Development
Studying
Personal Care and Style Fashion Hair Care Personal Hygiene
Youth Personal Care School Stuff Dating
All Categories
Arts and Entertainment Finance and Business Home and Garden Relationship
Quizzes
Cars & Other Vehicles Food and Entertaining Personal Care and Style Sports
and Fitness
Computers and Electronics Health Pets and Animals Travel
Education & Communication Hobbies and Crafts Philosophy and Religion Work
World
Family Life Holidays and Traditions Relationships Youth
* LOG IN
### Log in
Social login does not work in incognito and private browsers. Please log in
with your username or email to continue.
Facebook
Google
wikiHow Account
No account yet? Create an account
* RANDOM
* Home
* Random
* Browse Articles
* Learn Something New
* Guides
* Courses
* Quizzes Hot
* Play Games
* This Or That Game New
* Train Your Brain
* Explore More
* Support wikiHow
* About wikiHow
* Log in / Sign up
Terms of Use
wikiHow is where trusted research and expert knowledge come together. Learn
why people trust wikiHow
* Categories
* Personal Care and Style
* Makeup
* Eye Makeup
# How to Determine Eye Shape
Download Article
Explore this Article
methods
1 Checking for Different Characteristics
2 Applying Makeup for Your Eye Shape
Other Sections
Expert Q&A
Video WATCH NOW
Tips and Warnings
Related Articles
References
Article Summary
Co-authored by Alana Schober
Last Updated: April 11, 2024 Fact Checked
Download Article
ARTICLE
VIDEO
X
This article was co-authored by Alana Schober . Alana Schober is a
Professional Makeup Artist at Pout by Lipsticknick, an inclusive beauty bar in
Los Angeles, California. With over a decade of experience, Alana specializes
in hair and makeup. As a Licensed Comselotologist, she has worked for both MAC
Cosmetics and Paul Mitchell. Alana's celebrity clientele includes Iggy Azalea,
Solange, YG, and ASAP Nast. She has worked on campaigns with Nike, Adidas,
Morphe, Jaclyn Hill Cosmetics, and Quay. Alana has also worked on countless
music videos and photoshoots and her work has been featured on two billboards
in Los Angeles.
There are 22 references cited in this article, which can be found at the
bottom of the page.
This article has been fact-checked, ensuring the accuracy of any cited facts
and confirming the authority of its sources.
This article has been viewed 1,340,021 times.
It’s pretty easy to determine your eye shape—just find a mirror and a few
spare minutes! Look for different characteristics to determine if your eyes
are creased, hooded, upturned or downturned, round or almond, close- or wide-
set, prominent or deep-set. All eye shapes are beautiful, and once you know
yours, you’ll be able to accentuate the natural shape of your eyes with the
right kind of eye makeup .
##
Steps
Method 1
Method 1 of 2:
### Checking for Different Characteristics
Download Article
1. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/3\/3b\/Determine-Eye-Shape-Step-1-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-1-Version-2.jpg","bigUrl":"\/images\/thumb\/3\/3b\/Determine-Eye-Shape-Step-1-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-1-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
1
Note that you have monolid eyes if your eyelid doesn’t have a crease. Check
your eye in the mirror if you’re not sure. Look for a crease in the center of
your upper eyelid . If you don't have a crease, you have monolid eyes. You
may also have flatter eyelids and less prominent brow bones than people with
creases in their lids. [1] X Research source
* Monolid eyes are most common in people of Asian descent.
2. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/6\/69\/Determine-Eye-Shape-Step-2-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-2-Version-2.jpg","bigUrl":"\/images\/thumb\/6\/69\/Determine-Eye-Shape-Step-2-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-2-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
2
Figure out if you have hooded eyes by examining the crease, if you have one.
In hooded eyes, skin hangs down over the crease, making your upper eyelid look
smaller. If you can’t see the crease in your eye when your eyes are open, then
you have hooded eyes. [2] X Research source
* Many people are born with hooded eyes, and also people’s eyes often become hooded as they age. [3] X Research source
Advertisement
3. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/9\/9b\/Determine-Eye-Shape-Step-3-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-3-Version-2.jpg","bigUrl":"\/images\/thumb\/9\/9b\/Determine-Eye-Shape-Step-3-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-3-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
3
Study the tilt of your eyes to figure out if they are upturned or downturned.
Imagine that there is a straight, horizontal line extending through the
centers of both eyes. Ask yourself if the outer corners of your eyes lie above
or below this center line. [4] X Research source
* If the outer corners are above this line, you have "upturned" eyes.
* If the outer corners are below this line, you have "downturned" eyes.
4. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/e\/ee\/Determine-Eye-Shape-Step-4-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-4-Version-2.jpg","bigUrl":"\/images\/thumb\/e\/ee\/Determine-Eye-Shape-Step-4-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-4-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
4
Note that you have almond eyes if you can’t see white around the iris. Keep
your eyes relaxed as you look into the mirror. In almond eyes, both the top
and bottom of the iris and slightly covered by the eyelid. Almond eyes tend to
be oval shaped with narrow corners. [5] X Research source
* The outside corners of your eyes might turn up slightly. [6] X Research source
5. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/2\/28\/Determine-Eye-Shape-Step-5-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-5-Version-2.jpg","bigUrl":"\/images\/thumb\/2\/28\/Determine-Eye-Shape-Step-5-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-5-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
5
Recognize that you have round eyes if you can see white below your iris.
Look straight ahead at a mirror. If you can see any white around the top or
bottom of your iris in this position, you have "round" eyes. Round eyes are
less tapered than almond eyes and appear more open. [7] X Research source
* There doesn’t have to be a lot of white under your iris, even a thin sliver counts as round eyes.
6. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/2\/2d\/Determine-Eye-Shape-Step-6-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-6-Version-2.jpg","bigUrl":"\/images\/thumb\/2\/2d\/Determine-Eye-Shape-Step-6-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-6-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
6
Measure the gap between your eyes to determine if they are wide- or close-set.
Measure your eye with your thumb and index finger, and then hold that same
space between your eyes. If the space between your eye is less than one eye
length in size, you have close-set eyes, but if the gap is larger than one eye
length, you have wide-set eyes. [8] X Research source
* If the gap between your eyes is the same width as your eyes, then you have an average spacing.
7. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/c\/ca\/Determine-Eye-Shape-Step-7-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-7-Version-2.jpg","bigUrl":"\/images\/thumb\/c\/ca\/Determine-Eye-Shape-Step-7-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-7-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
7
Examine the depth of your eyes to determine if they are deep-set or
prominent. Deep-set eyes are tucked further back into the socket, causing the
upper eyelid to appear short and small. Protruding eyes, on the other hand,
stick outward from the socket and toward the upper lash line. [9] X
Research source
* Monolid eyes are usually not deep-set.
* In deep-set eyes, it might look like your brow bone is bigger, simply because your eyes are further back. [10] X Research source
8. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/6\/6a\/Determine-Eye-Shape-Step-8-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-8-Version-2.jpg","bigUrl":"\/images\/thumb\/6\/6a\/Determine-Eye-Shape-Step-8-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-8-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
8
Compare your eyes with your nose and mouth size to figure out your eye size.
Eyes that are "average" in size are similar to that of your mouth or nose, if
not a little smaller. If your eyes are significantly smaller, though, you have
small eyes . If they are larger than your other features, you have large
eyes. [11] X Research source
* Many people have one eye that’s slightly larger than the other, which is totally normal.
Advertisement
Method 2
Method 2 of 2:
### Applying Makeup for Your Eye Shape
Download Article
1. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/f\/fd\/Determine-Eye-Shape-Step-9-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-9-Version-2.jpg","bigUrl":"\/images\/thumb\/f\/fd\/Determine-Eye-Shape-Step-9-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-9-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
1
Create a gradient of eye-shadow for monolid eyes. Prep your eyes with
eyeshadow primer by rubbing it over the top lids. Brush on a dark color
close to the lash line, a soft neutral toward the middle, and a shiny color
near the brow. Put your eyeshadow on with your eyes still open, so you can see
how it will look. [12] X Research source
* Winged or cat-eye eyeliner is also a great look for monolid eyes.
* You can also create floating eyeliner by drawing the line at your lid line, rather than your lash line. [13] X Research source
2. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/b\/bc\/Determine-Eye-Shape-Step-10-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-10-Version-2.jpg","bigUrl":"\/images\/thumb\/b\/bc\/Determine-Eye-Shape-Step-10-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-10-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
2
Extend eye shadow beyond the crease for hooded eyes . Before you put on
your eyeshadow, prime your eyelids with eyeshadow primer and or a light
concealer. Then, using a medium shade, brush the shadow on from your lashes up
onto the hooded part of your eye, beyond the crease. Elongate the eyeshadow in
the outer corner to create an eyeshadow wing to open up the appearance of your
eye. [14] X Research source
* Tightline your eyes with a kohl eyeliner to make your upper eyelashes look thicker.
* Wing-tipped eyeliner doesn’t always work on hooded eyes, because the flicked part can be hidden by the hood. [15] X Research source Try keeping the wing really thin, and do the lid first. [16] X Expert Source Alana Schober
Pout Pro Makeup Artist Expert Interview. 15 June 2021.
3. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/c\/c5\/Determine-Eye-Shape-Step-11-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-11-Version-2.jpg","bigUrl":"\/images\/thumb\/c\/c5\/Determine-Eye-Shape-Step-11-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-11-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
3
Emphasize the tilt of upturned eyes with smokey eyeshadow . Apply an
eyeshadow primer first, if you want this look to last all day. Then apply a
light shade of eyeliner to the inner half of your eyelid and a medium-tinted
eyeshadow to the outer half. Blend the two shades together slightly in the
middle, and you’ll have your smokey look. [17] X Research source
* Use a dark eyeliner and line the outer corners of your eyes, both the top and bottom.
4. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/6\/66\/Determine-Eye-Shape-Step-12-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-12-Version-2.jpg","bigUrl":"\/images\/thumb\/6\/66\/Determine-Eye-Shape-Step-12-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-12-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
4
Try a cat-eye look for downturned eyes. You can do many different eyeliner
techniques with a downturned eye, but a cat eye will enhance the outer corner
of your eye. Line the entire upper lashline with a dark eyeliner, and diffuse
it upwards at the outer corner. You don’t need to line your bottom lashes.
[18] X Research source
* Many different eyeshadow techniques look good with cat-eye liner, so experiment to see what you like.
5. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/d\/dc\/Determine-Eye-Shape-Step-13-Version-2.jpg\/v4-460px-Determine-Eye-Shape-Step-13-Version-2.jpg","bigUrl":"\/images\/thumb\/d\/dc\/Determine-Eye-Shape-Step-13-Version-2.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-13-Version-2.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
5
Add length to round, small, or deep-set eyes with a gradient of eyeshadow.
Place the lightest shade of eyeliner closest to the inner corner, and darken
the shadow as you move outwards. Play up the corners of your eyeliner by
pulling it slightly out and up. Smudge the eyeliner a bit if you want the look
to seem a little softer. [19] X Research source
* Use a clean brush to blend the different shades together to create a smooth, cohesive look.
* Apply mascara to all your upper lashes, or focus on the outer corner.
6. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/4\/4e\/Determine-Eye-Shape-Step-14.jpg\/v4-460px-Determine-Eye-Shape-Step-14.jpg","bigUrl":"\/images\/thumb\/4\/4e\/Determine-Eye-Shape-Step-14.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-14.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
6
Extend close-set eyes with smokey shadow to emphasize the outer corners.
Apply light eyeshadow over the entire eyelid, then add medium shadow to the
outer third. Add dark eyeshadow to the outer corner to create a dark shape
that lifts at the corner. Line the outer corner with eyeliner and put a little
bit of mascara in the outer corner. [20] X Research source
* Any dark color on the inner part of your eyes will make them seem closer together.
7. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/8\/88\/Determine-Eye-Shape-Step-15.jpg\/v4-460px-Determine-Eye-Shape-Step-15.jpg","bigUrl":"\/images\/thumb\/8\/88\/Determine-Eye-Shape-Step-15.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-15.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
7
Try a halo look for almond eyes. Almond eyes pair well with many different
eye makeup formations, but a halo look will help mix things up a bit. First,
apply a light transition color to the crease of your eye. Then, put a mid-
toned color on the inner and outer thirds, and apply a light color to the
center of the lid. Repeat this process on the lower lash line and highlight
the inner corner. [21] X Research source
* Add liner and lashes for a more defined look.
8. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/4\/4d\/Determine-Eye-Shape-Step-16.jpg\/v4-460px-Determine-Eye-Shape-Step-16.jpg","bigUrl":"\/images\/thumb\/4\/4d\/Determine-Eye-Shape-Step-16.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-16.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
8
Put dark eyeshadow and liner on the inner corners of wide set eyes. First,
apply your eyeshadow primer by rubbing it on with a finger over your upper
eyelid. Apply a darker shade closer to your tear duct and on the edge of your
nose, to create the illusion that your eyes are closer together. Then,
highlight the outer corner with a light shade of eyeshadow. Make sure you
blend the shades together. Apply eyeliner in a straight line, as thick as you
would like, with no wing. [22] X Research source
* You can also try extending your brows inwards by letting them grow out or filling them in with brow makeup. [23] X Research source
9. {"smallUrl":"https:\/\/www.wikihow.com\/images\/thumb\/5\/5c\/Determine-Eye-Shape-Step-17.jpg\/v4-460px-Determine-Eye-Shape-Step-17.jpg","bigUrl":"\/images\/thumb\/5\/5c\/Determine-Eye-Shape-Step-17.jpg\/aid4490530-v4-728px-Determine-Eye-Shape-Step-17.jpg","smallWidth":460,"smallHeight":345,"bigWidth":728,"bigHeight":546,"licensing":"<div class=\"mw-parser-output\"><p>\u00a9 2024 wikiHow, Inc. All rights reserved. wikiHow, Inc. is the copyright holder of this image under U.S. and international copyright laws. This image is <b>not<\/b> licensed under the Creative Commons license applied to text content and some other images posted to the wikiHow website. This image may not be used by other entities without the express written consent of wikiHow, Inc.<br>\n<\/p><p><br \/>\n<\/p><\/div>"}
9
Use darker shadow for large or protruding eyes. Prime your eyelid with
eyeshadow primer or a neutral color of eyeshadow. Then, apply a coat of dark
eyeshadow on your top lid under the crease. Blend out the dark color with a
medium color above the crease. Apply a black liner to your waterline. [24] X
Research source
* A layer of black eyeliner all around the eyes will also help close off your eye’s appearance. [25] X Research source
Advertisement
## Expert Q&A
Search
Add New Question
* Question
How do you apply winged eyeliner for different eye shapes?
Alana Schober
Pout Pro Makeup Artist
Alana Schober is a Professional Makeup Artist at Pout by Lipsticknick, an
inclusive beauty bar in Los Angeles, California. With over a decade of
experience, Alana specializes in hair and makeup. As a Licensed
Comselotologist, she has worked for both MAC Cosmetics and Paul Mitchell.
Alana's celebrity clientele includes Iggy Azalea, Solange, YG, and ASAP Nast.
She has worked on campaigns with Nike, Adidas, Morphe, Jaclyn Hill Cosmetics,
and Quay. Alana has also worked on countless music videos and photoshoots and
her work has been featured on two billboards in Los Angeles.
Alana Schober
Pout Pro Makeup Artist
Expert Answer
With almond and round eyes, you can get away with a thicker wing. With hooded
eyes, you should make the wing thinner.
Thanks! We're glad this was helpful.
Thank you for your feedback.
If wikiHow has helped you, please consider a small contribution to support us
in helping more readers like you. We’re committed to providing the world with
free how-to resources, and even $1 helps us in our mission. Support wikiHow
Yes No
Not Helpful 7 Helpful 31
* Question
Is it possible for my eye to be in between almond and round shaped?
Community Answer
Yes, it's possible. If that is the case, take both make-up techniques into
consideration.
Thanks! We're glad this was helpful.
Thank you for your feedback.
If wikiHow has helped you, please consider a small contribution to support us
in helping more readers like you. We’re committed to providing the world with
free how-to resources, and even $1 helps us in our mission. Support wikiHow
Yes No
Not Helpful 8 Helpful 91
* Question
Is it possible to have two different eye shapes? I think my left one is hooded
and the other almond.
Community Answer
Yes, it is. Some people even have one double-eyelid on one eye and a mono-lid
on the other.
Thanks! We're glad this was helpful.
Thank you for your feedback.
If wikiHow has helped you, please consider a small contribution to support us
in helping more readers like you. We’re committed to providing the world with
free how-to resources, and even $1 helps us in our mission. Support wikiHow
Yes No
Not Helpful 9 Helpful 83
See more answers
Ask a Question
200 characters left
Include your email address to get a message when this question is answered.
Submit
Advertisement
##
Video
Read Video Transcript
##
Tips
* It’s possible to have more than one descriptor for your eyes. For example, you could have deep-set, almond, downturned eyes.
Thanks
Helpful 18 Not Helpful 1
* Once you know your eye shape, you can look up tips for eye make-up that will work best for you.
Thanks
Helpful 9 Not Helpful 1
* If you want your eyeshadow to last longer, use an eyeshadow primer first.
Thanks
Helpful 14 Not Helpful 5
Submit a Tip
All tip submissions are carefully reviewed before being published
Submit
Thanks for submitting a tip for review!
Advertisement
## You Might Also Like
How to
Determine Your Body Shape
How to
Determine Your Face Shape
How to
Measure Jacket Size
How to
Look Shorter if You're Tall
How to
Dress Well
How to
Dress Well As a Tall Girl
How to
Part Your Hair for Your Face Shape
How to
Choose a Haircut That Flatters Your Facial Shape
How to
Look Taller
How to
Choose a Winter Jacket Size
How to
Fix Imbalanced Breast Sizes
How to
Make Your Face Look Thinner
How to
Tell if Your Face Is Well Suited to Bangs
How to
Dress in Your 20s
Advertisement
##
References
1. ↑ https://sciencetrends1.com/what-are-monolid-eyes-epicanthic-fold/
2. ↑ https://www.vogue.com.au/beauty/makeup/how-to-determine-your-eye-shape-and-master-your-eyeshadow-application/image-gallery/d7b6b543bf9dafebce8b78d3f402251e?pos=5
3. ↑ https://youtu.be/aXQ-jVDeHMg?t=132
4. ↑ https://imabeautygeek.com/2014/01/08/how-to-apply-makeup-for-your-eye-shape-courtesy-of-smashbox//
5. ↑ https://coba.edu/blog/eyeliner-tips-and-techniques-for-different-eye-shapes/
6. ↑ https://coba.edu/blog/eyeliner-tips-and-techniques-for-different-eye-shapes/
7. ↑ https://www.vogue.com.au/beauty/makeup/how-to-determine-your-eye-shape-and-master-your-eyeshadow-application/image-gallery/d7b6b543bf9dafebce8b78d3f402251e?pos=6
8. ↑ https://youtu.be/ER_4sZK-Ju8?t=36
9. ↑ https://www.beautylish.com/a/vcacj/whats-your-eye-shape
More References (16)
10. ↑ https://youtu.be/uUPV6bnYmHE?t=209
11. ↑ https://www.cosmopolitan.com/style-beauty/beauty/a25394115/different-eye-shapes/
12. ↑ https://youtu.be/I_0iz-prrJk?t=169
13. ↑ https://www.allure.com/story/monolid-eye-makeup-tutorial
14. ↑ https://youtu.be/OoCWxK1153E?t=255
15. ↑ https://youtu.be/OoCWxK1153E?t=297
16. ↑ Alana Schober. Pout Pro Makeup Artist. Expert Interview. 15 June 2021.
17. ↑ https://imabeautygeek.com/2014/01/08/how-to-apply-makeup-for-your-eye-shape-courtesy-of-smashbox/
18. ↑ https://imabeautygeek.com/2014/01/08/how-to-apply-makeup-for-your-eye-shape-courtesy-of-smashbox/
19. ↑ https://imabeautygeek.com/2014/01/08/how-to-apply-makeup-for-your-eye-shape-courtesy-of-smashbox/
20. ↑ https://www.youtube.com/watch?v=d1Td4lMMCiE
21. ↑ https://www.ogleschool.edu/blog/eyelash-extensions-different-shape-eyes/
22. ↑ https://youtu.be/bJ1VPAi9iWs?t=125
23. ↑ https://youtu.be/zKbjHf7bDJE?t=192
24. ↑ https://youtu.be/hgiRSgqdiu8?t=168
25. ↑ https://youtu.be/SlcH3ODFecE?t=156
## About This Article
Co-authored by:
Alana Schober
Pout Pro Makeup Artist
This article was co-authored by Alana Schober . Alana Schober is a
Professional Makeup Artist at Pout by Lipsticknick, an inclusive beauty bar in
Los Angeles, California. With over a decade of experience, Alana specializes
in hair and makeup. As a Licensed Comselotologist, she has worked for both MAC
Cosmetics and Paul Mitchell. Alana's celebrity clientele includes Iggy Azalea,
Solange, YG, and ASAP Nast. She has worked on campaigns with Nike, Adidas,
Morphe, Jaclyn Hill Cosmetics, and Quay. Alana has also worked on countless
music videos and photoshoots and her work has been featured on two billboards
in Los Angeles. This article has been viewed 1,340,021 times.
67 votes - 79%
Co-authors: 33
Updated: April 11, 2024
Views: 1,340,021
Categories: Eye Makeup | Body Type and Fashion
Article Summary X
To determine eye shape, imagine a straight, horizontal line extending through
the centers of both eyes. If the outer corners of your eyes are above the
line, you have upturned eyes. If the corners are below it, you have downturned
eyes. Next, check to see if your eyelid crease is visible or hidden when your
eyes are open. If the crease is hidden, you have hooded eyes. Lastly, if you
can see any white around the top or bottom of your iris, you have round eyes.
If you can't, you have almond eyes. For tips on understanding the position of
your eyes, read on!
Did this summary help you? Yes No
In other languages
Russian
Italian
French
German
Spanish
Indonesian
Dutch
Portuguese
Czech
Arabic
Vietnamese
Japanese
Hindi
Chinese
* Print
* Send fan mail to authors
Thanks to all authors for creating a page that has been read 1,340,021 times.
## Reader Success Stories
* Anonymous
Nov 6, 2016
"The descriptions were really helpful. A lot of websites just use pictures to
help identify eye shapes and expect the person to instantly know what their
eye shape is! Here, certain "measurements" and sectioning off differing
attributes helped identify eye shape. " ..." more
More reader stories Hide reader stories
Share your story
## Did this article help you?
Yes No
Advertisement
Cookies make wikiHow better. By continuing to use our site, you agree to our
cookie policy .
Co-authored by:
Alana Schober
Pout Pro Makeup Artist
67 votes - 79%
Click a star to vote
% of people told us that this article helped them.
Co-authors: 33
Updated: April 11, 2024
Views: 1,340,021
Anonymous
Nov 6, 2016
"The descriptions were really helpful. A lot of websites just use pictures to
help identify eye shapes and expect the person to instantly know what their
eye shape is! Here, certain "measurements" and sectioning off differing
attributes helped identify eye shape. " ..." more
Angelina Vasconcelos
Jul 21, 2020
"I told my family I had big eyes, my family was like no you don't. I slowly
started to look in the mirror to closely see that I have big eyes I thought
to myself I have anime eyes. I do I guess, so I am kinda happy about that!"
..." more
Karina Santiago
Apr 5, 2016
"It has a very detailed description. The pictures helped to determine the kind
of eye shape I had. It taught me more than the YouTube tutorials." ..."
more
Pamela Cavanaugh
Apr 24, 2017
"I really liked the areas of how to dress depending on your shape. I'm an
apple shape, and it showed me the difference on what to wear. " ..." more
Claire Molyneux
Aug 19, 2021
"This has helped me find out what shape eyes I have and good tips on how to
apply eye make up."
Share yours! More success stories Hide success stories
### Quizzes
Am I a Narcissist or an Empath Quiz
Take Quiz
Does He Like Me Quiz
Take Quiz
What Is My Aesthetic Quiz
Take Quiz
Am I Gay Quiz
Take Quiz
Why Can't I Sleep Quiz
Take Quiz
Zodiac Compatibility Quiz
Take Quiz
### You Might Also Like
How to
Determine Your Body Shape
How to
Determine Your Face Shape
How to
Measure Jacket Size
How to
Look Shorter if You're Tall
### Featured Articles
How to
Relive the 1970s (for Kids)
How to
Avoid Drama
How to
Be Who You Want to Be
How to
Make Quick Money Online
The Best Ways to Catch a Cheater on an iPhone
7 Possible Reasons Why She Responds Quickly (and How to Reply)
### Trending Articles
How to Celebrate Passover: Rules, Rituals, Foods, & More
Am I Pretty Quiz
How to
Choose the Right Food for You
50+ Things to Do With Friends When You’re Bored: Free, Creative, & Exciting
Activities
Plant and Grow an Avocado Seed in Soil: A Step-by-Step Guide
How to
Stop a Wedding
### Featured Articles
How to
Be Outgoing
How to
Get Whiter Teeth at Home
How to Get to Know Yourself: A Guide to Self-Discovery
90+ Hilarious Happy Birthday Messages for Husbands
How to
Read Body Language
How to Relieve & Prevent Headaches & Migraines Fast
### Featured Articles
19 Ways to Build Character Every Day
What Does “What's Cooking, Good Looking” Mean? (Plus, How to Reply)
How to
Calm an Aggressive Cat
Feeling Weird? Why It Happens & How to Cope
How to
Be Funny
10 Easy Ways to Braid Your Hair
### Watch Articles
How to
Fold Boxer Briefs
How to
Suspend an Object in Jello
An Absolute Beginners' Guide to Applying Makeup Like a Pro
How to
Make Gummy Bears
How to
Store Cut Ginger
How to Draw a Bunny Step by Step: Easy and Realistic Methods
### Trending Articles
How to Search for a Word or Phrase in a PDF Document
What “Mid” Means and How to Use It (With Examples)
What Fruit Am I Quiz
Is It Okay to Wash Towels and Clothes in One Laundry Load?
How to
Make Samosas
One Piece Trivia Quiz
* Categories
* Personal Care and Style
* Makeup
* Eye Makeup
wikiHow Newsletter
You're all set!
Helpful how-tos delivered to
your inbox every week!
Sign me up!
By signing up you are agreeing to receive emails according to our privacy
policy.
* Home
* About wikiHow
* Experts
* Jobs
* Contact Us
* Site Map
* Terms of Use
* Privacy Policy
* Do Not Sell or Share My Info
* Not Selling Info
* Contribute
Follow Us
×
Get all the best how-tos!
Sign up for wikiHow's weekly email newsletter
Subscribe
You're all set!
X
####
\-
\-
722
| biology | 3830893 | https://sv.wikipedia.org/wiki/Xylonaeus%20breviatus | Xylonaeus breviatus | Xylonaeus breviatus är en skalbaggsart som beskrevs av Heinrich Bickhardt 1916. Xylonaeus breviatus ingår i släktet Xylonaeus och familjen stumpbaggar. Inga underarter finns listade i Catalogue of Life.
Källor
Stumpbaggar
breviatus | swedish | 1.374128 |
eye_capture_shape/how-the-eye-capture-images.txt | ×
* Sign in
* Sign up
* Welcome
* English Spanish Español Russian русский French français Italian italiano Korean 한국어 Japanese 日本語 Polish
* FAQ
* Switch to Night Mode
* Stolen Accounts Recovery
* Change Account Password
* Vote for Witnesses
* Steem Proposals
* Third-party exchanges:
* Poloniex
* Advertise
* Jobs at Steemit
* Developer Portal
* Steem Bluepaper
* SMT Whitepaper
* Steem Whitepaper
* Privacy Policy
* Terms of Service
Home
Login Sign up
* All posts
* Trending Communities
* SteemitCryptoAcademy
* Newcomers' Community
* আমার বাংলা ব্লগ
* Korea • 한국 • KR • KO
* AVLE 일상
* STEEM CN/中文
* WORLD OF XPILAR
* Steem POD Team
* Steem Alliance
* Comunidad Latina
* Incredible India
* Tron Fan Club
* Explore communities …
# How the eye captures images
mgibson ( 66 ) in #science • 7 years ago (edited)
The eye is a visual instrument in its own right and perhaps more perfect than
any instrument that men have ever devised. The complete visual mechanism
involves two steps:
(i) The eye has a lens which being convex in nature produces a real and
inverted image of an object on a light-sensitive membrane, called retina, at
the back of the eye-ball;
(ii) The sensation of the image is then sent through optic nerves to the
brain, which read what the image means.
* * *
Image credit The human eye
* * *
Structure of the human eye
The shape of the human eye is almost spherical. It is placed in bony socket in
skull and can be circulated within this socket extensively with the help of
the complex action of six muscles. The eye has a white unclear coating made of
tough fibrous tissue called the sclerotic. This is the ‘white’ of the eye. The
front division of the sclerotic, called the cornea, is transparent with bigger
curvature. Light gets in the eye through the cornea. The next layer inside the
sclerotic is called the choroid. It is a dark thin membrane made of many blood
vessels. The innermost layer of the eye is called the retina where the images
are formed. It is also a light sensitive membrane which directly has
connection with the optic nerve. The presence of black pigments in the choroid
prevents lights from being reflected back on to the retina again.
* * *
Image credit The diagram of the human eye
* * *
A narrow slit like anterior chamber, which contains aqueous humor, is found
behind the cornea. Aqueous humor is a clear salt solution which helps the eye
be inflated. Iris lies just behind the anterior chamber. It is a circular
diaphragm of pigmented membrane and has an adjustable central hole, called the
pupil. The size of the pupil can be adjusted via iris muscles. With
contraction of these muscles, the pupil extends to admit more light to the
interior of the eye and vice-versa. Iris muscles acts involuntary. The
contraction and expansion of the pupil depends on the external light whether
it is strong or weak. The color of iris increases the quality color of the
eye. The color of the pupil always looks black because all kinds of light that
enter the eye are absorbed by the retina.
* * *
Image credit Optical center in the brain
* * *
A transparent biconvex body just lies at the back of the iris, called the
crystalline lens composed of transparent flexible material, so that the lens
can adjust its shape. It is fixed with the support of suspensory ligaments
which are controlled by ciliary muscles. When these muscles contract or
expand, the curvature of the lens increases or decreases accordingly. The
ciliary muscles also act involuntary. We see different objects at different
distances at different moments. Without our knowing, immediately, the
curvature of the lens changes its shape to adjust its focal length, so that
the image can be formed on the retina.
There is a large space between the lens and the retina, called posterior
chamber. It is filled with the vitreous humor which is a transparent colorless
jelly-like substance.
Formation of the images on the retina
Rays from the object get in the eye through the cornea. These rays then pass
through the aqueous humor, the lens and the vitreous humor and finally fall on
the retina to form the images. The images that form on the retina are real and
inverted due to the converging optical system of the eye. The inverted images
is corrected and identified in the optical center of the brain.
* * *
Image credit The image formation on the retina
* * *
A slight depression is found in the retina, called the yellow spot or the
mocula lutea. Its diameter is about 2 mm. the diameter of the center of the
mocula lutea is about 0.25 mm., a minute region, called the fovea centralis.
It is most active part of the retina. The vision appears most clear when the
images form at the fovea. The muscles that control the eye always tend to
rotate the eye ball until the image forms on the fovea. Vision formed through
the fovea is called the direct vision and the vision which is formed via the
other parts of the retina, called the indirect vision.
* * *
Follow me
#biology #steemstem #eye
7 years ago in #science by mgibson ( 66 )
$ 9 .06
* Past Payouts $9.06, 0.00 TRX
* \- Author $7.53, 0.00 TRX
* \- Curators $1.53, 0.00 TRX
98 votes
* \+ val-a
* \+ witness.svk
* \+ michael-b
* \+ michael-a
* \+ wang
* \+ benjojo
* \+ created
* \+ roelandp
* \+ asmolokalo
* \+ joseph
* \+ aurel
* \+ exyle
* \+ michaeldodridge
* \+ justtryme90
* \+ svk
* \+ sauravrungta
* \+ biodragon
* \+ etcmike
* \+ picokernel
* \+ theyeti
* and 78 more
Reply 0
* * * *
### Coin Marketplace
STEEM 0.29
TRX 0.12
JST 0.033
BTC 63630.77
ETH 3179.32
USDT 1.00
SBD 3.95
| biology | 449353 | https://sv.wikipedia.org/wiki/Geometrisk%20optik | Geometrisk optik | Geometrisk optik, även kallad strålgångsoptik, är studiet av egenskaper hos ljus i form av raka strålar. Det görs med enkla geometriska modeller, därav namnet. Vanligtvis färdas ljuset inte enbart i form av raka strålar och det vet vi bland annat genom fenomen som diffraktion. Det är dock en rimlig approximation när ljuset passerar genom stora öppningar, där öppningens storlek, d, är mycket större än våglängden, , det vill säga d>>. Inom geometrisk optik ignorerar man alltså fenomenet med att ljus böjs på grund av diffraktion. Det kan kännas ganska naturligt: en klar och solig dag kastar objekt skarpa skuggor, och på morgonen kan man se att ljus färdas i linjer mellan träden.
Se även
Lins
Spegel
Stråle (optik) | swedish | 0.757834 |
eye_capture_shape/eye-photography-how-to-take-macro-pictures-of-eyes.txt | * Our products
* Luminar Neo
* Luminar for iPad
* Features
Features
* Enhance AI
* Relight AI
* Erase
* AI Masking
* Layers
* Portrait Background
* Sky AI
* Structure AI
* Skin AI
* Supercontrast
* Composition AI
* Studio Light
* Neon&Glow
* GenErase
* GenSwap
* GenExpand
* Water Enhancer AI
* Twilight Enhancer AI
* See All 30+ Features
Extensions
* Extensions Pack
* Supersharp AI
* Focus Stacking
* Background Removal AI
* Upscale AI
* Noiseless AI
* HDR Merge
* Magic Light AI
* Panorama Stitching
Use Cases
* Landscape Photography
* Wildlife Photography
* Portrait Photography
* Family Photography
* Wedding Photography
* Newborn Photography
* Photoediting for beginners
* AI Photo Editor
* Real Estate
* E-commerce Photography
* Food Photography
* Pricing
* What’s new
* Marketplace
* More
Company
* About Skylum
* Careers
* Newsroom
For Business
* Technology Licensing
Other
* Photo Tours
* Blog
* Ambassadors
* Affiliate Program
* Need help?
* Log In
English
* English
Log In
New tools. New look. New possibilities.
View plans
Back to all posts
December 29, 2022
# Eye Photography: How to Take Macro Pictures of Eyes?
Javier Pardina
Commercial Photographer
*
People have been photographing eyes since the moment they learned how to take
pictures. As we know from ancient times, the eyes are the mirror of the soul.
And it is whether or not you can take an amazing picture of a person's eyes
that the portrait will turn out.
Certain subtleties can help you make a beautiful photo, where the soul and
mood of the model will be expressed to the fullest extent. In our tutorial, we
will show you how to take a photo of your eyes correctly.
## Light for Macro Pictures of Eyes
Eyes look best when they are well-lit with beautiful highlights. Glare is the
reflection of the light source in your eyes. The size, shape, and brightness
of the glare depend on the catchlights you are using. Two large frosted
windows from the studio create beautiful glare. A diffused flash on your
camera creates a softer rectangular reflected glare. A medium-sized softbox
positioned off-camera at 2 o'clock gives a soft, natural glare. Poorly lit
eyeball photography without glare looks cold and a little creepy.
In addition to glare, you can capture various reflections in the model's eyes.
This is a great way to give portraits a certain artistic atmosphere. There are
many options - you can capture another person, an object, or even a landscape.
Capturing reflections can be especially interesting in close-up eyes
photography – try experimenting with it!
Avoid the red-eye effect. Red-eye in portrait photography is a common red-
eye phenomenon in flash photography, and it happens when the flash is get too
close to the camera. This effect is easily avoided by following a few
photography eye close-up tips:
* Remove the flash from the camera. Use only remote on-camera flashes;
* Reflect the flash off the ceiling or walls;
* Use special diffusers and reflectors;
* Use a light disk to diffuse the light from the flash;
* Turn on the camera's red-eye reduction mode;
* If nothing helps, red-eye can be removed during post-processing as a last resort.
## Camera Settings for Eye Macro Photography
Use the zoom to bring the eye in the frame closer. Adjust the zoom to get
exactly the frame you want. If you widen the frame to include additional
details, it can provide the photo with additional context. However, it can
shift the focus away from the subject you originally wanted to photograph.
Ensure that the camera is stationary by placing it on a tripod or other secure
surface. When shooting at close range, the slightest shake of the hand can
blur the picture. Using a tripod or shooting from a stationary surface that
will support the camera will help prevent blur. Consider using a macro ring
that reduces the focal length of the lens. The macro ring is mounted between
the camera body and the back of the lens. Using the macro ring allows you to
zoom in on the eye in the frame, thereby capturing more detail.
Attach a macro lens to your camera. A macro lens will allow you to capture the
eye in the most detail. Macro lenses range in focal length from 50 mm to 200
mm. Of course, you can find a perfect moment to shoot off the eye with a
normal lens but it won't fill the entire frame and be as detailed as you want
it to be.
If you don't have a macro lens and don't want to spend the money to buy one,
try using a magnifying filter as an alternative.
Reduce the camera aperture to increase the depth of field. Close-up shots
look better with good sharpness. Reduce the aperture from f/5.6 to f/11.
The specific aperture value will depend on what kind of eye detail you want to
emphasize in your photo. Experiment with the aperture to see how different
settings affect the frame. Now that you know direct camera settings let's talk
about how to take good pictures of your eyes.
## How to Do Beautiful Eye Macro Photography?
Let's start with the secrets that almost everyone knows. In bright light, the
pupils narrow, but in dim light or a special mood, they dilate, and you can
get an incredibly beautiful shot with "open" pupils. So you don't want to use
a flash. Experiment with natural light and auxiliary permanent sources, which
will help you create a correct and beautiful light pattern. While directing
the light, make sure not to direct it clearly into the wrinkles in the corners
of the eyes - such an effect will not be appreciated in the finished photo.
Now its time for a few other tips and eyes photography ideas:
* When you try several different light sources, keep in mind that the process can take a long time, because the color temperature of the eyes will change, depending on the lighting.
* It's no secret that human eyes are asymmetrical. There is a secret among photographers - to focus on one eye, so the photo will be more interesting making your model's gaze will attract the viewer's eye.
* An interesting trick is to take a photo of the look of your eyes photography with.... an object reflected in them. Place the light source, and the object that you think is most relevant and appropriate in front of the model. The photo will turn out magical and will arouse the viewer's interest.
* Another trick is macro photography of eyes. You will need a special camera or rings of attachments for macro, and also a quiet model who will not mind a long process. You will need to arrange a comfortable place for her to work, where she will be comfortable and quiet for a long time.
* Be sure to give the model a rest, as long, uninterrupted staring into the camera will make your eyes water and, of course, can be uncomfortable. So you need time for a short break, which will allow the model to recover.
* To make the eyes close-up photography session more comfortable, try advising the model to blink or use eye drops before each new shot.
* Use a slower shutter speed to avoid blurring the frame. The eye is constantly moving, which may result in a blurred image. To get a sharp image, need to adjust the shutter speed to 1/100th of a second or faster.
* Using a tripod will also allow you to use a shorter shutter speed.
* Reduce the ISO setting to avoid digital graininess. Increasing the ISO setting allows you to take photos when subjects are poorly lit, but the photos will become grainy. If you are taking photos in good light, keep the ISO setting to the lowest acceptable level. Manually adjust the focus. Autofocus may not focus the lens exactly on the details of the eye you want to capture, so it is best to turn it off and focus manually. To adjust the focus yourself, rotate the lens focus ring until the frame is completely blurred. Then start twisting it back until the parts you want to capture are in focus.
Now, that you know how to take eye pictures, let's talk about another aspect
of this case.
### Close-Up Filters
Close-up filters are filters for close-ups. Their action can be compared to
that of a magnifying glass. They scale the object, but the quality of the
image is somewhat reduced, as well as the amount of light entering the lens.
Filters are inexpensive and are good for fun experiments if you don't have a
macro lens. There is a gradation for them: +1, +2, +5, and so on, the higher
the number, the stronger the filters magnify, and the less light hits the
sensors.
### What Will Be Required of the Model?
Of course, not everything during the shoot is up to the photographer. How to
take eyes photography from the model side? The model will need to take care of
her eyes the day before the shoot. Not to spend sleepless nights or attend
noisy parties on the eve of the photo session. The model should get a good
rest, and gain strength. If the sleep is not good and prolonged - you'll get
on the photo yellowish whites, maybe even with red streaks, which absolutely
will not be too attractive. Of course, if the idea of the photo session is not
to capture a tired person. In other cases let your eyes rest. It is very
important to give instructions on sleep and rest regimens before taking an
eyes portrait photography session.
Of course, such advice may seem optional, as yellowish whites can be easily
corrected during photo retouching . But the mood and the inner state of the
model during the shooting cannot be corrected so easily.
Today we have learned what you can do to make the model's eyes more expressive
and express her soul.
### Post-Processing
Post-processing is very important in photography eyes. You can take your
portrait photo from just good to the best. The key to great post-processing is
not to overdo it.
It's easy to enjoy all the tricks and enhancements that post-production
software offers. When it comes to retouching, the photographer tries to use
the night rule. After Gina has worked on an image, she won't look at it again,
at least not during the day. More often than not, you find that your eyes are
constantly adapting to the modifications you make, so it becomes harder to see
when you've gone too far.
## Edit Your Macro Shots of Eyes in Luminar Neo
Now that you know how to take a picture of your eye, let's talk about how to
improve it!
The powerful photo editor Luminar Neo provides a great way to add surreal or
fantastical elements to your macro-eye sharp images! With the extensive
toolset of this app, your options are limited only by your imagination and the
time you're willing to spend on photo processing! Even if you are new to photo
editors, the Artificial Intelligence editing tools will help you improve your
photos.
## Conclusion
Macro photography is nothing new, but 10-15 years ago, it was quite a rare
phenomenon. Until now, macro photography is still something unusual for many
people, and even unattainable. With our tutorial, you can take such unusual
photos as macro-eye photography. Remember that it is not the final truth and
with experience, you will be able to improve it for yourself personally,
because the information here is for beginners. Don't be afraid to experiment
and you will be able to create your masterpieces of macro eyes pictures!
### Experience the power of Luminar Neo
Try free Try free
### Did you enjoy this post?
Share it on your social media
* * * *
### Advanced yet easy-to-use photo editor
view plans
## Most Popular
April 17, 2023
#### How to Use a Handheld Light Meter
If you've never used a handheld light meter to dial in your exposure settings,
you don't know what you're missing. Here's what it is and how to use it. Our
experts have prepared an excellent comprehensive guide on where is the light
meter on a camera and why you should not be afraid to work with it. Learn
More
May 24, 2023
#### How To Create HDR Images – Best HDR Photography Tutorial
In order to create HDR pictures you need to have a few RAW images, shot in a
special way, and HDR merging software to process a single final HDR photo.
Learn More
May 31, 2023
#### Monopod vs. Tripod: Which One Should You Use for Travel Photography?
Monopods and tripods can be useful for many different kinds of shooting, but
for others, they are absolutely indispensable. Learn More
### A Special Perk for Our Blog Readers
Get a 10% discount on Luminar Neo and dive into professional photo editing
today!
Please, enter valid email! Get the Discount
I agree to my personal data being stored and used to received newsletters and
commercial offers from Skylum.
### Thank you for subscribing.
Your gift is waiting in your inbox!
## Skylum Blog
The latest news and updates. direct from Skylum
READ MORE
Use Aurora HDR for free for 14 days.
Sent successfully!
Please check your inbox.
We've sent you a copy via email.
Sorry
Looks like you're subscribed already
Sorry
This is sad. Looks like you’ve earlier unsubscribed from Skylum emails.
Subscribe to news and offers.
Get my trial
Thank you for downloading
Aurora HDR
Preparing your download...
Did your download not start?
Dont worry, just click here to try again.
Oops! Something went wrong. Don't worry, just click here to try again.
Step 1
Step 1 Find & Open
Aurora HDR Installer
Step 2
Step 2 Follow the instructions
to install Aurora HDR
Step 3
Step 3 Enjoy new photo
editing experience
Beginners guide Video gallery
Get started for free
Try out Luminar Neo for free for 7 days. No credit card needed.
Sent successfully!
Please check your email
Get a download link
Make sure it's a valid email address
👆 This app is available for desktop only. We will send you a link when you’re
back on your computer.
### Unlock Pro-Quality iPhone Photos with Our Free Guide! 📸
Gain instant access to simple yet powerful tips for enhancing composition,
lighting, and editing—everything you need to transform your photos
effortlessly!
👇 Fill out the form below to receive your guide directly via email.
I agree to my personal data being stored and used to receive the newsletter
Get it free now
### Curiosity is the doorway to skill
Thank you for your interest! Discover the secrets to pro-quality photos in
your guide, waiting in your inbox.
### Try the request again later. If the error does not resolve, contact
support.
Try Later
Please wait
Products
Luminar Neo Luminar for iPad Online Photo Editor Luminar Neo Trial
Extensions Features Discounts Photo Tours
Marketplace
Skies Luminar presets Presets Bundles Sky Objects E-books LUTs Textures
Company
About Skylum Careers Ambassadors Affiliate Program Terms of use Privacy
Policy AI Guidelines Alternatives
Help
Contact Support FAQs User Guide Change Choice on Cookies
For Business
Technology Licensing
Learn More
Blog How To Glossary Newsroom Insiders Community Luminar for Creators
Top On Blog
Best Photo Editing Tools for PC Best Free Photo Editing Software for Mac
Best Free Photoshop Alternatives Best Free Lightroom Alternatives What is
HDR photography Best Photo Sharpening Softwares What is a Contrast in
Photography?
Top How To Tips
How to Capture Rain in a Photo How to Create a Black Background How to
Resize Photos for Facebook Profile How to Make a Digital Photo Look Vintage
How to Make a Picture Bigger How to Make Blurry Image Clear and Sharp How to
Open RAW Files
© 2010 - 2024 Skylum ® . All Rights Reserved.
Language
English
* English
#### Extensions
Supersharp AI Focus Stacking Background Removal AI Upscale AI
Noiseless AI HDR Merge Magic Light AI Panorama Stitching
#### Features
Image Brightener
Photo Enhancer
Skin Retouching
Image Eraser
Sky Replacement
Auto Photo Editor
Color Photo Editor
Image Resizer
B&W Photo Editor
Photo Restoration
Selective Color
RAW Photo Editor
Photo Filters
Grain Filter
Accent AI
Image Saturation
Blending Images
Golden Hour Tool
Photo Unblur
Removing Peoples
Dehaze Filter
Blur Backgrounds
Image Color Replacer
Sepia Filter
Change Background Color
Neon & Glow Editor
Transparent Background
Clone Tool
Straighten Image
Precise Image Recolor
Photo Blemish Remover
Body Photo Editor
Motion Blur Effect
4K Image Upscaler
Flip Image
Image Tinter
Fade Images
Add Noise to Image
Darken Image
Batch Photo Editor
Portrait Bokeh Effect
Tilt Shift Filter
AI Photo Retouching
Smooth Photo Editor
#### Alternatives
Lightroom
Photoshop
Topaz
Affinity
ON1
Capture One
| biology | 155298 | https://no.wikipedia.org/wiki/Kamera | Kamera | Et kamera er et apparat til å ta bilder med, enten fotografier (fotografiapparat, fotoapparat, stillbildekamera), film (filmkamera) eller video (videokamera). Denne siden omhandler fotografiapparater.
Et kamera består av et lystett kammer (i kamerahuset) hvor det i den ene enden er en linse (objektiv) eller tilsvarende og i den andre et lysfølsomt opptaksmedium (film eller bildebrikke). Lys slippes inn gjennom objektivet og begrenses til riktig mengde med hjelp av lukkeren, blender og opptaksmedium eksponering følsomheten. Lyset projiseres på det lysfølsomme opptaksmedium, og skaper et bilde, dette bildet kan reproduseres til et fotografi.
Historie
Navnet kommer av det latinske uttrykket camera obscura, som betyr «mørkt kammer».
Fotografiets historie går tilbake til den arabisk vitenskapsmann Ibn al-Haytham (965–1039) som skrev svært innflytelsesrike bøker om optikk, inkludert eksperimenter med lys gjennom en liten åpning i et mørklagt rom.
Prinsippet er at i et mørkt rom med et lite hull i den ene veggen vil det dukket opp på motsatt vegg et opp-ned bilde av landskapet utenfor. En anvendelse av dette prinsippet er det enkle hullkameraet, hvor "linsen" består av et lite hull, stort som en nålestikk.
På 1700-tallet ble det oppdaget at sølvsalter var følsomme for lys. På 1800-tallet kunne bilder fra kameraet fremkalles på ulike måter for å gjøre bildet permanent, og de første fotografiene ble laget.
Franskmannen Joseph Nicéphore Niépce var den første som laget et kamera i 1826, men det tok åtte timer å eksponere bildet og det ble veldig uskarpt, han brukte en boks som kamera. Niépce selv var egentlig ikke så interessert i kamera, han var mest interessert i å kopiere. Han hadde tidligere kopiert grafiske trykk på metallplater.
I 1839 oppdagert franskmannen Louis Jacques Mande Daguerre at en sølvbelagt kobberplate kunne gjøres lysfølsom i joddamp. Han var den første som fikk et akseptabelt resultat på denne måten. Bildet kunne dessverre ikke kopieres. Daguerre hadde tidligere samarbeidet med Niépce.
I 1835 laget William Henry Fox Talbot et papir lysfølsomt i klor-sølvløsning. Han fikk et negativt resultat, det vil si at hvit ble fremstilt som svart og svart ble fremstilt som hvit. Farger i mellom ble avbildet i forskjellige gråtoner, lyse farger ble i virkeligheten mørke på fotografiet og omvendt. Da han hørte om Daguerres oppdagelse i 1839, begynte han å eksperimentere med klorsølvløsning og kom til slutt på en spesiell prosess for å kopiere bildet. Dette skjedde i 1840 og teknikken brukes fortsatt i dag for å fremkalle bilder. Det kalles kalotype eller taletype.
To av firmaene som gjorde fotografering tilgjengelig for alle var Kodak og Leica. Georg Eastman lanserte i 1890 et Kodak kamera under mottoet: Trykk på knappen – vi ordner resten. Med dette kamera trengte man ikke tungvinte glassplater og kjemisk kompetanse fordi det hadde en filmrull med 100 bilder. Etter at bildene var tatt sendte man kameraet tilbake til fabrikken som fremkalte filmen og sendte apparatet tilbake til eieren sammen med kopiene og med ny film satt inn i kameraet. Oskar Barnack lanserte Leica 1 i 1925 et lite kamera som man kunne benytte til landskapsfotografering uten å bære på tungt utstyr. Kamera benyttet standard 35 mm kinofilm.
Gjennom årene har de lysfølsomme emulsjonene blitt foredlet og er i stand til å avbilde fargene enten i en gråskala som oppfattes som normal, eller å avbilde fargene trofast.
Enkle kameraer til privat brukKameraer til privat og mer profesjonell bruk
Kameratyper
Kameraer er delt inn på ulike måter, for eksempel etter fototeknologi, bildeformat/sensorstørrelse, kamera størrelse eller bruksmåte.
Digitalkameraet har en lysfølsom bildebrikke og det analoge kameraet bruker en fotografisk film.
Enøyde speilreflekskameraer
Det enøyde speilreflekskameraet er det vanligste kameraet for pressefotografer og amatører. Det enøyde speilreflekskameraet er nå også vanlig i sin digitale form. De har et speil som gjør at når man ser i søkeren ser man gjennom objektivet. Når bildet tas vippes speilet vekk og lyset gjennom objektivet treffer filmen eller biledbrikken.
Analoge kamera benytter 35 mm film.
Toøyde speilreflekskameraer
Denne kameratypen var vanlig frem til 1960-tallet. Den har et objektiv for søker og et for filmen eller biledbrikken.
Denne kameratypen er i prinsippet alltid et mellomformatkamera med et bildeformat på 6 × 6 cm.
Storformatkameraer
Et storformatkamera har et bildeformat som er større enn mellomformatkameraet, vanligvis 9 × 12 cm eller større. Denne typen kamera har mange innstillingsmuligheter, inkludert front og bak kan vippes og forskyves. Det moderne storformatkameraet er designet slik at ulike deler (objektiver, bakside, matte plater, søkere osv.) kan byttes ut for å passe til ulike opptakssituasjoner.
Kompaktkameraer
Kompaktkameraer er som navnet sier mindre og letter enn speilreflekskameraene. De har ikke utskiftbare objektiver men har ofte et zoomobjektiv.
Noen kompaktkameraer er veldig enkle, andre har omtrent samme innstillingsmuligheter som for eksempel det enøyde speilreflekskameraet. Noen digitale kompaktkameraer har ingen optisk søker i det hele tatt, bare en skjerm.
De analoge kameraene har vanligvis en gjennomsiktsøker. Analoge kamera benytter 35 mm film.
Konstruksjon
Kameraet består av et lystett kamerahus med et objektiv på den ene veggen og et lysømfintlig medium (fotografisk film eller bildebrikke) på motsatt vegg. Lyset passerer gjennom objektivet og avtegner et bilde på filmen.
Mer avanserte kameraer, som speilreflekskamera, har et utskiftbart objektiv. På eldre speilreflekskamera ble objektivene skrudd på, skrufatning, på nyere kamera kan man klikke dem på, bajonettfatning.
Bilde av et gjennomskåret spejlreflekskamera Film eller bildebrikke (Electronic sensor) – filmen eller en billedbrikke som omdanner lys til elektriske signaler
Utløser (Shutter release ) – et bilde tas når den trykkes helt ned. Automatisk fokusering oppnås ved å trykke den delvis ned
Data skjerm (Data display) - en LCD-skjerm som lar brukeren se gjeldende innstilling, på digitale kamera er denne informasjonen på skjerm på baksiden av kamera eller i søker.
Søker (Viewfinder) – lar brukeren se motivet som fokuseres på filmen eller bildebrikken
Linser (Lens Elements) – kan justeres frem eller tilbake for å bringe objekter i fokus på ulike avstander
Blender (Aperture) – øker eller minker i diameter for å slippe mer eller mindre lys gjennom linsen til filmen eller den elektroniske sensorenFoto og filmkameraer benytter vanligvis synlig lys, men det finnes også andre typer kameraer som arbeider med andre deler av det elektromagnetiske spekteret, som ved infrarød fotografering.
Lukker
Utdypende artikkel: Lukker
Lukkerens oppgave er å åpne kameraet for lysinnslipp i et bestemt tidsintervall - lukkertida. Sammen med blenderåpningen avgjør lukkertida hvor mye lys filmen eller bildebrikken skal eksponeres for. Lukkertida kan varieres i trinn fra kortere enn 1/1000 sekund til flere sekunder. Hvert trinn tilsvarer en dobling eller halvering av lukkertida. For eksponeringen er dermed et trinn på lukkertida likeverdig med et blendertrinn, og til sammen kalles disse eksponeringstrinn.
Ved håndholdt kamera og lange lukkertider er det en risiko for at bildet ikke blir skarpt fordi kameraet beveger seg under eksponeringen. Noen moderne objektiver er utstyrt med optisk bildestabilisator som registrerer og kompenserer for rystelser i kameraet under opptak, og demper virkningen av kamerabevegelsene. En del digitale kameraer har i stedet innebygd bildestabilisator i form av en bevegelig bildebrikke som kompenserer tilsvarende for bevegelsesuskarphet. Bruk av bildestabilisator tillater skarpe bilder med lengre lukkerhastigheter (kan variere opp til 4-5 eksponeringstrinn). Bildestabilisatoren kan normalt ikke kompensere for motiv i bevegelse, men noen typer bildestabilisatorer kan registrere at kameraet følger motivbevegelsen i én retning (følgefokusering) og vil i praksis gi skarpere bilder i denne retningen når fotografen klarer å følge motivet.
På kompaktkameraer sitter lukkeren i objektivet (sentrallukker) og er konstruert som, eller deler funksjon med, en irisblender. Systemkameraer med utbyttbart objektiv har oftest lukkeren inne i huset, nær filmen/bildebrikka. En slik fokalplanlukker er konstruert som et gardin som trekkes til side under eksponeringstida, eller den kan bestå av lameller. På moderne kameraer beveger lukkeren seg gjerne vertikalt, fordi en vertikallukker tillater kortere lukkertider enn en horisontallukker som må bevege seg en lengre avstand. Noen profesjonelle systemkameraer har sentrallukker i objektivet, andre har fokalplanlukker med mulighet for å bruke objektiver med sentrallukker.
Ved bruk av lang lukkertid må man benytte et fotostativ for å holde kamera helt stille.
Søker
Utdypende artikkel: Søker
Søkerens oppgave er å gi fotografen en oppfatning av bildeutsnittet og hvordan bilde vil bli før eksponering skjer.
I speilreflekskameraer fungerer objektivet også som søker, slik at det bildet som ses i søkeren er det samme som eksponeres på filmen. Det vil se at det man ser i søkeren kommer via et speil fra objektivet.
I kompaktkameraer er søkeren et eget vindu på utsida av kamerahuset, litt til side for objektivet. I en slik gjennomsiktssøker vil det ikke være fullt samsvar mellom det som vises i søkeren og det som eksponeres på filmen.
Digitale kompaktkameraer har elektronisk søker som viser på en LCD-skjerm det som til enhver tid registreres i bildebrikka.
På moderne kamera vises også mye informasjon i søkeren, som for eksempel gjenværende batterikapasitet,
Eksponering
Utdypende artikkel: Eksponering
Hvor mye lys filmen eller bildebrikka eksponeres for, bestemmes ved valg av blenderåpning og lukkerhastighet. For øvrig bestemmes eksponeringa av lysfølsomheten i opptaksmediet. Mediets lysfølsomhet måles i ISO-verdier. For fotografisk film er følsomheten bestemt ved den måten filmens belegg av lysfølsomme korn er bygd opp. I digitalkameraer kan følsomheten - dvs. hvor mye signalene som går inn på brikka skal forsterkes - varieres fra bilde til bilde.
Mest brukt er film med middels følsomhet (ISO 200). I digitalkameraer kan følsomheten reguleres fra ISO 50 (lav) til 1600 (meget høy). En film med høye ISO-verdier har tykkere hinne med større krystaller, og krever mindre belysning eller kortere lukkertid før bildet dannes. Til gjengjeld kan svært raske filmer gi et kornet bilde med øker risikoen for bildestøy.
De fleste digitalkameraer er utstyrt med eksponeringsautomatikk som sørger for at den lysmengden som når bildebrikka er i samsvar med brikkas følsomhet. Det er vanligvis fire programmer for eksponeringskontroll:
A (Aperture) – blenderprioritert automatikk: Brukeren stiller inn blenderåpning, kameraet beregner øvrige variabler.
S (Shutter) eller T (Time) – lukkerprioritert automatikk: Brukeren velger lukkertid.
P – programmert automatikk: Kameraet velger sjøl lukkertid og blenderåpning.
M – manuell innstilling: Brukeren velger lukkertid og blenderåpning.
I tillegg kan kameraet ha forhåndsprogrammerte innstillinger for visse motiver og situasjoner, f.eks. sport, nærfotografering, kveldsbelysning, motlys osv. Kameraet gir mulighet for å over- eller undereksponere bildet i forhold til de innstillingene som automatikken har valgt.
Lysmåling
For at fotografen eller eksponeringsautomatikken skal kunne velge riktige innstillinger, er kameraet utstyrt med en innebygd lysmåler som måler intensiteten i det lyset som reflekteres av motivet. En kan velge mellom punktmåling, der lyset måles innafor en liten sirkel i motivet, sentrumsdominert lysmåling, der lyset måles innafor en større sirkel i sentrum av motivet, eller flerfelts lysmåling, der lyset måles på punkter fordelt over hele motivet.
Nyere kameraer gir mulighet for evaluerende lysmåling. Her blir en flerfelts lysmåling automatisk jamført med et større antall forhåndsprogrammerte motiver og belysninger, slik at riktig eksponering kan beregnes for det foreliggende motivet med den gitte belysninga.
Ny teknologi har gjort det mulig å fremvise bilder på andre medier, for eksempel på TV eller lerret. Denne fremvisningsteknikken, på engelsk kalt «slideshow», ble blant annet demonstrert av fotograf Even Totland på Geilo i 2011.
Hvitbalanse
På digitale kameraer kompenseres det for den varierende fargetemperaturen (hvitbalanse) på forskjellige lyskilder elektronisk. Dette sørger for at hvitt lys registreres som nettopp det på det lagrede bildet og at forskjellen fra det blå lyset fra lysstoffrør og det gule fra glødepærer blir mindre framtredende på det ferdige bildet. Hvis man har et filmkamera kompenserer man med forskjellige typer film eller fargefiltre på objektivet. Hvitbalanse kan også justeres for estetikkens skyld ved å gjøre lyset enda gulere og varmere eller blåere og kaldere enn det egentlig er. Fargetemperatur defineres i Kelvin-grader; 5000 K vil passe i normalt dagslys. Ved lavere K-grader vil lyset bli blått, noe som kompenserer for for eksempel glødelampe. Ved høyere K-grader blir lyset rødere, noe som kompenserer for lysstoffrør og lignende.
Objektiv
Utdypende artikkel: Objektiv
De enkleste objektivene består av én konveks linse; slike finnes i gamle kassekameraer og i enkle mobiltelefonkameraer. De fleste moderne objektiver består av 5-10 linser, i visse fall mange flere. Kompaktkameraer har objektivet fast montert til huset, mens systemkameraer har utskiftbare objektiv.
Brennvidde
Objektivets brennvidde (f) bestemmer bildevinkelen, dvs. hvor stort område som fanges inn. I et kamera med vanlig 35 mm film vil et objektiv med en brennvidde på 50 mm ha en bildevinkel på ca. 45° og gi et bilde som noenlunde tilsvarer øyets perspektiv. Et slikt objektiv (50 mm) kalles derfor et normalobjektiv.
Objektiv med kortere brennvidde gir større bildevinkel og kalles vidvinkelobjektiv, det vil si at man får med mer på bilde enn øyets perspektiv (fra 40 til 20 mm, med vanlig 35 mm film). Dette er for eksempel mye benyttet til innendørs fotografering når man ønsker å få med et stort område på bildet, eller når motivet er et vidt landskapsmotiv.
Spesial objektiver med svært stor bildevinkel, gjerne hele 180°, disse har da en ennå mindre brennvidde (fra 18 mm og nedover, med vanlig 35 mm film) får stor forvrengning, disse blir kalt fisheyeobjektiv.
Objektiv med lengre brennvidde (gjerne i området 85 til 135 mm eller spesial objektiver helt opp til 1200 mm, med vanlig 35 mm film) gir et forstørra utsnitt av et mindre område og kalles teleobjektiv. Disse fungere som en kikkert og benyttes når man skal fotografere motiver langt unna, som dyr eller fugler.
Alternativt kan objektiver bygges slik at linseelementene kan forskyves innbyrdes, slik at brennvidden kan varieres etter behov. Slike kalles zoomobjektiv. De fleste kompaktkameraer er utstyrt med zoomobjektiv.
Digitale kompaktkameraer og de fleste digitale systemkameraer har ei bildebrikke som er mindre enn 35 mm. Forholdet mellom objektivets brennvidde og bildevinkelen blir derfor et annet enn ved tradisjonelle 35 mm-kameraer. For å få sammenliknbare verdier, oppgis for slike kameraer både den faktiske brennvidden og "brennvidden" omregna til 35 mm-format. Brennvidden er imidlertid uendret selv om bildeformatet endres.
Fokusering
Fokusering (skarphetsinnstilling) for å få bildet skarpt skjer ved at linseelementene flyttes i forhold til filmen. Man må stille/skru på objektivet for å få bildet i fokus. Jo nærmere motivet kameraet er, jo lenger frem må du flytte objektivet. Derfor er det av praktiske årsaker som regel en grense hvor det ikke lenger er mulig å forlenge linsen, og dette avgjør også hvor nærme motivet kan gå. Den minste avstanden til motivet kalt nærgrense.
De fleste moderne kameraer har autofokus, dvs. at elektronikk i kameraet beregner fokus og at fokusering skjer ved hjelp av en innebygd elektrisk motor. På kameraer uten autofokus er det ofte et splittbilde, eller annen løsning, som vises i søkeren for å hjelpe til med fokuseringen.
Enklere kameraer (for eksempel de fleste mobiltelefonkameraer) har fast fokus. Dette betyr at objektivet er låst til en viss fotograferingsavstand.
Blender
Utdypende artikkel: Irisblender
For å regulere hvor mye lys som slippes inn på filmen eller bildebrikken og dybdeskarpheten er kameraet som regel utstyrt med en irisblender (i enkelte tilfeller kan blenderen være av en enklere konstruksjon). Blenderen er innbygd i objektivet, og har lameller som gir en nær sirkelrund åpning med variabel diameter. Blenderåpningen angis som en brøk (hele trinn: f/1,0 - f/1,4 - f/2 - f/2,8 - f/4 - f/5,6 - f/8 - f/11 osv.) som angir forholdet mellom objektivets brennvidde (f) og diameteren på åpningen. En endring i ett helt blendertrinn betyr en dobling eller halvering av lysmengden gjennom objektivet. Blenderåpningen påvirker dybdeskarpheten i bildet, et lavt blendertall betyr mindre dybdeskarphet. Blender 16 gir således mye større dybdeskarphet enn blender 2,8 f.eks, men dette avhenger også i stor grad av brennvidden for objektivet. Som eksempel gir blender 2,8 på et vidvinkelobjektiv mye større dybdeskarphet enn 2,8 på ett teleobjektiv. Avhengig av konstruksjonen og kvaliteten av linsen, gir en nedblending fra største blender ned 2-3 hakk også større skarphet og kontrast, slik at optimal kvalitet ofte oppnås ved blenderverdier fra f/5,6 til f/11.
Blits
Utdypende artikler: Blits
Når det er mørkt kan man kan benytte en lyskilde for å lyse opp motivet. Kompaktkamera og dikitalkamera har ofte en innebygget blits for å kompensere for manglende eller uheldig belysning på motivet. Blitsen sender ut et kraftig lysblink i det lukkeren på kameraet åpnes, eller rett før lukkeren lukkes.
På speilreflekskameraer er det en blitssko hvor man kan feste en ekstern blits, skoen har en kontakt slik at kamera og blits blir synkronisert. Også kamera med innebygget blits kan være utstyrt med blitssko fordi de innebyggede blitsene er svake, ca. 3 m, og sitter nære objektivet.
Tidligere hadde blitsen engangs blitspærer, senere kom det blitskuber med 4 pærer og andre løsninger med flere engangs-pærer.
Opptaksmedium
Utdypende artikler: Fotografisk film og Bildebrikke
Fotografisk film
Fotografisk film lages plastremse (filmrull) belagt med et lysfølsomt lag bestående av sølvforbindelser som er innkapslet i et gelatinlag. De aller første fotografiske filmene var plater laget av metall eller glass, belagt med et lysfølsomt lag.
Når det lysfølsomet belegget blir utsatt for lys skjer en kjemisk prosess der sølvsaltkrystallene omdannes til rent sølv. Det mønsteret som derved dannes på filmen, utgjør et latent bilde av motivet. Ved en kjemisk prosess som kalles framkalling, dannes et synlig bilde på filmen.
Når vanlig/negativ film er fremkalt vil man ha et negativt bilde, det vil si at på et sort-hvit bilde er det som var sort når bilde ble tatt hvit på filmen, det som var hvit blir sort på filmen, lyst blir mørkt og mørkt blir lyst. På negativ fargefilm opptrer fargene med sine komplementærfarger. Den negative filmen kopieres til papirbilder og får da sine riktige/opprinnelige farger.
Det finnes også positiv film som ikke gir et negativt bilde. Denne benyttes til lysbilder, også kalt slides, som plasseres i en framviser og projiseres på lerret, eller betraktes gjennom ei lyslupe.
Kameraet ble åpnet og filmen satt inn, for hvert bilde som ble tatt ble filmen trukket frem manuelt eller automatisk om kameraet hadde denne funksjonen. 35 mm filmer fås kjøpt med 36 eller 24 bilder. Når alle bildene på filmen var tatt måtte den sveives manuelt eller automatisk tilbake inn i filmboksen slik at den ikke ble utsatt for lys når kameraet ble åpnet for å bytte film.
Lysfølsomheten, «hastighet», tidligere spesifisert i ASA eller DIN, er nå spesifisert i ISO-grader, se avsnitt om eksponering.
Instantfilm
Instantfilm, umiddelbar film eller direktefilm (Polaroid) krever et spesialkamra, instant kamra. Bildet blir fremkalt med en gang etter at det er tatt.
Med det tradisjonelle instant kamra blir bildet fremkalles gradvis etter å ha blitt matet ut av kameraet, men det finnes også en moderne kamera hvor bilde fremkalls ved hjelp av varmebehandling inne i kamera før det blir matet ut.
Bildebrikke
I digitale kameraer benyttes ikke film men en bildebrikke som er en komponent i kameraet . Den omdanner lys til elektriske signaler som kan avleses digitalt. Brikken består av en matrise av kvadratiske, lysfølsomme sensorer, som blir bildepunkter eller piksler i det ferdige bildet. Antall bildepunkter utgjør brikkens og bildets oppløsning, og måles i megapiksler (millioner piksler). Antall bildepunkter ligger på moderne kameraer mellom 4 og 16 megapiksler.
Informasjonen fra bildebrikken lagres i et minnekort.
Se også
Forskjellige typer kamera:
Camera obscura
Digitalkamra
Filmkamera
GoPro kamera
Instant kamera
Kameraovervåking
Kjøretøykamera
Nettkamera
Polaroidkamera
Speilreflekskamra
Undervannskamera
Varmekamera
Viltkamera
Fotografering:
Digital fotografi
Flyfotografi
Fotografering
Holografi
Lysbilde
Makrofotografi
Ortofotografi
Panoramafotografering
Undervannsfotografering
Fototeknikk | norwegian_bokmål | 0.609311 |
eye_capture_shape/21823-eyes.txt | Locations :
Abu Dhabi | Canada | Florida | London | Nevada | Ohio |
* 800.223.2273
* 100 Years of Cleveland Clinic
* MyChart
* Need Help?
* Giving
* Careers
* Search
* ClevelandClinic.org
* Find A Doctor
* Locations & Directions
* Patients & Visitors
* Health Library
* Institutes & Departments
* Appointments
Home /
Health Library /
Body Systems & Organs /
Eyes
Advertisement Advertisement
Advertisement
# Eyes
Your eyes are a key sensory organ, feeding information to your brain about the
outside world. Your eyes do the “physical” part of seeing. The signals they
send allow your brain to “build” the picture that you see. Eye-related
symptoms are also key clues to issues affecting your whole body, so experts
recommend making eye health a priority.
Contents Arrow Down Overview Function Anatomy Conditions and Disorders
Care Additional Common Questions
Contents Arrow Down Overview Function Anatomy Conditions and Disorders
Care Additional Common Questions
## Overview
Your eye is made up of many structures that work together so you can see.
### What are eyes?
Your eyes are the sensory organs that allow you to see. Your eyes capture
visible light from the world around you and turn it into a form your brain
uses to create your sense of vision. Your brain doesn’t have sensory abilities
of its own. It needs your eyes (and other senses, like hearing and touch) to
gather information about the world around you.
Most people are born with two eyes. Working together, they give you a field of
view about 200 degrees wide and 135 degrees tall. When your eyes work together
correctly, they give you depth perception and 3D vision. They also give you
color vision.
It’s also important to remember that sight and vision aren’t necessarily the
same thing, even though many people — including eye care specialists and
healthcare professionals — use those terms interchangeably. Sight is what your
eyes do. Vision is the entire process that starts with sight and ends with
your brain processing what your eyes see into a form your brain can use and
understand.
Care at Cleveland Clinic
Find a Primary Care Provider
Schedule an Appointment
Advertisement
Cleveland Clinic is a non-profit academic medical center. Advertising on our
site helps support our mission. We do not endorse non-Cleveland Clinic
products or services. Policy
## Function
### How do your eyes work?
Everything your eyes do starts with light from the outside world. Your eye
structure lets light enter and pass through a series of clear components and
sections, including the cornea, aqueous humor, lens and vitreous humor. Those
structures bend and focus light, adjusting how far the light beams travel
before they come into focus.
The focus needs to be precise. If it isn’t, what you’re looking at appears
blurry. Your eye has muscles that can make subtle changes to the shape of your
eye, moving the focus point so it lands correctly on the retina.
When light lands on the cells of your retinas, those cells send signals to
your brain. The signals are like coded messages describing everything they can
about the light. That includes the color, how intense it is and any other
relevant details. Your brain decodes and processes the signals and uses them
to “build” the image you see.
## Anatomy
### How do eyes work?
Human eyes are complex, and it takes many parts working together correctly for
you to see.
#### Eye anatomy
The parts of your eye include the:
* Cornea . This protects the inside of your eye like a windshield. Your tear fluid lubricates your corneas. The corneas also do part of the work bending light as it enters your eyes.
* Sclera . This is the white part of your eye that forms the general shape and structure of your eyeball.
* Conjunctiva . This clear, thin layer covers the sclera and lines the inside of your eyelids.
* Aqueous humor . This is fluid that fills a space called the anterior chamber. The pressure of the aqueous humor helps maintain your eye’s shape.
* Iris . This part contains the muscles that control the size of your pupil. It’s also responsible for eye color . The iris can be brown, blue, green or hazel (a blend of brown, yellow and green).
* Pupil . This is the black circle inside the iris. It’s like an adjustable window to the inside of your eye. It widens and narrows to control how much light enters your eye.
* Lens . This focuses light that enters your eye and directs it to the back of your eye.
* Vitreous humor . This clear, gel-like fluid fills the space between the lens and retina. It helps your eye hold its shape. It’s also sometimes known simply as “the vitreous.”
* Retina . This thin layer of light-sensitive cells at the back of your eyes converts light into electrical signals. It contains rods (which help you see in low light) and cones (which help you see colors).
* Macula . This small area of your retina is key to your vision. It’s responsible for the center of your visual field. It also helps you see color and fine details.
* Optic nerve . This connects your retinas to your brain. It’s like the data cable that carries signals from your eyes, with connection points linking to multiple brain areas.
* External muscles. These control your eye’s position, alignment and movement. They also contribute to your eye’s shape, which is part of your ability to switch your vision’s focus between near and far objects.
Advertisement
## Conditions and Disorders
### What are the common conditions and disorders that affect your eyes?
The types of conditions that can affect your eyes vary depending on the
specific part(s) involved. That’s because your eyes include a variety of
tissue types. It has muscle, connective tissue, nerves, blood vessels and
more.
Some of the different types of eye conditions include — but aren’t limited to
— the following:
* Refractive errors . These are problems with how you see because light isn’t coming into focus on your retinas correctly. Refractive errors can take many forms, such as focusing too soon ( nearsightedness ) or too late ( farsightedness ). They can also involve distortions in your sight, like with astigmatism .
* Corneal disorders . These are conditions that affect the cornea itself. They can happen for many reasons, ranging from congenital conditions (which you have at birth) to conditions that don’t develop until later in life.
* Retinal disorders . These conditions can happen because of problems that affect the retina directly. They can also be secondary effects of another disease, like how lattice degeneration can lead to a retinal detachment .
* Optic nerve-related conditions . These affect the nerve that links the eyes and brain. Examples include optic neuritis and optic atrophy .
* Age-related eye disorders . These conditions are more likely to happen as you get older, especially after age 65. They range from minor concerns like age-related loss of near vision ( presbyopia ) to serious concerns like cataracts. Some age-related eye diseases, like macular degeneration or glaucoma , are severe enough to cause permanent vision loss.
Your eyes are also susceptible to more general conditions and issues. Examples
of these include:
* Infections .
* Injuries and trauma .
* Cancer .
* Congenital malformations .
### What are some common signs or symptoms of eye conditions?
Signs and symptoms of an eye condition can vary greatly. One reason for that
is the many different parts that affect or contribute to your vision. A common
example of this is how a metabolic and circulatory condition like Type 2
diabetes can lead to vision loss over time.
Some symptoms affect the surface of your eye only. Others affect the inside of
your eye. Some key types of eye symptoms include:
* Eye surface issues.
* Eye appearance/alignment.
* Eye function and sight.
#### Eye surface issues
These symptoms affect your eyes’ surface or the area immediately surrounding
them. They include:
* Irritation .
* Red eye .
* Watery eyes .
* Discharge, ooze or “goop” coming from your eyes.
#### Eye appearance/alignment
* Changes in the color of the sclera (such as blue or yellow sclera ) or noticeable bleeding into the conjunctiva.
* Reflex or appearance changes affecting the iris or pupil (like leukocoria, coloboma , etc.).
* Eye misalignment (strabismus) , including inward ( esotropia ), outward ( exotropia ), upward ( hypertropia ) or downward (hypotropia).
#### Eye function and sight
Symptoms from eye-related conditions can also affect your sight itself. The
eyes are often a sense you rely on heavily, so sight- or vision-related
symptoms are often easier to notice.
Some of them involve changes or disruptions in how or what you see. Examples
include — but aren’t limited to — the following:
* Trouble seeing clearly or properly : Blurred vision from refractive errors, or specific vision issues like night blindness or color blindness .
* Decrease or loss of vision : Sudden vision loss or low vision and blindness .
* Disrupted vision : Double vision or visible auras (bright haze or splotches in your field of view) like from ocular migraines .
* Light sensitivity (photosensitivity or photophobia) : This is when bright light causes you pain or discomfort severe enough that you try to avoid brighter surroundings and prefer dimmer areas or light levels.
#### How the link between your eyes and brain influences symptoms
It’s also important to remember that many causes of eye symptoms — especially
sight/vision changes or eye movement control symptoms — may not be due to an
eye condition. Some might happen due to a condition elsewhere in your body. An
example of this is yellowing of the sclera when you have jaundice.
The familiar saying is that “the eyes are a window to the soul.” But from the
medical perspective, they’re also like a window to your brain. Eye-related
symptoms are a key way for healthcare providers to find brain conditions and
issues. That’s why vision changes can be telltale indicators of brain-related
issues like concussions or strokes .
#### What are some common tests to check eye health?
Many tests can detect conditions that affect your eyes directly or that cause
eye symptoms. The most important of them is an eye exam . Regular eye exams
can detect many eye conditions or concerns before you ever have symptoms. And
eye exams can help prevent long-term vision damage or issues when you have
other conditions like Type 2 diabetes.
Other common tests include:
* Color blindness test .
* General imaging tests, like ultrasound or magnetic resonance imaging (MRI) .
* Glaucoma tests .
* Intraocular pressure test .
* Retinal imaging tests .
* Slit lamp exam .
* Visual acuity test .
* Visual field tests .
There are many tests your eye care specialist or other healthcare provider may
recommend depending on your symptoms and the suspected cause(s). They may also
recommend tests for other body systems that might influence or cause eye
symptoms. Your specialist or provider is the best source of information about
test options, what they recommend and why.
Advertisement
### What are some common treatments for eye conditions?
There are many possible treatments for eye conditions, and the treatments can
vary widely. Some conditions or concerns that are common or not severe may
have simpler treatments. Other conditions or concerns need more advanced care
options.
Some examples of types of eye care include:
* Vision correction . This is the main approach to treating eyesight issues like nearsightedness, farsightedness or age-related near vision loss. Eyeglasses and contact lenses are the most common options. Some people need prescription glasses or contacts, while others may only need reading glasses (sometimes known as “cheaters”). Others may choose to undergo vision correction surgery .
* Medications . Medications can treat many conditions that — either directly or indirectly — affect your eyes. The type of medication depends on the specific condition(s) involved and other factors. These include medicated drops or ointments you apply to your eyes or medications you take other ways (by mouth, via injection or infusion, etc.).
* Surgery . Many eye conditions are treatable with surgery. These can include a variety of methods, including phacoemulsification (which uses ultrasound to break up cataracts), cryotherapy (which uses intense cold), radiofrequency ablation (which uses intense heat) and laser surgery.
Many other possible treatments can play a role in treating eye conditions or
symptoms. Because there are many influencing factors, your eye care specialist
or healthcare provider is the best person to tell you more about treatment
options. They can explain the options and help you choose one that’s most
likely to help you.
## Care
### How can I care for and keep my eyes healthy?
There are several things you can do to maintain your eye health. You can:
* Get regular eye exams . Everyone should get an eye exam every one to two years, regardless of whether or not they need glasses or contacts. And if you have a higher risk of eye disease, you may need more frequent eye exams. Your eye specialist can advise you on how often you should get an exam.
* Wear eye protection . Safety glasses or goggles can make all the difference in avoiding eye injuries or damage. While putting them on might seem like a hassle or unnecessary step, they can spare you a lot of pain and avoid irreversible damage.
* Quit using tobacco , or never start . Nicotine-containing products, including vaping or smokeless (chewing) tobacco, can contribute to circulatory problems over time. This is especially true of the smaller, more delicate blood vessels in your eyes. Ask your provider for resources to help you quit.
* Make nutrition a priority . Getting enough essential vitamins, minerals and other nutrients is a big help to your eye health.
* Reach and maintain a weight that’s healthy for you . Your weight and overall health can influence your eye health.
* Don’t ignore eye-related symptoms . Changes in your vision or symptoms that otherwise affect your eyes are often the earliest signs of a greater issue. Talk to an eye care specialist or healthcare provider about your concerns. It’s better to ask and not have an issue than not ask and have an existing issue worsen.
## Additional Common Questions
### When should I get medical attention for conditions or issues related to
my eyes?
Several eye-related changes or symptoms mean you (or someone you care for)
need to get medical attention. Some examples of these include:
* Sudden vision loss (either partial or total) that affects one or both eyes.
* An injury where you have something stuck in your eye, or you feel like something is stuck on the surface of your eye, even though you can’t see anything there.
* Burns around or on your eyes (even minor burns).
* Any injury that makes your eye red, swollen, bruised or bleed.
* Any condition or injury that causes the eye socket area around your eyeball to swell.
* If your eyes bulge or stick out noticeably farther than usual.
* If your vision becomes noticeably cloudy, hazy or foggy (especially if this happens suddenly).
* If you have eye symptoms along with vomiting, chills, fever or other infection symptoms.
* Bright flashes of light in your vision, a sudden increase in floaters , or a loss of vision that looks like a dark curtain or wall covering part of your vision.
* Kaleidoscope vision .
* If you have trouble moving your eyes a certain way or direction they can usually move.
* Sudden tunnel vision .
* Distortions or loss of central vision (with or without changes to your peripheral vision).
There are many more reasons other than those listed above. When in doubt, the
safest choice is to get medical attention quickly. Doing so could help protect
your eyesight and vision or could even help save your life.
### What are the most common eye problems?
The most common eye problems (other than temporary conditions like eye
infections or irritation) include:
* Refractive errors.
* Age-related cataracts.
* Age-related macular degeneration.
* Retinal diseases (especially diabetes-related retinopathy).
* Glaucoma.
* Dry eye disease .
### Which organ is connected to the eyes?
Your optic nerve is a direct connection between your eyes and brain. How your
eyes develop also means your retinas are technically part of your central
nervous system , brain and spinal cord .
A note from Cleveland Clinic
Your eyes are one of your brain’s windows to the world. They gather light from
your surroundings and help your brain build the picture you see. It’s easy to
take this sense for granted when it works properly and easy to miss when it
doesn’t. You can do several things to help protect and maintain your eyes,
many of which involve simple steps you can take every day.
If you have concerns about your vision and any related health effects, it’s a
good idea to talk to an eye care specialist or healthcare provider. They can
help you learn more and take steps to safeguard your vision. And when in
doubt, talk to a medical professional or seek medical care. Doing so without
delay could protect you from long-term vision issues.
Medically Reviewed
Last reviewed by a Cleveland Clinic medical professional on 11/15/2023.
Learn more about our editorial process .
#### References
Advertisement
Cleveland Clinic is a non-profit academic medical center. Advertising on our
site helps support our mission. We do not endorse non-Cleveland Clinic
products or services. Policy
Care at Cleveland Clinic
Find a Primary Care Provider
Schedule an Appointment
Ad
Advertisement Advertisement
Advertisement
Appointments 216.444.2020
Appointments & Locations
Request an Appointment
Facebook Twitter YouTube Instagram LinkedIn Pinterest Snapchat
##### Actions
Appointments & Access Accepted Insurance Events Calendar Financial
Assistance Give to Cleveland Clinic Pay Your Bill Online Refer a Patient
Phone Directory Virtual Second Opinions Virtual Visits
##### Blog, News & Apps
Consult QD Health Essentials Newsroom MyClevelandClinic MyChart
##### About Cleveland Clinic
100 Years of Cleveland Clinic About Us Locations Quality & Patient Safety
Office of Diversity & Inclusion Patient Experience Research & Innovations
Community Commitment Careers For Employees Resources for Medical
Professionals
##### Site Information & Policies
Send Us Feedback Site Map About this Website Copyright, Reprint & Licensing
Website Terms of Use Privacy Policy Notice of Privacy Practices Non-
Discrimination Notice
9500 Euclid Avenue, Cleveland, Ohio 44195 | 800.223.2273 | © 2024 Cleveland Clinic. All Rights Reserved.
| biology | 81705 | https://da.wikipedia.org/wiki/Listepunkt | Listepunkt | Et listepunkttegn ( • ) er et typografisk symbol, der bruges til at angive elementer i listepunktform.
Punktform
Et sædvanlig liste af punkter på punktform kan se ud som:
Dette er et element i en liste. Se punkttegnet forrest.
Dette er et nyt element – derfor et nyt punkt.Dette er anden linje af det andet element. Der er ikke noget nyt punkt hertil.
Dette er det tredje og sidste element.
Indholdet ud for de enkelte punkter (indholdet kaldes ofte også for punkter) kan være enkelte ord eller længere sætninger. Hvis det er sætninger vil man som regel afslutte hvert enkelt punkt med punktum. Dog, hvis det er enkelt-ord eller tilsvarende (for eksempel en opskrift) kan afsluttende tegnsætning udelades eller forblive som var det en komma-separeret liste:
Til maden skal bruges:
2 spsk. olie
1 valnød
Eller som var det en liste, der ikke var på punktform:
For et nå målet skal vi benytte:
gå-på-mod,
opfindsomhed samt
samarbejde
I datalogi
Det sædvanlige typografiske punkttegn er at finde som Unicode-tegnet U+2022: •. Der findes også et triangulært punkt som U+2023: ‣.
I HTML
I HTML kan det angives som • eller •, der bliver til • henholdsvist •. Dog vil man som oftest lave lister på punktform ved at lave liste-elementer – eng.: list item: <li> – inden i en ikkeordnet liste – eng.: unordered list: <ul>.
Det triangulære punkt laves som ‣: ‣.
I typografi
Ovenstående sædvanlige punkt fra Unicode (som det ser ud i denne font) er højst sandsynligt en lille rund udfyldt prik, der kaldes punkttegn. Men punkttegnet behøver ikke bestå af runde prikker, men kan være en vilkårlig geometrisk figur (firkanter, trekanter, streger, pile) eller helt andre figurer (diverse små piktogrammer).
Tegnsætning
Typografi | danish | 1.262697 |
shower_muscles/327461.txt | * Health Conditions
* Health Conditions
* Alzheimer's & Dementia
* Anxiety
* Asthma & Allergies
* Atopic Dermatitis
* Breast Cancer
* Cancer
* Cardiovascular Health
* COVID-19
* Diabetes
* Environment & Sustainability
* Exercise & Fitness
* Eye Health
* Headache & Migraine
* Health Equity
* HIV & AIDS
* Human Biology
* Leukemia
* LGBTQIA+
* Men's Health
* Mental Health
* Multiple Sclerosis (MS)
* Nutrition
* Parkinson's Disease
* Psoriasis
* Psoriatic Arthritis
* Sexual Health
* Ulcerative Colitis
* Women's Health
* Health Products
* Health Products
* Nutrition & Fitness
* Vitamins & Supplements
* CBD
* Sleep
* Mental Health
* At-Home Testing
* Men’s Health
* Women’s Health
* Discover
* News
* Latest News
* Original Series
* Medical Myths
* Honest Nutrition
* Through My Eyes
* New Normal Health
* Podcasts
* 2023 in medicine
* Why exercise is key to living a long and healthy life
* What do we know about the gut microbiome in IBD?
* My podcast changed me
* Can 'biological race' explain disparities in health?
* Why Parkinson's research is zooming in on the gut
* Tools
* General Health
* Drugs A-Z
* Health Hubs
* Health Tools
* Find a Doctor
* BMI Calculators and Charts
* Blood Pressure Chart: Ranges and Guide
* Breast Cancer: Self-Examination Guide
* Sleep Calculator
* Quizzes
* RA Myths vs Facts
* Type 2 Diabetes: Managing Blood Sugar
* Ankylosing Spondylitis Pain: Fact or Fiction
* Connect
* About Medical News Today
* Who We Are
* Our Editorial Process
* Content Integrity
* Conscious Language
* Newsletters
* Sign Up
* Follow Us
Medical News Today
* Health Conditions
* Health Products
* Discover
* Tools
* Connect
Subscribe
# What are the benefits of cold and hot showers?
Medically reviewed by Debra Rose Wilson, Ph.D., MSN, R.N., IBCLC, AHN-BC,
CHT — By Jessica Caporuscio, PharmD — Updated on March 29, 2023
* Cold showers
* Hot showers
* When to take
* Potential risks
* Summary
Cold and hot showers may have different benefits. A cold shower may help
reduce pain, inflammation, and swelling, while a hot shower may boost
cardiovascular health and improve sleep.
Most studies into the health benefits of cold and hot water have used water
immersion therapy. During water immersion therapy, a person immerses their
body, or a part of their body, in a vat of water for a specified amount of
time.
This article outlines the results of such studies, including the potential
health benefits and risks of cold and hot showers.
##
Benefits of cold showers
Although taking a cold shower may not be a pleasant experience, it can provide
certain health benefits. One 2014 review , for example, suggests that cold
water can:
* decrease inflammation and swelling
* decrease muscle spasms
* have pain-relieving effects
An older study, this time from 2000 , investigated the effects of different
water temperatures on bodily reactions. In the cold water condition, the
researchers submerged participants in 57.2°F (14°C) water for 1 hour. The
participants experienced the following effects (some of which may not
necessarily be beneficial to health):
* increased metabolism
* increased heart rate and blood pressure
* increased levels of the neurochemicals dopamine and norepinephrine
* lower levels of cortisol , the “ stress hormone”
Some beneficial effects of cold water immersion include:
### Improved circulation
Exposure to cold water causes the blood vessels on the surface of the skin to
constrict. This diverts blood flow away from the surface of the skin. In fact,
one small 2019 study found that taking a cold shower after exercise may
improve general hydration by cooling down the body.
As blood travels away from the skin, blood vessels in deeper body tissues
dilate. This improves circulation in the deeper tissues.
### Lower cortisol levels
Cortisol is the “fight-or-flight” hormone that the body releases in response
to stress .
Lowering cortisol levels in the blood may therefore help reduce stress levels.
Researchers note that when people immerse themselves in a bath of cold water,
their cortisol levels drop.
### Reduced pain
According to a 2014 review , cold water exposure triggers an automatic pain
response called stress-induced analgesia (SIA).
SIA is a reduced pain response during or following exposure to a stressful
stimulus, such as cold water immersion.
### Reduced sports-related muscle soreness and fatigue
One older study from 2009 tested the effects of cold water immersion on
physical performance among athletes. Following sports, some of the athletes
received cold water immersion, while others received tepid water immersion.
The researchers did not find any differences in subsequent physical
performance between the two groups, nor did they find any differences in the
amount of muscle damage or inflammation the participants experienced.
However, perceptions of muscle soreness and general fatigue were
significantly lower in the group that received the cold water immersion. This
psychological effect could prove beneficial during competitive sports.
##
Benefits of hot showers
Hot showers may also have beneficial effects for health. Some examples of the
potential health benefits include:
### Improved cardiovascular health
As one 2014 review of hydrotherapy notes, warm water bathing appears to
improve blood flow in people with chronic heart failure . This is due to a
natural widening of the blood vessels when exposed to high temperatures.
A 2012 study explored the effects of warm water immersion on arterial
stiffness, which occurs when the arteries within the body become less
flexible. Arterial stiffness plays a major role in the cardiovascular
condition atherosclerosis , which can lead to high blood pressure .
The study found that participants who submerged their feet and lower legs in
warm water for 30 minutes showed reduced arterial stiffness. Participants who
did not receive warm water immersion did not show this effect.
### Improved muscle and joint health
As mentioned above, hot showers can enhance blood flow, helping soothe stiff
joints and tired muscles. Cold showers, meanwhile, can reduce inflammation and
help numb pain.
One 2017 study investigated the effect of hot and cold therapies on knee
osteoarthritis. The researchers divided 96 participants into three groups. A
control group received only standard osteoarthritis treatment. The two other
groups received standard osteoarthritis treatment along with either heat
therapy or cold therapy.
The heat therapy group applied heat to the affected knee twice daily for 3
weeks, while the cold therapy group applied cold to the affected knee twice
daily for 3 weeks.
At the end of the study, both groups showed a mild reduction in pain and a
mild improvement in knee function. However, these effects were not
significantly stronger than those the control group experienced.
### Improved brain health
One 2018 study investigated the effects of hot water immersion on brain-
derived neurotrophic factor (BDNF). BDNF is a protein that has several
important functions in the brain and spinal cord, including:
* promoting the survival of nerve cells
* promoting the growth, maturation, and maintenance of nerve cells
* promoting learning and memory
The study was small, involving just eight men. One group took a 20-minute bath
in hot water with a temperature of 107.6°F (42°C). The other group took a
20-minute bath in warm water with a temperature of 95°F (35°C).
Participants who took the hot bath had significantly higher BDNF levels. The
researchers concluded that hyperthermia induced by the hot bath increased the
production of BDNF.
### Improved sleep
Organizations such as the National Heart, Lung, and Blood Institute
recommend taking a hot shower or bath before bed as a way to improve sleep.
A hot shower may improve sleep quality due to the combination of how the body
relaxes while bathing and the fall in body temperature afterward.
##
When to take hot or cold showers
Doctors sometimes recommend heat or cold therapy for people with muscle or
bone injuries. However, it is not yet clear whether or not a cold or hot
shower has the same benefits as applying an ice or heating pad.
Some people with arthritis enjoy hot showers in the morning because it helps
them feel mobile. However, cold showers are beneficial for injuries
accompanied by inflammation.
For taking a hot shower to improve sleep, a 2019 study found that the best
time to take a shower or bath was 1–2 hours before sleeping.
People with eczema and other rashes should avoid hot showers altogether, as
these can further dry out the skin.
##
Potential risks
Long baths or showers that are too cold or too hot could excessively lower or
raise a person’s body temperature.
If a person’s body temperature drops below 89.96°F (32.2°C) , a person may
experience:
* a decreased breathing rate
* a drop in blood pressure
* heartbeat irregularities
* decreased consciousness
If the water is too hot, people are at risk of burns and heat stroke .
As long as a person avoids extreme temperatures, taking cold and hot showers
can provide certain health benefits. Hydrotherapy is generally a safe
strategy and does not cause dependence or any noticeable side effects.
Further studies will be necessary to investigate and confirm the long-term
effects of taking cold and hot showers.
##
Summary
Most of the research into the health effects of cold and hot water has
involved immersion therapy. Studies seem to suggest that both cold and hot
temperatures can have beneficial, but differing, effects on the body.
It is not clear if taking a shower produces the same effects as immersion
therapy. If it does, a person should consider the type of effects they want to
achieve before deciding whether to take a cold or hot shower.
As with water immersion therapies, taking a cold or hot shower is likely to be
safe. However, people should avoid taking extremely cold or extremely hot
showers, as doing so could be harmful to health.
Last medically reviewed on January 13, 2020
* Public Health
* Immune System / Vaccines
* Sports Medicine / Fitness
* Complementary Medicine / Alternative Medicine
### How we reviewed this article:
Sources
Medical News Today has strict sourcing guidelines and draws only from peer-
reviewed studies, academic research institutions, and medical journals and
associations. We avoid using tertiary references. We link primary sources —
including studies, scientific references, and statistics — within each article
and also list them in the resources section at the bottom of our articles. You
can learn more about how we ensure our content is accurate and current by
reading our editorial policy .
* Aciksoz, S., et al. (2017). The effect of self-administered superficial local hot and cold application methods on pain, functional status and quality of life in primary knee osteoarthritis patients [Abstract].
https://www.ncbi.nlm.nih.gov/pubmed/28880416
* An, J., et al. (2019). The thermal effects of water immersion on health outcomes: An integrative review.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6479732/
* Atopic dermatitis clinical guideline. (n.d.).
https://www.aad.org/practicecenter/quality/clinical-guidelines/atopic-
dermatitis
* Brenner, I. K. M., et al. (1999). Immune changes in humans during cold exposure: Effects of prior heating and exercise.
https://www.physiology.org/doi/full/10.1152/jappl.1999.87.2.699?url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&rfr_dat=cr_pub%3Dpubmed
* Campa, F., et al. Stabilizing bioimpedance-vector-analysis measures with a 10-minute cold shower after running exercise to enable assessment of body hydration [Abstract].
https://journals.humankinetics.com/view/journals/ijspp/14/7/article-p1006.xml
* Eczema and bathing. (n.d.).
https://nationaleczema.org/eczema/treatment/bathing/
* Haghayegh, S., et al. (2019). Before-bedtime passive body heating by warm shower or bath to improve sleep: A systematic review and meta-analysis [Abstract].
https://www.sciencedirect.com/science/article/abs/pii/S1087079218301552?via%3Dihub
* Hu, Q., et al. (2012). Acute effects of warm footbath on arterial stiffness in healthy young and older women [Abstract].
https://link.springer.com/article/10.1007%2Fs00421-011-2066-1
* Kojima, D., et al. (2018). Head-out immersion in hot water increases serum BDNF in healthy males.
https://www.tandfonline.com/doi/full/10.1080/02656736.2017.1394502
* Mooventhan, A., & Nivethitha, L. (2014). Scientific evidence-based effects of hydrotherapy on various systems of the body.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4049052/
* Rowsell, G. J., et al. (2009). Effects of cold-water immersion on physical performance between successive matches in high-performance junior male soccer players [Abstract].
https://www.ncbi.nlm.nih.gov/pubmed/19308790
* Rowsell, G. J., et al. (2011). Effect of post-match cold-water immersion on subsequent match running performance in junior soccer players during tournament play [Abstract].
https://www.ncbi.nlm.nih.gov/pubmed/21077001
* Shevchuk, N. A. (2008). Hydrotherapy as a possible neuroleptic and sedative treatment [Abstract].
https://www.ncbi.nlm.nih.gov/pubmed/17640827
* Šrámek, P., et al. (2000). Human physiological responses to immersion into water of different temperatures [Abstract].
https://www.ncbi.nlm.nih.gov/pubmed/10751106
Share this article
Medically reviewed by Debra Rose Wilson, Ph.D., MSN, R.N., IBCLC, AHN-BC,
CHT — By Jessica Caporuscio, PharmD — Updated on March 29, 2023
## Latest news
* Wearable tech uses AI to detect cardiac arrhythmia 30 minutes before onset
* Artificial sweetener neotame may have potential to damage gut, lead to IBS
* AI tool may help detect cancer in a few minutes with a drop of blood
* Regularly eating avocado linked to lower diabetes risk in women
* Misplaced your keys? How to distinguish dementia from normal age-related memory loss
### Related Coverage
* Are there any health benefits to a cold shower?
Medically reviewed by Alana Biggers, M.D., MPH
The potential health benefits of taking a cold shower include strengthening
the immune system, feeling invigorated, and boosting mood. Learn more…
READ MORE
* * What to know about poor circulation
Medically reviewed by Angelica Balingit, MD
Poor circulation has a range of potential causes, including diabetes and
atherosclerosis. Learn more about poor circulation and how to improve it here.
READ MORE
* Are there any benefits of apple cider vinegar baths?
Medically reviewed by Cynthia Cobb, DNP, APRN, WHNP-BC, FAANP
Taking an apple cider vinegar bath may help with the symptoms of some skin
conditions. They could also help keep skin and hair healthy. Read this…
READ MORE
* What's the best shower frequency?
Medically reviewed by J. Keith Fisher, M.D
The ideal frequency of showering depends on the season and a person’s age and
lifestyle. It is possible to shower too much and cause skin issues…
READ MORE
* What causes itching after a shower?
Medically reviewed by Debra Sullivan, Ph.D., MSN, R.N., CNE, COI
Several things can cause itching after a shower, including eczema and certain
skin care products. Learn more about the causes, treatments, and…
READ MORE
* About Us
* Contact Us
* Terms of Use
* Privacy Policy
* Privacy Settings
* Advertising Policy
* Health Topics
* Health Hubs
* Medical Affairs
* Content Integrity
* Newsletters
* © 2024 Healthline Media UK Ltd, Brighton, UK. All rights reserved. MNT is the registered trade mark of Healthline Media. Any medical information published on this website is not intended as a substitute for informed medical advice and you should not take any action before consulting with a healthcare professional. See additional information .
© 2024 Healthline Media UK Ltd, Brighton, UK. All rights reserved. MNT is the
registered trade mark of Healthline Media. Any medical information published
on this website is not intended as a substitute for informed medical advice
and you should not take any action before consulting with a healthcare
professional. See additional information .
About Careers Advertise with us
OUR BRANDS
Healthline Medical News Today Greatist Psych Central Bezzy
| biology | 4683688 | https://sv.wikipedia.org/wiki/V%C3%A4rmevallning | Värmevallning | Värmevallning eller blodvallning är tillfälliga och ofta återkommande besvär före och efter menopaus, med plötsliga värmevågor i kroppen, ofta tillsammans med rodnad och svettning. En klassisk värmevallning förknippas allmänt med klimakteriet. Tillstånd som liknar eller är identiska med värmevallningar kan ingå i symtombilden vid panikångest, giftstruma, diabetes, carcinoider, carcinom, berusning, med mera. Om en kvinna i fertil ålder tar läkemedel som sänker östrogennivåerna, kan hon också drabbas.
Tillståndet kan förväxlas med feber, men kroppstemperaturen höjs inte, dock höjs hudens temperatur. Kroppstemperaturen under en värmevallning brukar understiga 37C. Precis innan en värmevallning kan kroppstemperaturen stiga, men detta sker så hastigt att det inte hinner uppmätas med muntermometer eller rektal termometer, och örontermometer kan ge falska värden tillföljd av hettande ansikte.
En klimaktierisk värmevallning börjar uppe i huvudet och sprider sig ner i fötterna, eller är begränsad till överkroppen och går upp mot ansiktet och halsen. Samtidigt med värmen börjar vanligen ansiktet rodna och kroppen svettas, och somliga får hjärtrusningar (med palpitationer). Tillståndet varar som regel någon minut, men enskilda kan ha mycket kortare (några sekunder) eller mycket längre (flera minuter) värmevallningar. Värmevallningen åtföljs ofta av frossa. Somliga får framför allt nattliga svettattacker. Värmevallningarna kan vara återkommande i flera månader, men några få kvinnor har återkommande värmevallningar under flera år runt menopaus. De kan återkomma en gång i timmen, någon gång i veckan, eller några enstaka gånger under hela klimakteriet.
Vad som orsakar tillståndet är inte helt klarlagt, men det tycks endokrint sett bero på de förändrade nivåerna av östrogen (östrogenbrist) under klimaktieret, samt på förändrad aktivitet på serotonin och noradrenalin. Neurologiskt har värmevallningarna att göra med förändringar i termoregleringen via hypotalamus. Att huden hettar och rodnar beror på vasodilation, och de neurologiska betingelserna kan uppstå i reaktion på detta, ett missförstånd att kroppstemperaturen skulle vara förhöjd när det i själva verket är blodgenomströmningen i huden som ökar. Risken att drabbas av värmevallningar ökar av rökning, fetma, undervikt, stillasittande, tidigt och hastigt klimaktierium, och att ha haft bröstcancer.
Den som drabbas av värmevallningar har ofta koncentrationsproblem, minnesproblem, och ibland depressioner. Nattliga svettningar ger ofta sömnproblem.
Källor
http://www.medscape.com/viewarticle/540531_11
http://www.patient.co.uk/doctor/Hot-Flushes.htm
http://www.healthline.com/health/menopause/hot-flash-causes#1
https://web.archive.org/web/20140606043232/http://menopausecentre.com.au/Symptoms-Hotflushes-menopause
https://web.archive.org/web/20140903130206/http://www.whria.com.au/page.aspx?docid=115
Klimakteriet
Sjukdomar i svettkörtlarna
Medicinska symtom
Kroppstemperaturförändringar | swedish | 0.788742 |
shower_muscles/hot-vs-cold-showers-health-benefits.txt | Skip to content
* Alien Gear
* Coaching
* Client Testimonials
* Blog
* Exerprise APP
Need help?
[email protected]
Follow Us
* Facebook
* Twitter
* Instagram
* YouTube
* TikTok
* LinkedIn
Back
* Alien Gear
* Supplements
* Ice Shakers
* Equipment
* Apparel & Accessories
Back
* Coaching
* 90 Day Coaching Experience
* Free Workout & Coaching Session
* Personalized Fitness Programs
* Alien Class Pass
Clear
All categories All categories Alien Bottle Amino Acids & Blends Apparel
Fitness Equipment Lifting Accessories Natural Extracts Proteins & Blends
Specialty Supplements Sweatpants T-Shirts Tank Vitamins & Minerals
Close
Anabolic Aliens
Login / Signup
My account
## Login to my account
Enter your e-mail and password:
Email
Password
Login
New customer? Create your account
Lost password? Recover password
## Recover password
Enter your email:
Email
Recover
Remembered your password? Back to login
0
Cart
* Alien Gear
* Supplements
* Ice Shakers
* Equipment
* Apparel & Accessories
* Coaching
* 90 Day Coaching Experience
* Free Workout & Coaching Session
* Personalized Fitness Programs
* Alien Class Pass
* Client Testimonials
* Blog
* Exerprise APP
Next 90-Day Coaching Round Starts May 13, 2024
###### Join us on Next Free Fitness Class on April 28th, 2024
# Hot Vs. Cold Showers Health Benefits | The Ultimate Guide!
.Mike Rosa Anabolic Aliens Sep 29, 2020
Share
* * * *
Some people take steaming hot showers but other people swear by the magical
healing properties of ice cold showers. No matter how you like to shower,
there are proven health benefits to both.
Today we’re going to go over the different health benefits of both hot and
cold showers!
## M uscle Effects
### H ot Showers
Let’s start off with the effects on our muscles. Hot showers are great for
relieving tension. So if your muscles are really sore or tight, hot showers
help relax that tightness and alleviate pain and discomfort, just like the
jets of a jacuzzi.
In fact, if you know you have an intense workout later in the day and you wake
up super stiff, a hot shower in the morning is a great way to help loosen up
and rid yourself of those stubborn knots.
Also, hot showers can help reduce joint inflammation, which can improve your
mobility. Overall, if you are experiencing any form of tightness, pain, or
discomfort, go take a hot shower.
## C old Showers
Cold water can actually help speed up muscle recovery. So taking a cold shower
right after a workout is a great idea. It’s been proven that cold showers
enhance muscle repair and recovery, which reduces delayed onset muscle
soreness.
Cold showers act like an ice pack— they reduce muscular swelling or
inflammation. When we workout and break down those muscle fibers, they swell,
and the coldness of the shower helps reduce that swelling, which speeds up the
repair of the new muscle fibers for an overall quicker recovery.
In general, hot showers can be viewed as a pre-workout tension reliever and
cold showers like a post-workout recovery.
If you’re not used to cold showers, you may have to build a little tolerance
toward them by cycling between hot and cold. The hot shower will open the
blood vessels then switching to cold increases circulation by the blood
rushing toward the organs.
This method will also help remove some of the lactic acid built up from
exercise. However, if you use the cycling method, it is important to end with
the cold shower.
### S kin/Hair Effects
#### C old Showers
Cold water hydrates the skin. So if you’re someone experiencing discomfort
from dry skin, a cold shower is a great fix. The more hydrated the skin, the
healthier it will be.
Also, cold showers will help close skin pores, which protects against dirt,
unnatural oils, germs and other debris getting in them. The combination of the
hydration and the pores getting closed is a solid method of preventing
blackheads as well as other forms of acne. Overall, cold showers significantly
strengthen and protect our skin cells.
This process is also extremely beneficial for hair health, keeping your hair
stronger, shinier and overall healthier. Cold showers can prevent the loss of
natural hair oils, helping with split ends as well as increasing follicle grip
to the scalp. This can help prevent hair loss, especially for men with
receding hairlines.
#### H ot Showers
Hot showers help cleanse the skin like an exfoliator. The steam from the hot
water does the opposite of cold showers and opens up the pores, which can
rejuvenate the skin. This cleanse can help get rid of dead skin, dirt, and
unnatural oils that have built up, which can help eliminate pimples and acne.
Regarding the hair, hot water helps rejuvenate nerve endings for a fuller,
healthier look. Now on the contrary, hot showers can lead directly to dry
skin. If you’re someone who takes long hot showers to relax, you’re actually
hurting your skin and hair health.
If you like your hot showers, the best method to get the best benefits would
be to start with the hot shower for the exfoliating and rejuvenating effects
and end with the cold shower for the hydration and skin cell protection.
### I mmune System/Body Effects
#### H ot Showers
Hot showers are well known for their decongestant effects. The steam from the
hot water relaxes and opens up the nasal passages, allowing the sinuses to
drain out and unclog as well as loosen up phlegm.
This is a great way to help eliminate sinus pressure, overall congestion, and
coughing, thus relieving cold symptoms. Also, the steam from hot water opens
up the lungs for better overall air flow, which can significantly help those
who suffer from asthma to breathe easier.
If you are someone who suffers from migraines, hot water opens up the blood
vessels. Considering most headaches are caused by decreased blood flow, hot
water can help bring blood to your head and ease your pain.
Another property of hot water is that it makes your body sweat which helps rid
itself of bad toxins. Also, the heat destroys germs and bacteria, which is a
great method to prevent sickness.
One last benefit of hot showers is they can actually help with healing. If you
have a small wound that’s open, the hot water can enhance blood circulation to
that specific area expediting the healing process.
#### C old Showers
Cold showers have been proven to strengthen the immune system. Exposure to
cold water results in increased lymphatic movement, ridding the body of large
amounts of waste.
This can prevent infection, increase testosterone and even sperm cell count,
clear blocked arteries, decrease blood pressure, increase the amount of white
blood cells fighting against disease and improve heart health by the promotion
of more efficient blood circulation.
Therefore, if you’re healthy, cold showers are for you. If you’re sick or
experiencing any symptoms of sickness, a hot shower may be more beneficial to
you.
### M ental Health Effects
#### C old Showers
If you’re someone who has a hard time waking up in the morning, take a cold
shower. It is proven that cold showers significantly increase mental alertness
and even reduce body fatigue. One fascinating effect of cold showers is they
may relieve depression.
Essentially, the shock of the cold water sends an electrical impulse to the
brain, igniting the serotonin neurotransmitter as well as releasing
noradrenalin, which regulates mood and alleviates depression.
It’s also been proven that cold showers increase willpower and resilience,
which positively affects the nervous system and can reduce stress. Along with
that, cold showers result in an increase in uric acid and blood glutathione
which lowers stress levels.
This reduced stress can also treat insomnia. Taking a cold shower before bed
has been proven to aid in deeper sleep by lowering body temperature. Also,
cold showers have been shown to enhance deeper and slower breathing to control
stress levels.
#### H ot Showers
On the other hand, hot showers have been shown to stimulate the brain to
release more oxytocin which significantly eases anxiety and can reduce stress
as well. Hot showers have also been proven to act as a sedative to help induce
sleep. The tension relief from hot showers results in a more optimal calming
effect for the mind and body.
### W eight Effects
#### Hot Showers
Hot showers have been shown to increase the body’s metabolism. A boost in
metabolism helps promote weight loss.
#### C old Showers
Cold showers have also been shown to increase metabolism due to the body’s
shock effect. However, cold showers can actually promote fat loss.
There are two types of fat in the body: brown fat and white fat. Brown fat is
the healthy fat that our bodies need for warmth. White fat is the stubborn fat
that is stored when we consume more calories than we burn.
Brown fat is actually more optimally activated by cold showers. This increases
calories burned, which keeps your body warm and promotes the loss of white
fat.
### The Bottom Line
In conclusion, hot and cold showers both have their health benefits. Overall,
cold showers are more than likely the way to go if you are to choose just one.
If you want to get benefits from both hot and cold showers, try a method of
starting with 3 minutes of hot shower and end with 5 minutes of cold shower.
You can also do the alternating back and forth method while ending with a cold
shower.
Lastly, there is no need to shower for a long period of time. Save some water
and keep your showers to under 10 minutes!
### J oin the Invasion!
This Anabolic Aliens membership will grant you access to workout classes,
rehab programs, diet plans, and more exclusive content to help you achieve
sustainable success!
Tags: .mike rosa , Exercises & fitness tips
Share
* * * *
Previous article Next article
Blog Categories
* Workouts
* Exercises & Fitness Tips
* Nutrition
Popular posts
1. Health Benefits of Gensing Root & More | Anabolic Aliens
2. 9 Intense Transverse Abdominis Exercises | Strengthen Your Core Today!
3. How Much Cardio Is Too Much
4. The Pronated Pull Up: A Complete Guide | How To Build A Bigger Back!
5. Raw Food Diet: Everything You Need To Know
Featured products
Anabolic Aliens 6 Week Fitness Program
Anabolic Aliens 8 Week Fitness Program
Anabolic Aliens 10 Week Fitness Program
Anabolic Aliens 12 Week Fitness Program
## Join Us
For exclusive recipes, workouts, discounts, opportunities, new product
announcements - plus a few surprises!
Your email
Subscribe
100% free, Unsubscribe any time!
WORKOUTS
EXERCISES & FITNESS TIPS
NUTRITION
Quick Links
* Terms & Condition
* Privacy Policy
* Shipping policy
* Return and Refund Policy
* Affiliate Program
* Contact Us
* Invasion Membership
Keep Following Us
* * * * * *
Country/region
United States (USD $)
Country/region
* Afghanistan (AFN ؋)
* Åland Islands (EUR €)
* Albania (ALL L)
* Algeria (DZD د.ج)
* Andorra (EUR €)
* Angola (USD $)
* Anguilla (XCD $)
* Antigua & Barbuda (XCD $)
* Argentina (USD $)
* Armenia (AMD դր.)
* Aruba (AWG ƒ)
* Ascension Island (SHP £)
* Australia (AUD $)
* Austria (EUR €)
* Azerbaijan (AZN ₼)
* Bahamas (BSD $)
* Bahrain (USD $)
* Bangladesh (BDT ৳)
* Barbados (BBD $)
* Belarus (USD $)
* Belgium (EUR €)
* Belize (BZD $)
* Benin (XOF Fr)
* Bermuda (USD $)
* Bhutan (USD $)
* Bolivia (BOB Bs.)
* Bosnia & Herzegovina (BAM КМ)
* Botswana (BWP P)
* Brazil (USD $)
* British Indian Ocean Territory (USD $)
* British Virgin Islands (USD $)
* Brunei (BND $)
* Bulgaria (BGN лв.)
* Burkina Faso (XOF Fr)
* Burundi (BIF Fr)
* Cambodia (KHR ៛)
* Cameroon (XAF Fr)
* Canada (CAD $)
* Cape Verde (CVE $)
* Caribbean Netherlands (USD $)
* Cayman Islands (KYD $)
* Central African Republic (XAF Fr)
* Chad (XAF Fr)
* Chile (USD $)
* China (CNY ¥)
* Christmas Island (AUD $)
* Cocos (Keeling) Islands (AUD $)
* Colombia (USD $)
* Comoros (KMF Fr)
* Congo - Brazzaville (XAF Fr)
* Congo - Kinshasa (CDF Fr)
* Cook Islands (NZD $)
* Costa Rica (CRC ₡)
* Côte d’Ivoire (XOF Fr)
* Croatia (EUR €)
* Curaçao (ANG ƒ)
* Cyprus (EUR €)
* Czechia (CZK Kč)
* Denmark (DKK kr.)
* Djibouti (DJF Fdj)
* Dominica (XCD $)
* Dominican Republic (DOP $)
* Ecuador (USD $)
* Egypt (EGP ج.م)
* El Salvador (USD $)
* Equatorial Guinea (XAF Fr)
* Eritrea (USD $)
* Estonia (EUR €)
* Eswatini (USD $)
* Ethiopia (ETB Br)
* Falkland Islands (FKP £)
* Faroe Islands (DKK kr.)
* Fiji (FJD $)
* Finland (EUR €)
* France (EUR €)
* French Guiana (EUR €)
* French Polynesia (XPF Fr)
* French Southern Territories (EUR €)
* Gabon (XOF Fr)
* Gambia (GMD D)
* Georgia (USD $)
* Germany (EUR €)
* Ghana (USD $)
* Gibraltar (GBP £)
* Greece (EUR €)
* Greenland (DKK kr.)
* Grenada (XCD $)
* Guadeloupe (EUR €)
* Guatemala (GTQ Q)
* Guernsey (GBP £)
* Guinea (GNF Fr)
* Guinea-Bissau (XOF Fr)
* Guyana (GYD $)
* Haiti (USD $)
* Honduras (HNL L)
* Hong Kong SAR (HKD $)
* Hungary (HUF Ft)
* Iceland (ISK kr)
* India (INR ₹)
* Indonesia (IDR Rp)
* Iraq (USD $)
* Ireland (EUR €)
* Isle of Man (GBP £)
* Israel (ILS ₪)
* Italy (EUR €)
* Jamaica (JMD $)
* Japan (JPY ¥)
* Jersey (USD $)
* Jordan (USD $)
* Kazakhstan (KZT 〒)
* Kenya (KES KSh)
* Kiribati (USD $)
* Kosovo (EUR €)
* Kuwait (USD $)
* Kyrgyzstan (KGS som)
* Laos (LAK ₭)
* Latvia (EUR €)
* Lebanon (LBP ل.ل)
* Lesotho (USD $)
* Liberia (USD $)
* Libya (USD $)
* Liechtenstein (CHF CHF)
* Lithuania (EUR €)
* Luxembourg (EUR €)
* Macao SAR (MOP P)
* Madagascar (USD $)
* Malawi (MWK MK)
* Malaysia (MYR RM)
* Maldives (MVR MVR)
* Mali (XOF Fr)
* Malta (EUR €)
* Martinique (EUR €)
* Mauritania (USD $)
* Mauritius (MUR ₨)
* Mayotte (EUR €)
* Mexico (USD $)
* Moldova (MDL L)
* Monaco (EUR €)
* Mongolia (MNT ₮)
* Montenegro (EUR €)
* Montserrat (XCD $)
* Morocco (MAD د.م.)
* Mozambique (USD $)
* Myanmar (Burma) (MMK K)
* Namibia (USD $)
* Nauru (AUD $)
* Nepal (NPR ₨)
* Netherlands (EUR €)
* New Caledonia (XPF Fr)
* New Zealand (NZD $)
* Nicaragua (NIO C$)
* Niger (XOF Fr)
* Nigeria (NGN ₦)
* Niue (NZD $)
* Norfolk Island (AUD $)
* North Macedonia (MKD ден)
* Norway (USD $)
* Oman (USD $)
* Pakistan (PKR ₨)
* Palestinian Territories (ILS ₪)
* Panama (USD $)
* Papua New Guinea (PGK K)
* Paraguay (PYG ₲)
* Peru (PEN S/.)
* Philippines (PHP ₱)
* Pitcairn Islands (NZD $)
* Poland (PLN zł)
* Portugal (EUR €)
* Qatar (QAR ر.ق)
* Réunion (EUR €)
* Romania (RON Lei)
* Russia (USD $)
* Rwanda (RWF FRw)
* Samoa (WST T)
* San Marino (EUR €)
* São Tomé & Príncipe (STD Db)
* Saudi Arabia (SAR ر.س)
* Senegal (XOF Fr)
* Serbia (RSD РСД)
* Seychelles (USD $)
* Sierra Leone (SLL Le)
* Singapore (SGD $)
* Sint Maarten (ANG ƒ)
* Slovakia (EUR €)
* Slovenia (EUR €)
* Solomon Islands (SBD $)
* Somalia (USD $)
* South Africa (USD $)
* South Georgia & South Sandwich Islands (GBP £)
* South Korea (KRW ₩)
* South Sudan (USD $)
* Spain (EUR €)
* Sri Lanka (LKR ₨)
* St. Barthélemy (EUR €)
* St. Helena (SHP £)
* St. Kitts & Nevis (XCD $)
* St. Lucia (XCD $)
* St. Martin (EUR €)
* St. Pierre & Miquelon (EUR €)
* St. Vincent & Grenadines (XCD $)
* Sudan (USD $)
* Suriname (USD $)
* Svalbard & Jan Mayen (USD $)
* Sweden (SEK kr)
* Switzerland (CHF CHF)
* Taiwan (TWD $)
* Tajikistan (TJS ЅМ)
* Tanzania (TZS Sh)
* Thailand (THB ฿)
* Timor-Leste (USD $)
* Togo (XOF Fr)
* Tokelau (NZD $)
* Tonga (TOP T$)
* Trinidad & Tobago (TTD $)
* Tristan da Cunha (GBP £)
* Tunisia (USD $)
* Türkiye (USD $)
* Turkmenistan (USD $)
* Turks & Caicos Islands (USD $)
* Tuvalu (AUD $)
* U.S. Outlying Islands (USD $)
* Uganda (UGX USh)
* Ukraine (UAH ₴)
* United Arab Emirates (AED د.إ)
* United Kingdom (GBP £)
* United States (USD $)
* Uruguay (UYU $)
* Uzbekistan (UZS )
* Vanuatu (VUV Vt)
* Vatican City (EUR €)
* Venezuela (USD $)
* Vietnam (VND ₫)
* Wallis & Futuna (XPF Fr)
* Western Sahara (MAD د.م.)
* Yemen (YER ﷼)
* Zambia (USD $)
* Zimbabwe (USD $)
© 2024 Anabolic Aliens
We Accept
American Express Apple Pay Diners Club Discover Meta Pay Google Pay
Mastercard Shop Pay Visa
© 2024 Anabolic Aliens
| biology | 650323 | https://no.wikipedia.org/wiki/Clifford%20Shull | Clifford Shull | Clifford Glenwood Shull (født 23. september 1915 i Pittsburgh, Pennsylvania, død 31. mars 2001) var en fysiker fra USA.
Han fikk Nobelprisen i fysikk i 1994 sammen med Bertram Brockhouse for deres arbeid med nøytroner. Det utgis en pris i hans navn: The Clifford G. Shull Prize in Neutron Physics, fra The Neutron Scattering Society of America.
Priser
Oliver E. Buckley Condensed Matter Prize, American Physical Society, 1956
Innvalgt i American Academy of Arts and Sciences, 1956
Innvalgt i The National Academy of Sciences, 1975
Gregori Aminoff prisen, Kungliga Vetenskapsakademien, 1993
Nobelprisen i fysikk, 1994, sammen med Bertram Brockhouse
Publikasjoner
Davidson, W. L., Morton, G. A., Shull, C. G. & E. O. Wollan. Neutron Diffraction Analysis of NaH and NaD, United States Department of Energy , (28. april 1947).
Shull, C. G.; Wollan, E. O. The Diffraction of Neutrons by Crystalline Powders, Oak Ridge National Laboratory (ORNL),United States Department of Energy (28. april 1947).
Shull, C.G., Wollan, E.O. & M.C. Marney. Neutron Diffraction Studies, Oak Ridge National Laboratory (ORNL),United States Department of Energy (22. oktober 1948).
Rundle, R.E., Shull, C.G. & E.O. Wollan. The Crystal Structure of Thorium and Zirconium Dihydrides by X-ray and Neutron Diffraction, Ames Laboratory, Oak Ridge National Laboratory (ORNL), United States Department of Energy (20. april 1951).
Nathans, R., Riste, T., Shirane, G. & C.G. Shull. Polarized Neutron Studies on Antiferromagnetic Single Crystals: Technical Report No. 4, Massachusetts Institute of Technology (MIT), Brookhaven National Laboratory, United States Department of Energy, National Security Agency (NSA), Air Force Office of Scientific Research (AFOSR), (26. november 1958).
Shull, C.G. Low Temperature and Neutron Physics Studies: Final Progress Report, March 1, 1986--May 31, 1987, Massachusetts Institute of Technology (MIT), United States Department of Energy, (27. juli 1989).
Referanser
Eksterne lenker
Fysikere fra USA
Kjernefysikere
Nobelprisvinnere fra USA
Nobelprisvinnere (fysikk)
Personer fra Pittsburgh
Fødsler i 1915
Dødsfall i 2001 | norwegian_bokmål | 1.08951 |
shower_muscles/cold-shower-vs-hot-shower.txt | * Health Conditions
* Featured
* Breast Cancer
* IBD
* Migraine
* Multiple Sclerosis (MS)
* Rheumatoid Arthritis
* Type 2 Diabetes
* Sponsored Topics
* Articles
* Acid Reflux
* ADHD
* Allergies
* Alzheimer's & Dementia
* Bipolar Disorder
* Cancer
* Crohn's Disease
* Chronic Pain
* Cold & Flu
* COPD
* Depression
* Fibromyalgia
* Heart Disease
* High Cholesterol
* HIV
* Hypertension
* IPF
* Osteoarthritis
* Psoriasis
* Skin Disorders and Care
* STDs
* Discover
* Wellness Topics
* Nutrition
* Fitness
* Skin Care
* Sexual Health
* Women's Health
* Mental Well-Being
* Sleep
* Product Reviews
* Vitamins & Supplements
* Sleep
* Mental Health
* Nutrition
* At-Home Testing
* CBD
* Men’s Health
* Original Series
* Fresh Food Fast
* Diagnosis Diaries
* You’re Not Alone
* Present Tense
* Video Series
* Youth in Focus
* Healthy Harvest
* Through An Artist's Eye
* Future of Health
* Plan
* Health Challenges
* Mindful Eating
* Sugar Savvy
* Move Your Body
* Gut Health
* Mood Foods
* Align Your Spine
* Find Care
* Primary Care
* Mental Health
* OB-GYN
* Dermatologists
* Neurologists
* Cardiologists
* Orthopedists
* Lifestyle Quizzes
* Weight Management
* Am I Depressed? A Quiz for Teens
* Are You a Workaholic?
* How Well Do You Sleep?
* Tools & Resources
* Health News
* Find a Diet
* Find Healthy Snacks
* Drugs A-Z
* Health A-Z
* Connect
* Find Your Bezzy Community
* Breast Cancer
* Inflammatory Bowel Disease
* COPD
* Migraine
* Multiple Sclerosis
* Psoriasis
* Follow us on social media
Healthline
* Health Conditions
* Discover
* Plan
* Connect
Subscribe
# Cold Showers vs. Hot Showers: Which One Is Better?
Medically reviewed by Alana Biggers, M.D., MPH — By Sara Lindberg —
Updated on March 21, 2023
Both cold and hot showers have pros and cons for your health. Cold showers may
help prevent dry skin while hot showers can help your muscles feel relaxed.
If a hot shower is what your body craves in the morning, you’re not alone. The
majority of people crank the handle all the way up in order to feel the warm
water all over their body.
But did you know that cold showers should also have a place in your daily
routine?
That’s right — cold showers. The ones you dread to take when you’re the last
person to get up in the morning. But if you give them a fair chance, you might
find that you actually like how you feel after taking one.
Regardless of how you feel about either type of shower, research shows that
both hot and cold showers have health benefits you should be aware of.
##
What’s so great about cold showers?
Benefits of taking a cold shower include:
* calming itchy skin
* waking you up
* increasing circulation
* reducing muscle soreness post-workout
* potentially boosting weight loss
* glowing hair and skin
### Cold showers calm itchy skin
Adam Friedman, MD , says if you have itchy skin or skin conditions that
cause you to itch, cold showers can help you overcome the sensation to
scratch.
### Cold showers help you wake up in the morning
When that cold spray hits your body, there’s a bit of shock. This shock
increases:
* oxygen intake
* heart rate
* alertness
### Cold showers increase your circulation
Increased circulation is one of the top reasons experts recommend cold
showers.
As cold water hits your body and external limbs, it constricts circulation on
the surface of your body. This causes blood in your deeper tissues to
circulate at faster rates to maintain ideal body temperature .
In that sense, a cold shower has the opposite effect of a hot shower for
someone with hypertension or cardiovascular disease , since exposure to
cold temperatures triggers the circulatory system to reduce inflammation and
can help prevent cardiovascular disease.
### Cold showers help reduce muscle soreness after intense workouts
Since cold water has regenerative properties, your muscles will relax and
repair after a tough workout.
### Cold showers may help boost weight loss
Some fat cells, such as brown fat , can generate heat by burning fat. They
do this when your body is exposed to cold conditions like in a shower.
Gerrit Keferstein, MD, says these cells are mostly situated around the neck
and shoulder area. So, perfect for showers!
### Cold showers give your skin and hair a healthy glow
Though scientific research is limited regarding the effect cold water has on
your skin and hair, anecdotal evidence points to positive effects.
Wellness expert Dr. Jacqueline Schaffer, MD , says that cold water tightens
and constricts the blood flow which gives your skin a healthier glow .
According to an article published on the website NaturallyCurly.com , cold
water closes and strengthens your hair cuticles.
Also, cold water, unlike hot water, doesn’t dry out the sebum layer, a
naturally lubricated barrier that provides protection for your skin and hair.
As a result of the effects of cold water, your hair may be more likely to
become stronger and healthier over time.
If you’re convinced a cold shower is totally out of the question, you might
want to rethink your philosophy. Unlike the long list of benefits that come
with taking a cold shower, the list of cons is surprisingly quite short.
#### The cons of cold showers:
* Cold showers might not be a good idea if you’re already cold, since the cooler temperature isn’t going to help warm you up by any means. It could actually make you even colder and increase the amount of time it will take for your body to warm back up.
* They may not be a good idea if you’re sick, either. Initially, the cold temperature might be too hard on your immune system, so it’s best to ease into the cooler temperatures.
##
Why do we like hot showers?
If you have trouble relaxing or falling asleep at night, you might be tempted
to take a hot shower to ease the stress of the day.
This is a common practice for muscle relaxation before going to sleep because
hot showers activate the parasympathetic nervous system which makes us tired,
says Keferstein.
Other benefits of hot showers include:
* providing relief from respiratory symptoms
* helping with blemishes
* helping with muscle relaxation
### Hot showers provide relief from cold or respiratory symptoms
Standing in a hot shower with the steam surrounding you has long been used as
a natural remedy to reduce cold and cough symptoms. The heat from the water
and the steam can help to:
* open airways
* loosen up phlegm
* clear out your nasal passages
### Hot showers help with blemishes
Hot showers can help open up the pores of the skin, which allows you to clean
out the trapped dirt and oil.
### Hot showers are good for muscle relaxation
Being in hot water effectively helps relieve body tension and can help soothe
muscle fatigue.
But, yes, beloved a hot shower does have some downsides.
However, the good news is, you don’t have to give them up completely. You just
need to turn down the temperature a bit and take care of your skin afterward.
#### The cons of hot showers include:
* Hot showers can dry out and irritate your skin. Schaffer says the hot water causes damage to the keratin cells that are located on the most outer layer of our skin — the epidermis. By disrupting these cells, it creates dry skin and prevents the cells from locking in moisture.
* They can also make certain skin conditions worse. Higher temperatures make it easier for the skin to dry out and worsen conditions like eczema .
* Hot showers can cause you to itch. Friedman says the heat can cause mast cells (which contain histamine) to release their contents in the skin and cause itching.
* They can increase your blood pressure, too. If you have problems with high blood pressure or cardiovascular disease, taking a shower that’s too hot can make these conditions worse.
##
So, which type is better?
There are obvious benefits to both hot and cold showers, so what should you
do?
Well, in an ideal world, Friedman says you should take a lukewarm shower — so
it’s tolerable — and apply a moisturizer to damp skin after bathing.
Another approach to try is what Keferstein describes as a contrast shower,
which is an age-old technique Dr. Sebastian Kneipp developed.
Basically, you get the water as cold as possible and stand in it for one
minute. When the minute is up, you then change the water to as hot as you can
handle for an additional minute.
Alternate between one minute each of cold and hot for three to five cycles.
He said the health benefits come from the cold water constricting the blood
vessels. This means all the blood will go to the middle of the body.
The hot water will open the blood vessels and all the blood comes rushing out
again. This can pump the blood completely through the muscles and organs and
is great for helping with regeneration and detoxification.
Last medically reviewed on March 23, 2020
### How we reviewed this article:
Sources
History
Healthline has strict sourcing guidelines and relies on peer-reviewed studies,
academic research institutions, and medical associations. We avoid using
tertiary references. You can learn more about how we ensure our content is
accurate and current by reading our editorial policy .
* Cypess AM, et al. (2009). Identification and importance of brown adipose tissue in adult humans. DOI:
https://doi.org/10.1056/NEJMoa0810780
* Friedman A. (2018). Personal interview.
* Keferstein G. (2018). Personal interview.
* Mooventhan A, et al. (2014). Scientific evidence-based effects of hydrotherapy on various systems of the body. DOI:
https://dx.doi.org/10.4103%2F1947-2714.132935
* Schaffer J. (2018). Personal interview.
Our experts continually monitor the health and wellness space, and we update
our articles when new information becomes available.
Current Version
Mar 21, 2023
Written By
Sara Lindberg
Copy Edited By
Delores Smith-Johnson
Mar 23, 2020
Medically Reviewed By
Alana Biggers, MD, MPH
Share this article
Medically reviewed by Alana Biggers, M.D., MPH — By Sara Lindberg —
Updated on March 21, 2023
### Read this next
* How a Cold Shower Before Bed Affects Your Sleep
Medically reviewed by Deborah Weatherspoon, Ph.D., MSN
Showering before bed may be part of your overall better-sleep strategy, but
there isn’t a lot of research to support the idea that a cold shower is…
READ MORE
* * Cold Shower Benefits for Your Health
Medically reviewed by Debra Rose Wilson, Ph.D., MSN, R.N., IBCLC, AHN-BC, CHT
READ MORE
* Is It Better to Take a Bath or a Shower?
Medically reviewed by Deborah Weatherspoon, Ph.D., MSN
Your personal hygiene priorities should determine whether you take a bath or a
shower. Here are the benefits of each one.
READ MORE
* What to Know About Cold Water Therapy
Medically reviewed by Gregory Minnis, DPT
Cold water therapy may benefit your health in several ways. This type of
therapy can include cold showers, ice baths, or swimming in cold water. Learn…
READ MORE
* Should You Wash Your Face with Cold Water?
Washing your face with cold water does have some benefits, but warm water may
be best.
READ MORE
* How to Prepare for a Blood Test
Medically reviewed by Jenneh Rishe, RN
Following your doctor’s instructions is the most important part of preparing
for a blood test. Other tips include sleeping well, staying hydrated, and…
READ MORE
* What Is Extracorporeal Membrane Oxygenation?
Extracorporeal Membrane Oxygenation (ECMO) bypasses the heart and lungs of
critically ill people. Learn how it works, how long it's used, and what to…
READ MORE
* What You Need to Know About Abdominal Swelling
Medically reviewed by Saurabh Sethi, M.D., MPH
A swollen abdomen is sometimes known as a distended abdomen or swollen belly.
A swollen abdomen is often uncomfortable or even painful, but it’s a…
READ MORE
* Everything You Need to Know About Magnetic Resonance Imaging (MRI) Scans
Medically reviewed by Jenneh Rishe, RN
An MRI scan is a noninvasive medical test that uses magnets, radio waves, and
a computer to produce images of soft tissues.
READ MORE
* What Is Immunohistochemistry Staining?
Learn why immunohistochemistry is the gold standard in testing for certain
diseases, including several cancers.
READ MORE
* About Us
* Contact Us
* Privacy Policy
* Privacy Settings
* Advertising Policy
* Health Topics
* Sitemap
* Medical Affairs
* Content Integrity
* Newsletters
* © 2024 Healthline Media LLC. All rights reserved. Our website services, content, and products are for informational purposes only. Healthline Media does not provide medical advice, diagnosis, or treatment. See additional information. See additional information .
© 2024 Healthline Media LLC. All rights reserved. Our website services,
content, and products are for informational purposes only. Healthline Media
does not provide medical advice, diagnosis, or treatment. See additional
information. See additional information .
About Careers Advertise with us
OUR BRANDS
Healthline Medical News Today Greatist Psych Central Bezzy
| biology | 497513 | https://no.wikipedia.org/wiki/Kroppstemperatur | Kroppstemperatur | Kroppstemperaturen hos mennesket er normalt rundt 37 ℃. Mennesket kan holde den temmelig stabil til tross for betydelige endringer i temperaturen i omgivelsene. Det er nødvendig for at kroppen skal fungere normalt. Altfor høy eller altfor lav temperatur kan likevel være direkte skadelig for kroppen. Når vi blir syke øker temperaturen.
Regulering
Kroppstemperaturen styres av et senter i hjernen som fungerer som en fininnstilt termostat. Det viktigste den gjør for at kroppen skal holde temperaturen normal, er kanskje å få mennesket til å føle at det fryser når det er kaldt, slik at mennesket kler på seg, og føle varmen når det er varmt, slik at mennesket kler av seg. Men kroppen har også andre måter å regulere varmetapet og varmeproduksjonen på. Dette styres også av «termostaten» i hjernen.
Huden spiller en viktig rolle i temperaturreguleringen, sammen med blodet. Når blodet strømmer gjennom muskler og de indre organer, varmes det opp. Når blodet strømmer gjennom huden, kjøles det ned igjen. Huden kan nemlig tape varme til omgivelsene på mange forskjellige måter. Jo mer blod som strømmer gjennom huden, desto større er varmetapet.
Når det er kaldt, må varmetapet begrenses. Da bør lite blod strømme gjennom huden. Derfor trekker ringformede snøremuskler i blodårene i huden seg sammen og reduserer blodtilførselen til huden. Fettet i underhuden virker som isolasjon. I tillegg strammes små muskler som er festet til dunhårene, slik at de reiser seg. Dette kalles gjerne gåsehud. Håret reiser seg når det er kaldt, for å lage et stabilt, isolerende luftlag rundt kroppen. Dette er viktig hos dyr med pels. Menneskene har så lite hår på kroppen at det bare er på hodet at håret har betydning som beskyttelse mot kulde.
Hvis kroppen taper mer varme enn den produserer, må den øke varmeproduksjonen. Kroppen har liten mulighet til å øke forbrenningen, og dermed varmeproduksjonen, i hvile. Derimot kan den øke aktiviteten. Da øker også varmeproduksjonen. Mennesket kan for eksempel bevege seg, slå floke eller bruke musklene på annen måte. Dessuten kan «termostaten» i hjernen sette i gang raske, små bevegelser i skjelettmusklene. Dette kalles skjelving. Skjelvingen øker kroppens varmeproduksjon mange ganger i løpet av kort tid. Den er ikke viljestyrt.
Når det er varmt rundt kroppen, må den sørge for at kroppstemperaturen holdes i balanse, ved å tape nok varme til omgivelsene. Da slappes musklene rundt blodårene i huden av, slik at blodtilførselen øker. Dermed kan mye varme passere fra blodet ut gjennom huden. Dunhårene ligger inntil huden, slik at det ikke skal bli noe isolerende luftlag. Når luft som varmes opp av huden, raskt skiftes ut (for eksempel når det blåser), kan varmetapet bli stort. Svetteproduksjonen øker i varmen. Når vannet i svetten fordamper, avkjøles huden mer effektivt, og mennesket taper mer varme.
Kroppen klarer å holde kroppstemperaturen i balanse hos en person uten klær, selv om lufttemperaturen varierer fra 15 til 55 grader, så lenge luften er tørr.
Fysiologi
Temperatur | norwegian_bokmål | 0.757596 |
shower_muscles/post-workout-recovery-shower.txt | Skip to content
Enjoy free shipping over $40.
* Shop
Shop All
Smart showerhead
Fuse
Repair infusion
Relax infusion
Energy Boost Infusion
Discovery Set
* Our Story
* Our Impact
* Search
Search
* Account
* Cart 0
0
Search
* Shop
Smart showerhead
Repair infusion
Relax infusion
Energy Boost Infusion
Discovery Set
Fuse
Shop All
* Our Story
* Our Impact
Log in
* Read the blog
* Twitter
* Facebook
* Instagram
* TikTok
## Item added to your cart
Check out Continue shopping
Share on Facebook Tweet on Twitter Pin on Pinterest Share on Tumblr Share
via Email
# How to Fuel Your Post-Workout Recovery with a Shower
In the world of sports, there's a phrase that's as common as a high-five after
a victory – "Hit the showers." Usually hollered by a coach to players post-
practice or game, this proclamation tends to be the universal signal for one
thing in particular: recovery.
Chances are, you're already familiar with the post-workout shower's general
benefits – plain and simple: it gets you clean. But beyond preventing the
accumulation of sweat-induced dirt and bacteria, there is research around the
notion of ‘contrast showering’ that supports the shower’s role as a recovery
tool. Some experts suggest that showers can potentially alleviate muscle
tension due to their impact on blood circulation, vouching for the benefits of
using either hot or cold water during their cleansing routine.
So, hit the showers and get your recovery started.
## Why shower after a workout for recovery?
After an intense workout, our muscles undergo stress and micro-tears, a
natural part of the muscle-building process. During the post-workout period,
our bodies initiate the recovery phase, where damaged muscle fibers are
repaired and strengthened. This repair process is crucial for muscle growth
and improved performance. One of the best ways to facilitate this recovery
right at home is through contrast showers or alternating between hot and cold
water.
What is Contrast Showering?
Contrast showering, also known as contrast hydrotherapy , is a technique
where an individual alternates between hot and cold water during a shower.
This practice is believed to have various benefits for the body, particularly
in terms of promoting circulation, reducing muscle soreness, and potentially
boosting recovery after exercise.
The typical pattern involves switching between hot and cold water multiple
times. The process can vary, but a common approach is to start with warm or
hot water for a few minutes, then switch to cold water for a shorter duration,
typically around 30 seconds to a minute. This cycle is repeated several times,
ending with cold water.
Advocates for contrast showering and hydrotherapy suggest that hot water
helps dilate blood vessels and improve blood flow, while cold water
constricts blood vessels, reducing inflammation and potentially aiding in the
removal of waste products from muscles. This contrast between hot and cold
temperatures is believed to stimulate the body's natural responses and promote
a sense of rejuvenation.
Benefits of cold post-workout showers
In a 2014 review spanning multiple studies, it was discovered that cold
water immersion not only alleviates soreness and inflammation but also imparts
a revitalizing boost to nearly every system in your body.
This advantage positions a post-workout cold shower as a great way to amplify
the endorphin rush from your exercise, potentially reducing post-training
muscle discomfort.
\- Elevates endorphin release
\- Boosts metabolic performance
\- Improves circulation
\- Strengthens the immune system
Benefits of hot post-workout showers:
Indulging in a warm and comforting shower enhances blood circulation,
rejuvenates cells, and promotes relaxation post rigorous exercise.
\- Loosens up stiff muscles
\- Reduces anxiety
\- Clears nasal cavities
\- Promotes healthy skin by clearing toxins
## The Benefits of Showering Post-Workout
Training and competition create an overload to stress the body. What everyone
from weekend warriors to pro athletes do after their exercise and workout
regime can affect their muscle recovery.
Muscle Recovery and Relief:
Post-workout showers can be your secret weapon when it comes to relieving
muscle soreness and stiffness. The warm water helps dilate blood vessels,
enhancing blood circulation and promoting the delivery of vital nutrients to
your muscles. This process aids in the removal of metabolic waste products,
like lactic acid, which accumulates during exercise and contributes to that
familiar post-workout ache.
Soothing Inflammation:
The aftermath of an intense workout can lead to micro-inflammation within your
muscles. A post-workout shower, especially one that alternates between warm
and cold water, triggers a process known as vasoconstriction and vasodilation.
This phenomenon helps reduce inflammation, supporting the body's natural
healing mechanisms.
Mental Relaxation and Stress Relief:
Physical recovery is not the only benefit a post-workout shower offers.
Exercise can be mentally taxing, especially if you've had a demanding day. The
shower can be a sanctuary for relaxation, providing a moment of solitude and
tranquility. The warm water soothes the nervous system, releasing tension and
promoting a sense of calmness. This not only helps in recovery but also
improves sleep quality, a crucial aspect of overall fitness and well-being.
How to optimize your post-workout shower with hai
Optimize your cool-down routine with the hai smart showerhead for the ultimate
post-workout rejuvenation.
Spray control: Bring the gym sauna home to you with our spa-like mist spray,
a blissful way to decompress after a hard workout. The warm steam helps relax
muscles, soothe joints, and alleviate tension. Beyond the physical, the
tranquil environment encourages mental relaxation, reduces stress, and
provides a meditative escape from the demands of daily life.
Handheld shower: For alleviating discomfort in joints like the shoulder,
elbow, wrist, knee, hip, and ankle, direct our potent massage spray precisely
onto the targeted joint region. Hold it for up to 2 minutes, gently performing
small rotations. This technique aims to ease soreness and tension, offering
relief to the affected joint area. This will also help increase blood flow to
the area.
Temperature notifications & LED light alerts: Whether you’re a cold plunger
or sauna goer, set your ideal temp in the hai app and our LED light will let
you know when your shower is ready to be hopped into. No more waiting around
for your water to warm up—we’ll notify you via app notification when your
ideal temp is reached.
Vitamin and aromatherapy-rich shower infusions: Designed specifically for
your post-workout shower, our repair shower infusion jumpstarts your recovery
by infusing eucalyptus, citric acid, and vitamin D3 into your shower’s water
stream. Incorporating essential oils like eucalyptus into your post-workout
shower provides both mental and physical benefits. The invigorating aroma
promotes mental clarity and relaxation, while its natural properties help open
airways and enhance respiratory function. Eucalyptus also offers a cooling
sensation, soothing muscles, and contributing to a refreshing after-workout
experience.
Shop Repair
When is it best to take a shower post-workout?
Just finished a HIIT class, bike ride, or Pilates session? It's shower time!
But hold on a second before you hop in. If you usually rush into the shower
post-workout, consider giving your tired body a brief pause. Your body
typically takes around five minutes to cease sweat production. If you step
into the shower during this window, it's advisable to switch to cold water for
a bit. This helps as your pores and veins, having expanded during exercise,
can now close back up during this cooling phase.
Elevate your post-workout routine by adding a refreshing shower to your
arsenal. Beyond the physical cleanse, a well-timed shower can promote muscle
recovery, enhance circulation, and provide a rejuvenating mental break after
intense exercise, setting the stage for a more balanced and revitalized you.
So go, hit the showers.
Back to blog
## Keep Reading
##
12.12.23
### Celebrating hai's Award-Winning Smart Showerhead
hai is making waves in the world of home innovation, proving itself as an
industry innovator.
12.12.23
### 6 Stretches You Can Do in the Shower
A few simple shower stretches to get your body moving.
12.12.23
### How to Clean Your hai Smart Showerhead
Simple tips and tricks to make cleaning your hai a breeze.
12.12.23
### World Water Day 2023: hai Announces Partnership with Clean the World
Foundation
Clean the World Foundation is leading a global hygiene revolution.
#### Join the hai community
Email
Subscribe
* Shop
* Our Story
* The Source
* Ambassadors
* Trade Program
* Contact
* FAQ
* Privacy Policy
* Terms and Conditions
* Terms of Service
* Refund Policy
* * * Twitter
* Instagram
* Facebook
* Screenshot_2 Translation missing: en.general.social.links.spotify
* TikTok
Cart
YOUR CART IS EMPTY. Continue
Spends $ more and receive Free shipping
Shipping
Free shipping over $40
Subtotal
Checkout
Best deal!
Upgrade model
* Custom spray
* Unique color
* Water-powered smart tech
* Free app membership
* Fuse add-on
* Infusions-ready
Update Cart
* Choosing a selection results in a full page refresh.
Apr 29, 2024 22:32
Product Tag :
Product Collection :
| biology | 396181 | https://da.wikipedia.org/wiki/Sportsdrik | Sportsdrik | Sportsdrik er en drik som indeholder næringsstoffer. Den bruges ofte af sportsfolk under træning og konkurrencer, med det formål at erstatte den væske og de mineraler, som tabes under hård fysisk udfoldelse, og tilføre energi i form af f.eks. kulhydrater. Der findes en lang række kommercielle produkter, som indeholder bl.a. husholdningssalt og sukker, og ofte fås de som et pulver der skal blandes op i vand.
Se også
Energidrik | danish | 0.876497 |
shower_muscles/.txt | Get ` 3 months of Peloton App One or App+ for the price of 1, ` starting at
$12.99. Offer ends 5/1. Terms apply. Explore App
Train Recover Level Up Connect Shop
Train
* Bike
* Row
* Run
* Sculpt
* Strength Train
* Beginners
* Yoga
* Cardio
Recover
* Nutrition
* Meditation
* Sleep
* Stretching + Mobility
Level Up
* Mindset
* Mental Health
* Health
* Marathon + Racing
* Equipment
Connect
* Member Tips
* Member Stories
* Meet Instructors
* Inspiration
* Peloton News
Social
Lusyaya / iStock / Getty Images Plus via Getty Images
1. Home
2. Level Up
3. Health
# Should You Take a Hot or Cold Shower After a Workout?
This is why your post-exercise shower temperature doesn’t matter too much.
By Michele Ross • March 4, 2024
* Stretching + Mobility
Share:
In this article
* Does It Matter Whether You Take a Hot or Cold Shower After a Workout?
* The Benefits of Taking a Hot Shower After a Workout
* The Benefits of Taking a Cold Shower After a Workout
* How to Decide Between a Hot or Cold Shower After a Workout
* The Takeaway
* * *
Whether you’re a casual exerciser or a seasoned athlete, chances are you
wouldn’t mind adopting a simple hack or two to help your body recover faster.
Some people particularly swear by taking a hot or cold shower after a
workout—or even alternating between the two extremes—to try to expedite the
recovery process.
But does the temperature of your shower really make that much of a
difference for your post-exercise rest and recuperation? We spoke with experts
and dug into the latest research to learn more.
## Does It Matter Whether You Take a Hot or Cold Shower After a Workout?
There’s no direct proof indicating that hot or cold showers will make or break
your post-workout recovery, says Corey Simon, PhD , an associate professor
in the department of orthopaedic surgery at Duke University, a senior fellow
in the Duke Aging Center, and a physical therapist.
In many cases, the perks are anecdotal and mainly proliferate on social media
rather than peer-reviewed journals. “I see multiple posts per week on cold
showers and hot-to-cold contrast showers, touting therapeutic effects that
include faster workout recovery, better skin, etc.,” Simon says. However, he’s
yet to see these claims backed by scientific evidence.
In case you’ve heard otherwise—with sources claiming that research does, in
fact, support the notion that shower temperature affects post-workout
recovery—it’s worth taking a closer look at the quality of the referenced
studies. They may be decades old or include small sample sizes, and they may
not be highly controlled or specific to exercise, Simon explains. “This means
that the findings may be a product of a number of factors aside from the
intervention itself,” he says.
### Related Articles
#### Health
4 Ways Infrared Saunas Can Give Your Mind and Body a Boost
#### Stretching + Mobility
Could a Cold Plunge Speed Up Your Workout Recovery?
#### Stretching + Mobility
Still Sore Days After Your Workout? Learn How to Relieve DOMS Pain
For instance, Simon often sees one 2016 study cited in which participants
who took hot-to-cold contrast showers for 30, 60, or 90 seconds daily for 30
days noted fewer self-reported sick days at work compared to a control group.
However, there are a few gaps in this study:
* Only the sick days from work were logged—not actual days of illness.
* The sample groups weren’t blinded, so a placebo effect could have played a role.
* Even if the benefits of hot-to-cold contrast showers were legitimate for staving off illness, the findings don’t correspond to post-workout recovery.
Plus, most of the research on hot or cold therapy as it relates to fitness
focuses on cold water immersions , cryotherapy, saunas , and other
environments that don’t mimic the exact conditions of a casual shower after
exercising. While some of these recovery methods may have their merits, the
common denominator of temperature doesn’t mean that a hot or cold shower after
a workout will inevitably offer the same recovery perks.
So why isn’t shower temperature all that powerful of a muscle recovery tool?
“The tissue most affected by thermal interventions is the skin, and skin
temperature is poorly correlated with muscle temperature,” Simon explains.
“Add adipose tissue (fat) and the correlation is even worse.” In other words,
changing the temperature of your skin with a hot or cold shower after a
workout won’t necessarily translate to temperature changes in your
muscles—meaning it probably won’t have a big impact on post-exercise recovery.
(Cold water immersion, on the other hand, may reduce delayed onset muscle
soreness, aka DOMS , by way of temporarily restricting blood flow, allowing
oxygenated blood to circulate after exposure.)
All that said, while your muscle recovery likely won’t be impacted by a cold
or hot shower after a workout, your post-exercise shower temperature can
offer other potential wellness benefits.
## The Benefits of Taking a Hot Shower After a Workout
Some studies suggest exercise-related benefits of hot water immersion via
baths, from increased blood flow to improved endurance while training in
the heat. However, it’s worth reiterating that this doesn’t mean that hot
showers will yield the same results.
Taking a hot shower after working out might not directly expedite recovery—but
it can potentially complement your exercise regimen and overall well-being.
Here are a couple of potential benefits of the practice:
### Less Stress
A hot shower won’t only help you wash away your sweat and grime, but it can
also help lower your stress levels. “Those who have taken a warm shower after
stressful days or high-impact workouts can attest to its calming effects,”
Simon says. “The negative impact of stress on health is well established, as
are the positive health benefits from its removal.”
### Better Sleep
Getting enough high-quality shuteye is crucial, day in and day out. Per a
2019 entry in Sleep Medicine Reviews , even a 10-minute hot shower, bath,
or foot bath taken an hour or two before bedtime can improve sleep. Better
sleep is only one component of post-workout recovery, but a crucial one
nonetheless.
## The Benefits of Taking a Cold Shower After a Workout
Showers aside, there’s more evidence on the benefits of thermal interventions
such as cold water immersion and whole- or partial-body cryo-stimulation ,
particularly for improving muscle soreness, Simon explains. Still, cold
showers may stand to benefit some people after exercise, though more robust
evidence is needed.
With that in mind, here are a couple of possible benefits of taking a cold
shower after a workout:
### Less Stress
Similar to hot showers, cold showers may also reduce stress levels—so long as
it’s paired with breathwork. Per a 2022 randomized trial published in the
journal Current Psychology , participants who paired pranayama-style
breathing exercises with a 15- to 60-second cold shower daily over 14 days
reported lower stress levels compared to a control group. “However,
participants performing breathing only, or cold shower only, also showed
improved stress levels; and the breathing-only group had larger effects than
the cold-shower-only group,” Simon explains.
In short, a cold shower might help you feel less stressed out—but mindfulness
practices may be even more effective.
### Feel-Good Chemical Release
Braving colder temps will test your limits and push you out of your comfort
zone. Overcoming the discomfort can garner a sense of accomplishment,
potentially eliciting a placebo effect that can make you feel powerful. Simon
is careful to note that this placebo effect isn’t “nothing” and can actually
have therapeutic effects: Even expecting an intervention to work can influence
the release of feel-good chemicals in your body, he says.
## How to Decide Between a Hot or Cold Shower After a Workout
More high-quality studies are needed to determine if taking a cold or hot
shower (or switching between the two) after a workout can actually promote
healing and recovery. With that in mind, you can simply stick to whichever
shower temperature you prefer most.
Feeling adventurous? You can test the waters (literally) with cold showers to
see if a placebo effect takes hold, or try out more vetted types of cold
exposure therapy such as cold plunging. “As mentioned, cold water immersion or
body cryo-stimulation show promise for reducing muscle soreness ,” Simon
reiterates. However, he adds that more high-quality research is needed to
gauge safety and efficacy for people of different ages, health statuses,
levels of physical fitness, and the like. (At the same time, you’ll want to
avoid going overboard, as excessive cold exposure may run counter to your
goals. For instance, “there is a growing sentiment among researchers that
using cold interventions for too long or too frequently can actually delay
healing,” Simon says.)
If recovery is your main goal, experts recommend prioritizing other aspects of
your regimen besides your post-exercise shower temperature. “My advice would
be to worry about things such as following a proper training program, eating
well, and resting, rather than the temperature of your shower,” says Jasmine
Marcus , a physical therapist based in Ithaca, New York. To alleviate sore
muscles after a workout , you can also benefit from the likes of cooldown
exercises , foam rolling , and massage. Staying hydrated and
incorporating rest days is crucial, too.
## The Takeaway
Whether you prefer a hot or cold shower after a workout, the water temperature
probably won’t impact your exercise recovery timeline much. While other
thermal interventions such as cold plunging or cryotherapy have been shown to
improve muscle recovery, your shower temperature probably won’t have a big
impact. That said, cold or hot showers after a workout may offer other
potential wellness benefits, such as less stress or better Zzzs. But if muscle
recovery is your top priority, experts advise focusing on other recovery best
practices, such as making time for rest, staying hydrated, fueling your body
with healthy foods, and following an appropriate training program.
Try the Peloton App for Free
This content is for informational and educational purposes only and does not
constitute individualized advice. It is not intended to replace professional
medical evaluation, diagnosis, or treatment. Seek the advice of your physician
for questions you may have regarding your health or a medical condition. If
you are having a medical emergency, call your physician or 911 immediately.
Share:
## Get our latest health stories straight to your inbox
Enter your email to get articles, expert-backed tips, and updates from Peloton
sent to your inbox.
Email
Sign Up
By providing your email address, you agree to receive marketing communications
from Peloton.
For more about how we use your information, see our Privacy Policy.
Learn
* Instructors
* Membership
* Bike Rental
* Home Trial
About
* Company
* Company Team
* Careers
* Press
* Global
* Investors
* Our Pledge
Visit Us
* Showrooms
* Commercial Bike
* Hotel Finder
* Booking
Support
* Contact Peloton
* Member Support Center
* Return Policy
* Warranties & Protection Plans
* Shipping
* Product Recalls
* Security
Sign up to get the latest
Email
Submit
By providing your email address, you agree to receive marketing communications
from Peloton.
For more about how we use your information, see our Privacy Policy.
SALES: 1-866-650-1996 Visit Support Center
* Privacy Policy
* Terms of Service
* Membership Terms
* IP Policy
* Cookie Settings
* Accessibility
* California Transparency Act
* © Peloton 2012-2023, Peloton Interactive, Inc. All rights reserved.
| biology | 4525249 | https://sv.wikipedia.org/wiki/V%C3%A4rmeutmattning | Värmeutmattning | Värmeutmattning är ett värmerelaterad sjukdomstillstånd som beror på uttorkning och/eller saltbrist vid mycket varmt väder. Det yttrar sig till en början som ymnig svettning och snabb puls till följd av kroppslig överhettning.
Värmeutmattning kan betraktas som ett förstadium till värmeslag, dock kan värmeslag utvecklas hastigt utan att personen först drabbats av värmeutmattning. Värmeutmattning har diffusa och allmänna symtom, såsom illamående, mindre urinmängd (oliguri), mörkare urin, anorexi, trötthet, samt tecken på begynnande cirkulationssvikt såsom något förhöjd kroppstemperatur (hög temperatur tyder däremot på värmeslag), snabb puls, blodtrycksfall, suddig syn, svimningskänslor och blekhet eller rodnad. Utmattning kan för många hänga samman med att inte ha druckit tillräckligt vid hett väder eller ansträngande fysiska aktiviteter. Vid saltbrist märks förutom föregående yrsel. Saltbrist är extra allvarligt eftersom det kan leda till krampanfall (värmekramp), och är dödligt om det inte behandlas.
Risk för värmeutmattning finns när personer inte dricker tillräckligt, och när personer frivilligt utsätter sig för hyperhidros (t.ex. solbadning vid varmt väder, bastubad). Det är lättare att drabbas av värmeutmattning vid hög luftfuktighet, hastiga värmeomslag eller klimatbyte, eller fysisk aktivitet. Vissa sjukdomar och mediciner gör personer mindre värmetåliga och ökar risken för värmeutmattning. Barn, äldre och personer med funktionsnedsättningar har svårt att själva reglera de faktorer som utlöser tillståndet. De är också fysiologiskt sett sämre anpassade att reglera den inre temperaturen.
Förutom vätskeersättning behöver kroppen kylas ner och omedelbar vila. Tillståndet kan behöva sjukvård för att häva ett begynnande värmeslag och andra potentiellt dödliga konsekvenser av de ingående faktorerna.
Noter
Värmerelaterade sjukdomstillstånd | swedish | 0.837137 |
shower_muscles/Vasodilation.txt |
Vasodilation, also known as vasorelaxation, is the widening of blood vessels. It results from relaxation of smooth muscle cells within the vessel walls, in particular in the large veins, large arteries, and smaller arterioles. Blood vessel walls are composed of endothelial tissue and a basal membrane lining the lumen of the vessel, concentric smooth muscle layers on top of endothelial tissue, and an adventitia over the smooth muscle layers. Relaxation of the smooth muscle layer allows the blood vessel to dilate, as it is held in a semi-constricted state by sympathetic nervous system activity. Vasodilation is the opposite of vasoconstriction, which is the narrowing of blood vessels.
When blood vessels dilate, the flow of blood is increased due to a decrease in vascular resistance and increase in cardiac output. Vascular resistance is the amount of force circulating blood must overcome in order to allow perfusion of body tissues. Narrow vessels create more vascular resistance, while dilated vessels decrease vascular resistance. Vasodilation acts to increase cardiac output by decreasing afterload, −one of the four determinants of cardiac output.
By expanding available area for blood to circulate, vasodilation decreases blood pressure. The response may be intrinsic (due to local processes in the surrounding tissue) or extrinsic (due to hormones or the nervous system). In addition, the response may be localized to a specific organ (depending on the metabolic needs of a particular tissue, as during strenuous exercise), or it may be systemic (seen throughout the entire systemic circulation).
Endogenous substances and drugs that cause vasodilation are termed vasodilators. Many of these substances are neurotransmitters released by perivascular nerves of the autonomic nervous system Baroreceptors sense blood pressure and allow adaptation via the mechanisms of vasoconstriction or vasodilation to maintain homeostasis.
Function[edit]
The primary function of vasodilation is to increase blood flow in the body to tissues that need it most. This is often in response to a localized need for oxygen but can occur when the tissue in question is not receiving enough glucose, lipids, or other nutrients. Vasodilation, both localized and systemic, also facilitates immune response. Localized tissues have multiple ways to increase blood flow, including releasing vasodilators, primarily adenosine, into the local interstitial fluid, which diffuses to capillary beds, provoking local vasodilation. Some physiologists have suggested that it is the lack of oxygen itself that causes capillary beds to vasodilate by the smooth muscle hypoxia of the vessels in the region. This latter hypothesis is posited due to the presence of precapillary sphincters in capillary beds. These approaches to the mechanism of vasodilation have not been found to be mutually exclusive.
Immune system[edit]
Vasodilation plays a major role in immune system function. Wider blood vessels allow more blood containing immune cells and proteins to reach the infection site. Vasodilation occurs as part of the process of inflammation, which is caused by several factors including presence of a pathogen, injury to tissues or blood vessels, and immune complexes. In severe cases, inflammation can lead to sepsis or distributive shock. Vasodilation is also a major component of anaphylaxis.
Inflammation causes not only vasodilation but also causes increased vascular permeability, allowing neutrophils, complement proteins, and antibodies to reach the site of infection or damage. Elevated vascular permeability can allow excess fluid to leave blood vessels and collect in tissues resulting in edema; vasodilation prevents blood vessels from constricting to adapt to reduced volume in the vessels, causing low blood pressure and septic shock.
In the case of inflammation, vasodilation is caused by cytokines. Interferon gamma, TNF-a, interleukin 1 beta, and interleukin 12 are a few examples of some inflammatory cytokines produced by immune cells such as natural killer cells, B cells, T cells, mast cells and macrophages. Anti-inflammatory cytokines that regulate inflammation and help prevent negative results such as septic shock are also produced by these immune cells. Vasodilation and increased vascular permeability also allow immune effector cells to leave blood vessels and follow chemoattractants to the infection site via a process called leukocyte extravasation. Vasodilation allows the same volume of blood to move more slowly according to the flow rate equation Q = Av, where Q represents flow rate, A represents cross-sectional area, and v represents velocity. Immune effector cells can more easily attach to selectins expressed on endothelial cells when blood is flowing slowly, enabling these cells to exit the blood vessel via diapedesis.
Anaphylaxis is a severe allergic reaction characterized by elevated vascular permeability, systemic vasodilation, gastrointestinal dysfunction, and respiratory dysfunction. Anaphylatoxins, specifically complement proteins C3a and C5a, bind to receptors on mast cells and basophils causing degranulation. Granules in these cells contain histamine, platelet-activating factor, and other compounds causing clinical manifestation of anaphylaxis- including systemic vasodilation causing dangerously low blood pressure. Immunoglobulin E, an antibody produced by plasma cells, also binds to receptors on mast cells and basophils causing degranulation.
Mechanism[edit]
A basic understanding of cardiac output, vascular resistance , and blood pressure is necessary to understand the causes and impacts of vasodilation. Cardiac output is defined as the amount of blood pumped through the heart over 1 minute, in units of liters per minute, equal to heart rate multiplied by stroke volume. It is directly related to heart rate, myocardial contractility, and preload, and inversely related with afterload. Elevated vascular resistance due to constricted blood vessels causes in increase in afterload, the amount of force against which the heart must contract. Vasodilation therefore decreases vascular resistance, which decreases afterload, elevating cardiac output and allowing perfusion of tissues. Blood pressure measures how much pressure blood exerts on blood vessel walls; systolic blood pressure measures pressure while the heart contracts (systole), and diastolic blood pressure reflects pressure between contractions (diastole). Mean arterial pressure (MAP)is a weighted average of systolic and diastolic blood pressures, and is a better measurement of perfusion over the duration of the cardiac cycle. Vasodilation works to decrease vascular resistance and blood pressure through relaxation of smooth muscle cells in the tunica media layer of large arteries and smaller arterioles. When vasodilation causes systolic blood pressure to fall below 90 mmHg, circulatory shock is observed.
Vascular resistance depends on several factors, including the length of the vessel, the viscosity of blood (determined by hematocrit) and the diameter of the blood vessel. The latter is the most important variable in determining resistance, with the vascular resistance changing by the fourth power of the radius. An increase in either of these physiological components (cardiac output or vascular resistance) causes a rise in MAP. Arterioles create the most vascular resistance of any blood vessel type, as they are very narrow and possess concentric layers of smooth muscle unlike venules and capillaries.
Vasodilation occurs in superficial blood vessels of warm-blooded animals when their ambient environment is hot; this process diverts the flow of heated blood to the skin of the animal, where heat can be more easily released to the environment. The opposite physiological process is vasoconstriction. These processes are naturally modulated by local paracrine agents from endothelial cells (e.g., nitric oxide, bradykinin, potassium ions, and adenosine), and by the autonomic nervous system and the adrenal glands, both of which secrete catecholamines, such as norepinephrine and epinephrine, respectively.
Smooth muscle physiology[edit]
The tunica media of the walls of arteries, arterioles, and veins is composed of smooth muscle and causes vasodilation and vasoconstriction. Contraction of smooth muscle cells causes vasoconstriction, and relaxation of smooth muscle causes vasodilation. Smooth muscle is innervated by the autonomic nervous system and is non-striated (does not contain sarcomeres). Contraction is dependent on concentrations of Ca in the cytosol, either via Ca,Mg-ATPase from the sarcoplasmic reticulum or voltage-gated calcium channels from the extracellular matrix. Calcium ions bind with calmodulin, activating myosin light-chain kinase which phosphorylates the myosin light-chain. Phosphorylated light-chain myosin interacts with actin filaments forming a cross-bridge, allowing muscle contraction causing vasoconstriction. Vasodilation is caused by myosin-light-chain phosphatase, which dephosphorylates the myosin light chain causing muscle relaxation. Smooth muscle cells can remain contracted without use of ATP due to action of the myosin-binding subunit of myosin light-chain phosphatase. Phosphorylation of this subunit by Rho-kinase prevents it from binding to and dephosphorylating the myosin light-chain, allowing the cell to remain contracted.
Vasodilation is the result of relaxation in smooth muscle surrounding the blood vessels. This relaxation, in turn, relies on removing the stimulus for contraction, which depends on intracellular calcium ion concentrations and is tightly linked with phosphorylation of the light chain of the contractile protein myosin. Thus, vasodilation works mainly either by lowering intracellular calcium concentration or by dephosphorylation (really substitution of ATP for ADP) of myosin. Dephosphorylation by myosin light-chain phosphatase and induction of calcium symporters and antiporters that pump calcium ions out of the intracellular compartment both contribute to smooth muscle cell relaxation and therefore vasodilation. This is accomplished through reuptake of ions into the sarcoplasmic reticulum via exchangers and expulsion across the plasma membrane. There are three main intracellular stimuli that can result in the vasodilation of blood vessels. The specific mechanisms to accomplish these effects vary from vasodilator to vasodilator.
Class
Description
Example
Hyperpolarization-mediated (Calcium channel blocker)
Changes in the resting membrane potential of the cell affects the level of intracellular calcium through modulation of voltage-sensitive calcium channels in the plasma membrane.
adenosine
cAMP-mediated
Adrenergic stimulation results in elevated levels of cAMP and protein kinase A, which results in increasing calcium removal from the cytoplasm.
prostacyclin
cGMP-mediated (Nitrovasodilator)
Through stimulation of protein kinase G.
nitric oxide
PDE5 inhibitors and potassium channel openers can also have similar results.
Compounds that mediate the above mechanisms may be grouped as endogenous and exogenous.
Causes[edit]
Endogenous[edit]
Vasodilators
Receptor (↑ = opens. ↓ = closes) On vascular smooth muscle cells if not otherwise specified
Transduction (↑ = increases. ↓ = decreases)
EDHF
?
hyperpolarization → ↓VDCC → ↓intracellular Ca
PKG activity →
phosphorylation of MLCK → ↓MLCK activity → dephosphorylation of MLC
↑SERCA → ↓intracellular Ca
NO receptor on endothelium
↓endothelin synthesis
epinephrine (adrenaline)
β-2 adrenergic receptor
↑Gs activity → ↑AC activity → ↑cAMP → ↑PKA activity → phosphorylation of MLCK → ↓MLCK activity → dephosphorylation of MLC
histamine
histamine H2 receptor
prostacyclin
IP receptor
prostaglandin D2
DP receptor
prostaglandin E2
EP receptor
VIP
VIP receptor
↑Gs activity → ↑AC activity → ↑cAMP → ↑PKA activity →
phosphorylation of MLCK → ↓MLCK activity → dephosphorylation of MLC
open Ca-activated and voltage-gated Kchannels → hyperpolarization → close VDCC → ↓intracellular Ca
(extracellular) adenosine
A1, A2a and A2b adenosine receptors
↑ATP-sensitive K channel → hyperpolarization → close VDCC → ↓intracellular Ca
(extracellular) ATP
(extracellular) ADP
↑P2Y receptor
activate Gq → ↑PLC activity → ↑intracellular Ca → ↑NOS activity → ↑NO → (see nitric oxide)
L-arginine
imidazoline and α-2 receptor?
Gi → ↓cAMP → activation of Na/K-ATPase → ↓intracellular Na → ↑Na/Ca exchanger activity → ↓intracellular Ca
bradykinin
bradykinin receptor
substance P
niacin (as nicotinic acid only)
platelet-activating factor (PAF)
CO2
-
↓interstitial pH → ?
interstitial lactic acid (probably)
-
muscle work
-
↑vasodilators:
↑ATP consumption → ↑adenosine
↑glucose usage → CO2
↑interstitial K
↑(extracellular) ATP
↑(extracellular) ADP
↑interstitial K
↓vasoconstrictors:
↑ATP consumption → ↓ ATP (intracellular)
↓oxygen → ↓oxidative phosphorylation → ↓ ATP (intracellular)
natriuretic peptides
prostaglandin I2
prostaglandin E2
heparin
various receptors on endothelium
↓endothelin synthesis
The vasodilating action of activation of beta-2 receptors (such as by adrenaline) appears to be endothelium-independent.
Autonomic nervous system control[edit]
As referenced in the explanation of smooth muscle physiology, smooth muscle within the tunica media is innervated by the autonomic nervous system. The autonomic nervous system (ANS) controls essential involuntary body functions and originates as nerves leaving the brain stem or spinal cord; it contains both sensor and motor nerves. The two divisions of the ANS, the sympathetic nervous system (SNS) and the parasympathetic nervous system (PSNS), impact blood vessels differently. Traditionally we understand that these two divisions work against each other, the SNS producing "fight or flight" and the PSNS producing "rest and digest", but in the case of vascular innervation this line becomes blurred ANS nerves do not directly innervate the vasculature via synapses with muscle cells; instead, they release neurotransmitters that reach target cells and effect smooth muscle contraction or relaxation. Physical characteristics of the SNS and PSNS cause the SNS to have a prolonged, systemic impact on blood vessels, while the PSNS causes short-lived, localized change. SNS stimulation causes a base level of vasoconstriction often referred to as basal neural tone, maintaining blood pressure. Often vasodilation is simply the result of insufficient neurotransmitter to maintain basal neural tone, without the presence of a compound directly causing vasodilation.
Neurotransmitters can act by binding directly to smooth muscle cells or by binding to endothelial cells mediating the effects of the neurotransmitter. Below is a table summarizing major neurotransmitters involved in regulation of the vasculature.
Neurotransmitter
Sympathetic or Parasympathetic
Target Cells and Receptors
Impact on Vasculature
norepinephrine (NE)
sympathetic (mostly)
adrenergic receptors α1, α2, β1, β2
α1- smooth muscle α2- endothelial β1, β2- smooth muscle
α1- increase concentration calcium ions, vasoconstricton
α2- inhibit cAMP, release NO, vasodilation β1, β2- possible vasodilation
Acetylcholine (Ach)
parasympathetic
nicotonic Ach receptors (nAchRs)
muscanaric Ach receptors (mAchRs) - on both endothelial and smooth muscle cells
nAchRs- modulate cytokines, counteract inflammation
mAchRs- endothelial M3 AchR release NO, vasodlation smooth muscle M2 and M3 AchRs reduce release NO, vasoconstriction Note: Ach is quickly broken down, diffused, or undergoes reuptake, impacts are brief and localized
Adenosine triphosphate (ATP)
sympathetic
purinergic receptors on smooth muscle and endothelial cells
smooth muscle- increase calcium ion concentration, vasoconstriction
endothelium- possible role as mediator of hyperpolarization of smooth muscle cells co-released with norepinephrine
Neuropeptide Y (NPY)
sympathetic
receptors on endothelial cells
causes vasoconstriction when co-released with norepinephrine
CGRP
?
CGRP1, CGRP2 receptors in endothelium
vasodilation, role in vascular dysfunction if levels are abnormal
Also worthy of mention when discussing neural control of vasodilation is the renin-angiotensin-aldosterone system, or RAAS. The kidneys retain water by reabsorbing sodium ions, or eliminate water by eliminating sodium ions. Sympathetic nervous system activity, reduced blood volume or reduced arterial pressure trigger β-adrenergic receptors in select kidney cells to release renin, which converts facilitates formation of angiotensin II from its substrate angiotensin. Angiotensin II triggers adrenal glands to secrete aldosterone, a potent vasoconstrictor.
Epinephrine, either exogenous or endogenous, is another vasoconstrictor released by the adrenal glands in response to stress. It binds to α and β adrenergic receptors like norepinephrine, causing vasodilation and vasoconstriction in different body parts to redistribute circulation to critical areas.
Cold-induced[edit]
Cold-induced vasodilation (CIVD) occurs after cold exposure, possibly to reduce the risk of injury. It can take place in several locations in the human body but is observed most often in the extremities. The fingers are especially common because they are exposed most often.
When the fingers are exposed to cold, vasoconstriction occurs first to reduce heat loss, resulting in strong cooling of the fingers. Approximately five to ten minutes after the start of the cold exposure of the hand, the blood vessels in the finger tips will suddenly vasodilate. This is probably caused by a sudden decrease in the release of neurotransmitters from the sympathetic nerves to the muscular coat of the arteriovenous anastomoses due to local cold. The CIVD increases blood flow and subsequently the temperature of the fingers. This can be painful and is sometimes known as the 'hot aches' which can be painful enough to bring on vomiting.
A new phase of vasoconstriction follows the vasodilation, after which the process repeats itself. This is called the Hunting reaction. Experiments have shown that three other vascular responses to immersion of the finger in cold water are possible: a continuous state of vasoconstriction; slow, steady, and continuous rewarming; and a proportional control form in which the blood vessel diameter remains constant after an initial phase of vasoconstriction. However, the vast majority of responses can be classified as the Hunting reaction.
Miscellaneous[edit]
This section needs more reliable medical references for verification or relies too heavily on primary sources. Please review the contents of the section and add the appropriate references if you can. Unsourced or poorly sourced material may be challenged and removed. Find sources: "Vasodilation" – news · newspapers · books · scholar · JSTOR (March 2022)
Other suggested vasodilators or vasodilating factors include:
absence of high levels of environmental noise
adenosine - adenosine agonist, used primarily as an anti-arrhythmic
alpha blockers (block the vasoconstricting effect of adrenaline)
atrial natriuretic peptide (ANP) - a weak vasodilator
ethanol (alcohol) causes immediate vasodilation followed by increase in blood pressure
nitric oxide inducers
l-arginine (a key amino acid)
citrulline (causes increased levels of L-arginine in the body)
glyceryl trinitrate (commonly known as nitroglycerin)
isosorbide mononitrate and isosorbide dinitrate
pentaerythritol tetranitrate (PETN)
sodium nitroprusside
PDE5 inhibitors: these agents indirectly increase the effects of nitric oxide
sildenafil (Viagra)
tadalafil (Cialis)
vardenafil (Levitra)
tetrahydrocannabinol (THC), the principal psychoactive constituent of cannabis
theobromine, the principal alkaloid found in Theobroma cacao, specifically in cocoa solids (which is found in chocolate, especially dark chocolate)
minoxidil
papaverine an alkaloid found in the opium poppy papaver somniferum
estrogen
Treatment[edit]
Direct vasodilation drugs[edit]
These drugs can keep vessels staying opened or help vessels refrain from being narrowed.
Angiotensin II receptor blockers
ACE inhibitors
Calcium channel blockers
Alpha-2A adrenergic receptor agonists[edit]
Drugs that appear to work by activating the α2A receptors in the brain thereby decreasing sympathetic nervous system activity.
methyldopa
According to American Heart Association, Alpha-methyldopa may cause Orthostatic syncope as it exerts a greater blood pressure lowering effect when one is standing upright which may lead to feeling weak or fainting if the blood pressure has been lowered too far. Methyldopa's prominent side effects include drowsiness or sluggishness, dryness of the mouth, fever or anemia. Additionally to these, male patients may experience impotence.
clonidine hydrochloride
guanabenz acetate
guanfacine hydrochloride
Clonidine, guanabenz or guanfacine may give rise to severe dryness of the mouth, constipation or drowsiness. Abrupt cessation taking may raise blood pressure quickly to dangerously high levels.
Blood vessel muscle relaxants[edit]
Directly relax the muscle in the walls of the blood vessels (especially the arterioles), allowing the vessel to dilate (widen).
hydralazine
minoxidil
Hydralazine may cause headaches, swelling around the eyes, heart palpitations or aches and pains in the joints. In clinical setting, hydralazine is not usually used alone.
Minoxidil is a potent direct vasodilator used only in resistant severe high blood pressure or when kidney failure is present. Noted adverse effects comprise fluid retention (marked weight gain) and excessive hair growth.
Therapeutic applications[edit]
Vasodilators are used to treat conditions such as hypertension, wherein the patient has an abnormally high blood pressure, as well as angina, congestive heart failure, and erectile dysfunction, and where maintaining a lower blood pressure reduces the patient's risk of developing other cardiac problems.
Flushing may be a physiological response to vasodilators. Some phosphodiesterase inhibitors such as sildenafil, vardenafil and tadalafil, work to increase blood flow in the penis through vasodilation. They may also be used to treat pulmonary arterial hypertension (PAH).
See also[edit]
Arteriolar vasodilator
Nitrophorin
Vasodilatory shock | biology | 789934 | https://no.wikipedia.org/wiki/Vasokonstriksjon | Vasokonstriksjon | Vasokonstriksjon er innsnevring av blodårene som følge av sammentrekning av muskulære veggen av årene, spesielt de store arterier, små arterioler og vener. Prosessen er det motsatte av vasodilatasjon, utvidelsen av blodårene. Prosessen er spesielt viktig for å stoppe blødning og akutt blodtap. Når blodkar trekker seg sammen, blir blodstrømmen redusert, og dermed holdes kroppsvarme tilbake og vaskulær motstand øker. Dette gjør huden blekere fordi mindre blod når overflaten, og dette hindrer utstrålingen av varme. Generelt er vasokonstriksjon en mekanisme der kroppen regulerer og opprettholder gjennomsnittlig arterieblodtrykk.
Medikamenter som forårsaker vasokonstriksjon, kalles vasokonstriktorer eller vasopressorer . Generalisert vasokonstriksjon resulterer vanligvis i en økning av systemisk blodtrykk, men det kan også oppstå i bestemte vev og forårsaker da en lokalisert reduksjon i blodstrømmen. Omfanget av vasokonstriksjon kan være lite eller alvorlig, avhengig av medikament eller omstendighet. Mange vasokonstriktorer kan også forårsake pupilldilatasjon. Medikamenter som forårsaker vasokonstriksjon inneholder antihistaminer, decongestants og sentralstimulerende midler som brukes til behandling av ADHD.
Generell mekanisme
Mekanismen som fører til vasokonstriksjon er et resultat av den økte konsentrasjonen av kalsium (Ca 2 + ioner) i blodårenes glatte muskelceller. Spesifikke mekanismer for å generere en økt intracellulær konsentrasjon av kalsium avhenger av vasokonstriktor. Glatte muskelceller er i stand til å generere aksjonspotensialer, men denne mekanismen blir sjeldent brukt til kontraksjon i blodkar. Hormonelle eller farmakokinetiske komponenter er fysiologisk mer relevante. To vanlige stimuli for å fremkalle sammentrekninger av glatt muskulatur er sirkulerende adrenalin og aktivering av det sympatiske nervesystemet som direkte innerverer muskelen (gjennom utskillelse av noradrenalin). Disse forbindelsene samhandler med celleoverflatens adrenerge reseptorer. Slike stimuli resulterer i en signaloverføringskaskade som fører til økt intracellulært kalsium fra sarkoplasmatisk retikulum gjennom IP3- mediert kalsium-utskillelse, samt økt kalsium-innstrømming over sarcolemma gjennom kalsium-kanaler. Økningen i intracellulære kalsium-komplekser med calmodulin, som igjen aktiverer myosin lett-kjede kinase. Dette enzymet er ansvarlig for fosforylering av den lette kjeden i myosin slik at den stimulerer til sirkulasjon over broen.
Når den har blitt hevet, synker den intracellulære kalsiumkonsentrasjonen tilbake til sitt basale nivå igjen gjennom en rekke proteinpumper og kalsium-vekslere lokalisert på cellemembranen og sarkoplasmatisk retikulum. Denne reduksjonen i kalsium fjerner stimulans nødvendig for kontraksjon, slik at cellen kommer tilbake til utgangspunktet.
Faktorer og individuelle mekanismer
Faktorer som utløser vasokonstriksjon kan være av eksogen (ytre) eller endogen (indre) opprinnelse. Omgivelsestemperatur er et eksempel på førstnevnte. Kutan vasokonstriksjon vil oppstå når kroppen eksponeres for streng kulde. Eksempler på endogene faktorer er det autonome nervesystemet, sirkulerende hormoner og iboende mekanismer knyttet til blodkarene selv (også referert til som myogen respons).
Eksogene medisiner
Eksempler kan være amfetaminer, antihistaminer og kokain. Mange er brukt i medisinen til å behandle hypotensjon og som lokal decongestant. Vasokonstriktorer brukes også klinisk til å øke blodtrykket eller for å redusere den lokale blodgjennomstrømningen.
Administrasjonsmåte varierer. Medikamentene kan gis både systemisk og utvortes. For eksempel er pseudoefedrin tilgjengelig systemisk (dvs. oralt inntatte tabletter som Sudafed) og lokalt (for eksempel nesespray som fenylefrin Neo-synephrine og øyedråper til pupilldilatasjon)
Eksempler er:
Endogene medisiner
Vasokonstriksjon er en kroppsmekanisme som forhindrer ortostatisk hypotensjon. Det er en del av en negativ feed back loop hvor kroppen forsøker å gjenopprette homeostase.
Vasokonstriksjon forekommer også i overfladiske blodkar hos varmblodige dyr når deres omgivende miljø er kaldt. Denne prosessen sender oppvarmet blod til sentrum av dyret, og hindrer derved tap av varme.
Patologi
Vasokonstriksjon kan være en medvirkende årsak til erektil dysfunksjon. En økning i blodstrømmen til penis forårsaker en ereksjon.
Uønsket vasokonstriksjon kan også spille en rolle i sekundær hypertensjon.
Referanser
Se også
Addisons sykdom
Inotrope
Hypertensjon
Nitrogenoksid
Feokromocytom
Sjokk
Vasodilatasjon
Postural ortostatisk takykardi-syndrom
Eksterne lenker
Definition of Vasoconstriction on HealthScout
Cannabis arteritis revisited--ten new case reports
Are coronary heart disease and peripheral arterial disease associated with tobacco or cannabis consumption
Vasoconstrictor effects of Cannabis appear to inhibit Migraine (headache) attacks
Kardiologi
Fysiologi | norwegian_bokmål | 0.419178 |
brain_no_pain_receptors_headache/Headache.txt | Headache, also known as cephalalgia, is the symptom of pain in the face, head, or neck. It can occur as a migraine, tension-type headache, or cluster headache. There is an increased risk of depression in those with severe headaches.
Headaches can occur as a result of many conditions. There are a number of different classification systems for headaches. The most well-recognized is that of the International Headache Society, which classifies it into more than 150 types of primary and secondary headaches. Causes of headaches may include dehydration; fatigue; sleep deprivation; stress; the effects of medications (overuse) and recreational drugs, including withdrawal; viral infections; loud noises; head injury; rapid ingestion of a very cold food or beverage; and dental or sinus issues (such as sinusitis).
Treatment of a headache depends on the underlying cause, but commonly involves pain medication (especially in case of migraine or cluster headaches). A headache is one of the most commonly experienced of all physical discomforts.
About half of adults have a headache in a given year. Tension headaches are the most common, affecting about 1.6 billion people (21.8% of the population) followed by migraine headaches which affect about 848 million (11.7%).
There are more than 200 types of headaches. Some are harmless and some are life-threatening. The description of the headache and findings on neurological examination, determine whether additional tests are needed and what treatment is best.
Headaches are broadly classified as "primary" or "secondary". Primary headaches are benign, recurrent headaches not caused by underlying disease or structural problems. For example, migraine is a type of primary headache. While primary headaches may cause significant daily pain and disability, they are not dangerous from a physiological point of view. Secondary headaches are caused by an underlying disease, like an infection, head injury, vascular disorders, brain bleed, stomach irritation, or tumors. Secondary headaches can be dangerous. Certain "red flags" or warning signs indicate a secondary headache may be dangerous.
Ninety percent of all headaches are primary headaches. Primary headaches usually first start when people are between 20 and 40 years old. The most common types of primary headaches are migraines and tension-type headaches. They have different characteristics. Migraines typically present with pulsing head pain, nausea, photophobia (sensitivity to light) and phonophobia (sensitivity to sound). Tension-type headaches usually present with non-pulsing "bandlike" pressure on both sides of the head, not accompanied by other symptoms. Such kind of headaches may be further classified into-episodic and chronic tension type headaches Other very rare types of primary headaches include:
Headaches may be caused by problems elsewhere in the head or neck. Some of these are not harmful, such as cervicogenic headache (pain arising from the neck muscles). The excessive use of painkillers can paradoxically cause worsening painkiller headaches.
More serious causes of secondary headaches include the following:
Gastrointestinal disorders may cause headaches, including Helicobacter pylori infection, celiac disease, non-celiac gluten sensitivity, irritable bowel syndrome, inflammatory bowel disease, gastroparesis, and hepatobiliary disorders. The treatment of the gastrointestinal disorders may lead to a remission or improvement of headaches.
Migraine headaches are also associated with Cyclic Vomiting Syndrome (CVS). CVS is characterized by episodes of severe vomiting, and often occur alongside symptoms similar to those of migraine headaches (photophobia, abdominal pain, etc.).
The brain itself is not sensitive to pain, because it lacks pain receptors. However, several areas of the head and neck do have pain receptors and can thus sense pain. These include the extracranial arteries, middle meningeal artery, large veins, venous sinuses, cranial and spinal nerves, head and neck muscles, the meninges, falx cerebri, parts of the brainstem, eyes, ears, teeth, and lining of the mouth. Pial arteries, rather than pial veins are responsible for pain production.
Headaches often result from traction or irritation of the meninges and blood vessels. The pain receptors may be stimulated by head trauma or tumours and cause headaches. Blood vessel spasms, dilated blood vessels, inflammation or infection of meninges and muscular tension can also stimulate pain receptors. Once stimulated, a nociceptor sends a message up the length of the nerve fibre to the nerve cells in the brain, signalling that a part of the body hurts.
Primary headaches are more difficult to understand than secondary headaches. The exact mechanisms which cause migraines, tension headaches and cluster headaches are not known. There have been different hypotheses over time that attempt to explain what happens in the brain to cause these headaches.
Migraines are currently thought to be caused by dysfunction of the nerves in the brain. Previously, migraines were thought to be caused by a primary problem with the blood vessels in the brain. This vascular theory, which was developed in the 20th century by Wolff, suggested that the aura in migraines is caused by constriction of intracranial vessels (vessels inside the brain), and the headache itself is caused by rebound dilation of extracranial vessels (vessels just outside the brain). Dilation of these extracranial blood vessels activates the pain receptors in the surrounding nerves, causing a headache. The vascular theory is no longer accepted. Studies have shown migraine head pain is not accompanied by extracranial vasodilation, but rather only has some mild intracranial vasodilation.
Currently, most specialists think migraines are due to a primary problem with the nerves in the brain. Auras are thought to be caused by a wave of increased activity of neurons in the cerebral cortex (a part of the brain) known as cortical spreading depression followed by a period of depressed activity. Some people think headaches are caused by the activation of sensory nerves which release peptides or serotonin, causing inflammation in arteries, dura and meninges and also cause some vasodilation. Triptans, medications that treat migraines, block serotonin receptors and constrict blood vessels.
People who are more susceptible to experiencing migraines without headaches are those who have a family history of migraines, women, and women who are experiencing hormonal changes or are taking birth control pills or are prescribed hormone replacement therapy.
Tension headaches are thought to be caused by the activation of peripheral nerves in the head and neck muscles.
Cluster headaches involve overactivation of the trigeminal nerve and hypothalamus in the brain, but the exact cause is unknown.
Most headaches can be diagnosed by the clinical history alone. If the symptoms described by the person sound dangerous, further testing with neuroimaging or lumbar puncture may be necessary. Electroencephalography (EEG) is not useful for headache diagnosis.
The first step to diagnosing a headache is to determine if the headache is old or new. A "new headache" can be a headache that has started recently, or a chronic headache that has changed character. For example, if a person has chronic weekly headaches with pressure on both sides of his head, and then develops a sudden severe throbbing headache on one side of his head, they have a new headache.
It can be challenging to differentiate between low-risk, benign headaches and high-risk, dangerous headaches since symptoms are often similar. Headaches that are possibly dangerous require further lab tests and imaging to diagnose.
The American College for Emergency Physicians published criteria for low-risk headaches. They are as follows:
A number of characteristics make it more likely that the headache is due to potentially dangerous secondary causes which may be life-threatening or cause long-term damage. These "red flag" symptoms mean that a headache warrants further investigation with neuroimaging and lab tests.
In general, people complaining of their "first" or "worst" headache warrant imaging and further workup. People with progressively worsening headache also warrant imaging, as they may have a mass or a bleed that is gradually growing, pressing on surrounding structures and causing worsening pain. People with neurological findings on exam, such as weakness, also need further workup.
The American Headache Society recommends using "SSNOOP", a mnemonic to remember the red flags for identifying a secondary headache:
Other red flag symptoms include:
Old headaches are usually primary headaches and are not dangerous. They are most often caused by migraines or tension headaches. Migraines are often unilateral, pulsing headaches accompanied by nausea or vomiting. There may be an aura (visual symptoms, numbness or tingling) 30–60 minutes before the headache, warning the person of a headache. Migraines may also not have auras. Tension-type headaches usually have bilateral "bandlike" pressure on both sides of the head usually without nausea or vomiting. However, some symptoms from both headache groups may overlap. It is important to distinguish between the two because the treatments are different.
The mnemonic 'POUND' helps distinguish between migraines and tension-type headaches. POUND stands for:
One review article found that if 4–5 of the POUND characteristics are present, a migraine is 24 times as likely a diagnosis than a tension-type headache (likelihood ratio 24). If 3 characteristics of POUND are present, migraine is 3 times more likely a diagnosis than tension type headache (likelihood ratio 3). If only 2 POUND characteristics are present, tension-type headaches are 60% more likely (likelihood ratio 0.41). Another study found the following factors independently each increase the chance of migraine over tension-type headache: nausea, photophobia, phonophobia, exacerbation by physical activity, unilateral, throbbing quality, chocolate as a headache trigger, and cheese as a headache trigger.
Cluster headaches are relatively rare (1 in 1000 people) and are more common in men than women. They present with sudden onset explosive pain around one eye and are accompanied by autonomic symptoms (tearing, runny nose and red eye).
Temporomandibular jaw pain (chronic pain in the jaw joint), and cervicogenic headache (headache caused by pain in muscles of the neck) are also possible diagnoses.
For chronic, unexplained headaches, keeping a headache diary can be useful for tracking symptoms and identifying triggers, such as association with menstrual cycle, exercise and food. While mobile electronic diaries for smartphones are becoming increasingly common, a recent review found most are developed with a lack of evidence base and scientific expertise.
Cephalalgiaphobia is fear of headaches or getting a headache.
New headaches are more likely to be dangerous secondary headaches. They can, however, simply be the first presentation of a chronic headache syndrome, like migraine or tension-type headaches.
One recommended diagnostic approach is as follows. If any urgent red flags are present such as visual loss, new seizures, new weakness, new confusion, further workup with imaging and possibly a lumbar puncture should be done (see red flags section for more details). If the headache is sudden onset (thunderclap headache), a computed tomography test to look for a brain bleed (subarachnoid hemorrhage) should be done. If the CT scan does not show a bleed, a lumbar puncture should be done to look for blood in the CSF, as the CT scan can be falsely negative and subarachnoid hemorrhages can be fatal. If there are signs of infection such as fever, rash, or stiff neck, a lumbar puncture to look for meningitis should be considered. If there is jaw claudication and scalp tenderness in an older person, a temporal artery biopsy to look for temporal arteritis should be performed and immediate treatment should be started.
The US Headache Consortium has guidelines for neuroimaging of non-acute headaches. Most old, chronic headaches do not require neuroimaging. If a person has the characteristic symptoms of a migraine, neuroimaging is not needed as it is very unlikely the person has an intracranial abnormality. If the person has neurological findings, such as weakness, on exam, neuroimaging may be considered.
All people who present with red flags indicating a dangerous secondary headache should receive neuroimaging. The best form of neuroimaging for these headaches is controversial. Non-contrast computerized tomography (CT) scan is usually the first step in head imaging as it is readily available in Emergency Departments and hospitals and is cheaper than MRI. Non-contrast CT is best for identifying an acute head bleed. Magnetic Resonance Imaging (MRI) is best for brain tumors and problems in the posterior fossa, or back of the brain. MRI is more sensitive for identifying intracranial problems, however it can pick up brain abnormalities that are not relevant to the person's headaches.
The American College of Radiology recommends the following imaging tests for different specific situations:
A lumbar puncture is a procedure in which cerebral spinal fluid is removed from the spine with a needle. A lumbar puncture is necessary to look for infection or blood in the spinal fluid. A lumbar puncture can also evaluate the pressure in the spinal column, which can be useful for people with idiopathic intracranial hypertension (usually young, obese women who have increased intracranial pressure), or other causes of increased intracranial pressure. In most cases, a CT scan should be done first.
Headaches are most thoroughly classified by the International Headache Society's International Classification of Headache Disorders (ICHD), which published the second edition in 2004. The third edition of the International Headache Classification was published in 2013 in a beta version ahead of the final version. This classification is accepted by the WHO.
Other classification systems exist. One of the first published attempts was in 1951. The US National Institutes of Health developed a classification system in 1962.
The International Classification of Headache Disorders (ICHD) is an in-depth hierarchical classification of headaches published by the International Headache Society. It contains explicit (operational) diagnostic criteria for headache disorders. The first version of the classification, ICHD-1, was published in 1988. The current revision, ICHD-2, was published in 2004. The classification uses numeric codes. The top, one-digit diagnostic level includes 14 headache groups. The first four of these are classified as primary headaches, groups 5-12 as secondary headaches, cranial neuralgia, central and primary facial pain and other headaches for the last two groups.
The ICHD-2 classification defines migraines, tension-types headaches, cluster headache and other trigeminal autonomic headache as the main types of primary headaches. Also, according to the same classification, stabbing headaches and headaches due to cough, exertion and sexual activity (sexual headache) are classified as primary headaches. The daily-persistent headaches along with the hypnic headache and thunderclap headaches are considered primary headaches as well.
Secondary headaches are classified based on their cause and not on their symptoms. According to the ICHD-2 classification, the main types of secondary headaches include those that are due to head or neck trauma such as whiplash injury, intracranial hematoma, post craniotomy or other head or neck injury. Headaches caused by cranial or cervical vascular disorders such as ischemic stroke and transient ischemic attack, non-traumatic intracranial hemorrhage, vascular malformations or arteritis are also defined as secondary headaches. This type of headache may also be caused by cerebral venous thrombosis or different intracranial vascular disorders. Other secondary headaches are those due to intracranial disorders that are not vascular such as low or high pressure of the cerebrospinal fluid pressure, non-infectious inflammatory disease, intracranial neoplasm, epileptic seizure or other types of disorders or diseases that are intracranial but that are not associated with the vasculature of the central nervous system.
ICHD-2 classifies headaches that are caused by the ingestion of a certain substance or by its withdrawal as secondary headaches as well. This type of headache may result from the overuse of some medications or exposure to some substances. HIV/AIDS, intracranial infections and systemic infections may also cause secondary headaches. The ICHD-2 system of classification includes the headaches associated with homeostasis disorders in the category of secondary headaches. This means that headaches caused by dialysis, high blood pressure, hypothyroidism, cephalalgia and even fasting are considered secondary headaches. Secondary headaches, according to the same classification system, can also be due to the injury of any of the facial structures including teeth, jaws, or temporomandibular joint. Headaches caused by psychiatric disorders such as somatization or psychotic disorders are also classified as secondary headaches.
The ICHD-2 classification puts cranial neuralgias and other types of neuralgia in a different category. According to this system, there are 19 types of neuralgias and headaches due to different central causes of facial pain. Moreover, the ICHD-2 includes a category that contains all the headaches that cannot be classified.
Although the ICHD-2 is the most complete headache classification there is and it includes frequency in the diagnostic criteria of some types of headaches (primarily primary headaches), it does not specifically code frequency or severity which are left at the discretion of the examiner.
The NIH classification consists of brief definitions of a limited number of headaches.
The NIH system of classification is more succinct and only describes five categories of headaches. In this case, primary headaches are those that do not show organic or structural causes. According to this classification, primary headaches can only be vascular, myogenic, cervicogenic, traction, and inflammatory.
Primary headache syndromes have many different possible treatments. In those with chronic headaches the long term use of opioids appears to result in greater harm than benefit.
Migraine can be somewhat improved by lifestyle changes, but most people require medicines to control their symptoms. Medications are either to prevent getting migraines, or to reduce symptoms once a migraine starts.
Preventive medications are generally recommended when people have more than four attacks of migraine per month, headaches last longer than 12 hours or the headaches are very disabling. Possible therapies include beta blockers, antidepressants, anticonvulsants and NSAIDs. The type of preventive medicine is usually chosen based on the other symptoms the person has. For example, if the person also has depression, an antidepressant is a good choice.
Abortive therapies for migraines may be oral, if the migraine is mild to moderate, or may require stronger medicine given intravenously or intramuscularly. Mild to moderate headaches should first be treated with acetaminophen (paracetamol) or NSAIDs, like ibuprofen. If accompanied by nausea or vomiting, an antiemetic such as metoclopramide (Reglan) can be given orally or rectally. Moderate to severe attacks should be treated first with an oral triptan, a medication that mimics serotonin (an agonist) and causes mild vasoconstriction. If accompanied by nausea and vomiting, parenteral (through a needle in the skin) triptans and antiemetics can be given.
Sphenopalatine ganglion block (SPG block, also known nasal ganglion block or pterygopalatine ganglion blocks) can abort and prevent migraines, tension headaches and cluster headaches. It was originally described by American ENT surgeon Greenfield Sluder in 1908. Both blocks and neurostimulation have been studied as treatment for headaches.
Several complementary and alternative strategies can help with migraines. The American Academy of Neurology guidelines for migraine treatment in 2000 stated relaxation training, electromyographic feedback and cognitive behavioral therapy may be considered for migraine treatment, along with medications.
Tension-type headaches can usually be managed with NSAIDs (ibuprofen, naproxen, aspirin), or acetaminophen. Triptans are not helpful in tension-type headaches unless the person also has migraines. For chronic tension type headaches, amitriptyline is the only medication proven to help. Amitriptyline is a medication which treats depression and also independently treats pain. It works by blocking the reuptake of serotonin and norepinephrine, and also reduces muscle tenderness by a separate mechanism. Studies evaluating acupuncture for tension-type headaches have been mixed. Overall, they show that acupuncture is probably not helpful for tension-type headaches.
Abortive therapy for cluster headaches includes subcutaneous sumatriptan (injected under the skin) and triptan nasal sprays. High flow oxygen therapy also helps with relief.
For people with extended periods of cluster headaches, preventive therapy can be necessary. Verapamil is recommended as first line treatment. Lithium can also be useful. For people with shorter bouts, a short course of prednisone (10 days) can be helpful. Ergotamine is useful if given 1–2 hours before an attack.
Treatment of secondary headaches involves treating the underlying cause. For example, a person with meningitis will require antibiotics. A person with a brain tumor may require surgery, chemotherapy or brain radiation.
Peripheral neuromodulation has tentative benefits in primary headaches including cluster headaches and chronic migraine. How it may work is still being looked into.
Literature reviews find that approximately 64–77% of adults have had a headache at some point in their lives. During each year, on average, 46–53% of people have headaches. However, the prevalence of headache varies widely depending on how the survey was conducted, with studies finding lifetime prevalence of as low as 8% to as high as 96%. Most of these headaches are not dangerous. Only approximately 1–5% of people who seek emergency treatment for headaches have a serious underlying cause.
More than 90% of headaches are primary headaches. Most of these primary headaches are tension headaches. Most people with tension headaches have "episodic" tension headaches that come and go. Only 3.3% of adults have chronic tension headaches, with headaches for more than 15 days in a month.
Approximately 12–18% of people in the world have migraines. More women than men experience migraines. In Europe and North America, 5–9% of men experience migraines, while 12–25% of women experience migraines.
Cluster headaches are relatively uncommon. They affect only 1–3 per thousand people in the world. Cluster headaches affect approximately three times as many men as women.
The first recorded classification system was published by Aretaeus of Cappadocia, a medical scholar of Greco-Roman antiquity. He made a distinction between three different types of headache: i) cephalalgia, by which he indicates a sudden onset, temporary headache; ii) cephalea, referring to a chronic type of headache; and iii) heterocrania, a paroxysmal headache on one side of the head.
Another classification system that resembles the modern ones was published by Thomas Willis, in De Cephalalgia in 1672. In 1787 Christian Baur generally divided headaches into idiopathic (primary headaches) and symptomatic (secondary ones), and defined 84 categories.
In general, children experience the same types of headaches as adults do, but their symptoms may be slightly different. The diagnostic approach to headaches in children is similar to that of adults. However, young children may not be able to verbalize pain well. If a young child is fussy, they may have a headache.
Approximately 1% of emergency department visits for children are for headache. Most of these headaches are not dangerous. The most common type of headache seen in pediatric emergency rooms is headache caused by a cold (28.5%). Other headaches diagnosed in the emergency department include post-traumatic headache (20%), headache related to a problem with a ventriculoperitoneal shunt (a device put into the brain to remove excess CSF and reduce pressure in the brain) (11.5%) and migraine (8.5%). The most common serious headaches found in children include brain bleeds (subdural hematoma, epidural hematoma), brain abscesses, meningitis and ventriculoperitoneal shunt malfunction. Only 4–6.9% of kids with a headache have a serious cause.
Just as in adults, most headaches are benign, but when head pain is accompanied with other symptoms such as speech problems, muscle weakness, and loss of vision, a more serious underlying cause may exist: hydrocephalus, meningitis, encephalitis, abscess, hemorrhage, tumor, blood clots, or head trauma. In these cases, the headache evaluation may include CT scan or MRI in order to look for possible structural disorders of the central nervous system. If a child with a recurrent headache has a normal physical exam, neuroimaging is not recommended. Guidelines state children with abnormal neurologic exams, confusion, seizures and recent onset of worst headache of life, change in headache type or anything suggesting neurologic problems should receive neuroimaging.
When children complain of headaches, many parents are concerned about a brain tumor. Generally, headaches caused by brain masses are incapacitating and accompanied by vomiting. One study found characteristics associated with brain tumor in children are: headache for greater than 6 months, headache related to sleep, vomiting, confusion, no visual symptoms, no family history of migraine and abnormal neurologic exam.
Some measures can help prevent headaches in children. Drinking plenty of water throughout the day, avoiding caffeine, getting enough and regular sleep, eating balanced meals at the proper times, and reducing stress and excess of activities may prevent headaches. Treatments for children are similar to those for adults, however certain medications such as narcotics should not be given to children.
Children who have headaches will not necessarily have headaches as adults. In one study of 100 children with headache, eight years later 44% of those with tension headache and 28% of those with migraines were headache free. In another study of people with chronic daily headache, 75% did not have chronic daily headaches two years later, and 88% did not have chronic daily headaches eight years later.
Causes[edit]
There are more than 200 types of headaches. Some are harmless and some are life-threatening. The description of the headache and findings on neurological examination, determine whether additional tests are needed and what treatment is best.
Headaches are broadly classified as "primary" or "secondary". Primary headaches are benign, recurrent headaches not caused by underlying disease or structural problems. For example, migraine is a type of primary headache. While primary headaches may cause significant daily pain and disability, they are not dangerous from a physiological point of view. Secondary headaches are caused by an underlying disease, like an infection, head injury, vascular disorders, brain bleed, stomach irritation, or tumors. Secondary headaches can be dangerous. Certain "red flags" or warning signs indicate a secondary headache may be dangerous.
Primary[edit]
Ninety percent of all headaches are primary headaches. Primary headaches usually first start when people are between 20 and 40 years old. The most common types of primary headaches are migraines and tension-type headaches. They have different characteristics. Migraines typically present with pulsing head pain, nausea, photophobia (sensitivity to light) and phonophobia (sensitivity to sound). Tension-type headaches usually present with non-pulsing "bandlike" pressure on both sides of the head, not accompanied by other symptoms. Such kind of headaches may be further classified into-episodic and chronic tension type headaches Other very rare types of primary headaches include:
cluster headaches: short episodes (15–180 minutes) of severe pain, usually around one eye, with autonomic symptoms (tearing, red eye, nasal congestion) which occur at the same time every day. Cluster headaches can be treated with triptans and prevented with prednisone, ergotamine or lithium.
trigeminal neuralgia or occipital neuralgia: shooting face pain
hemicrania continua: continuous unilateral pain with episodes of severe pain. Hemicrania continua can be relieved by the medication indomethacin.
primary stabbing headache: recurrent episodes of stabbing "ice pick pain" or "jabs and jolts" for 1 second to several minutes without autonomic symptoms (tearing, red eye, nasal congestion). These headaches can be treated with indomethacin.
primary cough headache: starts suddenly and lasts for several minutes after coughing, sneezing or straining (anything that may increase pressure in the head). Serious causes (see secondary headaches red flag section) must be ruled out before a diagnosis of "benign" primary cough headache can be made.
primary exertional headache: throbbing, pulsatile pain which starts during or after exercising, lasting for 5 minutes to 24 hours. The mechanism behind these headaches is unclear, possibly due to straining causing veins in the head to dilate, causing pain. These headaches can be prevented by not exercising too strenuously and can be treated with medications such as indomethacin.
primary sex headache: dull, bilateral headache that starts during sexual activity and becomes much worse during orgasm. These headaches are thought to be due to lower pressure in the head during sex. It is important to realize that headaches that begin during orgasm may be due to a subarachnoid hemorrhage, so serious causes must be ruled out first. These headaches are treated by advising the person to stop sex if they develop a headache. Medications such as propranolol and diltiazem can also be helpful.
hypnic headache: a moderate-severe headache that starts a few hours after falling asleep and lasts 15–30 minutes. The headache may recur several times during the night. Hypnic headaches are usually in older women. They may be treated with lithium.
Secondary[edit]
This section needs additional citations for verification. Please help improve this article by adding citations to reliable sources in this section. Unsourced material may be challenged and removed. (February 2021) (Learn how and when to remove this template message)
Headaches may be caused by problems elsewhere in the head or neck. Some of these are not harmful, such as cervicogenic headache (pain arising from the neck muscles). The excessive use of painkillers can paradoxically cause worsening painkiller headaches.
More serious causes of secondary headaches include the following:
meningitis: inflammation of the meninges which presents with fever and meningismus, or stiff neck
bleeding inside the brain (intracranial hemorrhage)
subarachnoid hemorrhage (acute, severe headache, stiff neck without fever)
ruptured aneurysm, arteriovenous malformation, intraparenchymal hemorrhage (headache only)
brain tumor: dull headache, worse with exertion and change in position, accompanied by nausea and vomiting. Often, the person will have nausea and vomiting for weeks before the headache starts.
temporal arteritis: inflammatory disease of arteries common in the elderly (average age 70) with fever, headache, weight loss, jaw claudication, tender vessels by the temples, polymyalgia rheumatica
acute closed-angle glaucoma (increased pressure in the eyeball): a headache that starts with eye pain, blurry vision, associated with nausea and vomiting. On physical exam, the person will have red eyes and a fixed, mid-dilated pupil.
Post-ictal headaches: Headaches that happen after a convulsion or other type of seizure, as part of the period after the seizure (the post-ictal state)
Gastrointestinal disorders may cause headaches, including Helicobacter pylori infection, celiac disease, non-celiac gluten sensitivity, irritable bowel syndrome, inflammatory bowel disease, gastroparesis, and hepatobiliary disorders. The treatment of the gastrointestinal disorders may lead to a remission or improvement of headaches.
Migraine headaches are also associated with Cyclic Vomiting Syndrome (CVS). CVS is characterized by episodes of severe vomiting, and often occur alongside symptoms similar to those of migraine headaches (photophobia, abdominal pain, etc.).
Pathophysiology[edit]
The brain itself is not sensitive to pain, because it lacks pain receptors. However, several areas of the head and neck do have pain receptors and can thus sense pain. These include the extracranial arteries, middle meningeal artery, large veins, venous sinuses, cranial and spinal nerves, head and neck muscles, the meninges, falx cerebri, parts of the brainstem, eyes, ears, teeth, and lining of the mouth. Pial arteries, rather than pial veins are responsible for pain production.
Headaches often result from traction or irritation of the meninges and blood vessels. The pain receptors may be stimulated by head trauma or tumours and cause headaches. Blood vessel spasms, dilated blood vessels, inflammation or infection of meninges and muscular tension can also stimulate pain receptors. Once stimulated, a nociceptor sends a message up the length of the nerve fibre to the nerve cells in the brain, signalling that a part of the body hurts.
Primary headaches are more difficult to understand than secondary headaches. The exact mechanisms which cause migraines, tension headaches and cluster headaches are not known. There have been different hypotheses over time that attempt to explain what happens in the brain to cause these headaches.
Migraines are currently thought to be caused by dysfunction of the nerves in the brain. Previously, migraines were thought to be caused by a primary problem with the blood vessels in the brain. This vascular theory, which was developed in the 20th century by Wolff, suggested that the aura in migraines is caused by constriction of intracranial vessels (vessels inside the brain), and the headache itself is caused by rebound dilation of extracranial vessels (vessels just outside the brain). Dilation of these extracranial blood vessels activates the pain receptors in the surrounding nerves, causing a headache. The vascular theory is no longer accepted. Studies have shown migraine head pain is not accompanied by extracranial vasodilation, but rather only has some mild intracranial vasodilation.
Currently, most specialists think migraines are due to a primary problem with the nerves in the brain. Auras are thought to be caused by a wave of increased activity of neurons in the cerebral cortex (a part of the brain) known as cortical spreading depression followed by a period of depressed activity. Some people think headaches are caused by the activation of sensory nerves which release peptides or serotonin, causing inflammation in arteries, dura and meninges and also cause some vasodilation. Triptans, medications that treat migraines, block serotonin receptors and constrict blood vessels.
People who are more susceptible to experiencing migraines without headaches are those who have a family history of migraines, women, and women who are experiencing hormonal changes or are taking birth control pills or are prescribed hormone replacement therapy.
Tension headaches are thought to be caused by the activation of peripheral nerves in the head and neck muscles.
Cluster headaches involve overactivation of the trigeminal nerve and hypothalamus in the brain, but the exact cause is unknown.
Diagnosis[edit]
Differential diagnosis of headaches
Tension headache
New daily persistent headache
Cluster headache
Migraine
mild to moderate dull or aching pain
severe pain
moderate to severe pain
duration of 30 minutes to several hours
duration of at least four hours daily
duration of 30 minutes to 3 hours
duration of 4 hours to 3 days
Occur in periods of 15 days a month for three months
may happen multiple times in a day for months
periodic occurrence; several per month to several per year
located as tightness or pressure across head
located on one or both sides of the head
located one side of head focused at eye or temple
located on one or both sides of head
consistent pain
pain describable as sharp or stabbing
pulsating or throbbing pain
no nausea or vomiting
nausea, perhaps with vomiting
no aura
no aura
auras
uncommonly, light sensitivity or noise sensitivity
may be accompanied by running nose, tears, and drooping eyelid, often only on one side
sensitivity to movement, light, and noise
exacerbated by regular use of acetaminophen or NSAIDS
may exist with tension headache
Most headaches can be diagnosed by the clinical history alone. If the symptoms described by the person sound dangerous, further testing with neuroimaging or lumbar puncture may be necessary. Electroencephalography (EEG) is not useful for headache diagnosis.
The first step to diagnosing a headache is to determine if the headache is old or new. A "new headache" can be a headache that has started recently, or a chronic headache that has changed character. For example, if a person has chronic weekly headaches with pressure on both sides of his head, and then develops a sudden severe throbbing headache on one side of his head, they have a new headache.
Red flags[edit]
It can be challenging to differentiate between low-risk, benign headaches and high-risk, dangerous headaches since symptoms are often similar. Headaches that are possibly dangerous require further lab tests and imaging to diagnose.
The American College for Emergency Physicians published criteria for low-risk headaches. They are as follows:
age younger than 30 years
features typical of primary headache
history of similar headache
no abnormal findings on neurologic exam
no concerning change in normal headache pattern
no high-risk comorbid conditions (for example, HIV)
no new concerning history or physical examination findings
A number of characteristics make it more likely that the headache is due to potentially dangerous secondary causes which may be life-threatening or cause long-term damage. These "red flag" symptoms mean that a headache warrants further investigation with neuroimaging and lab tests.
In general, people complaining of their "first" or "worst" headache warrant imaging and further workup. People with progressively worsening headache also warrant imaging, as they may have a mass or a bleed that is gradually growing, pressing on surrounding structures and causing worsening pain. People with neurological findings on exam, such as weakness, also need further workup.
The American Headache Society recommends using "SSNOOP", a mnemonic to remember the red flags for identifying a secondary headache:
Systemic symptoms (fever or weight loss)
Systemic disease (HIV infection, malignancy)
Neurologic symptoms or signs
Onset sudden (thunderclap headache)
Onset after age 40 years
Previous headache history (first, worst, or different headache)
Other red flag symptoms include:
Red Flag
Possible causes
The reason why a red flag indicates possible causes
Diagnostic tests
New headache after age 50
Temporal arteritis, mass in brain
Temporal arteritis is an inflammation of vessels close to the temples in older people, which decreases blood flow to the brain and causes pain. May also have tenderness in temples or jaw claudication. Some brain cancers are more common in older people.
Erythrocyte sedimentation rate (diagnostic test for temporal arteritis), neuroimaging
Very sudden onset headache (thunderclap headache)
Brain bleed (subarachnoid hemorrhage, hemorrhage into mass lesion, vascular malformation), pituitary apoplexy, mass (especially in posterior fossa)
A bleed in the brain irritates the meninges which causes pain. Pituitary apoplexy (bleeding or impaired blood supply to the pituitary gland at the base of the brain) is often accompanied by double vision or visual field defects, since the pituitary gland is right next to the optic chiasm (eye nerves).
Neuroimaging, lumbar puncture if computed tomography is negative
Headaches increasing in frequency and severity
Mass, subdural hematoma, medication overuse
As a brain mass gets larger, or a subdural hematoma (blood outside the vessels underneath the dura) it pushes more on surrounding structures causing pain. Medication overuse headaches worsen with more medication taken over time.
Neuroimaging, drug screen
New onset headache in a person with possible HIV or cancer
Meningitis (chronic or carcinomatous), brain abscess including toxoplasmosis, metastasis
People with HIV or cancer are immunosuppressed so are likely to get infections of the meninges or infections in the brain causing abscesses. Cancer can metastasize, or travel through the blood or lymph to other sites in the body.
Neuroimaging, lumbar puncture if neuroimaging is negative
Headache with signs of total body illness (fever, stiff neck, rash)
Meningitis, encephalitis (inflammation of the brain tissue), Lyme disease, collagen vascular disease
A stiff neck, or inability to flex the neck due to pain, indicates inflammation of the meninges. Other signs of systemic illness indicates infection.
Neuroimaging, lumbar puncture, serology (diagnostic blood tests for infections)
Papilledema
Brain mass, benign intracranial hypertension (pseudotumor cerebri), meningitis
Increased intracranial pressure pushes on the eyes (from inside the brain) and causes papilledema.
Neuroimaging, lumbar puncture
Severe headache following head trauma
Brain bleeds (intracranial hemorrhage, subdural hematoma, epidural hematoma), post-traumatic headache
Trauma can cause bleeding in the brain or shake the nerves, causing a post-traumatic headache
Neuroimaging of brain, skull, and possibly cervical spine
Inability to move a limb
Arteriovenous malformation, collagen vascular disease, intracranial mass lesion
Focal neurological signs indicate something is pushing against nerves in the brain responsible for one part of the body
Neuroimaging, blood tests for collagen vascular diseases
Change in personality, consciousness, or mental status
Central nervous system infection, intracranial bleed, mass
Change in mental status indicates a global infection or inflammation of the brain, or a large bleed compressing the brainstem where the consciousness centers lie
Blood tests, lumbar puncture, neuroimaging
Headache triggered by cough, exertion or while engaged in sexual intercourse
Mass lesion, subarachnoid hemorrhage
Coughing and exertion increases the intra cranial pressure, which may cause a vessel to burst, causing a subarachnoid hemorrhage. A mass lesion already increases intracranial pressure, so an additional increase in intracranial pressure from coughing etc. will cause pain.
Neuroimaging, lumbar puncture
Old headaches[edit]
Old headaches are usually primary headaches and are not dangerous. They are most often caused by migraines or tension headaches. Migraines are often unilateral, pulsing headaches accompanied by nausea or vomiting. There may be an aura (visual symptoms, numbness or tingling) 30–60 minutes before the headache, warning the person of a headache. Migraines may also not have auras. Tension-type headaches usually have bilateral "bandlike" pressure on both sides of the head usually without nausea or vomiting. However, some symptoms from both headache groups may overlap. It is important to distinguish between the two because the treatments are different.
The mnemonic 'POUND' helps distinguish between migraines and tension-type headaches. POUND stands for:
Pulsatile quality of headacheOne-day duration (four to 72 hours)Unilateral locationNausea or vomitingDisabling intensity
One review article found that if 4–5 of the POUND characteristics are present, a migraine is 24 times as likely a diagnosis than a tension-type headache (likelihood ratio 24). If 3 characteristics of POUND are present, migraine is 3 times more likely a diagnosis than tension type headache (likelihood ratio 3). If only 2 POUND characteristics are present, tension-type headaches are 60% more likely (likelihood ratio 0.41). Another study found the following factors independently each increase the chance of migraine over tension-type headache: nausea, photophobia, phonophobia, exacerbation by physical activity, unilateral, throbbing quality, chocolate as a headache trigger, and cheese as a headache trigger.
Cluster headaches are relatively rare (1 in 1000 people) and are more common in men than women. They present with sudden onset explosive pain around one eye and are accompanied by autonomic symptoms (tearing, runny nose and red eye).
Temporomandibular jaw pain (chronic pain in the jaw joint), and cervicogenic headache (headache caused by pain in muscles of the neck) are also possible diagnoses.
For chronic, unexplained headaches, keeping a headache diary can be useful for tracking symptoms and identifying triggers, such as association with menstrual cycle, exercise and food. While mobile electronic diaries for smartphones are becoming increasingly common, a recent review found most are developed with a lack of evidence base and scientific expertise.
Cephalalgiaphobia is fear of headaches or getting a headache.
New headaches[edit]
New headaches are more likely to be dangerous secondary headaches. They can, however, simply be the first presentation of a chronic headache syndrome, like migraine or tension-type headaches.
One recommended diagnostic approach is as follows. If any urgent red flags are present such as visual loss, new seizures, new weakness, new confusion, further workup with imaging and possibly a lumbar puncture should be done (see red flags section for more details). If the headache is sudden onset (thunderclap headache), a computed tomography test to look for a brain bleed (subarachnoid hemorrhage) should be done. If the CT scan does not show a bleed, a lumbar puncture should be done to look for blood in the CSF, as the CT scan can be falsely negative and subarachnoid hemorrhages can be fatal. If there are signs of infection such as fever, rash, or stiff neck, a lumbar puncture to look for meningitis should be considered. If there is jaw claudication and scalp tenderness in an older person, a temporal artery biopsy to look for temporal arteritis should be performed and immediate treatment should be started.
Neuroimaging[edit]
Old headaches[edit]
The US Headache Consortium has guidelines for neuroimaging of non-acute headaches. Most old, chronic headaches do not require neuroimaging. If a person has the characteristic symptoms of a migraine, neuroimaging is not needed as it is very unlikely the person has an intracranial abnormality. If the person has neurological findings, such as weakness, on exam, neuroimaging may be considered.
New headaches[edit]
All people who present with red flags indicating a dangerous secondary headache should receive neuroimaging. The best form of neuroimaging for these headaches is controversial. Non-contrast computerized tomography (CT) scan is usually the first step in head imaging as it is readily available in Emergency Departments and hospitals and is cheaper than MRI. Non-contrast CT is best for identifying an acute head bleed. Magnetic Resonance Imaging (MRI) is best for brain tumors and problems in the posterior fossa, or back of the brain. MRI is more sensitive for identifying intracranial problems, however it can pick up brain abnormalities that are not relevant to the person's headaches.
The American College of Radiology recommends the following imaging tests for different specific situations:
Clinical Features
Recommended neuroimaging test
Headache in immunocompromised people (cancer, HIV)
MRI of head with or without contrast
Headache in people older than 60 with suspected temporal arteritis
MRI of head with or without contrast
Headache with suspected meningitis
CT or MRI without contrast
Severe headache in pregnancy
CT or MRI without contrast
Severe unilateral headache caused by possible dissection of carotid or arterial arteries
MRI of head with or without contrast, magnetic resonance angiography or Computed Tomography Angiography of head and neck.
Sudden onset headache or worst headache of life
CT of head without contrast, Computed Tomography Angiography of head and neck with contrast, magnetic resonance angiography of head and neck with and without contrast, MRI of head without contrast
Lumbar puncture[edit]
A lumbar puncture is a procedure in which cerebral spinal fluid is removed from the spine with a needle. A lumbar puncture is necessary to look for infection or blood in the spinal fluid. A lumbar puncture can also evaluate the pressure in the spinal column, which can be useful for people with idiopathic intracranial hypertension (usually young, obese women who have increased intracranial pressure), or other causes of increased intracranial pressure. In most cases, a CT scan should be done first.
Classification[edit]
Headaches are most thoroughly classified by the International Headache Society's International Classification of Headache Disorders (ICHD), which published the second edition in 2004. The third edition of the International Headache Classification was published in 2013 in a beta version ahead of the final version. This classification is accepted by the WHO.
Other classification systems exist. One of the first published attempts was in 1951. The US National Institutes of Health developed a classification system in 1962.
ICHD-2[edit]
Main article: International Classification of Headache Disorders
The International Classification of Headache Disorders (ICHD) is an in-depth hierarchical classification of headaches published by the International Headache Society. It contains explicit (operational) diagnostic criteria for headache disorders. The first version of the classification, ICHD-1, was published in 1988. The current revision, ICHD-2, was published in 2004. The classification uses numeric codes. The top, one-digit diagnostic level includes 14 headache groups. The first four of these are classified as primary headaches, groups 5-12 as secondary headaches, cranial neuralgia, central and primary facial pain and other headaches for the last two groups.
The ICHD-2 classification defines migraines, tension-types headaches, cluster headache and other trigeminal autonomic headache as the main types of primary headaches. Also, according to the same classification, stabbing headaches and headaches due to cough, exertion and sexual activity (sexual headache) are classified as primary headaches. The daily-persistent headaches along with the hypnic headache and thunderclap headaches are considered primary headaches as well.
Secondary headaches are classified based on their cause and not on their symptoms. According to the ICHD-2 classification, the main types of secondary headaches include those that are due to head or neck trauma such as whiplash injury, intracranial hematoma, post craniotomy or other head or neck injury. Headaches caused by cranial or cervical vascular disorders such as ischemic stroke and transient ischemic attack, non-traumatic intracranial hemorrhage, vascular malformations or arteritis are also defined as secondary headaches. This type of headache may also be caused by cerebral venous thrombosis or different intracranial vascular disorders. Other secondary headaches are those due to intracranial disorders that are not vascular such as low or high pressure of the cerebrospinal fluid pressure, non-infectious inflammatory disease, intracranial neoplasm, epileptic seizure or other types of disorders or diseases that are intracranial but that are not associated with the vasculature of the central nervous system.
ICHD-2 classifies headaches that are caused by the ingestion of a certain substance or by its withdrawal as secondary headaches as well. This type of headache may result from the overuse of some medications or exposure to some substances. HIV/AIDS, intracranial infections and systemic infections may also cause secondary headaches. The ICHD-2 system of classification includes the headaches associated with homeostasis disorders in the category of secondary headaches. This means that headaches caused by dialysis, high blood pressure, hypothyroidism, cephalalgia and even fasting are considered secondary headaches. Secondary headaches, according to the same classification system, can also be due to the injury of any of the facial structures including teeth, jaws, or temporomandibular joint. Headaches caused by psychiatric disorders such as somatization or psychotic disorders are also classified as secondary headaches.
The ICHD-2 classification puts cranial neuralgias and other types of neuralgia in a different category. According to this system, there are 19 types of neuralgias and headaches due to different central causes of facial pain. Moreover, the ICHD-2 includes a category that contains all the headaches that cannot be classified.
Although the ICHD-2 is the most complete headache classification there is and it includes frequency in the diagnostic criteria of some types of headaches (primarily primary headaches), it does not specifically code frequency or severity which are left at the discretion of the examiner.
NIH[edit]
Main article: NIH classification of headaches
The NIH classification consists of brief definitions of a limited number of headaches.
The NIH system of classification is more succinct and only describes five categories of headaches. In this case, primary headaches are those that do not show organic or structural causes. According to this classification, primary headaches can only be vascular, myogenic, cervicogenic, traction, and inflammatory.
Management[edit]
An old advertisement for a headache medicine
See also: Management of chronic headaches
Primary headache syndromes have many different possible treatments. In those with chronic headaches the long term use of opioids appears to result in greater harm than benefit.
Migraines[edit]
Migraine can be somewhat improved by lifestyle changes, but most people require medicines to control their symptoms. Medications are either to prevent getting migraines, or to reduce symptoms once a migraine starts.
Preventive medications are generally recommended when people have more than four attacks of migraine per month, headaches last longer than 12 hours or the headaches are very disabling. Possible therapies include beta blockers, antidepressants, anticonvulsants and NSAIDs. The type of preventive medicine is usually chosen based on the other symptoms the person has. For example, if the person also has depression, an antidepressant is a good choice.
Abortive therapies for migraines may be oral, if the migraine is mild to moderate, or may require stronger medicine given intravenously or intramuscularly. Mild to moderate headaches should first be treated with acetaminophen (paracetamol) or NSAIDs, like ibuprofen. If accompanied by nausea or vomiting, an antiemetic such as metoclopramide (Reglan) can be given orally or rectally. Moderate to severe attacks should be treated first with an oral triptan, a medication that mimics serotonin (an agonist) and causes mild vasoconstriction. If accompanied by nausea and vomiting, parenteral (through a needle in the skin) triptans and antiemetics can be given.
Sphenopalatine ganglion block (SPG block, also known nasal ganglion block or pterygopalatine ganglion blocks) can abort and prevent migraines, tension headaches and cluster headaches. It was originally described by American ENT surgeon Greenfield Sluder in 1908. Both blocks and neurostimulation have been studied as treatment for headaches.
Several complementary and alternative strategies can help with migraines. The American Academy of Neurology guidelines for migraine treatment in 2000 stated relaxation training, electromyographic feedback and cognitive behavioral therapy may be considered for migraine treatment, along with medications.
Tension-type headaches[edit]
Tension-type headaches can usually be managed with NSAIDs (ibuprofen, naproxen, aspirin), or acetaminophen. Triptans are not helpful in tension-type headaches unless the person also has migraines. For chronic tension type headaches, amitriptyline is the only medication proven to help. Amitriptyline is a medication which treats depression and also independently treats pain. It works by blocking the reuptake of serotonin and norepinephrine, and also reduces muscle tenderness by a separate mechanism. Studies evaluating acupuncture for tension-type headaches have been mixed. Overall, they show that acupuncture is probably not helpful for tension-type headaches.
Cluster headaches[edit]
Abortive therapy for cluster headaches includes subcutaneous sumatriptan (injected under the skin) and triptan nasal sprays. High flow oxygen therapy also helps with relief.
For people with extended periods of cluster headaches, preventive therapy can be necessary. Verapamil is recommended as first line treatment. Lithium can also be useful. For people with shorter bouts, a short course of prednisone (10 days) can be helpful. Ergotamine is useful if given 1–2 hours before an attack.
Secondary headaches[edit]
Treatment of secondary headaches involves treating the underlying cause. For example, a person with meningitis will require antibiotics. A person with a brain tumor may require surgery, chemotherapy or brain radiation.
Neuromodulation[edit]
Peripheral neuromodulation has tentative benefits in primary headaches including cluster headaches and chronic migraine. How it may work is still being looked into.
Epidemiology[edit]
Literature reviews find that approximately 64–77% of adults have had a headache at some point in their lives. During each year, on average, 46–53% of people have headaches. However, the prevalence of headache varies widely depending on how the survey was conducted, with studies finding lifetime prevalence of as low as 8% to as high as 96%. Most of these headaches are not dangerous. Only approximately 1–5% of people who seek emergency treatment for headaches have a serious underlying cause.
More than 90% of headaches are primary headaches. Most of these primary headaches are tension headaches. Most people with tension headaches have "episodic" tension headaches that come and go. Only 3.3% of adults have chronic tension headaches, with headaches for more than 15 days in a month.
Approximately 12–18% of people in the world have migraines. More women than men experience migraines. In Europe and North America, 5–9% of men experience migraines, while 12–25% of women experience migraines.
Cluster headaches are relatively uncommon. They affect only 1–3 per thousand people in the world. Cluster headaches affect approximately three times as many men as women.
History[edit]
An 1819 caricature by George Cruikshank depicting a headache
The first recorded classification system was published by Aretaeus of Cappadocia, a medical scholar of Greco-Roman antiquity. He made a distinction between three different types of headache: i) cephalalgia, by which he indicates a sudden onset, temporary headache; ii) cephalea, referring to a chronic type of headache; and iii) heterocrania, a paroxysmal headache on one side of the head.
Another classification system that resembles the modern ones was published by Thomas Willis, in De Cephalalgia in 1672. In 1787 Christian Baur generally divided headaches into idiopathic (primary headaches) and symptomatic (secondary ones), and defined 84 categories.
Children[edit]
In general, children experience the same types of headaches as adults do, but their symptoms may be slightly different. The diagnostic approach to headaches in children is similar to that of adults. However, young children may not be able to verbalize pain well. If a young child is fussy, they may have a headache.
Approximately 1% of emergency department visits for children are for headache. Most of these headaches are not dangerous. The most common type of headache seen in pediatric emergency rooms is headache caused by a cold (28.5%). Other headaches diagnosed in the emergency department include post-traumatic headache (20%), headache related to a problem with a ventriculoperitoneal shunt (a device put into the brain to remove excess CSF and reduce pressure in the brain) (11.5%) and migraine (8.5%). The most common serious headaches found in children include brain bleeds (subdural hematoma, epidural hematoma), brain abscesses, meningitis and ventriculoperitoneal shunt malfunction. Only 4–6.9% of kids with a headache have a serious cause.
Just as in adults, most headaches are benign, but when head pain is accompanied with other symptoms such as speech problems, muscle weakness, and loss of vision, a more serious underlying cause may exist: hydrocephalus, meningitis, encephalitis, abscess, hemorrhage, tumor, blood clots, or head trauma. In these cases, the headache evaluation may include CT scan or MRI in order to look for possible structural disorders of the central nervous system. If a child with a recurrent headache has a normal physical exam, neuroimaging is not recommended. Guidelines state children with abnormal neurologic exams, confusion, seizures and recent onset of worst headache of life, change in headache type or anything suggesting neurologic problems should receive neuroimaging.
When children complain of headaches, many parents are concerned about a brain tumor. Generally, headaches caused by brain masses are incapacitating and accompanied by vomiting. One study found characteristics associated with brain tumor in children are: headache for greater than 6 months, headache related to sleep, vomiting, confusion, no visual symptoms, no family history of migraine and abnormal neurologic exam.
Some measures can help prevent headaches in children. Drinking plenty of water throughout the day, avoiding caffeine, getting enough and regular sleep, eating balanced meals at the proper times, and reducing stress and excess of activities may prevent headaches. Treatments for children are similar to those for adults, however certain medications such as narcotics should not be given to children.
Children who have headaches will not necessarily have headaches as adults. In one study of 100 children with headache, eight years later 44% of those with tension headache and 28% of those with migraines were headache free. In another study of people with chronic daily headache, 75% did not have chronic daily headaches two years later, and 88% did not have chronic daily headaches eight years later.
See also[edit]
Eye strain | biology | 1181741 | https://no.wikipedia.org/wiki/Head%20%26%20Shoulders | Head & Shoulders | Head & Shoulders er et merke av anti-flass sjampo som produseres av Procter & Gamble. Det ble først introdusert i USA i november 1961 som en blågrønn sjampo. Det finnes nå ni forskjellige Head & Shoulders-sjampoer for forskjellige hårtyper, og fargen har blitt endret til hvit.
Eksterne lenker
Varemerker
Hårpleie | norwegian_bokmål | 1.472537 |
brain_no_pain_receptors_headache/Brain.txt |
The brain is an organ that serves as the center of the nervous system in all vertebrate and most invertebrate animals. In vertebrates, a small part of the brain called the hypothalamus is the neural control center for all endocrine systems. The brain is the largest cluster of neurons in the body and is typically located in the head, usually near organs for special senses such as vision, hearing and olfaction.
It is the most energy-consuming organ of the body, and the most specialized, responsible for endocrine regulation, sensory perception, motor control, and the development of intelligence.
While invertebrate brains arise from paired segmental ganglia (each of which is only responsible for the respective body segment) of the ventral nerve cord, vertebrate brains develop axially from the midline dorsal nerve cord as a vesicular enlargement at the rostral end of the neural tube, with centralized control over all body segments. All vertebrate brains can be embryonically divided into three parts: the forebrain (prosencephalon, subdivided into telencephalon and diencephalon), midbrain (mesencephalon) and hindbrain (rhombencephalon, subdivided into metencephalon and myelencephalon). The spinal cord, which directly interacts with somatic functions below the head, can be considered a caudal extension of the myelencephalon enclosed inside the vertebral column. Together, the brain and spinal cord constitute the central nervous system in all vertebrates.
In humans, the cerebral cortex contains approximately 14–16 billion neurons, and the estimated number of neurons in the cerebellum is 55–70 billion. Each neuron is connected by synapses to several thousand other neurons, typically communicating with one another via root-like protrusions called dendrites and long fiber-like extensions called axons, which are usually myelinated and carry trains of rapid micro-electric signal pulses called action potentials to target specific recipient cells in other areas of the brain or distant parts of the body. The prefrontal cortex, which controls executive functions, is particularly well developed in humans.
Physiologically, brains exert centralized control over a body's other organs. They act on the rest of the body both by generating patterns of muscle activity and by driving the secretion of chemicals called hormones. This centralized control allows rapid and coordinated responses to changes in the environment. Some basic types of responsiveness such as reflexes can be mediated by the spinal cord or peripheral ganglia, but sophisticated purposeful control of behavior based on complex sensory input requires the information integrating capabilities of a centralized brain.
The operations of individual brain cells are now understood in considerable detail but the way they cooperate in ensembles of millions is yet to be solved. Recent models in modern neuroscience treat the brain as a biological computer, very different in mechanism from a digital computer, but similar in the sense that it acquires information from the surrounding world, stores it, and processes it in a variety of ways.
This article compares the properties of brains across the entire range of animal species, with the greatest attention to vertebrates. It deals with the human brain insofar as it shares the properties of other brains. The ways in which the human brain differs from other brains are covered in the human brain article. Several topics that might be covered here are instead covered there because much more can be said about them in a human context. The most important that are covered in the human brain article are brain disease and the effects of brain damage.
The shape and size of the brain varies greatly between species, and identifying common features is often difficult. Nevertheless, there are a number of principles of brain architecture that apply across a wide range of species. Some aspects of brain structure are common to almost the entire range of animal species; others distinguish "advanced" brains from more primitive ones, or distinguish vertebrates from invertebrates.
The simplest way to gain information about brain anatomy is by visual inspection, but many more sophisticated techniques have been developed. Brain tissue in its natural state is too soft to work with, but it can be hardened by immersion in alcohol or other fixatives, and then sliced apart for examination of the interior. Visually, the interior of the brain consists of areas of so-called grey matter, with a dark color, separated by areas of white matter, with a lighter color. Further information can be gained by staining slices of brain tissue with a variety of chemicals that bring out areas where specific types of molecules are present in high concentrations. It is also possible to examine the microstructure of brain tissue using a microscope, and to trace the pattern of connections from one brain area to another.
The brains of all species are composed primarily of two broad classes of cells: neurons and glial cells. Glial cells (also known as glia or neuroglia) come in several types, and perform a number of critical functions, including structural support, metabolic support, insulation, and guidance of development. Neurons, however, are usually considered the most important cells in the brain.
The property that makes neurons unique is their ability to send signals to specific target cells over long distances. They send these signals by means of an axon, which is a thin protoplasmic fiber that extends from the cell body and projects, usually with numerous branches, to other areas, sometimes nearby, sometimes in distant parts of the brain or body. The length of an axon can be extraordinary: for example, if a pyramidal cell (an excitatory neuron) of the cerebral cortex were magnified so that its cell body became the size of a human body, its axon, equally magnified, would become a cable a few centimeters in diameter, extending more than a kilometer. These axons transmit signals in the form of electrochemical pulses called action potentials, which last less than a thousandth of a second and travel along the axon at speeds of 1–100 meters per second. Some neurons emit action potentials constantly, at rates of 10–100 per second, usually in irregular patterns; other neurons are quiet most of the time, but occasionally emit a burst of action potentials.
Axons transmit signals to other neurons by means of specialized junctions called synapses. A single axon may make as many as several thousand synaptic connections with other cells. When an action potential, traveling along an axon, arrives at a synapse, it causes a chemical called a neurotransmitter to be released. The neurotransmitter binds to receptor molecules in the membrane of the target cell.
Synapses are the key functional elements of the brain. The essential function of the brain is cell-to-cell communication, and synapses are the points at which communication occurs. The human brain has been estimated to contain approximately 100 trillion synapses; even the brain of a fruit fly contains several million. The functions of these synapses are very diverse: some are excitatory (exciting the target cell); others are inhibitory; others work by activating second messenger systems that change the internal chemistry of their target cells in complex ways. A large number of synapses are dynamically modifiable; that is, they are capable of changing strength in a way that is controlled by the patterns of signals that pass through them. It is widely believed that activity-dependent modification of synapses is the brain's primary mechanism for learning and memory.
Most of the space in the brain is taken up by axons, which are often bundled together in what are called nerve fiber tracts. A myelinated axon is wrapped in a fatty insulating sheath of myelin, which serves to greatly increase the speed of signal propagation. (There are also unmyelinated axons). Myelin is white, making parts of the brain filled exclusively with nerve fibers appear as light-colored white matter, in contrast to the darker-colored grey matter that marks areas with high densities of neuron cell bodies.
Except for a few primitive organisms such as sponges (which have no nervous system) and cnidarians (which have a diffuse nervous system consisting of a nerve net), all living multicellular animals are bilaterians, meaning animals with a bilaterally symmetric body plan (that is, left and right sides that are approximate mirror images of each other). All bilaterians are thought to have descended from a common ancestor that appeared late in the Cryogenian period, 700–650 million years ago, and it has been hypothesized that this common ancestor had the shape of a simple tubeworm with a segmented body. At a schematic level, that basic worm-shape continues to be reflected in the body and nervous system architecture of all modern bilaterians, including vertebrates. The fundamental bilateral body form is a tube with a hollow gut cavity running from the mouth to the anus, and a nerve cord with an enlargement (a ganglion) for each body segment, with an especially large ganglion at the front, called the brain. The brain is small and simple in some species, such as nematode worms; in other species, such as vertebrates, it is a large and very complex organ. Some types of worms, such as leeches, also have an enlarged ganglion at the back end of the nerve cord, known as a "tail brain".
There are a few types of existing bilaterians that lack a recognizable brain, including echinoderms and tunicates. It has not been definitively established whether the existence of these brainless species indicates that the earliest bilaterians lacked a brain, or whether their ancestors evolved in a way that led to the disappearance of a previously existing brain structure.
This category includes tardigrades, arthropods, molluscs, and numerous types of worms. The diversity of invertebrate body plans is matched by an equal diversity in brain structures.
Two groups of invertebrates have notably complex brains: arthropods (insects, crustaceans, arachnids, and others), and cephalopods (octopuses, squids, and similar molluscs). The brains of arthropods and cephalopods arise from twin parallel nerve cords that extend through the body of the animal. Arthropods have a central brain, the supraesophageal ganglion, with three divisions and large optical lobes behind each eye for visual processing. Cephalopods such as the octopus and squid have the largest brains of any invertebrates.
There are several invertebrate species whose brains have been studied intensively because they have properties that make them convenient for experimental work:
The first vertebrates appeared over 500 million years ago (Mya), during the Cambrian period, and may have resembled the modern hagfish in form. Jawed fish appeared by 445 Mya, amphibians by 350 Mya, reptiles by 310 Mya and mammals by 200 Mya (approximately). Each species has an equally long evolutionary history, but the brains of modern hagfishes, lampreys, sharks, amphibians, reptiles, and mammals show a gradient of size and complexity that roughly follows the evolutionary sequence. All of these brains contain the same set of basic anatomical components, but many are rudimentary in the hagfish, whereas in mammals the foremost part (the telencephalon) is greatly elaborated and expanded.
Brains are most commonly compared in terms of their size. The relationship between brain size, body size and other variables has been studied across a wide range of vertebrate species. As a rule, brain size increases with body size, but not in a simple linear proportion. In general, smaller animals tend to have larger brains, measured as a fraction of body size. For mammals, the relationship between brain volume and body mass essentially follows a power law with an exponent of about 0.75. This formula describes the central tendency, but every family of mammals departs from it to some degree, in a way that reflects in part the complexity of their behavior. For example, primates have brains 5 to 10 times larger than the formula predicts. Predators tend to have larger brains than their prey, relative to body size.
All vertebrate brains share a common underlying form, which appears most clearly during early stages of embryonic development. In its earliest form, the brain appears as three swellings at the front end of the neural tube; these swellings eventually become the forebrain, midbrain, and hindbrain (the prosencephalon, mesencephalon, and rhombencephalon, respectively). At the earliest stages of brain development, the three areas are roughly equal in size. In many classes of vertebrates, such as fish and amphibians, the three parts remain similar in size in the adult, but in mammals the forebrain becomes much larger than the other parts, and the midbrain becomes very small.
The brains of vertebrates are made of very soft tissue. Living brain tissue is pinkish on the outside and mostly white on the inside, with subtle variations in color. Vertebrate brains are surrounded by a system of connective tissue membranes called meninges that separate the skull from the brain. Blood vessels enter the central nervous system through holes in the meningeal layers. The cells in the blood vessel walls are joined tightly to one another, forming the blood–brain barrier, which blocks the passage of many toxins and pathogens (though at the same time blocking antibodies and some drugs, thereby presenting special challenges in treatment of diseases of the brain).
Neuroanatomists usually divide the vertebrate brain into six main regions: the telencephalon (cerebral hemispheres), diencephalon (thalamus and hypothalamus), mesencephalon (midbrain), cerebellum, pons, and medulla oblongata. Each of these areas has a complex internal structure. Some parts, such as the cerebral cortex and the cerebellar cortex, consist of layers that are folded or convoluted to fit within the available space. Other parts, such as the thalamus and hypothalamus, consist of clusters of many small nuclei. Thousands of distinguishable areas can be identified within the vertebrate brain based on fine distinctions of neural structure, chemistry, and connectivity.
Although the same basic components are present in all vertebrate brains, some branches of vertebrate evolution have led to substantial distortions of brain geometry, especially in the forebrain area. The brain of a shark shows the basic components in a straightforward way, but in teleost fishes (the great majority of existing fish species), the forebrain has become "everted", like a sock turned inside out. In birds, there are also major changes in forebrain structure. These distortions can make it difficult to match brain components from one species with those of another species.
Here is a list of some of the most important vertebrate brain components, along with a brief description of their functions as currently understood:
The most obvious difference between the brains of mammals and other vertebrates is in terms of size. On average, a mammal has a brain roughly twice as large as that of a bird of the same body size, and ten times as large as that of a reptile of the same body size.
Size, however, is not the only difference: there are also substantial differences in shape. The hindbrain and midbrain of mammals are generally similar to those of other vertebrates, but dramatic differences appear in the forebrain, which is greatly enlarged and also altered in structure. The cerebral cortex is the part of the brain that most strongly distinguishes mammals. In non-mammalian vertebrates, the surface of the cerebrum is lined with a comparatively simple three-layered structure called the pallium. In mammals, the pallium evolves into a complex six-layered structure called neocortex or isocortex. Several areas at the edge of the neocortex, including the hippocampus and amygdala, are also much more extensively developed in mammals than in other vertebrates.
The elaboration of the cerebral cortex carries with it changes to other brain areas. The superior colliculus, which plays a major role in visual control of behavior in most vertebrates, shrinks to a small size in mammals, and many of its functions are taken over by visual areas of the cerebral cortex. The cerebellum of mammals contains a large portion (the neocerebellum) dedicated to supporting the cerebral cortex, which has no counterpart in other vertebrates.
The brains of humans and other primates contain the same structures as the brains of other mammals, but are generally larger in proportion to body size. The encephalization quotient (EQ) is used to compare brain sizes across species. It takes into account the nonlinearity of the brain-to-body relationship. Humans have an average EQ in the 7-to-8 range, while most other primates have an EQ in the 2-to-3 range. Dolphins have values higher than those of primates other than humans, but nearly all other mammals have EQ values that are substantially lower.
Most of the enlargement of the primate brain comes from a massive expansion of the cerebral cortex, especially the prefrontal cortex and the parts of the cortex involved in vision. The visual processing network of primates includes at least 30 distinguishable brain areas, with a complex web of interconnections. It has been estimated that visual processing areas occupy more than half of the total surface of the primate neocortex. The prefrontal cortex carries out functions that include planning, working memory, motivation, attention, and executive control. It takes up a much larger proportion of the brain for primates than for other species, and an especially large fraction of the human brain.
The brain develops in an intricately orchestrated sequence of stages. It changes in shape from a simple swelling at the front of the nerve cord in the earliest embryonic stages, to a complex array of areas and connections. Neurons are created in special zones that contain stem cells, and then migrate through the tissue to reach their ultimate locations. Once neurons have positioned themselves, their axons sprout and navigate through the brain, branching and extending as they go, until the tips reach their targets and form synaptic connections. In a number of parts of the nervous system, neurons and synapses are produced in excessive numbers during the early stages, and then the unneeded ones are pruned away.
For vertebrates, the early stages of neural development are similar across all species. As the embryo transforms from a round blob of cells into a wormlike structure, a narrow strip of ectoderm running along the midline of the back is induced to become the neural plate, the precursor of the nervous system. The neural plate folds inward to form the neural groove, and then the lips that line the groove merge to enclose the neural tube, a hollow cord of cells with a fluid-filled ventricle at the center. At the front end, the ventricles and cord swell to form three vesicles that are the precursors of the prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain). At the next stage, the forebrain splits into two vesicles called the telencephalon (which will contain the cerebral cortex, basal ganglia, and related structures) and the diencephalon (which will contain the thalamus and hypothalamus). At about the same time, the hindbrain splits into the metencephalon (which will contain the cerebellum and pons) and the myelencephalon (which will contain the medulla oblongata). Each of these areas contains proliferative zones where neurons and glial cells are generated; the resulting cells then migrate, sometimes for long distances, to their final positions.
Once a neuron is in place, it extends dendrites and an axon into the area around it. Axons, because they commonly extend a great distance from the cell body and need to reach specific targets, grow in a particularly complex way. The tip of a growing axon consists of a blob of protoplasm called a growth cone, studded with chemical receptors. These receptors sense the local environment, causing the growth cone to be attracted or repelled by various cellular elements, and thus to be pulled in a particular direction at each point along its path. The result of this pathfinding process is that the growth cone navigates through the brain until it reaches its destination area, where other chemical cues cause it to begin generating synapses. Considering the entire brain, thousands of genes create products that influence axonal pathfinding.
The synaptic network that finally emerges is only partly determined by genes, though. In many parts of the brain, axons initially "overgrow", and then are "pruned" by mechanisms that depend on neural activity. In the projection from the eye to the midbrain, for example, the structure in the adult contains a very precise mapping, connecting each point on the surface of the retina to a corresponding point in a midbrain layer. In the first stages of development, each axon from the retina is guided to the right general vicinity in the midbrain by chemical cues, but then branches very profusely and makes initial contact with a wide swath of midbrain neurons. The retina, before birth, contains special mechanisms that cause it to generate waves of activity that originate spontaneously at a random point and then propagate slowly across the retinal layer. These waves are useful because they cause neighboring neurons to be active at the same time; that is, they produce a neural activity pattern that contains information about the spatial arrangement of the neurons. This information is exploited in the midbrain by a mechanism that causes synapses to weaken, and eventually vanish, if activity in an axon is not followed by activity of the target cell. The result of this sophisticated process is a gradual tuning and tightening of the map, leaving it finally in its precise adult form.
Similar things happen in other brain areas: an initial synaptic matrix is generated as a result of genetically determined chemical guidance, but then gradually refined by activity-dependent mechanisms, partly driven by internal dynamics, partly by external sensory inputs. In some cases, as with the retina-midbrain system, activity patterns depend on mechanisms that operate only in the developing brain, and apparently exist solely to guide development.
In humans and many other mammals, new neurons are created mainly before birth, and the infant brain contains substantially more neurons than the adult brain. There are, however, a few areas where new neurons continue to be generated throughout life. The two areas for which adult neurogenesis is well established are the olfactory bulb, which is involved in the sense of smell, and the dentate gyrus of the hippocampus, where there is evidence that the new neurons play a role in storing newly acquired memories. With these exceptions, however, the set of neurons that is present in early childhood is the set that is present for life. Glial cells are different: as with most types of cells in the body, they are generated throughout the lifespan.
There has long been debate about whether the qualities of mind, personality, and intelligence can be attributed to heredity or to upbringing. Although many details remain to be settled, neuroscience shows that both factors are important. Genes determine both the general form of the brain and how it reacts to experience, but experience is required to refine the matrix of synaptic connections, resulting in greatly increased complexity. The presence or absence of experience is critical at key periods of development. Additionally, the quantity and quality of experience are important. For example, animals raised in enriched environments demonstrate thick cerebral cortices, indicating a high density of synaptic connections, compared to animals with restricted levels of stimulation.
The functions of the brain depend on the ability of neurons to transmit electrochemical signals to other cells, and their ability to respond appropriately to electrochemical signals received from other cells. The electrical properties of neurons are controlled by a wide variety of biochemical and metabolic processes, most notably the interactions between neurotransmitters and receptors that take place at synapses.
Neurotransmitters are chemicals that are released at synapses when the local membrane is depolarised and Ca enters into the cell, typically when an action potential arrives at the synapse – neurotransmitters attach themselves to receptor molecules on the membrane of the synapse's target cell (or cells), and thereby alter the electrical or chemical properties of the receptor molecules. With few exceptions, each neuron in the brain releases the same chemical neurotransmitter, or combination of neurotransmitters, at all the synaptic connections it makes with other neurons; this rule is known as Dale's principle. Thus, a neuron can be characterized by the neurotransmitters that it releases. The great majority of psychoactive drugs exert their effects by altering specific neurotransmitter systems. This applies to drugs such as cannabinoids, nicotine, heroin, cocaine, alcohol, fluoxetine, chlorpromazine, and many others.
The two neurotransmitters that are most widely found in the vertebrate brain are glutamate, which almost always exerts excitatory effects on target neurons, and gamma-aminobutyric acid (GABA), which is almost always inhibitory. Neurons using these transmitters can be found in nearly every part of the brain. Because of their ubiquity, drugs that act on glutamate or GABA tend to have broad and powerful effects. Some general anesthetics act by reducing the effects of glutamate; most tranquilizers exert their sedative effects by enhancing the effects of GABA.
There are dozens of other chemical neurotransmitters that are used in more limited areas of the brain, often areas dedicated to a particular function. Serotonin, for example—the primary target of many antidepressant drugs and many dietary aids—comes exclusively from a small brainstem area called the raphe nuclei. Norepinephrine, which is involved in arousal, comes exclusively from a nearby small area called the locus coeruleus. Other neurotransmitters such as acetylcholine and dopamine have multiple sources in the brain but are not as ubiquitously distributed as glutamate and GABA.
As a side effect of the electrochemical processes used by neurons for signaling, brain tissue generates electric fields when it is active. When large numbers of neurons show synchronized activity, the electric fields that they generate can be large enough to detect outside the skull, using electroencephalography (EEG) or magnetoencephalography (MEG). EEG recordings, along with recordings made from electrodes implanted inside the brains of animals such as rats, show that the brain of a living animal is constantly active, even during sleep. Each part of the brain shows a mixture of rhythmic and nonrhythmic activity, which may vary according to behavioral state. In mammals, the cerebral cortex tends to show large slow delta waves during sleep, faster alpha waves when the animal is awake but inattentive, and chaotic-looking irregular activity when the animal is actively engaged in a task, called beta and gamma waves. During an epileptic seizure, the brain's inhibitory control mechanisms fail to function and electrical activity rises to pathological levels, producing EEG traces that show large wave and spike patterns not seen in a healthy brain. Relating these population-level patterns to the computational functions of individual neurons is a major focus of current research in neurophysiology.
All vertebrates have a blood–brain barrier that allows metabolism inside the brain to operate differently from metabolism in other parts of the body. The neurovascular unit regulates cerebral blood flow so that activated neurons can be supplied with energy. Glial cells play a major role in brain metabolism by controlling the chemical composition of the fluid that surrounds neurons, including levels of ions and nutrients.
Brain tissue consumes a large amount of energy in proportion to its volume, so large brains place severe metabolic demands on animals. The need to limit body weight in order, for example, to fly, has apparently led to selection for a reduction of brain size in some species, such as bats. Most of the brain's energy consumption goes into sustaining the electric charge (membrane potential) of neurons. Most vertebrate species devote between 2% and 8% of basal metabolism to the brain. In primates, however, the percentage is much higher—in humans it rises to 20–25%. The energy consumption of the brain does not vary greatly over time, but active regions of the cerebral cortex consume somewhat more energy than inactive regions; this forms the basis for the functional brain imaging methods of PET, fMRI, and NIRS. The brain typically gets most of its energy from oxygen-dependent metabolism of glucose (i.e., blood sugar), but ketones provide a major alternative source, together with contributions from medium chain fatty acids (caprylic and heptanoic acids), lactate, acetate, and possibly amino acids.
Information from the sense organs is collected in the brain. There it is used to determine what actions the organism is to take. The brain processes the raw data to extract information about the structure of the environment. Next it combines the processed information with information about the current needs of the animal and with memory of past circumstances. Finally, on the basis of the results, it generates motor response patterns. These signal-processing tasks require intricate interplay between a variety of functional subsystems.
The function of the brain is to provide coherent control over the actions of an animal. A centralized brain allows groups of muscles to be co-activated in complex patterns; it also allows stimuli impinging on one part of the body to evoke responses in other parts, and it can prevent different parts of the body from acting at cross-purposes to each other.
The human brain is provided with information about light, sound, the chemical composition of the atmosphere, temperature, the position of the body in space (proprioception), the chemical composition of the bloodstream, and more. In other animals additional senses are present, such as the infrared heat-sense of snakes, the magnetic field sense of some birds, or the electric field sense mainly seen in aquatic animals.
Each sensory system begins with specialized receptor cells, such as photoreceptor cells in the retina of the eye, or vibration-sensitive hair cells in the cochlea of the ear. The axons of sensory receptor cells travel into the spinal cord or brain, where they transmit their signals to a first-order sensory nucleus dedicated to one specific sensory modality. This primary sensory nucleus sends information to higher-order sensory areas that are dedicated to the same modality. Eventually, via a way-station in the thalamus, the signals are sent to the cerebral cortex, where they are processed to extract the relevant features, and integrated with signals coming from other sensory systems.
Motor systems are areas of the brain that are involved in initiating body movements, that is, in activating muscles. Except for the muscles that control the eye, which are driven by nuclei in the midbrain, all the voluntary muscles in the body are directly innervated by motor neurons in the spinal cord and hindbrain. Spinal motor neurons are controlled both by neural circuits intrinsic to the spinal cord, and by inputs that descend from the brain. The intrinsic spinal circuits implement many reflex responses, and contain pattern generators for rhythmic movements such as walking or swimming. The descending connections from the brain allow for more sophisticated control.
The brain contains several motor areas that project directly to the spinal cord. At the lowest level are motor areas in the medulla and pons, which control stereotyped movements such as walking, breathing, or swallowing. At a higher level are areas in the midbrain, such as the red nucleus, which is responsible for coordinating movements of the arms and legs. At a higher level yet is the primary motor cortex, a strip of tissue located at the posterior edge of the frontal lobe. The primary motor cortex sends projections to the subcortical motor areas, but also sends a massive projection directly to the spinal cord, through the pyramidal tract. This direct corticospinal projection allows for precise voluntary control of the fine details of movements. Other motor-related brain areas exert secondary effects by projecting to the primary motor areas. Among the most important secondary areas are the premotor cortex, supplementary motor area, basal ganglia, and cerebellum. In addition to all of the above, the brain and spinal cord contain extensive circuitry to control the autonomic nervous system which controls the movement of the smooth muscle of the body.
Many animals alternate between sleeping and waking in a daily cycle. Arousal and alertness are also modulated on a finer time scale by a network of brain areas. A key component of the sleep system is the suprachiasmatic nucleus (SCN), a tiny part of the hypothalamus located directly above the point at which the optic nerves from the two eyes cross. The SCN contains the body's central biological clock. Neurons there show activity levels that rise and fall with a period of about 24 hours, circadian rhythms: these activity fluctuations are driven by rhythmic changes in expression of a set of "clock genes". The SCN continues to keep time even if it is excised from the brain and placed in a dish of warm nutrient solution, but it ordinarily receives input from the optic nerves, through the retinohypothalamic tract (RHT), that allows daily light-dark cycles to calibrate the clock.
The SCN projects to a set of areas in the hypothalamus, brainstem, and midbrain that are involved in implementing sleep-wake cycles. An important component of the system is the reticular formation, a group of neuron-clusters scattered diffusely through the core of the lower brain. Reticular neurons send signals to the thalamus, which in turn sends activity-level-controlling signals to every part of the cortex. Damage to the reticular formation can produce a permanent state of coma.
Sleep involves great changes in brain activity. Until the 1950s it was generally believed that the brain essentially shuts off during sleep, but this is now known to be far from true; activity continues, but patterns become very different. There are two types of sleep: REM sleep (with dreaming) and NREM (non-REM, usually without dreaming) sleep, which repeat in slightly varying patterns throughout a sleep episode. Three broad types of distinct brain activity patterns can be measured: REM, light NREM and deep NREM. During deep NREM sleep, also called slow wave sleep, activity in the cortex takes the form of large synchronized waves, whereas in the waking state it is noisy and desynchronized. Levels of the neurotransmitters norepinephrine and serotonin drop during slow wave sleep, and fall almost to zero during REM sleep; levels of acetylcholine show the reverse pattern.
For any animal, survival requires maintaining a variety of parameters of bodily state within a limited range of variation: these include temperature, water content, salt concentration in the bloodstream, blood glucose levels, blood oxygen level, and others. The ability of an animal to regulate the internal environment of its body—the milieu intérieur, as the pioneering physiologist Claude Bernard called it—is known as homeostasis (Greek for "standing still"). Maintaining homeostasis is a crucial function of the brain. The basic principle that underlies homeostasis is negative feedback: any time a parameter diverges from its set-point, sensors generate an error signal that evokes a response that causes the parameter to shift back toward its optimum value. (This principle is widely used in engineering, for example in the control of temperature using a thermostat.)
In vertebrates, the part of the brain that plays the greatest role is the hypothalamus, a small region at the base of the forebrain whose size does not reflect its complexity or the importance of its function. The hypothalamus is a collection of small nuclei, most of which are involved in basic biological functions. Some of these functions relate to arousal or to social interactions such as sexuality, aggression, or maternal behaviors; but many of them relate to homeostasis. Several hypothalamic nuclei receive input from sensors located in the lining of blood vessels, conveying information about temperature, sodium level, glucose level, blood oxygen level, and other parameters. These hypothalamic nuclei send output signals to motor areas that can generate actions to rectify deficiencies. Some of the outputs also go to the pituitary gland, a tiny gland attached to the brain directly underneath the hypothalamus. The pituitary gland secretes hormones into the bloodstream, where they circulate throughout the body and induce changes in cellular activity.
The individual animals need to express survival-promoting behaviors, such as seeking food, water, shelter, and a mate. The motivational system in the brain monitors the current state of satisfaction of these goals, and activates behaviors to meet any needs that arise. The motivational system works largely by a reward–punishment mechanism. When a particular behavior is followed by favorable consequences, the reward mechanism in the brain is activated, which induces structural changes inside the brain that cause the same behavior to be repeated later, whenever a similar situation arises. Conversely, when a behavior is followed by unfavorable consequences, the brain's punishment mechanism is activated, inducing structural changes that cause the behavior to be suppressed when similar situations arise in the future.
Most organisms studied to date use a reward–punishment mechanism: for instance, worms and insects can alter their behavior to seek food sources or to avoid dangers. In vertebrates, the reward-punishment system is implemented by a specific set of brain structures, at the heart of which lie the basal ganglia, a set of interconnected areas at the base of the forebrain. The basal ganglia are the central site at which decisions are made: the basal ganglia exert a sustained inhibitory control over most of the motor systems in the brain; when this inhibition is released, a motor system is permitted to execute the action it is programmed to carry out. Rewards and punishments function by altering the relationship between the inputs that the basal ganglia receive and the decision-signals that are emitted. The reward mechanism is better understood than the punishment mechanism, because its role in drug abuse has caused it to be studied very intensively. Research has shown that the neurotransmitter dopamine plays a central role: addictive drugs such as cocaine, amphetamine, and nicotine either cause dopamine levels to rise or cause the effects of dopamine inside the brain to be enhanced.
Almost all animals are capable of modifying their behavior as a result of experience—even the most primitive types of worms. Because behavior is driven by brain activity, changes in behavior must somehow correspond to changes inside the brain. Already in the late 19th century theorists like Santiago Ramón y Cajal argued that the most plausible explanation is that learning and memory are expressed as changes in the synaptic connections between neurons. Until 1970, however, experimental evidence to support the synaptic plasticity hypothesis was lacking. In 1971 Tim Bliss and Terje Lømo published a paper on a phenomenon now called long-term potentiation: the paper showed clear evidence of activity-induced synaptic changes that lasted for at least several days. Since then technical advances have made these sorts of experiments much easier to carry out, and thousands of studies have been made that have clarified the mechanism of synaptic change, and uncovered other types of activity-driven synaptic change in a variety of brain areas, including the cerebral cortex, hippocampus, basal ganglia, and cerebellum. Brain-derived neurotrophic factor (BDNF) and physical activity appear to play a beneficial role in the process.
Neuroscientists currently distinguish several types of learning and memory that are implemented by the brain in distinct ways:
The field of neuroscience encompasses all approaches that seek to understand the brain and the rest of the nervous system. Psychology seeks to understand mind and behavior, and neurology is the medical discipline that diagnoses and treats diseases of the nervous system. The brain is also the most important organ studied in psychiatry, the branch of medicine that works to study, prevent, and treat mental disorders. Cognitive science seeks to unify neuroscience and psychology with other fields that concern themselves with the brain, such as computer science (artificial intelligence and similar fields) and philosophy.
The oldest method of studying the brain is anatomical, and until the middle of the 20th century, much of the progress in neuroscience came from the development of better cell stains and better microscopes. Neuroanatomists study the large-scale structure of the brain as well as the microscopic structure of neurons and their components, especially synapses. Among other tools, they employ a plethora of stains that reveal neural structure, chemistry, and connectivity. In recent years, the development of immunostaining techniques has allowed investigation of neurons that express specific sets of genes. Also, functional neuroanatomy uses medical imaging techniques to correlate variations in human brain structure with differences in cognition or behavior.
Neurophysiologists study the chemical, pharmacological, and electrical properties of the brain: their primary tools are drugs and recording devices. Thousands of experimentally developed drugs affect the nervous system, some in highly specific ways. Recordings of brain activity can be made using electrodes, either glued to the scalp as in EEG studies, or implanted inside the brains of animals for extracellular recordings, which can detect action potentials generated by individual neurons. Because the brain does not contain pain receptors, it is possible using these techniques to record brain activity from animals that are awake and behaving without causing distress. The same techniques have occasionally been used to study brain activity in human patients with intractable epilepsy, in cases where there was a medical necessity to implant electrodes to localize the brain area responsible for epileptic seizures. Functional imaging techniques such as fMRI are also used to study brain activity; these techniques have mainly been used with human subjects, because they require a conscious subject to remain motionless for long periods of time, but they have the great advantage of being noninvasive.
Another approach to brain function is to examine the consequences of damage to specific brain areas. Even though it is protected by the skull and meninges, surrounded by cerebrospinal fluid, and isolated from the bloodstream by the blood–brain barrier, the delicate nature of the brain makes it vulnerable to numerous diseases and several types of damage. In humans, the effects of strokes and other types of brain damage have been a key source of information about brain function. Because there is no ability to experimentally control the nature of the damage, however, this information is often difficult to interpret. In animal studies, most commonly involving rats, it is possible to use electrodes or locally injected chemicals to produce precise patterns of damage and then examine the consequences for behavior.
Computational neuroscience encompasses two approaches: first, the use of computers to study the brain; second, the study of how brains perform computation. On one hand, it is possible to write a computer program to simulate the operation of a group of neurons by making use of systems of equations that describe their electrochemical activity; such simulations are known as biologically realistic neural networks. On the other hand, it is possible to study algorithms for neural computation by simulating, or mathematically analyzing, the operations of simplified "units" that have some of the properties of neurons but abstract out much of their biological complexity. The computational functions of the brain are studied both by computer scientists and neuroscientists.
Computational neurogenetic modeling is concerned with the study and development of dynamic neuronal models for modeling brain functions with respect to genes and dynamic interactions between genes.
Recent years have seen increasing applications of genetic and genomic techniques to the study of the brain and a focus on the roles of neurotrophic factors and physical activity in neuroplasticity. The most common subjects are mice, because of the availability of technical tools. It is now possible with relative ease to "knock out" or mutate a wide variety of genes, and then examine the effects on brain function. More sophisticated approaches are also being used: for example, using Cre-Lox recombination it is possible to activate or deactivate genes in specific parts of the brain, at specific times.
The oldest brain to have been discovered was in Armenia in the Areni-1 cave complex. The brain, estimated to be over 5,000 years old, was found in the skull of a 12 to 14-year-old girl. Although the brains were shriveled, they were well preserved due to the climate found inside the cave.
Early philosophers were divided as to whether the seat of the soul lies in the brain or heart. Aristotle favored the heart, and thought that the function of the brain was merely to cool the blood. Democritus, the inventor of the atomic theory of matter, argued for a three-part soul, with intellect in the head, emotion in the heart, and lust near the liver. The unknown author of On the Sacred Disease, a medical treatise in the Hippocratic Corpus, came down unequivocally in favor of the brain, writing:
The Roman physician Galen also argued for the importance of the brain, and theorized in some depth about how it might work. Galen traced out the anatomical relationships among brain, nerves, and muscles, demonstrating that all muscles in the body are connected to the brain through a branching network of nerves. He postulated that nerves activate muscles mechanically by carrying a mysterious substance he called pneumata psychikon, usually translated as "animal spirits". Galen's ideas were widely known during the Middle Ages, but not much further progress came until the Renaissance, when detailed anatomical study resumed, combined with the theoretical speculations of René Descartes and those who followed him. Descartes, like Galen, thought of the nervous system in hydraulic terms. He believed that the highest cognitive functions are carried out by a non-physical res cogitans, but that the majority of behaviors of humans, and all behaviors of animals, could be explained mechanistically.
The first real progress toward a modern understanding of nervous function, though, came from the investigations of Luigi Galvani (1737–1798), who discovered that a shock of static electricity applied to an exposed nerve of a dead frog could cause its leg to contract. Since that time, each major advance in understanding has followed more or less directly from the development of a new technique of investigation. Until the early years of the 20th century, the most important advances were derived from new methods for staining cells. Particularly critical was the invention of the Golgi stain, which (when correctly used) stains only a small fraction of neurons, but stains them in their entirety, including cell body, dendrites, and axon. Without such a stain, brain tissue under a microscope appears as an impenetrable tangle of protoplasmic fibers, in which it is impossible to determine any structure. In the hands of Camillo Golgi, and especially of the Spanish neuroanatomist Santiago Ramón y Cajal, the new stain revealed hundreds of distinct types of neurons, each with its own unique dendritic structure and pattern of connectivity.
In the first half of the 20th century, advances in electronics enabled investigation of the electrical properties of nerve cells, culminating in work by Alan Hodgkin, Andrew Huxley, and others on the biophysics of the action potential, and the work of Bernard Katz and others on the electrochemistry of the synapse. These studies complemented the anatomical picture with a conception of the brain as a dynamic entity. Reflecting the new understanding, in 1942 Charles Sherrington visualized the workings of the brain waking from sleep:
The invention of electronic computers in the 1940s, along with the development of mathematical information theory, led to a realization that brains can potentially be understood as information processing systems. This concept formed the basis of the field of cybernetics, and eventually gave rise to the field now known as computational neuroscience. The earliest attempts at cybernetics were somewhat crude in that they treated the brain as essentially a digital computer in disguise, as for example in John von Neumann's 1958 book, The Computer and the Brain. Over the years, though, accumulating information about the electrical responses of brain cells recorded from behaving animals has steadily moved theoretical concepts in the direction of increasing realism.
One of the most influential early contributions was a 1959 paper titled What the frog's eye tells the frog's brain: the paper examined the visual responses of neurons in the retina and optic tectum of frogs, and came to the conclusion that some neurons in the tectum of the frog are wired to combine elementary responses in a way that makes them function as "bug perceivers". A few years later David Hubel and Torsten Wiesel discovered cells in the primary visual cortex of monkeys that become active when sharp edges move across specific points in the field of view—a discovery for which they won a Nobel Prize. Follow-up studies in higher-order visual areas found cells that detect binocular disparity, color, movement, and aspects of shape, with areas located at increasing distances from the primary visual cortex showing increasingly complex responses. Other investigations of brain areas unrelated to vision have revealed cells with a wide variety of response correlates, some related to memory, some to abstract types of cognition such as space.
Theorists have worked to understand these response patterns by constructing mathematical models of neurons and neural networks, which can be simulated using computers. Some useful models are abstract, focusing on the conceptual structure of neural algorithms rather than the details of how they are implemented in the brain; other models attempt to incorporate data about the biophysical properties of real neurons. No model on any level is yet considered to be a fully valid description of brain function, though. The essential difficulty is that sophisticated computation by neural networks requires distributed processing in which hundreds or thousands of neurons work cooperatively—current methods of brain activity recording are only capable of isolating action potentials from a few dozen neurons at a time.
Furthermore, even single neurons appear to be complex and capable of performing computations. So, brain models that do not reflect this are too abstract to be representative of brain operation; models that do try to capture this are very computationally expensive and arguably intractable with present computational resources. However, the Human Brain Project is trying to build a realistic, detailed computational model of the entire human brain. The wisdom of this approach has been publicly contested, with high-profile scientists on both sides of the argument.
In the second half of the 20th century, developments in chemistry, electron microscopy, genetics, computer science, functional brain imaging, and other fields progressively opened new windows into brain structure and function. In the United States, the 1990s were officially designated as the "Decade of the Brain" to commemorate advances made in brain research, and to promote funding for such research.
In the 21st century, these trends have continued, and several new approaches have come into prominence, including multielectrode recording, which allows the activity of many brain cells to be recorded all at the same time; genetic engineering, which allows molecular components of the brain to be altered experimentally; genomics, which allows variations in brain structure to be correlated with variations in DNA properties and neuroimaging.
Animal brains are used as food in numerous cuisines.
Some archaeological evidence suggests that the mourning rituals of European Neanderthals also involved the consumption of the brain.
The Fore people of Papua New Guinea are known to eat human brains. In funerary rituals, those close to the dead would eat the brain of the deceased to create a sense of immortality. A prion disease called kuru has been traced to this.
Anatomy
Cross section of the olfactory bulb of a rat, stained in two different ways at the same time: one stain shows neuron cell bodies, the other shows receptors for the neurotransmitter GABA.
The shape and size of the brain varies greatly between species, and identifying common features is often difficult. Nevertheless, there are a number of principles of brain architecture that apply across a wide range of species. Some aspects of brain structure are common to almost the entire range of animal species; others distinguish "advanced" brains from more primitive ones, or distinguish vertebrates from invertebrates.
The simplest way to gain information about brain anatomy is by visual inspection, but many more sophisticated techniques have been developed. Brain tissue in its natural state is too soft to work with, but it can be hardened by immersion in alcohol or other fixatives, and then sliced apart for examination of the interior. Visually, the interior of the brain consists of areas of so-called grey matter, with a dark color, separated by areas of white matter, with a lighter color. Further information can be gained by staining slices of brain tissue with a variety of chemicals that bring out areas where specific types of molecules are present in high concentrations. It is also possible to examine the microstructure of brain tissue using a microscope, and to trace the pattern of connections from one brain area to another.
Cellular structure
Neurons generate electrical signals that travel along their axons. When a pulse of electricity reaches a junction called a synapse, it causes a neurotransmitter chemical to be released, which binds to receptors on other cells and thereby alters their electrical activity.
The brains of all species are composed primarily of two broad classes of cells: neurons and glial cells. Glial cells (also known as glia or neuroglia) come in several types, and perform a number of critical functions, including structural support, metabolic support, insulation, and guidance of development. Neurons, however, are usually considered the most important cells in the brain.
The property that makes neurons unique is their ability to send signals to specific target cells over long distances. They send these signals by means of an axon, which is a thin protoplasmic fiber that extends from the cell body and projects, usually with numerous branches, to other areas, sometimes nearby, sometimes in distant parts of the brain or body. The length of an axon can be extraordinary: for example, if a pyramidal cell (an excitatory neuron) of the cerebral cortex were magnified so that its cell body became the size of a human body, its axon, equally magnified, would become a cable a few centimeters in diameter, extending more than a kilometer. These axons transmit signals in the form of electrochemical pulses called action potentials, which last less than a thousandth of a second and travel along the axon at speeds of 1–100 meters per second. Some neurons emit action potentials constantly, at rates of 10–100 per second, usually in irregular patterns; other neurons are quiet most of the time, but occasionally emit a burst of action potentials.
Axons transmit signals to other neurons by means of specialized junctions called synapses. A single axon may make as many as several thousand synaptic connections with other cells. When an action potential, traveling along an axon, arrives at a synapse, it causes a chemical called a neurotransmitter to be released. The neurotransmitter binds to receptor molecules in the membrane of the target cell.
Synapses are the key functional elements of the brain. The essential function of the brain is cell-to-cell communication, and synapses are the points at which communication occurs. The human brain has been estimated to contain approximately 100 trillion synapses; even the brain of a fruit fly contains several million. The functions of these synapses are very diverse: some are excitatory (exciting the target cell); others are inhibitory; others work by activating second messenger systems that change the internal chemistry of their target cells in complex ways. A large number of synapses are dynamically modifiable; that is, they are capable of changing strength in a way that is controlled by the patterns of signals that pass through them. It is widely believed that activity-dependent modification of synapses is the brain's primary mechanism for learning and memory.
Most of the space in the brain is taken up by axons, which are often bundled together in what are called nerve fiber tracts. A myelinated axon is wrapped in a fatty insulating sheath of myelin, which serves to greatly increase the speed of signal propagation. (There are also unmyelinated axons). Myelin is white, making parts of the brain filled exclusively with nerve fibers appear as light-colored white matter, in contrast to the darker-colored grey matter that marks areas with high densities of neuron cell bodies.
Evolution
Main article: Evolution of the brain
Generic bilaterian nervous system
Nervous system of a generic bilaterian animal, in the form of a nerve cord with segmental enlargements, and a "brain" at the front
Except for a few primitive organisms such as sponges (which have no nervous system) and cnidarians (which have a diffuse nervous system consisting of a nerve net), all living multicellular animals are bilaterians, meaning animals with a bilaterally symmetric body plan (that is, left and right sides that are approximate mirror images of each other). All bilaterians are thought to have descended from a common ancestor that appeared late in the Cryogenian period, 700–650 million years ago, and it has been hypothesized that this common ancestor had the shape of a simple tubeworm with a segmented body. At a schematic level, that basic worm-shape continues to be reflected in the body and nervous system architecture of all modern bilaterians, including vertebrates. The fundamental bilateral body form is a tube with a hollow gut cavity running from the mouth to the anus, and a nerve cord with an enlargement (a ganglion) for each body segment, with an especially large ganglion at the front, called the brain. The brain is small and simple in some species, such as nematode worms; in other species, such as vertebrates, it is a large and very complex organ. Some types of worms, such as leeches, also have an enlarged ganglion at the back end of the nerve cord, known as a "tail brain".
There are a few types of existing bilaterians that lack a recognizable brain, including echinoderms and tunicates. It has not been definitively established whether the existence of these brainless species indicates that the earliest bilaterians lacked a brain, or whether their ancestors evolved in a way that led to the disappearance of a previously existing brain structure.
Invertebrates
Fruit flies (Drosophila) have been extensively studied to gain insight into the role of genes in brain development.
This category includes tardigrades, arthropods, molluscs, and numerous types of worms. The diversity of invertebrate body plans is matched by an equal diversity in brain structures.
Two groups of invertebrates have notably complex brains: arthropods (insects, crustaceans, arachnids, and others), and cephalopods (octopuses, squids, and similar molluscs). The brains of arthropods and cephalopods arise from twin parallel nerve cords that extend through the body of the animal. Arthropods have a central brain, the supraesophageal ganglion, with three divisions and large optical lobes behind each eye for visual processing. Cephalopods such as the octopus and squid have the largest brains of any invertebrates.
There are several invertebrate species whose brains have been studied intensively because they have properties that make them convenient for experimental work:
Fruit flies (Drosophila), because of the large array of techniques available for studying their genetics, have been a natural subject for studying the role of genes in brain development. In spite of the large evolutionary distance between insects and mammals, many aspects of Drosophila neurogenetics have been shown to be relevant to humans. The first biological clock genes, for example, were identified by examining Drosophila mutants that showed disrupted daily activity cycles. A search in the genomes of vertebrates revealed a set of analogous genes, which were found to play similar roles in the mouse biological clock—and therefore almost certainly in the human biological clock as well. Studies done on Drosophila, also show that most neuropil regions of the brain are continuously reorganized throughout life in response to specific living conditions.
The nematode worm Caenorhabditis elegans, like Drosophila, has been studied largely because of its importance in genetics. In the early 1970s, Sydney Brenner chose it as a model organism for studying the way that genes control development. One of the advantages of working with this worm is that the body plan is very stereotyped: the nervous system of the hermaphrodite contains exactly 302 neurons, always in the same places, making identical synaptic connections in every worm. Brenner's team sliced worms into thousands of ultrathin sections and photographed each one under an electron microscope, then visually matched fibers from section to section, to map out every neuron and synapse in the entire body. The complete neuronal wiring diagram of C.elegans – its connectome was achieved. Nothing approaching this level of detail is available for any other organism, and the information gained has enabled a multitude of studies that would otherwise have not been possible.
The sea slug Aplysia californica was chosen by Nobel Prize-winning neurophysiologist Eric Kandel as a model for studying the cellular basis of learning and memory, because of the simplicity and accessibility of its nervous system, and it has been examined in hundreds of experiments.
Vertebrates
The brain of a shark
The first vertebrates appeared over 500 million years ago (Mya), during the Cambrian period, and may have resembled the modern hagfish in form. Jawed fish appeared by 445 Mya, amphibians by 350 Mya, reptiles by 310 Mya and mammals by 200 Mya (approximately). Each species has an equally long evolutionary history, but the brains of modern hagfishes, lampreys, sharks, amphibians, reptiles, and mammals show a gradient of size and complexity that roughly follows the evolutionary sequence. All of these brains contain the same set of basic anatomical components, but many are rudimentary in the hagfish, whereas in mammals the foremost part (the telencephalon) is greatly elaborated and expanded.
Brains are most commonly compared in terms of their size. The relationship between brain size, body size and other variables has been studied across a wide range of vertebrate species. As a rule, brain size increases with body size, but not in a simple linear proportion. In general, smaller animals tend to have larger brains, measured as a fraction of body size. For mammals, the relationship between brain volume and body mass essentially follows a power law with an exponent of about 0.75. This formula describes the central tendency, but every family of mammals departs from it to some degree, in a way that reflects in part the complexity of their behavior. For example, primates have brains 5 to 10 times larger than the formula predicts. Predators tend to have larger brains than their prey, relative to body size.
The main subdivisions of the embryonic vertebrate brain (left), which later differentiate into structures of the adult brain (right)
All vertebrate brains share a common underlying form, which appears most clearly during early stages of embryonic development. In its earliest form, the brain appears as three swellings at the front end of the neural tube; these swellings eventually become the forebrain, midbrain, and hindbrain (the prosencephalon, mesencephalon, and rhombencephalon, respectively). At the earliest stages of brain development, the three areas are roughly equal in size. In many classes of vertebrates, such as fish and amphibians, the three parts remain similar in size in the adult, but in mammals the forebrain becomes much larger than the other parts, and the midbrain becomes very small.
The brains of vertebrates are made of very soft tissue. Living brain tissue is pinkish on the outside and mostly white on the inside, with subtle variations in color. Vertebrate brains are surrounded by a system of connective tissue membranes called meninges that separate the skull from the brain. Blood vessels enter the central nervous system through holes in the meningeal layers. The cells in the blood vessel walls are joined tightly to one another, forming the blood–brain barrier, which blocks the passage of many toxins and pathogens (though at the same time blocking antibodies and some drugs, thereby presenting special challenges in treatment of diseases of the brain).
Neuroanatomists usually divide the vertebrate brain into six main regions: the telencephalon (cerebral hemispheres), diencephalon (thalamus and hypothalamus), mesencephalon (midbrain), cerebellum, pons, and medulla oblongata. Each of these areas has a complex internal structure. Some parts, such as the cerebral cortex and the cerebellar cortex, consist of layers that are folded or convoluted to fit within the available space. Other parts, such as the thalamus and hypothalamus, consist of clusters of many small nuclei. Thousands of distinguishable areas can be identified within the vertebrate brain based on fine distinctions of neural structure, chemistry, and connectivity.
The main anatomical regions of the vertebrate brain, shown for shark and human. The same parts are present, but they differ greatly in size and shape.
Although the same basic components are present in all vertebrate brains, some branches of vertebrate evolution have led to substantial distortions of brain geometry, especially in the forebrain area. The brain of a shark shows the basic components in a straightforward way, but in teleost fishes (the great majority of existing fish species), the forebrain has become "everted", like a sock turned inside out. In birds, there are also major changes in forebrain structure. These distortions can make it difficult to match brain components from one species with those of another species.
Here is a list of some of the most important vertebrate brain components, along with a brief description of their functions as currently understood:
See also: List of regions in the human brain
The medulla, along with the spinal cord, contains many small nuclei involved in a wide variety of sensory and involuntary motor functions such as vomiting, heart rate and digestive processes.
The pons lies in the brainstem directly above the medulla. Among other things, it contains nuclei that control often voluntary but simple acts such as sleep, respiration, swallowing, bladder function, equilibrium, eye movement, facial expressions, and posture.
The hypothalamus is a small region at the base of the forebrain, whose complexity and importance belies its size. It is composed of numerous small nuclei, each with distinct connections and neurochemistry. The hypothalamus is engaged in additional involuntary or partially voluntary acts such as sleep and wake cycles, eating and drinking, and the release of some hormones.
The thalamus is a collection of nuclei with diverse functions: some are involved in relaying information to and from the cerebral hemispheres, while others are involved in motivation. The subthalamic area (zona incerta) seems to contain action-generating systems for several types of "consummatory" behaviors such as eating, drinking, defecation, and copulation.
The cerebellum modulates the outputs of other brain systems, whether motor-related or thought related, to make them certain and precise. Removal of the cerebellum does not prevent an animal from doing anything in particular, but it makes actions hesitant and clumsy. This precision is not built-in but learned by trial and error. The muscle coordination learned while riding a bicycle is an example of a type of neural plasticity that may take place largely within the cerebellum. 10% of the brain's total volume consists of the cerebellum and 50% of all neurons are held within its structure.
The optic tectum allows actions to be directed toward points in space, most commonly in response to visual input. In mammals, it is usually referred to as the superior colliculus, and its best-studied function is to direct eye movements. It also directs reaching movements and other object-directed actions. It receives strong visual inputs, but also inputs from other senses that are useful in directing actions, such as auditory input in owls and input from the thermosensitive pit organs in snakes. In some primitive fishes, such as lampreys, this region is the largest part of the brain. The superior colliculus is part of the midbrain.
The pallium is a layer of grey matter that lies on the surface of the forebrain and is the most complex and most recent evolutionary development of the brain as an organ. In reptiles and mammals, it is called the cerebral cortex. Multiple functions involve the pallium, including smell and spatial memory. In mammals, where it becomes so large as to dominate the brain, it takes over functions from many other brain areas. In many mammals, the cerebral cortex consists of folded bulges called gyri that create deep furrows or fissures called sulci. The folds increase the surface area of the cortex and therefore increase the amount of gray matter and the amount of information that can be stored and processed.
The hippocampus, strictly speaking, is found only in mammals. However, the area it derives from, the medial pallium, has counterparts in all vertebrates. There is evidence that this part of the brain is involved in complex events such as spatial memory and navigation in fishes, birds, reptiles, and mammals.
The basal ganglia are a group of interconnected structures in the forebrain. The primary function of the basal ganglia appears to be action selection: they send inhibitory signals to all parts of the brain that can generate motor behaviors, and in the right circumstances can release the inhibition, so that the action-generating systems are able to execute their actions. Reward and punishment exert their most important neural effects by altering connections within the basal ganglia.
The olfactory bulb is a special structure that processes olfactory sensory signals and sends its output to the olfactory part of the pallium. It is a major brain component in many vertebrates, but is greatly reduced in humans and other primates (whose senses are dominated by information acquired by sight rather than smell).
Birds
Main article: Avian brain
This section is an excerpt from Avian brain.[edit]
Brains of an emu, a kiwi, a barn owl, and a pigeon, with visual processing areas labelled
The avian brain is the central organ of the nervous system in birds. Birds possess large, complex brains, which process, integrate, and coordinate information received from the environment and make decisions on how to respond with the rest of the body. Like in all chordates, the avian brain is contained within the skull bones of the head.
The bird brain is divided into a number of sections, each with a different function. The cerebrum or telencephalon is divided into two hemispheres, and controls higher functions. The telencephalon is dominated by a large pallium, which corresponds to the mammalian cerebral cortex and is responsible for the cognitive functions of birds. The pallium is made up of several major structures: the hyperpallium, a dorsal bulge of the pallium found only in birds, as well as the nidopallium, mesopallium, and archipallium. The bird telencephalon nuclear structure, wherein neurons are distributed in three-dimensionally arranged clusters, with no large-scale separation of white matter and grey matter, though there exist layer-like and column-like connections. Structures in the pallium are associated with perception, learning, and cognition. Beneath the pallium are the two components of the subpallium, the striatum and pallidum. The subpallium connects different parts of the telencephalon and plays major roles in a number of critical behaviours. To the rear of the telencephalon are the thalamus, midbrain, and cerebellum. The hindbrain connects the rest of the brain to the spinal cord.
The size and structure of the avian brain enables prominent behaviours of birds such as flight and vocalization. Dedicated structures and pathways integrate the auditory and visual senses, strong in most species of birds, as well as the typically weaker olfactory and tactile senses. Social behaviour, widespread among birds, depends on the organisation and functions of the brain. Some birds exhibit strong abilities of cognition, enabled by the unique structure and physiology of the avian brain.
Mammals
The most obvious difference between the brains of mammals and other vertebrates is in terms of size. On average, a mammal has a brain roughly twice as large as that of a bird of the same body size, and ten times as large as that of a reptile of the same body size.
Size, however, is not the only difference: there are also substantial differences in shape. The hindbrain and midbrain of mammals are generally similar to those of other vertebrates, but dramatic differences appear in the forebrain, which is greatly enlarged and also altered in structure. The cerebral cortex is the part of the brain that most strongly distinguishes mammals. In non-mammalian vertebrates, the surface of the cerebrum is lined with a comparatively simple three-layered structure called the pallium. In mammals, the pallium evolves into a complex six-layered structure called neocortex or isocortex. Several areas at the edge of the neocortex, including the hippocampus and amygdala, are also much more extensively developed in mammals than in other vertebrates.
The elaboration of the cerebral cortex carries with it changes to other brain areas. The superior colliculus, which plays a major role in visual control of behavior in most vertebrates, shrinks to a small size in mammals, and many of its functions are taken over by visual areas of the cerebral cortex. The cerebellum of mammals contains a large portion (the neocerebellum) dedicated to supporting the cerebral cortex, which has no counterpart in other vertebrates.
Primates
See also: Human brain
Encephalization Quotient
Species
EQ
Human
7.4–7.8
Common chimpanzee
2.2–2.5
Rhesus monkey
2.1
Bottlenose dolphin
4.14
Elephant
1.13–2.36
Dog
1.2
Horse
0.9
Rat
0.4
The brains of humans and other primates contain the same structures as the brains of other mammals, but are generally larger in proportion to body size. The encephalization quotient (EQ) is used to compare brain sizes across species. It takes into account the nonlinearity of the brain-to-body relationship. Humans have an average EQ in the 7-to-8 range, while most other primates have an EQ in the 2-to-3 range. Dolphins have values higher than those of primates other than humans, but nearly all other mammals have EQ values that are substantially lower.
Most of the enlargement of the primate brain comes from a massive expansion of the cerebral cortex, especially the prefrontal cortex and the parts of the cortex involved in vision. The visual processing network of primates includes at least 30 distinguishable brain areas, with a complex web of interconnections. It has been estimated that visual processing areas occupy more than half of the total surface of the primate neocortex. The prefrontal cortex carries out functions that include planning, working memory, motivation, attention, and executive control. It takes up a much larger proportion of the brain for primates than for other species, and an especially large fraction of the human brain.
Development
Main article: Neural development
Brain of a human embryo in the sixth week of development
The brain develops in an intricately orchestrated sequence of stages. It changes in shape from a simple swelling at the front of the nerve cord in the earliest embryonic stages, to a complex array of areas and connections. Neurons are created in special zones that contain stem cells, and then migrate through the tissue to reach their ultimate locations. Once neurons have positioned themselves, their axons sprout and navigate through the brain, branching and extending as they go, until the tips reach their targets and form synaptic connections. In a number of parts of the nervous system, neurons and synapses are produced in excessive numbers during the early stages, and then the unneeded ones are pruned away.
For vertebrates, the early stages of neural development are similar across all species. As the embryo transforms from a round blob of cells into a wormlike structure, a narrow strip of ectoderm running along the midline of the back is induced to become the neural plate, the precursor of the nervous system. The neural plate folds inward to form the neural groove, and then the lips that line the groove merge to enclose the neural tube, a hollow cord of cells with a fluid-filled ventricle at the center. At the front end, the ventricles and cord swell to form three vesicles that are the precursors of the prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain). At the next stage, the forebrain splits into two vesicles called the telencephalon (which will contain the cerebral cortex, basal ganglia, and related structures) and the diencephalon (which will contain the thalamus and hypothalamus). At about the same time, the hindbrain splits into the metencephalon (which will contain the cerebellum and pons) and the myelencephalon (which will contain the medulla oblongata). Each of these areas contains proliferative zones where neurons and glial cells are generated; the resulting cells then migrate, sometimes for long distances, to their final positions.
Once a neuron is in place, it extends dendrites and an axon into the area around it. Axons, because they commonly extend a great distance from the cell body and need to reach specific targets, grow in a particularly complex way. The tip of a growing axon consists of a blob of protoplasm called a growth cone, studded with chemical receptors. These receptors sense the local environment, causing the growth cone to be attracted or repelled by various cellular elements, and thus to be pulled in a particular direction at each point along its path. The result of this pathfinding process is that the growth cone navigates through the brain until it reaches its destination area, where other chemical cues cause it to begin generating synapses. Considering the entire brain, thousands of genes create products that influence axonal pathfinding.
The synaptic network that finally emerges is only partly determined by genes, though. In many parts of the brain, axons initially "overgrow", and then are "pruned" by mechanisms that depend on neural activity. In the projection from the eye to the midbrain, for example, the structure in the adult contains a very precise mapping, connecting each point on the surface of the retina to a corresponding point in a midbrain layer. In the first stages of development, each axon from the retina is guided to the right general vicinity in the midbrain by chemical cues, but then branches very profusely and makes initial contact with a wide swath of midbrain neurons. The retina, before birth, contains special mechanisms that cause it to generate waves of activity that originate spontaneously at a random point and then propagate slowly across the retinal layer. These waves are useful because they cause neighboring neurons to be active at the same time; that is, they produce a neural activity pattern that contains information about the spatial arrangement of the neurons. This information is exploited in the midbrain by a mechanism that causes synapses to weaken, and eventually vanish, if activity in an axon is not followed by activity of the target cell. The result of this sophisticated process is a gradual tuning and tightening of the map, leaving it finally in its precise adult form.
Similar things happen in other brain areas: an initial synaptic matrix is generated as a result of genetically determined chemical guidance, but then gradually refined by activity-dependent mechanisms, partly driven by internal dynamics, partly by external sensory inputs. In some cases, as with the retina-midbrain system, activity patterns depend on mechanisms that operate only in the developing brain, and apparently exist solely to guide development.
In humans and many other mammals, new neurons are created mainly before birth, and the infant brain contains substantially more neurons than the adult brain. There are, however, a few areas where new neurons continue to be generated throughout life. The two areas for which adult neurogenesis is well established are the olfactory bulb, which is involved in the sense of smell, and the dentate gyrus of the hippocampus, where there is evidence that the new neurons play a role in storing newly acquired memories. With these exceptions, however, the set of neurons that is present in early childhood is the set that is present for life. Glial cells are different: as with most types of cells in the body, they are generated throughout the lifespan.
There has long been debate about whether the qualities of mind, personality, and intelligence can be attributed to heredity or to upbringing. Although many details remain to be settled, neuroscience shows that both factors are important. Genes determine both the general form of the brain and how it reacts to experience, but experience is required to refine the matrix of synaptic connections, resulting in greatly increased complexity. The presence or absence of experience is critical at key periods of development. Additionally, the quantity and quality of experience are important. For example, animals raised in enriched environments demonstrate thick cerebral cortices, indicating a high density of synaptic connections, compared to animals with restricted levels of stimulation.
Physiology
The functions of the brain depend on the ability of neurons to transmit electrochemical signals to other cells, and their ability to respond appropriately to electrochemical signals received from other cells. The electrical properties of neurons are controlled by a wide variety of biochemical and metabolic processes, most notably the interactions between neurotransmitters and receptors that take place at synapses.
Neurotransmitters and receptors
Neurotransmitters are chemicals that are released at synapses when the local membrane is depolarised and Ca enters into the cell, typically when an action potential arrives at the synapse – neurotransmitters attach themselves to receptor molecules on the membrane of the synapse's target cell (or cells), and thereby alter the electrical or chemical properties of the receptor molecules. With few exceptions, each neuron in the brain releases the same chemical neurotransmitter, or combination of neurotransmitters, at all the synaptic connections it makes with other neurons; this rule is known as Dale's principle. Thus, a neuron can be characterized by the neurotransmitters that it releases. The great majority of psychoactive drugs exert their effects by altering specific neurotransmitter systems. This applies to drugs such as cannabinoids, nicotine, heroin, cocaine, alcohol, fluoxetine, chlorpromazine, and many others.
The two neurotransmitters that are most widely found in the vertebrate brain are glutamate, which almost always exerts excitatory effects on target neurons, and gamma-aminobutyric acid (GABA), which is almost always inhibitory. Neurons using these transmitters can be found in nearly every part of the brain. Because of their ubiquity, drugs that act on glutamate or GABA tend to have broad and powerful effects. Some general anesthetics act by reducing the effects of glutamate; most tranquilizers exert their sedative effects by enhancing the effects of GABA.
There are dozens of other chemical neurotransmitters that are used in more limited areas of the brain, often areas dedicated to a particular function. Serotonin, for example—the primary target of many antidepressant drugs and many dietary aids—comes exclusively from a small brainstem area called the raphe nuclei. Norepinephrine, which is involved in arousal, comes exclusively from a nearby small area called the locus coeruleus. Other neurotransmitters such as acetylcholine and dopamine have multiple sources in the brain but are not as ubiquitously distributed as glutamate and GABA.
Electrical activity
Brain electrical activity recorded from a human patient during an epileptic seizure
As a side effect of the electrochemical processes used by neurons for signaling, brain tissue generates electric fields when it is active. When large numbers of neurons show synchronized activity, the electric fields that they generate can be large enough to detect outside the skull, using electroencephalography (EEG) or magnetoencephalography (MEG). EEG recordings, along with recordings made from electrodes implanted inside the brains of animals such as rats, show that the brain of a living animal is constantly active, even during sleep. Each part of the brain shows a mixture of rhythmic and nonrhythmic activity, which may vary according to behavioral state. In mammals, the cerebral cortex tends to show large slow delta waves during sleep, faster alpha waves when the animal is awake but inattentive, and chaotic-looking irregular activity when the animal is actively engaged in a task, called beta and gamma waves. During an epileptic seizure, the brain's inhibitory control mechanisms fail to function and electrical activity rises to pathological levels, producing EEG traces that show large wave and spike patterns not seen in a healthy brain. Relating these population-level patterns to the computational functions of individual neurons is a major focus of current research in neurophysiology.
Metabolism
All vertebrates have a blood–brain barrier that allows metabolism inside the brain to operate differently from metabolism in other parts of the body. The neurovascular unit regulates cerebral blood flow so that activated neurons can be supplied with energy. Glial cells play a major role in brain metabolism by controlling the chemical composition of the fluid that surrounds neurons, including levels of ions and nutrients.
Brain tissue consumes a large amount of energy in proportion to its volume, so large brains place severe metabolic demands on animals. The need to limit body weight in order, for example, to fly, has apparently led to selection for a reduction of brain size in some species, such as bats. Most of the brain's energy consumption goes into sustaining the electric charge (membrane potential) of neurons. Most vertebrate species devote between 2% and 8% of basal metabolism to the brain. In primates, however, the percentage is much higher—in humans it rises to 20–25%. The energy consumption of the brain does not vary greatly over time, but active regions of the cerebral cortex consume somewhat more energy than inactive regions; this forms the basis for the functional brain imaging methods of PET, fMRI, and NIRS. The brain typically gets most of its energy from oxygen-dependent metabolism of glucose (i.e., blood sugar), but ketones provide a major alternative source, together with contributions from medium chain fatty acids (caprylic and heptanoic acids), lactate, acetate, and possibly amino acids.
Function
Model of a neural circuit in the cerebellum, as proposed by James S. Albus
Information from the sense organs is collected in the brain. There it is used to determine what actions the organism is to take. The brain processes the raw data to extract information about the structure of the environment. Next it combines the processed information with information about the current needs of the animal and with memory of past circumstances. Finally, on the basis of the results, it generates motor response patterns. These signal-processing tasks require intricate interplay between a variety of functional subsystems.
The function of the brain is to provide coherent control over the actions of an animal. A centralized brain allows groups of muscles to be co-activated in complex patterns; it also allows stimuli impinging on one part of the body to evoke responses in other parts, and it can prevent different parts of the body from acting at cross-purposes to each other.
Perception
Diagram of signal processing in the auditory system
The human brain is provided with information about light, sound, the chemical composition of the atmosphere, temperature, the position of the body in space (proprioception), the chemical composition of the bloodstream, and more. In other animals additional senses are present, such as the infrared heat-sense of snakes, the magnetic field sense of some birds, or the electric field sense mainly seen in aquatic animals.
Each sensory system begins with specialized receptor cells, such as photoreceptor cells in the retina of the eye, or vibration-sensitive hair cells in the cochlea of the ear. The axons of sensory receptor cells travel into the spinal cord or brain, where they transmit their signals to a first-order sensory nucleus dedicated to one specific sensory modality. This primary sensory nucleus sends information to higher-order sensory areas that are dedicated to the same modality. Eventually, via a way-station in the thalamus, the signals are sent to the cerebral cortex, where they are processed to extract the relevant features, and integrated with signals coming from other sensory systems.
Motor control
Motor systems are areas of the brain that are involved in initiating body movements, that is, in activating muscles. Except for the muscles that control the eye, which are driven by nuclei in the midbrain, all the voluntary muscles in the body are directly innervated by motor neurons in the spinal cord and hindbrain. Spinal motor neurons are controlled both by neural circuits intrinsic to the spinal cord, and by inputs that descend from the brain. The intrinsic spinal circuits implement many reflex responses, and contain pattern generators for rhythmic movements such as walking or swimming. The descending connections from the brain allow for more sophisticated control.
The brain contains several motor areas that project directly to the spinal cord. At the lowest level are motor areas in the medulla and pons, which control stereotyped movements such as walking, breathing, or swallowing. At a higher level are areas in the midbrain, such as the red nucleus, which is responsible for coordinating movements of the arms and legs. At a higher level yet is the primary motor cortex, a strip of tissue located at the posterior edge of the frontal lobe. The primary motor cortex sends projections to the subcortical motor areas, but also sends a massive projection directly to the spinal cord, through the pyramidal tract. This direct corticospinal projection allows for precise voluntary control of the fine details of movements. Other motor-related brain areas exert secondary effects by projecting to the primary motor areas. Among the most important secondary areas are the premotor cortex, supplementary motor area, basal ganglia, and cerebellum. In addition to all of the above, the brain and spinal cord contain extensive circuitry to control the autonomic nervous system which controls the movement of the smooth muscle of the body.
Major areas involved in controlling movement
Area
Location
Function
Ventral horn
Spinal cord
Contains motor neurons that directly activate muscles
Oculomotor nuclei
Midbrain
Contains motor neurons that directly activate the eye muscles
Cerebellum
Hindbrain
Calibrates precision and timing of movements
Basal ganglia
Forebrain
Action selection on the basis of motivation
Motor cortex
Frontal lobe
Direct cortical activation of spinal motor circuits
Premotor cortex
Frontal lobe
Groups elementary movements into coordinated patterns
Supplementary motor area
Frontal lobe
Sequences movements into temporal patterns
Prefrontal cortex
Frontal lobe
Planning and other executive functions
Sleep
Main article: Sleep
See also: Arousal
Many animals alternate between sleeping and waking in a daily cycle. Arousal and alertness are also modulated on a finer time scale by a network of brain areas. A key component of the sleep system is the suprachiasmatic nucleus (SCN), a tiny part of the hypothalamus located directly above the point at which the optic nerves from the two eyes cross. The SCN contains the body's central biological clock. Neurons there show activity levels that rise and fall with a period of about 24 hours, circadian rhythms: these activity fluctuations are driven by rhythmic changes in expression of a set of "clock genes". The SCN continues to keep time even if it is excised from the brain and placed in a dish of warm nutrient solution, but it ordinarily receives input from the optic nerves, through the retinohypothalamic tract (RHT), that allows daily light-dark cycles to calibrate the clock.
The SCN projects to a set of areas in the hypothalamus, brainstem, and midbrain that are involved in implementing sleep-wake cycles. An important component of the system is the reticular formation, a group of neuron-clusters scattered diffusely through the core of the lower brain. Reticular neurons send signals to the thalamus, which in turn sends activity-level-controlling signals to every part of the cortex. Damage to the reticular formation can produce a permanent state of coma.
Sleep involves great changes in brain activity. Until the 1950s it was generally believed that the brain essentially shuts off during sleep, but this is now known to be far from true; activity continues, but patterns become very different. There are two types of sleep: REM sleep (with dreaming) and NREM (non-REM, usually without dreaming) sleep, which repeat in slightly varying patterns throughout a sleep episode. Three broad types of distinct brain activity patterns can be measured: REM, light NREM and deep NREM. During deep NREM sleep, also called slow wave sleep, activity in the cortex takes the form of large synchronized waves, whereas in the waking state it is noisy and desynchronized. Levels of the neurotransmitters norepinephrine and serotonin drop during slow wave sleep, and fall almost to zero during REM sleep; levels of acetylcholine show the reverse pattern.
Homeostasis
Cross-section of a human head, showing location of the hypothalamus
For any animal, survival requires maintaining a variety of parameters of bodily state within a limited range of variation: these include temperature, water content, salt concentration in the bloodstream, blood glucose levels, blood oxygen level, and others. The ability of an animal to regulate the internal environment of its body—the milieu intérieur, as the pioneering physiologist Claude Bernard called it—is known as homeostasis (Greek for "standing still"). Maintaining homeostasis is a crucial function of the brain. The basic principle that underlies homeostasis is negative feedback: any time a parameter diverges from its set-point, sensors generate an error signal that evokes a response that causes the parameter to shift back toward its optimum value. (This principle is widely used in engineering, for example in the control of temperature using a thermostat.)
In vertebrates, the part of the brain that plays the greatest role is the hypothalamus, a small region at the base of the forebrain whose size does not reflect its complexity or the importance of its function. The hypothalamus is a collection of small nuclei, most of which are involved in basic biological functions. Some of these functions relate to arousal or to social interactions such as sexuality, aggression, or maternal behaviors; but many of them relate to homeostasis. Several hypothalamic nuclei receive input from sensors located in the lining of blood vessels, conveying information about temperature, sodium level, glucose level, blood oxygen level, and other parameters. These hypothalamic nuclei send output signals to motor areas that can generate actions to rectify deficiencies. Some of the outputs also go to the pituitary gland, a tiny gland attached to the brain directly underneath the hypothalamus. The pituitary gland secretes hormones into the bloodstream, where they circulate throughout the body and induce changes in cellular activity.
Motivation
Components of the basal ganglia, shown in two cross-sections of the human brain. Blue: caudate nucleus and putamen. Green: globus pallidus. Red: subthalamic nucleus. Black: substantia nigra.
The individual animals need to express survival-promoting behaviors, such as seeking food, water, shelter, and a mate. The motivational system in the brain monitors the current state of satisfaction of these goals, and activates behaviors to meet any needs that arise. The motivational system works largely by a reward–punishment mechanism. When a particular behavior is followed by favorable consequences, the reward mechanism in the brain is activated, which induces structural changes inside the brain that cause the same behavior to be repeated later, whenever a similar situation arises. Conversely, when a behavior is followed by unfavorable consequences, the brain's punishment mechanism is activated, inducing structural changes that cause the behavior to be suppressed when similar situations arise in the future.
Most organisms studied to date use a reward–punishment mechanism: for instance, worms and insects can alter their behavior to seek food sources or to avoid dangers. In vertebrates, the reward-punishment system is implemented by a specific set of brain structures, at the heart of which lie the basal ganglia, a set of interconnected areas at the base of the forebrain. The basal ganglia are the central site at which decisions are made: the basal ganglia exert a sustained inhibitory control over most of the motor systems in the brain; when this inhibition is released, a motor system is permitted to execute the action it is programmed to carry out. Rewards and punishments function by altering the relationship between the inputs that the basal ganglia receive and the decision-signals that are emitted. The reward mechanism is better understood than the punishment mechanism, because its role in drug abuse has caused it to be studied very intensively. Research has shown that the neurotransmitter dopamine plays a central role: addictive drugs such as cocaine, amphetamine, and nicotine either cause dopamine levels to rise or cause the effects of dopamine inside the brain to be enhanced.
Learning and memory
Almost all animals are capable of modifying their behavior as a result of experience—even the most primitive types of worms. Because behavior is driven by brain activity, changes in behavior must somehow correspond to changes inside the brain. Already in the late 19th century theorists like Santiago Ramón y Cajal argued that the most plausible explanation is that learning and memory are expressed as changes in the synaptic connections between neurons. Until 1970, however, experimental evidence to support the synaptic plasticity hypothesis was lacking. In 1971 Tim Bliss and Terje Lømo published a paper on a phenomenon now called long-term potentiation: the paper showed clear evidence of activity-induced synaptic changes that lasted for at least several days. Since then technical advances have made these sorts of experiments much easier to carry out, and thousands of studies have been made that have clarified the mechanism of synaptic change, and uncovered other types of activity-driven synaptic change in a variety of brain areas, including the cerebral cortex, hippocampus, basal ganglia, and cerebellum. Brain-derived neurotrophic factor (BDNF) and physical activity appear to play a beneficial role in the process.
Neuroscientists currently distinguish several types of learning and memory that are implemented by the brain in distinct ways:
Working memory is the ability of the brain to maintain a temporary representation of information about the task that an animal is currently engaged in. This sort of dynamic memory is thought to be mediated by the formation of cell assemblies—groups of activated neurons that maintain their activity by constantly stimulating one another.
Episodic memory is the ability to remember the details of specific events. This sort of memory can last for a lifetime. Much evidence implicates the hippocampus in playing a crucial role: people with severe damage to the hippocampus sometimes show amnesia, that is, inability to form new long-lasting episodic memories.
Semantic memory is the ability to learn facts and relationships. This sort of memory is probably stored largely in the cerebral cortex, mediated by changes in connections between cells that represent specific types of information.
Instrumental learning is the ability for rewards and punishments to modify behavior. It is implemented by a network of brain areas centered on the basal ganglia.
Motor learning is the ability to refine patterns of body movement by practicing, or more generally by repetition. A number of brain areas are involved, including the premotor cortex, basal ganglia, and especially the cerebellum, which functions as a large memory bank for microadjustments of the parameters of movement.
Research
Main article: Neuroscience
"Brain research" redirects here. For the scientific journal, see Brain Research.
The Human Brain Project is a large scientific research project, starting in 2013, which aims to simulate the complete human brain.
The field of neuroscience encompasses all approaches that seek to understand the brain and the rest of the nervous system. Psychology seeks to understand mind and behavior, and neurology is the medical discipline that diagnoses and treats diseases of the nervous system. The brain is also the most important organ studied in psychiatry, the branch of medicine that works to study, prevent, and treat mental disorders. Cognitive science seeks to unify neuroscience and psychology with other fields that concern themselves with the brain, such as computer science (artificial intelligence and similar fields) and philosophy.
The oldest method of studying the brain is anatomical, and until the middle of the 20th century, much of the progress in neuroscience came from the development of better cell stains and better microscopes. Neuroanatomists study the large-scale structure of the brain as well as the microscopic structure of neurons and their components, especially synapses. Among other tools, they employ a plethora of stains that reveal neural structure, chemistry, and connectivity. In recent years, the development of immunostaining techniques has allowed investigation of neurons that express specific sets of genes. Also, functional neuroanatomy uses medical imaging techniques to correlate variations in human brain structure with differences in cognition or behavior.
Neurophysiologists study the chemical, pharmacological, and electrical properties of the brain: their primary tools are drugs and recording devices. Thousands of experimentally developed drugs affect the nervous system, some in highly specific ways. Recordings of brain activity can be made using electrodes, either glued to the scalp as in EEG studies, or implanted inside the brains of animals for extracellular recordings, which can detect action potentials generated by individual neurons. Because the brain does not contain pain receptors, it is possible using these techniques to record brain activity from animals that are awake and behaving without causing distress. The same techniques have occasionally been used to study brain activity in human patients with intractable epilepsy, in cases where there was a medical necessity to implant electrodes to localize the brain area responsible for epileptic seizures. Functional imaging techniques such as fMRI are also used to study brain activity; these techniques have mainly been used with human subjects, because they require a conscious subject to remain motionless for long periods of time, but they have the great advantage of being noninvasive.
Design of an experiment in which brain activity from a monkey was used to control a robotic arm
Another approach to brain function is to examine the consequences of damage to specific brain areas. Even though it is protected by the skull and meninges, surrounded by cerebrospinal fluid, and isolated from the bloodstream by the blood–brain barrier, the delicate nature of the brain makes it vulnerable to numerous diseases and several types of damage. In humans, the effects of strokes and other types of brain damage have been a key source of information about brain function. Because there is no ability to experimentally control the nature of the damage, however, this information is often difficult to interpret. In animal studies, most commonly involving rats, it is possible to use electrodes or locally injected chemicals to produce precise patterns of damage and then examine the consequences for behavior.
Computational neuroscience encompasses two approaches: first, the use of computers to study the brain; second, the study of how brains perform computation. On one hand, it is possible to write a computer program to simulate the operation of a group of neurons by making use of systems of equations that describe their electrochemical activity; such simulations are known as biologically realistic neural networks. On the other hand, it is possible to study algorithms for neural computation by simulating, or mathematically analyzing, the operations of simplified "units" that have some of the properties of neurons but abstract out much of their biological complexity. The computational functions of the brain are studied both by computer scientists and neuroscientists.
Computational neurogenetic modeling is concerned with the study and development of dynamic neuronal models for modeling brain functions with respect to genes and dynamic interactions between genes.
Recent years have seen increasing applications of genetic and genomic techniques to the study of the brain and a focus on the roles of neurotrophic factors and physical activity in neuroplasticity. The most common subjects are mice, because of the availability of technical tools. It is now possible with relative ease to "knock out" or mutate a wide variety of genes, and then examine the effects on brain function. More sophisticated approaches are also being used: for example, using Cre-Lox recombination it is possible to activate or deactivate genes in specific parts of the brain, at specific times.
History
See also: History of neuroscience
Illustration by René Descartes of how the brain implements a reflex response
The oldest brain to have been discovered was in Armenia in the Areni-1 cave complex. The brain, estimated to be over 5,000 years old, was found in the skull of a 12 to 14-year-old girl. Although the brains were shriveled, they were well preserved due to the climate found inside the cave.
Early philosophers were divided as to whether the seat of the soul lies in the brain or heart. Aristotle favored the heart, and thought that the function of the brain was merely to cool the blood. Democritus, the inventor of the atomic theory of matter, argued for a three-part soul, with intellect in the head, emotion in the heart, and lust near the liver. The unknown author of On the Sacred Disease, a medical treatise in the Hippocratic Corpus, came down unequivocally in favor of the brain, writing:
Men ought to know that from nothing else but the brain come joys, delights, laughter and sports, and sorrows, griefs, despondency, and lamentations. ... And by the same organ we become mad and delirious, and fears and terrors assail us, some by night, and some by day, and dreams and untimely wanderings, and cares that are not suitable, and ignorance of present circumstances, desuetude, and unskillfulness. All these things we endure from the brain, when it is not healthy...— On the Sacred Disease, attributed to Hippocrates
Andreas Vesalius' Fabrica, published in 1543, showing the base of the human brain, including optic chiasma, cerebellum, olfactory bulbs, etc.
The Roman physician Galen also argued for the importance of the brain, and theorized in some depth about how it might work. Galen traced out the anatomical relationships among brain, nerves, and muscles, demonstrating that all muscles in the body are connected to the brain through a branching network of nerves. He postulated that nerves activate muscles mechanically by carrying a mysterious substance he called pneumata psychikon, usually translated as "animal spirits". Galen's ideas were widely known during the Middle Ages, but not much further progress came until the Renaissance, when detailed anatomical study resumed, combined with the theoretical speculations of René Descartes and those who followed him. Descartes, like Galen, thought of the nervous system in hydraulic terms. He believed that the highest cognitive functions are carried out by a non-physical res cogitans, but that the majority of behaviors of humans, and all behaviors of animals, could be explained mechanistically.
The first real progress toward a modern understanding of nervous function, though, came from the investigations of Luigi Galvani (1737–1798), who discovered that a shock of static electricity applied to an exposed nerve of a dead frog could cause its leg to contract. Since that time, each major advance in understanding has followed more or less directly from the development of a new technique of investigation. Until the early years of the 20th century, the most important advances were derived from new methods for staining cells. Particularly critical was the invention of the Golgi stain, which (when correctly used) stains only a small fraction of neurons, but stains them in their entirety, including cell body, dendrites, and axon. Without such a stain, brain tissue under a microscope appears as an impenetrable tangle of protoplasmic fibers, in which it is impossible to determine any structure. In the hands of Camillo Golgi, and especially of the Spanish neuroanatomist Santiago Ramón y Cajal, the new stain revealed hundreds of distinct types of neurons, each with its own unique dendritic structure and pattern of connectivity.
Drawing by Santiago Ramón y Cajal of two types of Golgi-stained neurons from the cerebellum of a pigeon
In the first half of the 20th century, advances in electronics enabled investigation of the electrical properties of nerve cells, culminating in work by Alan Hodgkin, Andrew Huxley, and others on the biophysics of the action potential, and the work of Bernard Katz and others on the electrochemistry of the synapse. These studies complemented the anatomical picture with a conception of the brain as a dynamic entity. Reflecting the new understanding, in 1942 Charles Sherrington visualized the workings of the brain waking from sleep:
The great topmost sheet of the mass, that where hardly a light had twinkled or moved, becomes now a sparkling field of rhythmic flashing points with trains of traveling sparks hurrying hither and thither. The brain is waking and with it the mind is returning. It is as if the Milky Way entered upon some cosmic dance. Swiftly the head mass becomes an enchanted loom where millions of flashing shuttles weave a dissolving pattern, always a meaningful pattern though never an abiding one; a shifting harmony of subpatterns.— Sherrington, 1942, Man on his Nature
The invention of electronic computers in the 1940s, along with the development of mathematical information theory, led to a realization that brains can potentially be understood as information processing systems. This concept formed the basis of the field of cybernetics, and eventually gave rise to the field now known as computational neuroscience. The earliest attempts at cybernetics were somewhat crude in that they treated the brain as essentially a digital computer in disguise, as for example in John von Neumann's 1958 book, The Computer and the Brain. Over the years, though, accumulating information about the electrical responses of brain cells recorded from behaving animals has steadily moved theoretical concepts in the direction of increasing realism.
One of the most influential early contributions was a 1959 paper titled What the frog's eye tells the frog's brain: the paper examined the visual responses of neurons in the retina and optic tectum of frogs, and came to the conclusion that some neurons in the tectum of the frog are wired to combine elementary responses in a way that makes them function as "bug perceivers". A few years later David Hubel and Torsten Wiesel discovered cells in the primary visual cortex of monkeys that become active when sharp edges move across specific points in the field of view—a discovery for which they won a Nobel Prize. Follow-up studies in higher-order visual areas found cells that detect binocular disparity, color, movement, and aspects of shape, with areas located at increasing distances from the primary visual cortex showing increasingly complex responses. Other investigations of brain areas unrelated to vision have revealed cells with a wide variety of response correlates, some related to memory, some to abstract types of cognition such as space.
Theorists have worked to understand these response patterns by constructing mathematical models of neurons and neural networks, which can be simulated using computers. Some useful models are abstract, focusing on the conceptual structure of neural algorithms rather than the details of how they are implemented in the brain; other models attempt to incorporate data about the biophysical properties of real neurons. No model on any level is yet considered to be a fully valid description of brain function, though. The essential difficulty is that sophisticated computation by neural networks requires distributed processing in which hundreds or thousands of neurons work cooperatively—current methods of brain activity recording are only capable of isolating action potentials from a few dozen neurons at a time.
Furthermore, even single neurons appear to be complex and capable of performing computations. So, brain models that do not reflect this are too abstract to be representative of brain operation; models that do try to capture this are very computationally expensive and arguably intractable with present computational resources. However, the Human Brain Project is trying to build a realistic, detailed computational model of the entire human brain. The wisdom of this approach has been publicly contested, with high-profile scientists on both sides of the argument.
In the second half of the 20th century, developments in chemistry, electron microscopy, genetics, computer science, functional brain imaging, and other fields progressively opened new windows into brain structure and function. In the United States, the 1990s were officially designated as the "Decade of the Brain" to commemorate advances made in brain research, and to promote funding for such research.
In the 21st century, these trends have continued, and several new approaches have come into prominence, including multielectrode recording, which allows the activity of many brain cells to be recorded all at the same time; genetic engineering, which allows molecular components of the brain to be altered experimentally; genomics, which allows variations in brain structure to be correlated with variations in DNA properties and neuroimaging.
Society and culture
As food
Main article: Brain as food
Gulai otak, beef brain curry from Indonesia
Animal brains are used as food in numerous cuisines.
In rituals
Some archaeological evidence suggests that the mourning rituals of European Neanderthals also involved the consumption of the brain.
The Fore people of Papua New Guinea are known to eat human brains. In funerary rituals, those close to the dead would eat the brain of the deceased to create a sense of immortality. A prion disease called kuru has been traced to this.
See also
Philosophy portal
Brain–computer interface
Central nervous system disease
List of neuroscience databases
Neurological disorder
Optogenetics
Outline of neuroscience
Aging brain | biology | 322049 | https://da.wikipedia.org/wiki/Neuron | Neuron | Et neuron (fra græsk: sene, nerve), også kaldet en nervecelle, er en celletype i nervesystemet.
De adskiller sig fra andre celler ved deres mange udløbere, kaldet dendritter og aksoner, og ved at være specialiseret i transmittering af signaler, dels elektrisk intraneuronalt og dels ved hjælp af transmitterstoffer interneuronalt, der virker på andre neuroners eller effektorvævenes receptorer (se for eksempel G-protein-koblede receptorer, GPCR og den motoriske endeplade). Af neurotransmittere kan nævnes serotonin, acetylkolin, dopamin, adrenalin og noradrenalin.
Formålet med hjernens netværk af neuroner er kommunikation og informationsbehandling.
Opbygning
Neuroner er de centrale komponenter i hjernen og rygmarven i centralnervesystemet (CNS) samt ganglier i det perifere nervesystem (PNS). En typisk neuron består af et cellelegeme (soma), dendritter og et axon. Dendritter er tynde strukturer, der udspringer fra cellekroppen og ofte strækker sig flere hundrede mikrometer og forgrener sig flere gange. En Axon, også kaldet en nervefiber når de er myelinerede, udspringer fra cellen ved et sted kaldet axon højen og rejser op til en en meter i mennesker og endnu længere i andre arter. En neurons cellekroppe har ofte flere dendritter, men aldrig til mere end ét Axon. Neuronets lange aksoner kan være omgivet af en isolerende fedtskede kaldet myelinskeden, som har til funktion at øge signaleringshastigheden.
Myelinskeden produceres og vedligeholdes i centralnervesystemet af oligodendrocytter og af Schwannske celler i det perifere nervesystem.
I de fleste tilfælde er neuroner genereret af særlige typer af stamceller. Det menes generelt, at neuroner ikke undergår celledeling, men nyere forskning hos hunde viser, at det i nogle tilfælde sker i nethinden. Astrocytter er stjerneformede gliaceller, der er også blevet observeret at blive til neuroner. Hos mennesker ophører neurogenese stort set i voksenalderen, men i to områder i hjernen, hippocampus og den olfaktoriske pære, er der beviser for generering af nye neuroner.
Kommunikation mellem neuroner
En neuron behandler og sender oplysninger gennem elektriske og kemiske signaler. Nervecellernes signaler bygger på en spændingsforskel mellem nervecellens indre og ydre, kaldet membranpotentialet. Er neuronet i hvile, kaldes spændingsforskellen for hvilemembranpotentialet. Er ændringen i membranpotentialet tilstrækkelig stor, genereres en (alt-eller-intet) elektrokemisk puls kaldet et aktionspotentiale, som bevæger sig hurtigt langs cellens axon og aktiverer synaptiske forbindelser til andre celler.
Neurontyper
Neuroner kan overordnet inddeles i projektionsneuroner Golgi type 1 med et enkelt langt akson, der kan kommunikere med andre regioner af nervesystemet, og interneuroner Golgi type 2 som kun befinder sig i en hjerneregion. Der er dog ikke skarp adskillelse mellem disse to typer.
Specialiserede neuroner omfatter: 1) sensoriske neuroner, der reagerer på berøring, lyd, lys og alle andre stimuli, der påvirker cellerne via vores sanser, som derefter sender signaler til rygmarven og hjernen; 2) motoriske neuroner, der modtager signaler fra hjernen og rygmarven, som bevirker muskelsammentrækninger og påvirker glandulare udgange; 3) interneuroner, der forbinder neuroner i den samme region i hjernen eller rygmarven.
Se også
Dendrit, akson, soma, gliacelle
Kranienerver
Neuropeptid
Eksterne henvisninger | danish | 0.434241 |
brain_no_pain_receptors_headache/Psychogenic_pain.txt | Psychogenic pain is physical pain that is caused, increased, or prolonged by mental, emotional, or behavioral factors, without evidence of physical injury or illness.
Headache, back pain, or stomach pain are some of the most common types of psychogenic pain. It is commonly accompanied by social rejection, broken heart, grief, lovesickness, regret, or other such emotional events. This pain can also be caused by psychological disorders such as anxiety and depression, which can affect the onset and severity of pain experienced.
The International Association for the Study of Pain (IASP) defines pain as "an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage." This definition was revised for the first time since 1979 in 2020, and was officially published in the ICD-11. The IASP broadens this definition to include psychogenic pain with the following points:
Furthermore, the ICD-11 removed the previous classification for psychogenic pain (persistent somatoform pain disorder) from the handbook in favor of understanding pain as a combination of physical and psychosocial factors. This is reflected in the definition for chronic primary pain, which acknowledges that pain stems from multiple personal and environmental factors and should be diagnosed "independently of identified biological or psychological contributors."
Some specialists believe that psychogenic chronic pain exists as a protective distraction to keep dangerous repressed emotions such as anger or rage unconscious. It remains controversial, however, that chronic pain might arise purely from emotional causes.
Diagnosis[edit]
There is no specific way of testing for psychogenic pain making it difficult to assess. There are many different criteria and factors considered for psychogenic pain diagnosis.
Presence of pain
Intense pain or suffering
Impairment of everyday functions
Symptoms ruled as unintentional
Symptoms do not fit criteria for other potential somatic or mental disorders
Treatment[edit]
For many patients a combination of psychotherapy and pharmacotherapy can help to alleviate or treat the symptoms of psychogenic pain. These treatments can include Cognitive behavioral therapy, acceptance and commitment therapy, or forms commonly used for chronic pain treatments. Interventional techniques can also be used. Treatments can address underlying feelings and emotional conflicts that can lead to psychogenic pain, as well as other potential causes of dysfunction with behavior, affect, and coping that can be seen in patients. In cases where therapy and medication do not show results, some may consider surgical intervention. These surgeries target portions of the brain associated with mood disorders and pain. Deep Brain Stimulation (DBS) is another possible treatment that works by stimulating parts of the brain related to behavior and emotion to relieve the psychological cause of the pain.
Controversy[edit]
The term "psychogenic pain" has begun to fall out of relevance in the scientific community, due to the implication that the pain is entirely psychological and thus not "real". The change in preferred nomenclature can be traced to 1994 when the DSM-IV removed the term in favor of the more holistic "Pain Disorder" section. The ICD-11 made a similar change, as mentioned above. It is important to note that this change is not universal, and is mostly confined to the English-speaking medical community. The term psychogenic pain is still used in non-English literature.
To fill the new gap in terminology left by the declining use of psychogenic pain, the term "nociplastic pain" was coined in 2016. Nociplastic pain is defined as chronic pain that cannot be classified as nociceptive (pain caused by the activation of nociceptors) or neuropathic (pain caused by damage to the nervous system). Nociplastic pain is functionally defined in one article as "pain arising from the altered function of pain-related sensory pathways in the periphery and CNS (Central Nervous System)",and, unlike psychogenic pain, can be diagnosed in conjunction with other types of pain.
See also[edit]
Pain disorder
Psychogenic disease
Psychological trauma
Psychoneuroimmunology
Psychosomatic medicine
Tension myositis syndrome | biology | 912867 | https://da.wikipedia.org/wiki/Posttraumatisk%20hjernesyndrom | Posttraumatisk hjernesyndrom | Posttraumatisk hjernesyndrom, også kendt som Post-concussion syndrome eller PCS, en en gruppe af symptomer der kan fortsætte i uger, måneder eller et år eller mere efter en hjernerystelse - en mild form for traumatisk hjerneskade (TBI). Antallet af ramte af PCS varierer, men de fleste studier viser at omkring 15 % af alle der tidligere har haft en enkelt hjernerystelse udvikler vedholdende symptomer associeret med skaden. En diagnose kan laves når symptomer fra hjernerystelse varer i mere end tre måneder efter skaden. Tab af bevidsthed er ikke et krav for diagnose af hjernerystelse eller PCS.
Tilstanden er associeret med en større vifte af symptomer: fysiske, såsom hovedpine; kognitive, såsom koncentrationsproblemer; og emotionelle og adfærdsmæssige, såsom irritabilitet. Mange af symptomer associeret med PCS er almindelig eller kan forekomme ved andre lidelser, så der er en risiko for forkert diagnose. Hovedpiner der opstår efter en hjernerystelse kan følelse som migræne eller spændingshovedpiner. Flest hovedpiner er spændingshovedpiner, hvilket kan associeres med en nakkeskade der opstod på samme tid som hovedskaden.
Selvom der ikke er nogen behandling for PCS, kan symptomerne blive behandlet; medikamenter og fysisk og adfærdsterapi kan blive brugt, og den enkelte kan blive klogere på symptomerne og få information om forventningerne til helingsprocessen. Majoriteten af PCS-patienter oplever af symptomer forsvinder helt efter en periode.
Det vides ikke hvad der får PCS til at opstå og forblive, eller hvorfor nogle mennesker der får en mindre traumatisk hjerneskab senere udvikler PCS, mens andre ikke gør. Lidelsens natur og diagnosen i sig selv har været genstand for intens debat siden det 19. århundrede. Men nogle risikofaktorer er blevet identificeret: allerede eksisterende medicinske eller psykologiske lidelser, forventninger om handicap, at være kvinde og høj alder øger alle risikoen for at lide af PCS. Fysiologiske og psykologiske faktorer tilstede før, under og efter skaden er alle mistænkt for at være involveret i udviklingen af PCS.
Nogle eksperter mener af posttraumatisk hjernesyndrom skyldes strukturel skade på hjernen, forstyrrelser af neurotrasmittersystemer eller kronisk overaktivering af hjernens egen immunforsvar. Andre mener at PCS-symptomer relaterer sig til almindelige psykologiske faktorer. Mest almindelige symptomer, såsom hovedpine, svimmelhed og søvnproblemer er lig dem der oftest oplevelse af personer diagnosticeret med depression, angst eller posttraumatisk stress lidelse. I mange tilfælde spiller både de fysiologiske effekter af hjernetrauma og emotionelle reaktioner til disse begivenheder spiller en rolle i udviklingen af symptomer.
Historie
Symptomerne der opstår efter en hjernerystelse er blevet beskrevet i flere forskellige rapport og skrifter i hundrede af år. Ideen om at dette sæt af symptomer danner en særskilt enhed begyndte at få bredere anderkendelse i den sidste del af det 19. århundrede. John Erichsen, en kirurg fra London, spillede en vigtig rolle i udviklingen af studiet af PCS. Kontroverserne omkring årsagen til PCS begyndte i 1866 da Erichsen udgav en artikel omkring vedholdende symptomer efter at have fået et mildt hovedtrauma. Han foreslog at lidelse skyldtes "molekulær disarragement" på rygsøjlen. Lidelsen blev oprindeligt kaldt "jernbane rygsøjle", fordi de fleste skader der var undersøgt, skete hos jernbanearbejdere. Selvom nogle af hans samtidige var enige i at lidelsen var organisk baseret, tilskrev andre symptomerne en psykologisk faktor, eller ligefrem foregivelse. I 1879 blev ideen om at det var et fysisk problem der var ansvarlig, udfordret af Rigler, der foreslog at årsagen til de vedvarende symptomer faktisk var "kompensationsneuroser": jernbanens praksis med at kompenserer arbejdere der var blev skadet fik flere klager frem. Senere, blev ideen om at hysteri var ansvarlig for symptomerne efter en mild hovedskade fremsagt af Charcot. Kontroverser omkring lidelsen fortsætte gennem det 20. århundrede. Under 1. verdenskrig led mange soldater af gådefulde symptomer efter at være tæt på en detonation, men uden noget tegn på et sår i hovedet. Lidelsen blev kaldt granatchok, og en psykologisk forklaring blev til sidst foretrukket. I 1934 havde den nuværende tanke om PCS erstattet ideer om hysteri som årsagen til post-hjernerystelsessymptomer. Britiske autoriteter forbød termen granatchok under 2. verdenskrig, for at undgå et epidemi af tilfælde, og termen posttrauma hjernerystelses stadie blev opfundet i 1939 for at beskrive "forstyrrelsen i bevidstheden, uden nogen umiddelbar eller indlysende patologiske ændringer i hjernen". Termen posttraumatisk hjernesyndrom var i brug fra 1941
I 1961 var H. Miller den første til at bruge termen "uheldsneurose" for at referere til syndromet der nu kaldes PCS og hævdede af lidelsen kun opstår i situationer hvor folk har mulighed for at blive kompenseret for skaden. De reelle årsager til lidelsen er stadig uklare.
Tegn og symptomer
Tidligere blev termen PCS også brugt til at referere til umiddelbare fysiske symptomer eller symptomer efter hjernerystelse, efter en mindre TBI eller hjernerystelse. Sværhedsgraden af disse symptomer formindskes typisk hurtigt. Yderligere kan symptomernes karakter kan ændrer sig over tid: akutte symptomer er mest almindeligt af fysiologisk karakter, mens vedvarende symptomer gerne er mestendels psykologisk. Symptomer såsom lydfølsomhed, koncentrations- og hukommelsesproblemer, irritabilitet, depression, og angst kan kaldes 'sene symptomer' fordi de generelt ikke opstår umiddelbart efter skaden, men oftere i dagene eller ugerne efter skaden. Kvalme og døsighed opstår ofte akut efter hjernerystelse. Hovedpine og svimmelhed opstår umiddelbart efter skaden, men kan også være vedvarende.
Fysisk
En almindelige lidelse ved PCS er hovedpine. Selvom de fleste har samme type hovedpine som de oplevede før skaden, rapporterer folk med PCS ofte flere eller længerevarende hovedpiner. Mellem 30 og 90 % af folk der blive behandlet for PCS rapporterer oftere hovedpine, og mellem otte og 32 % har dem stadig efter et år.
Svimmelhed er et andet almindeligt symptom, som halvdelen af folk diagnosticeret med PCS rapporterer, og det er stadig til stede i op til 25 % et år efter skaden. Ældre mennesker har især en høj risiko for svimmelhed, hvilket kan være med til at at skabe yderligere skader og højere rater af dødelighed på grund af fald.
Omkring 10 % af patienter med PCS udvikler lys- eller lydoverfølsomhed, omkring fem procent oplever en nedsat smags- eller lugtesans, og omkring 14 % rapporterer om slørret syn. Der er også folk med dobbeltsyn eller ringen for ørerne. PCS kan forårsage insomni, udkørthed, og andre problemer med søvnen.
Psykologiske og adfærdsmæssige
Psykologiske lidelser, der er tilstede hos halvdelen af PCS-ramte, kan inkludere irritabilitet, angst, depression, og en ændringer i personlighed. Andre emotionelle og adfærdsmæssige symptomer kan inkludere rastløshed, aggression, og humørsvingninger. Nogle symptomer, såsom apati, insomni, irritabilitet og manglende motivation, kan skyldes andre samtidige lidelser, såsom depression.
Højere mentalfunktioner
Almindelige symptomer associeret med PCS-diagnosen er relateret til kognition, opmærksomhed, og hukommelse, især kortidshukommelse, hvilket også kan gøre andre problemer værre, såsom at glemme aftaler eller svært ved at gennemføre arbejde. I et studie, havde en ud af fire med PCS hukommelsesproblemer et år efter skaden, men de fleste eksperter er enige om at de kognitive symptomer forsvinder inden for seks måneder til et år efter skaden hos langt størstedelen.
Årsager
Spørgsmålet om årsagen eller årsagerne til PCS er blevet voldsomt debatteret i mange år, og er stadig kontroversiel. Det er ikke kendt hvilken grad symptomerne er på grund af fysiologiske ændringer eller på grund af andre faktorer som præeksisterende psykiatriske lidelser eller faktorer relateret til sekundær gain eller handicapkompensation. Subjektiviteten af klagerne komplicerer bedømmelsen og gør det svært at fastslå om symptomer overdrives eller lades som om.
Mens årsagerne til symptomerne der opstår umiddelbart efter en hjernerystelse formentlig er fysiologiske, er det mindre klart at vedholdende post-hjernerystelsessymptomer har en helt organisk basis, og ikkeorganiske faktorer formentlig er involveret i symptomer der varer i mere end tre måneder. PCS kan også forværres af psykosociale faktorer, kronisk smerte eller en interaktion af nogle eller alle af disse. Størstedelen af eksperter mener at PCS resulterer i en mikstur af faktorerne, herunder præeksisterende psykologiske faktorer, og dem direkte relateret til den fysiske skade.
Fysiologisk
inflaKoventionelle hjernescanningsstudier efter hjernerystelse viser typisk en normal tilstand. Men, studier har fundet nogle diskrete fysiologiske ændringer forbundet med PCS ved at bruge flere nye billeddiagnostiske metoder. Studier der har brugt positronemissionstomografi har forbundet PCS med en reduktion af glukoseforbrug i hjernen. Ændringer i cerebral blodgennemstrømning er også blevet set op til tre år efter en hjernerystelse i studier der har brugt single-foton tomografi (SPECT). Mindst et studie med funktionel magnetresonanstomografi (fMRI) har vist forskelle i hjernefunktion under opgaver der involverede hukommelse, efter en mild traumatisk hjerneskade (mTBI), selvom de ikke specifikt undersøgte PCS. Yderligere studier har vist, ved brug af forskellige MRI-teknikker (såsom Diffusionstensorafbildning (DTI) MRI), at individer med PCS har forskellige abnormaliteter i deres hjernestruktur. Lignende fund er blevet rapporteret hos soldater med eksplosionsinduceret mTBI/PCS.
Et nyt dansk studie viser, at hjernens immunceller bliver aktiveret efter hjernerystelse, og at aktiveringen er størst og mest tydeligt persisterende efter 3-4 måneder i de patienter der udvikler post-commotio syndrom.
Ikke alle med PCS har abnormaliteter på billedescanninger, og abnormalitet fundet i studier såsom fMRI, PET og SPECT kan resultere i fra andre komorbide lidelser såsom klinisk depression, kronisk smerte eller posttraumatisk belastningsreaktion (PTSD). Fortalere for det synspunkt at PCS har en fysiologisk base, peger på fund der viser at børn demonstrerer mangler i standardiserede test af kognitiv funktion efter en mild TBI. Nogle få studier har vist at folk med PCS scorer lavere end kontrolgrupper i neuropsykologiske test der måler opmærksomhed, verbal læring, ræsonnement og informationsbearbejdning, men problemer relateret til indsats og sekundær gevinst kan ikke udelukkes som medvirkende til disse forskelle. Heling, som målt af resultater af hyppige kognitive test korrelerer med bedring af symptomer; individer diagnosticeret med PCS kan stadig rapporterer subjektive symptomer efter deres præstation i test af kognitiv funktion er vendt tilbage til normalen. Et andet studie fandt at selvom børn med PCS havde dårligere resultater i test af kognitiv funktion efter skaden, havde de også dårligere adfærdsmæssig justering før skaden, en børn uden nogen vedholdende symptomer; disse fund støtter ideen om at PCS kan være resultatet af en kombination af faktorer, såsom hjernedysfunktion som resultat af hovedskade og præeksisterende psykologiske eller sociale problemer. Forskellige symptomer kan forudsiges af forskellige faktorer, eksempelvis fandt et studie at kognitive og fysiske symptomer ikke var forudsagt af den måde som forældre og familiemedlemmer agerede i forhold til skaden og til dens effekter, men psykologiske og adfærdsmæssige symptomer var.
Psykologisk
Det er blevet fremført at psykologiske faktorer spiller en vigtig rolle i tilstedeværelsen af PCS. Udviklingen af PCS kan være på grund af en kombination af faktorer som tilvænning til effekter af skaden, præeksisterende sårbarheder og hjernedysfunktion. Tilbagefald relateret til skaden, eksempelvis problemer på arbejdet eller med fysisk eller social funktionsevne, kan fungere som en stressfaktor der interagerer med præeksisterende faktorer som personlighed og mentale lidelser der forårsager og fastholder PCS. I et studie, var de daglige stressniveauer korreleret med PCS-symptomer hos både ramte af hjernerystelse og kontrolgruppen, men i andre studier var stress ikke signifikant relateret til symptomerne.
Iatrogen effekter (dem der skyldes medicinsk intervention) kan også ske når individer får misvisende eller ukorrekt information relateret til bedring af symptomer. Denne information kan få folk til at fokusere og dvæle ved ideen om at deres hjerner er permanent skadet. Det ladet til at selv forventningen om symptomer kan hjælpe på udviklingen af PCS ved at få folk med mTBI til at fokusere på symptomer og derfor opfatte dem mere intensive, at tilegne symptomer fra andre årsager til skaden, og at underestimere mængden af symptomer før skaden.
Diagnose
International Statistical Classification of Diseases and Related Health Problems (ICD-10) og American Psychiatric Associations Diagnostic and Statistical Manual of Mental Disorders har nogle kriterier for posttraumatisk hjernesyndrom og posttraumatisk hjernelidelse (PCD).
ICD-10 etablerede et sæt diagnosekriterier for PCS i 1992. For at opfylde disse kriterier, skulle en patient have haft en hovedskade "oftest voldsom nok til at have skabt bevidstløshed" og derefter udvikle tre ud af otte symptomer markeret med et flueben i tabellen til højre under "ICD-10" inden for fire uger.
Omkring 38 % der har fået en hovedskader med symptomer på hjernerystelse og ingen radiologisk bevis for hjernelæsioner opfylder disse kriterier. Udover disse symptomer, frygter nogle af dem der opfylder ICD-10-kriterierne for PCS, vil give dem permanent hjerneskade, hvilket kan forværre de oprindelige symptomer. Optagethed af skaden, kan medfølges af antagelse af en "sygerolle" og hypokondri. Kriterierne fokusere på subjektive symptomer, og nævner at neuropsykologiske evidens for signifikant forringelse ikke er tilstede. Med deres fokus på psykologiske faktorer, deler ICD-10-kriterierne ideen om at årsagen til PCS er funktionel. Ligesom ICD-10, definere ICD-9-CM PCS med hensyn til subjektive symptomer, og diskuterer den større hyppighed af PCS hos personer med en historik der involverer mentale lidelse eller et financielt incitament for en diagnose.
DSM-IV oplister kriterier for diagnose af PCD hos personer der har fået et hovedtrauma med vedholden posttraumatisk amnesi, tab af bevidsthed, eller posttraumatisk anfald. Disse symptomer skal være tilstede i tre måneder efter skaden, og skal have været fravarende eller mindre mindre før skaden. Yderligere skal patienten opleve sociale problemer som resultat heraf, om må ikke opfylde kriterier for andre lidelser der forklare symptomerne bedre.
Neuropsykologiske test eksisterer for at måle forskelle i kognitiv funktion der kan resultere fra PCS. Stroop Farvetest og 2&7 Processing Speed Test (der begge måler mangler i hastigheden af mental bearbejdning) kan forudsige udviklingen af kognitive problemer fra PCS. En test kaldet Rivermead Postconcussion Symptoms Questionnaire, et sæt spørgsmål der måler graden af 16 forskellige post-hjernerystelsessymptomer, kan blev lavet selv eller ved hjælp fra en interviewer. Andre test der kan forudsige udviklingen af PCS inkluderer Hopkins Verbal Learning A (HVLA) og Digit Span Forward eksamination. HVLA tester verbal læring og hukommelse ved at præsentere en serie af ord og giver point baseret på antallet der huskes, og digit span måler opmærksomhedseffektivitet ved at få eksaminanden til at gentage de cifre testeren siger, i den samme rækkefølge som de bliver sagt. Hertil kan neuropsykologiske test blive udført for at måle simulering (overdrivelse af eller finde på symptomer).
Differentialdiagnostik
PCS, der deler mange symptomer med en bred vifte af andre lidelser, vil meget sandsynligt blive misdiagnosticeret hos personer med disse lidelser. Kognitive og affektive symptomer der opstår efter en traumatisk skade kan tilskrives en mTBI, men faktisk kan den være på grund af posttraumatisk stress-lidelse, hvilket nemt kan misdiagnosticeres som PCS og visa versa. Affektive lidelse såsom depression har nogle symptomer der kan ligne dem fra PCS og føre til en forkert diagnose af sidstnævnte; disse inkludere problemer med koncentration, emotionel labilitet, angst og søvnproblemer- Depression, der er meget almindelig hos vedholden PCS, kan forværre andre PCS-symptomer, såsom hovedpine og problemer med koncentrationen, hukommelse og søvn. PCS kan også dele symptomer med kronisk træthedssyndrom, fibromyalgi og udsættelse for nogle toksiner. Traumatisk hjerneskade kan være skyld i hypothalamus eller hypofysen, og mangler i hypofysehormoner (hypopituitarism) kan skabe lignende symptomer på PCS; i disse tilfælde, symptomer kan behandles ved at udskifte ethvert hormon der er i mangel.
Behandling
Behandling af posttraumatisk hjernesyndrom involverer typisk behandlinger der adresserer specifikke symptomer; eksempelvis kan smertestillende tages for hovedpine, eller medicin mod depression eller insomni. Hvile anbefales, men er kun lidt effektivt. Fysio- og adfærdsterapi kan også bruges mod problemer såsom tab af balance, og problemer med koncentrationen.
Medicinering
Selvom der ikke eksisterer en farmakologisk behandling for PCS, udskriver læger medikamenter for symptomer der findes i andre lidelser; eksempelvis antidepressiver der bruges mod depression der ofte efterfølger mTBI. Bivirkninger af medikamenter kan påvirke folk der lider af konsekvenserne af mTBI voldsommere end de påvirker andre, og derfor anbefales det at medikamenter undgås hvis det er muligt; der kan være en fordel ved at undgå narkotiske medikamenter. Yderligere, kan nogle smertestillende udskrevet mod hovedpine, give tilbagevendende hovedpine når de ikke længere bruges.
Psykoterapi
Psykologisk behandling, til hvilket omkring 40 % af PCS-patienter henvises til, har vist sig at reducere problemerne. Vedvarende handicap kan blive behandlet med terapi for at forbedre funktionsevnen på arbejde, socialt eller i andre kontekster. Målet med terapi er at hjælpe med en gradvis tilbagevenden til arbejde, eller andre aktiviteter fra før skaden, i det omfang symptomerne tillader det. En protokol til PCS-behandling er blevet designet på baggrund af principperne bag kognitiv adfærdsterapi (KAT), en psykoterapi målrettet mod at påvirke forstyrrede følelser ved at forbedre tanker og adfærd. KAT kan hjælpe med at forhinde iatrogene symptomer. – dem der opstår fordi sundhedspersonel skaber en forventning om at de vil. Der er en risiko for at "forslagets kraft" kan forværre symptomerne eller skabe langstidshandicap; derfor, skal en terapeut tænke det psykologiske ophav med også, og ikke gå ud fra at alle symptomer er et direkte resultat af den neurologiske skade fra skaden.
I situationer såsom uheld med motorkøretøjer eller efter en voldeligt angreb, kan posttraumatisk hjernesyndrom blev ledsaget af posttraumatisk belastningsreaktion, der er vigtigt at genkende og behandle separat fra PCS. Folk med PTSD, depression eller angst kan blive behandlet med medicin og psykoterapi.
Undervisning
Information om symptomerne og deres almindelige tidsforløb er en del af den psykologiske terapi, og er mest effektiv når den gives kort efter skaden. Siden stress forværrer PCS-symptomer, og vise versa, er en vigtig del af behandlingen en forsikring om at symptomerne er normale og information om hvordan man håndterer hæmningerne. Et studie fandt at PCS-patienter der blev coachet til gradvist at vende tilbage til aktiviteter, fik fortalt hvilke symptomer de skulle forvente, og trænede i hvordan de skulle håndteres havde en reduktion i symptomer sammenlignet med en kontrolgruppe af ikkeskadet personer. Early education has been found to reduce symptoms in children as well.
Neuroterapi
Neuroterapi er en operant konditioneringstest hvor patienter gives en konditionel audio/visuel belønning efter at have produceret en bestemt type af hjernebølgeaktivitet. Nylig neuroterapeutiske forbedringer i QEEG kan identificere specifikke hjernebølgemønstre der har behov for at blive rettet til. Studier har vist at neuroterapi er en effektiv behandling af PCS og andre lidelser med lignende symptomer.
Prognose
Prognosen for PCS anses generelt for at være positiv, men en fuldstænding opløsning af symptomerne i mange, men ikke alle, tilfælde. For 50 % af de ramte, vil PCS-symptomer vare fra nogle få dage til flere uger efter den oprindelige skade skete. Hos andre kan symptomerne vedvare i tre til seks måneder, men evidens indikerer at mange tilfælde er fuldstændigt helbredt inden for seks måneder. Majoriteten af symptomer er stort set væk i halvdelen af folk med hjernerystelsen en måned efter skade, og omkring to tredjedele af folk med et mindre hovedtrauma are næste symptomfri inden for tre måneder. Vedvarende, ofte voldsomme hovedpiner, er det længstblivende symptom i mange tilfælde, og er det symptom der oftest ikke forsvinder helt. Det bliver ofte angivet i litteraturen at 15-30 % af PCS-ramte ikke er kommet sig et år efter skaden, men dette estimat er upræcist fordi det er baseret på studier af folk indlagt på et hospital, hvorfor metodologien er blevet kritiseret. Hos cirka 15 % ramte, kan symptomerne vare ved i år eller være permanente. Hvis symptomerne ikke er forsvundet inden for et år, er de formentlig permanente, men forbedringer kan stadig ske selv efter to eller tre år, eller de kan pludselig opstår efter lang tid uden større forbedringer. Ældre mennesker, og dem der tidligere har lidt af en hovedskade, vil formentlig bruge længere tid på at hele.
Måde hvorpå børn kaperer med skaden efter den opstår kan have en større indflydelse end faktorer der eksisterede før skaden. Børn mekanisme for at håndtere deres skader kan have en effekt på symptomvarigheden, og forældre der ikke håndtere angst effektivt i forhold til deres børn post-skade funktionsevne kan være dårligere til at hjælpe deres børn med at komme sig.
Hvis endnu et slag til hovedet forekommer efter en hjernerystelse, før symptomerne er væk, er der en mindre risiko for at udvikle den alvorlige second-impact syndrome (SIS). Ved SIS vil hjernen hurtigt hæve, hvilket kraftigt øger det intrakraniale tryk. Folk der har haft gentagne milde hovedskader over en længere periode, såsom boksere og gridiron-fodboldspillere, har en risiko for CTE (eller den relaterede variant dementia pugilistica), en sværd kronisk lidelse der involverer en nedgang i mentale og fysiske evner.
Epidemiologi
Det vides ikke hvor almindelig PCS er. Estimater af prævalensen tre måneder efter skaden er mellem 24 og 84 %, en variation der muligvis er skabt af forskellige populationer eller studiemetoder. Den estimerede incidens af PPCS (vedvarende PCS ) er omkring 10 % af mTBI-tilfælde. Siden PCS per definition kun eksisterer hos personer der har haft en hovedskade, er demografien og risikofaktorerne meget lig dem for hovedskader; eksempelvis har yngre voksne en højere risiko en andre for at pådrage sig en hovedskade, og, som konsekvens heraf, udvikle PCS.
Eksistensen af PCS hos børn er kontroversiel. Det er muligt at børns hjerner har nok plasticitet, så de ikke påvirkes af langvarige konsekvenser af hjernerystlese (selvom sådanne konsekvenser er kendt for at stamme fra moderate til svære hovedtraumaer). På den anden side, så er børns hjerner mere udsatte for skader, da de stadig er under udvikling og har færre evner der kan kompenserer for manglerne. Klinisk forskning har fundet flere tilfælde af PCS-symptomer hos børn med TBI, end hos dem med skader på andre dele af kroppen, og at symptomerne er mere almindelige hos ængstelige børn. Symptomer hos børn er lig dem hos voksne, men børn oplever færre af dem. Evidens fra kliniske studier har vist at atleter i gymnasiealderen havde langsommere helingsproces fra hjernerystelse målt ved neuropsykologiske test, end atleter i universitetsalderen og voksne. PCS er sjældent hos små børn.
Risikofaktorer
En bred vifte af faktorer er blevet identificeret som værende forudsigende for PCS, herunder lav socioøkonomisk status, tidligere mTBI, en alvorlig associeret skade, hovedpine, en aktiv retssag, og det kvindelige køn. At være mere end 40 og kvinde er også blevet identificeret som prædiktiv for en PCS-diagnose, og kvinder har en tendens til at rapportere flere værre symptomer. Yderligere kan udviklingen af PCS blive forudsagt ved at have en historik med alkoholmisbrug, dårlige kognitive evner før skaden, en personlighedslidelse, eller en medicinsk lidelse ikke relateret til skaden. PCS er også fremherskende hos folk der har en historie med psykiatriske lidelser, såsom klinisk depression eller angst før skaden.
mTBI-relaterede faktorer der øger risikoen for vedvarende PCS-symptomer inkluderer skader associeret med akut hovedpine, svimmelhed, eller kvalme; en akut Glasgow Coma Score på 13 eller 14; og få en ny hovedskade før man er helet fra den første. Risikoen for at udvikle PCS viser sig også at øges hos folk der har traumatiske minder om skaden, eller forventer at blive handicappet som følge af skaden.
Se også
Hjernerystelsesforeningen
Referencer
Eksterne henvisninger
"Viden om hjernerystlse" Hjernerystelsesforeningen
Hjernerystelse | danish | 0.499851 |
brain_no_pain_receptors_headache/Nerve.txt | A nerve is an enclosed, cable-like bundle of nerve fibers (called axons) in the peripheral nervous system.
Nerves have historically been considered the basic units of the peripheral nervous system. A nerve provides a common pathway for the electrochemical nerve impulses called action potentials that are transmitted along each of the axons to peripheral organs or, in the case of sensory nerves, from the periphery back to the central nervous system. Each axon, within the nerve, is an extension of an individual neuron, along with other supportive cells such as some Schwann cells that coat the axons in myelin.
Within a nerve, each axon is surrounded by a layer of connective tissue called the endoneurium. The axons are bundled together into groups called fascicles, and each fascicle is wrapped in a layer of connective tissue called the perineurium. Finally, the entire nerve is wrapped in a layer of connective tissue called the epineurium. Nerve cells (often called neurons) are further classified as sensory, motor, or mixed nerves.
In the central nervous system, the analogous structures are known as nerve tracts.
Structure[edit]
Cross-section of a nerve
Each nerve is covered on the outside by a dense sheath of connective tissue, the epineurium. Beneath this is a layer of fat cells, the perineurium, which forms a complete sleeve around a bundle of axons. Perineurial septae extend into the nerve and subdivide it into several bundles of fibres. Surrounding each such fibre is the endoneurium. This forms an unbroken tube from the surface of the spinal cord to the level where the axon synapses with its muscle fibres, or ends in sensory receptors. The endoneurium consists of an inner sleeve of material called the glycocalyx and an outer, delicate, meshwork of collagen fibres. Nerves are bundled and often travel along with blood vessels, since the neurons of a nerve have fairly high energy requirements.
Within the endoneurium, the individual nerve fibres are surrounded by a low-protein liquid called endoneurial fluid. This acts in a similar way to the cerebrospinal fluid in the central nervous system and constitutes a blood-nerve barrier similar to the blood–brain barrier. Molecules are thereby prevented from crossing the blood into the endoneurial fluid. During the development of nerve edema from nerve irritation (or injury), the amount of endoneurial fluid may increase at the site of irritation. This increase in fluid can be visualized using magnetic resonance neurography, and thus MR neurography can identify nerve irritation and/or injury.
Categories[edit]
Nerves are categorized into three groups based on the direction that signals are conducted:
Afferent nerves conduct signals from sensory neurons to the central nervous system, for example from the mechanoreceptors in skin.
Efferent nerves conduct signals from the central nervous system along motor neurons to their target muscles and glands.
Mixed nerves contain both afferent and efferent axons, and thus conduct both incoming sensory information and outgoing muscle commands in the same bundle. All spinal nerves are mixed nerves, and some of the cranial nerves are also mixed nerves.
Nerves can be categorized into two groups based on where they connect to the central nervous system:
Spinal nerves innervate (distribute to/stimulate) much of the body, and connect through the vertebral column to the spinal cord and thus to the central nervous system. They are given letter-number designations according to the vertebra through which they connect to the spinal column.
Cranial nerves innervate parts of the head, and connect directly to the brain (especially to the brainstem). They are typically assigned Roman numerals from 1 to 12, although cranial nerve zero is sometimes included. In addition, cranial nerves have descriptive names.
Terminology[edit]
Main article: Anatomical terms of neuroanatomy
Specific terms are used to describe nerves and their actions. A nerve that supplies information to the brain from an area of the body, or controls an action of the body is said to innervate that section of the body or organ. Other terms relate to whether the nerve affects the same side ("ipsilateral") or opposite side ("contralateral") of the body, to the part of the brain that supplies it.
Development[edit]
Nerve growth normally ends in adolescence, but can be re-stimulated with a molecular mechanism known as "Notch signaling".
Regeneration[edit]
If the axons of a neuron are damaged, as long as the cell body of the neuron is not damaged, the axons can regenerate and remake the synaptic connections with neurons with the help of guidepost cells. This is also referred to as neuroregeneration.
The nerve begins the process by destroying the nerve distal to the site of injury allowing Schwann cells, basal lamina, and the neurilemma near the injury to begin producing a regeneration tube. Nerve growth factors are produced causing many nerve sprouts to bud. When one of the growth processes finds the regeneration tube, it begins to grow rapidly towards its original destination guided the entire time by the regeneration tube. Nerve regeneration is very slow and can take up to several months to complete. While this process does repair some nerves, there will still be some functional deficit as the repairs are not perfect.
Function[edit]
A nerve conveys information in the form of electrochemical impulses (as nerve impulses known as action potentials) carried by the individual neurons that make up the nerve. These impulses are extremely fast, with some myelinated neurons conducting at speeds up to 120 m/s. The impulses travel from one neuron to another by crossing a synapse, where the message is converted from electrical to chemical and then back to electrical.
Nerves can be categorized into two groups based on function:
An afferent nerve fiber conducts sensory information from a sensory neuron to the central nervous system, where the information is then processed. Bundles of fibres or axons, in the peripheral nervous system are called nerves, and bundles of afferent fibers are known as sensory nerves.
An efferent nerve fiber conducts signals from a motor neuron in the central nervous system to muscles. Bundles of these fibres are known as efferent nerves.
Nervous system[edit]
Main article: Nervous system
The nervous system is the part of an animal that coordinates its actions by transmitting signals to and from different parts of its body. In vertebrates it consists of two main parts, the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain, brainstem and spinal cord. The PNS consists mainly of nerves, which are enclosed bundles of the long fibers or axons, that connect the CNS to all remaining body parts.
Nerves that transmit signals from the CNS are called motor or efferent nerves, while those nerves that transmit information from the body to the CNS are called sensory or afferent. Spinal nerves serve both functions and are called mixed nerves. The PNS is divided into three separate subsystems, the somatic, autonomic, and enteric nervous systems. Somatic nerves mediate voluntary movement.
The autonomic nervous system is further subdivided into the sympathetic and the parasympathetic nervous systems. The sympathetic nervous system is activated in cases of emergencies to mobilize energy, while the parasympathetic nervous system is activated when organisms are in a relaxed state. The enteric nervous system functions to control the gastrointestinal system. Both autonomic and enteric nervous systems function involuntarily. Nerves that exit from the cranium are called cranial nerves while those exiting from the spinal cord are called spinal nerves.
Clinical significance[edit]
Micrograph demonstrating perineural invasion of prostate cancer. H&E stain.
Cancer can spread by invading the spaces around nerves. This is particularly common in head and neck cancer, prostate cancer and colorectal cancer.
Nerves can be damaged by physical injury as well as conditions like carpal tunnel syndrome (CTS) and repetitive strain injury. Autoimmune diseases such as Guillain–Barré syndrome, neurodegenerative diseases, polyneuropathy, infection, neuritis, diabetes, or failure of the blood vessels surrounding the nerve all cause nerve damage, which can vary in severity.
Multiple sclerosis is a disease associated with extensive nerve damage. It occurs when the macrophages of an individual's own immune system damage the myelin sheaths that insulate the axon of the nerve.
A pinched nerve occurs when pressure is placed on a nerve, usually from swelling due to an injury, or pregnancy and can result in pain, weakness, numbness or paralysis, an example being CTS. Symptoms can be felt in areas far from the actual site of damage, a phenomenon called referred pain. Referred pain can happen when the damage causes altered signalling to other areas.
Neurologists usually diagnose disorders of nerves by a physical examination, including the testing of reflexes, walking and other directed movements, muscle weakness, proprioception, and the sense of touch. This initial exam can be followed with tests such as nerve conduction study, electromyography (EMG), and computed tomography (CT).
Other animals[edit]
A neuron is called identified if it has properties that distinguish it from every other neuron in the same animal—properties such as location, neurotransmitter, gene expression pattern, and connectivity—and if every individual organism belonging to the same species has exactly one neuron with the same set of properties. In vertebrate nervous systems, very few neurons are "identified" in this sense. Researchers believe humans have none—but in simpler nervous systems, some or all neurons may be thus unique.
In vertebrates, the best known identified neurons are the gigantic Mauthner cells of fish. Every fish has two Mauthner cells, located in the bottom part of the brainstem, one on the left side and one on the right. Each Mauthner cell has an axon that crosses over, innervating (stimulating) neurons at the same brain level and then travelling down through the spinal cord, making numerous connections as it goes. The synapses generated by a Mauthner cell are so powerful that a single action potential gives rise to a major behavioral response: within milliseconds the fish curves its body into a C-shape, then straightens, thereby propelling itself rapidly forward. Functionally this is a fast escape response, triggered most easily by a strong sound wave or pressure wave impinging on the lateral line organ of the fish. Mauthner cells are not the only identified neurons in fish—there are about 20 more types, including pairs of "Mauthner cell analogs" in each spinal segmental nucleus. Although a Mauthner cell is capable of bringing about an escape response all by itself, in the context of ordinary behavior other types of cells usually contribute to shaping the amplitude and direction of the response.
Mauthner cells have been described as command neurons. A command neuron is a special type of identified neuron, defined as a neuron that is capable of driving a specific behavior all by itself. Such neurons appear most commonly in the fast escape systems of various species—the squid giant axon and squid giant synapse, used for pioneering experiments in neurophysiology because of their enormous size, both participate in the fast escape circuit of the squid. The concept of a command neuron has, however, become controversial, because of studies showing that some neurons that initially appeared to fit the description were really only capable of evoking a response in a limited set of circumstances.
In organisms of radial symmetry, nerve nets serve for the nervous system. There is no brain or centralised head region, and instead there are interconnected neurons spread out in nerve nets. These are found in Cnidaria, Ctenophora and Echinodermata.
History[edit]
Further information: History of neurology and neurosurgery
Herophilos (335–280 BC) described the functions of the optic nerve in sight and the oculomotor nerve in eye movement. Analysis of the nerves in the cranium enabled him to differentiate between blood vessels and nerves (Ancient Greek: νεῦρον (neûron) "string, plant fiber, nerve").
Modern research has not confirmed William Cullen's 1785 hypothesis associating mental states with physical nerves, although popular or lay medicine may still invoke "nerves" in diagnosing or blaming any sort of psychological worry or hesitancy, as in the common traditional phrases "my poor nerves",
"high-strung", and "nervous breakdown".
See also[edit]
Biology portal
Connective tissue in the peripheral nervous system
Dermatome (anatomy)
List of nerves of the human body
Nerve injury
Neuropathy
Nerve injury classification | biology | 322049 | https://da.wikipedia.org/wiki/Neuron | Neuron | Et neuron (fra græsk: sene, nerve), også kaldet en nervecelle, er en celletype i nervesystemet.
De adskiller sig fra andre celler ved deres mange udløbere, kaldet dendritter og aksoner, og ved at være specialiseret i transmittering af signaler, dels elektrisk intraneuronalt og dels ved hjælp af transmitterstoffer interneuronalt, der virker på andre neuroners eller effektorvævenes receptorer (se for eksempel G-protein-koblede receptorer, GPCR og den motoriske endeplade). Af neurotransmittere kan nævnes serotonin, acetylkolin, dopamin, adrenalin og noradrenalin.
Formålet med hjernens netværk af neuroner er kommunikation og informationsbehandling.
Opbygning
Neuroner er de centrale komponenter i hjernen og rygmarven i centralnervesystemet (CNS) samt ganglier i det perifere nervesystem (PNS). En typisk neuron består af et cellelegeme (soma), dendritter og et axon. Dendritter er tynde strukturer, der udspringer fra cellekroppen og ofte strækker sig flere hundrede mikrometer og forgrener sig flere gange. En Axon, også kaldet en nervefiber når de er myelinerede, udspringer fra cellen ved et sted kaldet axon højen og rejser op til en en meter i mennesker og endnu længere i andre arter. En neurons cellekroppe har ofte flere dendritter, men aldrig til mere end ét Axon. Neuronets lange aksoner kan være omgivet af en isolerende fedtskede kaldet myelinskeden, som har til funktion at øge signaleringshastigheden.
Myelinskeden produceres og vedligeholdes i centralnervesystemet af oligodendrocytter og af Schwannske celler i det perifere nervesystem.
I de fleste tilfælde er neuroner genereret af særlige typer af stamceller. Det menes generelt, at neuroner ikke undergår celledeling, men nyere forskning hos hunde viser, at det i nogle tilfælde sker i nethinden. Astrocytter er stjerneformede gliaceller, der er også blevet observeret at blive til neuroner. Hos mennesker ophører neurogenese stort set i voksenalderen, men i to områder i hjernen, hippocampus og den olfaktoriske pære, er der beviser for generering af nye neuroner.
Kommunikation mellem neuroner
En neuron behandler og sender oplysninger gennem elektriske og kemiske signaler. Nervecellernes signaler bygger på en spændingsforskel mellem nervecellens indre og ydre, kaldet membranpotentialet. Er neuronet i hvile, kaldes spændingsforskellen for hvilemembranpotentialet. Er ændringen i membranpotentialet tilstrækkelig stor, genereres en (alt-eller-intet) elektrokemisk puls kaldet et aktionspotentiale, som bevæger sig hurtigt langs cellens axon og aktiverer synaptiske forbindelser til andre celler.
Neurontyper
Neuroner kan overordnet inddeles i projektionsneuroner Golgi type 1 med et enkelt langt akson, der kan kommunikere med andre regioner af nervesystemet, og interneuroner Golgi type 2 som kun befinder sig i en hjerneregion. Der er dog ikke skarp adskillelse mellem disse to typer.
Specialiserede neuroner omfatter: 1) sensoriske neuroner, der reagerer på berøring, lyd, lys og alle andre stimuli, der påvirker cellerne via vores sanser, som derefter sender signaler til rygmarven og hjernen; 2) motoriske neuroner, der modtager signaler fra hjernen og rygmarven, som bevirker muskelsammentrækninger og påvirker glandulare udgange; 3) interneuroner, der forbinder neuroner i den samme region i hjernen eller rygmarven.
Se også
Dendrit, akson, soma, gliacelle
Kranienerver
Neuropeptid
Eksterne henvisninger | danish | 0.434241 |
brain_no_pain_receptors_headache/Headache_(EP).txt | Headache is the fourth EP by American post-hardcore band Big Black. The record generated some controversy due to a cover photograph of a shotgun suicide victim whose head was split in half; it only appeared on a very limited edition of the record and was later replaced with a drawing by Savage Pencil. The identity of the dead man in the original album cover remains unknown.
Original pressings bore a sticker reading, "Warning! Not as good as Atomizer, so don't get your hopes up, cheese!" as a frank communication from the members of Big Black to their fans that they did not regard the Headache EP to be as strong as the band's previous release. This also came at a time when major labels were first starting to warn parents that albums contained "explicit lyrics", and while all of Big Black's records contained such lyrics, Headache's sticker was an ironic dig at mainstream music. Copies of the sticker also appeared on the vinyl release of Sonic Youth's Master=Dik EP as an independent music in-joke.
Track listing[edit]
"My Disco" – 2:51
"Grinder" – 2:22
"Ready Men" – 3:50
"Pete, King of all Detectives" – 2:40
Notes[edit]
^ Kellman, Andy. "allmusic ((( Headache > Review )))". Allmusic. Retrieved March 17, 2010.
^ Brackett, Nathan. "Big Black". The New Rolling Stone Album Guide. November 2004. pg. 69, cited March 17, 2010
^ (in English) Selected Savage Pencil Bibliography, Exhibitions, Discography, Record Covers
External links[edit]
Headache/Heartbeat package (Dementlieu, "ARBEIT MACHT FREI: Big Black Discography") - includes links to photos of limited cover
vteBig Black
Steve Albini
Jeff Pezzati
Santiago Durango
Dave Riley
Studio albums
Atomizer
Songs About Fucking
Live albums
Sound of Impact
Pigpile
Compilation albums
The Hammer Party
The Rich Man's Eight Track Tape
EPs
Lungs
Bulldozer
Racer-X
Headache
Singles
"He's a Whore" / "The Model"
Related articles
Naked Raygun
Rapeman
Shellac
Category
Authority control databases
MusicBrainz release group
This 1980s punk rock album–related article is a stub. You can help Wikipedia by expanding it.vte | biology | 635080 | https://sv.wikipedia.org/wiki/Ooooooohhh%E2%80%A6%20On%20the%20TLC%20Tip | Ooooooohhh… On the TLC Tip | Ooooooohhh... On the TLC Tip är debutalbumet av den amerikanska R&B-gruppen TLC, utgivet den 25 februari 1992 på LaFace Records. Albumet, som producerades under ledning av skivbolagets ägare Babyface och L.A. Reid, inrymmer hiphop och new jack swing. Albumet nådde som bäst plats 14 på Billboard 200 i USA, där det senare har sålts i över fyra miljoner exemplar (4xplatina).
Titeln Ooooooohhh... On the TLC Tip kommer från den sista raden ur Lisa Lopes rapdel i singeln "Ain't 2 Proud 2 Beg", en låt om sexuell befrielse ur ett kvinnligt, humoristiskt perspektiv. I låtens musikvideo, som främjade säkrare sex, hade Lopes fäst ett inslaget kondompaket på sitt vänstra öga och fick därav smeknamnet Left Eye. Med uppföljarsingeln "Baby-Baby-Baby" nådde gruppen andra plats på Billboard Hot 100. Flera låtar bygger på samplingar från andra artisters låtar.
Bakgrund
TLC skrev kontrakt med skivbolaget LaFace Records den 16 augusti 1991 och började producera sitt debutalbum. Bandet arbetade tillsammans med Babyface, L.A. Reid, Dallas Austin, De Funky Bunch, Jermaine Dupri och Marley Marl, som hjälpte Lopes med låtskrivandet. Albumet spelades in vid Doppler Studios, Studio LaCoCo och Bosstown Recording Studios i Atlanta samt vid House Of Hits i New York. Produktionen slutfördes i december 1991.
Mottagande
Steve Huey på Allmusic betygsatte albumet 3/5 och skrev att "fastän det är ojämnt förtjänar de bästa stunderna på On the TLC Tip sin popularitet, och banade vägen för gruppens dundersuccé vid nästa tillfälle".
Låtlista
* betecknar en medproducent
Samplingar
Flera av låtarna på albumet innehåller samplingar från andra artisters låtar.
Ain't 2 Proud 2 Beg
"School Boy Crush" av Average White Band
"Jungle Boogie" av Kool and the Gang
"Escape-ism" av James Brown
"Fly, Robin, Fly" av Silver Convention
"Take Me to the Mardi Gras" av Bob James
"I Want to Take You Higher" av Sly and the Family Stone
"Get Me Back on Time, Engine Number 9" av Wilson Pickett
Shock Dat Monkey
"Get Me Back on Time, Engine Number 9" av Wilson Pickett
"Funky Drummer" av James Brown
"Funky President (People It's Bad)" av James Brown
"Slippin' Into the Darkness" av War
"The Champ" av The Mohawks
"Impeach the President" av The Honey Drippers
"God Make Me Funky" av The Headhunters feat. The Pointer Sisters
"Kool is Back" av Funk, Inc.
"Different Stroke" av Syl Johnson
"Shock the Monkey" av Peter Gabriel
Hat 2 da Back
"Big Ole Butt" av LL Cool J
"What Makes You Happy" av KC & the Sunshine Band
"Here We Go (Live at the Funhouse)" av Run-D.M.C.
"Shut the Eff Up! (Hoe)" av MC Lyte
"Synthetic Substitution" av Melvin Bliss
"Poetry" av Boogie Down Productions
"Hihache" av Lafayette Afro Rock Band
Das Da Way We Like 'Em
"Think (About It)" av Lyn Collins
"UFO" av ESG
What About Your Friends
"Blues and Pants" av James Brown
"Sing a Simple Song" av Sly and the Family Stone
Bad by Myself
"Welcome to the Terrordome" av Public Enemy
"Peter Piper" av Run-D.M.C.
"Last Night Changed It All" av Esther Williams
"Peace is Not the Word to Play" av Main Source
This is How It Should Be Done
"We're a Winner" av The Impressions
"I Know You Got Soul" av Eric B. & Rakim
Depend on Myself
"Son of Shaft" av The Bar-Kays
"Humpin'" av The Bar-Kays
"The Assembly Line" av Commodores
"Scorpio" bavy Dennis Coffey and The Detroit Guitar Band
Listplaceringar
Referenser
Album av TLC
Amerikanska debutalbum
Engelskspråkiga musikalbum
Musikalbum 1992
Album producerade av Dallas Austin | swedish | 0.370641 |
below_temperature_muscles_not_work/benefitsofextremetem.txt | Edition:Available editions
United States
Donate
Get newsletter
Become an author
Sign up as a reader
Sign in
The Conversation
Search analysis, research, academics…
Academic rigor, journalistic flair
Arts + CultureEconomyEducationEnvironment + EnergyEthics + ReligionHealthPolitics + SocietyScience + TechPodcasts
Anna Ewa Bieniek/Shutterstock
Benefits of extreme temperature workouts – not as great as you might think
Published: September 10, 2019 6:38am EDT
Authors
Lindsay Bottoms
Principal Lecturer, Exercise Physiology, University of Hertfordshire
Daniel Muniz
Senior Lecturer, Exercise Physiology, University of Hertfordshire
Disclosure statement
The authors do not work for, consult, own shares in or receive funding from any company or organization that would benefit from this article, and have disclosed no relevant affiliations beyond their academic appointment.
Partners
View all partners
CC BY ND
We believe in the free flow of information
Republish our articles for free, online or in print, under a Creative Commons license.
Email
X (Twitter)7
Facebook312
LinkedIn
Print
First there were heated fitness studios, now the latest trend is working out in frigid temperatures. Although there are some health benefits associated with each of these regimes, there are also some risks. Here’s what you need to know.
Hot workouts
The ideal body temperature is around 37⁰C. When you exercise, your muscles are very inefficient and only 25% of the energy is used for movement. The other 75% of the energy muscles produce is lost as heat, increasing the temperature of your body. If it exceeds 40⁰C, it can be dangereous, so your body tries to keep the temperature at around 37⁰C.
One strategy to prevent body temperature from rising is getting more blood to the skin, which is people’s faces flush during an intense workout. If you exercise in an already hot environment, the difference between body temperature and room temperature is small, and your heart needs to work harder trying to get more blood to the skin.
Another way your body gets rid of excess heat is by warming up sweat to the point at which it evaporates, which then takes heat with it when it evaporates to the air. You can lose up to two litres of water every hour through sweat. As you sweat more during exercise in the heat, it is important to replace the fluid you have lost or your blood can become thicker, which also puts a strain on the heart.
Both ways of keeping the body under 40⁰C add an extra burden on the heart. As a result, cardio workouts are more difficult in hot environments and endurance performance is decreased in hot environments.
But exercising in a hot environment might be good for short activities that need powerful muscle contractions. There is also a school of thought that light-intensity workouts, such as yoga, can benefit because the extra strain on the heart during exercise in the heat can increase the number of calories you burn. But research by Central Michigan University showed that there was no difference in exercise intensity, so the amount of calories burnt doing yoga at 20⁰C and 35⁰C were the same.
Increasing muscle temperature can increase flexibility and reduce the risk of injury. But a hot environment does not necessarily mean increased muscle temperature.
Regularly undertaking workouts in a hot environment can help endurance performance in the heat, but your overall fitness and health may not get anything extra.
Working out in the heat can improve endurance in the heat, but that’s about all. Klemen K. Misic/Shutterstock
Cold workouts
In the cold, your body is hotter than the surrounding environment and can easily get rid of the heat produced in the muscles during exercise. This stops the temperature of the body from increasing during prolonged exercise and performance in your cardio session will usually be better on cold days. Researchers from the University of Aberdeen found endurance to be best at around 10⁰C (compared with 4⁰C, 21⁰C and 31⁰C).
When it is really cold, the heat produced by your muscles is not enough to maintain a core temperature at 37⁰C. The body has ways to cope with extreme cold environments, such as shivering. Shivering is essentially muscles contracting to produce heat, not movement. As with any muscle contraction, shivering requires energy and burns calories, so at rest you may burn more calories in the cold than in normal temperatures.
Another strategy to cope with cold is to use fat to produce heat. Researchers in the US have shown repeated workouts in the cold increases the amount of brown fat in your body. Brown fat is known as “good fat” as it burns calories.
Both shivering and burning fat consume calories, and studios that hold workout classes at 7⁰C have begun to appear, with the idea that they can help people lose weight. Researchers from Spain found that exposure to a progressively cold environment can increase energy expenditure by up to 30%, which corresponds to around 500 calories for 24 hours of cold exposure. This means exposure to cold can help you lose weight faster, but the effect would be small for a one-hour workout in the cold. And feeling cold is unpleasant.
If you want to increase your muscle strength, doing workouts in the cold probably won’t help. Muscles work best at hot temperatures. When muscles get cold, the force they can produce decreases. You are also more likely to get injured in the cold, but the extra risk of injury is reduced if you warm up properly.
Take-home message
The small extra amount of calories burnt in extreme environments (hot or cold) may help you lose weight, but it could be outweighed by the increased health risks and reduced performance. Doing 150 minutes of physical activity a week is the key to good health, regardless of the temperature.
Exercise
Fitness
Cold workout
Want to write?
Write an article and join a growing community of more than 182,600 academics and researchers from 4,946 institutions.
Register now
Editorial Policies
Community standards
Republishing guidelines
Friends of The Conversation
Analytics
Our feeds
Donate
Get newsletter
Who we are
Our charter
Our team
Partners and funders
Resource for media
Contact us
En Español
Privacy policy
Terms and conditions
Corrections
Copyright © 2010–2024, The Conversation US, Inc. | biology | 1364233 | https://no.wikipedia.org/wiki/Treningsfysiologi | Treningsfysiologi | Treningsfysiologi er studiet av de akutte reaksjoner og kroniske tilpasninger av fysisk trening. I tillegg studerer mange treningsfysiologer effekten av trening på patologi, og mekanismene som gjør at trening kan redusere eller reversere sykdom. En treningsfysiologs studiefelt kan kan omfatte, men er ikke begrenset til, biokjemi, bioenergi, hjerte-funksjon, hematologi, biomekanikk, skjelettmuskelfysiologi, nevroendokrin funksjon, og sentrale og perifere nervesystemet. Treningsfysiologi jobber i spennet fra grunnforskere, til kliniske forskere, klinikere, til idrettstrenere.
Energiforbruk
Mennesket har kapasitet til å bruke energi i mange timer med vedvarende anstrengelse. For eksempel, en person som sykler med en hastighet på 26,4 km / t (16.4 mph) gjennom 8204 km (5098 miles) over 50 påfølgende dager bruker totalt 1145 MJ (273,850 kcal; 273850 kalorier) med en gjennomsnittlig utgangseffekt på 182,5 W
Skjelettmuskler forbrenner 90 mg (0,5 mmol) glukose hvert minutt under kontinuerlig aktivitet (for eksempel under gjentatte bevegelser i kneet), genererer ≈24 W mekanisk energi, og siden muskelenergi konverterterer bare 22-26% effektivt, ≈76 W varmeenergi. Hvileskjelettmuskulatur har en basal forbrenning (hvilende energiforbruk) på 0,63 W / kg som utgjør 160 gangers forskjell mellom energiforbruket for inaktive og aktive muskler. For kort varighet av muskulær anstrengelse, kan energiforbruket bli langt større: når en voksen mann hopper opp fra en knebøy kan han mekanisk generere 314 W / kg. Slike raske bevegelser kan hos andre dyr som for eksempel sjimpansen, og noen små øgler, generere den dobbelte energimengden av dette.
Dette energiforbruk er svært stor sammenlignet med basal hvilemetabolisme hos den voksne menneskekroppen. Denne mengden varierer noe med størrelse, kjønn og alder, men er vanligvis mellom 45 W og 85 W. Totalt energiforbruk (TEE) på grunn av energi musklene har brukt er mye høyere og avhenger av det gjennomsnittlige nivået av fysisk arbeid og mosjon gjort i løpet av en dag. Mosjon, særlig hvis over svært lange perioder, dominerer energimetabolismen i kroppen. Energiforbruket ved fysisk aktivitet korrelerer sterkt med kjønn, alder, vekt, puls, og V [O.sub.2max] til en person, under fysisk aktivitet.
Referanser
Evolusjonsbiologi
Menneskelig evolusjon
Treningsteori | norwegian_bokmål | 0.55114 |
below_temperature_muscles_not_work/documentrepidrep1typ.txt | J. exp. Biol. 115,333-344 (1985)
Printed in Great Britain The Company of Biologists Limited 1985
TEMPERATURE AND MUSCLE
BY ALBERT F. BENNETT
School of Biological Sciences, University of California, Irvine, California 9271 7,
USA.
SUMMARY
Rates of force development, contraction and relaxation of vertebrate
skeletal muscle are temperature dependent with Qlo values of approximately 2. Maximal forces developed have a low or negative thermal dependence. The functional basis of these patterns is poorly understood. Muscle
performance generally does not acclimate. There appears to have been
some evolutionary adaptation among species and classes to different
thermal regimes, such that muscles from cold-adapted species maintain
better mechanical performance at low temperatures than do those from
warm-adapted animals. However, rate processes remain strongly thermally dependent even in animals with low or variable body temperatures.
This thermal dependence of muscle in vitro is reflected in behavioural
performance: maximal force generation in vivo is temperature independent and time-dependent activities are more rapid at higher muscle temperatures.
INTRODUCTION
Mechanical performance of muscle is greatly influenced by temperature, as are
most biological processes. Maximal forces developed by muscles and their rates
of force generation, contraction, relaxation and power output are all altered when
body temperature varies. As these muscular forces and rate processes underlie
behavioural capacities, these may also be thermally dependent. Such factors as
maximal locomotor speed and reaction rates may change with temperature and
be so slow in the cold that effective escape or pursuit by an animal is curtailed.
In animals that are subjected to varying or low body temperatures, we might expect
to find adaptations which minimize the thermal dependence of muscle performance.
The influence of temperature on the mechanical performance of vertebrate skeletal
muscle is reviewed in this paper, first for one species of lizard and then for vertebrates
more generally. Adaptations of muscle performance to temperature are examined,
underlying mechanisms of thermal dependence are reviewed and implications for
behaviour are discussed.
Key words: Acclimation, adaptation, mechanical properties.
TEMPERATURE AND MECHANICAL PERFORMANCE
First, we will examine the influence of temperature on the contractile performance
of muscle from a lizard. Lizards are a particularly useful group of animals for such
studies as they may naturally experience a wide range of body temperatures daily.
Many species also have very high thermal tolerance. Data are presented here (Fig. 1)
on the influence of temperature on several aspects of force generation and contraction
of skeletal muscle of the lizard Dipsosaums dorsalis (Marsh & Bennett, 1985).
Dipsosaums inhabits the hot desert regions of California and regulates field body
temperatures at approximately 40°C. However, it must maintain the capacity to
react and move even when its body temperature is far below these diurnal levels.
The reported data were measured on the white portion of the iliofibularis muscle of
the hind limb. This preparation is composed almost exclusively of fast glycolytic
fibres (Gleeson, Putnam & Bennett, 1980), which constitute the large majority of
fibres in all the locomotory muscles of this species (Putnam, Gleeson & Bennett,
1980).
All contractile rate processes in this muscle, both isometric and isotonic, are greatly
accelerated by increasing temperature, even up to 44°C. In isometric twitch, the rates
of both tension development (measured as the inverse of time-to-peak tension, TPT)
and relaxation (measured as the inverse of time to return to 50 % of maximal twitch
tension, I/~RT) have Qlo values of 2.36 and 2.82, respectively, between 20 and
30°C. Maximal rate of isometric tetanic tension development (dpo/dt) has a Qlo of
2-22 over this range. The maximal velocity of shortening (V,,) and maximal power
output (w,,) during isotonic contractions are similarly thermally dependent with
Qlo values of 1.95 and 2-42, respectively. All these rates have a thermal dependence
similar to those of most other biological processes (i.e. Qlo=2-3, Precht,
Christophersen, Hensel & Larcher, 1973; Prosser, 1973).
In contrast, the temperature dependence of force exerted during isometric
contraction is substantially different from that of contractile rate processes. Tetanic
tension (Po) has a significant but very low thermal dependence: its thermal ratio
(Rlo, the ratio of two quantities measured over a 10°C interval, Bennett, 1984) is
1.2 between 20 and 30°C. Maximal Po is attained at 40°C. Twitch tension (P,) is
maximal at 15"C, declining at higher temperatures (Rlo = 0.62 between 20 and
30°C).
Force and its rate of development thus have very different thermal sensitivities in this
lizard. Nearly maximal muscular performance is obtained at normal field activity
temperature (40°C): contraction and relaxation rates are rapid and tetanic force is
maximal. Twitch tension is the only factor that is not near its maximal value at this
temperature. Exposure to low temperatures greatly retards the speed of muscle
contraction in Dipsosaums. This dependence may restrict locomotory responses at low
body temperature: the speed of limb movement during burst escape speed in this lizard is
limited by the time course of the muscle twitch at 25°C and below (Marsh & Bennett,
1985). Low temperature does not similarly affect force output. Nearly the same tetanic
tension can be produced and twitch tension is even increased.
Temperature and muscle 335
How representative is the thermal dependence of muscle function in Dipsosaums?
Qlo and Rlo values measured approximately between 20 and 30°C from all other
studies available on vertebrate skeletal muscle are given in Fig. 2. Most of these
-
20
:, /, lG/:7
OD
5 A en Z
= 10 200 *
;
2 -3
0 0
10 20 30 40 10 20 30 40
Temperature ("C)
Fig. 1. Contractile performance of the iliofibularis muscle of the lizard Dipsosaums dorsalis as a
function of temperature. (A) twitch tension, P,; (B) tetanic tension, Po; (C) time-to-peak twitch
tension, TPT; (D) time of half relaxation from peak twitch tension, 112 RT; (E) rate of rise of tetanic
tension dPo/dt; (F) maximal velocity of shortening, V,,; (G) maxipal power output, IT,,,. Data
from Marsh & Bennett (1985).
observations were made on amphibian and mammalian muscle. Although the variance
in the data is high, due to such factors as the diversity of animals examined, differing
fibre type composition of the muscles and different measurement techniques, the
pattern is clear. As in the lizard muscle, rate processes in general are thermally
dependent with Qlo values of approximately 2, tetanic tension has a very low thermal
dependence and twitch tension often decreases over this temperature range.
ADAPTATIONS TO TEMPERATURE
This general pattern of high thermal dependence of rate processes might be
expected to pose problems for poikilothermic organisms. As muscle temperature
changes, so do contractile speeds and possibly reaction rates or locomotor ability.
Poikilotherms with low body temperatures might not attain maximal performance of
which their muscles are capable due to an extrinsic factor, temperature, rather than an
intrinsic structural or biochemical limitation. For example, the lizard Gerrhonotus
multicarinatus has field active body temperatures of about 25 "C, but maximal rates of
muscle contraction and burst escape speed at 35-40°C (Bennett, 1980; Putnam &
Bennett, 1982), body temperatures far in excess of those encountered under any
Force Rate
TPT 112 RT
Fig. 2. Thermal dependence of force and rate of contraction of vertebrate skeletal muscle between 20
and 30°C. Wide horizontal bar indicates median value; narrow horizontal bars, 25th and 75th
percentile observations; vertical bar, range. Number of observations given in parentheses. Data from
summary by Bennett (1984). For details of abbreviations see legend to Fig. 1.
Temperature and muscle 337
natural conditions. Such a pattern of thermal dependence does not appear particularly
adaptive. To what extent has there been adjustment to minimize the thermal
perturbation of contractile rate processes in animals with low and/or variable body
temperatures? Few data on this topic exist, so no confident generalizations are
possible, but the studies available suggest some interesting adaptive patterns.
Acclimation
When an individual animal is exposed to a new thermal regime, it often shows
compensatory changes in its physiological reactions (acclimation). Biological rate
processes are initially altered in accordance with their Qlo upon acute temperature
exposure. Over several days or weeks, these rates often return partially or completely
to their original levels, even while the new thermal regime is maintained (Precht et al.
1973; Prosser, 1973). Several comprehensive studies on the effect of long-term
temperature exposure have been done on the mechanical performance of skeletal
muscles of anuran amphibians. As illustrated for twitch kinetics (Fig. 3), no
acclimation of either force generation (P,, Po) or contractile rate (TPT, ~/ZRT,
dpo/dt, V,,,, w,,,) has been demonstrated (Renaud & Stevens, 1981a,b; Rome,
1983). The initial depression of contraction rates by cold exposure is maintained
indefinitely. A similar lack of acclimation occurs in locomotor capacity of these
animals (Putnam & Bennett, 1981). In fish, myofibrillar ATPase activity, which
should be reflected in V, (BBrBny, 1967), has been shown to acclimate in goldfish
(Johnston, 1979) but not in killifish (Sidell, Johnston, Moerland & Goldspink, 1983).
TPT 112 RT
0J , I 1 I I 1
5 15 25 5 15 25
Experimental temperature ("C)
Fig. 3. Mean values of time-to-peak tension (TPT) and half relaxation time (1/2RT) for twitch
contractions of the sartorius muscle of Ranapipiens acclimated to 5°C (filled circles), 15OC (open
circles) and 25OC (squares). The only significant difference among acclimation groups is a longer
112 RT for 25°C-acclimated frogs measured at 15T. Data from Renaud & Stevens (1981~).
The lack of acclimation of muscle function is a very puzzling result in view of its
obvious importance to behavioural capacity and may reflect a constraint on
adaptation.
Interspecz$c compan'sons
Studies comparing species naturally exposed to different thermal regimes show a
different pattern of adjustment of muscle function over evolutionary time. In lizards,
species with lower activity temperatures have lower Qlo values for TPT, 112 RT and
dpo/dt and faster twitch responses measured at any common temperature (Putnam &
Bennett, 1982). Maximization of P, at preferred thermal levels previously reported
(Licht, 1964) has not been confirmed (Putnam & Bennett, 1982). The range of
temperatures over which lizard muscles can function is clearly affected by their
thermal regimes (Ushakov, 1964; Licht, 1964; Putnam & Bennett, 1982): muscles
from more thermophilic species lose contractile ability and undergo irreversible heat
damage at substantially higher temperatures than do those of more cryophilic animals.
In fish, actomyosin ATPases of species from cold environments have both lower Qlo
values and greater activities at any common temperature than those from warmadapted fish (Fig. 4) (Johnston, Walesby, Davison & Goldspink, 1977; Johnston &
Walesby, 1977, 1979). In both fish and lizards, evolutionary adaptation to temperature has evidently proceeded with both a shift (translation) and rotation of the
rate-temperature curve.
r I I I I I i
0 10 20 30
Temperature ("C)
Fig. 4. Activity of MgZ+, Caz+-activated actomyosin ATPase from teleost fish adapted to different
thermal regimes. Cold-adapted species: Salvelinus alpinus, arctic (open squares) ; Champsocephalus
gunnan', antarctic (open circles); Cottus bubalis, North Sea (open triangles). Warm-adapted species:
Dascyllus aruanus (filled circles) and Pomatocentrus pulchem'mus (filled squares), both tropical.
Note log axis of enzyme activity. Data from Johnston & Walesby (1979).
Temperature and muscle 339
It should be emphasized, however, that these interspecific differences are only
relative. In absolute terms, rate processes are still very thermally dependent even in
cold-adapted species.
Comparison of muscle from homeothems and poikilothems
Are there major differences in the thermal dependence of muscle function in
homeotherms and poikilotherms? One might anticipate that the latter would show
adaptations to minimize thermal dependence and the former would optimize function
over a narrow range of high temperatures. It should be noted, however, that even
muscles from homeotherms may undergo major temperature changes depending on
ambient conditions and work intensity (e.g. Saltin, Gagge & Stolwijk, 1968).
Some functional differences are apparent between muscle from anuran amphibians
and mammals (Bennett, 1984). Anuran muscle develops maximal P, at O°C;
mammalian peak P, usually occurs at 20°C. Maximal Po is maintained at lower
temperatures in anurans: Po of anuran muscle declines below approximately lS°C,
that of mammalian muscle always decreases below 25 OC (Fig. 5). The time course of a
muscle twitch, both TPT and 112 RT, has a lower Qlo in anurans than in mammals.
Anuran muscle is thus capable of producing maximal tension at lower temperatures
than is mammalian muscle and its rate processes are less temperature sensitive, at least
in regard to twitch kinetics. However, the distinction between the thermal
dependence of saurian and mammalian muscle function is much less clear (Bennett,
1984). Although maximal P, is developed at lower temperatures in lizard than in
mammalian muscle, the thermal dependencies of Po and contractile rate processes are
almost identical in these groups. The anuran-mammalian differences probably reflect
100 -
Mammalian
80 -
-
d
// Amphibian
10 20 30
Temperature ("C)
Fig. 5. Thermal dependence of tetanic tension (Po) in mammalian and anuran amphibian skeletal
muscle. Data summarized from the literature by Bennett (1984).
adaptation to very different thermal regimes rather than a dichotomy in the functional
capacity of muscle from poikilotherms and homeotherms.
It is apparent from the foregoing that some adjustments of vertebrate skeletal
muscle function have evolved with respect to temperature. These adaptations in rate
processes, however, have not been substantial. Muscle from poikilotherms still has a
marked thermal dependence and rate processes are often faster at temperatures above
those normally encountered. Lack of acclimation of these properties in individual
animals further points to a lack of plasticity in these systems in regard to temperature
adjustment. 1n view of the small number of studies, it should be evident that more
comparative work is required to substantiate or alter these conclusions.
FUNCTIONAL BASIS OF THERMAL DEPENDENCE
The biochemical and functional properties that underlie these patterns of thermal
dependence are not well understood. The pronounced thermal dependence of
contractile rate processes are expected, as several of the steps underlying contraction
and relaxation are enzymatically catalysed and have rates with Qlo values of 2.0 or
more. For instance, strong thermal dependencies have been demonstrated for the rate
of ca2+ release (Blinks, Riidel & Taylor, 1978; Rall, 1979), the activity of actomyosin
ATPase (Bendall, 1964; Hartshorne, Barns, Parker & Fuchs, 1972) and ca2+ uptake
by sarcoplasmic reticulum (Yamamoto & Tonomura, 1967; Blinks et al. 1978).
However, the correspondence between the thermal dependence of contractile events
and isolated enzyme systems associated with them is often poor. For example,
although actomyosin ATPase is thought to be an important determinant of V,,,, the
thermal dependence of the former is much greater than that of the latter (Biriny,
1967). An attempt to determine limiting factors of contractile events by comparisons
of thermal dependencies would probably be no more successful than previous
attempts to elucidate controlling reactions of biochemical pathways by measurement
of Arrhenius activation energies.
Twitch tension is generally maximal at 0-20°C, depending on species and muscle
type, and declines at higher temperatures. In some sense, the muscle is not as fully
activated by a single stimulus at temperatures above 20°C (Ranatunga, 1977). The
ratio of P,/P~ approaches 1.0 at low temperatures but is less than 0.5 at higher
temperatures (Putnam & Bennett, 1982). One possible explanation for this response is
insufficient ca2+ release into fibres at higher temperatures. This is evidently not the
case, however, as activation heat, which represents the energetic cost of ca2+ release
and removal, is thermally independent, indicating no deficiency of release at higher
temperatures (Homsher, Mommaerts, Ricchiuti & Wallner, 1972; Rall, 1979).
Additionally, measurements with aequorin indicate high levels of ca2+ within the
fibre at higher temperatures (Blinks et al. 1978). The latter measurements also
indicate a much shorter time course for the presence of ca2+ within the fibre at higher
temperatures and a consequently shorter period of activation. At high temperatures,
insufficient time may be available to attain maximal force. Twitch contraction may be
looked upon as a competition between actomyosin ATPase activity and ca2+ removal
Temperature and muscle 341
by the sarcoplasmic reticulum (Josephson, 1981). These processes evidently have
different thermal dependencies, with the latter being more temperature sensitive; the
Qlo of 1/2RT is generally greater than that of TPT measured on the same muscle
(Hill, 1951; Walker, 1960; Bennett, 1984). Thus at low temperatures, greater force
can be attained by the contractile proteins, even though their intrinsic contractile
velocity is also slowed, before deactivation processes become effective. This
explanation of the thermal dependence of P,, while attractive, has not been critically
tested.
Tetanic tension generally has a low but significant thermal dependence below
15-25"C, depending on species (Fig. 5). As the muscle is supposedly completely
activated by repetitive stimuli, one might expect Po to be thermally independent over
the entire temperature range. One explanation for lower Po at low temperatures is a
smaller number of cross-bridge attachments at those temperatures. However,
measurements of instantaneous stiffness, which is dependent upon the number of
cross-bridges attached, do not support this hypothesis (Ford, Huxley & Simmons,
1977; Kuhn et al. 1979; Bressler, 198 1). Instantaneous stiffness is either thermally
independent or has a different thermal dependence from that of Po. Consequently, the
number of cross-bridges attached or a thermally-dependent bridge cycling does not
appear to account for the observed pattern. Bressler (1981) suggests that increasing
temperature may increase the tension per cross-bridge during tetanus.
It should be clear that further work is required to explain the bases of the thermal
dependence of muscle function. Investigations exploiting thermal dependence may
well lead to further hypotheses and insights into the nature of the contractile process
itself (e.g. Mittenthal, 1975, on the distance of cross-bridge movement). Muscle from
animals and/or animal groups with very different thermal histories could be
particularly useful in these studies. For example, the different response of Po to
temperature in anuran and mammalian muscle (Fig. 5) may be helpful to
understanding patterns of cross-bridge tension generation and cycling during tetany.
Or, the differing thermal dependencies of myofibrillar ATPase from animals adapted
to different temperatures (e.g. Fig. 4) may help elucidate their role in determining
intrinsic shortening velocities.
THERMAL DEPENDENCE OF MUSCLE PERFORMANCE IN VIVO
Is the thermal dependence of muscle function reflected in animal behavioural
capacities? On the basis of experiments on isolated muscle, one would expect
performance capacity for behaviour involving rates of contraction (e.g. running speed)
to improve with increasing temperature. One would also expect performance involving
maximal force generation by an animal to be relatively temperature independent.
Physical performance involving rate-dependent factors, such as maximal power
output, does improve significantly with increasing muscle temperature (e.g.
Asmussen & Bgje, 1945; Binkhorst, Hoofd & Vissers, 1977; Bergh & Ekblom, 1979).
Consequently, 'warming up', in the literal sense, does have a positive effect on
performance speed, and animals with higher body temperatures do in fact have
greater maximal speeds (Webb, 1978; Bennett, 1980; Putnam & Bennett, 1981).
However, the thermal dependence of this performance is distinctly lower than that of
rate processes in isolated muscle. For example, in the lizard Dipsosaums, maximal
running velocity and limb cycling frequency have Qlo values of 1.3-1.4 from 25 to
40°C, while Vma, and Wm, have Qlo values of 1.7 and 2.0, respectively (Fig. 6)
(Marsh & Bennett, 1985). Similarly low Qlo values of 1.0-1.6 have been reported in
other studies on rate-dependent performance cited above. This lower thermal
dependence of behavioural performance compared to that of isolated muscle function
could be attributable to several factors, including storage of energy in elastic
structures of low thermal sensitivity (Marsh & Bennett, 1985). However, its basis is
unknown at present.
Maximal force generation by muscles in vivo is almost independent of muscle
temperature from 25 to 40°C (Binkhorst et al. 1977; Bergh & Ekblom, 1979;
Petrofsky, Burse & Lind, 1981). This pattern accords very well with the observed
thermal independence of Po over this temperature range. Endurance, measured as the
time of maintenance of constant force, is maximal at approximately 30°C in both in
0 J, I I I I I I
15 25 35 45
Temperature ("C)
Fig. 6. The thermal dependence of burst locomotory speed and isotonic contractile performance
of isolated skeletal muscle of the lizard Dipsosaurus dorsalis. V,, maximal running velocity (filled
squares); f, limb cycling frequency (open squares); V,,, maximal velocity of shortening
(open circles); W,,,, maximal power output (filled circles). Data are normalized to maximal value
observed (V, = 4.3 ms-I; f = 13.5 s-'; V,,, = 20.1 lengths s-I; W,, = 505 W kg-'). Data from
Marsh & Bennett (1985).
Temperature and muscle 343
vivo performance (Clarke, Hellon & Lind, 1958; Petrofsky & Lind, 1969; Edwardset
al. 1972) and in isolated muscle (Petrofsky & Lind, 1981; Segal & Faulkner, 1982).
Endurance declines at both higher and lower temperatures.
As the foregoing data indicate, behavioural performance involving both speed and
force generation reflects the underlying patterns of thermal dependence of muscle
function. Higher muscle temperatures may be expected to result in improved rate
performance in vivo, although maximal force application may be little affected and
endurance may decline.
Financial support for this work was provided by NSF Grant PCM 81-02331. I
thank Roger Seymour for his helpful comments on the manuscript.
REFERENCES
ASMUSSEN, E. & B~JE, 0. (1945). Body temperature and capacity for work. Actaphysiol. scand. 10, 1-22.
BARANY, M. (1967). ATPase activity of myosin correlated with speed of muscle shortening. J. gen. Physiol. 50,
197-218.
BENDALL, J. R. (1964). The myofibrillar ATPase activity of various animals in relation to ionic strength and
temperature. In Biochemistry ofMuscle Contraction, (ed. J. Gergely), pp. 448-452. Boston: Little, Brown &
Co.
BENNEIT, A. F. (1980). The thermal dependence of lizard behaviour. Anim. Behav. 28, 752-762.
BENNEIT, A. F. (1984). The thermal dependence of muscle function. Am. J. Physiol. 247 (Reg. Integ. comp.
Physiol. 16), R217-R229.
BERGH, U. & EKBLOM, B. (1979). Influence of muscle temperature on maximal muscle strength and power
output in human skeletal muscle. Actaphysiol. scand. 107, 33-37.
BINKHORST, R. A., HOO~D, L. & VISSERS, A. C. A. (1977). Temperature and force-velocity relationship of
human musc1es.J. appl. Physiol. 42,471-475.
BLINKS, J. R., RODEL, R. & TAYLOR, S. R. (1978). Calcium transients in isolated amphibian skeletal muscle
fibres: detection with aequ0rin.J. Physiol., Lond. 277,291-323.
BRESSLER, B. H. (1981). Isometric contractile properties and instantaneous stiffness of amphibian skeletal
muscle in the temperature range of 0 to 20°C. Can. J. Physiol. Phannacol. 59, 548-554.
CLARKE, R. S. J., HELLON, R. F. & LIND, A. R. (1958). The duration of sustained contractions in the human
forearm at different muscle temperatures. J. Physiol., Lond. 143, 454-473.
EDWARDS, R. H. T., HARRIS, R. C., HULTMAN, E., KAIJSER, L., KOH, D. & NORDESJO, L.-0. (1972). Effect of
temperature on muscle energy metabolism and endurance during successive isometric contractions sustained
to fatigue, of the quadriceps muscle in man. J. Physiol., Lond. 220, 335-352,
FORD, L. E., HUXLEY, A. F. & SIMMONS, R. M. (1977). Tension responses to sudden length changes in
stimulated frog muscle fibres near slack length. J. Physiol., Lond. 269, 441-515.
GLEESON, T. T., PUTNAM, R. W. & BENNEIT, A. F. (1980). Histochemical, enzymatic, and contractile
properties of skeletal muscle fibres in the lizard Dipsosaums dorsa1is.J. exp. 2001. 214, 293-302.
HARTSHORNE, D. J., BARNS, E. M., PARKER, L. & FUCHS, F. (1972). The effect of temperatureonactomyosin.
Biochim. biophys. Acta 267, 190-202.
HILL, A. V. (195 1). The influence of temperature on the tension developed in an isometric twitch. Pmc. R. Soc. B
138,349-354.
HOMSHER, E., MOMMAERTS, W. F. H. M., RICCHIUTI, N. V. & WALLNER, A.(1972). Activation heat, activation
metabolism and tension-related heat in frog semitendinosus musc1es.J. Physiol., Lond. 220, 601-625.
JOHNSTON, I.A. (1979). Calcium regulatory proteins and temperature acclimation of actomyosin ATPase from a
eurythermal teleost (Carassius auratus L.). J. comp Physiol. 129, 163-167.
JOHNSTON, I.A. & WALBSBY, N. J. (1977). Molecular mechanisms of temperature adaptation in fish myofibrillar
adenosine triph0sphatase.J. comp. Physiol. 119, 195-206.
JOHNSTON, I. A. & WALESBY, N. J. (1979). Evolutionary temperature adaptation and the calcium regulation of
fish actomyosin ATPases.J. comp. Physiol. 129, 169-177.
JOHNSTON, I. A,, WALESBY, N. J., DAVISON, W. & GOLDSPINK, G.(1977). Further studies on the adaptation of
fish myofibrillar ATPases to different cell temperatures. PJiigers Arch. ges. Physiol. 371, 257-262.
JOSEPHSON, R. K. (1981). Temperature and the mechanical performance of insect muscle. In Insect Thennoregulation, (ed. B. Heinrich), pp. 19-44. New York: John Wiley & Sons.
344 A. F. BENNETT
KUHN, H. J., GOTH, K., DREXLER, B., BERBERICH, W. & ROEGG, J. C. (1979). lnvestigation of the temperature
dependence of the cross-bridge parameters for attachment, force generation and detachment as deduced from
mechano-chemical studies in glycerinated single fibers from the dorsal longitudinal muscle of Lethocerus
maxemus. Biophys. Struct. Mech. 6, 1-29.
LICHT, P. (1964). A comparative study of the thermal dependence of contractility in saurian skeletal muscle.
Comp. Biochem. Physiol. 13,27-34.
MARSH, R. L. & BENNETT, A. F. (1985). Thermal properties of isotonic contractile properties of skeletal muscle
and sprint performance of the lizard Dipsosaurus dorsalis. J. comp. Physiol. (in press).
MI~TENTHAL, J. E. (1975). A sliding filament model for skeletal muscle: dependence of isometric dynamics on
temperature and sarcomere length. J. theor. Bwl. 52, 1-16.
PETROFSKY, J. S., BURSE, R. L. & LIND, A. R. (1981). The effect of deep muscle temperature on the
cardiovascular responses of man to static effort. Eur. J. appl. Physiol. 47,7-16.
PETROFSKY, J.S. & LIND, A. R. (1969). Insulative power of body fat on deep muscle temperatures and isometric
endurance.J. appl. Physiol. 39,629-642.
PETROFSKY, J. S. & LIND, A. R. (1981). The influence of temperature on the isometric characteristics of fast and
slow muscle of the cat. Ppiigers Arch. ges. Physiol. 389, 149-154.
PRECHT, H., CHRISTOPHERSEN, J., HENSEL, H. & LARCHER, W. (1973). Temperature and Life. Berlin:
Springer-Verlag.
PROSSER, C. L. (1973). Comparative Animal Physiology, 3rd edn. Philadelphia: W. B. Saunders Co.
PUTNAM, R. W. & BENNE~, A. F. (1981). Thermal dependence of behavioural performance of anuran
amphibians. Anim. Behav. 29, 502-509.
PUTNAM, R. W. & BENNETT, A. F. (1982). Thermal dependence of isometric contractile properties of lizard
musc1e.J. comp. Physiol. 147, 11-20.
PUTNAM, R. W., GLEESON, T. T. & BENN~, A. F. (1980). Histochemical determination of the fiber
composition of locomotory muscles in a lizard, Dipsosaurus dorsalis. J. exp. Zool. 214, 303-309.
RALL, J. R. (1979). Effects of temperature on tension, tension-dependent heat, and activation heat in twitchesof
frog skeletal muscle. J. Physiol., Lond. 291, 265-275.
RANATUNGA, K.W. (1977). Influence of temperature on the characteristics of summation of isometric
mechanical responses of mammalian skeletal muscle. Expl Neuml. 54, 513-532.
RENAUD, J. M. & STEVENS, E. D. (1981~). Effect of acclimation temperature and pH on contraction of frog
sartorius muscle. Am. J. Physiol. 240 (Reg. Znteg. comp Physiol. 9), R301-R309.
RENAUD, J. M. & STEVENS, E. D. (1981b). The interactive effects of temperature and pH on the isometric
contraction of toad sartorius muscle. J. comp. Physiol. 145, 67-71.
ROME, L. C. (1983). The effect of long-term exposure to different temperatures on the mechanical performance
of frog muscle. Physiol. Zool 56, 33-40.
SALTIN, B., GAGGE, A. P. & STOLWIJK, J. A. J. (1968). Muscle temperature duringsubmaximal exercise in man.
J. appl. Physiol. 25,679-688.
SEGAL, S. S. & FAULKNER, J.A. (1982). Static endurance of rat skeletal muscles at different temperatures in
vitm. Med. Sci. Sports Exer. 14, 106.
SIDELL, B. D., JOHNSTON, I. A., MOERLAND, T. S. & GOLDSPINK, G. (1983). The eurythermal myofibrillar
protein complex of the mummichog (Fundulus hetemclitus) : adaptation to a fluctuating thermal environment.
J. comp. Physiol. 153, 167-173.
USHAKOV, B. (1964). Thermostability of cells and proteins of poikilothems and its significance in speciation.
Physiol. Rev. 44,518-560.
WALKER, S. M. (1960). The relation of stretch and of temperature to contraction of skeletal muscle. Am.J.phys.
Med. 39, 234-258.
WEBB, P. W. (1978). Temperature effects on acceleration of rainbow trout, Salmogairdneri. J. Fish. Res. Bd
Can. 35, 1417-1422.
YAMAMOTO, T.& TONOMURA, Y.(1967). Reaction mechanism of the Caz+-dependent ATPase of sarcoplasmic
reticulum from skeletal muscle. J. Biochem., Tokyo 62, 558-575. | biology | 4079561 | https://sv.wikipedia.org/wiki/Stenospermation%20arborescens | Stenospermation arborescens | Stenospermation arborescens är en kallaväxtart som beskrevs av Michael T. Madison. Stenospermation arborescens ingår i släktet Stenospermation och familjen kallaväxter. IUCN kategoriserar arten globalt som starkt hotad.
Artens utbredningsområde är Ecuador. Inga underarter finns listade.
Källor
Kallaväxter
arborescens | swedish | 1.333338 |
below_temperature_muscles_not_work/japplphysiol01107200.txt |
PHYSIOLOGY.ORG SKIP MAIN NAVIGATION
JOURNALS
All journals
Quick Search anywhere
Search all content
Search all content
Advanced Search
SIGN IN
Journal of Applied PhysiologyVol. 94, No. 6
Journal of Applied Physiology Logo
JOURNAL HOME
ISSUES
COLLECTIONSINFORMATIONABOUT
SUBMITSUBSCRIBE
QUICK SEARCH IN JOURNALS
SEARCH THIS JOURNAL
Search this journal
ARTICLE
Muscle temperature transients before, during, and after exercise measured using an intramuscular multisensor probe
G. P. Kenny,F. D. Reardon,W. Zaleski,M. L. Reardon,F. Haman, andM. B. Ducharme
01 JUN 2003https://doi.org/10.1152/japplphysiol.01107.2002
This is the final version - click for previous version
Sections
PDF (216 KB)
TOOLS
SHARE
Abstract
Seven subjects (1 woman) performed an incremental isotonic test on a Kin-Com isokinetic apparatus to determine their maximal oxygen consumption during bilateral knee extensions (V˙o 2 sp). A multisensor thermal probe was inserted into the left vastus medialis (middiaphysis) under ultrasound guidance. The deepest sensor (tip) was located ∼10 mm from the femur and deep femoral artery (Tmu 10), with additional sensors located 15 (Tmu 25) and 30 mm (Tmu 40) from the tip. Esophageal temperature (Tes) was measured as an index of core temperature. Subjects rested in an upright seated position for 60 min in an ambient condition of 22°C. They then performed 15 min of isolated bilateral knee extensions (60% of V˙o 2 sp) on a Kin-Com, followed by 60 min of recovery. Resting Tes was 36.80°C, whereas Tmu 10, Tmu 25, and Tmu 40 were 36.14, 35.86, and 35.01°C, respectively. Exercise resulted in a Tes increase of 0.55°C above preexercise resting, whereas muscle temperature of the exercising leg increased by 2.00, 2.37, and 3.20°C for Tmu 10, Tmu 25, and Tmu 40, respectively. Postexercise Tes showed a rapid decrease followed by a prolonged sustained elevation ∼0.3°C above resting. Muscle temperature decreased gradually over the course of recovery, with values remaining significantly elevated by 0.92, 1.05, and 1.77°C for Tmu 10, Tmu 25, and Tmu 40, respectively, at end of recovery (P < 0.05). These results suggest that the transfer of residual heat from previously active musculature may contribute to the sustained elevation in postexercise Tes.
a number of studies have examined muscle temperature (Tmu) profiles for resting conditions under different thermal conditions (4, 7, 8, 17, 20-22, 24, 27, 30, 31). The study by Ducharme and Tikuisis (8), however, was the only study to present a mean Tmu profile for a group of subjects, as opposed to single-depth measurements. Despite the large number of studies, there is still no consistent description of resting Tmu profile. There also have been a number of studies that have reported Tmu response during exercise (1, 2, 5,6, 19, 23-27), of which the study by Saltin et al. (25) seems to be the only one to examine changes in Tmu profile (i.e., Tmu measured at multiple depths). Although, these experiments were not designed to show the time course change in tissue temperature gradients, their measurement of individual intramuscular temperature response did show large variations in the temperature at the superficial, mid-, and deep muscle sites. Furthermore, they showed significant variation in the rates of temperature change during muscle activity.
There are no studies that have examined changes in Tmuduring the postexercise period. Several have reported postexercise Tmu response (1, 23, 25); however, none has specifically addressed these responses. In short, there remains a lack of information regarding the kinetics of heat exchange between muscle and the core of the body and within a given mass of muscle tissue. This information is critical to our understanding of the underlying mechanism responsible for the sustained, postexercise elevation in core temperature. In previous work, core temperature has been shown to remain elevated by ∼0.4°C for a prolonged period after cessation of exercise (16, 28) performed in different thermal environments. However, the actual mechanism responsible for this increase in core temperature remains unresolved.
Tissue temperature at any given time is ultimately determined by the relative rates of heat production and heat loss. For example, regional Tmu at any point in time is the result of regional differences in metabolic rate (9), conductive heat loss to adjacent tissue (9, 10), and deep and peripheral convective blood flow (9, 29). As such, it would be expected that both regional temperature profile and the rate of temperature change would differ during resting, exercise, and postexercise recovery. The following study was designed to measure intramuscular temperature profile during rest, exercise, and postexercise recovery. In contrast to the findings of previous studies, we hypothesized that the tissue temperature profile will be consistent among subjects as the probe position is standardized within the muscle of all subjects. Furthermore, in conjunction with a postexercise decrease in heat loss, subsequent to a decrease in skin blood perfusion, and an attenuation of sudomotor activity during the postexercise recovery (14), we hypothesized that convective heat transfer between muscle and core will significantly influence postexercise core temperature response.
METHODS
Subjects.
Subsequent to approval of the project by the University Human Research Ethics Committee, seven healthy subjects (6 men, 1 woman) consented to participate in the study. Mean values (±SD) of the subjects' age, height, body mass, maximal oxygen consumption during bilateral concentric knee extensions (V˙o 2 sp), and body fat content were 24 ± 5 yr, 1.8 ± 0.5 m, 85.6 ± 6.1 kg, 2.1 ± 0.9 l/min, and 10.9 ± 2.3%, respectively.
In each trial, esophageal temperature (Tes) was measured by using a thermocouple temperature probe (Mallinckrodt Medical) inserted through a nostril, into the esophagus, to the level of the heart. Regional Tmu of the vastus medialis was measured by using a flexible multithermocouple temperature probe (Physitemp Instruments, Clifton, NJ; model IT-17:3) inserted into the vastus medialis. Ultrasound imaging was used to discern the best perpendicular insertion tract and subsequently to place the probe at a position 10 mm and equidistant from the deep femoral artery and the femur. The implant site was approximately midway between, and medial to, a line joining the anterior superior iliac spine and the superior aspect of the center of the patella.
With the use of aseptic technique and under ultrasound guidance, the skin, subcutaneous tissue, and muscle were anesthetized to a maximum depth of 50 mm by infiltrating ∼2 ml of 1% lidocaine without epinephrine. The tip of this 25-gauge needle was placed at the proposed site for the deep temperature probe. Under full ultrasound imaging and with the use of the anesthetic needle as a guide, an 18-gauge, 50-mm polyethylene catheter (Cathlon and Critikon Canada, Markham, Ontario) was then inserted into the anesthetized tract to the required depth. The anesthetic needle and the catheter stylet were then withdrawn, and the temperature probe was inserted in the catheter shaft. When the probe was fully inserted, the catheter was carefully withdrawn, leaving the tip of the temperature probe ∼10 mm from the femur and deep femoral artery. Once the catheter was withdrawn, the final position of the probe was verified by using the ultrasound imaging. The average depth of the probe from the surface was 47.2 mm and within 11.0 mm of both the deep femoral artery and femur. The probe assembly was secured to the skin with sterile, waterproof transparent dressing (3M 1622W Tegaderm transparent dressing) and tape (total surface coverage ∼25 cm2). The Tegaderm transparent dressing consists of a thin polyurethane membrane coated with a layer of an acrylic adhesive. The dressing, which is permeable to both water vapor and oxygen, is impermeable to microorganisms, and, once in position, it provides an effective barrier to external contamination.
The temperature probe was a sterile Teflon-coated multisensor probe (model IT-17:3, Physitemp Instruments; thermal constant of 0.25 s). Each probe had three thermocouples: one positioned at the tip, one at 15 mm, and the third at 30 mm from the tip. The deepest temperature sensor (tip) was located ∼10 mm from the femur and deep femoral artery (Tmu 10), with two sensors located at 15 (Tmu 25) and 30 mm (Tmu 40) from the tip (Table 1). The internal position of the temperature sensor relative to the skin surface was calculated based on the ratio of the known depth of the probe (radius r) from the skin surface measured by ultrasound imaging and the radius of the thigh (r sk). Thusr/r sk is the relative radius (8). Although it was not possible to verify the final position of the probe after the completion of the experimental trial, the length of the probe within the limb tissue was measured during the removal of the probe. The depth of the probe was verified with the preexperiment depth, as determined by ultrasound imaging.
Enlarge table
Table 1. Mean (±SD) and individual data relating to the placement of the intramuscular multisensor thermal probe of the upper leg
Enlarge table
Skin temperature was monitored at 12 sites by using type T thermocouples integrated into heat-flow sensors (Concept Engineering, Old Saybrook, CT). The area-weighed mean skin temperature (T̄sk) and heat flux (H̄Fsk) were calculated by assigning the following regional percentages: 6% head, 9% upper arm, 6% forearm, 2% finger, 19% chest, 9.5% upper back, 9.5% lower back, 10% anterior thigh, 10% posterior thigh, 9.5% anterior calf, and 9.5% posterior calf (12). Temperature and heat flux data were collected and digitized (Hewlett Packard data-acquisition module, model 3497A) at 5-s intervals and simultaneously displayed and recorded in spreadsheet format on a hard disk (Hewlett Packard, model PC-312, 9000).
Oxygen consumption (V˙o 2) was determined by open-circuit analysis by using an automated gas-collection system (Quinton Instrument, Seattle, WA; model Q-Plex 1 cardiopulmonary exercise system). Skin blood flow was measured by laser-Doppler velocimetry (PeriFlux System 5000, main control unit; PF5010 LDPM, operating unit; Perimed, Stockholm, Sweden) from the left midanterior forearm. The laser-Doppler flow probes (PR 401 angled probe, Perimed) were taped to cleaned skin, in an area that superficially did not appear to be highly vascular and from where consistent readings were noted (18). Sweat rate was estimated from a 5.0-cm2 ventilated capsule placed on the upper back. Anhydrous compressed air was passed through the capsule over the skin surface at a rate of 1 l/min. Water content of the effluent air was measured at known barometric pressure by using the readings from an Omega HX93 humidity and temperature sensor (Omega Engineering, Stamford, CT). Sweat rate was calculated from the product of the difference in water content between effluent and influent air and the flow rate. This value was normalized for the skin surface area under the capsule and expressed in milligrams per minute per centimeter squared.
Subjects performed an incremental isotonic test (constant angular velocity, increases in force output) on the Kin-Com isokinetic apparatus to determine their V˙o 2 sp. The exercise consisted of bilateral, concentric knee extension over a range of 70° from perpendicular, with the subject sitting (hip angle between 90 and 110°) and the long axis of the thigh in the horizontal plane. The force output was increased by 15 N every 2 min until fatigue, whereas the angular velocity was maintained at 58.3°/s throughout the test. The results of the test were used to establish the work rate for the experimental trial. The experimental trial was conducted in the morning after a 24-h period without heavy or prolonged physical activity. On arrival at the laboratory at 0800, subjects were appropriately instrumented. Subjects then rested in a semirecumbent position for 60 min at an ambient temperature of 22°C, of which the final 20 min were recorded as representative of the baseline resting values. At 2 min before exercise, the subjects were secured to the Kin-Com isokinetic apparatus at the level of the torso and ankles. Subjects then performed 15 min of exercise, as described above, consisting of bilateral, concentric knee extension over a range of 70° from perpendicular against a dynamic resistance sufficient to elicit a heat load of 4.78 kJ/kg. Exercise was followed by 60 min of seated rest.
The total energy expended (Mtotal) as a result of exercise, during the period from onset of exercise until the time at whichV˙o 2 returned to preexercise values, was calculated from the sum of the energy expended by using the following equation (expressed in kJ)
𝑀total=∑𝑀˙Ex/rest
Equation 1
=∑{𝑉˙𝑂2·[((RER−0.7)0.3𝑒𝑐)+((1−RER)0.3𝑒𝑓)]}
where M˙Ex/rest is the rate of energy expenditure during exercise and recovery, RER is the respiratory exchange ratio, ec is the caloric equivalent (in kJ/l O2) for carbohydrates, and ef is the caloric equivalent (in kJ/l O2) for fat.
The minute values were summed for the entire period as described above.
The mechanical work (W) done during each contraction of the exercise phase was measured and recorded by using the Kin-Com isokinetic machine. This was calculated from the force exerted and the angular displacement during the knee extension
𝑊=𝑇𝜃
Equation 2
where T is rotational force or torque and θ is the angular displacement.
The total work done (Wtotal) was the sum of the work accomplished during each of the contractions during the 15 min of exercise.
Mechanical efficiency (ME) was defined as the Wtotalcompleted during the 15-min exercise period divided by the Mtotal minus the energy expended under resting conditions (Mrest) (Mtotal − Mrest). Thus
ME=𝑊total𝑀total−𝑀rest
Equation 3
The Mrest was calculated from the average rate ofV˙o 2 during the 5 min preceding the exercise bout. These values were calculated and expressed in kilojoules by using the aforementioned equation.
The total heat load generated by the exercise (HLex) for each subject was calculated by subtracting the Mrest and the energy equivalent of the total mechanical work done (Wtotal) from the Mtotal. Values are expressed in kilojoules
HLex=𝑀total−(𝑀rest+𝑊total)
Equation 4
The total dry heat loss by radiation, conduction, and convection from the skin surface during exercise (Hsk ex) and during recovery (Hsk rec) was estimated by subtracting the area-weighted H̄Fsk (as above) corrected for body surface area (AD) during rest from those values recorded during exercise and recovery, respectively. Thus
𝐻skex=(𝐻̄ 𝐹sk·𝐴𝐷)ex−(𝐻̄ 𝐹sk·𝐴𝐷)rest
Equation 5
and
𝐻skrec=(𝐻̄ 𝐹sk·𝐴𝐷)rec−(𝐻̄ 𝐹sk·𝐴𝐷)rest
Equation 6
where Hsk ex and Hsk rec are the total dry heat lost during the 15-min exercise and 60-min recovery periods, respectively.
Statistical analyses for Tes, Tmu,T̄sk, and H̄Fsk were performed by ANOVA for repeated measures to compare values for preexercise, end-exercise, and at 10-min intervals during postexercise recovery. Data are presented as means ± SD.
RESULTS
Baseline Tes and T̄sk were 36.80 ± 0.30 and 31.66 ± 0.89°C, respectively. Resting Tmu was significantly lower than Tes (i.e., 36.14 ± 0.29, 35.86 ± 0.31, and 35.01 ± 0.33°C for Tmu 10, Tmu 25, and Tmu 40, respectively) (Fig. 1). It should be noted that the increase in muscle tissue temperature before the onset of exercise was likely due to the preparation of the subject for the exercise portion of the experimental trial.
Fig. 1.
Fig. 1.
Mean (±SE) muscle [temperature sensor located 10 mm (Tmu 10; □), 15 mm (Tmu 25; ○), and 30 mm (Tmu 40; ▿) from femur and deep femoral artery] and esophageal (◊) temperature response during rest, exercise (Ex), and postexercise recovery. Vertical dotted lines represent the start (time = 0 min) and end (time = 15 min) of exercise. * Significant difference from baseline resting values,P < 0.05. 1 Value for esophageal temperature not significantly elevated from baseline.
Download figureDownload PowerPoint
The Tmu profiles, expressed as a function of the position of the placement of the temperature relative to the radius of the thigh (r/r sk), show a parabolic profile for mean resting tissue temperature profile (Fig.2). This parabolic form of Tmu profile was observed consistently in the data of all seven subjects (Fig. 3). During resting, the deep (36.14°C) and mid-Tmu (35.86°C) were significantly different from superficial (35.01°C) Tmu. As depicted in Fig. 4, the greatest tissue temperature difference (1.13°C) was between the deep and superficial sections of the muscle, with a 0.84°C temperature gradient between the mid- and superficial muscle. Mean tissue temperature difference between deep and midmuscle was only 0.28°C. Furthermore, the muscle-to-core temperature gradient was equal to −0.66, −0.94, and −1.79°C in relation to Tmu 10, Tmu 25, and Tmu 40, respectively (P < 0.05).
Fig. 2.
Fig. 2.
Mean (±SE) muscle temperature profile during baseline resting (●), Ex (□), end-exercise (End Ex; ■), and postexercise (post-Ex; ▵) recovery at selected periods as a function of the placement of the temperature sensors relative to the radius of the thigh. r, Radius (cm); r sk, radius of the thigh (cm);r/r sk, relative radius.
Download figureDownload PowerPoint
Fig. 3.
Fig. 3.
Mean (±SE) (●) and individual muscle temperature profiles during resting (A), end-exercise (B), and at 60 min postexercise resting (C). Note: individual subjects are represented by different symbols, and these symbols are the same for each time period inA–C. § Significantly different from superficial muscle, P < 0.05.
Download figureDownload PowerPoint
Fig. 4.
Fig. 4.
Mean (±SE) core-to-muscle temperature gradient (○, deep muscle to core; □, midmuscle to core; ▵, superficial muscle to core) (A) and intramuscular temperature gradients (✳, deep muscle to midmuscle; ◊, deep muscle to superficial muscle; ▿, midmuscle to superficial muscle) (B). Vertical dotted lines represent the start (time = 0 min) and end (time = 15 min) of exercise. P < 0.05: ‡ significantly different from the deep to midmuscle temperature gradient; § significantly different from baseline resting; * superficial-to-core temperature gradient significantly different from deep-to-core temperature gradient.
Download figureDownload PowerPoint
Exercise tissue temperature response.
After the onset of exercise, Tes increased gradually, reaching a maximum rate of increase of 0.05 ± 0.02°C/min between 6 and 9 min of exercise, and, subsequently, the rate decreased over the balance of the exercise period (Table2). In contrast, Tmu at all measured points increased rapidly during the initial period of exercise followed by a gradual reduction in the rate over the balance of the exercise period. Superficial muscle (Tmu 40) showed the greatest rate of temperature increase (0.61 ± 0.19°C/min). This value was significantly higher than the rate measured in deep muscle (0.22 ± 0.09°C/min). After the initial 3 min of exercise, the rate of Tmu change decreased gradually and was similar at all three intramuscular sites until the end of exercise. Exercise resulted in a 0.55°C (end-exercise Tes of 37.35°C) increase in core temperature above baseline resting values. In contrast, Tmu increased by 2.09, 2.37, and 3.20°C above baseline resting values for Tmu 10, Tmu 25, and Tmu 40 measurement points, respectively, with end-exercise values similar at all three measured sites (i.e., 38.23, 38.23, and 38.21°C for Tmu 10, Tmu 25, and Tmu 40, respectively). T̄sk increased continuously during exercise to an end-exercise value that was significantly elevated above baseline rest (32.79°C,P < 0.05). The increase in T̄sk was paralleled by an increase in nonevaporative heat loss (i.e., Fig.5). Forearm skin blood flow increased continuously during the course of the exercise.
Enlarge table
Table 2. Mean (±SD) rate of change of esophageal and muscle (Tmu 10, Tmu 25, and Tmu 40) temperatures during exercise and postexercise recovery
Enlarge table
Fig. 5.
Fig. 5.
Mean (±SD) heat load (⊙) and dry heat loss (⟐) responses during baseline resting, exercise, and postexercise recovery. Vertical dotted lines represent the start (time = 0 min) and end (time = 15 min) of exercise.
Download figureDownload PowerPoint
As depicted in Fig. 2, the tissue temperature profile evolved from a parabolic form typical of resting to a linear profile during the early stages of exercise. By the end of exercise, Tmu values across the radial axis were homogenous. As such, the large temperature gradient between the deep and superficial muscle was rapidly reduced such that, by the end of exercise, the temperature at all sites was similar. Furthermore, the muscle-to-core temperature gradient was reversed from resting, such that Tmu values at all sites were significantly elevated above Tes by 0.90, 0.90, and 0.89°C for Tmu 10, Tmu 25, and Tmu 40, respectively (P < 0.05).
Postexercise tissue temperature response.
Tes decreased rapidly (−0.04°C/min) during the initial minutes after the cessation of exercise (Table 2), after which there was a rapid decrease in the rate of temperature decrease to negligible values. At ∼5 min of recovery, Tes reached an elevated value 0.35°C above baseline resting values (P < 0.05). For the duration of recovery, the rate of decrease of Tes remained at ∼0.001°C/min. Tes decreased to 37.11°C by the end of the 60-min recovery period (∼0.3°C above baseline). Tmu showed a similar high rate of temperature decrease during the initial 5 min of exercise recovery, although the rates were ∼2 to 2.7 times greater than the rate measured for Tes. Unlike the response in Tes, Tmu for all measured sites decreased continuously during the initial 30 min of recovery. However, the rates of Tmudecay were reduced for the duration of recovery. In the final 15 min of recovery, superficial muscle demonstrated an elevated rate of temperature decrease above deep muscle (P < 0.05). Muscle tissue temperature at the end of the postexercise recovery period remained significantly elevated above baseline resting values by 0.92, 1.05, and 1.77°C for Tmu 10, Tmu 25, and Tmu 40, respectively (P <0.05) (Fig.1). T̄sk and whole body nonevaporative heat loss decreased to baseline resting values within ∼20–25 min of recovery. Similarly, forearm skin blood flow decreased to baseline resting values within 10 min of the termination of the exercise. In contrast, both thigh nonevaporative heat loss and thigh skin temperature remained significantly elevated from preexercise values for the duration of the recovery period (P < 0.05).
The temperature gradient between the different depths of muscle remained relatively unchanged during the postexercise recovery period, despite a slow decay in Tmu. The muscle-to-core temperature gradient decreased gradually over the course of the recovery. At ∼25 min into recovery, Tmu at all depths achieved similar values to that of Tes. For the duration of the recovery period, the muscle-to-core temperature gradient was increased, with superficial muscle demonstrating the largest temperature gradient by the end of recovery (∼0.3°C; P < 0.05) compared with that temperature gradient between deep muscle and core. Of note, the deep muscle-to-core temperature gradient remained relatively unchanged for the duration of the recovery period (∼0.02°C).
Heat load and heat loss response.
The workload resistance was adjusted for each subject, according to the individual heat load and mechanical efficiency of the leg-extension exercise. The mechanical efficiency varied from 5.98 to 15.96%, whereas the average for the group was 9.93 ± 1.32%. The average actual workload was one at which the heat production was 4.78 ± 0.38 kJ/kg. The Mtotal and Wtotal were 585.87 ± 53.72 and 41.48 ± 4.68 kJ, respectively. Thus the average heat load generated as a result of the exercise was 391.73 ± 38.93 kJ. The average total dry heat loss during the exercise period was 15.93 ± 5.98 kJ, whereas, during the 60 min of postexercise recovery, this value was 37.90 ± 18.80 kJ.
The kinetics of heat load generation at rest, over the 15 min of exercise, and over the first 10 min of recovery are presented in Fig.5. The corresponding evolution of dry heat loss is also shown. Thus during the 5 min preceding exercise, the dry heat loss defined relative to heat load, that is, minus the resting levels, was essentially zero. During exercise, the dry heat loss increased at a rate of 0.14 kJ/min to a maximal level of 2.19 kJ/min after 15 min of exercise. At cessation of exercise, the sensitive heat loss dropped exponentially to a level ∼1.0 kJ/min above initial resting values and remained elevated for the next 10 min. The heat production, on the other hand, increased to 17.08 kJ/min after 2 min of exercise and continued to rise at a rate of ∼0.78 kJ/min for the next 13 min to a maximum heat production of 27.71 kJ/min. Immediately postexercise, the heat load returned exponentially to resting levels within 5 min.
DISCUSSION
In this study, an attempt was made to specifically evaluate the kinetics of heat exchange in muscle tissue during and after exercise by using a multisensor thermal probe positioned at a predetermined internal marker. In contrast to previous studies, we observed similar individual and group Tmu profiles during resting, exercise, and subsequent resting recovery. Furthermore, we observed a sustained elevation of core temperature for the duration of the recovery period that is consistent with previous findings (16, 28). Specifically, Tes showed a rapid decrease in the first minutes of exercise recovery followed by a prolonged sustained elevation of ∼0.3°C. Of particular importance was the observation that deep Tmu decreased during the early stages of exercise recovery to values equal to that of Tes. Subsequently, deep Tmu remained relatively unchanged from Tes for the duration of recovery. This supports the hypothesis that the postexercise recovery of core temperature may be, to a large degree, influenced by the residual heat load of muscle.
Tissue temperature response: resting.
Different shapes of limb temperature profile have been reported for resting conditions during different thermal stresses between individuals, whereas we noted a consistent parabolic profile of Tmu in all subjects (4, 7, 8, 17, 20-22, 24, 27,31). This could arise from inconsistency in the specific placement of the internal probe in the muscle. There is a wide variation in recorded muscle tissue temperature due to the proximity of the probe to the surface and to such structures as large arteries and bone (10). This could have been a source of significant variation in the recorded internal temperature. The differences in the specific heat of these tissues, as well as the differing blood flow and hence the convective effect within these structures, influence the rates of temperature change in adjacent regions of the muscle. Therefore, consistent placement of the probe is critical. Thus an attempt was made to minimize the variation in temperature recording resulting from individual anatomic differences.
Tmu response: exercise.
From the onset of exercise until the late phases of exercise, there is a gradual change in the Tmu profile from one parabolic in the form seen at rest to a zero gradient or homogenous temperature profile across the muscle. As shown in Fig. 4, the large temperature gradient that existed between the deep and midportions of the muscle and the superficial muscle was rapidly eliminated during the first 5 min of exercise. By the end of exercise, the gradient between the different muscle depths was nonexistent, as temperatures across the radial axis of the muscle became homogenous. This effect was not observed earlier by Saltin et al. (24). In that case, it was noted that both mid- and superficial Tmu remained generally lower than deep Tmu. Similarly, superficial Tmu remained lower than mid-Tmu, whereas the temperature gradient between mid- and deep muscle seemed to remain relatively constant throughout exercise. The gradient between the superficial muscle increased relative to that between mid- and deep muscle. The different response to that observed in our study may be attributed to a number of factors, which may include the following: 1) differences in the measurement site (i.e., vastus lateralis); 2) differences in the muscle mass implicated in the exercise activity; 3) differences in ambient conditions; and 4) difference in work intensity. For example, it would be expected that the temperature profile of the vastus lateralis would be different from that of the medialis, as it is less affected by the circulation in the femoral artery and vein. The relative influence of convective heat exchange would be considerably different between these muscles.
The high rate of Tmu increase in the early stages of exercise is consistent with previous studies (1, 3, 24,25). Aulick et al. (3) showed, at the beginning of exercise, that heat gained by the leg (local metabolic heat production plus vascular heat delivery from the viscera) exceeded heat loss, and femoral vein blood temperature rose rapidly. In this study, the superficial regions of the muscle demonstrated the largest rate of Tmu increase (0.53°C/min) that was 1.4 and 1.6 times the rate of temperature increase for the deep and midmuscle, respectively. On the other hand, based on visual observation of the work by Saltin and coworkers (24), it would seem that the rate of temperature increase was greater in deep muscle compared with superficial muscle.
During the course of exercise, the muscle-to-core temperature gradient increased progressively (Fig. 4), from −1.15°C at rest to +0.90°C by the end of exercise. Also, despite the rapid increase in muscle heat content (as represented by increased Tmu) to values exceeding that for core, the rate of temperature increase of core remained consistently lower than that of muscle. Therefore, this would suggest that the rate of heat accumulation within the core region is attenuated to a large degree by an increase in the rate of whole body heat loss (i.e., evaporative and nonevaporative heat loss). For example, Aulick et al. (3) previously noted that, as limb sweat rate, cutaneous blood flow, and muscle-to-skin temperature differences increased during exercise, the active leg became a more effective vehicle for heat dissipation, and that femoral venous temperature eventually reached a plateau during steady state. Furthermore, Gisolfi and Robinson (11) showed that much of the heat produced by active leg muscles is rapidly transported to surface veins and that this muscle heat is potentially lost across the leg surface. In this study, muscle-to-skin temperature gradient remained elevated during the course of the exercise by ∼5.2°C, and skin blood flow and sweat rate increased gradually during the course of the exercise. Furthermore, it has previously been shown that, during leg work, the inactive upper limbs also act as an avenue for vascular heat loss from the central circulation (15), which would further attenuate the increase in core temperature.
Tmu response: postexercise.
Few studies have graphically presented muscle tissue temperature response during the postexercise period, and, even so, no specific discussion was presented with regard to these data (1, 23,25). It is clear in this study that, during the transition from exercise to postexercise resting recovery, the Tmu profile across the radial axis of the muscle remains constant (i.e., linear profile, see Figs. 2 and 3). During the course of the 60-min recovery, all three sites showed a similar rate of temperature change, although superficial muscle showed a significantly greater rate of temperature decrease toward the later stages of recovery (P < 0.05).
Deep Tmu decreased during the early stages of exercise recovery to values equal to that of Tes, after which deep Tmu remained relatively unchanged from Tes for the duration of recovery, with the deep muscle-to-core temperature gradient no greater than ∼0.02°C. The lack of a difference in temperature gradient between muscle and core suggests equilibration of heat distribution within the body. Thus changes in surface heat loss (i.e., evaporative and nonevaporative heat loss) will change the rate of whole body cooling. Therefore, the rate of core temperature decay is limited by the rate at which heat is lost at the skin-air interface.
All Tmu values remained significantly elevated above baseline resting values at the end of recovery. That was paralleled by a significant increase in Tes of ∼0.3°C (P < 0.05). Aikas et al. (1) have shown a similar postexercise increase in Tmu of the previously active muscle, although Tes showed a rapid decrease to values below baseline rest within a short period (∼15 min) after cessation of exercise. This is more difficult to explain if one believes that the convective arterial flow plays a major role in muscle cooling postexercise. Our results are more consistent with those of Saltin et al. (25), who, on the other hand, did observe a sustained increase in postexercise Tes, whereas Tmu remained significantly elevated above baseline resting values.
Thoden et al. (28) previously showed a prolonged postexercise elevation (0.4–0.5°C) in Tes after dynamic exercise. It was subsequently shown that an increase in the postexercise hypotensive response, induced by exercise of increasing intensity, was paralleled by an increase (∼0.4°C) in the magnitude of the postexercise elevation in Tes (13). It was suggested that the postexercise Tes response may be defined to a large degree by the gradient between the periphery and core and that the convective transfer of residual heat from previously active musculature may contribute to the sustained elevation in postexercise Tes. Our observation of a prolonged elevation in Tmu at values elevated above esophageal provides further evidence to support this conclusion. Thus, in the absence of a postexercise increase in heat loss response (14), Tes would remain elevated as long as 1) the heat content of muscle remains higher or equal to that of core;2) the postexercise hypotensive effect persists; or3) a combination of the two.
Summary.
In the present study, exercise was performed such that the dynamic resistance during the bilateral knee-extension exercise was sufficient to elicit a heat load of 4.78 kJ/kg. Thus it can be assumed that the rate of heat production and accumulation in muscle was comparable between subjects. Thus the variation in Tmu profile observed between the transition from rest to exercise and exercise to resting recovery was not only the result of the change in metabolic heat production but also the result of changes in the convective heat transfer between blood and muscle and conductive heat transfer within the muscle and skin surface. Furthermore, as with previous studies that have shown that tissue heat content and compartmental heat exchange are significantly influenced by convective heat exchange during rest (9) and exercise (15), our findings suggest that postexercise core temperature response (and the rate of temperature decay) is significantly influenced by convective heat transfer between muscle and core.
We acknowledge the technical support of Carolyn Proulx and Normand Boulé.
This research was supported by the Natural Sciences and Engineering Research Council of Canada (to G. P. Kenny).
FOOTNOTES
Address for reprint requests and other correspondence: G. P. Kenny, School of Human Kinetics, Univ. of Ottawa, Rm. 372, Montpetit Hall, P.O. Box 450 Station A, 125 Univ., Ottawa, Ontario, Canada K1N 6N5 (E-mail:[email protected]).
The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
First published February 21, 2003;10.1152/japplphysiol.01107.2002
Download PDF
PreviousBack to TopNext
Figures
References
Related
Information
Journal of Applied Physiology 94 6 cover image
Volume 94Issue 6
June 2003
Pages 2350-2357
Copyright & Permissions
Copyright © 2003 the American Physiological Society
https://doi.org/10.1152/japplphysiol.01107.2002
PubMed12598487
History
Received 3 December 2002
Accepted 7 February 2003
Published online 1 June 2003
Published in print 1 June 2003
Keywords
heat loadthermoregulationhyperthermiaheat contentheat balance
Metrics
Downloaded 4,154 times
Article has an altmetric score of 2
See more details
Posted by 1 X users
On 1 Facebook pages
117 readers on Mendeley
103103 total citations on Dimensions.
103 Total citations
14 Recent citations
23 Field Citation Ratio
2.72 Relative Citation Ratio
Publications
110
Supporting
4
Mentioning
91
Contrasting
3
Smart Citations
110
4
91
3
Citing Publications
Supporting
Mentioning
Contrasting
View Citations
See how this article has been cited at scite.ai
scite shows how a scientific paper has been cited by providing the context of the citation, a classification describing whether it supports, mentions, or contrasts the cited claim, and a label indicating in which section the citation was made.
We recommend
Postexercise hypotension causes a prolonged perturbation in esophageal and active muscle temperature recovery
Glen P. Kenny et al., American Journal of Physiology - Regulatory, Integrative and Comparative Physiology, 2006
Metabolic heat production during fatigue from voluntary repetitive isometric contractions in humans
E. Saugen et al., Journal of Applied Physiology, 1996
Effects of cold water immersion and active recovery on hemodynamics and recovery of muscle strength following resistance exercise
Llion A. Roberts et al., American Journal of Physiology - Regulatory, Integrative and Comparative Physiology, 2015
Acute head-down tilt decreases the postexercise resting threshold for forearm cutaneous vasodilation
Glen P. Kenny et al., Journal of Applied Physiology, 2000
Postexercise protein metabolism in older and younger men following moderate-intensity aerobic exercise.
M Sheffield-Moore et al., American Journal of Physiology - Endocrinology and Metabolism, 2004
Inhibiting the inhibitors: Development of the IAP inhibitor xevinapant for the treatment of locally advanced squamous cell carcinoma of the head and neck
Robert L. Ferris et al., Cancer Treatment Reviews, 2023
Powered by
BACK
Sign up for alertsSIGN UP
American Physiological Society
Twitter
Facebook
LinkedIn
YouTube
AMERICAN PHYSIOLOGICAL SOCIETY JOURNALS
American Journal of Physiology-Cell Physiology
American Journal of Physiology-Endocrinology and Metabolism
American Journal of Physiology-Gastrointestinal and Liver Physiology
American Journal of Physiology-Heart and Circulatory Physiology
American Journal of Physiology-Lung Cellular and Molecular Physiology
American Journal of Physiology-Regulatory, Integrative and Comparative Physiology
American Journal of Physiology-Renal Physiology
American Journal of Physiology (1898-1976)
Physiological Genomics
Journal of Applied Physiology
Journal of Neurophysiology
Advances in Physiology Education
Physiological Reviews
Physiology
Physiological Reports
Comprehensive Physiology
Function
INFORMATION FOR
Authors
Reviewers
Librarians
Advertisers
APS Members
Media/Public
PROGRAMS
APSselect
Legacy Collection
APS Books Program
APS
Visit the APS website
Become an APS member
I Spy Physiology Blog
STAY CONNECTED
Submit
Subscribe
About Us
Contact Us
Your Account
NEWS
APS Press Releases
Publication News
American Physiological Society logo Privacy Policy Disclaimer Terms & ConditionsCopyright © 2024 the American Physiological Society
PDF
Help | biology | 993573 | https://da.wikipedia.org/wiki/Tr%C3%A6ningsintensitet | Træningsintensitet | Træningsintensistet er beskrivelsen af hvor meget energi der forbruges når man bevæger sig. Selvopfattet intensitet varierer fra person til person. Man har fundet ud af, at intensiteten har en effekt på hvilket brændstof kroppen bruger, og hvilke former for tilpasninger kroppen laver efter fysisk aktivitet. Intensitet er mængden af fysisk kraft kroppen bruger når den udfører en aktivitet. Eksempelvis definerer træningsintensitet hvor meget kroppen skal arbejde for at gå en kilometer på fem minutter.
Målinger af intensitet
Pulsmåling bruges typisk til at måle træningsintensiteten. Pulsen kan være en indikator for hvor meget det kardiovaskulære system skal arbejde ved en given fysisk aktivitet.
Den mest præcise måling af intensitet er iltoptagelse (VO2). VO2 repræsenterer den samlede metaboliske udfordring ved en given fysisk aktivitet. Der findes et direkte lineært forhold mellem intensiteten af aerob aktivitet og VO2. Den maksimale intensitet er en spejling af den maksimale iltoptagelse (VO2max). En måling af denne repræsenterer det kardiovaskulære fitnessniveau.
VO2 er målt i METs (ml/kg/min). En MET, der er lige med 3,5 ml/kg/min, anses for at være det gennemsnitlige energiforbrug i hvile for et normal menneske. Den fysisk aktivitets intensitet kan udtrykkes som multiple af dette energiforbrug i hvile. En fysisk intensitet lige med 6 MET betyder at energiforbruget ved denne fysiske aktivitet er seks gange højere end ved hvile.
Fysisk aktivitets intensitet kan blive udtryk i absolutte eller relative terme. Eksempelvis vil to individer med forskellige VO2max-målinger, der begge løber med ti km/t, løbe ved samme absolutte intensitet (kilometer/time), men forskellig relativ intensitet (% af VO2-max). Individet med det højere VO2-max løber med en lavere intensitet ved denne hastighed, end individet med den lavere VO2-max er.
Nogle studier måler træningsintensitet ved at få subjekter til at udføre fysiske test for at fastslå maksimal kraftproduktion, hvilket kan måles i watt, puls, eller gennemsnitlig kadence hvis det er på en cykel. Her forsøger man at udregne den samlede arbejdsbelastning.
Referencer
Fysiologi | danish | 0.713684 |
below_temperature_muscles_not_work/PMC2269891.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List J Physiol v.524(Pt 2); 2000 Apr 15 PMC2269891
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of jphysiol
J Physiol. 2000 Apr 15; 524(Pt 2): 603–615.
doi: 10.1111/j.1469-7793.2000.00603.x
PMCID: PMC2269891
PMID: 10766936
Heat production in human skeletal muscle at the onset of intense dynamic exercise
José González-Alonso, Bjørn Quistorff, Peter Krustrup, Jens Bangsbo, and Bengt Saltin
Author information Article notes Copyright and License information PMC Disclaimer
Go to:
Abstract
We hypothesised that heat production of human skeletal muscle at a given high power output would gradually increase as heat liberation per mole of ATP produced rises when energy is derived from oxidation compared to phosphocreatine (PCr) breakdown and glycogenolysis.
Five young volunteers performed 180 s of intense dynamic knee-extensor exercise (≈80 W) while estimates of muscle heat production, power output, oxygen uptake, lactate release, lactate accumulation and ATP and PCr hydrolysis were made. Heat production was determined continuously by (i) measuring heat storage in the contracting muscles, (ii) measuring heat removal to the body core by the circulation, and (iii) estimating heat transfer to the skin by convection and conductance as well as to the body core by lymph drainage.
The rate of heat storage in knee-extensor muscles was highest during the first 45 s of exercise (70-80 J s−1) and declined gradually to 14 ± 10 J s−1 at 180 s. The rate of heat removal by blood was negligible during the first 10 s of exercise, rising gradually to 112 ± 14 J s−1 at 180 s. The estimated rate of heat release to skin and heat removal via lymph flow was < 2 J s−1 during the first 5 s and increased progressively to 24 ± 1 J s−1 at 180 s.
The rate of heat production increased significantly throughout exercise, being 107 % higher at 180 s compared to the initial 5 s, with half of the increase occurring during the first 38 s, while power output remained essentially constant.
The contribution of muscle oxygen uptake and net lactate release to total energy turnover increased curvilinearly from 32 % and 2 %, respectively, during the first 30 s to 86 % and 8 %, respectively, during the last 30 s of exercise. The combined energy contribution from net ATP hydrolysis, net PCr hydrolysis and muscle lactate accumulation is estimated to decline from 37 % to 3 % comparing the same time intervals.
The magnitude and rate of elevation in heat production by human skeletal muscle during exercise in vivo could be the result of the enhanced heat liberation during ATP production when aerobic metabolism gradually becomes dominant after PCr and glycogenolysis have initially provided most of the energy.
Quantification of energy fluxes in contracting muscle in man is difficult. In isometric contractions with a well-defined muscle group this has been attempted when the force of the contraction causes a complete mechanical hindrance to the blood flow, i.e. when aerobic energy yield plays no role (Edwards et al. 1972; Saugen & Vøllested, 1995). In intense dynamic exercise, however, oxidation is the primary energy-liberating pathway after ∼60 s of exercise, while at the onset of dynamic exercise anaerobic energy production predominates. At the whole body level oxygen consumption can be precisely determined in humans, but it is more of a problem to measure the amount of oxygen used by a specific muscle or muscle group. To, at least in part, overcome this problem the one-legged knee-extensor exercise model was developed, confining the dynamic contractions largely to the quadriceps muscle (Andersen et al. 1985; Ray & Dudley, 1998; Richardson et al. 1998). Blood flow to or from this muscle can be measured, as can the arterial-venous (a-v) difference for oxygen, giving the oxygen uptake by the Fick principle. During steady-state submaximal dynamic knee-extension exercise, the mechanical efficiency estimated according to the traditional concept (i.e. the ratio between power output and the sum of the caloric equivalent of oxygen consumption and power output; Benedict & Cathcart, 1913) was found to be ∼25 % (Andersen & Saltin, 1985), which is in the upper range of what has been observed at the whole body level with cycle ergometer exercise (Åstrand, 1960). With more intense exercise and at the onset of exercise not only the oxygen uptake, but also the anaerobic energy liberation has to be determined in order to obtain an estimate of energy turnover. Bangsbo et al. (1990) have made such an attempt using the knee-extensor model. The anaerobic processes were estimated from the reductions of muscle ATP and PCr, the accumulation of glycolytic intermediates and lactate, and the release of lactate. Although there was an acceptable match between this estimate of the anaerobic energy yield and the oxygen deficit (total energy turnover – aerobic energy turnover) it did not provide sufficient insight since total energy turnover during exercise remained unknown.
The appropriate approach, novel in humans performing dynamic exercise, would be to measure heat production, power output and aerobic and anaerobic energy turnover. This approach has a long and successful history in muscle energetics in the in vitro study of muscle fibres (Wilkie, 1960; Hill & Woledge, 1962; Kushmerick et al. 1969), in vivo in exercising animals (Ardevol et al. 1998) and in man performing isometric contractions (Edwards et al. 1972, 1975; Saugen & Vøllested, 1995, 1996). Furthermore, early work of Barcroft & Edholm (1942) and Pennes (1948) established the theoretical basis for the quantification of heat transfer in perfused tissue. Despite the complexity of the various tissues, muscle fibre and motor unit recruitment (Lexell et al. 1983), the knee-extensor model could provide a means to precisely determine total heat production in dynamic exercise by measuring heat stored in the contracting muscles and that dissipated from the muscle. If heat capacity and muscle mass are known, it is possible to determine heat storage via thermosensors placed in the various active muscle portions. Heat dissipation from dynamically contracting muscles consists of heat transfer to the core of the body (limb blood flow × v-a temperature difference according to the Fick principle) and to surrounding tissues or environment. Heat conductance through tissues in the human body is a slow process (Hensel & Bock, 1955) and in knee-extensor exercise it can be estimated based on temperature measurements of inactive tissues within the thigh, including the subcutis. Heat exchange with the surroundings of the exercising thigh (convection and conduction) can be minimised by a thermostat isolation system.
Thus, the aim of this study was to quantify energy liberation at the onset of and during short intense dynamic exercise by continuously measuring the rate of heat production and power output by the knee-extensors. This was done to test the hypothesis that in man the efficiency of conversion of chemical energy to mechanical power is high in the transition from rest to exercise, and then gradually declines corresponding to the change in source of ATP production. This hypothesis is based on the findings from in vitro studies that heat liberation during ATP utilisation varies from 35 to 72 kJ (mol ATP)−1 depending upon whether PCr, glycolysis or oxidative phosphorylation provide the energy for ATP resynthesis (Wilkie, 1968; Curtin & Woledge, 1978; Woledge & Reilly, 1988). Thus, an increase in heat production in the order of a factor of two would be anticipated with a shift in ATP resynthesis from pure PCr splitting to pure oxidative phosphorylation. Continuous measurements were performed of the aerobic metabolism whereas data on the anaerobic contribution could not be obtained simultaneously due to technical restraints. However, such information is available from parallel studies using the same exercise model and of the same intensity and duration.
Go to:
METHODS
Subjects
The five healthy, recreationally active males participating in this study possessed a mean age of 24 years (range 22–25 years), mean body weight of 76.6 kg (55-92 kg) and a mean height of 180 cm (169-192 cm). The peak oxygen uptake of the quadriceps muscle of one leg was 0.89 l min−1 (0.59-1.05 l min−1) (Table 1). The study was approved by the Ethics Committee of Copenhagen and Frederiksberg communities and was carried out in accordance with the Declaration of Helsinki. Subjects gave written informed consent before participating.
Table 1
Subject characteristics
Subject no. Age (years) Weight (kg) Height (cm) Mean power output (W) Quadriceps and tensor fasciae latae (kg) Hamstrings, sartorious and adductors (kg) Skin, subcutaneous fat, connective tissue (kg) Femur (kg) Total thigh (kg) Peak V̇O2 thigh (l min−1)
1 24 74 178 87 2.6 2.8 1.1 0.7 7.2 0.99
2 24 90 188 72 3.0 3.7 2.1 0.9 9.8 0.78
3 25 55 169 66 2.1 2.8 0.8 0.7 6.5 0.59
4 22 92 192 109 3.3 3.6 1.7 1.0 9.6 1.02
5 24 72 172 80 2.4 2.5 1.2 0.7 6.7 1.05
Mean 24 77 180 83 2.7 3.0 1.4 0.8 7.9 0.89
±s.d. 1 15 10 17 0.5 0.6 0.5 0.2 1.6 0.20
Open in a separate window
Mechanical power output
Power output was continuously recorded during exercise. The mean force produced by the knee-extensor muscles during each kick was estimated by integrating the area under the curve (over the active angle ∼80-170 deg of the kicking cycle) obtained from the changes in voltage detected by a strain-gauge placed between the ankle and the metal rod connecting the leg to the fly-wheel (Andersen et al. 1985). The strain-gauge was calibrated by placing weights of known mass (5 and 20 kg) on the metal rod. The duration of the kick was measured by optical encoders on the cranks of the cycle ergometer. Power output was calculated by estimating the external work done on the ergometer as well as the work done to lift the lower leg.
Oesophageal, arterial, skin and muscle temperatures
Commercially available thermistors were used to continuously monitor skin and oesophageal temperatures (MHC-40050-A, Ellab A/S, Rødovre, Denmark) and for muscle and vessel blood temperatures (Edslab, TD probe 94-030-2.5F).
Thigh blood flow
Femoral venous blood flow was measured by a modified version of the thermodilution technique originally described by Andersen & Saltin (1985). With the same experimental set-up, we obtained continuous measures of the infusate and femoral venous blood temperatures, allowing the calculation of femoral venous blood flow second by second during the time of infusion of cold saline (20 s; 1–3°C) after an initial stabilisation period of 5 s. This was achieved by connecting the thermistors (Edslab, TD probe 94-030-2.5F) via a custom-made interface to a Macintosh Performa computer using a MacLab 8:s data acquisition system (ADInstruments, Sydney, Australia). The data sampling frequency was 100 Hz. Before and during the second 3 min exercise bout, femoral venous blood flow (index of thigh blood flow; TBF) was measured at rest, during the passive acceleration phase, and at the time intervals 0–10, 40-55, 80–95, 120–135 and 170–180 s during the exercise. Infusate temperature was measured at the site where the infusate entered the venous catheter (∼8 cm from the tip of the catheter). The set-up was tested under in vitro conditions to determine the linearity of the system up to blood flows of 10 l min−1. The in vitro experiment was also used to determine the elevation in the infusate temperature occurring as the infusate travels from the measuring point to the tip of the catheter. Infusate temperature was corrected accordingly by 0.6°C with infusion rates of 1.866 ml s−1 (exercise conditions) and by 1.0°C with infusion rates of 0.766 ml s−1 (resting conditions and passive acceleration phase). TBF (expressed in ml s−1) was calculated according to the following formula, derived from a heat balance equation (Ganz & Swan, 1974; Andersen & Saltin, 1985):
equation image
where VI represents infusate rate (ml s−1), TB is blood temperature before saline infusion (°C), TI is temperature of infusate, TM is temperature of blood-saline mixture during steady state (after 5 s infusion), SI and SB are specific gravities of infusate and blood (1.005 and 1.045 g cm−3), respectively, and CI and CB are specific heats of infusate and blood (4.173 and 3.600 J g−1°C−1), respectively.
Under the present experimental conditions (non-steady state), the reference blood temperature increased 0.05-0.09°C during the 20 s of infusion. Furthermore, although venous blood temperature was restored in less than 1 s upon termination of the infusion period, repeated blood flow measurements reduced the venous blood temperature by 0.10-0.15°C after 3 min of exercise compared to that observed during the first bout (no blood flow measurements). Notwithstanding, correction for this effect only increased thigh blood flow values by < 1 %. This effect is therefore negligible compared to the oscillations of limb blood flow from kick to kick (see Results).
Data sampling
Muscle and blood temperatures as well as power output were recorded with a sampling frequency of 100 Hz. In addition to the thermistors in the blood vessels (femoral artery and vein) each of the muscle thermistors were connected via another custom-made interface and A/D converter to an IBM computer board. Data were stored on a hard disk and displayed on-line, using LabVIEW 4.0 software (National Instruments, Austin, TX, USA). Temperature data were then averaged per second and expressed in degrees Celsius. The seven skin thermistors and the oesophageal thermistor were connected to an eight-channel temperature monitor (Ellab CTF 9008 Precision thermometer; Ellab A/S) interfaced with an IBM-AT computer. Data were stored on a hard disk every 15 s and also simultaneously displayed on-line using the data acquisition software PCLINK92 (Ellab A/S).
Muscle mass
Muscle mass was determined by magnetic resonance imaging (MRI) performed on a Siemens 1.5 T MAGNETOM vision scanner (Siemens, Germany). For each subject, 30–33 parallel axial T1-weighted images (cross-sections) of the right thigh (i.e. from the anterior superior iliac spine to the patellar ligament) were obtained with a multi-slice spin-echo FLASH sequence (T1 is the spin-lattice relaxation time; repetition time (TR) = 500 ms, echo time (TE) = 15 ms) using a standard body coil. Slice thickness was 3 mm with 12 mm interslice distance. Pixel size was 1.2 mm2. This setting was selected to optimise image quality in order to most clearly separate muscle, bone, fat, and connective tissue.
Definitions of anatomical cross-sections were performed manually and the area calculations made using NIH Image software (Rasband & Bright, 1995). The proximal portion of the patella was considered as the zero-reference slice for all the subjects. The volume of the compartments was calculated as the sum of each anatomical cross-sectional area times the distance covered by each slice (1.5 cm). Volume values were converted into mass values using the specific density of each compartment (1.043 for muscle, 0.9 for fat, 1.791 for femur, 1.053 for skin, and 1.045 for soft tissue; von Döbeln, 1956). The anatomical compartments investigated were: (1) quadriceps plus tensor fasciae latae muscles (active during knee-extensor exercise), (2) hamstring, sartorious and adductor muscles complex (inactive), (3) bone (femur) and (4) skin, subcutaneous fat and connective tissue complex (Fig. 1A and B).
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f1.jpg
Figure 1
Anatomical compartments of the thigh and quantification of the knee-extensor muscle mass
A, MRI of cross-sections of upper-thigh (a) and mid-thigh (b) with the white line indicating borders of quadriceps femoris muscle, including vastus lateralis (v.l.), vastus intermedius (v.i.), vastus medialis (v.m.), rectus femoris (r.f.) and tendon of quadriceps femoris muscle (t). A cross-section of the distal end is shown in c. It was sometimes difficult to exactly determine the origin of the muscles at the proximal end (a). This was solved by plotting the serial individual cross-sections and determining the origin by extrapolation (see Ba). Mean values ±s.e.m. for all subjects’ serial cross-sections of the thigh are shown in Bb.
Procedures and protocol
In preparation for this study the subjects were familiarised with the exercise model by training at a desired cadence while minimising the involvement of the hamstring and gluteal muscles, and thereby confining the work to the knee-extensor muscles. In the preliminary trials they also became accustomed to a thermistor placed in the oesophagus at the level of the heart while exercising.
On the morning of the experiment, subjects arrived after a light breakfast. Three catheters were placed by the Seldinger technique at the level of the inguinal ligament. In the resting leg, a catheter was placed in the femoral artery for blood sampling. In the exercising leg, catheters were placed in both the femoral vein and artery. Through these latter two catheters, thermistors (Edslab probe 94-030-2.5F) were placed just at the tip of the catheters for the measurements of arterial-venous differences for temperature (°C). Following the placement of the arterial and venous catheters, seven thermistors of the same type were inserted into the thigh muscles through a venflon catheter (18G/32 mm). The tip of the thermistor probe used in the muscle was cut 2–3 mm beyond the thermistor location, which was advanced into the muscle 0.5-0.8 cm beyond the catheter tip (approximately 3.0-4.0 cm into the thigh, depending upon muscle group and size of subjects). Different inclinations (30, 45 and 60 deg) with respect to the length direction of the muscle fibres were used when the thermistors were inserted. This minimised their movement, reduced the risk of damaging the muscle or the thermistors and minimised the discomfort of subjects. Three thermistors were inserted in the vastus lateralis (v.l., proximal, medial and distal portions), two in the rectus femoris (r.f., medial and distal portions), one in the distal portion of the vastus medialis (v.m.), and one in the medial portion of the biceps femoris. The flexible venflon catheter was left in place to provide protection for the thermistor probe. The thermistor probes and the venflon catheters were fastened to the skin with tape. Due to the difficulty reaching the vastus intermedius (v.i.) muscle, which is located underneath the rectus femoris (Fig. 1A), thermistors were only placed in this muscle group in two separate additional experiments. Next to each of the seven muscle thermistor probes, a skin thermistor was placed and secured to the skin with tape. Thereafter, the oesophageal thermistor was inserted through the nostril down to the level of the heart.
To minimise the heat losses to the environment surrounding the exercising leg via convection and conductance, local muscle, skin, and blood temperatures were equalised to the core temperature (õesophageal ≈ 37°C) by perfusing a cuff surrounding the thigh with warm water (∼41°C). This procedure lasted 45–60 min until muscle and tissue temperatures were ∼37°C (Fig. 3). Thereafter, the temperature of the water was reduced to 37.5°C to minimise heat transfer to or from the surroundings.
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f3.jpg
Figure 3
Thigh temperature during the thermal equilibration procedure prior to exercise
The tissue temperatures of the thigh are depicted when warming it with a water-perfused wrapping set at 41 °C for ≈70 min to equalise these temperatures with the core temperatures (≈37 °C). Thereafter, the water temperature was adjusted to 37.5 °C for the remainder of the experiment.
The subject performed the 3 min knee-extensor exercise with the right leg in a recumbent position at a mean power output of 83 ± 9 W (mean kicking frequency of 1.03 ± 0.02 s−1) for 3 min. The work was close to exhaustive for the subject. During exercise, the left leg was resting. Over a period of 30 s prior to the voluntary exercise, the lower part of the right leg was accelerated passively up to the target cadence to ensure that the subject did not use extra energy to accelerate the fly-wheel. During the 3 min exercise period a cuff just below the knee was inflated to 240 mmHg to avoid any transport of heat to or from this part of the leg. Additionally, blood samples were withdrawn from the femoral artery and vein at rest, during passive exercise and at 13, 31, 50, 75, 128 and 176 s of the first exercise bout, for later analysis of oxygen saturation and haemoglobin concentration (OSM-3 Hemoximeter, Radiometer, Copenhagen, Denmark) as well as blood lactate (lactate analyser; Yellow Springs Instruments, Yellow Springs, OH, USA).
To determine the thigh blood flow the subjects repeated the same exercise after 1 h of recovery during which femoral venous blood flow was measured using the thermodilution method, as explained above. The reason for measuring blood flow during this second bout of exercise was to avoid the confounding effect of infusing cold saline on venous and arterial blood temperatures. Previous results from this laboratory indicate that the pattern and the magnitude of femoral venous blood flow are the same during a first and a second bout of intense knee-extensor exercise, separated by 1 h of recovery (Bangsbo et al. 1992). After each experiment all thermistors were immediately calibrated within the temperature range 35–40°C against a mercury thermometer having a precision of 0.01°C. The response time of the thermistors in the muscles and in the blood vessels, being Teflon imbedded, was ≤ 0.8 s.
Calculations (see Fig. 2)
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f2.jpg
Figure 2
Schematic model used for calculation of total energy turnover
Ta and Tv, arterial and venous temperature. Tv-a, venous-arterial temperature difference. ΔTm, mean increase in temperature of all muscles.
Rate of heat storage in active muscles The rate of heat storage (Hs) in the active muscles (i.e. quadriceps and tensor of fasciae latae) was calculated for 5 s intervals by multiplying the mean increase in temperature of all muscle portions in 5 s (ΔTm;°C) by the muscle mass (m; kg) and the specific heat of the muscle at 37.5°C (cm= 3590 J kg−1°C−1), assuming a 23 % solid content (based on an average water content in quadriceps muscle of 77 %). The resulting value was divided by 5 to express the rate of heat storage in joules per second. The active muscle mass was corrected by the estimated increases in muscle volume observed during intense knee-extensor exercise (Bangsbo et al. 1992; Ray & Dudley, 1998). The same principle was used to determine the rate of heat storage in the hamstrings muscle:
equation image (1)
Rate of heat removal by the blood The mean thigh blood flow (TBF; l s−1) during each 5 s interval was estimated from the individual curve fitting applying a 3rd-order polynomial model (r2= 0.84-0.93). The rate of heat removal by the blood (Hr) was calculated for 5 s intervals by multiplying the mean v-a temperature gradient (Tv-a;°C) by TBF and the specific heat of the blood at 37.5°C (blood specific heat (cb) = 3610 J l−1°C−1; haemotocrit ∼45 %):
equation image (2)
Additional heat loss
The additional rate of heat loss (Ha) was calculated by adding the estimated heat loss to the surrounding skin by the processes of conductance and convection as well as the heat transfer to the body core by the lymph drainage.
(1) Rate of heat loss by conductance. The heat transfer from the knee-extensor muscles to the skin through conductance (Hc) was estimated every 5 s by multiplying the temperature gradient between the muscle and skin (ΔTm-sk;°C) by the thermal conductivity of human muscle (ch= 4.8 J s−1 cm−1°C−1; Hensel & Bock, 1955) and the mean distance through which heat is conducted in the anterior thigh (l =∼3 cm):
equation image (3)
(2)Rate of heat loss by convection. Heat loss from the skin of the thigh by convection (Hcv) was calculated every 5 s by multiplying the estimated thigh skin blood flow (SkBF = 0.005 l s−1 measured in the saphenous vein) by the temperature gradient between the arterial blood and saphenous venous blood (equal to skin temperature under these conditions; Ta-v,°C) and the specific heat of the blood (cb= 3610 J l−1°C−1):
equation image (4)
(3) Rate of heat loss by the lymph flow. Heat loss by the lymph flow (Hl) was estimated by multiplying the estimated lymph flow (LF = 0.003 l s−1) by the arterial-to-lymph temperature gradient, assuming that the lymph leaving the thigh has the same temperature as the venous blood and the same specific heat as plasma (cp= 3930 J l−1°C−1):
equation image (5)
Rate of heat production Total rate of heat production (Ht) was calculated by adding the rate of heat storage in the active muscles, the rate of heat removal by the blood and the rate of heat transfer to the skin by conductance and convection as well as to the body core by the lymph drainage:
equation image (6)
Total energy turnover (Et) was calculated by summing Ht and mechanical power output (W):
equation image (7)
Mechanical efficiency Mechanical efficiency (MEf) was calculated by dividing the mechanical power output by the total energy turnover:
equation image (8)
Aerobic and anaerobic heat liberation Oxygen uptake (V̇O2 expressed in ml s−1) of the exercising thigh was obtained by multiplying the a-v O2 difference by thigh blood flow (Fick principle), and converted into moles of O2 assuming that 1 mole O2 equals 25.4-25.5 l O2 when temperature ranges from 37 to 38°C. V̇O2 was then converted into moles of ATP assuming a P:O ratio of 3.0 throughout exercise. Resting V̇O2 was subtracted from exercise V̇O2 to compare aerobic heat liberation to total energy turnover. Net PCr hydrolysis and lactate accumulation estimations per 30 s interval are based on the initial and final biopsy mean values (vastus lateralis) obtained in parallel studies with the same experimental protocol (J. Bangsbo, P. Krustrup, J. González-Alonso & B. Saltin, unpublished observations; ΔPCr 16.3 mmol (kg wet wt)−1 and Δlactate 22.3 mmol (kg wet wt)−1) and the kinetics described by Gollnick & Hermansen (Fig. 3, p. 12; 1973) and Sinclair et al. (1999). Lactate concentrations were converted into ATP concentrations by multiplying lactate concentrations by a factor of 1.5. Heat produced per mole of ATP used was assumed to be 35 kJ for ATP and PCr hydrolysis, 65 kJ for glycolysis and 72 kJ for oxidation (Curtin & Woledge, 1978; Woledge & Reilly, 1988).
Analysis
The mean response time for total heat production was estimated by fitting a two-component exponential model to the data, using a (non-linear) least squares regression technique.
Statistics
A one-way repeated measures analysis of variance (ANOVA) was performed to test significance over time. When the F value was significant, pairwise differences were identified using Tukey's honestly significant difference post hoc procedure. The significance level was set at P < 0.05. Data are presented as means ±s.e.m. unless otherwise stated.
Go to:
RESULTS
Pre-exercise thermal equilibration
The use of a water-perfused jacket was quite successful in equalising temperatures at ∼37.0°C within the thigh before the exercise (see Fig. 3). This was true for the various muscles except for the hamstrings. At the start of the exercise, the hamstrings had a slightly higher temperature than ∼37.0°C (i.e. 37.3-37.4°C), probably due to the direct contact with, and pressure of the posterior side of the thigh on, the water jacket.
Thigh size
The mean mass of the knee-extensor muscle including the tensor fasciae latae was 2.68 ± 0.46 kg (±s.d.) (Table 1). The mass of m. rectus femoris was ∼0.2-0.3 kg with the other three portions of the quadriceps being of similar weight (0.6-0.8 kg). After 180 s of knee-extension exercise, the active muscle volume is estimated to increase by 0.26 ± 0.04 kg to 2.94 ± 0.50 kg due to fluid gain.
Muscle temperature
At the onset of the voluntary dynamic exercise the temperature in the quadriceps muscles was promptly elevated at all sites, i.e. after the first kick (Fig 4A and Fig 5A, B). However, there were some differences in the magnitude of increase in temperature among the different muscle portions (v.l., r.f. and v.m.) and subjects. A faster increase was observed in rectus femoris with 0.26-0.31°C elevation over the first 30 s compared to the vastus medialis and vastus lateralis which reached 0.14-0.19°C over the same time interval (P < 0.05; Fig. 5A). At the end of the 3 min exercise bout, the temperature rise ranged between 0.89 and 1.03°C in v.l., v.m. and r.f. In two additional experiments in which thermistors were placed in the v.i. portion of the quadriceps femoris muscle and in the tensor fasciae latae, the temperature elevation was similar to that observed in the other portions of the knee-extensor muscles (Fig. 6A). This rendered the basis for using a mean value of temperature in v.l., v.m. and r.f. for the entire knee-extensor muscles. When comparing the rate of rise in muscle temperature, it was observed that knee-extensor muscle temperature increased 0.04 ± 0.01°C during the first 5 s to decline progressively to 0.01 ± 0.01°C during the last 5 s of exercise (P < 0.05). In contrast to the knee-extensor muscles, the temperature in the hamstrings barely increased during the first 120 s of exercise (Figs 5A and and6B),6B), but was at the end elevated by 0.19°C (range 0.13-0.42°C). Half of this elevation occurred during the last 30 s of exercise (Fig. 5A). In the additional experiments, it was observed that anterior thigh subcutaneous tissue temperature and hamstrings temperature in three different sites only increased slightly during exercise (Fig. 6B).
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f6.jpg
Figure 6
Temperature in various knee-extensor and hamstring muscles as well as subcutaneous tissue during intense knee-extensor exercise
A, mean increases in the two additional experiments when thermistors were placed in vastus intermedius (v.i.) and tensor fasciae latae (t.f.l.) in addition to other knee-extensors. B, data from another additional experiment. Three thermistor probes were placed in different hamstring muscles (biceps femoris, semitendinosus and semimembranosus), and one in the subcutaneous fat (≈5 mm under the skin) next to a thermistor probe placed in the rectus femoris (depth ≈3 cm).
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f5.jpg
Figure 5
Temperature and thigh blood flow during intense dynamic knee-extensor exercise
A, mean values (n = 5) for temperature increases in the rectus femoris (r.f.; 2 sites), vastus medialis (v.m.; 1 site) and vastus lateralis (v.l.; 2 sites) of the quadriceps muscle and hamstrings muscle (1 site; biceps femoris medial) during dynamic knee-extensor exercise. s.e.m. bars are not included for reasons of clarity (s.e.m. range 0.00-0.10 °C; mean s.e.m. values 0.04-0.07 °C for all muscle portions). B, the mean (±s.e.m.) increases (n = 5) in arterial and venous blood temperatures are given from start to end of the exercise. For comparison mean (±s.e.m.) quadriceps, oesophageal (core) and skin temperature are given as well. C, mean (±s.e.m.) values for thigh blood flow during exercise are presented. The mean elevation (n = 5) observed during the exercise fits the polynomial equation:
equation image
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f4.jpg
Figure 4
Mean quadriceps muscle temperature, arterial and femoral blood temperature (A) and thigh blood flow (B) for all subjects at rest and during passive exercise, dynamic exercise and recovery
Of note is the observation that the decline in muscle temperature during recovery can be accounted for almost completely by the on-going convective heat removal to the body core.
Blood temperature
Femoral venous temperature increased after the first kick with the arterial temperature lagging behind (Figs 4A and and5B).5B). The v-a temperature difference was nil during the first 5 s but increased to 0.09 ± 0.06°C after 15 s (Fig. 5B). Thereafter, v-a temperature difference increased further to reach 0.39 ± 0.04°C at the end of exercise. The absolute values at the termination of exercise for femoral artery and venous blood temperatures were 37.65 ± 0.06 and 37.21 ± 0.08°C, respectively. The rate of rise in oesophageal temperature (Toes) was only slightly lower than that of femoral artery temperature, reaching a value of 37.1 ± 0.1°C at the end of exercise (Fig. 5B). The observation that the rise in Toes (index of body core or central blood temperature) throughout exercise was similar to that of arterial temperature, indicates that countercurrent heat exchange between the femoral vein and femoral artery, if it indeed occurred, was very small.
Thigh blood flow
Although the passive acceleration of the leg did not cause any alteration in temperature of the muscle or the arterial blood (Fig. 4A), it increased the thigh blood flow from its resting level of 7.7 ± 1.5 ml s−1 to 33.7 ± 1.2 ml s−1 (Figs 4B and and5C).5C). A fast further elevation occurred in the blood flow during the voluntary exercises reaching 56.7 ± 10.3 ml s−1 at 45 s and 76.7 ± 16 ml s−1 at the end of the exercise.
Heat production
The rate of heat storage in the knee-extensors (Hs) was largest early in the exercise, amounting to 70–80 J s−1 after the first 30 s, after which it gradually declined to 14 ± 10 J s−1 for the last 30 s of exercise (Fig. 7). The rate of heat removal by the blood (Hr) was not significant until after 10 s of exercise, then being 8 ± 10 J s−1. Thereafter, convective heat removal increased progressively to reach a value of 112 ± 14 J s−1 during the last 5 s of exercise (Fig. 7). The decline in Hs was less than the elevation in Hr, especially during the first 60–75 s of exercise, in keeping with an increased heat production during the exercise (Fig. 7). The rate of heat production was 70 ± 10 J s−1 during the first 5 s of the exercise, and approached a value just above 100 J s−1 at 60–75 s (P < 0.05). Thereafter, the increase in the rate of heat production was linear reaching 126 ± 26 J s−1 at the end of the exercise. Therefore, heat production was 77 % higher (P < 0.05) during the last compared to the first 5 s. After 180 s of exercise, total heat storage in the knee-extensors amounted to 9276 J (range 7822–11776 J), whereas total heat removal by the blood amounted to 10993 J (range 6808–20403 J).
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f7.jpg
Figure 7
Heat production during dynamic knee-extensor exercise
Mean values (±s.e.m.; n = 5) of total heat production (Ht) are depicted for each 5 s period of the exercise as well as its subdivision in terms of storage in the quadriceps muscle (Hs) and removal by the blood from the thigh (Hr).
Additional heat loss
To the above reported heat production should be added the small heat losses by convection, conduction and convection via the lymph flow (Ha). The amount of heat conducted to the skin of the anterior thigh during exercise was estimated to range from ∼2 J s−1 during the first 5 s of exercise to 6 J s−1 during the last 5 s of exercise, with almost no difference among subjects. The corresponding calculation for the posterior portion of the thigh demonstrated that it can be neglected due to the temperatures in the hamstring and the skin being similar. With no temperature gradient, heat transfer to the thigh skin through convection was estimated to be zero at the beginning of exercise, increasing progressively throughout exercise to a value of 12 ± 4 J s−1. Heat transfer to the body core via the lymph might amount to a maximum of 6 J s−1. Therefore, during the first 5 s of exercise, Ha was negligible (< 2 J s−1), and increased throughout exercise to a value of 24 J s−1 (range 6–48 J s−1) at 180 s. The total value for Ha is 2446 J (range 334–3096 J), which represents a maximum of 10 % of Ht (22726 J; range 16612–33878 J) or 6 % of Et (37652 J; range 31650–44769 J). When including Ha, Ht during the last 5 s of exercise was twice as high as that observed during the first 5 s of exercise (P < 0.05) (Fig. 7), with half of the increase occurring during the first 38 ± 12 s.
Mechanical power output
During the first 30 s of exercise the mean power output was 83 W (range 58–114 W). During the subsequent 90 s, power output was within 1 % of this level (Fig. 8C). Overall there was a trend for a reduction in power output which was due to a progressive decline in kicking frequency from 1.08 ± 0.03 s−1 during the first 5 s to 0.96 ± 0.03 s−1 during the last 5 s (P < 0.05; Fig. 8A and B). After 120 s of exercise, power output dropped somewhat more in two subjects. This coincided with a progressive increase in the work performed by the hamstrings as evidenced by the force tracing.
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f8.jpg
Figure 8
Mechanical power output during dynamic knee-extensor exercise
Mean values for mechanical work per kick (A) and the kicking frequency (B) as well as the mean power output over 30 s intervals (C) for n = 5.
Mechanical efficiency
With heat production doubling over the 180 s of the exercise while power output was in essence constant (Fig. 8C), the estimated mechanical efficiency declined from an initial value of 53 ± 6 % to 36 ± 5 % at the end of exercise (P < 0.05).
Oxygen consumption and lactate release
Thigh V̇O2 increased in a curvilinear fashion from 0.055 ± 0.023 mmol s−1 immediately before exercise to 0.464 ± 0.035 mmol s−1 at the end of exercise, with one-half of the increase occurring during the first 28 ± 3 s (Fig. 9A). Net lactate release from the thigh increased from 0.01 ± 0.01 mmol s−1 immediately before exercise to 0.15 ± 0.03 mmol s−1 (P < 0.05) after 70 s and 0.21 ± 0.02 mmol s−1 at the end of exercise (P < 0.05; Fig. 9B).
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f9.jpg
Figure 9
Oxygen consumption (A) and lactate release (B) during exercise
Data are means ±s.e.m. for 5 subjects.
Go to:
DISCUSSION
The major finding of this study was that heat production by contracting human skeletal muscle doubled over 3 min of intense dynamic exercise at essentially constant power output. Half of this increase in rate of heat production occurred during the first 38 s of exercise. This elevated heat production in contracting skeletal muscle appears to be tightly coupled with changes in heat liberation during ATP production in the metabolic reactions involved early in exercise.
Measurement in man of heat production in dynamically contracting muscle with a free blood flow is complex, requiring a high time resolution and precise measurements. This was accomplished in this study and one major finding was that after 60 s of exercise, when aerobic metabolism provided 82–89 % of the ATP resynthesis, the match between total energy turnover (sum of rate of heat production and power output) and total metabolic input (sum of net PCr and net ATP hydrolysis, glycolysis and oxidative phosphorylation) was quite close. However, the critical time period is the very first minute of exercise where the rate of heat production is much smaller than in the later phase of the exercise bout. Multiple thermistors were placed in all major muscle portions developing the force as well as in one adjacent inactive muscle and temperature was recorded continuously. As evidenced in Figs 4A and and5A,5A, an elevation in temperature in the active muscle can be observed during the first few contractions (1-3 s). The observation that passive exercise did not elevate quadriceps muscle temperature and that knee-extensor exercise did not increase hamstring temperature early in exercise indicates that movement friction of the thermistor did not add to the heat gain in the muscle. Therefore, the precise muscle temperature and volume measurements in the present study provided highly valid estimates of heat accumulated in the muscle, including the initial phase of the exercise.
Some uncertainty, however, is involved in the estimation of the heat removal from the muscle by the blood flow. The inflow and outflow temperatures are obtained with the same time resolution and sensitivity as the measurements of the changes in muscle temperature and have an error of ∼0.01°C. The largest uncertainty in the measurement of heat removal by the blood is the blood flow determinations. The thermodilution technique does not allow for continuous measurements. Thus, thigh blood flow in the present study was measured for 20 s every ∼30 s and mean thigh blood flow during each 5 s interval was estimated using a curve-fitting polynomial model. The present rate of rise and absolute values in thigh blood flow correspond closely with continuous Doppler measurements using the same exercise model and exercise intensity (Rådegran & Saltin, 1998). In this study, a rapid increase in thigh blood flow was found with the first contractions. However, in this time interval the v-a temperature difference was very small, indicating that heat removal by the blood was negligible and that even a large error in blood flow measurement would only constitute a very small error in the estimation of convective heat removal. Furthermore, in the present experimental conditions with no or small temperature gradients between muscle and skin during the first 30 s of exercise, the additional heat loss to the skin by convection and conductance as well as to the body core by lymph drainage amounted to < 3 % of total heat production. Taken together, these findings indicate that most (> 90 %) of the heat produced during the first 10 s of exercise accumulated in the contracting muscles whereas, at the end of exercise, most of the heat produced was transported by the blood to the body core (see Fig. 7). Therefore, it is very unlikely that the heat accounting during the initial phase of the exercise bout is missing significant amounts of heat and vice versa for the later part of the exercise bout. Consequently, since work output is maintained nearly constant throughout the exercise, the overall efficiency of the muscle work performed is decreasing proportionally to the additional heat output.
There are several possible mechanisms explaining our observation at the onset of exercise of progressive increasing rate of heat production during generally maintained power output. One likely explanation is based on Wilkie's work in the 1960 s (Wilkie, 1968) and that of Woledge & Reilly (1988). They studied the heat produced during muscle contraction when the ATP resynthesis was powered by a net breakdown of PCr and found that the immediate heat liberated was only ∼35 kJ per mole of ATP used due to the near-equilibrium state of the creatine kinase reaction (Wilkie, 1968; Woledge & Reilly, 1988) as compared to the ∼72 kJ per mole of ATP used when ATP is resynthesised via oxidation (Curtin & Woledge, 1978). ATP resynthesis from the anaerobic usage of glycogen (glucose) appears to have an intermediate value for heat liberation compared to PCr breakdown and aerobic metabolism (i.e. ∼65 kJ per mole of ATP used; Curtin & Woledge, 1978). We measured thigh V̇O2 and lactate release and observed that oxidation accounted for ∼30 % of the total energy turnover during the first 30 s of exercise, increasing to > 82 % after 60 s of exercise. PCr degradation and lactate accumulation in muscle were not assessed in this study; however, these parameters have been measured repeatedly in the same model with similar work rate and exercise regimen in other studies (Bangsbo et al. 1990; Sinclair et al. 1999). Based upon measured absolute values in parallel studies and well-documented PCr and lactate kinetics (Gollnick & Hermansen, 1973; Meyer, 1988; Bangsbo et al. 1990; Sinclair et al. 1999), a likely model can be described for the energy contribution of these pathways (see Table 2). Accordingly, PCr and anaerobic glycogenolysis would account for 15 and 25 % of the total energy turnover, respectively, and 20 and 33 % of the total ATP production, respectively, during the first 30 s of exercise. During this 30 s interval the anaerobic processes therefore provided approximately two-thirds of the ATP resynthesis (Table 2; see also Fig. 10 for an indirect account of the anaerobic heat liberation). The observation that the metabolic rate of heat liberation was 53–68 % higher in all the 30 s intervals following the initial 30 s of exercise strongly suggests lesser heat liberation per ATP provided by ATP-PCr hydrolysis and glycogenolysis compared to ATP provided by oxidative phosphorylation.
Table 2
Muscle energetics during 3 min of intense dynamic kneeextension exercise
Time interval (s)
Energy source 0–30 30–60 60–90 90–120 120–150 150–180 0–180
(1) Oxygen consumption (J s−1) 57.6 ± 7.5 132.8 ± 9.7 168.0 ± 16.9 176.8 ± 22.0 182.0 ± 22.6 196.7 ± 19.6 152.3 ± 15.3
(2) Net PCr hydrolysis (J s−1)a 27.0 15.0 6.3 1.7 0.8 0.3 8.6
(3) Net ATP hydrolysis (J s−1)b 0.4 0.6 0.6 0.6 0.6 0.6 0.6
(4) Lactate accumulation (J s−1)a 40.6 64.3 12.6 7.9 6.3 5.0 21.7
(5) Lactate release (J s−1) 4.1 ± 1.2 11.4 ± 2.1 17.1 ± 2.6 19.0 ± 2.9 18.2 ± 2.7 18.4 ± 2.4 14.7 ± 2.2
Σ(1–5) Total metabolic input (J s−1) 133.8 224.1 204.6 206.0 207.9 221.0 197.9
Total energy turnover (J s−1)c 181.9 ± 13.6 197.2 ± 16.3 211.0 ± 17.4 216.0 ± 21.0 220.8 ± 20.4 228.1 ± 23.2 209.2 ± 15.3
Open in a separate window
Data represent mean rates for 30 s time intervals and the overall mean for 180 s. Measured data are depicted as means ±s.e.m. for 5 subjects. aNet PCr hydrolysis and lactate accumulation estimations are based on the initial and final values observed in biopsy samples from vastus lateralis obtained in parallel studies with the same experimental protocol (J. Bangsbo, P. Krustrup, J. Gonz.alezález-Alonso & B. Saltin, unpublished; ΔPCr 16.3 mmol (kg wet wt)−1 and Δlactate 22.3 mmol (kg wet wt)−1; see also Bangsbo et al. 1990, for similar measurements) and the kinetics described by Gollnick & Hermansen (Fig. 3, p. 12; 1973) and Sinclair et al. (1999). Heat produced per mole of ATP used was assumed to be 35 kJ for ATP and PCr hydrolysis, 65 kJ for glycogenolysis and 72 kJ for oxidation (Curtin & Woledge, 1978; Woledge & Reilly, 1988). P:O ratio was assumed to be constant throughout exercise and equal to theoretical 3.0. Total metabolic input was calculated as the sum of the energy input from oxygen consumption, net ATP and net PCr hydrolysis and lactate production. bNet ATP hydrolysis estimation (ΔATP 1.1 mmol (kg wet wt)−1) is based on data from Hellsten et al. (1999). cTotal energy turnover was calculated by summing power output and rate of heat production.
An external file that holds a picture, illustration, etc.
Object name is tjp0524-0603-f10.jpg
Figure 10
Total and aerobic energy turnover during dynamic exercise
Total energy turnover (Et) vs. aerobic heat liberation (HVO2) during 180 s of exercise. Note that the difference in heat between Et and HVO2 is accounted for by anaerobic heat liberation.
Oxygen uptake of the thigh gradually increased throughout the exercise period. However, it did not reach a plateau value, although the work rate and thus energy turnover was in each individual above the aerobic capacity. To what extent this continuous elevation in oxygen uptake is due to (i) altered motor unit and fibre type recruitment, (ii) declining mitochondrial P:O ratio, (iii) reduced free energy (ΔG) from the hydrolysis of ATP, (iv) declining crossbridge cycling efficiency, and (v) increasing non-crossbridge ATPase activity (Crow & Kushmerick, 1982; Curtin & Woledge, 1991, 1993; Barclay et al. 1993; Willis & Jackman, 1994; Barclay, 1996) requires further investigations. The possibility cannot be excluded that one or all five of these mechanisms could play a role. The knee-extensor muscles are mixed with regard to their fibre type distribution, usually containing similar amounts of type I and type II muscle fibres (Saltin et al. 1977; Lexell et al. 1983). As the efficiency of type I muscle fibres is likely to be greater than that of type II fibres when contraction velocity is < 25 % of the maximal value (Crow & Kushmerick, 1982; Curtin & Woledge, 1991, 1993; Aagaard et al. 1994; Barclay, 1996), the question arises as to whether a shift in fibre recruitment and/or enhanced motor unit recruitment contributed to the observed increases in oxygen uptake and heat production. Based on T2-weighted MRI images obtained before and after exercise and the glycogen depletion pattern, all knee-extensor muscles are engaged when the intensity of contractions is high (Bangsbo et al. 1992; Richardson et al. 1998; Ray & Dudley, 1998). The rapid increase in muscle temperature in all the locations in the knee-extensor muscles after the first contractions is consistent with this notion. The glycogen depletion pattern reveals a recruitment of all fibre types at this intensity of exercise (Bangsbo et al. 1992). Thus, it is most likely that the relative role of type I fibres is enhanced rather than reduced in the present experiment. Therefore, it appears unlikely that an altered motor unit recruitment pattern influenced to a significant extent the slow increase in muscle V̇O2 and heat production during exercise.
The free energy (ΔG) for ATP hydrolysis will decrease from rest to maximal exercise, since the contribution from the RTlnK′ term of the equation ΔG =ΔG°+RTlnK′ (where ΔG° is the free energy under standard conditions), will decline with the increase in free ADP, Pi and H+, occurring during contraction. It may be estimated that the K′ will typically increase by a factor of 300 or more, comparing resting conditions and end-exercise conditions (see e.g. Ratkevicius et al. 1998), corresponding to a decrease in ΔG by about 15 kJ mol−1. Whether or not the P:O ratio declines with intense exercise is unknown at present; however, in vitro measurements do not suggest that this is the case (Hinckle & Yu, 1979; Willis & Jackman, 1994). Whether a decreasing efficiency of the crossbridge coupling during maximal exercise occurs is also unknown. Judging from NMR measurements of the cost of contraction (moles of ATP hydrolysis per watt of power output) under anaerobic conditions, this does not appear to be the case (Y. Nakagawa, A. Ratkevicius, M. Mizuno & B. Quistorff, unpublished observations). Finally, the possibility cannot be excluded that the non-crossbridge ATPase activity could contribute to the observed elevated heat production (Barclay et al. 1993; Barclay, 1996).
In conclusion, the total skeletal muscle heat production in humans performing intense work was accurately quantified during dynamic knee-extension exercise by summing: (a) heat storage in the contracting muscles, (b) heat removal to the body core by the circulation, and (c) heat release to the skin and environment. At constant power output, heat production increased progressively throughout exercise. This meant that heat production was ∼100 % larger during the final as compared to the initial 5 s of exercise, with half of the increase occurring during the first 38 s. This allows the comparison of metabolic energy input with heat production. Concomitant measurements of aerobic energy metabolism and net lactate release supplemented with data on PCr depletion and muscle lactate accumulation in similar exercise made possible the accurate estimation of the magnitude of energy yield from these three metabolic pathways throughout exercise. The match was found to be very close provided correct values for the heat liberated from the breakdown of PCr, anaerobic glycolysis and oxidative phosphorylation were applied.
Go to:
Acknowledgments
Special thanks are given to the subjects in this study. The excellent engineering assistance of Flemming Jensen is acknowledged. The authors also thank Markus Novak (Rigshospitalet) for his work collecting the MRI scans. This study was supported by a grant from The Danish National Research Foundation (504-14). J.G.-A. was supported by a Marie Curie Research Training Grant (FMBICT950007).
Go to:
References
Aagaard P, Simonsen EB, Trolle M, Bangsbo J, Klausen K. Moment and power generation during maximal knee extensions performed at low and high speeds. European Journal of Applied Physiology. 1994;69:376–381. [PubMed] [Google Scholar]
Andersen P, Adams RP, Sjøgaard G, Thorboe A, Saltin B. Dynamic knee extension as model for study of isolated exercising muscle in humans. Journal of Applied Physiology. 1985;59:1647–1653. [PubMed] [Google Scholar]
Andersen P, Saltin B. Maximal perfusion of skeletal muscle in man. The Journal of Physiology. 1985;366:233–249. [PMC free article] [PubMed] [Google Scholar]
Ardevol A, Adan C, Remesar X, Fernández-López JA, Alemany M. Hind leg heat balance in obese Zucker rats during exercise. Pflügers Archiv. 1998;435:454–464. [PubMed] [Google Scholar]
Åstrand I. Aerobic work capacity in men and women with special reference to age. Acta Physiologica Scandinavica. 1960;49(suppl. 169):67–158. [PubMed] [Google Scholar]
Bangsbo J, Gollnick PD, Graham TE, Juel C, Kiens B, Mizuno M, Saltin B. Anaerobic energy production and O2 deficit debt relationship during exhaustive exercise in humans. The Journal of Physiology. 1990;422:539–559. [PMC free article] [PubMed] [Google Scholar]
Bangsbo J, Graham TE, Kiens B, Saltin B. Elevated muscle glycogen and anaerobic energy production during exhaustive exercise in man. The Journal of Physiology. 1992;451:205–227. [PMC free article] [PubMed] [Google Scholar]
Barclay CJ. Mechanical efficiency and fatigue of fast and slow muscles of the mouse. The Journal of Physiology. 1996;497:781–794. [PMC free article] [PubMed] [Google Scholar]
Barclay CJ, Curtin NA, Woledge RC. Changes in crossbridge and non-crossbridge energetics during moderate fatigue of frog muscle fibres. The Journal of Physiology. 1993;468:543–555. [PMC free article] [PubMed] [Google Scholar]
Barcroft H, Edholm OG. The effect of temperature on blood flow and deep temperature in the human forearm. The Journal of Physiology. 1942;102:5–20. [PMC free article] [PubMed] [Google Scholar]
Benedict FG, Cathcart EP. Muscular Work: A Metabolic Study with Special Reference to the Efficiency of the Human Body as a Machine. 1913. Carnegie Institution of Washington, Publication No. 187, Washington, DC, USA.
Crow MT, Kushmerick MJ. Chemical energetics of slow- and fast-twitch muscles of the mouse. Journal of General Physiology. 1982;79:147–166. [PMC free article] [PubMed] [Google Scholar]
Curtin NA, Woledge RC. Energy changes and muscular contraction. Physiological Reviews. 1978;58:690–761. [PubMed] [Google Scholar]
Curtin NA, Woledge RC. Efficiency of energy conversion during shortening of muscle fibres from the dogfish Scyliorhinus canicula. Journal of Experimental Biology. 1991;158:343–353. [PubMed] [Google Scholar]
Curtin NA, Woledge RC. Efficiency of energy conversion during sinusoidal movement of white muscle fibres from dogfish Scyliorhinus canicula. Journal of Experimental Biology. 1993;183:137–147. [PubMed] [Google Scholar]
Edwards RHT, Harris RC, Hultman E, Kaijser L, Koh D, Nordesjö L-O. Effect of temperature on muscle energy metabolism and endurance during successive isometric contractions, sustained to fatigue, of the quadriceps muscle in man. The Journal of Physiology. 1972;220:335–352. [PMC free article] [PubMed] [Google Scholar]
Edwards RHT, Hill DK, Jones DA. Heat production and chemical changes during isometric contractions of the human quadriceps muscle. The Journal of Physiology. 1975;251:303–315. [PMC free article] [PubMed] [Google Scholar]
Ganz W, Swan HJC. Measurements of blood flow by the thermodilution technique. In: Bloomfield DA, editor. Dye Curves: The Theory and Practice of Indicator Dilution. Baltimore: University Park Press; 1974. pp. 245–266. [Google Scholar]
Gollnick PD, Hermansen L. Biochemical adaptations to exercise: anaerobic metabolism. In: Wilmore JH, editor. Exercise and Sport Sciences Reviews. Vol. 1. New York: Academic Press; 1973. pp. 1–43. [PubMed] [Google Scholar]
Hellsten Y, Richter EA, Kiens B, Bangsbo J. AMP deamination and purine exchange in human skeletal muscle during and after intense exercise. The Journal of Physiology. 1999;520:909–919. [PMC free article] [PubMed] [Google Scholar]
Hensel H, Bock KD. Durchblutung und Wärmeleiffähigkeit des menschlichen Muskels. Pflügers Archiv. 1955;260:361–367. [PubMed] [Google Scholar]
Hill AV, Woledge RC. An examination of absolute values in myothermic measurements. The Journal of Physiology. 1962;162:311–333. [PMC free article] [PubMed] [Google Scholar]
Hinckle PC, Yu ML. The phosphorous/oxygen ratio of mitochondrial oxidative phosphorylation. Journal of Biological Chemistry. 1979;254:2450–2455. [PubMed] [Google Scholar]
Kushmerick MJ, Larsen RE, Davies RE. The chemical energetics of muscle contraction. I. Activation heat, heat of shortening and ATP utilization for contraction. Proceedings of the Royal Society. 1969;B 174:293–313. [PubMed] [Google Scholar]
Lexell J, Henriksson-Larsén K, Sjöstrom M. Distribution of different fibre types in human skeletal muscles. 2. A study of cross-sections of whole m. vastus lateralis. Acta Physiologica Scandinavica. 1983;117:115–122. [PubMed] [Google Scholar]
Meyer RA. A linear model of muscle respiration explains monoexponential phosphocreatine changes. American Journal of Physiology. 1988;254:C548–553. [PubMed] [Google Scholar]
Pennes HH. Analysis of tissue and arterial blood temperatures in the resting human forearm. Journal of Applied Physiology. 1948;1:93–122. [PubMed] [Google Scholar]
Rådegran G, Saltin B. Muscle blood flow at onset of dynamic exercise in humans. American Journal of Physiology. 1998;274:H314–322. [PubMed] [Google Scholar]
Rasband WS, Bright DS. NIH Image: A public domain image processing program for the Macintosh. Microbeam Analysis Society Journal. 1995;4:137–149. [Google Scholar]
Ratkevicius A, Mizuno M, Povilonis E, Quistorff B. Energy metabolism of the gastrocnemius and soleus muscles during isometric voluntary and electrically induced contractions in man. The Journal of Physiology. 1998;507:593–602. [PMC free article] [PubMed] [Google Scholar]
Ray CA, Dudley GA. Muscle use during dynamic knee extension: implication for perfusion and metabolism. Journal of Applied Physiology. 1998;85:1194–1197. [PubMed] [Google Scholar]
Richardson RS, Frank LR, Haseler LJ. Dynamic knee-extensor and cycle exercise: functional MRI of muscular activity. International Journal of Sports Medicine. 1998;19:182–187. [PubMed] [Google Scholar]
Saltin B, Henriksson J, Nygaard E, Andersen P, Jansson E. Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Annals of the New York Academy of Sciences. 1977;301:3–29. [PubMed] [Google Scholar]
Saugen E, Vøllested NK. Non-linear relationship between heat production and force during voluntary contractions in man. Journal of Applied Physiology. 1995;79:2043–2049. [PubMed] [Google Scholar]
Saugen E, Vøllested NK. Metabolic heat production during fatigue from voluntary repetitive isometric contractions in humans. Journal of Applied Physiology. 1996;81:1323–1330. [PubMed] [Google Scholar]
Sinclair AS, Montain SJ, Matott RP, Zientara GP, Jolesz FA, Fielding RA. Effects of creatine supplementation on the energy cost of muscle contraction: a 31P-MRS study. Journal of Applied Physiology. 1999;87:116–123. [PubMed] [Google Scholar]
von Döbeln W. Human standard and maximal metabolic rate in relation to fat-free body mass. Acta Physiologica Scandinavica. 1956;37(suppl. 126):1–79. [PubMed] [Google Scholar]
Wilkie DR. Thermodynamics and interpretations of biological heat measurements. Progress in Biophysics and Biophysical Chemistry. 1960;10:259–289. [PubMed] [Google Scholar]
Wilkie DR. Heat work and phosphorylcreatine breakdown in muscle. The Journal of Physiology. 1968;195:157–183. [PMC free article] [PubMed] [Google Scholar]
Willis WT, Jackman MR. Mitochondrial function during heavy exercise. Medicine and Science in Sports and Exercise. 1994;26:1347–1354. [PubMed] [Google Scholar]
Woledge RG, Reilly PJ. Molar enthalpy change for hydrolysis of phosphorylcreatine under conditions in muscle cells. Biophysical Journal. 1988;54:97–104. [PMC free article] [PubMed] [Google Scholar]
Articles from The Journal of Physiology are provided here courtesy of The Physiological Society
OTHER FORMATS
PDF (436K)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 1364233 | https://no.wikipedia.org/wiki/Treningsfysiologi | Treningsfysiologi | Treningsfysiologi er studiet av de akutte reaksjoner og kroniske tilpasninger av fysisk trening. I tillegg studerer mange treningsfysiologer effekten av trening på patologi, og mekanismene som gjør at trening kan redusere eller reversere sykdom. En treningsfysiologs studiefelt kan kan omfatte, men er ikke begrenset til, biokjemi, bioenergi, hjerte-funksjon, hematologi, biomekanikk, skjelettmuskelfysiologi, nevroendokrin funksjon, og sentrale og perifere nervesystemet. Treningsfysiologi jobber i spennet fra grunnforskere, til kliniske forskere, klinikere, til idrettstrenere.
Energiforbruk
Mennesket har kapasitet til å bruke energi i mange timer med vedvarende anstrengelse. For eksempel, en person som sykler med en hastighet på 26,4 km / t (16.4 mph) gjennom 8204 km (5098 miles) over 50 påfølgende dager bruker totalt 1145 MJ (273,850 kcal; 273850 kalorier) med en gjennomsnittlig utgangseffekt på 182,5 W
Skjelettmuskler forbrenner 90 mg (0,5 mmol) glukose hvert minutt under kontinuerlig aktivitet (for eksempel under gjentatte bevegelser i kneet), genererer ≈24 W mekanisk energi, og siden muskelenergi konverterterer bare 22-26% effektivt, ≈76 W varmeenergi. Hvileskjelettmuskulatur har en basal forbrenning (hvilende energiforbruk) på 0,63 W / kg som utgjør 160 gangers forskjell mellom energiforbruket for inaktive og aktive muskler. For kort varighet av muskulær anstrengelse, kan energiforbruket bli langt større: når en voksen mann hopper opp fra en knebøy kan han mekanisk generere 314 W / kg. Slike raske bevegelser kan hos andre dyr som for eksempel sjimpansen, og noen små øgler, generere den dobbelte energimengden av dette.
Dette energiforbruk er svært stor sammenlignet med basal hvilemetabolisme hos den voksne menneskekroppen. Denne mengden varierer noe med størrelse, kjønn og alder, men er vanligvis mellom 45 W og 85 W. Totalt energiforbruk (TEE) på grunn av energi musklene har brukt er mye høyere og avhenger av det gjennomsnittlige nivået av fysisk arbeid og mosjon gjort i løpet av en dag. Mosjon, særlig hvis over svært lange perioder, dominerer energimetabolismen i kroppen. Energiforbruket ved fysisk aktivitet korrelerer sterkt med kjønn, alder, vekt, puls, og V [O.sub.2max] til en person, under fysisk aktivitet.
Referanser
Evolusjonsbiologi
Menneskelig evolusjon
Treningsteori | norwegian_bokmål | 0.55114 |
below_temperature_muscles_not_work/Shivering.txt | Shivering (also called shuddering) is a bodily function in response to cold and extreme fear in warm-blooded animals. When the core body temperature drops, the shivering reflex is triggered to maintain homeostasis. Skeletal muscles begin to shake in small movements, creating warmth by expending energy. Shivering can also be a response to fever, as a person may feel cold. During fever, the hypothalamic set point for temperature is raised. The increased set point causes the body temperature to rise (pyrexia), but also makes the patient feel cold until the new set point is reached. Severe chills with violent shivering are called rigors. Rigors occur because the patient's body is shivering in a physiological attempt to increase body temperature to the new set point.
Biological basis[edit]
Located in the posterior hypothalamus near the wall of the third ventricle is an area called the primary motor center for shivering. This area is normally inhibited by signals from the heat center in the anterior hypothalamic-preoptic area but is excited by cold signals from the skin and spinal cord. Therefore, this center becomes activated when the body temperature falls even a fraction of a degree below a critical temperature level.
Increased muscular activity results in the generation of heat as a byproduct. Most often, when the purpose of the muscle activity is to produce motion, the heat is wasted energy. In shivering, the heat is the main intended product and is utilized for warmth.
Newborn babies, infants, and young children experience a greater (net) heat loss than adults because of greater surface-area-to-volume ratio. As they cannot shiver to maintain body heat, they rely on non-shivering thermogenesis. Children have an increased amount of brown adipose tissue (increased vascular supply, and high mitochondrial density), and, when cold-stressed, will have greater oxygen consumption and will release norepinephrine. Norepinephrine will react with lipases in brown fat to break down fat into triglycerides. Triglycerides are then metabolized to glycerol and non-esterified fatty acids. These are then further degraded in the needed heat-generating process to form CO2 and water. Chemically, in mitochondria, the proton gradient producing the proton electromotive force that is ordinarily used to synthesize ATP is instead bypassed to produce heat directly.
Shivering can also appear after surgery. This is known as postanesthetic shivering.
In humans, shivering can also be caused by mere cognition. This is known as psychogenic shivering.
Shivering and the elderly[edit]
The functional capacity of the thermoregulatory system alters with aging, reducing the resistance of elderly people to extreme external temperatures. The shiver response may be greatly diminished or even absent in the elderly, resulting in a significant drop in mean deep body temperature upon exposure to cold. Standard tests of thermoregulatory function show a markedly different rate of decline of thermoregulatory processes in different individuals with ageing.
See also[edit]
Goose bumps
Myoclonus
Post micturition convulsion syndrome
Chattering teeth
Tremor | biology | 6338308 | https://sv.wikipedia.org/wiki/Studnice%20%28ort%20i%20Tjeckien%2C%20lat%2050%2C42%2C%20long%2016%2C09%29 | Studnice (ort i Tjeckien, lat 50,42, long 16,09) | Studnice är en ort i Tjeckien. Den ligger i den nordöstra delen av landet, km öster om huvudstaden Prag. Studnice ligger meter över havet och antalet invånare är .
Terrängen runt Studnice är platt åt sydväst, men åt nordost är den kuperad. Den högsta punkten i närheten är meter över havet, km öster om Studnice. Runt Studnice är det tätbefolkat, med invånare per kvadratkilometer. Närmaste större samhälle är Trutnov, km nordväst om Studnice. Trakten runt Studnice består till största delen av jordbruksmark.
Trakten ingår i den hemiboreala klimatzonen. Årsmedeltemperaturen i trakten är °C. Den varmaste månaden är juli, då medeltemperaturen är °C, och den kallaste är januari, med °C.
Kommentarer
Källor
Externa länkar
Orter i Hradec Králové (region) | swedish | 1.17197 |
shut_down_conciousness/PMC2923650.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List HHS Author Manuscripts PMC2923650
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of nihpa
Emerg Med Clin North Am. Author manuscript; available in PMC 2011 Aug 1.
Published in final edited form as:
Emerg Med Clin North Am. 2010 Aug; 28(3): 571–594.
doi: 10.1016/j.emc.2010.03.003
PMCID: PMC2923650
NIHMSID: NIHMS193971
PMID: 20709244
Traumatic Alterations in Consciousness: Traumatic Brain Injury
Brian J. Blyth, MDa and Jeffrey J. Bazarian, MD, MPHb
Author information Copyright and License information PMC Disclaimer
The publisher's final edited version of this article is available at Emerg Med Clin North Am
Go to:
Abstract
Mild traumatic brain injury (mTBI) refers to the clinical condition of transient alteration of consciousness as a result of traumatic injury to the brain. The priority of emergency care is to identify and facilitate the treatment of rare but potentially life threatening intra-cranial injuries associated with mTBI through the judicious application of appropriate imaging studies and neurosurgical consultation. Although post-mTBI symptoms quickly and completely resolve in the vast majority of cases, a significant number of patients will complain of lasting problems that may cause significant disability. Simple and early interventions such as patient education and appropriate referral can reduce the likelihood of chronic symptoms. Although definitive evidence is lacking, mTBI is likely to be related to significant long-term sequelae such as Alzheimer's disease and other neurodegenerative processes.
Keywords: concussion, trauma, brain, injury
Go to:
Introduction
The purpose of this review is to provide an overview of mild traumatic brain injury (mTBI) in a form useful for Emergency Physicians. This is a disease with considerable public health impact and is the subject of a vast amount of current research. Our discussion of this disease is necessarily limited both by space constraints and the interests of our target audience. For more detailed discussions of various topics covered in this review the reader is directed to the publications listed in Table 1.
Table 1
Recommended Further Reading
Topic Reference Notes
Epidemiology Summers C.R., lvins B., and Schwab K. A.14 Brief review of literature pertaining to the epidemiology of both military and civilian TBI.
Biomechanics LaPlaca M.C., Simon C. M., Prado G. R., et al.18 Thorough review of basic biomechanics relevant to CNS trauma including discussion of experimental models of injury.
Mechanisms of LOC Shaw N. A.33 Detailed account of the historical development and current theories of altered conciousness resulting from TBI.
Acute Clinical Management Jagoda A. S., Bazarian J. J., Bruns J.J., Jr., et al.42 American College of Emergency Physicians and Centers for Disease Control clinical policies for the acute management of adult mTBI.
Sub-acute and Chronic Clinical Management VA/DoD Clinical Practice Guideline for Management of Concussion/mTBI 1 Definitive clinical guidelines for sub-acute and chronic management of adult mTBI (>1 week post injury).
Long Term Health Effects IOM (Institute of Medicine). Gulf War and Health, Volume 7 24 Comprehensive review of evidence for long term consequences of TBI.
Open in a separate window
Definition of Traumatic Brain Injury
Traumatic Brain Injury (TBI) is defined as any traumatically induced structural injury or physiological disruption of brain function as a result of an external force. It is manifested by one or more clinical signs occurring immediately afterwards including a loss, decreased, or altered level of consciousness, amnesia, neurologic deficit, or intracranial lesion 1. External forces may include direct impact of the head with another object, indirect forces from acceleration/deceleration, or a blast injury. The Glasgow Coma Score (GCS) has traditionally been used to classify TBI as mild (GCS 13-15), moderate (GCS 9-12), or severe (GCS 3-8). A more recent classification scheme for TBI uses length of loss of consciousness (LOC), alteration of consciousness (AOC), and post traumatic amnesia (PTA) as well as imaging findings to categorize TBI (Table 2) 1. It is important to stress that mild TBI (mTBI) is clinically defined based solely on self-reported or observed symptoms and often occurs with normal neuroimaging. Indeed, the mTBI definition included in Table 1 classifies TBI with positive neuroimaging as at least moderate in severity. Two older, but still commonly used mTBI definitions by the American Congress of Rehabilitation Medicine 2 and the Centers for Disease Control and Prevention 3 define patients as mild if they have positive neuroimaging findings but meet all other clinical criteria for mTBI. Patients who have abnormal intracranial imaging but otherwise meet a clinical definition of mTBI are referred to as complicated mTBI 4.
Table 2
Clinical Criteria for TBI Severity
Criteria Mild Moderate Severe
Structural imaging Definition dependent* Normal or abnormal Normal or abnormal
Loss of consciousness (LOC) 0-30 minutes > 30 min and < 24 hrs > 24hrs
Alteration of consciousness (AOC)** A moment up to 24 hours > 24 hours. Severity based on other criteria
Post Traumatic amnesia (PTA) 0-1 day > 1 and < 7 days > 7 days
GCS (best score in first 24 hours) 13-15 9-12 < 9
Open in a separate window
*Patients who otherwise meet the clinical criteria for mTBI but have intracranial imaging abnormalities may be classified as complicated mTBI or moderate TBI depending on the definition used.
**Alteration of mental status must be immediately related to the trauma to the head. Typical symptoms may include looking or feeling dazed, confusion, difficulty thinking clearly or responding appropriately to mental status questions, or inability to describe events immediately before or after the traumatic event.
Adapted from: IOM (Institute of Medicine). Gulf War and Health, Volume 7: Long-term Consequences of Traumatic Brain Injury. Washington, D.C.: The National Academies Press; 2009.
Concussion is a common term for mild traumatic brain injury (mTBI) and will be used interchangeably within this article. Our understanding of traumatic brain injury remains rudimentary relative to many other medical problems of similar magnitude. A symptom based classification uses the description of symptoms evident on history and physical exam to classify illness. This method is imprecise, often grouping disparate pathophysiological process together as single clinical entities. This is particularly problematic in moderate and severe TBI where multiple injury processes as evidenced by heterogeneous imaging findings are often present simultaneously. As knowledge of a disease increases and diagnostic tools improve, a more sophisticated classification emerges that may include anatomic, physiologic, metabolic, immunologic, and genetic factors. TBI has been the subject of intensive research in recent years and recommendations for improved classification of this diverse disease are beginning to appear in the literature 5.
Epidemiology
An estimated 1.4 million Americans presented to emergency departments for medical care after TBI each year between 1995 and 2001 6. Over 1 million of these patients had mTBI. The incidence of ED-attended concussion is 444/100,000 in the United States 7 however it is thought that nearly 40% of patients suffering mTBI do not seek hospital based care, and 25% do not report their injuries to health care providers at all 8. An estimated 5-25% of all patients with concussion have post-concussive symptoms or other cognitive deficits that persist beyond one year 9-11. This number is greater than the annual incidence of multiple sclerosis, Parkinson's disease, myasthenia gravis, and Huntington's disease combined 12. TBI is more common in men than women with 60% of TBI occurring in males 6. Young children, adolescents and the elderly suffer the highest rates of TBI 6. The most common mechanisms of TBI are falls, automobile accidents, being struck by or against an object, and assault. The rate of hospitalizations for TBI fell dramatically during the 1980's and 90's 6. Wounded veterans from the wars in Iraq and Afghanistan may represent a non-trivial increase in patients living with mTBI. Approximately 20% of troops returning from combat deployments in Iraq have clinician confirmed mTBI. These patients typically have TBI resulting from a blast 13 and often have comorbid PTSD or depression 14.
Biomechanics
Traumatic injury results from the transfer of energy from the environment to tissue above the amount that can be absorbed without dysfunction. Biomechanics is the study of the interaction of forces and physical responses in biological systems. Traumatic insults generally occur over short periods of time and are referred to as dynamic loading. Dynamic loading includes both direct or impact loading, as well as impulsive loading where no physical contact occurs. The loads absorbed by the brain after trauma generally include linear and rotational components called angular loads. The rate and duration of the insult are important because loads applied at high rates tend to result in more damage 18. For example, the force involved in punching a wall can also be applied by pressing your fist against that same wall for a few minutes: the former instance results in a boxer's fracture whereas the latter does not. Focal injury such as contusion results from direct loading and often occurs in the absence of widespread injury. In contrast, diffuse axonal injury (DAI) often occurs as a result of the rotational acceleration accompanying indirect loading 15. Humans are particularly susceptible given their large cranium connected to the trunk by relatively weak neck musculature. Rotational acceleration produces substantial and widespread strains within the brain resulting from both acceleration and deceleration. These diffuse strains lead to differential movement of the brain relative to the skull which can cause hemorrhage. Shear strain is most prominent after rotational injury, and brain tissue is particularly sensitive to this type of strain 16. In animal models, rotational acceleration is required to produce concussion whereas isolated linear acceleration produced contusions and subdural hematomas but no loss of consciousness 17.
Pathophysiology
The initial traumatic insult results in mechanical damage including rupture of cellular and vascular membranes with release of intracellular contents, ultrastructural damage of axons, and changes in cerebral blood flow 19-20. Subsequent metabolic derangement includes widespread release of excitatory neurotransmitters such as glutamate, severe dysregulation of calcium homeostasis, energy failure due to ATP depletion, free radical generation, and cell death by necrotic and apoptotic pathways 20-21. More global consequences of the traumatic insult include increased intra-cranial pressure, decreased cerebral blood flow, tissue ischemia, cerebral edema, and functional blood brain barrier dysfunction 22-23. Following the initial damage, repair and recovery processes begin through the removal of cellular debris, glial scar formation, and plastic changes in neural networks 24. Due to the difficulties studying human mTBI, the mechanisms described in this section were derived principally from animal studies and, to a lesser degree, from humans with severe TBI (sTBI). Similar processes are thought to occur in human mTBI
Putative Causes of Altered Consciousness in mTBI
The definitive causes of altered consciousness are not known. Loss of consciousness requires either loss of the function of both cerebral hemispheres or of the reticular activating system. Several plausible hypothetical mechanisms have been proposed for the alteration of consciousness that occurs with mTBI. These include the reticular, pontine-cholinergic system, centripetal, and convulsive hypotheses. The reticular activating system (RAS) resides in the brainstem reticular formation which extends from the top of the spinal column to the rostral midbrain with extensions into the thalamus and hypothalamus. The RAS is excited by input from surrounding sensory tracts and transmits this excitation to the cortex to induce generalized cortical and behavioral arousal. In the absence of input from the RAS, consciousness is impaired.
Under the reticular hypothesis of concussion, loss of consciousness after brain trauma results from a disturbance or depression of the activity of polysynaptic pathways within the RAS 25. It is not completely understood how a traumatic dysfunction of the RAS occurs however it is believed to result from shearing or tensile strains on RAS pathways at the cranio-cervical junction. Neuropathological evidence for this is limited. The hypothesis also fails to address traumatic amnesia. A further difficulty is that EEG findings do not support depression of the RAS in concussion.
The pontine-cholinergic system hypothesis differs from the reticular activating system hypothesis in that RAS dysfunction is thought to occur as a consequence of trauma-induced activation of the inhibitory cholinergic system of the dorsal pontine tegmentum 26. In animal models, injection of cholinergic agonists into the brainstem induces unconsciousness 27-28 whereas similar injections of cholinergic antagonists reduce the duration of traumatic unconsciousness 29. Furthermore, EEG studies show widespread neuronal discharge after concussion and elevated acetylcholine is found in the CSF of patients after TBI. However, it is not clear that activation of this system can produce loss of consciousness due to RAS suppression.
The centripetal hypothesis posits that sudden rotational forces cause shearing strains and stresses that result in functional decoupling of nerve fibers 17. The depth of this functional decoupling is directly related to the extent of rotational acceleration delivered to the brain. Also, with greater rotational acceleration the likelihood of mechanical injury to fibers increases. Lower inertial forces that result in functional decoupling between the subcortex or diencephalon and the cortex may result in amnesia or confusion without LOC. Furthermore, greater forces resulting in decoupling between more superficial structures and the mesenephalon result in LOC. This hypothesis nicely explains post-traumatic amnesia and dazed states, however it also requires very high energy injuries to cause full loss of consciousness. Consequently, patients with LOC would often have accompanying structural brain injury which is simply is not observed.
Patients with concussion have similar symptoms to those who have experienced generalized epileptic seizures or electro convulsive therapy (ECT). This overlap of symptoms has led to speculation that similar pathophysiological events occur in all three conditions. Close observation of human patients and animal models shows that concussive injury generally causes an initial convulsive event followed by a longer and more prominent paralytic phase 30-31. EEG recordings from concussed animals also show initial, transient epileptiform activity. According to the convulsive hypothesis the symptoms associated with concussion are due to direct injury to neurons resulting in hyperexcitability and widespread membrane depolarization followed by neuronal exhaustion 32. These two neuronal states correspond to the convulsive and paralytic phases, respectively.
The convulsive hypothesis is able to reasonably account for a broader range of post-concussive behaviors than its competitors including LOC, amnesia, convulsive movements, autonomic disturbances, and the dazed or “dinged” state 33. While this hypothesis does a better job than the others at providing a unified explanation for the broad range of symptoms observed as an acute result of mTBI, it does not account for the structural abnormalities that occur as a result of mTBI. In summary, none of the individual hypotheses currently available explain all the findings seen with mTBI. Given the often complimentary strengths and weakness of the four hypotheses discussed above, it seems likely that the mechanisms of altered consciousness after TBI may be due to a combination of processes. For a detailed explanation of these hypotheses, the reader is directed to the excellent review by Shaw 33.
Go to:
Diagnosis of mTBI
Clinical Presentation
A 28-year-old male presents to the ED after a motor vehicle accident. He was the restrained driver in a car that skidded of the road in icy conditions and collided head on with a tree at 50 mph. There was airbag deployment. The paramedic reports that he was unconscious initially but that he was alert and oriented during transport.
Differential Diagnosis The diagnosis of mTBI is made clinically and relies heavily on the history obtained from the patient and any witnesses. Obtaining a reliable history is often difficult because of post-traumatic amnesia, persistent altered mental status, or intoxication, a frequent comorbid factor in mTBI patients. Diagnoses with similar presentations include seizure, syncope, intoxication, malingering, anxiety, and other psychiatric conditions.
Clinical Criteria Several clinical criteria for the diagnosis of mTBI exist 1-3. Concussion or mTBI is defined as a loss of consciousness of less than 30 minutes or amnesia lasting less than 24 hours, or any period of altered mental status at the time of injury. In conjunction, patients must also have a GCS of 13-15 and normal structural imaging to meet the criteria for mTBI. Lower GCS scores classify the patient as having moderate (GCS 9-12) or severe (GCS 3-8) TBI.
Imaging The imaging study of choice in mTBI is non-contrast head computed tomography (CT). This study is preferred over others because it is sensitive for traumatic injuries that require neurosurgical intervention including acute bleeding, increased intra-cranial pressure, and skull fracture. Although as many as 15% of mTBI patients will have an acute injury detected by non-contrast head CT, only 1% of those abnormalities require neurosurgical intervention 34-40. Other imaging modalities are of limited use for the clinical evaluation of mTBI patients and are not recommended. Although magnetic resonance imaging (MRI) is 30% more sensitive than CT for the detection of traumatic abnormalities after mTBI 41, there is no evidence that it identifies more patients requiring neurosurgical intervention 42. More exotic imaging modalities including functional MRI, diffusion tensor imaging MRI, magnetic resonance spectroscopy, single photon emission computed tomography, and positron emission tomography are valuable research tools but do not have proven clinical utility.
Decision Rules The low rate of clinically important brain injury seen on head CT obtained acutely after mTBI has resulted in efforts to minimize unnecessary studies through the application of rigorously, validated, clinical decision rules. Two major decision rules applying to adult mTBI patients include the Canadian CT Head Rule 43 and the New Orleans Criteria 44. For patients with a GCS of 15, both of these rules have equivalent sensitivities for detecting injuries requiring neurosurgical intervention, however the Canadian CT Head Rule has a higher specificity for some clinical outcomes and its use may reduce imaging rates 45. In pediatric populations, increased concern for radiation exposure and the potential requirement for sedation make minimizing unnecessary CT after mTBI even more compelling than in adult populations. Kupperman and colleagues recently reported a very sensitive decision rule that identifies low risk children who do not require a head CT after mTBI 46. Indications for obtaining head CT after mTBI based on these decision rules are summarized in Figure 1.
An external file that holds a picture, illustration, etc.
Object name is nihms193971f1.jpg
Figure 1
Indications for Obtaining Non-contrast Head CT after TBI
Non-contrast head CT is the study of choice to evaluate TBI patients for clinically important neurotrauma. Clinically important neurotrauma is defined as any traumatically induced intracranial injury that requires neurosurgical intervention or requires hospital admission and neurosurgical follow-up. Clinically important TBI occurs rarely after mTBI, therefore minimizing unnecessary head CT scans is desirable. This figure integrates validated decision rules for both adult45 and pediatric patients46 designed to minimize unnecessary CT scans after mTBI.
Biomarkers There is substantial interest in developing protein biomarkers obtained from serum to aid the diagnosis and guide the treatment of TBI of all severities. While several potential biomarkers have been studied 47-53, to date only serum S100B has accepted clinical utility for mTBI. Specifically, elevated S100B has a high negative predictive value for clinically important injury on head CT after mTBI. In a large cohort, elevated S100B was 99% sensitive for the detection of injury on CT scan 54 prompting the use of this test as the clinical standard of care in several European countries. The test has the added advantage of not being affected by concomitant alcohol intoxication 55. Although not yet FDA approved in the US, the American College of Emergency Physicians recently issued a guideline stating that for mTBI patients with serum S100B concentrations of less than 0.1 μg/ml measured within 4 hours of injury, it is reasonable to consider not obtaining a head CT 42. Multi-center studies to evaluate the accuracy of this test in US patient populations are currently underway and may provide the data necessary for FDA approval.
Go to:
Treatment of mTBI
Clinical Presentation
After receiving the prehospital provider's report, you note that the patient is complaining of headache, back pain and abdominal pain. He has no significant past medical history and takes no medications. Pertinent findings on exam include a slight tachycardia, an abrasion on his forehead, mild tenderness in his upper abdomen, and diffuse tenderness over his lumbar spine. He is alert and oriented but cannot remember any events since the accident. Otherwise, he has a normal neurologic exam.
ED Priorities Initial assessment of the mTBI patient in the ED is focused on identifying patients who may require medical or neurosurgical intervention for the treatment of increased intracranial pressure or an expanding mass lesion. Patients with “red flag” conditions such as altered mental status, papillary asymmetry, seizures, repeated vomiting, double vision, worsening headache, motor or sensory deficits, or ataxia should have an emergent non-contrast head CT scan performed. See Figure 1 for further imaging recommendations.
Patients with intra-cranial imaging abnormalities or declining mental status require immediate neurosurgical consultation. Worsening mental status is typically due to increasing intracranial pressure (ICP) leading to compromised cerebral blood flow and oxygen delivery. For these patients, airway management with endotracheal intubation to protect against aspiration as well as to control ventilation should be considered. Non-surgical management also includes mitigating ICP increases by raising the head of the bed to 30 degrees and treatment with hyperosmolar agents such as IV mannitol. Finally, brief periods of hyperventilation can also reduce dangerous ICP increases. Mechanistically, hyperventilation causes vasoconstriction and reduces intracranial pressure by decreasing cerebral blood flow. Overaggressive hyperventilation can result in ICP decreases at the expense of adequate tissue perfusion. Therefore, prolonged hyperventilation should be used only when other therapies have failed.
Clinical Presentation
You obtain imaging studies, including computed tomography of head, neck, abdomen and pelvis which reveal no traumatic injuries. After returning from radiology, the patient does not recall meeting you. Although is head CT did not reveal a traumatic injury, he is admitted for observation overnight due to his persistent anterograde amnesia. He has an uneventful night and his amnesia resolves. He receives detailed discharge instructions that include a description of “red flag conditions”, common post-concussive symptoms, and reassurance that the vast majority of patients recover completely from concussion. He is advised to avoid activities that exacerbate his symptoms, to take acetaminophen as needed for headache, and to follow up with a local concussion clinic in one week if he is having any persistent discomfort from his concussion. He is accompanied home by his fiancée who will stay with him over the next 24 hours.
Acute Phase: Within One Week of Injury After evaluating for “red flag” signs and symptoms (see ED Priorities), a thorough history of symptoms including loss or alteration of consciousness, headache, irritability, unsteadiness, vertigo, photophobia, or phonophobia should be obtained. The physical exam includes a focused neurological exam including assessment of cranial nerves, postural instability, visual function, and mental status. Non-contrast head CT should be obtained when indicated (see Figure 1). Neurosurgical consultation is necessary for patients with imaging abnormalities. These patients are often admitted for 24 hours for ongoing mental status monitoring and repeat head CT prior to discharge. Patients in whom imaging was not indicated or with a normal head CT may be safely discharged 56-58.
Discharge instructions for mTBI patients include two principle elements: symptoms requiring immediate re-evaluation (see ED Priorities) and post-concussive symptom education. Post-concussive symptoms include headache, sleep disturbances, vertigo, nausea, fatigue, sensitivity to light or noise, attention and concentration problems, depression, and emotional lability. The vast majority of adults with post-concussive symptoms recover within 3-12 months 59. Early patient education that includes likely post-concussive symptoms and reassurance about an expected positive recovery has been shown to speed recovery and decrease post-concussive symptoms 60-62. Headache should be managed with acetaminophen. Non-steroidal anti-inflammatory drugs (NSAIDs) may be used in patients with negative neuroimaging but should be deferred until 48 hours after injury if imaging was not obtained. In addition, narcotics should be avoided in the treatment of post-traumatic headache.
Pharmacologic treatment of other post-concussive symptoms is not recommended in the acute phase. Rather, patients with symptoms other than headache should be advised to rest and encouraged to return to normal activity as soon as possible. However, individuals whose normal activity includes a high risk for re-injury should have careful evaluation of their symptoms and exam findings with consideration of their specific activities that result in a high injury risk. Specific limitations on activity may be recommended for these patients to mitigate their individual risk. Patients reporting fatigue may be given a graded return to work or activity. For patients with normal activities involving significant physical activity, exertional testing may be performed. If this results in a return of symptoms, a monitored progressive return to these activities as tolerated should be recommended 1.
Clinical Presentation
Two weeks after the initial injury, the patient continues to suffer from frequent headaches that are only slightly relieved by acetaminophen. He also complains of increased irritability, sleepiness, and difficulty concentrating. During an initial follow up visit, a detailed history and physical examination fails to reveal comorbid psychiatric or physical problems including PTSD, depression, substance abuse, hypertension, cervical spine abnormalities, sinus infections, or visual acuity deficits. However, the patient indicates that he has not been sleeping well due to persistent headache. He is started on an NSAID for his headaches, and provided with education regarding good sleep hygiene and relaxation techniques. He is also advised to begin a regular exercise program. A follow up appointment is scheduled in 4 weeks.
Initial Management of Post-concussive Symptoms This section provides an overview of treatment for the initial treatment of patients with mTBI and symptoms lasting more than one week after injury. Patients with a delayed initial presentation should also be treated according to these guidelines. Detailed recommendations for the evaluation and treatment of specific symptoms can be found in the VA/DoD Clinical Practice Guideline for Management of Concussion/mTBI 1.
Symptom classification and goals of therapy Post-concussive symptoms generally fall into three categories: physical, cognitive, and behavioral or emotional. Typical physical symptoms include headache, nausea, vomiting, dizziness, fatigue, blurred vision, sleep disturbances, light or noise sensitivity, balance problems, and transient neurologic abnormalities. Cognitive symptoms may occur with attention, concentration, memory, processing speed, judgment, and executive functioning. Behavioral/emotional symptoms include depression, anxiety, agitation, irritability, and aggression.
Because there is an incomplete understanding of the etiology of symptoms after mTBI, the goal of intervention for post-concussive symptoms is to improve identified problems rather than affect a cure. It is believed that symptoms resulting from mTBI are inter-related and alleviation of one symptom often leads to improvement in others. Post-concussive symptoms are also common to many other psychiatric ailments including depression, anxiety disorders, post-traumatic stress disorder, and substance abuse disorders. Indeed, there is substantial evidence that affective disorders, post traumatic stress disorder (PTSD), and substance abuse disorders are often associated with mTBI 59, 63-64. These disorders are also associated with higher rates of persistent post-concussive symptoms 65-66. Consequently, aggressive treatment of any comorbid psychiatric illness may help to improve post-concussive symptoms.
Patient Evaluation Patient evaluation should include a thorough history, physical exam, and review of the medical record. A review of sleep habits is particularly important as poor sleep may contribute to symptoms including headache, fatigue, anxiety, irritability, depressive thoughts, poor concentration, memory difficulties, and poor decision making. TBI patients should also be screened for psychiatric conditions including PTSD, depression, and substance abuse disorders. Low yield diagnostic testing should be minimized. There is limited evidence to support the utility of comprehensive neuropsychological/cognitive testing within the first 30 days of mTBI and a focused clinical interview is sufficient to assess for cognitive difficulties 67. Laboratory studies including electrolytes, a complete blood count, and thyroid function testing may be useful, particularly when evaluating behavioral and cognitive symptoms. Imaging studies are of limited use.
Physical symptoms should prompt a search for treatable causes. Screening patients with headaches for preexisting headache conditions, hypertension, cervical spine abnormalities, sinus infections, and visual acuity deficits may provide useful avenues of treatment. Symptoms related to dizziness including poor coordination, unsteadiness, vertigo, or loss of balance, may be due to medication effects, orthostatic hypotension, or peripheral vertigo. Nausea may be caused by medications or gastro-esophageal reflux disease. Nasal polyps, sinus infection, and traumatic injury to the lingual or olfactory nerves may cause appetite changes. Physical injuries to the eye including corneal abrasions, lens dislocation, retinal detachment, and optic nerve injury should be considered in the evaluation of post-concussive visual complaints. Ear abnormalities including infection, tympanic membrane rupture, and auditory nerve injury may lead to phonophobia.
General Treatment Guidelines Treatment of physical symptoms includes treating the underlying causative or contributory conditions. Interventions targeting specific patient complaints such as sleep hygiene education, physical therapy, relaxation, and modification of the environment should be used. Moreover, medications may be used to relieve pain, enable sleep, and reduce stress 1. Cognitive deficits are often measurable within 30 days of mTBI but generally return to normal within the same period 68-70. Unfortunately, many patients continue to have subjective cognitive complaints 9, 71-75. Educational and cognitive-behavioral interventions consistently improve subjective cognitive complaints 61, 76-79. Behavioral symptoms may improve with psychotherapeutic and pharmacological interventions. Treatment should be based upon severity and nature of the symptom presentation. Patients with atypical symptoms or with significant suspected or confirmed comorbid illnesses may benefit from specialty referral or consultation. Finally, “red flag” conditions indicating an acute neurologic condition requiring urgent neurologic or neurosurgical intervention should prompt emergent transfer to a medical facility with an appropriate level of care.
The primary goal of pharmacological therapies for mTBI is symptomatic improvement. Currently, disease altering therapies are not available. Drug therapy for mTBI symptoms should follow several general principles. Medications that lower the seizure threshold such as buproprion and some anti-psychotic medicines should be avoided. Similarly, medications such as lithium, anti-cholinergic agents, benzodiazepines and others can cause altered mental status and should also be avoided. Starting doses should be as low as possible and titrated to effect under close monitoring. Conversely, maximal tolerated dosing should be trialed before switching to a new agent to avoid under treatment. Patients should be advised to avoid alcohol, caffeine, and herbal supplements. Limited doses of medications with significant toxicity in intentional overdoses such as tri-cyclic antidepressants should also be considered because suicide risk is high in brain injured patients. Finally, patients should be monitored closely for medication interactions and toxicity.
Medication therapy for patients in the first week after injury should be reserved for the treatment of headache only. Acetaminophen is the agent of choice. NSAIDs should not be used until 48 hours after injury unless there is normal neuroimaging data for the patient. Other immediate post-concussive symptoms should not be treated as they typically resolve spontaneously within the first week of injury.
Headache Headache is the most common symptom after mTBI affecting over 90% of patients. Post-traumatic headaches commonly fall into one of three categories: tension, migraine, or a combination of the two. The evaluation of post-traumatic headache should include assessment for neurologic findings suggestive of serious intra-cranial abnormalities. Focal neurologic deficits should prompt additional urgent investigation with appropriate neuroimaging. A medication review for patients with symptoms lasting more than two weeks is also important as rebound headaches are common in with daily acetaminophen or NSAID use. Similarly, withdrawal from caffeine or nicotine may also result in headache. Patients who state that their headache improves only with opiates should be referred to a pain or headache specialist. Headache symptoms often improve after treatment of comorbid conditions such as sleep disturbances, anxiety, and depression.
Pharmacologic treatment should be selected based on the type of headache suspected. Similar to the opiate dependent patients above, those with symptoms that do not improve within 3 months of initiating therapy should be referred to a headache or pain specialist. Episodic tension type headaches may be treated with aspirin, acetaminophen, or NSAIDs. These medications typically work best when combined with other treatment modalities such as a regular exercise program, relaxation techniques, or biofeedback. Combination medications that include caffeine or a sedative may be more effective but also have a greater likelihood of rebound headache.
Migraine treatment is divided into the prevention and management of acute episodes. Awareness and avoidance of precipitating events should be encouraged. Abortive medications for acute episodes include sumatriptan and zolmatriptan. These medications are most effective when used early in the course of the episode. Some patients with established migraines may require rescue medications to break their headache. Examples of effective rescue medications include ketorolac, butorphanol, opiods, prochlorparazine, and promethazine. Patients with migraine headaches that occur more than once a month should be placed on prophylaxis. First line agents for prophylaxis include metroprolol and toprimate. Prophylactic agents may take as long as three months to become maximally effective. Finally, mixed headache types may require separate agents for treatment of the tension and migraine components.
Disequilibrium and Vertigo Up to 30% of patients with mTBI complain of disturbed equilibrium or vertigo 87. Despite this, symptoms do not correlate with objective evidence after the first week after injury 88. A thorough medication review should be performed for all mTBI patients complaining of dizziness. Medications such as stimulants, benzodiazepines, tricyclics, monoamine oxidase inhibitors, tetracyclics, neuroleptics, selective serotonin reuptake inhibitors, beta blockers, and cholinesterase inhibitors may cause or exacerbate dizziness. Vestibular suppression may be useful in the acute phase but has not proven effective for persistent symptoms 89. Vestibular suppressants should only be used if the symptoms significantly limit the patient's functional activities as they may result in delayed improvement 90-91. Meclizine is the first agent recommended. Scopolamine and dimenhydrinate may be used if meclizine fails. Benzodiazepines should only be used after careful consideration of their sedating and habit-forming properties. Trials should be limited to two weeks duration.
Fatigue and Sleep Disturbances Another common symptom after mTBI is fatigue, which may be due to central nervous system dysfunction, sleep disturbances or depression. Proper assessment of this symptom requires a thorough history of pre- and post-injury levels of activity. There are also several validated instruments to objectively measure fatigue 92-93. Physical causes of fatigue may also be assessed with laboratory testing including metabolic panel, a complete blood count, and thyroid function testing. Review of the patient's medication history, alcohol, caffeine, and illicit drug use should also be performed as all of these may result in fatigue. Prior to initiating medications for fatigue conservative measures such as education of the patient, initiating an exercise program, as well as referring the patient for physical or cognitive behavioral therapy should be trialed. There is limited evidence for the efficacy of stimulant treatment for fatigue after mTBI. Commonly used agents include modafanil, methylphenidate, and amantadine. These medications should only be used if symptoms have lasted more than 4 weeks, the patient does not have substance abuse issues, and addressing other factors mentioned in this section have failed to improve symptoms. Trials of these medications should last at least 3 months.
Sleep disturbances are common after mTBI. The goal of therapy is to restore a regular, unbroken night-time sleep pattern and improve the perception of sleep quality. Any drug therapy for sleep disturbances should be accompanied by education regarding good sleep hygiene. Furthermore, concomitant primary sleep disorders such as obstructive sleep apnea, restless legs syndrome, and narcolepsy should be appropriately treated. In the acute phase, short term treatment with non-benzodiazepine sleep medications such as zolpidem may be helpful. Prazosin may be used in patients with nightmares or agitation during sleep.
Clinical Presentation
The patient returns to the concussion clinic 4 weeks after his initial visit and 6 weeks after his accident. He reports that he instituted the sleep hygiene recommendations given to him on the prior visit and that his headaches, sleepiness, irritability, and concentration difficulties subsequently resolved. He has required NSAIDS with decreasing frequency and has not had a headache in the last two weeks. The patient is sent home with instructions to contact the clinic for a future appointment if symptoms return.
Follow Up All patients require a follow up assessment within 4-6 weeks of initiation of therapy. Patients can be grouped into three categories at this second assessment: those with complete symptom resolution, those with partial resolution, and those with no improvement or worsened symptoms. Patients whose symptoms completely resolve should be given contact information to make a future appointment if symptoms return. Patients with a partial response may benefit from augmentation or adjustment of their current therapy. Those patients whose symptoms are refractory to initial treatment should be considered to have persistent post-concussive symptoms and treated according to the guidelines below (Management of Persistent Post-concussive Symptoms).
Management of Persistent Post-concussive Symptoms This section is relevant for patients who have had an initial evaluation and failed a trial of treatment for mTBI related symptoms. Patients with delayed presentation for mTBI symptoms should first be treated according to the preceding section (Initial Management of Post-concussive Symptoms) regardless of the interval since injury. The definitive reference is the VA/DoD Clinical Practice Guideline for Management of Concussion/mTBI 1.
Patients with persistent post-concussive symptoms often have concomitant behavioral health, psychosocial support, or compensation and litigation issues. Attention should be given to addressing these issues as this may help mitigate symptoms refractory to initial treatment. The evaluation of the patient with persistent post-concussive symptoms should include an assessment of available support systems, a mental health history including pre-morbid conditions, co-occurring symptoms such as chronic pain or personality disorders, substance abuse disorders, secondary gain issues, job status, and other financial or legal difficulties 1. Finally, all patients presenting with persistent post-concussive symptoms should be assessed for any potential danger to themselves or others.
Less than 5% of patients have persistent symptoms one year or more after injury 11. Patients typically have more physical complaints within 4 weeks of injury after which emotional complaints predominate 80. Once a thorough assessment has been obtained, the principle goal is to identify appropriate referrals for management of the persistent symptoms. Patients with behavioral symptoms and possible co-morbid psychiatric conditions may benefit from referral to mental health professionals. Persistent physical symptoms should be evaluated by appropriate specialists. Persistent cognitive symptoms are rare and are frequently accompanied by comorbid conditions such as mood disorders, poor physical health, poor psychosocial support, or chronic pain. In addition to addressing these comorbid conditions, these patients should be referred for neuropsychiatric evaluation to determine appropriate treatment options. Cognitive rehabilitation may be helpful for patients with persistent difficulties in memory, executive function, or attention 81-83. A social work referral is appropriate for patients with poor psychosocial support, legal difficulties, or financial problems. While there is consistent evidence of an association between mTBI related compensation or litigation and increased symptom reporting and poor outcome 59, 84-86, there is no evidence to support a therapeutic benefit of attributing persistent symptoms to these secondary gain issues 1. Consequently, clinicians should not allow symptom exaggeration by patients seeking compensation to alter their care plans.
Given the diverse group of health professionals involved in the treatment of persistent post-concussive symptoms, a multi-disciplinary, team approach with the referring provider as the coordinator of care is required. A designated case manager can be very helpful for coordinating care. Typical tasks benefiting from case management includes coordination of referrals, ensuring appropriate patient and family education, participation in short and long-term goal setting, ensuring that appropriate social service and mental health screening is performed, and coordination with the multi-disciplinary team. Ongoing follow up visits should occur regularly with goals of monitoring symptom severity, reviewing symptom impact on activities, and the effectiveness of treatments.
Return to Play after Sports Injury Guidelines for returning to play in an athlete differ from general instructions for return to normal activities after mTBI in that they are designed to prevent a repeat mTBI while the patient is recovering from the initial injury. In general, the risks of suffering a second and third TBI are 3 fold and 8 to 9 fold greater than the risk of a first TBI, respectively 94. Furthermore, case reports suggest that athletes are at increased risk for concussion in the period immediately after their initial injury 95. Therefore, consideration of the concussion risk in a sports-specific manner is important 96. Return to play guidelines are consensus rather than evidence based. The most commonly used guidelines include those by R.C. Cantu and the American Academy of Neurology (AAN) and of Dr. Cantu 97-98. Both sets of guidelines use severity of the concussion and presence of post-concussive symptoms as the criteria for return to play decision making. The Cantu guidelines allow a player to return once he/she is asymptomatic for one week if post traumatic amnesia lasted less than 24 hours and the initial loss of unconsciousness was less than five minutes. Players with more severe symptoms at the time of their concussion should not play for one month and then can return after an additional one week without symptoms. In players with a history of multiple concussions, consideration should be given to sitting out for the remainder of the season.
While the consensus based back to play guidelines referenced in this section are clinically accepted as the standard of care they are not infallible. A recent example of a second injury despite scrupulous application of these guidelines is seen with Brian Westbrook, a professional football player in the National Football League (NFL). Mr. Westbrook suffered the first concussion of his 8 year NFL career on October 26th 2009, suffering a brief loss of consciousness with associated retrograde amnesia after being tackled in a game. He was held from play for three weeks due to lingering headache and then suffered a second concussion in his first game back from injury. While there was widespread speculation that he might not return that season or even that his football career was over, he did play the final two games of the season without further injury after being out for a total of 5 weeks. While no guideline can prevent all adverse events, it is possible that improved guidelines could result in fewer repeat injuries. Prospectively validated, evidence based return to play guidelines are needed.
Go to:
Sequelae of mTBI
Post Concussive Symptoms and Cognitive Deficits
Common post-concussive symptoms include headache, dizziness, fatigue, sleep disturbances, memory problems, balance problems, sensitivity to sound or tinnitus, concentration difficulties, and irritability. These symptoms are notably non-specific and are associated with many other diseases. Nonetheless, several studies have reported higher rates of these symptoms in patients after mTBI than in patients with no injury or extra-cranial trauma without TBI 64, 99-100. The percentage of patients who suffer from persistent post-concussive symptoms diminishes with time after injury. Less than 25% of patients are likely to have problems lasting more than 12 months after injury 11. Although cognitive complaints are fairly common after mTBI, measurable cognitive deficits are generally only present after severe or moderate TBI 101-102. There is little evidence of objective cognitive deficits after mTBI 103.
Motor, Balance, and Cranial Nerve Abnormalities
In general, objective findings after mTBI are absent. Balance problems are emerging as a promising exception to this rule. In one study of 37 mTBI patients, testing of saccades, oculomotor smooth pursuit, upper-limb visuomotor function and neuropsychologic domains was performed and the results compared to uninjured control patients. At one year after injury, eye and upper limb movement, but not cognitive function remained impaired in the mTBI patients 104. In a more recent study, the same group found that eye movement impairment was significantly worse in mTBI patients suffering from post-concussive syndrome relative to mTBI patients with good recovery 105.
Psychiatric Diagnoses
Many studies have found an association between TBI of all severities and major depressive disorder 63, 106-107. This observed association was not likely to be explained by depression prior to injury however prior mood disorder may be an increased risk for TBI 108-109. While there are few studies of relationship between mania or bipolar disorder and TBI, the existing evidence suggests that there is not a strong relationship between them 110-112. There is limited evidence supporting an association between mTBI and PTSD in military populations. In a study of 2,525 soldiers returning after a one year deployment to Iraq, researchers identified a clear association between PTSD and mild TBI with LOC (OR, 2.98; 95% CI, 1.70–5.24) 64. A second cross-sectional study of 2,235 Afghanistan and Iraq war veterans also found an association between PTSD and mTBI 113. However, two studies of civilian populations found no relationship between mTBI and PTSD 114-115.
Second Impact Syndrome
Second impact syndrome (SIS) is a dreaded, rare complication of mTBI that occurs after a patient suffers a second mTBI while remaining symptomatic from the first. Typically, a patient will suffer a head injury during play resulting in post-concussive symptoms. After returning to play while still suffering symptoms they sustain a second, apparently minor head trauma, and rapidly suffer depressed mental status resulting in death or a persistent vegetative state. It is postulated that this disorder is caused by disordered cerebral autoregulation resulting from the initial TBI. The condition has mainly been reported in young men who play contact sports. The term SIS was first coined by Saunders and Harbaugh 116, however a similar syndrome was previously described by Schneider 117.
While SIS has become firmly fixed in the minds of clinicians as an important complication of mTBI, there is some question regarding whether it is a true clinical entity 118. A critical review of reported cases of SIS found that most did not meet a reasonable clinical definition of SIS. Cases often lacked a neuropathologic evidence of unexplained cerebral swelling 119. Even more problematic, most of the reported cases of precipitous neurologic collapse after a seemingly minor trauma occurred in the absence of any documented “first impact”. Of the seventeen cases reviewed, only five where classified as “probable SIS”. Given this analysis it is reasonable to conclude that the term SIS is inaccurate. Diffuse cerebral swelling can very rarely occur after mTBI, principally in children and adolescents, however a second mTBI is not required.
Seizures
While there is sufficient evidence to support a causal relationship between moderate or severe TBI and the development of unprovoked seizures, the evidence is limited for an association between seizures and mTBI 24. In non-military TBI populations, there is a 3.6 fold increase in the incidence of seizures relative to non-injured patients after TBI of all severities. After severe TBI, there was a 17 fold increase in seizure incidence which declined to 2.9 fold in moderate TBI patients. For mTBI patients with loss of consciousness or post traumatic amnesia, the incidence of seizures was 1.5 times that of controls (95% CI 1.0 – 2.2) 94, 120. These studies were limited in that pediatric patients, who have a higher baseline incidence of seizures than adults, were not analyzed separately from adults. Post-traumatic seizure risk is greatest in the first year after injury. After 4 years, TBI patients are no longer at increased risk relative to uninjured subjects 121.
Dementia and Neurodegeneration
Alzheimer's disease is the most common neurodegenerative disease and results in progressive dementia and eventual death. Familial or early onset Alzheimer's disease is caused by specific mutations and comprises approximately 10% cases. The remaining 90% of cases are referred to as sporadic. Although the mechanisms of disease progression in sporadic Alzheimer's disease are not known, it likely results from a combination of genetic and environmental factors. TBI is the strongest known environmental exposure associated with subsequent development of sporadic Alzheimer's disease. A retrospective cohort study of World War II veterans with documented closed head injury demonstrated an increase risk of Alzheimer's type dementia relative to non-head injured controls (Hazard ratio 2.00, 95% CI 1.03-3.90) 122. A meta-analysis of seven case control studies revealed similar results 123.
Dementia pugilistica, also known as chronic traumatic encephalopathy, is a neurodegenerative condition that affects athletes in sports that involve repeated head trauma such as boxing and mixed martial arts 124. Characteristic neuropathologic changes include cerebellar damage, cortical damage, and other scarring of the brain; substantia nigral degeneration; neurofibrillary tangles in the cerebral cortex and temporal horn areas; and abnormalities of the septum pellucidum. Autopsy of professional football players who died in their forties after developing dementia also showed neurodegenerative changes consistent with chronic traumatic encephalopathy 125-127. Neuropsychologic deficits associated with dementia pugilistica have been found in some studies 128-129 but not others 130-131.
Parkinsonism is a constellation of symptoms including tremor, rigidity, and bradikinesia, and postural instability and is caused by loss of central dopamine. Very little has been reported regarding association between TBI and parkinsonism, however, several case-control studies have shown an increased risk after mTBI with LOC or post-traumatic amnesia 132-133. The risk for the development of parkinsonism appears to increase with severity of TBI 132, 134.
Go to:
Summary
Mild traumatic brain injury is a widespread problem. Because of our limited understanding of the injury pathophysiology, the diagnosis of mTBI is based entirely on clinical symptoms, and often occurs in the absence of objective findings. The central feature of mTBI is a transiently altered state of consciousness after a traumatic injury to the head. The priority of emergency care is to identify potentially life threatening intra-cranial injuries through the judicious application of appropriate imaging studies and neurosurgical consultation. Although post-mTBI symptoms quickly and completely resolve in the vast majority of cases, a significant number of patients will complain of lasting problems. Post-concussive complaints tend to be inter-related and relief of one may have beneficial effects on others. Although the evidence is not definitive, longer term sequelae of mTBI may include seizure disorders and neurodegeneration. Recognizing the potentially life changing aspects of mTBI should be an important priority for the emergency physician because simple, early interventions such as education regarding the expected positive outcome from the injury and prompt treatment can prevent chronic symptoms from occurring.
Go to:
Acknowledgments
This work was supported by Grant No. 5R01HD051865-03 from the National Institutes of Health (Drs. Bazarian and Blyth) and a Jahnigen Career Development Scholars Award (Dr. Blyth).
Go to:
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Go to:
References
1. The Concussion/mTBI Working Group. VA/DoD Clinical Practice Guideline for Management of Concussion/mTBI. [October, 2009]. Available at: http://www.dvbic.org/images/pdfs/Providers/VADoD-CPG---Concussion-mTBI.aspx.
2. ACRM. American Congress of Rehabilitation Medicine Mild Traumatic Brain Injury Committee of the Head Injury Interdisciplinary Special Interest Group. Definition of mild traumatic brain injury. The Journal of Head Trauma Rehabilitation. 1993;8(3):86–87. [Google Scholar]
3. National Center for Injury Prevention and Control. Report to Congress on Mild Traumatic Brain Injury in the United States: Steps to Prevent a Serious Public Health Problem. Atlanta, GA: Centers for Disease Control and Prevention; 2003. [Google Scholar]
4. Lange RT, Iverson GL, Franzen MD. Neuropsychological functioning following complicated vs. uncomplicated mild traumatic brain injury. Brain Inj. 2009;23(2):83–91. [PubMed] [Google Scholar]
5. Saatman KE, Duhaime AC, Bullock R, et al. Classification of traumatic brain injury for targeted therapies. J Neurotrauma. 2008;25(7):719–38. [PMC free article] [PubMed] [Google Scholar]
6. Langlois JA, Rutland-Brown W, Thomas KE. Traumatic Brain Injury in the United States: Emergency Department Visits, Hospitalizations, and Deaths. Atlanta, GA: Centers for Disease Control and Prevention National Center for Injury Prevention and Control; 2006. [Google Scholar]
7. Jager TE, Weiss HB, Coben JH, et al. Traumatic brain injuries evaluated in U.S. emergency departments, 1992-1994. Acad Emerg Med. 2000;7(2):134–40. [PubMed] [Google Scholar]
8. Sosin DM, Sniezek JE, Thurman DJ. Incidence of mild and moderate brain injury in the United States, 1991. Brain Inj. 1996;10(1):47–54. [PubMed] [Google Scholar]
9. Alves WM, Macciocchi SN, Barth JT. Postconcussive symptoms after uncomplicated mild head injury. The Journal of Head Trauma Rehabilitation. 1993;8(3):48–59. [Google Scholar]
10. Middleboe T, Andersen HS, Birket-Smith M, et al. Minor head injury: impact on general health after 1 year. A prospective follow-up study. Acta Neurol Scand. 1992;85(1):5–9. [PubMed] [Google Scholar]
11. Iverson GL. Post-concussive disorder. New York, NY: Demos Medical Publishing LLC; 2007. [Google Scholar]
12. Alexander MP. Mild traumatic brain injury: pathophysiology, natural history, and clinical management. Neurology. 1995;45(7):1253–60. [PubMed] [Google Scholar]
13. Gondusky JS, Reiter MP. Protecting military convoys in Iraq: an examination of battle injuries sustained by a mechanized battalion during Operation Iraqi Freedom II. Mil Med. 2005;170(6):546–49. [PubMed] [Google Scholar]
14. Summers CR, Ivins B, Schwab KA. Traumatic brain injury in the United States: an epidemiologic overview. Mt Sinai J Med. 2009;76(2):105–10. [PubMed] [Google Scholar]
15. Gennarelli TA, Thibault LE, Adams JH, et al. Diffuse axonal injury and traumatic coma in the primate. Ann Neurol. 1982;12(6):564–74. [PubMed] [Google Scholar]
16. Holbourn AH. Mechanics of Head Injuries. Lancet. 1943;242(6267):438–41. [Google Scholar]
17. Ommaya AK, Gennarelli TA. Cerebral concussion and traumatic unconsciousness. Correlation of experimental and clinical observations of blunt head injuries. Brain. 1974;97(4):633–54. [PubMed] [Google Scholar]
18. LaPlaca MC, Simon CM, Prado GR, et al. CNS injury biomechanics and experimental models. Prog Brain Res. 2007;161:13–26. [PubMed] [Google Scholar]
19. McIntosh TK. Neurochemical sequelae of traumatic brain injury: therapeutic implications. Cerebrovasc Brain Metab Rev. 1994;6(2):109–62. [PubMed] [Google Scholar]
20. Werner C, Engelhard K. Pathophysiology of traumatic brain injury. Br J Anaesth. 2007;99(1):4–9. [PubMed] [Google Scholar]
21. Thompson HJ, Lifshitz J, Marklund N, et al. Lateral fluid percussion brain injury: a 15-year review and evaluation. J Neurotrauma. 2005;22(1):42–75. [PubMed] [Google Scholar]
22. Statler KD, Jenkins LW, Dixon CE, et al. The simple model versus the super model: translating experimental traumatic brain injury research to the bedside. J Neurotrauma. 2001;18(11):1195–206. [PubMed] [Google Scholar]
23. Marklund N, Bakshi A, Castelbuono DJ, et al. Evaluation of pharmacological treatment strategies in traumatic brain injury. Curr Pharm Des. 2006;12(13):1645–80. [PubMed] [Google Scholar]
24. IOM (Institute of Medicine) Gulf War and Health, Volume 7: Long-term Consequences of Traumatic Brain Injury. Washington, D.C.: The National Academies Press; 2009. [PubMed] [Google Scholar]
25. Adams RD, Victor M, Ropper AH. Principles of Neurology. 6th. New York: McGraw-Hill; 1997. [Google Scholar]
26. Hayes RL, Lyeth BG, Jenkins LW. Neurochemical mechanisms of mild and moderate head injury: implications for treatment. In: Levin HS, Eisenberg HM, Benton AL, editors. Mild Head Injury. Oxford: Oxford University Press; 1989. pp. 54–79. [Google Scholar]
27. Katayama Y, Watkins LR, Becker DP, et al. Evidence for involvement of cholinoceptive cells of the parabrachial region in environmentally induced nociceptive suppression in the cat. Brain Res. 1984;299(2):348–53. [PubMed] [Google Scholar]
28. Hayes RL, Pechura CM, Katayama Y, et al. Activation of pontine cholinergic sites implicated in unconsciousness following cerebral concussion in the cat. Science. 1984;223(4633):301–03. [PubMed] [Google Scholar]
29. Lyeth BG, Dixon CE, Hamm RJ, et al. Effects of anticholinergic treatment on transient behavioral suppression and physiological responses following concussive brain injury to the rat. Brain Res. 1988;448(1):88–97. [PubMed] [Google Scholar]
30. Ishige N, Pitts LH, Hashimoto T, et al. Effect of hypoxia on traumatic brain injury in rats: Part 1. Changes in neurological function, electroencephalograms, and histopathology. Neurosurgery. 1987;20(6):848–53. [PubMed] [Google Scholar]
31. Marmarou A, Foda MA, van den Brink W, et al. A new model of diffuse brain injury in rats. Part I: Pathophysiology and biomechanics. J Neurosurg. 1994;80(2):291–300. [PubMed] [Google Scholar]
32. Walker AE, Kollros JJ, Case TJ. The Physiological Basis of Concussion*. Journal of Neurosurgery. 1944;1(2):103–16. [Google Scholar]
33. Shaw NA. The neurophysiology of concussion. Prog Neurobiol. 2002;67(4):281–344. [PubMed] [Google Scholar]
34. Stein SC, Ross SE. The value of computed tomographic scans in patients with low-risk head injuries. Neurosurgery. 1990;26(4):638–40. [PubMed] [Google Scholar]
35. Nagurney JT, Borczuk P, Thomas SH. Elder patients with closed head trauma: a comparison with nonelder patients. Acad Emerg Med. 1998;5(7):678–84. [PubMed] [Google Scholar]
36. Miller EC, Holmes JF, Derlet RW. Utilizing clinical factors to reduce head CT scan ordering for minor head trauma patients. J Emerg Med. 1997;15(4):453–57. [PubMed] [Google Scholar]
37. Jeret JS, Mandell M, Anziska B, et al. Clinical predictors of abnormality disclosed by computed tomography after mild head trauma. Neurosurgery. 1993;32(1):9–15. discussion 15-16. [PubMed] [Google Scholar]
38. Jennett B, Teasdale G, Galbraith S, et al. Severe head injuries in three countries. J Neurol Neurosurg Psychiatry. 1977;40(3):291–98. [PMC free article] [PubMed] [Google Scholar]
39. Harad FT, Kerstein MD. Inadequacy of bedside clinical indicators in identifying significant intracranial injury in trauma patients. J Trauma. 1992;32(3):359–61. discussion 61-63. [PubMed] [Google Scholar]
40. Borczuk P. Predictors of intracranial injury in patients with mild head trauma. Ann Emerg Med. 1995;25(6):731–36. [PubMed] [Google Scholar]
41. Mittl RL, Grossman RI, Hiehle JF, et al. Prevalence of MR evidence of diffuse axonal injury in patients with mild head injury and normal head CT findings. AJNR Am J Neuroradiol. 1994;15(8):1583–89. [PMC free article] [PubMed] [Google Scholar]
42. Jagoda AS, Bazarian JJ, Bruns JJ, Jr, et al. Clinical policy: neuroimaging and decisionmaking in adult mild traumatic brain injury in the acute setting. Ann Emerg Med. 2008;52(6):714–48. [PubMed] [Google Scholar]
43. Stiell IG, Wells GA, Vandemheen K, et al. The Canadian CT Head Rule for patients with minor head injury. Lancet. 2001;357(9266):1391–96. [PubMed] [Google Scholar]
44. Haydel MJ, Preston CA, Mills TJ, et al. Indications for computed tomography in patients with minor head injury. N Engl J Med. 2000;343(2):100–05. [PubMed] [Google Scholar]
45. Stiell IG, Clement CM, Rowe BH, et al. Comparison of the Canadian CT Head Rule and the New Orleans Criteria in patients with minor head injury. JAMA. 2005;294(12):1511–18. [PubMed] [Google Scholar]
46. Kuppermann N, Holmes JF, Dayan PS, et al. Identification of children at very low risk of clinically-important brain injuries after head trauma: a prospective cohort study. Lancet. 2009;374(9696):1160–70. [PubMed] [Google Scholar]
47. Tasci A, Okay O, Gezici AR, et al. Prognostic value of interleukin-1 beta levels after acute brain injury. Neurol Res. 2003;25(8):871–74. [PubMed] [Google Scholar]
48. Ringger NC, O'Steen BE, Brabham JG, et al. A novel marker for traumatic brain injury: CSF alphaII-spectrin breakdown product levels. J Neurotrauma. 2004;21(10):1443–56. [PubMed] [Google Scholar]
49. Pelinka LE, Kroepfl A, Schmidhammer R, et al. Glial fibrillary acidic protein in serum after traumatic brain injury and multiple trauma. J Trauma. 2004;57(5):1006–12. [PubMed] [Google Scholar]
50. Olsson A, Csajbok L, Ost M, et al. Marked increase of beta-amyloid(1-42) and amyloid precursor protein in ventricular cerebrospinal fluid after severe traumatic brain injury. J Neurol. 2004;251(7):870–76. [PubMed] [Google Scholar]
51. Missler U, Wiesmann M, Wittmann G, et al. Measurement of glial fibrillary acidic protein in human blood: analytical method and preliminary clinical results. Clin Chem. 1999;45(1):138–41. [PubMed] [Google Scholar]
52. Berger RP, Heyes MP, Wisniewski SR, et al. Assessment of the macrophage marker quinolinic acid in cerebrospinal fluid after pediatric traumatic brain injury: insight into the timing and severity of injury in child abuse. J Neurotrauma. 2004;21(9):1123–30. [PubMed] [Google Scholar]
53. Papa L, Akinyi L, Liu MC, et al. Ubiquitin C-terminal hydrolase is a novel biomarker in humans for severe traumatic brain injury*. Crit Care Med. 2009 [PMC free article] [PubMed] [Google Scholar]
54. Biberthaler P, Linsenmeier U, Pfeifer KJ, et al. Serum S-100B concentration provides additional information fot the indication of computed tomography in patients after minor head injury: a prospective multicenter study. Shock. 2006;25(5):446–153. [PubMed] [Google Scholar]
55. Biberthaler P, Mussack T, Wiedemann E, et al. Elevated serum levels of S-100B reflect the extent of brain injury in alcohol intoxicated patients after mild head trauma. Shock. 2001;16(2):97–101. [PubMed] [Google Scholar]
56. Nagy KK, Joseph KT, Krosner SM, et al. The utility of head computed tomography after minimal head injury. J Trauma. 1999;46(2):268–70. [PubMed] [Google Scholar]
57. Livingston DH, Loder PA, Hunt CD. Minimal head injury: is admission necessary? Am Surg. 1991;57(1):14–17. [PubMed] [Google Scholar]
58. Dunham CM, Coates S, Cooper C. Compelling evidence for discretionary brain computed tomographic imaging in those patients with mild cognitive impairment after blunt trauma. J Trauma. 1996;41(4):679–86. [PubMed] [Google Scholar]
59. Carroll LJ, Cassidy JD, Peloso PM, et al. Prognosis for mild traumatic brain injury: results of the WHO Collaborating Centre Task Force on Mild Traumatic Brain Injury. J Rehabil Med. 2004;(43 Suppl):84–105. [PubMed] [Google Scholar]
60. Holm L, Cassidy JD, Carroll LJ, et al. Summary of the WHO Collaborating Centre for Neurotrauma Task Force on Mild Traumatic Brain Injury. J Rehabil Med. 2005;37(3):137–41. [PubMed] [Google Scholar]
61. Comper P, Bisschop SM, Carnide N, et al. A systematic review of treatments for mild traumatic brain injury. Brain Inj. 2005;19(11):863–80. [PubMed] [Google Scholar]
62. Borg J, Holm L, Peloso PM, et al. Non-surgical intervention and cost for mild traumatic brain injury: results of the WHO Collaborating Centre Task Force on Mild Traumatic Brain Injury. J Rehabil Med. 2004;(43 Suppl):76–83. [PubMed] [Google Scholar]
63. Fann JR, Burington B, Leonetti A, et al. Psychiatric illness following traumatic brain injury in an adult health maintenance organization population. Arch Gen Psychiatry. 2004;61(1):53–61. [PubMed] [Google Scholar]
64. Hoge CW, McGurk D, Thomas JL, et al. Mild traumatic brain injury in U.S. Soldiers returning from Iraq. N Engl J Med. 2008;358(5):453–63. [PubMed] [Google Scholar]
65. Mooney G, Speed J. The association between mild traumatic brain injury and psychiatric conditions. Brain Inj. 2001;15(10):865–77. [PubMed] [Google Scholar]
66. Rapoport MJ, Kiss A, Feinstein A. The impact of major depression on outcome following mild-to-moderate traumatic brain injury in older adults. J Affect Disord. 2006;92(2-3):273–76. [PubMed] [Google Scholar]
67. Cassidy JD, Carroll LJ, Peloso PM, et al. Incidence, risk factors and prevention of mild traumatic brain injury: results of the WHO Collaborating Centre Task Force on Mild Traumatic Brain Injury. J Rehabil Med. 2004;(43 Suppl):28–60. [PubMed] [Google Scholar]
68. Belanger HG, Curtiss G, Demery JA, et al. Factors moderating neuropsychological outcomes following mild traumatic brain injury: a meta-analysis. J Int Neuropsychol Soc. 2005;11(3):215–27. [PubMed] [Google Scholar]
69. Belanger HG, Vanderploeg RD. The neuropsychological impact of sports-related concussion: a meta-analysis. J Int Neuropsychol Soc. 2005;11(4):345–57. [PubMed] [Google Scholar]
70. Schretlen DJ, Shapiro AM. A quantitative review of the effects of traumatic brain injury on cognitive functioning. Int Rev Psychiatry. 2003;15(4):341–49. [PubMed] [Google Scholar]
71. Deb S, Lyons I, Koutzoukis C. Neurobehavioural symptoms one year after a head injury. Br J Psychiatry. 1999;174:360–65. [PubMed] [Google Scholar]
72. Dikmen S, McLean A, Temkin N. Neuropsychological and psychosocial consequences of minor head injury. J Neurol Neurosurg Psychiatry. 1986;49(11):1227–32. [PMC free article] [PubMed] [Google Scholar]
73. Hartlage LC, Durant-Wilson D, Patch PC. Persistent neurobehavioral problems following mild traumatic brain injury. Arch Clin Neuropsychol. 2001;16(6):561–70. [PubMed] [Google Scholar]
74. Luis CA, Vanderploeg RD, Curtiss G. Predictors of postconcussion symptom complex in community dwelling male veterans. J Int Neuropsychol Soc. 2003;9(7):1001–15. [PubMed] [Google Scholar]
75. Powell TJ, Collin C, Sutton K. A follow-up study of patients hospitalized after minor head injury. Disabil Rehabil. 1996;18(5):231–37. [PubMed] [Google Scholar]
76. Anson K, Ponsford J. Evaluation of a coping skills group following traumatic brain injury. Brain Inj. 2006;20(2):167–78. [PubMed] [Google Scholar]
77. Bedard M, Felteau M, Mazmanian D, et al. Pilot evaluation of a mindfulness-based intervention to improve quality of life among individuals who sustained traumatic brain injuries. Disabil Rehabil. 2003;25(13):722–31. [PubMed] [Google Scholar]
78. Hinkle JL, Alves WM, Rimell RW, et al. Restoring social competence in minor head-injury patients. J Neurosci Nurs. 1986;18(5):268–71. [PubMed] [Google Scholar]
79. Mittenberg W, Tremont G, Zielinski RE, et al. Cognitive-behavioral prevention of postconcussion syndrome. Arch Clin Neuropsychol. 1996;11(2):139–45. [PubMed] [Google Scholar]
80. Yang CC, Tu YK, Hua MS, et al. The association between the postconcussion symptoms and clinical outcomes for patients with mild traumatic brain injury. J Trauma. 2007;62(3):657–63. [PubMed] [Google Scholar]
81. Cicerone KD. Remediation of “working attention” in mild traumatic brain injury. Brain Inj. 2002;16(3):185–95. [PubMed] [Google Scholar]
82. Cicerone KD, Dahlberg C, Malec JF, et al. Evidence-based cognitive rehabilitation: updated review of the literature from 1998 through 2002. Arch Phys Med Rehabil. 2005;86(8):1681–92. [PubMed] [Google Scholar]
83. Tiersky LA, Anselmi V, Johnston MV, et al. A trial of neuropsychologic rehabilitation in mild-spectrum traumatic brain injury. Arch Phys Med Rehabil. 2005;86(8):1565–74. [PubMed] [Google Scholar]
84. Binder LM, Rohling ML. Money matters: a meta-analytic review of the effects of financial incentives on recovery after closed-head injury. Am J Psychiatry. 1996;153(1):7–10. [PubMed] [Google Scholar]
85. Kashluba S, Paniak C, Casey JE. Persistent symptoms associated with factors identified by the WHO Task Force on Mild Traumatic Brain Injury. Clin Neuropsychol. 2008;22(2):195–208. [PubMed] [Google Scholar]
86. Suhr JA, Gunstad J. Postconcussive symptom report: the relative influence of head injury and depression. J Clin Exp Neuropsychol. 2002;24(8):981–93. [PubMed] [Google Scholar]
87. Cicerone KD, Kalmar K. Persistent postconcussion syndrome: The structure of subjective complaints after mild traumatic brain injury. The Journal of Head Trauma Rehabilitation. 1995;10(3):1–17. [Google Scholar]
88. Gottshall KR, Gray NL, Drake AI, et al. To investigate the influence of acute vestibular impairment following mild traumatic brain injury on subsequent ability to remain on activity duty 12 months later. Mil Med. 2007;172(8):852–57. [PubMed] [Google Scholar]
89. Zee DS. Perspectives on the pharmacotherapy of vertigo. Arch Otolaryngol. 1985;111(9):609–12. [PubMed] [Google Scholar]
90. Pyykko I, Magnusson M, Schalen L, et al. Pharmacological treatment of vertigo. Acta Otolaryngol Suppl. 1988;455:77–81. [PubMed] [Google Scholar]
91. Hain TC, Yacovino D. Pharmacologic treatment of persons with dizziness. Neurol Clin. 2005;23(3):831–53. vii. [PubMed] [Google Scholar]
92. Krupp LB, LaRocca NG, Muir-Nash J, et al. The fatigue severity scale. Application to patients with multiple sclerosis and systemic lupus erythematosus. Arch Neurol. 1989;46(10):1121–3. [PubMed] [Google Scholar]
93. Chalder T, Berelowitz G, Pawlikowska T, et al. Development of a fatigue scale. J Psychosom Res. 1993;37(2):147–53. [PubMed] [Google Scholar]
94. Annegers JF, Grabow JD, Kurland LT, et al. The incidence, causes, and secular trends of head trauma in Olmsted County, Minnesota, 1935-1974. Neurology. 1980;30(9):912–19. [PubMed] [Google Scholar]
95. Kelly JP, Nichols JS, Filley CM, et al. Concussion in sports. Guidelines for the prevention of catastrophic outcome. JAMA. 1991;266(20):2867–69. [PubMed] [Google Scholar]
96. Kissick J, Johnston KM. Return to play after concussion: principles and practice. Clin J Sport Med. 2005;15(6):426–31. [PubMed] [Google Scholar]
97. Anonymous. Practice Parameter: The management of concussion in sports (summary statement) Neurology. 1997;48(3):581–85. [PubMed] [Google Scholar]
98. Cantu RC. Return to play guidelines after a head injury. Clin Sports Med. 1998;17(1):45–60. [PubMed] [Google Scholar]
99. Gerber DJ, Schraa JC. Mild traumatic brain injury: Searching for the syndrome. The Journal of Head Trauma Rehabilitation. 1995;10(4):28–40. [Google Scholar]
100. Masson F, Maurette P, Salmi LR, et al. Prevalence of impairments 5 years after a head injury, and their relationship with disabilities and outcome. Brain Inj. 1996;10(7):487–97. [PubMed] [Google Scholar]
101. Incoccia C, Formisano R, Muscato P, et al. Reaction and movement times in individuals with chronic traumatic brain injury with good motor recovery. Cortex. 2004;40(1):111–15. [PubMed] [Google Scholar]
102. Ruff RM, Evans R, Marshall LF. Impaired verbal and figural fluency after head injury. Arch Clin Neuropsychol. 1986;1(2):87–101. [PubMed] [Google Scholar]
103. Vanderploeg RD, Curtiss G, Belanger HG. Long-term neuropsychological outcomes following mild traumatic brain injury. J Int Neuropsychol Soc. 2005;11(3):228–36. [PubMed] [Google Scholar]
104. Heitger MH, Jones RD, Dalrymple-Alford JC, et al. Motor deficits and recovery during the first year following mild closed head injury. Brain Inj. 2006;20(8):807–24. [PubMed] [Google Scholar]
105. Heitger MH, Jones RD, Macleod AD, et al. Impaired eye movements in post-concussion syndrome indicate suboptimal brain function beyond the influence of depression, malingering or intellectual ability. Brain. 2009;132(Pt 10):2850–70. [PubMed] [Google Scholar]
106. Jorge RE, Robinson RG, Moser D, et al. Major depression following traumatic brain injury. Arch Gen Psychiatry. 2004;61(1):42–50. [PubMed] [Google Scholar]
107. Vanderploeg RD, Curtiss G, Luis CA, et al. Long-term morbidities following self-reported mild traumatic brain injury. J Clin Exp Neuropsychol. 2007;29(6):585–98. [PubMed] [Google Scholar]
108. Fann JR, Leonetti A, Jaffe K, et al. Psychiatric illness and subsequent traumatic brain injury: a case control study. J Neurol Neurosurg Psychiatry. 2002;72(5):615–20. [PMC free article] [PubMed] [Google Scholar]
109. Vassallo JL, Proctor-Weber Z, Lebowitz BK, et al. Psychiatric risk factors for traumatic brain injury. Brain Inj. 2007;21(6):567–73. [PubMed] [Google Scholar]
110. Koponen S, Taiminen T, Portin R, et al. Axis I and II psychiatric disorders after traumatic brain injury: a 30-year follow-up study. Am J Psychiatry. 2002;159(8):1315–21. [PubMed] [Google Scholar]
111. Sagduyu K. Association of mild traumatic brain injury with bipolar disorder. J Clin Psychiatry. 2002;63(7):594. [PubMed] [Google Scholar]
112. Silver JM, Kramer R, Greenwald S, et al. The association between head injuries and psychiatric disorders: findings from the New Haven NIMH Epidemiologic Catchment Area Study. Brain Inj. 2001;15(11):935–45. [PubMed] [Google Scholar]
113. Schneiderman AI, Braver ER, Kang HK. Understanding sequelae of injury mechanisms and mild traumatic brain injury incurred during the conflicts in Iraq and Afghanistan: persistent postconcussive symptoms and posttraumatic stress disorder. Am J Epidemiol. 2008;167(12):1446–52. [PubMed] [Google Scholar]
114. Bryant RA, Harvey AG. The influence of traumatic brain injury on acute stress disorder and post-traumatic stress disorder following motor vehicle accidents. Brain Inj. 1999;13(1):15–22. [PubMed] [Google Scholar]
115. Creamer M, O'Donnell ML, Pattison P. Amnesia, traumatic brain injury, and posttraumatic stress disorder: a methodological inquiry. Behav Res Ther. 2005;43(10):1383–89. [PubMed] [Google Scholar]
116. Saunders RL, Harbaugh RE. The second impact in catastrophic contact-sports head trauma. JAMA. 1984;252(4):538–39. [PubMed] [Google Scholar]
117. Schneider RC. Head and neck injuries in football: mehanisms, treatment and prevention. Baltimore: Williams and Wilkins; 1973. pp. 35–43. [Google Scholar]
118. McCrory PR, Berkovic SF. Second impact syndrome. Neurology. 1998;50(3):677–83. [PubMed] [Google Scholar]
119. McCrory P. Does second impact syndrome exist? Clin J Sport Med. 2001;11(3):144–49. [PubMed] [Google Scholar]
120. Annegers JF, Hauser WA, Coan SP, et al. A population-based study of seizures after traumatic brain injuries. N Engl J Med. 1998;338(1):20–24. [PubMed] [Google Scholar]
121. Singer RB. Incidence of seizures after traumatic brain injury--a 50-year population survey. J Insur Med. 2001;33(1):42–45. [PubMed] [Google Scholar]
122. Plassman BL, Havlik RJ, Steffens DC, et al. Documented head injury in early adulthood and risk of Alzheimer's disease and other dementias. Neurology. 2000;55(8):1158–66. [PubMed] [Google Scholar]
123. Van Duijn CM, Clayton DG, Chandra V, et al. Interaction between genetic and environmental risk factors for Alzheimer's disease: a reanalysis of case-control studies. EURODEM Risk Factors Research Group. Genet Epidemiol. 1994;11(6):539–51. [PubMed] [Google Scholar]
124. Corsellis JA, Bruton CJ, Freeman-Browne D. The aftermath of boxing. Psychol Med. 1973;3(3):270–303. [PubMed] [Google Scholar]
125. Omalu BI, DeKosky ST, Minster RL, et al. Chronic traumatic encephalopathy in a National Football League player. Neurosurgery. 2005;57(1):128–34. discussion 28-34. [PubMed] [Google Scholar]
126. Omalu BI, DeKosky ST, Hamilton RL, et al. Chronic traumatic encephalopathy in a National Football League player: part II. Neurosurgery. 2006;59(5):1086–92. discussion 92-93. [PubMed] [Google Scholar]
127. McKee AC, Cantu RC, Nowinski CJ, et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 2009;68(7):709–35. [PMC free article] [PubMed] [Google Scholar]
128. Drew RH, Templer DI, Schuyler BA, et al. Neuropsychological deficits in active licensed professional boxers. J Clin Psychol. 1986;42(3):520–25. [PubMed] [Google Scholar]
129. Roberts AJ. Brain Damage in Boxers. London: Pitman Medical Scientific Publications; 1969. [Google Scholar]
130. Porter MD. A 9-year controlled prospective neuropsychologic assessment of amateur boxing. Clin J Sport Med. 2003;13(6):339–52. [PubMed] [Google Scholar]
131. Porter MD, Fricker PA. Controlled prospective neuropsychological assessment of active experienced amateur boxers. Clin J Sport Med. 1996;6(2):90–96. [PubMed] [Google Scholar]
132. Bower JH, Maraganore DM, Peterson BJ, et al. Head trauma preceding PD: a case-control study. Neurology. 2003;60(10):1610–15. [PubMed] [Google Scholar]
133. Goldman SM, Tanner CM, Oakes D, et al. Head injury and Parkinson's disease risk in twins. Ann Neurol. 2006;60(1):65–72. [PubMed] [Google Scholar]
134. Taylor CA, Saint-Hilaire MH, Cupples LA, et al. Environmental, medical, and family history risk factors for Parkinson's disease: a New England-based case control study. Am J Med Genet. 1999;88(6):742–49. [PubMed] [Google Scholar]
OTHER FORMATS
PDF (166K)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 553816 | https://da.wikipedia.org/wiki/Bokserdemens | Bokserdemens | Bokserdemens eller Punch Drunk syndrom, tidligere kendt som dementia pugilistica er en neurologisk sygdom eller demens, der kan ramme boksere og andre sportsudøvere, der rammes af hjernerystelser. Symptomer og tegn på bokserdemens udvikles løbende over en længere latensperiode, der kan vare i årtier. Den gennemsnitlige periode for udviklingen af bokserdemens er mellem 12 og 16 år efter boksekarrierens begyndelse. Tilstanden menes at ramme 15-20% af professionelle boksere.
Tilstanden er forårsaget af gentagne slag mod hovedet, uanset om det enkelte slag er kraftigt nok til at give hjernerystelse eller ej. Grundet bekymringen om, at boksning kan forårsage bokserdemens, er der blandt visse læger ønske om at forbyde boksning. Der har været fremsat ønske om sådant forbud så tidligt som i 1950'erne.
Symptomer
Tilstanden, der ses hos boksere, der har modtaget gentagne slag mod hovedet, manifesterer sig som demens eller reduceret intellektuelle færdigheder, hukommelsesproblemer, parkinsons sygdom, rystelser eller manglende koordinationsevne. Tilstanden kan også give sig udtryk i talevanskeligheder og usikker gang. Patienter, der lider af dementia pugilistica kan optræde affektlabilt og kan udvise patologisk jalousi eller paranoia.
Personer, der lider af dementia pugilistica kan behandles med medicin, der benyttes til behandling af Alzheimers sygdom og Parkinsons sygdom.
Mekanisme
Det er ikke ganske klart, hvorledes syndromet opstår. Mistede neuroner, arvæv i hjernen, hydrocephalus (vand i hovedet), reduktion af hjernebjælkens funktion, diffus axonal skade og skade på lillehjernen m.v. involveret i syndromet. Tilstanden kan være etiologisk relateret til Alzheimers sygdom. Der er ved obduktion fundet neurofibrilliare tangles i hjernevævet hos patienter med dementia pugilistica men ikke med samme hyppighed, der normalt konstateres hos patienter, der lider af Alzheimers. Undersøgelser af skiver af hjernevæv fra patienter, der havde været udsat for milde traumatiske hjerneskader, viste ændringer i cellernes cytoskelet, hvilket kan være forårsaget af skade på de cerebrale blodårer.
Historie
Dementia pugilistica blev først beskrevet i 1928 af patologen Dr. Harrison Stanford Martland i en artikel i Journal of the American Medical Association, hvori han bekrev rystelser, langsomme bevægelser, forvirring og taleproblemer, der ofte forbindes med sygdommen. I 1973 beskrev en gruppe under ledelse af J.A. Corsellis de typiske neuropatologiske fund, der ofte observeret i forbindelse med dementia pugilistica efter obduktion af hjernerne hos 15 tidligere boksere.
Kendte tilfælde
Dementia pugilistica er forholdsvist udbredt blandt boksere, der har haft lange karrierer og som har modtaget mange slag mod hovedet. Sygdommen er muligvis ikke indrapporteret så ofte, idet symptomerne ofte ikke viser sig før end patienten er kommet op i årene, ligesom symptomerne ofte er vanskelige at adskille fra Alzheimers sygdom. På den anden side er dementia pugilistica til tider blevet fejlagtigt rapporteret.
En lang række boksere er blevet diagnosticeret med dementia pugilistica. Dette omfatter brødrene Jerry og Mike Quarry, Wilfred Benitez Jimmy Ellis, Floyd Patterson og Sugar Ray Robinson. Det er omdiskuteret om Muhammad Alis Parkinsons sygdom er forårsaget af boksekarrieren og har en sammenhæng med dementia pugilistica
Se også
Kronisk traumatisk encefalopati
Referencer
Eksterne links
Chronic Traumatic Encephalopathy In The National Football League: Pages 223-225 (Courtesy of http://www.sportslegacy.org)
Pugilistic Dementia; Something old, something new – from Boxingwriter.co.uk
Offensive Play: How different are dogfighting and football?, Malcolm Gladwell, The New Yorker, October 19, 2009 – article on the serious dangers of suffering long-term chronic traumatic encephalopathy brain damage in American football
Boston University Center for the Study of Traumatic Encephalopathy
Boksning
Hjerneskade i sport | danish | 0.545615 |
shut_down_conciousness/PMC6703193.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List Front Syst Neurosci PMC6703193
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of frontsysneuro
Front Syst Neurosci. 2019; 13: 36.
Published online 2019 Aug 14. doi: 10.3389/fnsys.2019.00036
PMCID: PMC6703193
PMID: 31474839
General Anesthesia: A Probe to Explore Consciousness
Vincent Bonhomme,1,2,3,* Cécile Staquet,1,3 Javier Montupil,1,2,3 Aline Defresne,1,2,3 Murielle Kirsch,1,3 Charlotte Martial,4 Audrey Vanhaudenhuyse,5 Camille Chatelle,4 Stephen Karl Larroque,4 Federico Raimondo,4 Athena Demertzi,6 Olivier Bodart,4 Steven Laureys,4 and Olivia Gosseries4
Author information Article notes Copyright and License information PMC Disclaimer
Go to:
Abstract
General anesthesia reversibly alters consciousness, without shutting down the brain globally. Depending on the anesthetic agent and dose, it may produce different consciousness states including a complete absence of subjective experience (unconsciousness), a conscious experience without perception of the environment (disconnected consciousness, like during dreaming), or episodes of oriented consciousness with awareness of the environment (connected consciousness). Each consciousness state may potentially be followed by explicit or implicit memories after the procedure. In this respect, anesthesia can be considered as a proxy to explore consciousness. During the recent years, progress in the exploration of brain function has allowed a better understanding of the neural correlates of consciousness, and of their alterations during anesthesia. Several changes in functional and effective between-region brain connectivity, consciousness network topology, and spatio-temporal dynamics of between-region interactions have been evidenced during anesthesia. Despite a set of effects that are common to many anesthetic agents, it is still uneasy to draw a comprehensive picture of the precise cascades during general anesthesia. Several questions remain unsolved, including the exact identification of the neural substrate of consciousness and its components, the detection of specific consciousness states in unresponsive patients and their associated memory processes, the processing of sensory information during anesthesia, the pharmacodynamic interactions between anesthetic agents, the direction-dependent hysteresis phenomenon during the transitions between consciousness states, the mechanisms of cognitive alterations that follow an anesthetic procedure, the identification of an eventual unitary mechanism of anesthesia-induced alteration of consciousness, the relationship between network effects and the biochemical or sleep-wake cycle targets of anesthetic agents, as well as the vast between-studies variations in dose and administration mode, leading to difficulties in between-studies comparisons. In this narrative review, we draw the picture of the current state of knowledge in anesthesia-induced unconsciousness, from insights gathered on propofol, halogenated vapors, ketamine, dexmedetomidine, benzodiazepines and xenon. We also describe how anesthesia can help understanding consciousness, we develop the above-mentioned unresolved questions, and propose tracks for future research.
Keywords: general anesthesia, consciousness, mechanisms, brain function, brain networks
Go to:
Introduction: General Anesthesia Is More Complex than Simply “Absence of Consciousness”
General anesthesia aims at providing patients with a state where they can tolerate unpleasant and/or noxious interventions, usually during a surgical procedure. Routinely, this involves a cocktail of medications ensuring an alteration of consciousness (pharmacological hypnosis) with absence of awareness of the surrounding environment, explicit recall of undercurrent events, a limitation of the stress response to nociception (anti-nociception), as well as immobility (muscle relaxation). General anesthesia does not shut down the brain globally and does not always produce a complete absence of consciousness. Similarly to disorders of consciousness in patients suffering from a severe brain insult (Aubinet et al., 2018), consciousness may be altered to various degrees, and its alteration may concern different consciousness elements as a function of the equilibrium between the inherent pharmacodynamic properties of anesthetic agents, their concentration in the body, and the intensity of the underlying stimulation by surgery.
Hence, one may not really speak about the depth of anesthetic hypnosis (Bayne et al., 2016), but rather about the presence or absence of one consciousness element or the other. As opposed to concepts that prevailed previously proposing that the brain was simply switched off by anesthesia, it is now clear that subjects may retain several higher-order brain functions until high concentrations of anesthetic agents are attained (Sleigh et al., 2018). If consciousness is defined as reflecting subjective experience/selfhood, or alternatively the sense of being a distinct entity capable of agency, sentience, narrative identity in time, and other higher-order components, anesthesia is capable of suppressing some of these components while keeping others functional (Sleigh et al., 2018). From a more basic and operational point of view, consciousness states that can be observed during general anesthesia are: (i) unconsciousness; (ii) disconnected consciousness; and (iii) connected consciousness (Table 1, Sanders et al., 2012). Unconsciousness can be defined as the inability to achieve any subjective experience and is expected to be the most common anesthetic state. During unconsciousness, reflex motor responses to stimulation may occur, such as arm withdrawal in response to noxious stimulation, but they are not purposeful and voluntary and do not imply a conscious connection with the environment. Disconnected consciousness is characterized by the presence of a mental content, but no conscious perception of the environment. In that case, the mental content can be similar to dreaming (Radek et al., 2018), or more distorted like ketamine-induced psychedelic subjective experiences. Connected consciousness in anesthesia refers to the subjective experience of self, and perception of information from the environment, which may happen in episodes of variable duration, and are not that rare (Sanders et al., 2017; Linassi et al., 2018; Radek et al., 2018). Such episodes can be observed immediately after laryngoscopy and tracheal intubation in approximately 5% of patients (Sanders et al., 2017). Whereas disconnected consciousness refers to internal awareness, connected consciousness refers to both external and internal awareness during anesthesia. External awareness is defined as the perception of environmental sensory stimuli (e.g., auditory, visual, olfactory, or somesthetic), and internal awareness refers to all environmental stimuli-independent thoughts (e.g., inner speech, autobiographical memories, or mind-wandering; Vanhaudenhuyse et al., 2011).
Table 1
Possible consciousness states during general anesthesia and their cognitive and mnemonic characteristics.
Mental content
Consciousness state External awareness (perception of environmental sensory stimuli) Internal awareness (thoughts independent from the environment) and sense of self (agency, sentience, identity, …) Sensory processing Purposeful response to command Explicit memory Implicit memory
Unconsciousness No No Possible (not accessible from the conscious field) No No Possible
Disconnected consciousness No Yes Yes (not related to external stimulation, e.g., seeing or smelling something during a dream) No Possible Possible
Connected consciousness Yes Yes Yes Yes Possible (probably rare) Possible
Open in a separate window
Inspired from Vanhaudenhuyse et al. (2011), Sanders et al. (2012) and Sleigh et al. (2018).
The classic method to know in which states patients were during anesthesia is to ask them after recovery, but this delayed assessment misses a lot of events that are not followed by explicit memories. In anesthesia, both disconnected and connected consciousness is rarely followed by explicit recall after the procedure, but the possibility of implicit memories exists. The isolated forearm technique allows assessing connected consciousness “online” during general anesthesia with muscle relaxation, when the patient is unable to manifest consciousness because being paralyzed. This technique consists of isolating the patient’s forearm from the systemic blood circulation through a cuffed upper arm tourniquet, which is inflated before the administration of the neuromuscular blocking agent. In such a setup, the isolated forearm remains non-paralyzed, and the anesthesiologist may ask verbal instructions, such as squeezing the hand and observe the patient’s response (Sanders et al., 2017). The same assessment can be done without the need of a tourniquet when the procedure does not require the use of neuromuscular blocking agents. A debate exists regarding the real incidence and risk factors of connected consciousness episodes during routine anesthesia practice. Although higher incidences have already been reported, the recently published incidence of 5% (and higher in younger patients) immediately after tracheal intubation was claimed to be conservative by the authors, because unclear responses were not counted and because assessment of connected consciousness did not occur during the remaining procedure (Sanders et al., 2017). During experimental studies, it remains important to gather subjective experiences immediately after recovery, to increase the likelihood of catching connectedness during unresponsive periods.
Classical processed electroencephalogram (EEG) indexes of the depth of anesthesia such as the Bispectral Index are not sensitive and specific enough to distinguish between the possible consciousness states of anesthesia (Gao et al., 2018). When asked to cite the consciousness states they would consider acceptable during anesthesia, subjects diversely appreciate one situation or the other. The eventuality of recall or feeling pain are major determinants of their appreciation (Rowley et al., 2017). By reversibly splitting and selectively altering some components of consciousness and selfhood, anesthetic agents are unique tools to explore the associated functional correlates.
Aside from the prerequisite of cortical arousal, which is controlled by sub-cortical sleep-wake cycle regulating systems, the complex phenomena of consciousness and subjective experience/selfhood mainly entail within and between neural networks interactions to generate and integrate information (Tononi, 2004). These activities translate into recordable electrical and metabolic complex signals that can be analyzed using sophisticated techniques. They have been and continue to be used in mostly single anesthetic drug studies in humans, and will ultimately shed light on the specific drug-related functional changes occurring during the different consciousness states of anesthesia, the hysteresis occurring during state transitions in one direction or the other (Kim et al., 2018), sensory processing, explicit or implicit memorization, and the frequently observed functional brain disturbances during the recovery period (Numan et al., 2017). We hereby narratively review the current state of the art in this domain, including studies on propofol, halogenated vapors, ketamine, dexmedetomidine, benzodiazepines and xenon, and underline questions that are not resolved to date.
Go to:
Several Ways of Exploring the Effects of Anesthetic Medications on the Brain
Currently, the exploration of brain function in humans mainly involves EEG recordings, combined with transcranial magnetic stimulation (TMS) or not, and functional brain imaging techniques such as positron emission tomography (PET) and functional magnetic resonance imaging (fMRI). The EEG signal corresponds to the underlying neuronal activities, and the blood oxygen level-dependent (BOLD) signal of fMRI (or the emitted gamma-ray energy in PET) corresponds to changes in regional cerebral blood flow in response to changes in activity. In addition to classical time and frequency domain analyses of the EEG (Marchant et al., 2014) and to classical activation studies of fMRI and PET (Bonhomme et al., 2001), sophisticated analyses can be applied to the recorded signals, with the primary aim to characterize the interactions between different brain regions. These interactions are thought to be the core mechanism of sensory processing and mental content generation (Lee and Mashour, 2018a). The number of possible analysis techniques is high, and each of them addresses a specific aspect of within-brain communication (Figure 1; Mashour and Hudetz, 2018; Staquet et al., 2018). It is therefore important to know exactly what they look at, in order to correctly interpret the results.
An external file that holds a picture, illustration, etc.
Object name is fnsys-13-00036-g0001.jpg
Figure 1
Summary representation of the available types of studies of the functioning brain that are applicable to the exploration of anesthetic brain effects. ICA, independent component analysis; DCM, dynamic causal modeling; ERP, event-related potentials; TMS-EEG, combined electroencephalography and transcranial magnetic stimulation; PCI, perturbational complexity index.
The most basic witness of brain communication is between-region synchrony in activity, or functional connectivity. When using fMRI, functional connectivity may be evidenced using hypothesis-based approaches, where brain regions show synchronized (as well as anti-correlated) activity with a predefined seed region (Boveroux et al., 2010; Bonhomme et al., 2016). Hypothesis-free data-driven analyses, for example, Independent Component Analysis (ICA), where signal synchronization is sought globally around the brain (Ribeiro de Paula et al., 2017), can also be used. Effective connectivity refers to the causal influence and directionality of the activity of a brain region on the activity of another region, and may be evidenced using Granger causality (GC) and its derivatives (Nicolaou and Georgiou, 2014), or using dynamic causal modeling (DCM; Gómez et al., 2013; Crone et al., 2017). Another approach, based on graph theory, consists in depicting the architecture or topology of brain networks (Lee and Mashour, 2018a), which are defined as assemblies of nodes (specific brain regions) that are linked by edges (the communication route between two regions). For each network, a set of topological properties can be described, including the average node degree (average number of edges for each node), the clustering coefficient (a measure of segregation, which estimates the isolation of node groups from other groups, a high clustering coefficient corresponds to a high degree of functional specialization), the characteristic path length (average of the minimum number of edges needed to link two nodes, a short path length denotes high integration capacities or efficiency), the participation coefficient (also a measure of integration), the small-worldness (best compromise between functional specialization and efficiency, a small-world network has a high clustering coefficient and short path length), the centrality (index of the central character of a node), the major hubs (most influential nodes, with a high number of edges and a high degree of centrality), the modularity (estimates the separation of a network into modules, which are groups of nodes separated by a hub), and criticality (or metastability, a state between order and disorder resulting from a scale-free organization, allowing the maximum number of possible configurations; Lee et al., 2017). Another mode of investigating brain communication relates to the evolution of the between-region interrelations over time and space, or spatio-temporal dynamics, and their complexity (Untergehrer et al., 2014; Wang et al., 2017; Cavanna et al., 2018; Huang et al., 2018; Muthukumaraswamy and Liley, 2018; Thiery et al., 2018), which is linked to criticality (Barttfeld et al., 2015; Lee et al., 2018). Finally, the analysis of evoked responses to stimulation offers other insights. The oldest studies in that respect were event-related potential EEG studies (Uhl et al., 1980; Plourde and Boylan, 1991), and the fMRI or PET activation studies (Bonhomme et al., 2008). The most recent ones look into how the brain handles sensory information (Lichtner et al., 2018a,b; Nourski et al., 2018) and between-region communication (Darracq et al., 2018a), as well as the directionality of information transfer (Sanders et al., 2018), and sensory cross-modal interactions (Bekinschtein et al., 2009). Some mixed approaches exist, melting one mode of analysis with another, such as those measuring the spatio-temporal complexity of TMS-evoked cortical responses (Casali et al., 2013; Bodart et al., 2017). Most recent emerging techniques use machine learning/decoding approaches through, for example, multivariate pattern analyses, but these have not much been applied to the anesthesia paradigm, yet (Liang et al., 2018). It is also possible to combine structural connectivity analyses (i.e., the exploration of anatomical connections through the white matter), and other types of connectivity analyses such as functional connectivity (Agarwal et al., 2016).
Go to:
Current State of the Art—Integrating the Available Data
The process of characterizing changes in brain function during general anesthesia is still ongoing. To date, information has been obtained through studies investigating one single anesthetic agent, mostly during the installing/induction and maintenance phase of sedation, and not all analysis modalities have been applied to each of them. The studies looking at other moments in the anesthesia process (such as the specific aspects of the direction of anesthetic state transitions, recovery as compared to induction of anesthesia, or post-operative delirium), at sensory processing, at memory processes, and at less frequent consciousness states (such as connected consciousness without memorization) are still scarce. A summary of the currently available information follows for each commonly used anesthetic agent (Table 2).
Table 2
Summary of the known effects of anesthetic agents on brain function.
Functional connectivity Effective connectivity Topological properties Evoked responses—sensory processing Spatio-temporal dynamics
Propofol Disruption of within- and between-network functional connectivity in large-scale brain networks (particularly fronto-parietal connectivity; Boveroux et al., 2010) Reduced amplitude and complexity of long-distance cortical communication (Gómez et al., 2013; Sarasso et al., 2015) Increased local efficiency (parietal regions; Kim et al., 2016; Lee et al., 2017) Generalized alteration in short-latency evoked electrocorticographic responses to auditory novelty within higher-order cortical areas, outside the auditory cortex (Nourski et al., 2018) Alteration of dynamics and directionality of effective connectivity (Lee et al., 2009; Untergehrer et al., 2014; Sanders et al., 2018)
Suppression of the complexity of regions sparsely connected with large-scale brain networks (Pappas et al., 2019) Disruption of effective connectivity in large-scale brain networks (Lee et al., 2009; Boly et al., 2012; Lee et al., 2018; Untergehrer et al., 2014; Guldenmund et al., 2016; Sanders et al., 2018) Fewer small-world properties (Barttfeld et al., 2015) Suppression of long-latency responses to novelty (Nourski et al., 2018) Reduced complexity and randomness of the electroencephalographic signal (Wang et al., 2017; Darracq et al., 2018b)
Disruption of thalamo-cortical connectivity within higher-order networks (Boveroux et al., 2010) Alteration of effective connectivity in lower-order sensory networks (Gómez et al., 2013) Limitation of connectivity configuration repertoire (Barttfeld et al., 2015; Cavanna et al., 2018) Reconfiguration of cortical functional connectivity networks involved in nociception, despite activation of spinal cord and cortex by noxious stimulation (Lichtner et al., 2018a,b) Synchronization of local activity (Huang et al., 2018)
Preservation of connectivity in lower-order sensory networks (Boveroux et al., 2010) Traffic of information constrained to inflexible patterns (Mashour, 2018; Uhrig et al., 2018) Prolongation of long-distance communication timescales (Gómez et al., 2013; Huang et al., 2018)
Remoteness from criticality, with preserved scale-free organization (Liu et al., 2014; Tagliazucchi et al., 2016; Alonso et al., 2019)
Disturbance of posterior parietal hub activity (Lee et al., 2013)
Halogenated vapors Breakdown of functional connectivity in higher-order resting-state consciousness networks (Palanca et al., 2015) Disruption of fronto-parietal anterior to posterior effective connectivity (Lee et al., 2013) Limitation of connectivity configuration repertoire (Cavanna et al., 2018; Uhrig et al., 2018) No information Augmentation of temporal persistence in neuronal oscillation amplitude (Thiery et al., 2018)
Disruption of thalamo-cortical connectivity within higher-order networks (Palanca et al., 2015) Remoteness from criticality (Lee et al., 2018) Disruption of intermediate strength spatio-temporal patterns of functional connectivity within and between consciousness networks (Kafashan et al., 2016)
Preservation of connectivity in lower-order sensory networks (Ranft et al., 2016) Preservation of higher strength spatio-temporal patterns within networks (Kafashan et al., 2016)
Ketamine Global increase in functional connectivity, with network reorganization (Driesen et al., 2013b) Disruption of fronto-parietal anterior to posterior effective connectivity (Lee et al., 2013; Vlisides et al., 2017) No information TMS-evoked communication complexity close to the waking state (Sarasso et al., 2015) No information
Disruption of functional connectivity in all higher-order consciousness networks but not in the executive control network (Bonhomme et al., 2016) Reduced alpha power in the precuneus and temporo-parietal junction (possibly related to disconnected consciousness; Vlisides et al., 2018; Darracq et al., 2018a)
Preservation of functional connectivity in sensory networks (Bonhomme et al., 2016)
Long-term effect on the interactions between the default mode network and networks involved in depression? and restoration of the abnormal connectivity of depressed patients (Li et al., 2018; Vutskits, 2018)
Transient effect on working memory network (Driesen et al., 2013a)
Dexmedetomidine Reduced within-network and thalamic connectivity in higher-order consciousness networks (Guldenmund et al., 2017) No information Reduced local and global large-scale network efficiency (Hashmi et al., 2017) No information No information
Preservation of lower-order sensory networks functional connectivity (Guldenmund et al., 2017) Reduced large-scale network connectivity strength (Hashmi et al., 2017)
Better preservation of functional connectivity between thalamus, medial anterior cingulate cortex, and mesopontine area as compared to sleep and propofol unresponsiveness (Guldenmund et al., 2017) No impairment in node degree (Hashmi et al., 2017)
Benzodiazepines Disruption of higher-order consciousness networks (Greicius et al., 2008; Liang et al., 2018) Disruption of effective connectivity in large-scale brain networks (Greicius et al., 2008; Ferrarelli et al., 2010; Liang et al., 2018) No information Reduced auditory cortex activation by sounds(Frolich et al., 2017) No information
Preservation of lower-order sensory networks (Frolich et al., 2017) Reduced duration and propagation of evoked TMS cortical response (Ferrarelli et al., 2010)
Xenon No information No information Remoteness from criticality (Colombo et al., 2019) TMS-evoked high amplitude slow waves with low complexity (Sarasso et al., 2015) Slowing down and smoothing of the temporal profile of the EEG signal (Colombo et al., 2019)
Open in a separate window
Propofol
Thanks to its safety and ease of use, the γ-amino-butyric acid (GABA) neurotransmission promoting agent propofol has been the most widely studied anesthetic agent with respect to brain mechanisms in primates and healthy humans. Years after the pioneering works of Fiset and Alkire, who were the first to demonstrate region-specific and dose-dependent effect of propofol on brain activity (Alkire et al., 1995; Fiset et al., 1999), it came out that this agent diminishes the randomness of the spontaneous and evoked EEG signal (Wang et al., 2017; Darracq et al., 2018b), alters long-distance cortical communication (Gómez et al., 2013) and reduces its complexity (Sarasso et al., 2015), disrupts within- and between-network functional connectivity in large-scale brain networks that sustain consciousness (Boveroux et al., 2010), and particularly functional fronto-parietal connectivity. Recent findings also suggest the suppression of the complexity of regions sparsely connected with large-scale brain networks as a mechanism of propofol-induced alteration in oriented reactivity to stimulation (Pappas et al., 2019). Effective connectivity, its dynamics, and its directionality are altered by propofol (Lee et al., 2009, 2013; Boly et al., 2012; Untergehrer et al., 2014; Guldenmund et al., 2016; Sanders et al., 2018). Contrarily, propofol relatively preserves lower-order sensory networks connectivity but impedes the effective connectivity within them (Gómez et al., 2013). When looking at network topological properties, propofol-induced unresponsiveness is associated with increased local efficiency, particularly in parietal regions (Lee et al., 2017), reduced global efficiency (Kim et al., 2016), disturbance of the posterior parietal hub activity (Lee et al., 2013), fewer small-world properties (Barttfeld et al., 2015), a limitation of the repertoire of possible connectivity configurations (Barttfeld et al., 2015; Cavanna et al., 2018) by constraining the traffic of information to inflexible patterns (Mashour, 2018; Uhrig et al., 2018), and remoteness from criticality (Tagliazucchi et al., 2016) with preserved scale-free organization of networks (preservation of node size and node degree distribution; Liu et al., 2014). Under propofol, the dynamics of within-brain interactions become more stable (Alonso et al., 2019). Propofol also synchronizes local activity and prolongs the timescales of long-distance communications (Huang et al., 2018). The consequences of propofol infusion on sensory processing involve a generalized alteration in short-latency evoked EEG responses to auditory novelty (which are thought to reflect pre-attentive processing) within higher-order cortical areas, but outside the auditory cortex. Long-latency responses to novelty (which may reflect conscious processing) are markedly suppressed by propofol in all regions (Nourski et al., 2018). Noxious information still reaches the cortex through the spinal cord, even under high propofol concentrations (Lichtner et al., 2018a), and in the presence of remifentanil (Lichtner et al., 2018b), but the cortical functional connectivity networks usually involved in nociception are reconfigured (Lichtner et al., 2018a).
Halogenated Vapors
Similarly to propofol, the inhaled halogenated vapors such as sevoflurane have GABAergic properties in addition to other biochemical targets such as potassium channels (Bonhomme et al., 2011). They break down functional connectivity in higher-order resting-state large-scale networks such as the default-mode network, and the salience network, as well as the thalamo-cortical connectivity within them (Palanca et al., 2015), with a preservation of connectivity within sensory networks (Ranft et al., 2016). The fronto-parietal anterior to posterior effective connectivity is also reduced by sevoflurane at anesthetic doses (Lee et al., 2013; Ranft et al., 2016). Halogenates put networks aside from criticality (Lee et al., 2018), and limit the repertoire of possible network configurations (Cavanna et al., 2018; Uhrig et al., 2018), possibly through an augmentation of temporal persistence in neuronal oscillation amplitude (Thiery et al., 2018). The intermediate strength spatio-temporal patterns of functional connectivity are disrupted within and between networks, while higher strength patterns are preserved within networks (Kafashan et al., 2016), and fewer transitions in between-networks connectivity patterns occur (Golkowski et al., 2019).
Ketamine
Among anesthetic agents, ketamine can be considered as the black sheep, because it induces distinct behavioral and functional changes as compared to other agents. The N-methyl-D-aspartate (NMDA) glutamate receptor antagonist ketamine produces a dissociative state with disconnected consciousness, through an isolation of the individual from the environment, and the emergence of intense dreaming with hallucinations and distorted self-perceptions. Ketamine increases functional connectivity globally (Driesen et al., 2013b). This occurs through a reconfiguration of between-region communication that becomes ineffective for some networks. Indeed, ketamine disrupts functional connectivity in all higher-order networks but the executive control network (Bonhomme et al., 2016). Also, connectivity is further preserved in sensory networks. A disruption of the fronto-parietal effective connectivity, and particularly the connectivity going from the anterior to the posterior part of the brain, has been observed during ketamine anesthesia (Lee et al., 2013; Vlisides et al., 2017). It induces a degree of TMS-evoked communication complexity that is close to the waking state and is very different from other agents such as propofol or xenon in that respect (Sarasso et al., 2015). Indeed, ketamine increases the randomness of the EEG signal (Wang et al., 2017). Reduced alpha power in the precuneus and temporal-parietal junction, both regions involved in multisensory integration and body representation, has been proposed as a mechanism for the ketamine-induced dissociative altered consciousness state and disconnected consciousness (Darracq et al., 2018a; Vlisides et al., 2018). Effects of ketamine on other functional systems have also been evidenced, such as its long-term effect on the interactions between the default-mode network and other networks involved in depression pathophysiology (Li et al., 2018), the restoration of abnormal connectivity observed in depressed patients (Scheidegger et al., 2012; Vutskits, 2018), and a transient effect on a network involved in working memory (Driesen et al., 2013a).
Dexmedetomidine
At first glance, the brain functional connectivity profile of the alteration of consciousness induced by the α2-adrenoceptor agonist dexmedetomidine appears similar to the one induced by physiological sleep and propofol, with a reduced within-network and thalamic connectivity in the higher-order consciousness networks, and a preservation of lower-order sensory networks (Guldenmund et al., 2017). However, dexmedetomidine better preserves functional connectivity between the thalamus, the anterior cingulate cortex, and the mesopontine area than sleep or propofol sedation (Guldenmund et al., 2017), all regions that are parts of the salience network. This may be in relation with the ability of dexmedetomidine to induce a state where the subject retains the capacity of rapidly recovering oriented responsiveness to external stimulation. From a topological point of view, dexmedetomidine reduces local and global large-scale network efficiency and connectivity strength, without impairing node degree (Hashmi et al., 2017).
Benzodiazepines
Similarly to propofol, benzodiazepines, which are potent GABA receptor ligands, are known to preserve functional connectivity in lower-order sensory networks despite reduced direct auditory cortex activation by sounds (Frolich et al., 2017), but not in the higher-order consciousness networks (Greicius et al., 2008; Liang et al., 2015). They also reduce the duration and propagation of evoked TMS cortical responses (Ferrarelli et al., 2010). A long term and chronic administration of diazepam increase functional connectivity in areas of emotional processing (Pflanz et al., 2015).
Xenon
Brain functional studies using the noble gas xenon are still scarce. This agent has anti-NMDA properties and interferes with potassium channels (Bonhomme et al., 2011). It reduces the activity of specific brain regions including the orbito- and mesiofrontal cortex, cingulate gyrus, thalamus, hippocampus and bilateral cerebellum (Rex et al., 2008). Similarly to propofol, it slows down and smooths the temporal profile of the EEG signal, and slides the brain state away from criticality (Colombo et al., 2019). TMS-evoked cortical responses under xenon anesthesia correspond to high amplitude slow waves with low complexity as compared to the wake state (Sarasso et al., 2015).
Go to:
Emerging Issues
Despite huge progress in unraveling the modalities of within-brain interactions, their implications in consciousness generation, and defining concepts that were not even imagined a decade ago, each discovery in the domain of consciousness physiology and brain effects of anesthetic agents leads to new questions. The ones of relevance for the understanding of anesthetic action, and whose resolution will allow making progress in the understanding of consciousness itself, are discussed hereafter (Figure 2).
An external file that holds a picture, illustration, etc.
Object name is fnsys-13-00036-g0002.jpg
Figure 2
Summary of the currently emerging issues regarding the brain effects of anesthetic agents and their relationship with the postulated neural mechanisms of consciousness.
Substrate of Consciousness and Its Alteration
Different models of within-brain communication were proposed as being witnesses of conscious processing by the brain following their fade-out or modification during altered states of consciousness. The strength of certainty when proposing a model is always reinforced when similar modifications are observed during alterations of consciousness of different origins (Lee et al., 2018). However, making the distinction between the effective neural correlates of consciousness (those responsible for the mental content experience), the elements that may support consciousness but are not mandatory (such as the sub-cortical arousal systems), and the neural systems whose alteration is simply a consequence or independent correlate of unconsciousness is not always easy (Boly et al., 2017; Mashour and Hudetz, 2018). Some authors distinguish content-specific neural correlates of consciousness (related to a specific mental content but not necessarily involved in another one), and full neural correlates of consciousness (mandatory for a mental content to be present; Boly et al., 2017). Based on lesion data, electrical or magnetic stimulation data, and functional brain imaging data, they argue that both content-specific and full neural correlates of consciousness are mainly located in the posterior part of the brain, encompassing the parietal, occipital, and lateral temporal lobes (Boly et al., 2017). This view is opposed to the previously prevailing one that was attributing a major role to the prefrontal cortex in this respect. It seems, however, that the prefrontal cortex is important at regulating the level of consciousness, through its privileged reciprocal interactions with subcortical arousal systems, in addition to attention, working memory, verbal and motor report processes (Pal et al., 2018). Information integration would occur through between-brain regions correlation in activity or functional connectivity (Koch et al., 2016), while information differentiation would be linked to the spatio-temporal dynamics of within-brain interactions and the diversity of brain functional configurations (Mashour and Hudetz, 2018; Demertzi et al., 2019). During anesthesia, however, one may not consider consciousness as a whole, insofar as specific aspects of it can be altered separately.
Further, matching up the evidence coming from different study paradigms and consciousness alterations of different origins, and finding parallelisms in the modification of functional witnesses of within-brain interaction models according to a specified component of consciousness will certainly lead to progress in characterizing its neural correlates with precision. By precisely and reversibly modulating consciousness, anesthesia will continue to play a major role in this progress. However, when analyzing anesthesia studies, researchers should pay attention to not mixing up the specific anesthetic drug effects on these neural correlates and their non-specific effects on neural functional assemblies that sustain other brain functions. Some study designs may help to overcome the difficulty when they involve a modulation of the consciousness state without changing the anesthetic agent concentration. As suggested by Scheinin et al. (2018), this can be achieved using external stimulation or an additional pharmacological agent that promotes cortical arousal through the strengthening of some subcortical arousal neurotransmission systems. External stimulation is more likely to provoke a change in the consciousness state when sedation is provided by an α2-adrenoceptor agonist like dexmedetomidine, but this can also occur under propofol sedation (Scheinin et al., 2018), or during routine general anesthesia when noxious stimulation elicits an episode of connected consciousness (Sanders et al., 2017). Similarly, physostigmine may change the level of arousal through an increase in cholinergic neurotransmission (Xie et al., 2011). When comparing the different brain states obtained at the same anesthetic agent concentration, it is important to precisely define their phenomenological characteristics, in order to pertinently correlate the functional changes with the behavioral ones.
Specific Consciousness States of Anesthesia and the Associated Memory Processes
Dreaming and/or connectedness can occur frequently in apparently and behaviorally unresponsive subjects during anesthesia (Radek et al., 2018). The ability to sensitively and specifically distinguish between the different possible consciousness states of anesthesia, namely unconsciousness, disconnected consciousness, and connected consciousness, in addition to the ability of predicting implicit or explicit recall would be ideal. One must admit that this is currently not the case with classical EEG analyses and commercially available depth of anesthesia monitors, as shown in a study by Gaskell et al. (2017). In this isolated forearm technique study, only the alpha-slow wave phase-amplitude coupling in the EEG was able to discriminate between patients with connected consciousness and those not responding to command. The first step before achieving such ability is to develop an understanding of the involved mechanisms, so as to define recordable correlates. Up to now, the memory processes of anesthesia have been poorly studied from a functional point of view. For example, a low dose of sevoflurane blocks emotional memory by reducing functional connectivity between the amygdala and the hippocampus (Alkire et al., 2008), and ketamine reversibly interferes with the circuits of working memory (Driesen et al., 2013a). Brief episodes of connected consciousness following intense noxious events during anesthesia are rarely—if not never—followed by explicit recall, but could potentially lead to implicit memories (Sanders et al., 2017). The proportion of true unexpected awareness episodes with explicit recall over the total number of detected and undetected connected consciousness episodes is currently not known. Nevertheless, very few elements are available to characterize the functional status of the brain at the very moment of connected consciousness, and its ability to initiate and consolidate memories at that time. Again, hints of mechanistic explanations to the anesthesia-induced consciousness states begin to be obtained, essentially by comparing altered consciousness states of different origins, but with the same phenomenological characteristics, and by correlating the observations with implicit and explicit memory data. Hence, when simultaneously measuring the effects of different anesthetic agents on brain activity, it was shown that a steeper decay-rate of the resting EEG power spectral density was characteristic of the absence of mental content in unresponsive subjects under propofol or xenon anesthesia. At the same time, a power spectral density decay similar to wakefulness was indicative of a mental content in unresponsive subjects under ketamine anesthesia (Colombo et al., 2019). TMS-evoked alpha power was found to be reduced during disconnected consciousness (dreaming) as compared to wakefulness, whenever it occurs during ketamine anesthesia or rapid-eye-movement sleep (Darracq et al., 2018a). Decreased TMS-evoked gamma power was observed in unconsciousness states induced by propofol anesthesia or non-rapid eye movement sleep (Darracq et al., 2018a) as compared to wakefulness and the other states. Slow-wave EEG activity saturation also seems to be pathognomonic of disconnectedness from the environment (Warnaby et al., 2017). All mechanistic information obtainable through this approach has not been gathered yet, but will certainly be in the future.
Sensory Processing and Cross-modal Interactions During Anesthesia
Within the scope of defining the amount of environmental information that reaches the brain, and the extent of its processing during the different anesthetic consciousness states, the study of sensory processing and cross-modal interactions is important. An intriguing recurrent finding of time-averaged connectivity studies is the preservation of functional connectivity within sensory networks, including thalamo-cortical connectivity, even during anesthesia-induced unresponsive states (Boveroux et al., 2010; Guldenmund et al., 2013; Bonhomme et al., 2016). However, sensory information handling by the brain appears to be altered during anesthesia through anomalies in between-network sensory modality crosstalk (Boveroux et al., 2010), changes in effective connectivity (Gómez et al., 2013), and increases in local inhibitory connectivity (Gómez et al., 2013). The use of more dynamic analysis paradigms and the study of evoked responses (Nourski et al., 2018) will certainly bring new information in this domain soon.
Mixing Anesthetic Agents
As stated above, general anesthesia is rarely provided to patients using a single agent. Commonly, anesthetic agents with hypnotic properties are at least combined with opioids. It has been known for a long time that pharmacodynamic interactions between anesthetic agents exist, manifesting notably on routinely used processed EEG indexes of anesthetic depth (Bouillon et al., 2004). Very few information is currently available regarding those interactions at the level of within-brain communication, as well as information generation and handling by the brain. This topic merits further investigations using the existing functional brain imaging techniques.
Direction-Dependent Mechanistic Differences in Anesthetic State Transitions
A phenomenon of hysteresis, sometimes referred to as neural inertia (Warnaby et al., 2017), occurs during anesthetic state transitions, meaning that the sequence of functional changes is distinct during forward and reverse transition from one consciousness state to the other. Although neural inertia typically refers to the pharmacokinetics of anesthetic agents, the observed differences are not uniquely related to them (Lee and Mashour, 2018b). This phenomenon may differ between anesthetic agents and is larger with the more potent ones (Kuizenga et al., 2018). Several EEG-based pharmacodynamic measures have been used to track the anesthesia recovery process (Purdon et al., 2013), and a phenomenon of hysteresis has been observed for the power spectrum, connectivity measures, structure and strength of networks (Kim et al., 2018), sensory-evoked EEG responses (Lewis et al., 2018), and slow-wave EEG activity saturation (Warnaby et al., 2017). The proposed involved mechanisms remain elusive. Specific study paradigms that precisely look at the dynamics of the transition loops and using different functional brain imaging modalities will help to elucidate them.
Post-operative Remnant Effects of Anesthetic Agents
The effects of anesthetic agents on the brain do not cease when the syringe pump or the vaporizer are turned off. A series of perioperative neurocognitive disorders (NCD’s) may be encountered in patients beneficiating from surgery. The taxonomy of perioperative NCD’s has recently been redefined by a group of experts (Evered et al., 2018). When considering the NCD’s that may have a link with anesthetic agents themselves, the following entities can be described. Immediately after anesthesia and the recovery of consciousness, cognition may remain altered for a limited period of time, a condition called emergence delirium. New NCD’s that appear within the usually expected recovery period of 30 post-operative days are qualified as delirium or delayed neurocognitive recovery. The qualifier post-operative is added to delirium when new and persisting before discharge from the hospital, but not when appearing after discharge. After 30 days up to 12 months, persisting NCD’s are named mild or major post-operative NCD’s. They correspond to the previously used term of post-operative cognitive disorders (POCD). These entities may present over a wide variety of symptoms, ranging from an agitated and restless disorientation in time and space, to slight memory or other cognitive deficits (Mason et al., 2010). Very few information exists regarding their neural correlates and the underlying alterations in brain function. As compared to normal immediate post-operative recovery where functional and directed back-to-front connectivity is slowed down (Lee et al., 2013; van Dellen et al., 2014), hypoactive delirium is marked out by a less integrated network topology (Numan et al., 2017). Additional further studies are needed to fully understand the diversity of these cognitive disorders.
Unitary Mechanism of Anesthesia-Induced Alteration of Consciousness
The question of a unitary mechanism of anesthesia-induced unconsciousness is a falsehood. Anesthetic agents produce different altered states of consciousness as a function of agent type and dose. This cannot occur through a common pathway. Several brain functional changes seem to be shared by the majority of anesthetic agents with hypnotic properties, including disrupted fronto-parietal functional and effective connectivity, reduced efficiency, increased modularity, disrupted posterior major hub, reconfigured network structure, disrupted balance between integration and segregation, disrupted metastability, and constrained repertoire of possible configurations (Lee and Mashour, 2018a). Among the observed functional effects, a good approach would be to establish a correspondence between them and phenomenological observations such as response to command when using the isolated forearm technique, post hoc report of dreaming with details, explicit recall of connected consciousness, standardized questionnaires for the detection of awareness, or specific paradigms for the detection of implicit memories. By submitting agent-related data to all available types of functional analyses, and making between-agent comparisons while taking into account the phenomenological characteristics of the consciousness states, substantial progress will be made in the understanding of the mechanism of each agent, and the functional correlate of consciousness components.
Link Between Network Effects, Sleep-Wake Regulation, and Biochemical Targets
Anesthetic agents with hypnotic properties pertain to highly variable chemical families and have differing biochemical targets. They have also been shown to interfere with several subcortical neurotransmission systems that are involved in the regulation of the sleep-wake cycle (Bonhomme et al., 2011), although anesthesia is clearly distinct from physiological sleep (Akeju and Brown, 2017). As a corollary, the crux of the matter remains to link those observations with the evidenced network effects. Some postulate a primary cortical effect, particularly for those agents mainly promoting the inhibitory GABA neurotransmission (Brown et al., 2011). Others favor a primary subcortical site of action and cortical dysfunction as mainly originating from thalamo-cortical interaction changes (Hutt et al., 2018). Dexmedetomidine, whose main target is the subcortical noradrenergic system, is an exception (Nelson et al., 2003). Nevertheless, each agent probably has its own sequential scheme of action, primarily cortical or subcortical, and we are still far from having drawn the complete picture for each of them. Magnetic resonance spectroscopy studies looking at in vivo neurotransmission will probably offer new insights soon (Abdallah et al., 2018).
Dose, Administration Mode, Anesthetic State, and Comparisons
The published studies in the domain of anesthetic drug effects on brain function considerably vary in their design, sometimes rendering the comparison between attained sedation level (when comparing different anesthetic agents), anesthetic doses, and studied anesthetic states hazardous. The phenomenological assessment of behavioral changes can sometimes be uneasy to perform (like during disconnected consciousness) or must be made through post hoc interviews. Behavioral assessment tools may also differ between studies. In addition, the route and mode of administration, which depend on the nature (intravenous or inhaled) and pharmacokinetic properties of the anesthetic agent, may introduce confounders. For example, a single intravenous bolus produces a continuously evolving alteration of consciousness, while a computer-controlled target infusion allows steady-state conditions of recording. Looking at steady-state anesthetic conditions does not provide the same information as looking at transitions from one state to the other. As mentioned above, the direction of the transition is also important. Precisely measuring or estimating the attained plasma or effect-site concentration of the anesthetic agent at the moment of recording is not always possible or not always performed. Indeed, due to inter-individual variability, the same anesthetic agent concentration may produce a different effect from one subject to the other, and some authors prefer defining the studied anesthetic state behaviorally rather than in terms of attained concentration. As a consequence, and given the high variability in study protocols regarding dose, mode of administration, and achieved anesthetic states, it is not easy to have a clear idea of the exact dose-response relationships, either in terms of brain function modifications and correspondences with phenomenological observations. A continuum in the observed effects from light to very deep sedation is not necessarily a reality, these relationships may be different from one anesthetic agent to the other, and not all possible levels of sedation have been studied for all anesthetic agents. All types of knowledge derived from the published studies are informative, but caution should be paid to not drawing erroneous conclusions when comparing them.
Go to:
Conclusion
The relationship between general anesthesia, brain function, and consciousness mechanisms is complex. As a matter of fact, anesthetic agents do not blunt out brain function globally but exert specific and dose-dependent effects on brain systems that sustain internal consciousness and perception of the environment. Each agent has its own mechanism of action, and dose-dependently induces distinct phenomenological altered consciousness states. Answering the questions that have recently emerged following recent discoveries on anesthetic brain effects will probably permit new insights into the specific diagnoses of anesthesia-induced altered states of consciousness, and into the understanding of the different aspects of consciousness itself. In that respect, general anesthesia can be considered as a flexible probe to explore consciousness.
Go to:
Author Contributions
VB and OG wrote the first draft of this article, the other authors equally participated in the revision and editing of the manuscript.
Go to:
Conflict of Interest Statement
During the past 5 years, VB has received research grant from Orion Pharma and honoraria for consultancy from Medtronic. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Go to:
Acknowledgments
We thank all the patients and volunteers who participated in our studies.
Go to:
Footnotes
Funding. The writing of this article was financially supported by the University Department of Anesthesia and Intensive Care Medicine, Centre Hospitalier Régional de la Citadelle (CHR Citadelle), Liege, Belgium; the Department of Anesthesia and Intensive Care Medicine, University Hospital Center of Liège (CHU Liège), Belgium; and the GIGA-Consciousness Thematic Unit, GIGA-Research, Liege University, Liege, Belgium. The studies performed by our group were funded by University Hospital of Liege; the Belgian National Funds for Scientific Research (FRS-FNRS); the European Union’s Horizon 2020 Framework Program for Research and Innovation under the Specific Grant Agreement No. 720270 (Human Brain Project SGA1) and No. 785907 (Human Brain Project SGA2); the Luminous project (EU-H2020-fetopen-ga686764); the DOCMA project (EU-H2020-MSCA-RISE-778234); the Marie Sklodowska-Curie Actions (H2020-MSCA-IF-2016-ADOC-752686); the BIAL Foundation; the French Speaking Community Concerted Research Action (ARC 12-17/01); the James McDonnell Foundation; the Mind Science Foundation; the IAP research network P7/06 of the Belgian Government (Belgian Science Policy); the European Commission; the Public Utility Foundation “Université Européenne du Travail”; the “Fondazione Europea di Ricerca Biomedica”; Orion Pharma; the Society for Anesthesia and Resuscitation of Belgium; the Benoit Foundation; the Belgian Cancer Foundation (2017-064). SKL is a FRS-FNRS research fellow, AD is an associate researcher at FRS-FNRS and SL is research director at FRS-FNRS. The funders had no role in any study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Go to:
References
Abdallah C. G., De Feyter H. M., Averill L. A., Jiang L., Averill C. L., Chowdhury G. M. I., et al.. (2018). The effects of ketamine on prefrontal glutamate neurotransmission in healthy and depressed subjects. Neuropsychopharmacology 43, 2154–2160. 10.1038/s41386-018-0136-3 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Agarwal S., Stamatakis E. A., Geva S., Warburton E. A. (2016). Dominant hemisphere functional networks compensate for structural connectivity loss to preserve phonological retrieval with aging. Brain Behav. 6:e00495. 10.1002/brb3.495 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Akeju O., Brown E. N. (2017). Neural oscillations demonstrate that general anesthesia and sedative states are neurophysiologically distinct from sleep. Curr. Opin. Neurobiol. 44, 178–185. 10.1016/j.conb.2017.04.011 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Alkire M. T., Gruver R., Miller J., McReynolds J. R., Hahn E. L., Cahill L. (2008). Neuroimaging analysis of an anesthetic gas that blocks human emotional memory. Proc. Natl. Acad. Sci. U S A 105, 1722–1727. 10.1073/pnas.0711651105 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Alkire M. T., Haier R. J., Barker S. J., Shah N. K., Wu J. C., Kao Y. J. (1995). Cerebral metabolism during propofol anesthesia in humans studied with positron emission tomography. Anesthesiology 82, 393–403. 10.1097/00000542-199502000-00010 [PubMed] [CrossRef] [Google Scholar]
Alonso L. M., Solovey G., Yanagawa T., Proekt A., Cecchi G. A., Magnasco M. O. (2019). Single-trial classification of awareness state during anesthesia by measuring critical dynamics of global brain activity. Sci. Rep. 9:4927. 10.1038/s41598-019-41345-4 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Aubinet C., Murphy L., Bahri M. A., Larroque S. K., Cassol H., Annen J., et al.. (2018). Brain, behavior, and cognitive interplay in disorders of consciousness: a multiple case study. Front. Neurol. 9:665. 10.3389/fneur.2018.00665 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Barttfeld P., Uhrig L., Sitt J. D., Sigman M., Jarraya B., Dehaene S. (2015). Signature of consciousness in the dynamics of resting-state brain activity. Proc. Natl. Acad. Sci. U S A 112, 887–892. 10.1073/pnas.1418031112 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Bayne T., Hohwy J., Owen A. M. (2016). Are there levels of consciousness? Trends Cogn. Sci. 20, 405–413. 10.1016/j.tics.2016.03.009 [PubMed] [CrossRef] [Google Scholar]
Bekinschtein T. A., Dehaene S., Rohaut B., Tadel F., Cohen L., Naccache L. (2009). Neural signature of the conscious processing of auditory regularities. Proc. Natl. Acad. Sci. U S A 106, 1672–1677. 10.1073/pnas.0809667106 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Bodart O., Gosseries O., Wannez S., Thibaut A., Annen J., Boly M., et al.. (2017). Measures of metabolism and complexity in the brain of patients with disorders of consciousness. Neuroimage Clin. 14, 354–362. 10.1016/j.nicl.2017.02.002 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Boly M., Massimini M., Tsuchiya N., Postle B. R., Koch C., Tononi G. (2017). Are the neural correlates of consciousness in the front or in the back of the cerebral cortex? Clinical and neuroimaging evidence. J. Neurosci. 37, 9603–9613. 10.1523/JNEUROSCI.3218-16.2017 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Boly M., Moran R., Murphy M., Boveroux P., Bruno M.-A., Noirhomme Q., et al.. (2012). Connectivity changes underlying spectral EEG changes during propofol-induced loss of consciousness. J. Neurosci. 32, 7082–7090. 10.1523/JNEUROSCI.3769-11.2012 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Bonhomme V., Boveroux P., Vanhaudenhuyse A., Hans P., Brichant J. F., Jaquet O., et al.. (2011). Linking sleep and general anesthesia mechanisms: this is no walkover. Acta Anaesthesiol. Belg. 62, 161–171. [PubMed] [Google Scholar]
Bonhomme V., Fiset P., Meuret P., Backman S., Plourde G., Paus T., et al.. (2001). Propofol anesthesia and cerebral blood flow changes elicited by vibrotactile stimulation: a positron emission tomography study. J. Neurophysiol. 85, 1299–1308. 10.1152/jn.2001.85.3.1299 [PubMed] [CrossRef] [Google Scholar]
Bonhomme V., Maquet P., Phillips C., Plenevaux A., Hans P., Luxen A., et al.. (2008). The effect of clonidine infusion on distribution of regional cerebral blood flow in volunteers. Anesth. Analg. 106, 899–909. 10.1213/ane.0b013e3181619685 [PubMed] [CrossRef] [Google Scholar]
Bonhomme V., Vanhaudenhuyse A., Demertzi A., Bruno M. A., Jaquet O., Bahri M. A., et al.. (2016). Resting-state network-specific breakdown of functional connectivity during ketamine alteration of consciousness in volunteers. Anesthesiology 125, 873–888. 10.1097/aln.0000000000001275 [PubMed] [CrossRef] [Google Scholar]
Bouillon T. W., Bruhn J., Radulescu L., Andresen C., Shafer T. J., Cohane C., et al.. (2004). Pharmacodynamic interaction between propofol and remifentanil regarding hypnosis, tolerance of laryngoscopy, bispectral index, and electroencephalographic approximate entropy. Anesthesiology 100, 1353–1372. 10.1097/00000542-200406000-00006 [PubMed] [CrossRef] [Google Scholar]
Boveroux P., Vanhaudenhuyse A., Bruno M.-A., Noirhomme Q., Lauwick S., Luxen A., et al.. (2010). Breakdown of within- and between-network resting state functional magnetic resonance imaging connectivity during propofol-induced loss of consciousness. Anesthesiology 113, 1038–1053. 10.1097/ALN.0b013e3181f697f5 [PubMed] [CrossRef] [Google Scholar]
Brown E. N., Purdon P. L., Van Dort C. J. (2011). General anesthesia and altered states of arousal: a systems neuroscience analysis. Annu. Rev. Neurosci. 34, 601–628. 10.1146/annurev-neuro-060909-153200 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Casali A. G., Gosseries O., Rosanova M., Boly M., Sarasso S., Casali K. R., et al.. (2013). A theoretically based index of consciousness independent of sensory processing and behavior. Sci. Transl. Med. 5:198ra105. 10.1126/scitranslmed.3006294 [PubMed] [CrossRef] [Google Scholar]
Cavanna F., Vilas M. G., Palmucci M., Tagliazucchi E. (2018). Dynamic functional connectivity and brain metastability during altered states of consciousness. Neuroimage 180, 383–395. 10.1016/j.neuroimage.2017.09.065 [PubMed] [CrossRef] [Google Scholar]
Colombo M. A., Napolitani M., Boly M., Gosseries O., Casarotto S., Rosanova M., et al.. (2019). The spectral exponent of the resting EEG indexes the presence of consciousness during unresponsiveness induced by propofol, xenon, and ketamine. Neuroimage 189, 631–644. 10.1016/j.neuroimage.2019.01.024 [PubMed] [CrossRef] [Google Scholar]
Crone J. S., Lutkenhoff E. S., Bio B. J., Laureys S., Monti M. M. (2017). Testing proposed neuronal models of effective connectivity within the cortico-basal gangliathalamo-cortical loop during loss of consciousness. Cereb. Cortex 27, 2727–2738. 10.1093/cercor/bhw112 [PubMed] [CrossRef] [Google Scholar]
Darracq M., Funk C. M., Polyakov D., Riedner B., Gosseries O., Nieminen J. O., et al.. (2018a). Evoked α power is reduced in disconnected consciousness during sleep and anesthesia. Sci. Rep. 8:16664. 10.1038/s41598-018-34957-9 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Darracq M., Sleigh J., Banks M. I., Sanders R. D. (2018b). Characterising the effect of propofol on complexity and stability in the EEG power spectrum. Br. J. Anaesth. 121, 1368–1369. 10.1016/j.bja.2018.09.006 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Demertzi A., Tagliazucchi E., Dehaene S., Deco G., Barttfeld P., Raimondo F., et al.. (2019). Human consciousness is supported by dnamic patterns of brain signal coordination. Sci. Adv. 5:eaat7603. 10.1126/sciadv.aat7603 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Driesen N. R., McCarthy G., Bhagwagar Z., Bloch M. H., Calhoun V. D., D’Souza D. C., et al.. (2013a). The impact of NMDA receptor blockade on human working memory-related prefrontal function and connectivity. Neuropsychopharmacology 38, 2613–2622. 10.1038/npp.2013.170 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Driesen N. R., McCarthy G., Bhagwagar Z., Bloch M., Calhoun V., D’Souza D. C., et al.. (2013b). Relationship of resting brain hyperconnectivity and schizophrenia-like symptoms produced by the NMDA receptor antagonist ketamine in humans. Mol. Psychiatry 18, 1199–1204. 10.1038/mp.2012.194 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Evered L., Silbert B., Knopman D. S., Scott D. A., DeKosky S. T., Rasmussen L. S., et al.. (2018). Recommendations for the nomenclature of cognitive change associated with anaesthesia and surgery-2018. Br. J. Anaesth. 121, 1005–1012. 10.1016/j.bja.2017.11.087 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Ferrarelli F., Massimini M., Sarasso S., Casali A., Riedner B. A., Angelini G., et al.. (2010). Breakdown in cortical effective connectivity during midazolam-induced loss of consciousness. Proc. Natl. Acad. Sci. U S A 107, 2681–2686. 10.1073/pnas.0913008107 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Fiset P., Paus T., Daloze T., Plourde G., Meuret P., Bonhomme V., et al.. (1999). Brain mechanisms of propofol-induced loss of consciousness in humans: a positron emission tomographic study. J. Neurosci. 19, 5506–5513. 10.1523/JNEUROSCI.19-13-05506.1999 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Frolich M. A., Banks C., Ness T. J. (2017). The effect of sedation on cortical activation: a randomized study comparing the effects of sedation with midazolam, propofol, and dexmedetomidine on auditory processing. Anesth. Analg. 124, 1603–1610. 10.1213/ane.0000000000002021 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Gómez F., Phillips C., Soddu A., Boly M., Boveroux P., Vanhaudenhuyse A., et al.. (2013). Changes in effective connectivity by propofol sedation. PLoS One 8:e71370. 10.1371/journal.pone.0071370 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Gao W.-W., He Y.-H., Liu L., Yuan Q., Wang Y.-F., Zhao B. (2018). BIS monitoring on intraoperative awareness: a meta-analysis. Curr. Med. Sci. 38, 349–353. 10.1007/s11596-018-1886-1 [PubMed] [CrossRef] [Google Scholar]
Gaskell A. L. L., Hight D. F. F., Winders J., Tran G., Defresne A., Bonhomme V., et al.. (2017). Frontal α-delta EEG does not preclude volitional response during anaesthesia: prospective cohort study of the isolated forearm technique. Br. J. Anaesth. 119, 664–673. 10.1093/bja/aex170 [PubMed] [CrossRef] [Google Scholar]
Golkowski D., Larroque S. K., Vanhaudenhuyse A., Plenevaux A., Boly M., Di Perri C., et al.. (2019). Changes in whole brain dynamics and connectivity patterns during sevoflurane- and propofol-induced unconsciousness identified by functional magnetic resonance imaging. Anesthesiology 130, 898–911. 10.1097/aln.0000000000002704 [PubMed] [CrossRef] [Google Scholar]
Greicius M. D., Kiviniemi V., Tervonen O., Vainionpää V., Alahuhta S., Reiss A. L., et al.. (2008). Persistent default-mode network connectivity during light sedation. Hum. Brain Mapp. 29, 839–847. 10.1002/hbm.20537 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Guldenmund P., Demertzi A., Boveroux P., Boly M., Vanhaudenhuyse A., Bruno M.-A., et al.. (2013). Thalamus, brainstem and salience network connectivity changes during propofol-induced sedation and unconsciousness. Brain Connect. 3, 273–285. 10.1089/brain.2012.0117 [PubMed] [CrossRef] [Google Scholar]
Guldenmund P., Gantner I. S., Baquero K., Das T., Demertzi A., Boveroux P., et al.. (2016). Propofol-induced frontal cortex disconnection: a study of resting-state networks, total brain connectivity and mean BOLD signal oscillation frequencies. Brain Connect. 6, 225–237. 10.1089/brain.2015.0369 [PubMed] [CrossRef] [Google Scholar]
Guldenmund P., Vanhaudenhuyse A., Sanders R. D., Sleigh J., Bruno M. A., Demertzi A., et al.. (2017). Brain functional connectivity differentiates dexmedetomidine from propofol and natural sleep. Br. J. Anaesth. 119, 674–684. 10.1093/bja/aex257 [PubMed] [CrossRef] [Google Scholar]
Hashmi J. A., Loggia M. L., Khan S., Gao L., Kim J., Napadow V., et al.. (2017). Dexmedetomidine disrupts the local and global efficiencies of large-scale brain networks. Anesthesiology 126, 419–430. 10.1097/ALN.0000000000001509 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Huang Z., Liu X., Mashour G. A., Hudetz A. G. (2018). Timescales of intrinsic BOLD signal dynamics and functional connectivity in pharmacologic and neuropathologic states of unconsciousness. J. Neurosci. 38, 2304–2317. 10.1523/JNEUROSCI.2545-17.2018 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Hutt A., Lefebvre J., Hight D., Sleigh J. (2018). Suppression of underlying neuronal fluctuations mediates EEG slowing during general anaesthesia. Neuroimage 179, 414–428. 10.1016/j.neuroimage.2018.06.043 [PubMed] [CrossRef] [Google Scholar]
Kafashan M., Ching S., Palanca B. J. A. (2016). Sevoflurane alters spatiotemporal functional connectivity motifs that link resting-state networks during wakefulness. Front. Neural Circuits 10:107. 10.3389/fncir.2016.00107 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Kim H., Moon J.-Y., Mashour G. A., Lee U. (2018). Mechanisms of hysteresis in human brain networks during transitions of consciousness and unconsciousness: theoretical principles and empirical evidence. PLoS Comput. Biol. 14:e1006424. 10.1371/journal.pcbi.1006424 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Kim M., Mashour G. A., Moraes S.-B., Vanini G., Tarnal V., Janke E., et al.. (2016). Functional and topological conditions for explosive synchronization develop in human brain networks with the onset of anesthetic-induced unconsciousness. Front. Comput. Neurosci. 10:1. 10.3389/fncom.2016.00001 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Koch C., Massimini M., Boly M., Tononi G. (2016). Neural correlates of consciousness: progress and problems. Nat. Rev. Neurosci. 17, 307–321. 10.1038/nrn.2016.22 [PubMed] [CrossRef] [Google Scholar]
Kuizenga M. H., Colin P. J., Reyntjens K. M. E. M., Touw D. J., Nalbat H., Knotnerus F. H., et al.. (2018). Test of neural inertia in humans during general anaesthesia. Br. J. Anaesth. 120, 525–536. 10.1016/j.bja.2017.11.072 [PubMed] [CrossRef] [Google Scholar]
Lee H., Golkowski D., Jordan D., Berger S., Ilg R., Lee J., et al.. (2018). Relationship of critical dynamics, functional connectivity, and states of consciousness in large-scale human brain networks. Neuroimage 188, 228–238. 10.1016/j.neuroimage.2018.12.011 [PubMed] [CrossRef] [Google Scholar]
Lee U., Kim S., Noh G. J., Choi B. M., Hwang E., Mashour G. A. (2009). The directionality and functional organization of frontoparietal connectivity during consciousness and anesthesia in humans. Conscious. Cogn. 18, 1069–1078. 10.1016/j.concog.2009.04.004 [PubMed] [CrossRef] [Google Scholar]
Lee U., Ku S., Noh G., Baek S., Choi B., Mashour G. A. (2013). Disruption of frontal-parietal communication by ketamine, propofol, and sevoflurane. Anesthesiology 118, 1264–1275. 10.1097/ALN.0b013e31829103f5 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lee H., Mashour G. A., Noh G.-J., Kim S., Lee U. (2013). Reconfiguration of network hub structure after propofol-induced unconsciousness. Anesthesiology 119, 1347–1359. 10.1097/aln.0b013e3182a8ec8c [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lee U., Mashour G. A. (2018a). Role of network science in the study of anesthetic state transitions. Anesthesiology 129, 1029–1044. 10.1097/ALN.0000000000002228 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lee U., Mashour G. A. (2018b). Stochastic nature of neural inertia. Br. J. Anaesth. 121, 7–8. 10.1016/j.bja.2018.04.018 [PubMed] [CrossRef] [Google Scholar]
Lee M., Sanders R. D., Yeom S. K., Won D. O., Seo K. S., Kim H. J., et al.. (2017). Network properties in transitions of consciousness during propofol-induced sedation. Sci. Rep. 7:16791. 10.1038/s41598-017-15082-5 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lewis L. D., Piantoni G., Peterfreund R. A., Eskandar E. N., Harrell P. G., Akeju O., et al.. (2018). A transient cortical state with sleep-like sensory responses precedes emergence from general anesthesia in humans. Elife 7:e33250. 10.7554/elife.33250 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Li M., Woelfer M., Colic L., Safron A., Chang C., Jochen H. (2018). Default mode network connectivity change corresponds to ketamine’s delayed glutamatergic effects. Eur. Arch. Psychiatry Clin. Neurosci. [Epub ahead of print]. 10.1007/s00406-018-0942-y [PubMed] [CrossRef] [Google Scholar]
Liang Z., Huang C., Li Y., Hight D. F., Voss L. J., Sleigh J. W., et al.. (2018). Emergence EEG pattern classification in sevoflurane anesthesia. Physiol. Meas. 39:045006. 10.1088/1361-6579/aab4d0 [PubMed] [CrossRef] [Google Scholar]
Liang P., Zhang H., Xu Y., Jia W., Zang Y., Li K. (2015). Disruption of cortical integration during midazolam-induced light sedation. Hum. Brain Mapp. 36, 4247–4261. 10.1002/hbm.22914 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lichtner G., Auksztulewicz R., Kirilina E., Velten H., Mavrodis D., Scheel M., et al.. (2018a). Effects of propofol anesthesia on the processing of noxious stimuli in the spinal cord and the brain. Neuroimage 172, 642–653. 10.1016/j.neuroimage.2018.02.003 [PubMed] [CrossRef] [Google Scholar]
Lichtner G., Auksztulewicz R., Velten H., Mavrodis D., Scheel M., Blankenburg F., et al.. (2018b). Nociceptive activation in spinal cord and brain persists during deep general anaesthesia. Br. J. Anaesth. 121, 291–302. 10.1016/j.bja.2018.03.031 [PubMed] [CrossRef] [Google Scholar]
Linassi F., Zanatta P., Tellaroli P., Ori C., Carron M. (2018). Isolated forearm technique: a meta-analysis of connected consciousness during different general anaesthesia regimens. Br. J. Anaesth. 121, 198–209. 10.1016/j.bja.2018.02.019 [PubMed] [CrossRef] [Google Scholar]
Liu X., Ward B. D., Binder J. R., Li S. J., Hudetz A. G. (2014). Scale-free functional connectivity of the brain is maintained in anesthetized healthy participants but not in patients with unresponsive wakefulness syndrome. PLoS One 9:e92182. 10.1371/journal.pone.0092182 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Marchant N., Sanders R., Sleigh J., Vanhaudenhuyse A., Bruno M. A., Brichant J. F., et al.. (2014). How electroencephalography serves the anesthesiologist. Clin. EEG Neurosci. 45, 22–32. 10.1177/1550059413509801 [PubMed] [CrossRef] [Google Scholar]
Mashour G. A. (2018). Highways of the brain, traffic of the mind. Anesthesiology 129, 869–871. 10.1097/aln.0000000000002385 [PubMed] [CrossRef] [Google Scholar]
Mashour G. A., Hudetz A. G. (2018). Neural correlates of unconsciousness in large-scale brain networks. Trends Neurosci. 41, 150–160. 10.1016/j.tins.2018.01.003 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Mason S. E., Noel-Storr A., Ritchie C. W. (2010). The impact of general and regional anesthesia on the incidence of post-operative cognitive dysfunction and post-operative delirium: a systematic review with meta-analysis. J. Alzheimers Dis. 22, 67–79. 10.3233/jad-2010-101086 [PubMed] [CrossRef] [Google Scholar]
Muthukumaraswamy S. D., Liley D. T. (2018). 1/F electrophysiological spectra in resting and drug-induced states can be explained by the dynamics of multiple oscillatory relaxation processes. Neuroimage 179, 582–595. 10.1016/j.neuroimage.2018.06.068 [PubMed] [CrossRef] [Google Scholar]
Nelson L. E., Lu J., Guo T., Saper C. B., Franks N. P., Maze M. (2003). The α2-adrenoceptor agonist dexmedetomidine converges on an endogenous sleep-promoting pathway to exert its sedative effects. Anesthesiology 98, 428–436. 10.1097/00000542-200302000-00024 [PubMed] [CrossRef] [Google Scholar]
Nicolaou N., Georgiou J. (2014). Neural network-based classification of anesthesia/awareness using granger causality features. Clin. EEG Neurosci. 45, 77–88. 10.1177/1550059413486271 [PubMed] [CrossRef] [Google Scholar]
Nourski K. V., Steinschneider M., Rhone A. E., Kawasaki H., Howard M. A., III., Banks M. I. (2018). Auditory predictive coding across awareness states under anesthesia: an intracranial electrophysiology study. J. Neurosci. 38, 8441–8452. 10.1523/JNEUROSCI.0967-18.2018 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Numan T., Slooter A. J. C., van der Kooi A. W., Hoekman A. M. L., Suyker W. J. L., Stam C. J., et al.. (2017). Functional connectivity and network analysis during hypoactive delirium and recovery from anesthesia. Clin. Neurophysiol. 128, 914–924. 10.1016/j.clinph.2017.02.022 [PubMed] [CrossRef] [Google Scholar]
Pal D., Dean J. G., Liu T., Li D., Watson C. J., Hudetz A. G., et al.. (2018). Differential role of prefrontal and parietal cortices in controlling level of consciousness. Curr. Biol. 28, 2145.e5–2152.e5. 10.1016/j.cub.2018.05.025 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Palanca B. J. A., Mitra A., Larson-Prior L., Snyder A. Z., Avidan M. S., Raichle M. E. (2015). Resting-state functional magnetic resonance imaging correlates of sevoflurane-induced unconsciousness. Anesthesiology 123, 346–356. 10.1097/aln.0000000000000731 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Pappas I., Adapa R. M., Menon D. K., Stamatakis E. A. (2019). Brain network disintegration during sedation is mediated by the complexity of sparsely connected regions. Neuroimage 186, 221–233. 10.1016/j.neuroimage.2018.10.078 [PubMed] [CrossRef] [Google Scholar]
Pflanz C. P., Pringle A., Filippini N., Warren M., Gottwald J., Cowen P. J., et al.. (2015). Effects of seven-day diazepam administration on resting-state functional connectivity in healthy volunteers: a randomized, double-blind study. Psychopharmacology 232, 2139–2147. 10.1007/s00213-014-3844-3 [PubMed] [CrossRef] [Google Scholar]
Plourde G., Boylan J. F. (1991). The auditory steady state response during sufentanil anaesthesia. Br. J. Anaesth. 66, 683–691. 10.1093/bja/66.6.683 [PubMed] [CrossRef] [Google Scholar]
Purdon P. L., Pierce E. T., Mukamel E. A., Prerau M. J., Walsh J. L., Wong K. F. K., et al.. (2013). Electroencephalogram signatures of loss and recovery of consciousness from propofol. Proc. Natl. Acad. Sci. U S A 110, E1142–E1151. 10.1073/pnas.1221180110 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Radek L., Kallionpää R. E., Karvonen M., Scheinin A., Maksimow A., Långsjö J., et al.. (2018). Dreaming and awareness during dexmedetomidine- and propofol-induced unresponsiveness. Br. J. Anaesth. 121, 260–269. 10.1016/j.bja.2018.03.014 [PubMed] [CrossRef] [Google Scholar]
Ranft A., Golkowski D., Kiel T., Riedl V., Kohl P., Rohrer G., et al.. (2016). Neural correlates of sevoflurane-induced unconsciousness identified by simultaneous functional magnetic resonance imaging and electroencephalography. Anesthesiology 125, 861–872. 10.1097/ALN.0000000000001322 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rex S., Meyer P. T., Baumert J.-H., Rossaint R., Fries M., Bull U., et al.. (2008). Positron emission tomography study of regional cerebral blood flow and flow-metabolism coupling during general anaesthesia with xenon in humans. Br. J. Anaesth. 100, 667–675. 10.1093/bja/aen036 [PubMed] [CrossRef] [Google Scholar]
Ribeiro de Paula D., Ziegler E., Abeyasinghe P. M., Das T. K., Cavaliere C., Aiello M., et al.. (2017). A method for independent component graph analysis of resting-state fMRI. Brain Behav. 7:e00626. 10.1002/brb3.626 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rowley P., Boncyk C., Gaskell A., Absalom A., Bonhomme V., Coburn M., et al.. (2017). What do people expect of general anaesthesia? Br. J. Anaesth. 118, 486–488. 10.1093/bja/aex040 [PubMed] [CrossRef] [Google Scholar]
Sanders R. D., Banks M. I., Darracq M., Moran R., Sleigh J., Gosseries O., et al.. (2018). Propofol-induced unresponsiveness is associated with impaired feedforward connectivity in cortical hierarchy. Br. J. Anaesth. 121, 1084–1096. 10.1016/j.bja.2018.07.006 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Sanders R. D., Gaskell A., Raz A., Winders J., Stevanovic A., Rossaint R., et al.. (2017). Incidence of connected consciousness after tracheal intubation: a prospective, international, multicenter cohort study of the isolated forearm technique. Anesthesiology 126, 214–222. 10.1097/ALN.0000000000001479 [PubMed] [CrossRef] [Google Scholar]
Sanders R. D., Tononi G., Laureys S., Sleigh J. W. (2012). Unresponsiveness ≠ unconsciousness. Anesthesiology 116, 946–959. 10.1097/ALN.0b013e318249d0a7 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Sarasso S., Boly M., Napolitani M., Gosseries O., Charland-Verville V., Casarotto S., et al.. (2015). Consciousness and complexity during unresponsiveness induced by propofol, xenon and ketamine. Curr. Biol. 25, 3099–3105. 10.1016/j.cub.2015.10.014 [PubMed] [CrossRef] [Google Scholar]
Scheidegger M., Walter M., Lehmann M., Metzger C., Grimm S., Boeker H., et al.. (2012). Ketamine decreases resting state functional network connectivity in healthy subjects: implications for antidepressant drug action. PLoS One 7:e44799. 10.1371/journal.pone.0044799 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Scheinin H., Alkire E. C., Scheinin A., Alkire M. T., Kantonen O., Langsjo J. (2018). Using positron emission tomography in revealing the mystery of general anesthesia: study design challenges and opportunities. Meth. Enzymol. 603, 279–303. 10.1016/bs.mie.2018.01.025 [PubMed] [CrossRef] [Google Scholar]
Sleigh J., Warnaby C., Tracey I. (2018). General anaesthesia as fragmentation of selfhood: insights from electroencephalography and neuroimaging. Br. J. Anaesth. 121, 233–240. 10.1016/j.bja.2017.12.038 [PubMed] [CrossRef] [Google Scholar]
Staquet C., Vanhaudenhuyse A., Bonhomme V. (2018). Aware beside an unconscious patient, not the inverse! On the necessity of knowing how anesthesia modulates consciousness. Acta Anaesthesiol. Belg. 69, 137–145. [Google Scholar]
Tagliazucchi E., Chialvo D. R., Siniatchkin M., Amico E., Brichant J.-F., Bonhomme V., et al.. (2016). Large-scale signatures of unconsciousness are consistent with a departure from critical dynamics. J. R. Soc. Interface 13:20151027. 10.1098/rsif.2015.1027 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Thiery T., Lajnef T., Combrisson E., Dehgan A., Rainville P., Mashour G. A., et al.. (2018). Long-range temporal correlations in the brain distinguish conscious wakefulness from induced unconsciousness. Neuroimage 179, 30–39. 10.1016/j.neuroimage.2018.05.069 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Tononi G. (2004). An information integration theory of consciousness. BMC Neurosci. 5:42. 10.1186/1471-2202-5-42 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Uhl R. R., Squires K. C., Bruce D. L., Starr A. (1980). Variations in visual evoked potentials under anesthesia. Prog. Brain Res. 54, 463–466. 10.1016/s0079-6123(08)61662-3 [PubMed] [CrossRef] [Google Scholar]
Uhrig L., Sitt J. D., Jacob A., Tasserie J., Barttfeld P., Dupont M., et al.. (2018). Resting-state dynamics as a cortical signature of anesthesia in monkeys. Anesthesiology 129, 942–958. 10.1097/ALN.0000000000002336 [PubMed] [CrossRef] [Google Scholar]
Untergehrer G., Jordan D., Kochs E. F., Ilg R., Schneider G. (2014). Fronto-parietal connectivity is a non-static phenomenon with characteristic changes during unconsciousness. PLoS One 9:e87498. 10.1371/journal.pone.0087498 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
van Dellen E., van der Kooi A. W., Numan T., Koek H. L., Klijn F. A. M., Buijsrogge M. P., et al.. (2014). Decreased functional connectivity and disturbed directionality of information flow in the electroencephalography of intensive care unit patients with delirium after cardiac surgery. Anesthesiology 121, 328–335. 10.1097/ALN.0000000000000329 [PubMed] [CrossRef] [Google Scholar]
Vanhaudenhuyse A., Demertzi A., Schabus M., Noirhomme Q., Bredart S., Boly M., et al.. (2011). Two distinct neuronal networks mediate the awareness of environment and of self. J. Cogn. Neurosci. 23, 570–578. 10.1162/jocn.2010.21488 [PubMed] [CrossRef] [Google Scholar]
Vlisides P. E., Bel-Bahar T., Lee U. C., Li D., Kim H., Janke E., et al.. (2017). Neurophysiologic correlates of ketamine sedation and anesthesia: a high-density electroencephalography study in healthy volunteers. Anesthesiology 127, 58–69. 10.1097/ALN.0000000000001671 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Vlisides P. E., Bel-Bahar T., Nelson A., Chilton K., Smith E., Janke E., et al.. (2018). Subanaesthetic ketamine and altered states of consciousness in humans. Br. J. Anaesth. 121, 249–259. 10.1016/j.bja.2018.03.011 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Vutskits L. (2018). General anesthetics to treat major depressive disorder: clinical relevance and underlying mechanisms. Anesth. Analg. 126, 208–216. 10.1213/ANE.0000000000002594 [PubMed] [CrossRef] [Google Scholar]
Wang J., Noh G. J., Choi B. M., Ku S. W., Joo P., Jung W. S., et al.. (2017). Suppressed neural complexity during ketamine- and propofol-induced unconsciousness. Neurosci. Lett. 653, 320–325. 10.1016/j.neulet.2017.05.045 [PubMed] [CrossRef] [Google Scholar]
Warnaby C. E., Sleigh J. W., Hight D., Jbabdi S., Tracey I. (2017). Investigation of slow-wave activity saturation during surgical anesthesia reveals a signature of neural inertia in humans. Anesthesiology 127, 645–657. 10.1097/ALN.0000000000001759 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Xie G., Deschamps A., Backman S. B., Fiset P., Chartrand D., Dagher A., et al.. (2011). Critical involvement of the thalamus and precuneus during restoration of consciousness with physostigmine in humans during propofol anaesthesia: a positron emission tomography study. Br. J. Anaesth. 106, 548–557. 10.1093/bja/aeq415 [PubMed] [CrossRef] [Google Scholar]
Articles from Frontiers in Systems Neuroscience are provided here courtesy of Frontiers Media SA
OTHER FORMATS
PDF (1.6M)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 962003 | https://no.wikipedia.org/wiki/Arbeidshukommelsen | Arbeidshukommelsen | Arbeidshukommelsen (engelsk: Working memory) er en modell for en del av menneskets hukommelse. Ifølge denne teorien er vår umiddelbare hukommelse del av et system som midlertidig lagrer og bearbeider informasjon mens vi utfører kognitive oppgaver (Baddeley & Hitch, 1974). Mens den relaterte termen ”korttids-hukommelse” ifølge Atkinson og Shiffrin (1968) fungerte som et enhetlig system, tydet forskning på at input og output forekommer i ulike modaliteter (Baddeley & Hitch, 1974). Ut fra dette kom Baddeley og Hitch (1974) frem til modellen for det som nå blir kalt arbeidshukommelsen.
Komponenter
Før 60-tallet var det dominerende synet på hukommelsen at den bestod av to systemer: Kortidshukommelse og langtidshukommelse. Imidlertid mente Baddeley & Hitch (1974) at menneskets hukommelse er mer komplekst organisert. I deres modell er arbeidshukommelsen satt sammen av fire ulike komponenter, som er modalitets-spesifikke. Mens komponenten central executive står for prosessering, står de tre nedre komponentene for midlertidig lagring. Komponentene har begrenset kapasitet, og fungerer relativt uavhengig av hverandre. Ut fra modellen har man dannet to prediksjoner: 1) To oppgaver som krever samme komponent vil trolig føre til såkalt interferens (at oppgavene påvirker hverandre på en måte som gjør utførelsen dårligere), og 2) To oppgaver som krever ulike komponenter kan løses uten videre interferens.
Den fonologiske løkke (Phonological loop)
Den fonologiske løkke står for midlertidig lagring av verbalt og auditorisk materiale. Dette er den mest definerte komponenten i arbeidshukommelsen, ettersom den har vært gjenstand for mest forskning. Den fonologiske løkke antas å være tale-basert av natur, og består av to interrelaterte komponenter: Det passive fonologiske lager, og den aktive artikulatoriske løkke.
Det er særlig tre effekter som kommer frem fra forskning på den fonologiske løkke:
Fonologisk likhetseffekt (Phonological similarity effect)
Ordlengde-effekt (Word length effect)
Irrelevant tale-effekt (Irrelevant speech effect)
Nevrovitenskapelig forskning antyder at oppgaver som krever den fonologiske løkke aktiverer deler av frontallappen og temporallappen i venstre hemisfære (eks. Baddeley, 2006). Videre ser det ut til at den fonologiske løkke lagrer auditorisk informasjon i parietallappen.
Visuo-spatial skisseblokk (Visuo-spatial sketchpad)
Den visuo-spatiale skisseblokk lar deg lagre et sammenhengende bilde av objekters form, samt deres relative posisjon i en scene. (eks. Cornoldi & Vecchi, 2003). Komponenten behandler også visuelle fremstillinger man innkoder ut fra ord/fortellinger (Baddeley, 2006).
Det eksisterer mindre forskning på den visuo-spatiale skisseblokk enn på den fonologiske løkke. Et av problemene er at vi gjerne gir navn til visuelle stimuli, og dermed også behandler denne informasjonen med den fonologiske løkke.
Nevrovitenskapelig forskning antyder at visuelle og spatiale oppgaver i hovedsak aktiverer høyre hemisfære (Gazzaniga et al., 2002), samt baklappen (Baddeley, 2001).
Central executive
Central executive står for koordineringen av ulike prosesser, prosessering, resonnering og problemløsing. Hvor involvert denne komponenten er, avhenger av oppgavens kompleksitet – jo mer kompleks oppgave, jo høyere involvering. Man antar at denne komponenten i stor grad avhenger av frontallappen.
Central executive er fremdeles dårlig definert. Den antas å ha begrenset kapasitet, og grensene er uklare. Et vanlig mål for central executive-kapasitet er såkalt «random generation», hvor man skal produsere et tilfeldig mønster av et bestemt antall objekter, for eksempel tall. Dette antas å kreve bl.a. selv-monitorering.
Funn fra nevrovitenskapelig forskning tyder på at frontallappen er særlig viktig i oppgaver som krever involvering av central executive (eks. Baddeley, 2006).
Episodisk buffer
Arbeidshukommelsens siste komponent ble lagt til av Baddeley i 2000. Den episodiske buffer antas å integrere informasjon fra den fonologiske loop og den visuo-spatiale skisseblokk til helhetlige, multimodale episodiske representasjoner. Den spiller en nøkkelrolle i bindeprosesser i arbeidshukommelsen. Man antar at deler av informasjonen i den episodiske buffer går videre til å lagres i langtidshukommelsen (Matlin, 2009).
Den episodiske buffer er en relativt ny komponent, og det er gjennomført begrenset forskning rundt den. Den er dermed fremdeles vagt definert.
Forholdet mellom arbeidshukommelsen og langtidshukommelsen
Det er uvisst hvordan arbeidshukommelsen henger sammen med langtidshukommelsen. Man skiller hovedsakelig mellom gateway-hypotser og workspace-hypotser.
Ifølge gateway-hypoteser kommer informasjon til langtidshukommelsen via arbeidshukommelsen (eks. Baddeley & Hitch). Informasjon må dermed bearbeides i arbeidshukommelsen for å kunne lagres videre i langtidshukommelsen.
Ifølge workspace-hypoteser må ikke informasjon via arbeidshukommelsen til langtidshukommelsen. Derimot er arbeidshukommelsen den delen av langtidshukommelsen som vi til enhver tid har oppmerksomheten rettet mot (eks. Cowan).
Individuelle forskjeller
Individuelle forskjeller innen arbeidshukommelses-kapasitet ser blant annet ut til å henge sammen med intelligens (Cowan et al., 2007), om man har ADHD (Attention-deficit/hyperactivity disorder) (Karatekin, 2004), eller er deprimert (Christopher & MacDonald, 2005).
Referanser
Atkinson, R. C. & Shiffrin, R. M. (1968). Human memory: a proposed system and its control processes. In K. W. Spence & J. T. Spence (Eds.) The psychology of learning and motivation, vol. 2. New York: Academic Press. pp. 89-195.
Baddeley, A. D. (2000). The episodic buffer: A new component of working memory? Trends in Cognitive Sciences, 4(11). pp. 417-423
Baddeley, A. D. (2001). Is working memory still working? American psychologist, 56(11), pp. 849-864.
Baddeley, A. D. (2006). Working memory: An overview. In S. J. Pickering (Ed.), Working memory and education (pp. 3-31). Burlington, MA: Elsevier.
Baddeley, A. D. & Hitch, G. (1974). Working memory. In G. H. Bower (Ed.), The psychology of learning and motivation, vol. 8. New York: Academic Press, Inc.
Christopher, G. & MacDonald, J. (2005). The impact of clinical depression on working memory. Cognitive Neuropsychiatry, 10(5), pp. 379-399.
Cornoldi, C. & Vecchi, T. (2003). Visuo-spatial working memory and individual differences. New York: Psychology Press.
Cowan, N., Morey, C. C., Chen, Z. & Bunting, M. F. (2007). What do estimates of working memory capacity tell us? In N. Osaka & R. Logie (Eds), Working memory: Behavioural & neural correlates. New York: Oxford University Press.
Gazzaniga, M. S., Ivry, R. B. & Mangun, G. R. (2002). Cognitive neuroscience: The biology of the mind. New York: Norton.
Karatekin, C. (2004). A test of the integrity of the components of Baddeley’s model of working memory in attention-deficit/hyperactivity disorder (ADHD). Journal of Child Psychology and Psychiatry, 45(5), pp. 912-926.
Matlin, M. (2009). Cognition. The United States of America: John Wiley & Sons, Inc.
Menneskelig adferd
Problemløsning | norwegian_bokmål | 0.560539 |
shut_down_conciousness/syc20378557.txt |
Skip to content
Mayo Clinic Home Page
Care at
Mayo Clinic
Health
Library
For Medical
Professionals
Research & Education
at Mayo Clinic
Giving to
Mayo Clinic
Request appointment
Log in
Search
Diseases & Conditions
Traumatic brain injury
Request an appointment
Symptoms &
causes
Diagnosis &
treatment
Doctors &
departments
On this page
Overview
Symptoms
When to see a doctor
Causes
Risk factors
Complications
Prevention
Overview
Traumatic brain injury usually results from a violent blow or jolt to the head or body. An object that goes through brain tissue, such as a bullet or shattered piece of skull, also can cause traumatic brain injury.
Mild traumatic brain injury may affect your brain cells temporarily. More-serious traumatic brain injury can result in bruising, torn tissues, bleeding and other physical damage to the brain. These injuries can result in long-term complications or death.
Products & Services
A Book: Guide to the Comatose Patient
Symptoms
Traumatic brain injury can have wide-ranging physical and psychological effects. Some signs or symptoms may appear immediately after the traumatic event, while others may appear days or weeks later.
Mild traumatic brain injury
The signs and symptoms of mild traumatic brain injury may include:
Physical symptoms
Headache
Nausea or vomiting
Fatigue or drowsiness
Problems with speech
Dizziness or loss of balance
Sensory symptoms
Sensory problems, such as blurred vision, ringing in the ears, a bad taste in the mouth or changes in the ability to smell
Sensitivity to light or sound
Cognitive, behavioral or mental symptoms
Loss of consciousness for a few seconds to a few minutes
No loss of consciousness, but a state of being dazed, confused or disoriented
Memory or concentration problems
Mood changes or mood swings
Feeling depressed or anxious
Difficulty sleeping
Sleeping more than usual
Moderate to severe traumatic brain injuries
Moderate to severe traumatic brain injuries can include any of the signs and symptoms of mild injury, as well as these symptoms that may appear within the first hours to days after a head injury:
Physical symptoms
Loss of consciousness from several minutes to hours
Persistent headache or headache that worsens
Repeated vomiting or nausea
Convulsions or seizures
Dilation of one or both pupils of the eyes
Clear fluids draining from the nose or ears
Inability to awaken from sleep
Weakness or numbness in fingers and toes
Loss of coordination
Cognitive or mental symptoms
Profound confusion
Agitation, combativeness or other unusual behavior
Slurred speech
Coma and other disorders of consciousness
Children's symptoms
Infants and young children with brain injuries might not be able to communicate headaches, sensory problems, confusion and similar symptoms. In a child with traumatic brain injury, you may observe:
Change in eating or nursing habits
Unusual or easy irritability
Persistent crying and inability to be consoled
Change in ability to pay attention
Change in sleep habits
Seizures
Sad or depressed mood
Drowsiness
Loss of interest in favorite toys or activities
When to see a doctor
Always see your doctor if you or your child has received a blow to the head or body that concerns you or causes behavioral changes. Seek emergency medical care if there are any signs or symptoms of traumatic brain injury following a recent blow or other traumatic injury to the head.
The terms "mild," "moderate" and "severe" are used to describe the effect of the injury on brain function. A mild injury to the brain is still a serious injury that requires prompt attention and an accurate diagnosis.
Request an appointment
From Mayo Clinic to your inbox
Sign up for free and stay up to date on research advancements, health tips, current health topics, and expertise on managing health. Click here for an email preview.
Email *
Enter your email
Subscribe!
Learn more about Mayo Clinic’s use of data.
Causes
Traumatic brain injury is usually caused by a blow or other traumatic injury to the head or body. The degree of damage can depend on several factors, including the nature of the injury and the force of impact.
Common events causing traumatic brain injury include the following:
Falls. Falls from bed or a ladder, down stairs, in the bath, and other falls are the most common cause of traumatic brain injury overall, particularly in older adults and young children.
Vehicle-related collisions. Collisions involving cars, motorcycles or bicycles — and pedestrians involved in such accidents — are a common cause of traumatic brain injury.
Violence. Gunshot wounds, domestic violence, child abuse and other assaults are common causes. Shaken baby syndrome is a traumatic brain injury in infants caused by violent shaking.
Sports injuries. Traumatic brain injuries may be caused by injuries from a number of sports, including soccer, boxing, football, baseball, lacrosse, skateboarding, hockey, and other high-impact or extreme sports. These are particularly common in youth.
Explosive blasts and other combat injuries. Explosive blasts are a common cause of traumatic brain injury in active-duty military personnel. Although how the damage occurs isn't yet well understood, many researchers believe that the pressure wave passing through the brain significantly disrupts brain function.
Traumatic brain injury also results from penetrating wounds, severe blows to the head with shrapnel or debris, and falls or bodily collisions with objects following a blast.
Risk factors
The people most at risk of traumatic brain injury include:
Children, especially newborns to 4-year-olds
Young adults, especially those between ages 15 and 24
Adults age 60 and older
Males in any age group
Complications
Several complications can occur immediately or soon after a traumatic brain injury. Severe injuries increase the risk of a greater number of and more-severe complications.
Altered consciousness
Moderate to severe traumatic brain injury can result in prolonged or permanent changes in a person's state of consciousness, awareness or responsiveness. Different states of consciousness include:
Coma. A person in a coma is unconscious, unaware of anything and unable to respond to any stimulus. This results from widespread damage to all parts of the brain. After a few days to a few weeks, a person may emerge from a coma or enter a vegetative state.
Vegetative state. Widespread damage to the brain can result in a vegetative state. Although the person is unaware of surroundings, he or she may open his or her eyes, make sounds, respond to reflexes, or move.
It's possible that a vegetative state can become permanent, but often individuals progress to a minimally conscious state.
Minimally conscious state. A minimally conscious state is a condition of severely altered consciousness but with some signs of self-awareness or awareness of one's environment. It is sometimes a transitional state from a coma or vegetative condition to greater recovery.
Brain death. When there is no measurable activity in the brain and the brainstem, this is called brain death. In a person who has been declared brain dead, removal of breathing devices will result in cessation of breathing and eventual heart failure. Brain death is considered irreversible.
Physical complications
Seizures. Some people with traumatic brain injury will develop seizures. The seizures may occur only in the early stages, or years after the injury. Recurrent seizures are called post-traumatic epilepsy.
Fluid buildup in the brain (hydrocephalus). Cerebrospinal fluid may build up in the spaces in the brain (cerebral ventricles) of some people who have had traumatic brain injuries, causing increased pressure and swelling in the brain.
Infections. Skull fractures or penetrating wounds can tear the layers of protective tissues (meninges) that surround the brain. This can enable bacteria to enter the brain and cause infections. An infection of the meninges (meningitis) could spread to the rest of the nervous system if not treated.
Blood vessel damage. Several small or large blood vessels in the brain may be damaged in a traumatic brain injury. This damage could lead to a stroke, blood clots or other problems.
Headaches. Frequent headaches are very common after a traumatic brain injury. They may begin within a week after the injury and could persist for as long as several months.
Vertigo. Many people experience vertigo, a condition characterized by dizziness, after a traumatic brain injury.
Sometimes, any or several of these symptoms might linger for a few weeks to a few months after a traumatic brain injury. When a combination of these symptoms lasts for an extended period of time, this is generally referred to as persistent post-concussive symptoms.
Traumatic brain injuries at the base of the skull can cause nerve damage to the nerves that emerge directly from the brain (cranial nerves). Cranial nerve damage may result in:
Paralysis of facial muscles or losing sensation in the face
Loss of or altered sense of smell or taste
Loss of vision or double vision
Swallowing problems
Dizziness
Ringing in the ear
Hearing loss
Intellectual problems
Many people who have had a significant brain injury will experience changes in their thinking (cognitive) skills. It may be more difficult to focus and take longer to process your thoughts. Traumatic brain injury can result in problems with many skills, including:
Cognitive problems
Memory
Learning
Reasoning
Judgment
Attention or concentration
Executive functioning problems
Problem-solving
Multitasking
Organization
Planning
Decision-making
Beginning or completing tasks
Communication problems
Language and communications problems are common following traumatic brain injuries. These problems can cause frustration, conflict and misunderstanding for people with a traumatic brain injury, as well as family members, friends and care providers.
Communication problems may include:
Difficulty understanding speech or writing
Difficulty speaking or writing
Inability to organize thoughts and ideas
Trouble following and participating in conversations
Communication problems that affect social skills may include:
Trouble with turn taking or topic selection in conversations
Problems with changes in tone, pitch or emphasis to express emotions, attitudes or subtle differences in meaning
Difficulty understanding nonverbal signals
Trouble reading cues from listeners
Trouble starting or stopping conversations
Inability to use the muscles needed to form words (dysarthria)
Behavioral changes
People who've experienced brain injury may experience changes in behaviors. These may include:
Difficulty with self-control
Lack of awareness of abilities
Risky behavior
Difficulty in social situations
Verbal or physical outbursts
Emotional changes
Emotional changes may include:
Depression
Anxiety
Mood swings
Irritability
Lack of empathy for others
Anger
Insomnia
Sensory problems
Problems involving senses may include:
Persistent ringing in the ears
Difficulty recognizing objects
Impaired hand-eye coordination
Blind spots or double vision
A bitter taste, a bad smell or difficulty smelling
Skin tingling, pain or itching
Trouble with balance or dizziness
Degenerative brain diseases
The relationship between degenerative brain diseases and brain injuries is still unclear. But some research suggests that repeated or severe traumatic brain injuries might increase the risk of degenerative brain diseases. But this risk can't be predicted for an individual — and researchers are still investigating if, why and how traumatic brain injuries might be related to degenerative brain diseases.
A degenerative brain disorder can cause gradual loss of brain functions, including:
Alzheimer's disease, which primarily causes the progressive loss of memory and other thinking skills
Parkinson's disease, a progressive condition that causes movement problems, such as tremors, rigidity and slow movements
Dementia pugilistica — most often associated with repetitive blows to the head in career boxing — which causes symptoms of dementia and movement problems
Prevention
Follow these tips to reduce the risk of brain injury:
Seat belts and airbags. Always wear a seat belt in a motor vehicle. A small child should always sit in the back seat of a car secured in a child safety seat or booster seat that is appropriate for his or her size and weight.
Alcohol and drug use. Don't drive under the influence of alcohol or drugs, including prescription medications that can impair the ability to drive.
Helmets. Wear a helmet while riding a bicycle, skateboard, motorcycle, snowmobile or all-terrain vehicle. Also wear appropriate head protection when playing baseball or contact sports, skiing, skating, snowboarding or riding a horse.
Pay attention to your surroundings. Don't drive, walk or cross the street while using your phone, tablet or any smart device. These distractions can lead to accidents or falls.
Preventing falls
The following tips can help older adults avoid falls around the house:
Install handrails in bathrooms
Put a nonslip mat in the bathtub or shower
Remove area rugs
Install handrails on both sides of staircases
Improve lighting in the home, especially around stairs
Keep stairs and floors clear of clutter
Get regular vision checkups
Get regular exercise
Preventing head injuries in children
The following tips can help children avoid head injuries:
Install safety gates at the top of a stairway
Keep stairs clear of clutter
Install window guards to prevent falls
Put a nonslip mat in the bathtub or shower
Use playgrounds that have shock-absorbing materials on the ground
Make sure area rugs are secure
Don't let children play on fire escapes or balconies
Request an appointment
By Mayo Clinic Staff
Feb 04, 2021
Print
Show References
Diagnosis & treatment
Diseases & Conditions
Traumatic brain injury symptoms & causes
Patient Stories from Mayo Clinic
Unforeseen danger of a child's fall: Livia's remarkable recovery
Associated Procedures
Brain rehabilitation
CT scan
MRI
Products & Services
A Book: Guide to the Comatose Patient
CON-20378540
Mayo Clinic does not endorse companies or products. Advertising revenue supports our not-for-profit mission.
Advertising & Sponsorship
Policy
Opportunities
Ad Choices
Mayo Clinic Press
Check out these best-sellers and special offers on books and newsletters from Mayo Clinic Press.
Mayo Clinic on Incontinence
The Essential Diabetes Book
Mayo Clinic on Hearing and Balance
FREE Mayo Clinic Diet Assessment
Mayo Clinic Health Letter - FREE book
Find a doctor
Explore careers
Sign up for free e-newsletters
About Mayo Clinic
About this Site
Contact Us
Locations
Health Information Policy
Medicare Accountable Care Organization (ACO)
Media Requests
News Network
Price Transparency
Medical Professionals
AskMayoExpert
Clinical Trials
Mayo Clinic Alumni Association
Refer a Patient
Businesses
Executive Health Program
International Business Collaborations
Facilities & Real Estate
Supplier Information
Students
Admissions Requirements
Degree Programs
Student & Faculty Portal
Researchers
Research Faculty
Laboratories
International Patients
Appointments
Financial Services
International Locations & Offices
Charitable Care & Financial Assistance
Community Health Needs Assessment
Financial Assistance Documents – Arizona
Financial Assistance Documents – Florida
Financial Assistance Documents – Minnesota
Follow Mayo Clinic
X
Youtube
Facebook
Linkedin
Instagram
Get the Mayo Clinic app
Android App on Google Play
Apple Download on the App Store
Terms & Conditions
Privacy Policy
Notice of Privacy Practices
Notice of Nondiscrimination
Accessibility Statement
Advertising & Sponsorship Policy
Site Map
© 1998-2024 Mayo Foundation for Medical Education and Research (MFMER). All rights reserved.
Language:
English
Feedback | biology | 8732751 | https://sv.wikipedia.org/wiki/Traumatisk%20hj%C3%A4rnskada | Traumatisk hjärnskada | Traumatisk hjärnskada (engelska: traumatic brain injury, TBI) är en skada på hjärnan som orsakas av en yttre kraft. TBI är en huvudorsak till mortalitet och morbiditet över hela världen, särskilt hos barn och unga vuxna. Traumatiska hjärnskador är ungefär dubbelt så vanliga bland män som bland kvinnor. Under 1900-talet sågs en stor utveckling inom diagnostik och behandling som minskade dödstalen och förbättrade återhämtningen efter traumatisk hjärnskada. Förebyggande åtgärder inkluderar användning av säkerhetsbälten och hjälmar, att inte köra bil under alkoholpåverkan, fallförebyggande insatser hos äldre vuxna och säkerhetsåtgärder för barn.
TBI kan klassificeras baserat på svårighetsgrad, skademekanism, eller andra faktorer. Skadans svårighetsgrad sträcker sig från mild traumatisk hjärnskada [mTBI/hjärnskakning] till allvarlig traumatisk hjärnskada, ofta baserat på medvetandegrad enligt Glasgow coma scale. Skademekanismen delas primärt in i sluten eller penetrerande huvudskada. Huvudskada är en bredare kategori som kan innebära skador på andra strukturer som hårbotten och skallen.
Skademekanism
Orsaker till TBI inkluderar fall, fordonskollisioner och våld. Hjärnskada uppstår antingen som en konsekvens av en plötslig acceleration eller deceleration mellan hjärna och kraniet, eller av en komplex kombination av både rörelse och kollision mellan skallen och yttre föremål. Utöver de skador som orsakades i ögonblicket för skadan, kan en mängd olika händelser efter skadan resultera i ytterligare skada. Dessa processer inkluderar förändringar i hjärnans blodflöde och intrakraniellt tryck.
Diagnostik
Diagnos sker med hjälp av anamnes, statustagning och radiologisk avbildning. Några av de avbildningstekniker som används för diagnos inkluderar datortomografi (CT) och magnetisk resonanstomografi (MRI), där datortomografi dominerar i akutskedet.
Prognos
TBI kan resultera i kroppsliga, kognitiva, sociala, emotionella och beteendemässiga symtom, och återhämtningen efter TBI kan variera från fullständig återhämtning till permanent funktionsnedsättning eller död. Beroende på skadan kan det krävas olika behandling, från observation till att innefatta läkemedel, akut kirurgi eller kirurgi i senare skede. Sjukgymnastik, logopedi och arbetsterapi kan användas för rehabilitering.
Referenser
Neurotrauma
Akuta sjukdomstillstånd | swedish | 0.588841 |
shut_down_conciousness/Neocortex.txt | The neocortex, also called the neopallium, isocortex, or the six-layered cortex, is a set of layers of the mammalian cerebral cortex involved in higher-order brain functions such as sensory perception, cognition, generation of motor commands, spatial reasoning and language. The neocortex is further subdivided into the true isocortex and the proisocortex.
In the human brain, the cerebral cortex consists of the larger neocortex and the smaller allocortex, respectively taking up 90% and 10%. The neocortex is made up of six layers, labelled from the outermost inwards, I to VI.
Etymology[edit]
The term is from cortex, Latin, "bark" or "rind", combined with neo-, Greek, "new". Neopallium is a similar hybrid, from Latin pallium, "cloak". Isocortex and allocortex are hybrids with Greek isos, "same", and allos, "other".
Anatomy[edit]
The neocortex is the most developed in its organisation and number of layers, of the cerebral tissues. The neocortex consists of the grey matter, or neuronal cell bodies and unmyelinated fibers, surrounding the deeper white matter (myelinated axons) in the cerebrum. This is a very thin layer though, about 2–4 mm thick. There are two types of cortex in the neocortex, the proisocortex and the true isocortex. The pro-isocortex is a transitional area between the true isocortex and the periallocortex (part of the allocortex). It is found in the cingulate cortex (part of the limbic system), in Brodmann's areas 24, 25, 30 and 32, the insula and the parahippocampal gyrus.
Of all the mammals studied to date (including humans), a species of oceanic dolphin known as the long-finned pilot whale has been found to have the most neocortical neurons.
Geometry[edit]
The neocortex is smooth in rodents and other small mammals, whereas in elephants, dolphins and primates and other larger mammals it has deep grooves (sulci) and ridges (gyri). These folds allow the surface area of the neocortex to be greatly increased. All human brains have the same overall pattern of main gyri and sulci, although they differ in detail from one person to another. The mechanism by which the gyri form during embryogenesis is not entirely clear, and there are several competing hypotheses that explain gyrification, such as axonal tension, cortical buckling or differences in cellular proliferation rates in different areas of the cortex.
Layers[edit]
Neurons form distinct layers in mouse visual cortex. Layer II/III (green), Layer IV (purple), Layer V (red), Layer VI (yellow). 3D reconstructions from the MICrONS cubic millimeter.
The neocortex contains both excitatory (~80%) and inhibitory (~20%) neurons, named for their effect on other neurons. The human neocortex consists of hundreds of different types of cells. The structure of the neocortex is relatively uniform (hence the alternative names "iso-" and "homotypic" cortex), consisting of six horizontal layers segregated principally by cell type and neuronal connections. However, there are many exceptions to this uniformity; for example, layer IV is small or missing in the primary motor cortex. There is some canonical circuitry within the cortex; for example, pyramidal neurons in the upper layers II and III project their axons to other areas of neocortex, while those in the deeper layers V and VI often project out of the cortex, e.g. to the thalamus, brainstem, and spinal cord. Neurons in layer IV receive the majority of the synaptic connections from outside the cortex (mostly from thalamus), and themselves make short-range, local connections to other cortical layers. Thus, layer IV is the main recipient of incoming sensory information and distributes it to the other layers for further processing.
Cortical columns[edit]
The column is the function unit of computation in the cortex. Neurons are color-coded by their layer: Layer II/III (green), Layer IV (purple), Layer V (red), Layer VI (yellow).
The neocortex is often described as being arranged in vertical structures called cortical columns, patches of neocortex with a diameter of roughly 0.5 mm (and a depth of 2 mm, i.e., spanning all six layers). These columns are often thought of as the basic repeating functional units of the neocortex, but their many definitions, in terms of anatomy, size, or function, are generally not consistent with each other, leading to a lack of consensus regarding their structure or function or even whether it makes sense to try to understand the neocortex in terms of columns.
Function[edit]
The neocortex is derived embryonically from the dorsal telencephalon, which is the rostral part of the forebrain. The neocortex is divided, into regions demarcated by the cranial sutures in the skull above, into frontal, parietal, occipital, and temporal lobes, which perform different functions. For example, the occipital lobe contains the primary visual cortex, and the temporal lobe contains the primary auditory cortex. Further subdivisions or areas of neocortex are responsible for more specific cognitive processes. In humans, the frontal lobe contains areas devoted to abilities that are enhanced in or unique to our species, such as complex language processing localized to the ventrolateral prefrontal cortex (Broca's area). In humans and other primates, social and emotional processing is localized to the orbitofrontal cortex.
The neocortex has also been shown to play an influential role in sleep, memory and learning processes. Semantic memories appear to be stored in the neocortex, specifically the anterolateral temporal lobe of the neocortex. It is also involved in instrumental conditioning; responsible for transmitting sensory information and information about plans for movement to the basal ganglia. The firing rate of neurons in the neocortex also has an effect on slow-wave sleep. When the neurons are at rest and are hyperpolarizing, a period of inhibition occurs during a slow oscillation, called the down state. When the neurons of the neocortex are in the excitatory depolarizing phase and are firing briefly at a high rate, a period of excitation occurs during a slow oscillation, called the up state.
Clinical significance[edit]
Lesions that develop in neurodegenerative disorders, such as Alzheimer's disease, interrupt the transfer of information from the sensory neocortex to the prefrontal neocortex. This disruption of sensory information contributes to the progressive symptoms seen in neurodegenerative disorders such as changes in personality, decline in cognitive abilities, and dementia. Damage to the neocortex of the anterolateral temporal lobe results in semantic dementia, which is the loss of memory of factual information (semantic memories). These symptoms can also be replicated by transcranial magnetic stimulation of this area. If damage is sustained to this area, patients do not develop anterograde amnesia and are able to recall episodic information.
Evolution[edit]
See also: Pallium (neuroanatomy) § Evolution
The neocortex is the newest part of the cerebral cortex to evolve (hence the prefix neo meaning new); the other part of the cerebral cortex is the allocortex. The cellular organization of the allocortex is different from the six-layered neocortex. In humans, 90% of the cerebral cortex and 76% of the entire brain is neocortex.
For a species to develop a larger neocortex, the brain must evolve in size so that it is large enough to support the region. Body size, basal metabolic rate and life history are factors affecting brain evolution and the coevolution of neocortex size and group size. The neocortex increased in size in response to pressures for greater cooperation and competition in early ancestors. With the size increase, there was greater voluntary inhibitory control of social behaviors resulting in increased social harmony.
The six-layer cortex appears to be a distinguishing feature of mammals; it has been found in the brains of all mammals, but not in any other animals. There is some debate, however, as to the cross-species nomenclature for neocortex. In avians, for instance, there are clear examples of cognitive processes that are thought to be neocortical in nature, despite the lack of the distinctive six-layer neocortical structure. Evidence suggest the avian pallium to be broadly equivalent to the mammalian neocortex. In a similar manner, reptiles, such as turtles, have primary sensory cortices. A consistent, alternative name has yet to be agreed upon.
Neocortex ratio[edit]
The neocortex ratio of a species is the ratio of the size of the neocortex to the rest of the brain. A high neocortex ratio is thought to correlate with a number of social variables such as group size and the complexity of social mating behaviors. Humans have a large neocortex as a percentage of total brain matter when compared with other mammals. For example, there is only a 30:1 ratio of neocortical gray matter to the size of the medulla oblongata in the brainstem of chimpanzees, while the ratio is 60:1 in humans.
See also[edit]
List of regions in the human brain
Blue Brain, a project to produce a computer simulation of a neocortical column and eventually a whole neocortex
Memory-prediction framework, a theory of the neocortex function by Jeff Hawkins and related software models
Claustrum | biology | 4863944 | https://sv.wikipedia.org/wiki/Entorhinal%20cortex | Entorhinal cortex | Entorhinal cortex eller entorinala barken är en del av hjärnan som är lokaliserad till den mittersta ytan av tinningloben och en del av det limbiska systemet. Dess funktion är att förmedla information från neocortex till hippocampus och vice versa.
Entorinala cortex är ett område i hjärnans mediala temporallob som angränsar till hippocampus och med vilken den har både efferenta och afferenta kopplingar. Entorinala cortex har även kopplingar med en stor del av neocortex och bearbetar minne och spatial information aktivt. De två delområdena i entorinala barken benämns laterala entorinala cortex och mediala entorinala cortex. Gridceller är en av flera specialiserade neuron i entorinala cortex som bidrar till däggdjurs förmåga att navigera sig fram i tillvaron. Ett av de områden som drabbas värst vid Alzheimers sjukdom är entorinala cortex och redan tidigt i sjukdomsförloppet uppstår en markant degeneration av neuron.
Anatomi
Entorinala barken är beläget i temporallobens mediala del och är område 28 i Brodmanns uppdelning av hjärnan. Det består av mediala entorinala cortex och laterala entorinala cortex och utöver denna uppdelning så delas även entorinala cortex in i laterala, mellanliggande och mediala banden. Dessa tre band har kopplingar till olika delar av gyrus dentatus i hippocampus och även om det inte är klarlagt exakt hur dessa kopplingar är avgränsade så finns det andra tydligare beskrivningar av kopplingar mellan entorinala cortex och hippocampus. Den primära afferenta banan till hippocampus är perforant pathway där neuron från lager två i entorinala barken projiceras till dentate gyrus och område CA3 i hippocampus, neuron från lager tre i entorinala cortex projiceras till område CA1 i hippocampus. En alternativ nervbana som benämns temporoammonic pathway går från lager tre i entorinala cortex till CA1 och subiculum i hippocampus. Det finns även banor som projicerar tillbaka till entorinala cortex djupare lager från subiculum och CA1 i hippocampus.
Entorinala cortex verkar fungera som en förbindelselänk mellan hippocampus och andra kortikala och subkortikala delar av hjärnan. Detta verkar också vara i form av en aktiv sortering och bearbetning av spatial och semantisk information. Det finns ingående banor i laterala entorinala cortex till exempel från piriforma hjärnbarken, insula, amygdala och luktsinnet. Vad gäller mediala entorinala cortex så verkar det komma förhållandevis fler (än vad gäller laterala entorinala barken) inkommande banor från occipitalloben, parietalloben, gördelvindlingen och dorsala thalamus.
En faktor som påverkar hjärnans plasticitet är nervbanan från entorinala cortex till CA1 i hippocampus, eftersom dorsala hippocampus huvudsakliga efferenta flöde kommer från CA1. Det kan även finnas liknande kopplingar mellan mediala entorinala cortex och ventrala hippocampus. Även om det verkar som att laterala entorinala cortex i huvudsak sänder vidare icke spatial information och att mediala entorinala cortex är mer specialiserat på att sända vidare spatial information, så har de båda delarna av entorinala cortex interna förbindelser som blandar spatial och icke spatial information innan den skickas vidare till hippocampusformationen.
Celler
Upptäckten av gridceller gjordes 2005 av May-Britt Moser, Edvard Moser, Torkel Hafting, Marianne Fyhn och Sturla Molden och dessa celler hittades i dorsocaudala mediala entorinala barken. Gridceller (eller rutnätsceller) är neuron som fyrar av så att det formar triangulära eller hexagonala mönster i omgivningen som en individ rör sig inom. Detta är specifikt för entorinala cortex och skiljer sig till exempel från hur det ser ut i hippocampus (där platsceller fyrar av efter miljön på ett annat sätt men med hjälp av informationen från gridcellerna). Gridcellers hexagonala mönster sammanlänkar omgivningen genom att olika gridceller fyrar av (efter hexagonala mönster) vid olika platser i omgivningen. Gridceller verkar avfyra efter större avstånd i omgivningen ju mer centralt de ligger i dorsocaudala mediala entorinala barken. Gridcellerna behöver ingen visuell input för att kunna läsa av omgivningen. Rutnätsliknande celler har hittats i såväl råttor som i möss, fladdermöss och inte minst i människor. Eftersom de återfunnits hos så pass många olika arter kan de tänkas ha varit viktiga sedan långt tillbaka i däggdjurens historia. Rutnätsmönstren är påfallande lika mellan olika arter. Entorinala barken innehåller även head directionceller (huvudriktningsceller) som avfyrar efter individens huvudriktning. Border cells (gränsceller) läser av avstånd och gränser i omgivningen. Hastighetsceller finns i alla lager i entorinala barken, dessa är interneuron (så kallade fast spiking cells). Integreringen av head directionceller och hastighetsceller möjliggör rutnätsceller att avfyra vid specifika platser. Gridceller, head directionceller och borderceller projicerar från entorinala barken till hippocampus och informationen som förs över används av hippocampus platsceller för att läsa av en individs omgivning.
Den laterala entorinala barkens neuron skickar mest signaler om objekt till hippocampus och är därmed mer semantisk (och inte spatial) än mediala entorinala barken. Laterala entorinala barken skickar även information om doft till hippocampus. Det verkar som att framplockningssignaler från dofter om platser lagras i hippocampus men verkar vara beroende av input från laterala entorinala barken för att kunna plockas fram.
Celler i mediala entorinala cortex är modulärt organiserade men man vet ej om dessa moduler representerar den spatiala miljön. Modulsystemet i mediala entorinala barken kan underlätta vid minneslagring.
Forskning
Mycket av forskningen rörande entorinala barken är utförd genom djurstudier vilket innebär en problematik att dra slutsatser av människors hjärnor. Det är vanligt med studier på råttor och en anledning till det är att funktioner i entorinala barken verkar vara liknande mellan råttor och människor (trots stora skillnader i hur de respektive hjärnorna ser ut i övrigt). Ett annat problem är att laboratoriemiljöer ofta är begränsade vilket gör det svårt att förutsäga hur den spatiala förmågan hos den entorinala barken skulle fungera i en oändlig miljö.
Skador och defekter
H.M. är ett av neuropsykologins mest kända fall och efter svåra epileptiska problem bestämdes att delar av hans mediala temporallob skulle opereras bort. Efter operationen blev epilepsin lindrigare men han drabbades dessvärre av allvarlig anterograd amnesi vilket innebar en total oförmåga för honom att lagra in nya episodiska minnen. I operationen togs bland annat entorinala cortex bort bilateralt vilket i stor utsträckning tros ha bidragit till amnesin. Detta har jämförts med lesionsstudier på apor där denna teori har styrkts.
Entorinala cortex och Alzheimers sjukdom
Vid Alzheimers sjukdom uppges det vara en av de strukturer som lider de allra svåraste skadorna. Man har sett omfattande degeneration av neuron i entorinala barkens kortikala lager två hos personer i tidiga steg av Alzheimers sjukdom. Det kan vara i storleksordningen av 50 procent av cellerna och denna celldegeneration ses inte hos normalt åldrade personer. Det verkar vara viktigt att gå in och titta på specifika lager och inte förlita sig på den totalt minskade volymen av entorinala barken då skadorna kan vara starkt kopplade till specifika lager. Strukturell MRI är därmed en lämplig avbildningsmetod som kan användas för att undersöka tidiga stadier av Alzheimers sjukdom. Med strukturell MRI bedöms vidden av atrofi i de specifika cellager som kan vara drabbade, vilket gör det lättare att avgöra om det handlar om Alzheimers sjukdom eller försämrat minne på grund av åldrande.
Även diffusionstensoravbildning kan vara en lämplig metod att använda då den mäter nervbanor mellan olika hjärnstrukturer. Diffusionstensoravbildning utnyttjar vattenmolekylers egenskap att röra sig längs nervfibrers riktning till skillnad från mer slumpmässig molekulär rörelse i ventriklar och cellkroppar. Perforant pathway innehåller neuron från entorinala kortikala lager två och degeneration av neuron där kan vara kopplat till tidiga stadier av Alzheimer, med diffusionstensoravbildning kan en sådan degeneration upptäckas.
Nobelpris
Makarna May-Britt och Edvard Mosers upptäckt av gridcellerna resulterade 2014 i att de, tillsammans med John O˙Keefe, tilldelades Nobelpriset i fysiologi eller medicin.
Källor
Referenser
Moser E, Kropff E, Moser M (2008). ”Place cells, grid cells, and the brain's spatial representation system”. Annual review of neuroscience 31: 69-89. doi: 10.1146/annurev.neuro.31.061307.090723.
Kerr K, Agster K, Furtak S, Burwell R (2007). ”Functional neuroanatomy of the parahippocampal region: the lateral and medial entorhinal areas”. Hippocampus 17 (9): 697-708. doi: http://dx.doi.org/10.1002/hipo.20315.
Fyhn M, Molden S, Witter M, Moser E, Moser M (2004). ”Spatial representation in the entorhinal cortex”. Science 305 (5688): 1258-1264. doi: 10.1126/science.1099901.
Witter M, Moser E (2006). ”Spatial representation and the architecture of the entorhonal cortex”. Trends in neurosciences 29 (12): 671-678. doi: 10.1016/j.tins.2006.10.003.
Stensola H, Stensola T, Solstad T, Frøland K, Moser M, Moser E (2012). ”The entorhinal grid map is discretized”. Nature 492 (7427): 72-78. doi: 10.1038/nature11649.
Noter
Storhjärnan | swedish | 0.577259 |
shut_down_conciousness/PMC10539891.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List BJA Open v.8; 2023 Dec PMC10539891
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of bjaopen
BJA Open. 2023 Dec; 8: 100224.
Published online 2023 Sep 26. doi: 10.1016/j.bjao.2023.100224
PMCID: PMC10539891
PMID: 37780201
The nature of consciousness in anaesthesia
Javier Montupil,1,2,3,† Paolo Cardone,4,5,† Cécile Staquet,1,2 Arthur Bonhomme,4 Aline Defresne,1,2,3 Charlotte Martial,4,5 Naji L.N. Alnagger,4,5 Olivia Gosseries,4,5 and Vincent Bonhomme1,2,∗
Author information Article notes Copyright and License information PMC Disclaimer
Associated Data
Supplementary Materials
Go to:
Abstract
Neuroscientists agree on the value of locating the source of consciousness within the brain. Anaesthesiologists are no exception, and have their own operational definition of consciousness based on phenomenological observations during anaesthesia. The full functional correlates of consciousness are yet to be precisely identified, however rapidly evolving progress in this scientific domain has yielded several theories that attempt to model the generation of consciousness. They have received variable support from experimental observations, including those involving anaesthesia and its ability to reversibly modulate different aspects of consciousness. Aside from the interest in a better understanding of the mechanisms of consciousness, exploring the functional tenets of the phenomenological consciousness states of general anaesthesia has the potential to ultimately improve patient management. It could facilitate the design of specific monitoring devices and approaches, aiming at reliably detecting each of the possible states of consciousness during an anaesthetic procedure, including total absence of mental content (unconsciousness), and internal awareness (sensation of self and internal thoughts) with or without conscious perception of the environment (connected or disconnected consciousness, respectively). Indeed, it must be noted that unresponsiveness is not sufficient to infer absence of connectedness or even absence of consciousness. This narrative review presents the current knowledge in this field from a system-level, underlining the contribution of anaesthesia studies in supporting theories of consciousness, and proposing directions for future research.
Keywords: brain function, consciousness, general anaesthesia, mechanisms, theories
Contrary to common belief, consciousness does not simply disappear during general anaesthesia. The brain of anaesthetised patients goes through a series of different states with variable mental content and perception of the environment.1 As a consequence, the reversible alteration of consciousness by anaesthesia can serve as a basis for designing unique experimental paradigms aimed at exploring the neural correlates of different aspects of consciousness, including perception of the environment, self-awareness, and internal thoughts.2 Understanding these elements paves the way towards a better definition of the nature of human consciousness. Currently, the most prominent ideas about how anaesthesia changes consciousness postulate a disruption of the capacity of the brain to generate information, an unbinding of the integrative information processes, or both.3,4 These ideas emerged from studies exploring the spectrum of anaesthesia-induced changes in brain function, including functional or effective connectivity, topological properties of brain networks, evoked responses and sensory processing, and spatio-temporal dynamics of brain connectivity.2 Most of these studies compared the normal awake state with complete anaesthesia-induced unresponsiveness, thought to correspond to the absence of mental content. Work is underway, which not only explores the functional tenets of anaesthesia-induced unconsciousness, but other additional anaesthesia-induced states of consciousness. Identifying specific and recordable neural signatures of the internal and external awareness systems that underlie these states5 will facilitate the possibility of recognising them in individual patients during anaesthesia.
Parallel to anaesthesia research, and named the ‘hard problem’ of consciousness for neuroscientists,6 several theories have been proposed to model how consciousness is generated by brain activity. Merging knowledge of different origins into a unified framework is important, not only because theories of consciousness can guide anaesthesia research in the quest for identifying the signatures of the brain states of anaesthesia, but also because anaesthesia can help to confirm them. Consequently, this narrative review first aims at describing, from a system perspective, the most prominent theories explaining the generation of consciousness by the brain and how experiments involving anaesthesia, if any, have supported them. Secondly, we describe the current knowledge about the functional correlates of the different anaesthesia-induced brain states and their identified recordable signatures. Finally, we propose some future directions for research in this field.
Go to:
The neuroscientific conceptualisation of consciousness
The word ‘consciousness’ has various meanings, depending on the perspective from which it is approached, be it philosophy, religion, or neuroscience. Indeed, the study of consciousness has only recently entered the scope of neuroscience. As a vital starting point from which to launch testable scientific investigations, it is agreed that consciousness is generated by brain activity. This means that there must be neuroanatomical and neurophysiological substrates of consciousness, or neural correlates of consciousness (NCCs). NCCs are defined as the minimal set of neuronal mechanisms sufficient for any phenomenological aspect of consciousness to emerge.7 Theoretically, NCCs can be related to a specific mental content (content-specific NCC), or denote the minimal neural mechanisms that are together necessary and sufficient for presence of mental content (full NCC).8 The different phenomenological aspects of consciousness must first be precisely defined before identifying their corresponding NCC. Given the complexity of consciousness and its phenomenology, several teams of scientists have proposed theories to model how the different aspects of consciousness are generated in the brain.9 These theories are based on different features and constructs, but are not necessarily mutually exclusive.10 Before these are described below, we first give a description of several concepts related to consciousness and its contents in addition to outlining the current knowledge of brain functioning underlying consciousness.
One way of dissecting the components of consciousness is to differentiate phenomenal consciousness and access consciousness.11 The necessity of distinguishing these two entities arose from the observation that ‘unconscious’ processing exists, and that one can almost never explain the totality of an experience at any given time. Phenomenal consciousness characterises the experiential features of consciousness as a whole.12 It can be referred to as a global feeling of an experience (‘what it is like’10), of which not all aspects can be verbalised. Access consciousness is the content of consciousness that can be verbalised or manifested in behavioural terms, such as, for example, describing a good meal at the restaurant in one's own terms, or imperfectly mimicking the behaviour of a recently met friend. It occurs through the availability of conscious information from numerous cognitive processes which mediate functions such as working memory, verbal report or motor behaviours.13 In order for conscious access to arise, perceptual contents have to be available for these functions. Accordingly, it is possible to subdivide the NCC into phenomenal-NCC, that is the minimal neural basis of the phenomenal content of an experience that is different across experiences (e.g. between the experience of red or green), and access-NCC, referring to the neural information that can be consciously reported.11 Another matter of concern for scientists is to determine the features of the ‘phenomenal self’, which can be defined as the conscious experience of being someone. This led to the notion of minimal phenomenal selfhood, corresponding in its simplest form to the conscious experience of being oneself, a distinct entity with several capabilities such as global self-control and attention, and spatio-temporal orientation of own body.14,15 If an episode of consciousness occurs during anaesthesia, elements of access consciousness, phenomenal consciousness, and phenomenal selfhood are exhibited. However, there has been more scientific interest in access consciousness and phenomenal selfhood because of their reportability, which allows them to be studied more readily.
Consciousness can also be approached through its main clinical features, namely wakefulness, awareness, and responsiveness. This is a clinically useful and operational perspective for anaesthesiologists. Wakefulness (or the degree of arousal) can be defined as the prerequisite for the ability to open eyes, spontaneously or following a stimulus.16 It is sustained by complex neural systems emerging from the brainstem, projecting to the cortex, and controlling the sleep-wake cycle.17,18 Awareness results from the synergy of all functions mediated through cortical-subcortical interactions,19,20 including cognitive and affective features, external perceptions and, for humans, phenomenal selfhood.21 Awareness can itself be divided into two components, namely internal and external awareness. Internal awareness is composed of internal mental processes (inner speech or mind wandering). External awareness (connectedness) is the conscious perception of the environment through sensory modalities.5 These concepts enable a description of the majority of physiological, pathological or drug-induced consciousness states (Table 1). For example, in disconnected consciousness, there is internal but no external awareness25 (e.g. in dreams experienced during general anaesthesia or sleep). Connected consciousness is a state of connection with the environment, in which both external and internal awareness is present.2,22 Understanding and manipulating connectedness is particularly relevant during general anaesthesia, where we attempt to prevent patients perceiving their environment.
Table 1
Operational classification of phenomenological consciousness states related to access consciousness and phenomenological self, and the relative contribution of wakefulness, internal awareness, and external awareness to those states. Adapted with permission from Martial and colleagues.22 NREM, non-rapid eye movement sleep; REM, rapid eye moment sleep; MCS−, minimally conscious state −, with non-linguistic signs of conscious awareness; MCS+, minimally conscious state with clear evidence of expressive language functions. a Patients with partially preserved cortical activity compatible with MCS but not showing any behavioural sign of conscious awareness (although cognitive-motor dissociation may exist, and appropriate cortical responses to active paradigms be present as evidenced by functional neuroimaging). b Reportable internal consciousness experience with features that are specific to near-death experiences (out-of-body experience,…) but not associated to a critical event potentially leading to death.
Phenomenological state
Wakefulness
Internal awareness
External awareness (connectedness)
Responsiveness to verbal command
Normal awake consciousness ++++ ++++ ++++ ++++
General anaesthesia Deep 0 0 0 0
Disconnected consciousness (with dreams; e.g. ketamine anaesthesia) 0 ++ 0 0
Connected consciousness episodes ++ ++ ++ ++
Sleep NREM sleep without dream 0 0 0 0
NREM sleep with dreams 0 ++ 0 0
REM sleep with dreams 0 +++ 0 0
REM sleep with lucid dream23 0 +++ Unknown Possible
Severe brain injury Coma without dreams/near-death experiences 0 0 0 0
Coma with dreams/near-death experiences 0 0 Unknown 0
Unresponsive wakefulness syndrome + (fluctuating) 0 0 0
Minimally conscious state + (fluctuating) + + 0 (MCS−) or + (MCS+)
Minimally conscious state or non-behavioural minimally conscious statea,24 + (fluctuating) Possible Possible 0 (behaviourally, but appropriate cortical response may be present)
Emergence from minimally conscious state ++++ (fluctuating) ++ ++ +
Locked-in syndrome ++++ ++++ ++++ ++++
Others Drug-induced and psychotic hallucinations ++++ ++++ Variable Variable
Near-death experience 0 +++ Unknown Unknown
Near-death-like experiencesb Variable +++ Variable Variable
Open in a separate window
Connected consciousness must not be mistaken with responsiveness, the possibility to deliver a behavioural response. Volitional responsiveness, when present, is crucial to infer connected consciousness, however, even the absence of which does not equate to the absence of internal or external awareness.1,22,26 Additionally, it must not be confounded with reflex, automatic motor responses such as arm withdrawal in response to noxious stimulation. In fact, connected consciousness during anaesthesia is not necessarily associated with movements or responsiveness1,27, 28, 29 and it is therefore difficult to detect. Its incidence has been estimated to be close to 5% after laryngoscopy, with higher incidences in young patients (11%) and women (13%),30 using the isolated forearm technique. This technique preserves one arm from muscle paralysis via a tourniquet, which enables response to command in such responders through squeezing of the hand. Although rarely followed by explicit recall but frequently ignored, the real consequences for patients of this frequent event demand attention.31
Go to:
The regulation of wakefulness and the generation of a mental content
Wakefulness
The degree of wakefulness determines the level of cortical arousal, where the synergy of cortical functions generates awareness. Wakefulness is modulated by the interplay between brainstem nuclei and cortical regions.32 In the neurotypical individual, two factors drive wakefulness: sleep pressure, or the biological circadian rhythm indicating when to sleep according to clock time and environmental cues, and sleep homeostasis, or the sleep debt increasing with length of wakefulness and decreasing with accumulating sleep time.33 The main timekeeper of circadian rhythms is the suprachiasmatic nucleus of the hypothalamus.34 Circadian rhythm manifests at the cellular and network level, involving several pathways and neurotransmitters35 and modulating brain activity,36 cognition,37 and cortical excitability.38 Sleep pressure, or sleep homeostasis, is mostly mediated by the concentration of adenosine diphosphate (ADP). ADP is a waste product of neural activity,39 and high concentrations of ADP are correlated with deeper slow-wave sleep.40 Adenosine inhibits neurons involved in the arousal circuits and excites neurons that promote sleep.32,41
The balance between wakefulness and sleep is a bi-stable flip-flop system,42 mediated by a delicate and complex neuroanatomical organisation with mutually inhibiting arousal-promoting and arousal-inhibiting networks.43 The arousal system is a variegated circuit involving several neurotransmitter pathways including neurons with large and diffuse projections. The most critical are glutamatergic and gamma-aminobutyric acid (GABA)ergic ones, but their activity is regulated by neuromodulators such as norepinephrine, serotonin, histamine, orexin, and acetylcholine.44 The ascending reticular activation system or, as recently referred to, the ascending arousal network (AAN), is a key source of neuromodulators for maintenance of wakefulness.45 AAN includes, among others, the reticular formation and the mesencephalic reticular formation (excitatory glutamatergic input to the cortex), the raphe nuclei (serotonin), the tuberomammillary nucleus (histamine), and the locus coeruleus (norepinephrine).32 The AAN is composed of a dorsal and a ventral pathway. The dorsal pathway innervates the non-specific thalamo-cortical system, a glutamatergic system in which discharge rate differentiates wakefulness and rapid eye movement sleep from non-rapid eye movement sleep and is responsible for a cortical up-state.46 The ventral pathway involves the lateral hypothalamus and the basal forebrain, where neurons produce orexin and acetylcholine, respectively.47,48
Modulation of the AAN by anaesthetics has been mainly studied in animal models.49 It is still unknown whether the alterations of consciousness by anaesthetics are attributable to changes in cortical arousal after a direct effect on the AAN, or whether these are secondary to initial cortical effects. Propofol, sevoflurane, and xenon reproducibly decrease the activity of the thalamus and of the AAN,50 whereas halothane causes locus coeruleus hyperpolarisation.50,51 Histaminergic neurotransmission has also been proposed as a key target,52 and cholinergic neurotransmission.53,54 The activity of the locus coeruleus is inhibited by α2-adrenoceptor agonist sedatives such as dexmedetomidine. This inhibition in turn activates the inhibitory ventro-lateral preoptic nucleus in the basal forebrain, which then exerts its inhibitory activity on a series of cortical arousal-promoting nuclei.55 This is mechanistically similar to non-rapid eye movement sleep.52,56 Through the inhibition of inhibitory interneurons, ketamine promotes cholinergic, dopaminergic, and other aminergic neurotransmission emerging from subcortical nuclei.57 Other subcortical modulations by anaesthetic agents have been described, notably on thalamo-cortical interactions, which appear to be selective. The non-specific thalamo-cortical connectivity is affected first, whereas interactions related to sensory processing are altered at higher doses.58 Notably, anaesthesia-induced unconsciousness can be reversed by deep brain stimulation of the central thalamus in non-human primates,59, 60, 61 indicating the importance of thalamo-cortical communication for consciousness. However these observations of anaesthesia reversal may be linked to an unspecific arousal effect on the cortex, as is observed after indirect boosting of cholinergic neurotransmission.54,62
Aside from these observations, several arguments also support a direct, region-specific, and dose-dependent effect of anaesthetic agents on cortical function.2,63 Agents with a primarily GABAergic effect are known to directly influence the activity of cortical pyramidal neurons.64 Ketamine inhibits GABAergic inhibitory interneurons at the cortical level, and produces a hyperglutamatergic state leading to the activation of specific cortical regions, the limbic system, and the hippocampus.65,66 Additionally, through an effect on other types of interneurons, ketamine causes spontaneously active cortical neurons to become silent, whereas previously silent neurons become active.67
The truth likely resides between these extremities of primarily a cortical or subcortical effect of anaesthetics, and likely depends on the considered agent and dose. However, the net result translates into cortical consequences that lead to the different anaesthetic states of consciousness. A summary of the known effects of anaesthetic agents on the AAN and on the cortex is provided in Figure 1.
Figure 1
Figure 1
Schematic representation of the complex systems regulating cortical arousal and controlling the sleep-wake cycle. Each assembly of neurons interacts with a series of other nuclei through complex excitatory and inhibitory projections (which are not all drawn for the sake of clarity). The final outputs of this complex network are ascending arousal-promoting or arousal-inhibiting cortical projections. Top-down cortical control of the activity of the subcortical structures also occurs. Stars indicate structures the activity of which is thought to be influenced by the administration of anaesthetic agents. For example, agents with a predominant gamma-aminobutyric acid (GABA)ergic effect (propofol or halogenated vapours; red star) potentiate the inhibition of cortical pyramidal neurons by inhibitory interneurons, boost the inhibitory effect of TR neurons on the cortex, and increase the inhibition of cortical arousal-promoting nuclei by the Vl and MPo, including the histaminergic Tm, the noradrenergic LC, the serotonergic DR, and the cholinergic BF, Pp, and LdT. The effects of ketamine (green star) are complex, and dose dependent. Through N-methyl-D-aspartate (NMDA) glutamate receptor antagonism, ketamine is known to inhibit GABAergic inhibitory interneurons at the cortical level, producing a hyper-glutamatergic state and the activation of specific cortical regions (anterior cingulate, medial prefrontal cortex, insula, and precuneus), the limbic system, and the hippocampus.65 66 This inhibitory interneuron inhibition also promotes cholinergic, dopaminergic, and other aminergic neurotransmission emerging from subcortical nuclei (e.g. TM, BF, Pp, and LdT). Aside from these effects, ketamine also inhibits excitatory-to-excitatory coupling in the cortex, provoking the inhibition of specific cortical regions.68 Alpha2-adrenergic agonists such as dexmedetomidine block the release of norepinephrine by LC. This has a direct inhibitory effect on cortical arousal-promoting nuclei such as BF and IL, or promotes the inhibition of other arousal-promoting nuclei (e.g. Tm, DR, PeF, Pp, and LdT) by Vl and MPo.55 Inspired by Moody and colleagues48 and from Purdon and colleagues.64 Drawn with BioRender. AAN, ascending arousal network; BF, basal forebrain; DR, dorsal raphe; IL, intralaminar nucleus of the thalamus; LC, locus coeruleus; LH, lateral hypothalamus; LHb, lateral habenula; Pb, parabrachial nucleus; PeF, perifornical area; Pp and LdT, pedunculo-pontine and latero-dorsal tegmentum nuclei; ReF, reticular formation; RmT, rostro-medial tegmental nucleus; So, supra-optic nucleus; Tm, tubero-mamillary nucleus; TR, thalamic reticular nucleus; Vl and MPo, ventro-lateral and median preoptic nuclei; VTA, ventral tegmental area.
Awareness
It is now commonly accepted that the emergence of awareness results from the collaborative work of, or communication between brain regions within and between brain networks.69 Adequate functioning of those networks requires an aroused cortex, but also involves subcortical structures.19,20 This exchange of information is thought to correspond to the aforementioned-mentioned NCCs.
Go to:
Exploring brain communication and exchange of information
Exploring brain communication noninvasively is possible through recording physiological signals from regional brain activity. The signal can be spontaneous or evoked, and recorded through magnetoencephalography (change in the magnetic field produced by neuronal activity), positron emission tomography (regional oxygen consumption or glucose metabolism, or regional cerebral blood flow), functional magnetic resonance imaging (fMRI, blood oxygen level-dependent or BOLD signal, a marker of regional cerebral blood flow), electroencephalography (EEG, electrical signal produced by neuronal activity), coupled or not with transcranial magnetic stimulation (TMS, stimulation of a cortical region to study the nature and spread of the evoked response), or functional near-infrared spectroscopy (fNIRS).2 Various characterisations and properties of the recorded signals are thereby analysed. For example, the activity evoked by a stimulus,70 synchrony in activity (functional connectivity), directional connectivity (effective connectivity),71 cortical excitability,38 complexity of interactions,72,73 amount of integrated information,74 or spatio-temporal interaction dynamics75, 76, 77 between brain regions, and within or between identified brain networks can be studied. Such networks can be further characterised by methods derived from the graph theory, or topological properties.78 In this case, a region is considered as a node that is linked to other nodes by edges. The node degree corresponds to its number of connections with other nodes. Nodes with a high degree (i.e. highly connected) play a crucial role for information transfer.79 These nodes spontaneously and incessantly reconfigure themselves to maintain an optimised state of organisation.80 Rather than being composed of static functional brain networks such as the default mode, salience, dorsal attention, executive control, sensory, or motor networks, the functional organisation of the brain is thought to occupy complex spatio-temporal dynamic of states of connectivity within and between brain networks. The brain activity underlying these networks can be characterised as existing in a state of criticality, thus hovering between several unstable (or metastable) states within a multi-stable attractor landscape.81
As one can see, the measures thought to reflect exchange of information within the brain are numerous (see Supplementary Table S1 for some examples). Each of them has been tentatively proposed to correspond to an NCC, according to changes observed when passing from one consciousness state to the other. However, it is difficult to confirm their effective NCC nature, because some of them can support consciousness but are not mandatory (e.g. those related to the AAN activity), or can display concomitant changes that are indeed independent from changes in a consciousness state.8,82
Go to:
Theories of consciousness
Several theories framing how brain activity can generate conscious content have emerged. Each of them focuses on a different feature,83 but none encompass the totality of the aforementioned concepts. It is beyond the scope of this paper to review all of them.9 In this review, we primarily focus on those theories addressing the phenomenological aspects of consciousness (access consciousness, phenomenal consciousness, and phenomenal selfhood), that is the subjective experience of an individual, as subjective experience during anaesthesia is the most important aspect for our patients. We also limit our description to the ones that have gained most attention in recent years and may have direct applications to the domain of anaesthesia. According to the classification of Seth and Bayne,9 we detail most prominent examples of global workspace theories (global neuronal workspace theory; GNW), integrated information theories (integrated information theory; IIT), re-entry and predictive processing theories (recurrent processing theory; RPT; predictive processing; PP), and higher-order theories (higher-order theory; HOT).
The global neuronal workspace hypothesis
GNW is a framework that explains consciousness by a widespread broadcasting of information through the brain. GNW considers that information only becomes conscious once it is sent towards the independent processors of a global workspace. Therefore, any information that is not broadcasted remains unconscious (Fig 2). In brief, local cortical processors are linked by a core set of highly interconnected areas, which can select pieces of information generated by local processors, amplify them, and broadcast the resulting set of information to all other processors of the global workspace in a top-down manner, rendering it conscious and reportable verbally. Processes of information selection, amplification, and sustainability are critical to trigger ignition and occur through recurrent processing phenomena (feedforward and feedback transfer of information or connectivity). The core set of highly interconnected brain regions is mainly composed of prefrontal and parietal areas. If the feedforward signal generated by local processors (sensory information or information of another nature) is strong enough, it reaches the prefrontal cortex, which in turn triggers ignition into the reverberant network, involving not only the posterior parietal cortex, but also other cortical and subcortical regions, including the thalamus and cerebellum.13
Figure 2
Figure 2
The global neuronal workspace hypothesis. In this theory, information (sensory or of another nature) generated within local processors of the brain becomes conscious if the feedforward signal from those processors is strong enough to generate ignition. Ignition allows the reverberation of the information within a global workspace, which is composed of highly interconnected independent processors. The independent processors (and particularly the frontal ones) can select relevant information, amplify it, and sustain it, and therefore control the process of ignition through recurrent processing (feedforward and feedback communication). Once in the global workspace, the information is broadcasted to all other processors of the workspace, again through recurrent processing, and becomes conscious (like being in the spotlight of a theatre). The global workspace is mainly composed of prefrontal and parietal areas, but also of other cortical and subcortical regions, including the thalamus and cerebellum (not shown in the figure, for the sake of clarity). Evoked responses to sensory stimuli and their timing are thought to be witnesses of recurrent processing within the global workspace, and of information reaching the field of consciousness. Anaesthesia is known to disrupt recurrent processing. Inspired by Seth and Bayne9 and Sergent and colleagues.70 Drawn with BioRender. GW, global workspace.
In this model, not all perceptual content enters conscious awareness. This is because of the all-or-none ignition of workspace activity, which allows the availability of a limited number of representations at a time.84 One could consider a theatre where only the small portion of the stage shown by the spotlight is perceived, while the remaining parts are ignored until they, in turn, enter the spotlight. A global workspace framework also explains why something can be partially reported (access consciousness), even if the entire experience itself (phenomenal consciousness) is richer than the one expressed. There is a ‘winner-takes-all’ competition among representations to be broadcasted within the global workspace.11 This has led to a long history of research looking for a ‘global signature’ of such broadcasting activity.85
Neural signatures of ignition have been previously identified in human and non-human primates,13 which are suppressed by anaesthesia.86 The most recent finding in this area is that, when a stimulus is consciously perceived, independently from being reported or not, a late electroencephalographic evoked activity is observed 250–300 ms after the stimulus (P3). This late activity is first seen in frontal areas, and then transferred to parietal regions, where it remains sustained up to 800–900 ms after the stimulus. This type of activity could be a signature of ignition and reverberant activity within the global workspace.70 Its maintenance or disappearance during anaesthesia and its associated possible different phenomenological brain states (unconsciousness, disconnected consciousness, and connected consciousness) are beginning to be studied.
It has been suggested that general anaesthesia primarily occurs via disruption of the reverberant global workspace functioning, and therefore of conscious access to information. Supporting this, EEG studies of directional connectivity in anaesthesia have shown that the functional connections from the frontal cortex towards more posterior areas (reflecting a frontal broadcasting of information) are pertinent,82,87,88,89 and the large-scale network top-down organisation,90,91 feedback (from higher- to lower-order cortical networks),91, 92, 93, 94, 95, 96, 97 and feedforward (the opposite pathway) connectivity,98, 99, 100, 101, 102 underlining the importance of recurrent processing. These effects do not preclude from basic sensory processing by lower-order sensory networks, as attested by the preservation of their connectivity when subjects are unresponsive.56,87,88,89
However, recent findings contradict the P3 as being a signature of ignition.103 P3 is observed during an awake connected state in response to oddball tones, but a similar response occurs in disconnected individuals in response to standard tones, as if all incoming sensory stimuli were interpreted as equally surprising despite not reaching the field of consciousness. This is associated with an increase in feedback connectivity within the auditory network thought to generate the P3 response.103
Others advocate that the dynamic state of the brain is differently affected by anaesthetic agents, without ever disappearing completely.104,105 Suppression of consciousness by anaesthesia would constrain the brain state towards more stable fluctuations and less temporal complexity,104,106 remaining further from critical dynamics,80,111 particularly regarding the dynamics of the main nodes of GNW.107 However, this view hardly explains how anaesthetised patients can be disconnected from their environment (i.e. sensory information does not reach the global workspace), while a mental content is still present in the form of a dream, indicating adequate functioning of the reverberant network. In this case, the feedforward sensory signal must be sufficiently and selectively attenuated as not to reach the prefrontal cortex, hence precluding it from ignition, while other internal inputs can still undergo ignition and reach the global workspace, thus enabling consciousness.
The integrated information theory
IIT bases its assumption on the phenomenological individual experience: conscious experience exists intrinsically (it has cause-effect power, meaning that it is possible to change its state), is structured (i.e. composed of different phenomenal units), is specific (it corresponds to a precise assembly of phenomenal units), is unitary (the conscious experience cannot be subdivided into a subset of other independent experiences), and is definite (regarding its composing elements and in time).108 Symmetrically, the physical substrate of consciousness must have an irreducible intrinsic cause-effect power that has the same properties.108 Hence, conscious experience corresponds to an irreducible maximum of integrated information generated by its physical substrate.9 The amount of integrated information is symbolised by the information theoretic quantity Φ, a measure of complexity. Complexity is a discrete measure that can be used to differentiate conscious states (e.g. normal wakefulness and awareness, dreaming, and hallucinogenic states) from the unconscious ones (e.g. deep sleep without dreams, general anaesthesia),109 but its value is not easy to determine with precision.109 This has led to a major criticism of the theory, some arguing that it is untestable. The solution to the problem could lie in trying to link some elements of consciousness with measures of information dynamics.110 Hence, some proxies for Φ have been proposed, based on the EEG power, frequency, functional connectivity, and modularity,74 based on the information content of the brain's response to TMS (perturbational complexity index or PCI72,111, 112, 113, 114, 115, 116, 117, 118), or obtained through interpretable deep learning analysis of the EEG [explainable consciousness indicator or ECI, able to distinguish the level of arousal (ECIaro) and awareness (ECIawa)].73 PCI or ECI critical values are suggested to clinically differentiate patients as a function of their level of arousal and awareness, such as minimally conscious state patients (fluctuating level of arousal, partially preserved awareness), unresponsive wakefulness syndrome patients (fluctuating level of arousal, no awareness),73,112 patients anesthetised with propofol or xenon (low level of arousal, no awareness), or patients anaesthetised with ketamine (low level of arousal, distorted internal awareness).73
As opposed to GNW, IIT proposes that a circumscribed posterior zone of the brain plays a key regulating role in the integration of information, the ‘posterior hot zone’ (Fig 3). To consciously perceive the external world, a condition is first to be sensitive to incoming stimuli.113 The information related to incoming stimuli is separately processed in segregated modules before being integrated. Those processes are thought to be reflected in surrogate measures of brain activity such as the BOLD signal of fMRI. Indeed, BOLD signal frequency is found to be narrower, lower, and in a dynamically distinct range during segregation, while being wider, and dynamically coherent during integration.114 The lower-order processing of information would occur in content-specific, segregated NCCs (e.g. face recognition in the occipital cortex). The integration of all information emerging from the content-specific NCCs would occur in the full NCC complex, namely the ‘posterior hot zone’ composed of temporal, parietal, and occipital cortical regions. This functioning would be influenced by enabling factors (e.g. the AAN determining the level of arousal), and factors modulating the activity of content-specific NCCs (e.g. attention and working memory, and sensory input to the cortex). The full NCC would provide output for motor responses and verbal report.8
Figure 3
Figure 3
The information integration theory (IIT). IIT postulates that the physical substrates of consciousness (neural correlates of consciousness, NCC) are of different natures. It distinguishes content-specific NCCs, which process specific aspects of information (auditory, visual, …), and full NCCs, which integrate all pertinent information into a mental content. Before reaching the full NCC, information from content-specific NCCs is modulated by specific brain functions such as working memory and attention, and segregated. The ‘posterior hot zone’, a hub involving parietal, occipital, and temporal cortical regions, plays a key role in controlling information integration by the full NCC. Integration of information can be quantified by the value Φ, a measure of brain activity complexity. Proxies of Φ have been proposed (see main text for details). The full NCC generates behavioural outputs, including motor and verbal report ones. Inspired by Seth and Bayne9 and Boly and colleagues.8 Drawn with BioRender. NCC, neural correlate of consciousness.
Using high-density EEG data acquired during propofol-induced unresponsiveness, dynamic causal modelling showed a selective breakdown of posterior parietal and medial feedforward fronto-parietal connectivity, and of parietal inter-network connectivity. These factors were found to be associated with loss of consciousness.115 This constitutes an argument in favour of the ‘posterior hot zone’ functional reality, in addition to the fact that posterior regions appear most relevant for quantifying arousal and awareness using ECI.73 It also supports the importance of communication between this zone and more anterior regions such as the anterior cingulate cortex for the integration of information and conscious content generation. This is in line with other anaesthesia and sleep studies investigating disconnected consciousness.116, 117, 118
The recurrent processing theory
RPT assumes that a percept becomes conscious after several steps only, which are the ‘feedforward sweep’ and local ‘recurrent processing’119 (Fig 4). In other words, it assumes that consciousness necessitates reverberation of the activity in the sensory areas, and therefore that consciousness is sustained by the continuous interplay between low- and high-level areas. The originality here is the assumption that information reverberating locally is still available and experienced, even if not broadcasted. The theory still makes a difference between what can be reported and what is really experienced, in the same line as phenomenal and access consciousness. Partial report paradigms have supported RPT, through the exploration of the dissociation between the amount of remembered information, and its estimation.121,122 This theory has received little but significant support from anaesthesia studies in humans.123,124
Figure 4
Figure 4
The recurrent processing theory (RPT). RPT has emerged following the study of visual perception. After sensory information incoming to the cortex (through the optic pathways, for example), feedforward sweep (green arrows) of information between visual areas (V1, V2, V4, V5, V7, and TE) occurs very rapidly for the interpretation of all the elements of the information (shape, localisation, …). It can lead to motor responses without any conscious perception of visual information. Localised recurrent processing (orange arrows) between visual areas allows fine tuning of information content, and may lead to a conscious percept. The occurrence of a reportable conscious percept would necessitate widespread recurrent processing (red arrows), not only between visual areas, but also with and within the fronto-parietal network. Inspired by Lamme.119 Drawn with BioRender. TE, temporal visual area; V1, V2, V4, V5, and V7, visual areas 1, 2, 4, 5, and 7, respectively.
Predictive processing
PP is currently grasping the attention of neuroscientists. PP is not a pure theory of consciousness, but rather a framework for understanding how the mind is organised. According to such view, the brain is a Bayesian machine, the role of which is to minimise surprise or free energy.125 As such, perception can be thought of as a ‘controlled hallucination’, a model that the brain constructs to navigate its surrounding.126 Lately, PP has served as a starting point to investigate modified states of consciousness, such as psychedelic experiences.127 According to PP,120 higher-order functional units constantly generate hypotheses of the sensory world and compare them against incoming sensory information. A feedback of the prediction is returned to sensory lower-order networks, and, in case of mismatch, a feedforward error message is sent to higher-order units with estimation of the precision of this information. Connected consciousness would therefore require balanced feedforward and feedback communication between higher-order and lower-order functional units. Alterations in the prediction, feedback, or both to lower-order sensory systems would result in unconsciousness, and alterations of feedforward messages, precision estimates, or both would characterise disconnected consciousness (Fig 5).
Figure 5
Figure 5
Predictive processing. Higher-order functional units generate hypotheses of the sensory world within canonical predictive coding circuits, and confront them to incoming sensory information (also processed in canonical predictive coding circuits of lower-order networks). A feedback of the prediction is returned to sensory lower-order networks (orange arrows), and, in case of mismatch, a feedforward error message is sent to higher-order units with estimation of the precision of this information (blue thin arrows). Alteration of feedback to lower-order sensory systems would result in unconsciousness, and alteration of feedforward in disconnected consciousness. Inspired by Seth and Bayne9 and Sanders and colleagues.120 Drawn with BioRender.
The higher-order theory
In contrast to RPT, HOT128 is based on the assumption that the mere activation of sensory regions is not sufficient for the emergence of phenomenal consciousness, but that a hierarchically higher-order region is instead necessary (Supplementary Fig. S1). Lower-order representations, in the sensory areas, for example, would become conscious when targeted by the right type of meta-representation, which would come from higher-order functional regions. The prefrontal cortex would play a key role in this metacognition.9 Interestingly, HOT implies the presence of an inner awareness, which makes it different from other theories such as GNW. Additionally, part of the added value of HOT is an accounting of emotive feeling and mental states,129 which have strong influence on conscious experience.128 Like RPT, HOT has not been investigated in studies involving anaesthesia, but several studies exploring consciousness networks during anaesthesia make the distinction between lower-order (or sensory) networks, and higher-order ones.
Caveats of consciousness theories
Each proposed theory has been challenged. For example, in opposition to assertions of GNW and HOT, anterior regions of the brain (frontal and prefrontal) have been involved in behavioural report, subjective report, and executive control rather than consciousness per se.9 In addition, the identification of their neural correlates is impeded by the ability to validate the proposed models of brain communication, and to distinguish the real NCCs of mental content from those that are only prerequisites for consciousness, or simply independent correlates.8 Stakeholders of these theories have different conceptions about consciousness, and communication between them can be uneasy. This has been a limiting factor for the reproducibility of results. The development of collaborative and adversarial projects will likely progress the situation thereby allowing the direct testing of opposing hypotheses130 (see, e.g., https://www.templetonworldcharity.org/accelerating-research-consciousness-our-structured-adversarial-collaboration-projects).130,131 Such collaborations have already led to studies confirming some prediction of IIT and GNW, but also challenging them at the same time (questioning the reality of the posterior hot zone for IIT, and ignition and the role of the prefrontal cortex for GNW).132
Go to:
Future directions
Recent progress is incrementally elucidating the complex nature of the effects of anaesthetic agents on the functional properties of consciousness. The current scientific literature contains a significant number of publications detailing changes in physiological brain signals when comparing a consciousness state to another, including studies using anaesthesia to modify consciousness. Those findings do not necessarily confirm that the considered physiological signal property really corresponds to an NCC, for the aforementioned reasons, and are not necessarily linked to a specific theory of consciousness. Through its ability to precisely and reversibly modulate different aspects of consciousness, anaesthesia offers unique possibilities to further progress in this domain, provided that researchers correctly assess consciousness states, namely connectedness and the presence of a mental content in an otherwise unresponsive subject. Progresses in these domains could help to provide evidence for theories of consciousness, and lead to the development of monitors facilitating, in patients with altered consciousness and unresponsive (either after brain damage or during anaesthesia), the detection of awareness of the environment, or the detection of disconnected consciousness. Such innovations might be closer than initially expected.
Current knowledge about anaesthesia-induced brain functional changes
Most studies investigating the brain functional changes induced by general anaesthesia have focused on single-drug sedation and have compared the full waking state with a state of unresponsiveness, supposed to be reflective of an absence of mental content. Functional changes that are common to all anaesthetic agents include a breakdown of higher-order networks (those involved in higher-order cognitive functions), and particularly of fronto-parietal connectivity, a relative preservation of lower-order networks (mainly sensory-motor networks, although between-network communication with higher-order networks might be compromised), an alteration of long distance cortico-cortical communication, and an alteration of the spatio-temporal dynamics in network interactions. Changes in communication dynamics involve a limitation of the repertoire of possible configurations, reduced complexity, reconfiguration of network structure, reduced efficiency, increased clustering and segregation, and a breakdown of the posterior ‘hot zone’.78 Authors disagree on the involved changes in communication directionality, both feedback91, 92, 93, 94 and feedforward,98, 99, 100, 101 suggesting that specific changes in feedforward or feedback connectivity throughout the cortical hierarchy might be involved in the mechanisms leading to the different anaesthesia-related brain states. As most of these studies were focused on the differences between wakefulness and drug-induced unresponsiveness thought to reflect unconsciousness, they did not specifically attempt to identify correlates of residual mental content and covert connectedness during anaesthesia. Therefore, we must consider the possibility that observed unresponsiveness-related functional changes may not correspond to a state of unconsciousness. This consideration renders the identification of the functional elements of anaesthesia-related brain states even more challenging.
Functional correlates of the states of consciousness during anaesthesia as a basis to identify their recordable signatures
The phenomenological state of disconnection from the environment supposes that incoming sensory stimuli are not consciously perceived. Knowing that functional connectivity within lower-order sensory-motor networks is usually preserved during deep sedation with a series of anaesthetic agents,89,133 indicating possible residual information processing at that level, disconnected consciousness could involve a disruption of communication between low-order sensory systems and higher-order cognitive systems, while the latter keep on maintaining internal information processing for the generation of mental content. Arguments that support this hypothesis are still scarce. Interestingly, propofol titration to loss of overt responsiveness during the performance of an auditory verbal memory task increases functional connectivity between the precuneus and other cortical regions, particularly the dorsal prefrontal and visual cortices.134 This could be in relation to a disconnected endogenous mentation. Strong evidence of a disruption of communication between lower-order and higher-order networks during disconnected consciousness, for example through a topological reconfiguration, is not yet available. Another possibility could be that disconnected consciousness occurs when information processing in lower-order networks is no longer generating information content (as assessed by the entropy of the principal components of the regional BOLD signal), while information content within higher-order networks remains of a high enough quality to generate internal mentation.135 Contrarily to this, others have shown that connected—compared with disconnected consciousness—necessitates preserved regional activity in midline structures of the brain including the thalamus, cingulate cortices, and angular gyri, which are components of higher-order consciousness networks.136 However, regional brain activity does not necessarily account for connectivity, and generation and exchange of information, rendering the interpretation of this finding difficult. It must also be kept in mind that the functional integrity of the dorsal anterior insular cortex appears to be necessary for a behavioural response to sensory stimuli,97 but this does not necessarily mean that such integrity is related to the conscious perception of external information.
Along with functional brain imaging studies trying to identify the functional correlates of anaesthesia-related brain states, the quest for easily recordable physiological markers of those states has now started. Slow-wave activity saturation in the EEG has long been advocated as a signature of perception loss during anaesthesia,137 but is probably not specific enough to distinguish between unconsciousness and disconnected consciousness. A frontal α-δ EEG pattern is almost constantly observed during surgical anaesthesia, and was first seen as indicative of unconsciousness.64 However, patients with this pattern might still be responsive to command, whereas patients without it can be unresponsive.138 It has been hypothesised that evoked occipital TMS-EEG α power is a marker of sensory information processing and hence of connectedness, whereas evoked low-γ power is necessary for consciousness, either connected or disconnected.139 In another study, disconnected consciousness was associated with broad spatial and spectral EEG changes, whereas unconsciousness was associated with focal decreases in activity in anterior and posterior cingulate cortices.116 Other examples are the perturbational distance metric, which assesses the ‘dynamical stability’ of a conscious state against perturbation, and hence its reversibility,140 or the above-mentioned PCI and ECI, whose value above a threshold is thought to correspond to the presence of awareness and to the level of arousal.72,73 The above-mentioned evoked cortical EEG responses to an auditory oddball paradigm may serve to screen for disconnection from the environment. During normal connected consciousness, oddball tones produce a P3 scalp EEG response. This P3 is absent during standard tone delivery. When disconnection occurs during sedation (while internal consciousness is still present), a similar oddball-evoked P3 response is observed during standard tones delivery. This could be as a result of increased feedback signalling in the auditory network, provoking a disruption of normal predictive coding processes, in that all incoming auditory stimuli become similarly surprising.103 The importance of attention processes in regulating access of sensory information to consciousness is considered in some of the above-described theories of consciousness (mainly in GNW, where it is associated with ignition, and in IIT). Attention is thought to be regulated by some of the higher-order consciousness networks (executive control network, dorsal attention network, and their relationship with the default-mode network), which can be differently and selectively affected by anaesthetic agents.87 Attention has recently been proposed as a core element of consciousness,141 and plays a role in patients' ability to give a purposeful response to a stimulus during anaesthesia.142 It also determines the ability to recall events after the procedure, as shown by a newly developed EEG-based index of attention (the cognitive effort index; CEI), which displays higher values in sedated patients who will have recall after the procedure.143
Despite these initial explorations into the discovery of specific markers of anaesthetic brain states, further research using optimal models to reliably discriminate the consciousness states of anaesthesia is needed. For instance, the isolated forearm technique and iterative awakening to ask for the presence of dreams, associated or not to sensory stimulation paradigms will aid progression in this respect.
Go to:
Authors’ contributions
Literature search, writing and revising paper: all authors.
Accountable for all aspects of the work: all authors.
Go to:
Declaration of interest
VB has had financial relationships with the following companies: Orion Pharma, Medtronic, Edwards, and Elsevier. PC and NA are research fellows and OG is research associate at F.R.S-FNRS. The other authors declare that they have no conflicts of interest.
Go to:
Funding
The Department of Anesthesia and Intensive Care Medicine (Liege University Hospital, Liege, Belgium), the Belgian National Fund for Scientific Research (FNRS), the European Union's Horizon 2020 Framework Programme for Research and Innovation (945539) (Human Brain Project SGA3), the BIAL Foundation, the Mind Science Foundation, the Leon Fredericq Foundation, and the GIGA Doctoral School for Health Sciences.
Go to:
Notes
Handling Editor: Phil Hopkins
Go to:
Footnotes
Appendix ASupplementary data to this article can be found online at https://doi.org/10.1016/j.bjao.2023.100224.
Go to:
Appendix A. Supplementary data
The following are the Supplementary data to this article.
Multimedia component 1:
Click here to view.(222K, docx)Multimedia component 1
Multimedia component 2:
Click here to view.(33K, docx)Multimedia component 2
Go to:
References
1. Sanders R.D., Tononi G., Laureys S., Sleigh J.W. Unresponsiveness ≠ unconsciousness. Anesthesiology. 2012;116:946–959. [PMC free article] [PubMed] [Google Scholar]
2. Bonhomme V., Staquet C., Montupil J., et al. General anesthesia: a probe to explore consciousness. Front Syst Neurosci. 2019;13:36. [PMC free article] [PubMed] [Google Scholar]
3. Hudetz A.G. General anesthesia and human brain connectivity. Brain Connect. 2012;2:291–302. [PMC free article] [PubMed] [Google Scholar]
4. Mashour G.A. Cognitive unbinding: a neuroscientific paradigm of general anesthesia and related states of unconsciousness. Neurosci Biobehav Rev. 2013;37:2751–2759. [PMC free article] [PubMed] [Google Scholar]
5. Vanhaudenhuyse A., Demertzi A., Schabus M., et al. Two distinct neuronal networks mediate the awareness of environment and of self. J Cogn Neurosci. 2011;23:570–578. [PubMed] [Google Scholar]
6. Klein C., Barron A.B. How experimental neuroscientists can fix the hard problem of consciousness. Neurosci Conscious. 2020;2020:niaa009. [PMC free article] [PubMed] [Google Scholar]
7. Crick F., Koch C. Towards a neurobiological theory of consciousness. Semin Neurosci. 1990;2:263–275. [Google Scholar]
8. Boly M., Massimini M., Tsuchiya N., Postle B.R., Koch C., Tononi G. Are the neural correlates of consciousness in the front or in the back of the cerebral cortex? clinical and neuroimaging evidence. J Neurosci. 2017;37:9603–9613. [PMC free article] [PubMed] [Google Scholar]
9. Seth A.K., Bayne T. Theories of consciousness. Nat Rev Neurosci. 2022;23:439–452. [PubMed] [Google Scholar]
10. Northoff G., Lamme V. Neural signs and mechanisms of consciousness: is there a potential convergence of theories of consciousness in sight? Neurosci Biobehav Rev. 2020;118:568–587. [PubMed] [Google Scholar]
11. Block N. Two neural correlates of consciousness. Trends Cogn Sci. 2005;9:46–52. [PubMed] [Google Scholar]
12. Fazekas P., Overgaard M. Perceptual consciousness and cognitive access: an introduction. Philos Trans R Soc Lond B Biol Sci. 2018;373 [PMC free article] [PubMed] [Google Scholar]
13. Mashour G.A., Roelfsema P., Changeux J.-P., Dehaene S. Conscious processing and the global neuronal workspace hypothesis. Neuron. 2020;105:776–798. [PMC free article] [PubMed] [Google Scholar]
14. Blanke O., Metzinger T. Full-body illusions and minimal phenomenal selfhood. Trends Cogn Sci. 2009;13:7–13. [PubMed] [Google Scholar]
15. Seth A.K., Tsakiris M. Being a beast machine: the somatic basis of selfhood. Trends Cogn Sci. 2018;22:969–981. [PubMed] [Google Scholar]
16. Bernat J.L. Chronic consciousness disorders. Annu Rev Med. 2009;60:381–392. [PubMed] [Google Scholar]
17. Schiff N.D. Central thalamic contributions to arousal regulation and neurological disorders of consciousness. Ann N Y Acad Sci. 2008;1129:105–118. [PubMed] [Google Scholar]
18. Satpute A.B., Kragel P.A., Barrett L.F., Wager T.D., Bianciardi M. Deconstructing arousal into wakeful, autonomic and affective varieties. Neurosci Lett. 2019;693:19–28. [PMC free article] [PubMed] [Google Scholar]
19. Lutkenhoff E.S., Johnson M.A., Casarotto S., Massimini M., Monti M.M. Subcortical atrophy correlates with the perturbational complexity index in patients with disorders of consciousness. Brain Stimul. 2020;13:1426–1435. [PubMed] [Google Scholar]
20. Luppi A.I., Cain J., Spindler L.R.B., et al. Mechanisms underlying disorders of consciousness: bridging gaps to move toward an integrated translational science. Neurocrit Care. 2021;35:37–54. [PMC free article] [PubMed] [Google Scholar]
21. Naccache L. Visual consciousness explained by its impairments. Curr Opin Neurol. 2015;28:45–50. [PubMed] [Google Scholar]
22. Martial C., Cassol H., Laureys S., Gosseries O. Near-death experience as a probe to explore (disconnected) consciousness. Trends Cogn Sci. 2020;24:173–183. [PubMed] [Google Scholar]
23. Konkoly K.R., Appel K., Chabani E., et al. Real-time dialogue between experimenters and dreamers during REM sleep. Curr Biol. 2021;31:1417–1427.e6. [PMC free article] [PubMed] [Google Scholar]
24. Thibaut A., Panda R., Annen J., et al. Preservation of brain activity in unresponsive patients identifies MCS Star. Ann Neurol. 2021;90:89–100. [PMC free article] [PubMed] [Google Scholar]
25. Brown E.N., Lydic R., Schiff N.D. General anesthesia, sleep, and coma. N Engl J Med. 2010;363:2638–2650. [PMC free article] [PubMed] [Google Scholar]
26. Boly M., Sanders R.D., Mashour G.A., Laureys S. Consciousness and responsiveness: Lessons from anaesthesia and the vegetative state. Curr Opin Anaesthesiol. 2013;26:444–449. [PubMed] [Google Scholar]
27. Linassi F., Zanatta P., Tellaroli P., Ori C., Carron M. Isolated forearm technique: a meta-analysis of connected consciousness during different general anaesthesia regimens. Br J Anaesth. 2018;121:198–209. [PubMed] [Google Scholar]
28. Radek L., Kallionpää R.E., Karvonen M., et al. Dreaming and awareness during dexmedetomidine- and propofol-induced unresponsiveness. Br J Anaesth. 2018;121:260–269. [PubMed] [Google Scholar]
29. Sanders R.D., Gaskell A., Raz A., et al. Incidence of connected consciousness after tracheal intubation: a prospective, international, multicenter cohort study of the isolated forearm technique. Anesthesiology. 2017;126:214–222. [PubMed] [Google Scholar]
30. Lennertz R., Pryor K.O., Raz A., et al. Connected consciousness after tracheal intubation in young adults: an international multicentre cohort study. Br J Anaesth. 2023;130:e217–e224. [PMC free article] [PubMed] [Google Scholar]
31. Hudson A.E. Presumption of insensibility during general anaesthesia. Br J Anaesth. 2023;130:e209–e212. [PubMed] [Google Scholar]
32. Brown R.E., Basheer R., McKenna J.T., Strecker R.E., McCarley R.W. Control of sleep and wakefulness. Physiol Rev. 2012;92:1087–1187. [PMC free article] [PubMed] [Google Scholar]
33. Borbély A.A., Daan S., Wirz-Justice A., Deboer T. The two-process model of sleep regulation: a reappraisal. J Sleep Res. 2016;25:131–143. [PubMed] [Google Scholar]
34. Mohawk J.A., Green C.B., Takahashi J.S. Central and peripheral circadian clocks in mammals. Annu Rev Neurosci. 2012;35:445–462. [PMC free article] [PubMed] [Google Scholar]
35. Rosenwasser A.M., Turek F.W. Neurobiology of circadian rhythm regulation. Sleep Med Clin. 2015;10:403–412. [PubMed] [Google Scholar]
36. Muto V., Jaspar M., Meyer C., et al. Local modulation of human brain responses by circadian rhythmicity and sleep debt. Science. 2016;353:687–690. [PubMed] [Google Scholar]
37. Schmidt C., Collette F., Cajochen C., Peigneux P. A time to think: circadian rhythms in human cognition. Cogn Neuropsychol. 2007;24:755–789. [PubMed] [Google Scholar]
38. Ly J.Q.M., Gaggioni G., Chellappa S.L., et al. Circadian regulation of human cortical excitability. Nat Commun. 2016;7 [PMC free article] [PubMed] [Google Scholar]
39. Greene R.W., Bjorness T.E., Suzuki A. The adenosine-mediated, neuronal-glial, homeostatic sleep response. Curr Opin Neurobiol. 2017;44:236–242. [PMC free article] [PubMed] [Google Scholar]
40. Radulovacki M., Virus R.M., Djuricic Nedelson M., Green R.D. Adenosine analogs and sleep in rats. J Pharmacol Exp Ther. 1984;228:268–274. [PubMed] [Google Scholar]
41. Korkutata M., Saitoh T., Cherasse Y., et al. Enhancing endogenous adenosine A2A receptor signaling induces slow-wave sleep without affecting body temperature and cardiovascular function. Neuropharmacology. 2019;144:122–132. [PubMed] [Google Scholar]
42. McGinty D., Szymusiak R. The sleep-wake switch: a neuronal alarm clock. Nat Med. 2000;6:510–511. [PubMed] [Google Scholar]
43. Sulaman B.A., Wang S., Tyan J., Eban-Rothschild A. Neuro-orchestration of sleep and wakefulness. Nat Neurosci. 2023;26:196–212. [PubMed] [Google Scholar]
44. Jones B.E. Arousal and sleep circuits. Neuropsychopharmacology. 2020;45:6–20. [PMC free article] [PubMed] [Google Scholar]
45. Edlow B.L., Takahashi E., Wu O., et al. Neuroanatomic connectivity of the human ascending arousal system critical to consciousness and its disorders. J Neuropathol Exp Neurol. 2012;71:531–546. [PMC free article] [PubMed] [Google Scholar]
46. Gent T.C., Bandarabadi M., Herrera C.G., Adamantidis A.R. Thalamic dual control of sleep and wakefulness. Nat Neurosci. 2018;21:974–984. [PMC free article] [PubMed] [Google Scholar]
47. Jones B.E. Principal cell types of sleep–wake regulatory circuits. Curr Opin Neurobiol. 2017;44:101–109. [PMC free article] [PubMed] [Google Scholar]
48. Moody O.A., Zhang E.R., Vincent K.F., et al. The neural circuits underlying general anesthesia and sleep. Anesth Analg. 2021;132:1254–1264. [PMC free article] [PubMed] [Google Scholar]
49. Bao W.-W., Jiang S., Qu W.-M., Li W.-X., Miao C.-H., Huang Z.-L. Understanding the neural mechanisms of general anesthesia from interaction with sleep-wake state: a decade of discovery. Pharmacol Rev. 2023;75:532–553. [PubMed] [Google Scholar]
50. Franks N.P. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nat Rev Neurosci. 2008;9:370–386. [PubMed] [Google Scholar]
51. Sirois J.E., Lei Q., Talley E.M., Lynch C., Bayliss D.A. The TASK-1 two-pore domain K+ channel is a molecular substrate for neuronal effects of inhalation anesthetics. J Neurosci. 2000;20:6347–6354. [PMC free article] [PubMed] [Google Scholar]
52. Yu X., Franks N.P., Wisden W. Sleep and sedative states induced by targeting the histamine and noradrenergic systems. Front Neural Circuits. 2018;12:4. [PMC free article] [PubMed] [Google Scholar]
53. Meuret P., Backman S.B., Bonhomme V., Plourde G., Fiset P. Physostigmine reverses propofol-induced unconsciousness and attenuation of the auditory steady state response and bispectral index in human volunteers. Anesthesiology. 2000;93:708–717. [PubMed] [Google Scholar]
54. Plourde G., Chartrand D., Fiset P., Font S., Backman S.B. Antagonism of sevoflurane anaesthesia by physostigmine: effects on the auditory steady-state response and bispectral index. Br J Anaesth. 2003;91:583–586. [PubMed] [Google Scholar]
55. Nelson L.E., Lu J., Guo T., Saper C.B., Franks N.P., Maze M. The alpha2-adrenoceptor agonist dexmedetomidine converges on an endogenous sleep-promoting pathway to exert its sedative effects. Anesthesiology. 2003;98:428–436. [PubMed] [Google Scholar]
56. Guldenmund P., Vanhaudenhuyse A., Sanders R.D., et al. Brain functional connectivity differentiates dexmedetomidine from propofol and natural sleep. Br J Anaesth. 2017;119:674–684. [PubMed] [Google Scholar]
57. Kokkinou M., Ashok A.H., Howes O.D. The effects of ketamine on dopaminergic function: meta-analysis and review of the implications for neuropsychiatric disorders. Mol Psychiatry. 2018;23:59–69. [PMC free article] [PubMed] [Google Scholar]
58. Liu X., Lauer K.K., Ward B.D., Li S.J., Hudetz A.G. Differential effects of deep sedation with propofol on the specific and nonspecific thalamocortical systems: a functional magnetic resonance imaging study. Anesthesiology. 2013;118:59–69. [PMC free article] [PubMed] [Google Scholar]
59. Tasserie J., Uhrig L., Sitt J.D., et al. Deep brain stimulation of the thalamus restores signatures of consciousness in a nonhuman primate model. Sci Adv. 2022;8:eabl5547. [PMC free article] [PubMed] [Google Scholar]
60. Bastos A.M., Donoghue J.A., Brincat S.L., et al. Neural effects of propofol-induced unconsciousness and its reversal using thalamic stimulation. eLife. 2021;10 [PMC free article] [PubMed] [Google Scholar]
61. Afrasiabi M., Redinbaugh M.J., Phillips J.M., et al. Consciousness depends on integration between parietal cortex, striatum, and thalamus. Cell Syst. 2021;12:363–373.e11. [PMC free article] [PubMed] [Google Scholar]
62. Meuret P., Backman S.B.S.B., Bonhomme V., Plourde G., Fiset P. Physostigmine reverses propofol-induced unconsciousness and attenuation of the auditory steady state response and bispectral index in human volunteers. Anesthesiology. 2000;93:708–717. [PubMed] [Google Scholar]
63. Luppi A.I., Craig M.M., Pappas I., et al. Consciousness-specific dynamic interactions of brain integration and functional diversity. Nat Commun. 2019;10:4616. [PMC free article] [PubMed] [Google Scholar]
64. Purdon P.L., Sampson A., Pavone K.J., Brown E.N. Clinical electroencephalography for anesthesiologists. part i: background and basic signatures. Anesthesiology. 2015;123:937–960. [PMC free article] [PubMed] [Google Scholar]
65. Kraguljac N.V., Frölich M.A., Tran S., et al. Ketamine modulates hippocampal neurochemistry and functional connectivity: a combined magnetic resonance spectroscopy and resting-state fMRI study in healthy volunteers. Mol Psychiatry. 2017;22:562–569. [PMC free article] [PubMed] [Google Scholar]
66. Höflich A., Hahn A., Küblböck M., et al. Ketamine-dependent neuronal activation in healthy volunteers. Brain Struct Funct. 2017;222:1533–1542. [PubMed] [Google Scholar]
67. Cichon J., Wasilczuk A.Z., Looger L.L., Contreras D., Kelz M.B., Proekt A. Ketamine triggers a switch in excitatory neuronal activity across neocortex. Nat Neurosci. 2023;26:39–52. [PMC free article] [PubMed] [Google Scholar]
68. Sleigh J., Harvey M., Voss L., Denny B. Ketamine - more mechanisms of action than just NMDA blockade. Trends Anaesth Crit Care. 2014;4:76–81. [Google Scholar]
69. Smith S.M., Fox P.T., Miller K.L., et al. Correspondence of the brain’s functional architecture during activation and rest. Proc Natl Acad Sci U S A. 2009;106:13040–13045. [PMC free article] [PubMed] [Google Scholar]
70. Sergent C., Corazzol M., Labouret G., et al. Bifurcation in brain dynamics reveals a signature of conscious processing independent of report. Nat Commun. 2021;12:1149. [PMC free article] [PubMed] [Google Scholar]
71. Crone J.S., Lutkenhoff E.S., Bio B.J., Laureys S., Monti M.M. Testing proposed neuronal models of effective connectivity within the cortico-basal ganglia-thalamo-cortical loop during loss of consciousness. Cereb Cortex. 2017;27:2727–2738. [PubMed] [Google Scholar]
72. Casali A.G., Gosseries O., Rosanova M., et al. A theoretically based index of consciousness independent of sensory processing and behavior. Sci Transl Med. 2013;5:198ra105. [PubMed] [Google Scholar]
73. Lee M., Sanz L.R.D., Barra A., et al. Quantifying arousal and awareness in altered states of consciousness using interpretable deep learning. Nat Commun. 2022;13:1064. [PMC free article] [PubMed] [Google Scholar]
74. Kim H., Hudetz A.G., Lee J., et al. Estimating the integrated information measure phi from high-density electroencephalography during states of consciousness in humans. Front Hum Neurosci. 2018;12:42. [PMC free article] [PubMed] [Google Scholar]
75. Del Pozo S.M., Laufs H., Bonhomme V., Laureys S., Balenzuela P., Tagliazucchi E. Unconsciousness reconfigures modular brain network dynamics. Chaos. 2021;31 [PubMed] [Google Scholar]
76. Golkowski D., Larroque S.K., Vanhaudenhuyse A., et al. Changes in whole brain dynamics and connectivity patterns imaging. Anesthesiology. 2019;130:898–911. [PubMed] [Google Scholar]
77. Luppi A.I., Cabral J., Cofre R., Destexhe A., Deco G., Kringelbach M.L. Dynamical models to evaluate structure-function relationships in network neuroscience. Nat Rev Neurosci. 2022;23:767–768. [PubMed] [Google Scholar]
78. Lee U., Mashour G.A. Role of network science in the study of anesthetic state transitions. Anesthesiology. 2018;129:1029–1044. [PMC free article] [PubMed] [Google Scholar]
79. Blain-Moraes S., Tarnal V., Vanini G., et al. Network efficiency and posterior alpha patterns are markers of recovery from general anesthesia: a high-density electroencephalography study in healthy volunteers. Front Hum Neurosci. 2017;11:328. [PMC free article] [PubMed] [Google Scholar]
80. Lee H., Golkowski D., Jordan D., et al. Relationship of critical dynamics, functional connectivity, and states of consciousness in large-scale human brain networks. Neuroimage. 2019;188:228–238. [PubMed] [Google Scholar]
81. Demertzi A., Tagliazucchi E., Dehaene S., et al. Human consciousness is supported by dynamic complex patterns of brain signal coordination. Sci Adv. 2019;5:eaat7603. [PMC free article] [PubMed] [Google Scholar]
82. Mashour G.A., Hudetz A.G. Neural correlates of unconsciousness in large-scale brain networks. Trends Neurosci. 2018;41:150–160. [PMC free article] [PubMed] [Google Scholar]
83. Hanson J.R., Walker S.I. Formalizing falsification for theories of consciousness across computational hierarchies. Neurosci Conscious. 2021;2021 niab014. [PMC free article] [PubMed] [Google Scholar]
84. Dehaene S., Sergent C., Changeux J.P. A neuronal network model linking subjective reports and objective physiological data during conscious perception. Proc Natl Acad Sci U S A. 2003;100:8520–8525. [PMC free article] [PubMed] [Google Scholar]
85. Bekinschtein T.A., Dehaene S., Rohaut B., Tadel F., Cohen L., Naccache L. Neural signature of the conscious processing of auditory regularities. Proc Natl Acad Sci U S A. 2009;106:1672–1677. [PMC free article] [PubMed] [Google Scholar]
86. Noel J.-P., Ishizawa Y., Patel S.R., Eskandar E.N., Wallace M.T. Leveraging nonhuman primate multisensory neurons and circuits in assessing consciousness theory. J Neurosci. 2019;39:7485–7500. [PMC free article] [PubMed] [Google Scholar]
87. Bonhomme V., Vanhaudenhuyse A., Demertzi A., et al. Resting-state network-specific breakdown of functional connectivity during ketamine alteration of consciousness in volunteers. Anesthesiology. 2016;125:873–888. [PubMed] [Google Scholar]
88. Palanca B.J.A., Mitra A., Larson-Prior L., Snyder A.Z., Avidan M.S., Raichle M.E. Resting state functional magnetic resonance imaging correlates of sevoflurane-induced unconsciousness. Anesthesiology. 2015;123:346–356. [PMC free article] [PubMed] [Google Scholar]
89. Boveroux P., Vanhaudenhuyse A., Bruno M.-A., et al. Breakdown of within- and between-network resting state functional magnetic resonance imaging connectivity during propofol-induced loss of consciousness. Anesthesiology. 2010;113:1038–1053. [PubMed] [Google Scholar]
90. Ferrarelli F., Massimini M., Sarasso S., et al. Breakdown in cortical effective connectivity during midazolam-induced loss of consciousness. Proc Natl Acad Sci U S A. 2010;107:2681–2686. [PMC free article] [PubMed] [Google Scholar]
91. Boly M., Moran R., Murphy M., et al. Connectivity changes underlying spectral EEG changes during propofol-induced loss of consciousness. J Neurosci. 2012;32:7082–7090. [PMC free article] [PubMed] [Google Scholar]
92. Peltier S.J., Kerssens C.A.C., Hamann S.B., Sebel P.S., Byas-Smith M., Hu X.P. Functional connectivity changes with concentration of sevoflurane anesthesia. Neuroreport. 2005;16:285–288. [PubMed] [Google Scholar]
93. Lee U., Kim S., Noh G.J., Choi B.M., Hwang E., Mashour G.A. The directionality and functional organization of frontoparietal connectivity during consciousness and anesthesia in humans. Conscious Cogn. 2009;18:1069–1078. [PubMed] [Google Scholar]
94. Ku S.-W.W., Lee U., Noh G.-J.J., Jun I.-G.G., Mashour G.A. Preferential inhibition of frontal-to-parietal feedback connectivity is a neurophysiologic correlate of general anesthesia in surgical patients. PLoS One. 2011;6 [PMC free article] [PubMed] [Google Scholar]
95. Jordan D., Ilg R., Riedl V., et al. Simultaneous electroencephalographic and functional magnetic resonance imaging indicate impaired cortical top-down processing in association with anesthetic-induced unconsciousness. Anesthesiology. 2013;119:1031–1042. [PubMed] [Google Scholar]
96. Ranft A., Golkowski D., Kiel T., et al. Neural correlates of sevoflurane-induced unconsciousness identified by simultaneous functional magnetic resonance imaging and electroencephalography. Anesthesiology. 2016;125:861–872. [PMC free article] [PubMed] [Google Scholar]
97. Warnaby C.E., Seretny M., Mhuircheartaigh R.N., et al. Anesthesia-induced suppression of human dorsal anterior insula responsivity at loss of volitional behavioral response. Anesthesiology. 2016;124:766–778. [PubMed] [Google Scholar]
98. Barrett A.B., Murphy M., Bruno M.A., et al. Granger causality analysis of steady-state electroencephalographic signals during propofol-induced anaesthesia. PLoS One. 2012;7 [PMC free article] [PubMed] [Google Scholar]
99. Maksimow A., Silfverhuth M., Långsjö J., et al. Directional connectivity between frontal and posterior brain regions is altered with increasing concentrations of propofol. PLoS One. 2014;9 [PMC free article] [PubMed] [Google Scholar]
100. Nicolaou N., Hourris S., Alexandrou P., Georgiou J. EEG-based automatic classification of ‘awake’ versus ‘anesthetized’ state in general anesthesia using granger causality. PLoS One. 2012;7 [PMC free article] [PubMed] [Google Scholar]
101. Nicolaou N., Georgiou J. Neural network-based classification of anesthesia/awareness using granger causality features. Clin EEG Neurosci. 2014;45:77–88. [PubMed] [Google Scholar]
102. Sanders R.D., Banks M.I., Darracq M., et al. Propofol-induced unresponsiveness is associated with impaired feedforward connectivity in cortical hierarchy. Br J Anaesth. 2018;121:1084–1096. [PMC free article] [PubMed] [Google Scholar]
103. Casey C.P., Tanabe S., Farahbakhsh Z., et al. Dynamic causal modelling of auditory surprise during disconnected consciousness: the role of feedback connectivity. Neuroimage. 2022;263 [PMC free article] [PubMed] [Google Scholar]
104. Li D., Vlisides P.E., Kelz M.B., et al. Dynamic cortical connectivity during general anesthesia in healthy volunteers. Anesthesiology. 2019;130:870–884. [PubMed] [Google Scholar]
105. Vlisides P.E., Li D., Zierau M., et al. Dynamic cortical connectivity during general anesthesia in surgical patients. Anesthesiology. 2019;130:885–897. [PMC free article] [PubMed] [Google Scholar]
106. Tagliazucchi E., Chialvo D.R.D.R., Siniatchkin M., et al. Large-scale signatures of unconsciousness are consistent with a departure from critical dynamics. J R Soc Interface. 2016;13 [PMC free article] [PubMed] [Google Scholar]
107. Barttfeld P., Uhrig L., Sitt J.D., Sigman M., Jarraya B., Dehaene S. Signature of consciousness in the dynamics of resting-state brain activity. Proc Natl Acad Sci U S A. 2015;112:887–892. [PMC free article] [PubMed] [Google Scholar]
108. Tononi G., Boly M., Massimini M., Koch C. Integrated information theory: from consciousness to its physical substrate. Nat Rev Neurosci. 2016;17:450–461. [PubMed] [Google Scholar]
109. Sarasso S., Casali A.G., Casarotto S., Rosanova M., Sinigaglia C., Massimini M. Consciousness and complexity: a consilience of evidence. Neurosci Conscious. 2021;7:1–24. [Google Scholar]
110. Mediano P.A.M., Rosas F.E., Bor D., Seth A.K., Barrett A.B. The strength of weak integrated information theory. Trends Cogn Sci. 2022;26:646–655. [PubMed] [Google Scholar]
111. Comolatti R., Pigorini A., Casarotto S., et al. A fast and general method to empirically estimate the complexity of brain responses to transcranial and intracranial stimulations. Brain Stimul. 2019;12:1280–1289. [PubMed] [Google Scholar]
112. Casarotto S., Comanducci A., Rosanova M., et al. Stratification of unresponsive patients by an independently validated index of brain complexity. Ann Neurol. 2016;80:718–729. [PMC free article] [PubMed] [Google Scholar]
113. Lee H., Noh G.J., Joo P., et al. Diversity of functional connectivity patterns is reduced in propofol-induced unconsciousness. Hum Brain Mapp. 2017;38:4980–4995. [PMC free article] [PubMed] [Google Scholar]
114. Bharath R.D., Panda R., Saini J., Sriganesh K., Rao G.S.U. Dynamic local connectivity uncovers altered brain synchrony during propofol sedation. Sci Rep. 2017;7:8501. [PMC free article] [PubMed] [Google Scholar]
115. Ihalainen R., Gosseries O., de Steen F Van, et al. How hot is the hot zone? Computational modelling clarifies the role of parietal and frontoparietal connectivity during anaesthetic-induced loss of consciousness. Neuroimage. 2021;231 [PubMed] [Google Scholar]
116. Casey C.P., Tanabe S., Farahbakhsh Z., et al. Distinct EEG signatures differentiate unconsciousness and disconnection during anaesthesia and sleep. Br J Anaesth. 2022;128:1006–1018. [PMC free article] [PubMed] [Google Scholar]
117. Siclari F., Baird B., Perogamvros L., et al. The neural correlates of dreaming. Nat Neurosci. 2017;20:872–878. [PMC free article] [PubMed] [Google Scholar]
118. Siclari F., Bernardi G., Cataldi J., Tononi G. Dreaming in NREM sleep: a high-density EEG study of slow waves and spindles. J Neurosci. 2018;38:9175–9185. [PMC free article] [PubMed] [Google Scholar]
119. Lamme V.A.F. Towards a true neural stance on consciousness. Trends Cogn Sci. 2006;10:494–501. [PubMed] [Google Scholar]
120. Sanders R.D., Casey C., Saalmann Y.B. Predictive coding as a model of sensory disconnection: relevance to anaesthetic mechanisms. Br J Anaesth. 2021;126:37–40. [PubMed] [Google Scholar]
121. Pinto Y., Vandenbroucke A.R., Otten M., Sligte I.G., Seth A.K., Lamme V.A.F. Conscious visual memory with minimal attention. J Exp Psychol Gen. 2017;146:214–226. [PubMed] [Google Scholar]
122. van Gaal S., Lamme V.A.F. Unconscious high-level information processing: implication for neurobiological theories of consciousness. Neuroscientist. 2012;18:287–301. [PubMed] [Google Scholar]
123. Alkire M.T., Hudetz A.G., Tononi G. Consciusness and anesthesia. Science. 2008;7:876–880. [PMC free article] [PubMed] [Google Scholar]
124. Lamme V.A., Zipser K., Spekreijse H. Figure-ground activity in primary visual cortex is suppressed by anesthesia. Proc Natl Acad Sci U S A. 1998;95:3263–3268. [PMC free article] [PubMed] [Google Scholar]
125. Kilner J.M., Friston K.J., Frith C.D. Predictive coding: an account of the mirror neuron system. Cogn Process. 2007;8:159–166. [PMC free article] [PubMed] [Google Scholar]
126. Seth A.K. Interoceptive inference, emotion, and the embodied self. Trends Cogn Sci. 2013;17:565–573. [PubMed] [Google Scholar]
127. Carhart-Harris R.L., Friston K.J. REBUS and the anarchic brain: toward a unified model of the brain action of psychedelics. Pharmacol Rev. 2019;71:316–344. [PMC free article] [PubMed] [Google Scholar]
128. Brown R., Lau H., LeDoux J.E. Understanding the higher-order approach to consciousness. Trends Cogn Sci. 2019;23:754–768. [PubMed] [Google Scholar]
129. LeDoux J.E., Brown R. A higher-order theory of emotional consciousness. Proc Natl Acad Sci U S A. 2017;114:E2016–E2025. [PMC free article] [PubMed] [Google Scholar]
130. Melloni L., Mudrik L., Pitts M., et al. An adversarial collaboration protocol for testing contrasting predictions of global neuronal workspace and integrated information theory. PLoS One. 2023;18 [PMC free article] [PubMed] [Google Scholar]
131. Melloni L., Mudrik L., Pitts M., Koch C. Making the hard problem of consciousness easier. Science. 2021;372:911–912. [PubMed] [Google Scholar]
132. Consortium C., Ferrante O., Gorska-Klimowska U., et al. An adversarial collaboration to critically evaluate theories of consciousness. bioRxiv [Internet] Cold Spring Harbor Laboratory; 2023. https://www.biorxiv.org/content/early/2023/06/26/2023.06.23.546249 Available from: [Google Scholar]
133. Amico E., Marinazzo D., Di Perri C., et al. Mapping the functional connectome traits of levels of consciousness. Neuroimage. 2017;148:201–211. [PubMed] [Google Scholar]
134. Liu X., Li S.-J.J., Hudetz A.G. Increased precuneus connectivity during propofol sedation. Neurosci Lett. 2014;561:18–23. [PMC free article] [PubMed] [Google Scholar]
135. Liu X., Lauer K.K.K., Ward B.D., et al. Regional entropy of functional imaging signals varies differently in sensory and cognitive systems during propofol-modulated loss and return of behavioral responsiveness. Brain Imaging Behav. 2019;13:514–525. [PMC free article] [PubMed] [Google Scholar]
136. Scheinin A., Kantonen O., Alkire M., et al. Foundations of human consciousness: imaging the twilight zone. J Neurosci. 2021;41:1769–1778. [PMC free article] [PubMed] [Google Scholar]
137. Warnaby C.E., Sleigh J.W., Hight D., Jbabdi S., Tracey I. Investigation of slow-wave activity saturation during surgical anesthesia reveals a signature of neural inertia in humans. Anesthesiology. 2017;127:645–657. [PMC free article] [PubMed] [Google Scholar]
138. Gaskell A.L.L., Hight D.F.F., Winders J., et al. Frontal alpha-delta EEG does not preclude volitional response during anaesthesia: prospective cohort study of the isolated forearm technique. Br J Anaesth. 2017;119:664–673. [PubMed] [Google Scholar]
139. Darracq M., Funk C., Polyakov D., et al. Evoked alpha power is reduced in disconnected consciousness during sleep and anesthesia. Sci Rep. 2018;8:16664. [PMC free article] [PubMed] [Google Scholar]
140. Sanz Perl Y., Pallavicini C., Pérez Ipiña I., et al. Perturbations in dynamical models of whole-brain activity dissociate between the level and stability of consciousness. PLoS Comput Biol. 2021;17 [PMC free article] [PubMed] [Google Scholar]
141. Graziano M.S.A. A conceptual framework for consciousness. Proc Natl Acad Sci U S A. 2022;119 [PMC free article] [PubMed] [Google Scholar]
142. Deng F., Taylor N., Owen A.M., Cusack R., Naci L. Responsiveness variability during anaesthesia relates to inherent differences in brain structure and function of the frontoparietal networks. Hum Brain Mapp. 2023;44:2142–2157. [PMC free article] [PubMed] [Google Scholar]
143. Baron Shahaf D., Weissman A., Priven L., Shahaf G. Identifying recall under sedation by a novel EEG based index of attention-a pilot study. Front Med. 2022;9 [PMC free article] [PubMed] [Google Scholar]
Articles from BJA Open are provided here courtesy of Elsevier
OTHER FORMATS
PDF (2.1M)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 1088687 | https://sv.wikipedia.org/wiki/Kognitiv%20neurovetenskap | Kognitiv neurovetenskap | Kognitiv neurovetenskap är läran om hur hjärnan möjliggör psykologiska fenomen eller mentala processer, det vill säga perception, tankar, beteenden och känslor. Den teoretiska grunden är kognitiv (se även kognitiv psykologi), medan den metodologiska grunden bygger på psykofysiologi, neuropsykologi, elektrofysiologi, hjärnavbildning och i viss utsträckning djurförsök. Metoder inom kognitionsvetenskap inkluderar elektroencefalografi (EEG), magnetoencefalografi (MEG), elektromyografi (EMG), funktionell magnetisk resonanstomografi (eng. functional magnetic resonance imaging, fMRI), diffusionsbaserad magnetisk resonanstomografi (eng. diffusion tensor imaging, DTI), transkranial magnetisk stimulering (TMS) och neuropsykologiska observationer. Drivkraften bakom samtida kognitiv neurovetenskap är i stor utsträckning tack vare teknologiska framsteg inom hjärnavbildning.
Se även
Kognition
Kognitiv neuropsykologi
Kognitiv psykologi
Referenser
Delområden inom psykologi
Neurovetenskap | swedish | 0.535577 |
shut_down_conciousness/PMC3461283.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List Emerg Med Int v.2012; 2012 PMC3461283
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of emi
Emerg Med Int. 2012; 2012: 637171.
Published online 2012 Sep 20. doi: 10.1155/2012/637171
PMCID: PMC3461283
PMID: 23050154
Sedation in Traumatic Brain Injury
Oliver Flower 1 , 2 and Simon Hellings 2 ,*
Author information Article notes Copyright and License information PMC Disclaimer
Go to:
Abstract
Several different classes of sedative agents are used in the management of patients with traumatic brain injury (TBI). These agents are used at induction of anaesthesia, to maintain sedation, to reduce elevated intracranial pressure, to terminate seizure activity and facilitate ventilation. The intent of their use is to prevent secondary brain injury by facilitating and optimising ventilation, reducing cerebral metabolic rate and reducing intracranial pressure. There is limited evidence available as to the best choice of sedative agents in TBI, with each agent having specific advantages and disadvantages. This review discusses these agents and offers evidence-based guidance as to the appropriate context in which each agent may be used. Propofol, benzodiazepines, narcotics, barbiturates, etomidate, ketamine, and dexmedetomidine are reviewed and compared.
Go to:
1. Introduction
Several different classes of drugs are used as sedatives in patients with traumatic brain injury (TBI). Several of these agents may also have other uses, for example as anticonvulsants or analgesics. Whilst none are perfect, they all have potential roles in managing a condition that is a major cause of disability, death, and economic cost to society. This paper discusses and compares these agents and offers evidence-based guidance as to the appropriate context in which each agent may be used.
It is important to delineate the contexts in which sedative agents are used in the setting of TBI and what is considered a sedative. For the purposes of this paper, sedative agents are considered to be drugs that decrease consciousness and have therapeutic applications in the management of TBI. After primary brain injury, airway protection and control of ventilation are often required. Induction sedative agents (distinct from muscle relaxants) are used to safely facilitate endotracheal intubation in a manner that minimises haemodynamic instability and secondary brain injury. Maintenance of sedation is then employed as part of the overall management of TBI to permit manipulation of ventilation, optimisation of cerebral metabolic rate (CMRO2), cerebral blood flow (CBF), and intracranial pressure (ICP). See Table 1 for abbreviations with explanations. In TBI confounded by alcohol or illicit drug intoxication, sedative agents facilitate safe management whilst these confounding drugs wear off. For refractory, elevated ICP in severe TBI, sedative agents play a key role in the escalating tiers of therapy to reduce ICP. Sedative hypnotics are also employed in seizure control for refractory acute posttraumatic epilepsy. As with all ventilated patients, sedatives act as anxiolytics whilst patients are mechanically ventilated [1].
Table 1
Abbreviations and explanations
(i) AMPA: α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid, GABA: γ-Aminobutyric acid
(ii) EEG: Electroencephalogram
(iii) CMRO2: Cerebral Metabolic Rate of Oxygen
(iv) CPP: Cerebral Perfusion Pressure
(v) ICP: Intracranial Pressure
(vi) IV: Intravenous
(vii) MAP: Mean Arterial Pressure
(viii) t 1/2: Half-life
(ix) Context sensitive t 1/2: the time taken for blood plasma concentration of a drug to decline by one half after an infusion designed to maintain a steady state (i.e., a constant plasma concentration) has been stopped. The “context” is the duration of infusion [5].
Open in a separate window
The primary injury of TBI causes diffuse axonal injury, cerebral oedema, intracranial haematoma, elevated ICP, reduced cerebral perfusion pressure (CPP), and cerebral ischaemia. Therapeutic efforts focus on reducing the secondary insults of hypoxia, hypercapnea, systemic hypotension and intracranial hypertension. Sedatives address these issues in several ways. They allow optimisation of ventilation to prevent hypoxia and achieve normocapnea (and hypocapnea for brief episodes of elevated ICP); they reduce CMRO2 and therefore CBF and cerebral blood volume (CBV) and reduce ICP. However they may reduce systemic blood pressure, thereby reducing CPP, and have other adverse effects. Even a single episode of hypotension is a powerful predictor of outcome following TBI [2, 3].
There is limited evidence available to guide the choice of specific sedative agents in TBI. A recent systematic review examining a range of outcomes in TBI concluded that there was no convincing evidence that any one of the sedative agent was superior to another [4]. A number of these studies included patients with less severe traumatic brain injuries and spanned several decades, further limiting conclusions that can be made. Multiple sedative agents are often used synchronously, clouding assessment of individual agents. The guidelines from the Brain Trauma Foundation also highlight a lack of high quality evidence to recommend one sedative agent over another, with the exception of barbiturate use for refractory elevated ICP. Despite this, each agent has a potential role in TBI and clinicians must consider the advantages and disadvantages when deciding what to use in each context.
Go to:
2. Propofol
See Table 2. Propofol is a phenol derivative with high lipid solubility and a rapid onset of action. It has a very low solubility in water so is formulated as an emulsion in soya bean oil, glycerol, and egg phosphatide. A relatively rapid plasma clearance ensures a reliable recovery of consciousness even after prolonged administration, thereby facilitating neurological examination. However, the context-sensitive half time does increase with prolonged infusions, though to a much lesser extent than seen with many other sedatives.
Table 2
Propofol
Group Phenol Derivative
Mechanism of Action/Pharmakodynamics Potentiation GABAA receptors
Na+ channel blocker
Neuroprotective effects Reduces CBF, CMRO2 and ICP
Reduces MAP, therefore variable effect on CPP
Increases seizure threshold
Pharmacokinetics Rapid hepatic metabolised, with extra-hepatic metabolism
t 1/2 2–24 hours, but rapid peripheral distribution
Short context sensitive t 1/2
Advantages Favourable effects on CBF, CMRO2 and ICP
Rapid onset of action
Relatively short context sensitive t 1/2 facilitating neurological assessment
Disadvantages and major side effects Hypotension may worsen CPP
High lipid load
Associated with elevated liver enzymes & pancreatitis
Potential for PRIS, particularly with prolonged, high dose infusions
Formulation may support bacterial and fungal growth
Contraindicated if allergic to egg or soybeans
Dosage Induction: 1–2.5 mg/kg, 0.5–1.5 mg/kg in elderly or limited cardiovascular reserve
Maintenance of sedation: 1.5–4.5 mg/kg/hour, titrated to desired effect
Other significant facts Increased risk of PRIS at infusions >4 mg/kg/h for >48 h
Appropriate roles in TBI Induction agent, caution in hypotension
Continuous infusion to provide sedation in TBI
Refractory elevated ICP
Refractory seizures
Open in a separate window
Since its introduction in 1986, propofol has increasingly been used both as an induction agent and as a maintenance sedative in the neurointensive care unit. Several studies have demonstrated the favourable cerebral effects of propofol. ICP, CBF, and CMRO2, have all been shown to be reduced with propofol [8, 9]. However, a fall in mean arterial blood pressure (MAP) may reduce the CPP if this is not mitigated with adequate fluid resuscitation and vasopressors. When comparing propofol sedation with midazolam in medical and surgical ICU patients, propofol has been associated with improved quality of sedation and a faster recovery of consciousness on discontinuation of sedation [10].
There is increasing awareness in the literature of the central role of mitochondrial dysfunction and cerebral cell death in areas of the brain with high oxidative stress [11, 12]. Propofol may act as a neuroprotective agent through limitation of oxidative stress. An RCT employing cerebral microdialysis catheters compared sedation with midazolam and propofol, using several cerebral biomarkers as endpoints in the acute phase of TBI [13]. No difference was found between the two groups over a 72-hour period in the lactate to pyruvate ratio, a marker of cerebral oxidative stress. This was a relatively small study and the concentrations of propofol used may not have been sufficient to produce an antioxidant effect nevertheless this is an interesting and novel area of future research.
Aside from a reduction in MAP and the need for increased vasopressor requirements to preserve CPP, the lipid formulation of propofol may be associated with other adverse effects. Propofol infusion syndrome (PRIS) was initially described in case studies of children who were sedated with propofol infusions. Subsequently it has been reported in adults, both with long-term infusions in ICU patients and in the short term when used as a general anaesthetic. Clinically patients may present with a variety of findings including lactic acidosis, cardiac dysfunction, and Brugada-like electrocardiogram changes (see Figure 1), which may herald imminent malignant arrhythmias [14]. This can progress to rhabdomyolysis, renal failure, and cardiovascular collapse. The pathophysiology of PRIS is incompletely understood and involves multiple different pathways. An underlying imbalance between energy utilization and demand at the mitochondrial level and effects on lipid metabolism are postulated mechanisms.
An external file that holds a picture, illustration, etc.
Object name is EMI2012-637171.001.jpg
Figure 1
Brugada-like ECG changes that may be seen in propofol infusion syndrome. Coved ST elevation, at least 2 mm J point elevation and descending ST segment followed by a negative T wave (see [67]).
Importantly, it is thought that PRIS is more common in patients with TBI. In one retrospective cohort study of adult neurosurgical patients in ICU, 7 of 67 patients displayed signs of PRIS and died. There was an increased incidence of PRIS with higher doses [15]. PRIS may be more common in TBI because large doses of propofol can be used to control elevated ICP [16]. It has been argued that PRIS may limit the usefulness of propofol as a sedative agent in traumatic brain injury, particularly when used in higher doses.
Other potential complications associated with the use of propofol include an elevation in pancreatic enzymes and pancreatitis [17]. Concerns have also been raised that propofol offers a good medium for microbial growth [18], although this may be less significant with newer formulations. Propofol has a significant calorific content, and this should be taken into account when performing nutritional assessments.
Initial reports suggested that propofol may increase seizure activity in susceptible patients [19]. The extent to which this activity represented disordered muscle tone or true seizure activity is unclear [20]. Conversely, propofol has also been demonstrated to increase seizure threshold and has been successfully used in the treatment of status epilepticus. Much of the evidence for the use of propofol in refractory status epilepticus is derived from case series that demonstrated cessation of seizure activity with infusions of propofol [21]. Propofol has been demonstrated to achieve and maintain burst suppression, although at the expense of significant decreases in mean arterial pressure and cardiac index [22].
Therefore, propofol is indicated as a sedative agent in TBI. It has the advantage of a relatively quick onset and offset of action facilitating neurological assessment. Clinicians should be mindful of the risk of PRIS, particularly when using >4 mg/kg/hour for >48 hours [23]. As an induction agent it may cause a fall in MAP and thus CPP, and this should be mitigated through the judicious use of vasopressors and fluid boluses. Propofol may be indicated in the treatment of refractory status epilepticus. Its use as an agent to achieve burst suppression may come at the expense of worsening haemodynamics.
Go to:
3. Benzodiazepines
See Table 3. Benzodiazepines are commonly used as sedative agents in patients with TBI. They are nonselective CNS depressants that augment the action of GABA at GABAA receptors, causing increased conductance of chloride ions. They have anxiolytic, amnesic, and anticonvulsant properties. Prior to the advent of propofol, midazolam was the most frequently used sedative in TBI in the UK [24], with lorazepam frequently being used in the US [25]. Midazolam offers the most benefits of the benzodiazepines for sedation in TBI, due to its shorter context sensitive t 1/2 (2–2.5 hours) and faster onset and offset of action, compared to lorazepam (t 1/2 10–20 h) or diazepam (t 1/2 20–40 hours) [26]. It has a rapid onset as a result of high lipid solubility at physiological pH due to the closure of the imidazole ring. Its rapid hepatic metabolism accounts for its rapid offset of action [27] however some metabolites are active and accumulate with prolonged infusions. This may result in continued sedation even after drug cessation, particularly in the elderly or with liver impairment.
Table 3
Midazolam
Group Imadobenzodiazepine
Mechanism of Action/Pharmakodynamics GABAA receptor agonist
Chloride channel activation,
Kappa opioid agonist
Neuroprotective effects Reduces CBF, CMRO2 and ICP but minimal effect beyond that of sedation
Reduces MAP, variable effect on CPP
Raises seizure threshold
Pharmacokinetics Onset of action 2–4 minutes
94% protein bound
Highly lipid soluble
Hepatic metabolism
Renal excretion (some bile)
Short context sensitive t 1/2 (2.4 h)
Advantages Shorter t 1/2 than other benzodiazepines
Causes less hypotension than barbiturates or propofol
Disadvantages and major side effects Metabolites accumulate delaying neurological assessment post cessation of infusion
Boluses in TBI reduce MAP (and CPP)
Withdrawal syndrome
Delirium
Respiratory and cough reflex suppression
Tachyphylaxis after 72 hours
Plateau effect on reducing ICP, where increasing doses have no effect
Dosage Induction: 0.1 mg/kg
Maintenance of sedation: 0.01–0.2 mg/kg/hour
Other significant facts Interaction with peripheral benzodiazepine leucocyte receptors so may have immunosuppressant effect
Appropriate roles in TBI Induction of anaesthesia
Maintenance of sedation in hypotensive patients with TBI
Maintenance of sedation when imminent neurological assessment not required
Treatment of seizures
Open in a separate window
Whilst benzodiazepines reduce CBF, CMRO2, and ICP and increase seizure threshold, there is evidence that bolus doses significantly reduce MAP and CPP in severe TBI [28]. The depth of CMRO2 reduction possible with benzodiazepines is not as profound as barbiturates or etomidate, and burst suppression cannot be achieved [29].
Other disadvantages include significant respiratory depression and inhibition of the cough reflex, limiting its use in non-intubated patients. After prolonged sedation with benzodiazepines, tolerance develops, and on cessation, withdrawal symptoms including tremors, seizures, hypertension, and insomnia may occur, requiring ongoing longer acting benzodiazepines to be prescribed [30]. Benzodiazepines are a risk factor for ICU delirium [31], which is independently associated with poor outcomes [32].
There have been several studies comparing the safety and efficacy of benzodiazepines with other commonly used agents. In one RCT, 63 trauma patients, the majority with severe TBI, were randomised to receive either midazolam or 2% propofol infusions. Patients in both groups received morphine for analgesia. No significant difference in ICP or in wake-up time was demonstrated between the two groups. Similarly no significant differences were seen in haemodynamic variables between the two groups. Interestingly, there was a higher incidence of therapeutic failure in the propofol group either because of inadequate sedation or hypertriglyceridemia [33]. Other smaller, underpowered studies have also failed to demonstrate a difference in outcomes between these two agents [34].
Therefore benzodiazepines have a role in the sedation of patients where imminent neurological assessment is not required. They have significant disadvantages including an accumulation of metabolites, increasing tolerance with prolonged infusions, and an increased likelihood of delirium.
Go to:
4. Narcotics
See Table 4. Opioid narcotics primarily have analgesic properties, and their sedative action may even be considered a side effect. However, various opioids are used in the sedation of patients with TBI, usually in combination with hypnotic agents to ensure analgesia and reduce hypnotic dose requirements. Analgesia-based protocols are feasible, with certain advantages over hypnotic (propofol and midazolam) sedative regimens [35]. Intravenous opioids used include morphine, fentanyl, sufentanil, and more recently remifentanil.
Table 4
Morphine Fentanyl Alfentanil Sufentanil Remifentanil
Pharmacodynamics μ 1, μ 2, κ and Δ agonists
Elimination t 1/2 (h) 3 3.7 1.5 2.2 0.25
Distribution t 1/2 3–11 min 10–30 min 15 min 5 min 1 min
Neuroprotective effects May increase ICP Minimal effect beyond the analgesic effect on CBF and CMRO2
Pharmacokinetics
Onset 6 min
Peak effect 20 min (IV)
30% protein bound
Hepatically metabolised to active metabolites
Renal clearance 95% protein bound
High lipid solubility
75% first pass pulmonary uptake
Hepatically metabolised to active metabolites
Renal clearance Onset
Peak 90 s
Duration 5–10 min
90% protein bound
Hepatically metabolised
Renal clearance Hepatically metabolised
Renal clearance Peak 60 s
Small Vd
Rapid clearance
Rapid ester hydrolysis by plasma esterases to inactive metabolite
(Independent of renal & hepatic function)
Advantages Lower cost
Relative haemodynamic stability
Hypnotic agent sparing
Analgesic properties Lower cost
Relative haemodynamic stability
Hypnotic agent sparing
Analgesic properties Relative haemodynamic stability
Hypnotic agent sparing
Analgesic properties Relative haemodynamic stability
Hypnotic agent sparing
Analgesic properties Very rapid onset/offset
Less nausea
Relative haemodynamic stability
Hypnotic agent sparing
Analgesic properties
Disadvantages and major side effects Hypotension
Bradycardia
Respiratory depression
Cough reflex suppression
Seizures
Rigidity
Constipation
Spasm sphincter of Oddi
Nausea
Pruritis
Dosage 0.05–0.1 mg/kg/hr Induction: 1–3 mcg/kg
Maintenance: 0.5–2 mcg/kg/h Induction: 10–50 mcg/kg
Infusion: 0.5–1 mcg/kg/min Induction: 4 mcg/kg Bolus: 1 mcg/kg
Infusion: 0.0125–1 mcg/kg/min
Appropriate uses in TBI Long term analgesia
Palliation Co-Induction agent
Continuous infusion
Palliation Co-Induction agent Co-Induction agent Co-Induction agent
Continuous infusion
infusion
Open in a separate window
Opiates act on μ 1 receptors (supraspinal analgesia), μ 2 receptors (ventilatory depression, bradycardia, physical addiction), κ receptors (sedation, spinal analgesia), ε receptors (dysphoria, hallucinations, respiratory stimulation), and Δ receptors (analgesia, behavioural effects, and epileptogenic). The different opioids have variable effects on each receptor [26]. Opioids can produce hypotension by a number of mechanisms including a reduction in sympathetic tone and the stimulation of histamine release. This hypotension may be detrimental in patients with TBI in whom maintenance of cerebral perfusion pressure is vital.
Prior to the advent of newer agents morphine has been most commonly used as a narcotic in TBI. However, prolonged use of opioids such as morphine can lead to redistribution and accumulation, with potentially unpredictable delays in awakening. The t 1/2 of morphine is increased in renal failure, as a proportion of both the parent drug and an active metabolite, morphine-6-glucuronide, are excreted renally [36]. In addition, tachyphylaxis can lead to increasing dose requirements with subsequent withdrawal phenomena and a possible rebound increase in ICP on cessation.
Shorter acting opioids include fentanyl, alfentanil, sufentanil, and remifentanil. These are more lipid soluble than morphine and so have a faster onset of action [37]. Metabolism to inactive metabolites leads to less accumulation in renal failure. Nevertheless, with prolonged infusion shorter acting opioids can accumulate and impede neurological assessment. For example, with an increasing duration of fentanyl infusion, saturation of inactive tissue sites and a return of opioid from peripheral compartments mean that there is a prolonged context-sensitive half time relative to sufentanil.
Studies of the effects of opioids on ICP have been inconsistent. However, there is evidence that the administration of high bolus doses of opioids may have potentially deleterious effects in TBI, with some studies showing an increase in ICP and a fall in CPP. These effects occurred despite controlling PaCO2. Interestingly, in those studies that prevented hypotension, an increase in ICP was not seen. It is suggested that hypotension may increase ICP and decrease CPP through cerebral autoregulatory reflexes [9]. It is unclear to what extent opioids may induce seizure activity. Whilst there are numerous case reports of clinical seizure activity, it has been argued that many of these represent muscle rigidity associated with high doses of opioid rather than seizure activity per se [38].
There has been increased interest in remifentanil as an alternative opioid sedative in TBI. Remifentanil is a potent, synthetic opioid receptor agonist, which differs from other synthetic opioids in that it undergoes rapid hydrolysis by tissue and plasma esterases. This rapid metabolism and lack of accumulation facilitate faster waking and neurological assessment of patients with TBI [39]. An RCT on neuro-intensive care patients showed analgesia-based sedation with remifentanil offered faster and more predictable time to assessment of neurological function than a hypnotic-based technique (propofol or midazolam) [40]. Furthermore, remifentanil was well tolerated in patients with TBI, with a significantly shorter time to extubation in patients who had received remifentanil compared with patients who had received morphine [11].
Opioids have a role as an adjunct to other sedative agents, for example in combination with propofol. They may reduce sedative requirements of other agents and provide effective analgesia and anxiolysis. Prolonged infusions of opioids, particularly morphine, may accumulate and hinder neurological assessment. When opioids are administered as a bolus, there is a risk of increasing the ICP, particularly when the MAP is allowed to fall.
Go to:
5. Barbiturates
See Table 5. Barbiturates, particularly pentobarbital and thiopentone, have previously played a central role in the sedation of patients with TBI [41]. However, with the advent of newer agents with less disadvantages, thiopentone is largely confined to use as an induction agent, for the treatment of refractory elevated ICP and for status epilepticus. Barbiturates stimulate γ-aminobutyric acid (GABA) receptors and inhibit α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors in the CNS producing dose-dependent sedation and general anaesthesia.
Table 5
Thiopentone
Group Barbiturate
Mechanism of Action/Pharmacodynamics Stimulate GABA receptors
Inhibit AMPA receptors
Neuroprotective effects Reduces CBF, CMRO2 and ICP
Reduces MAP, therefore variable effect on CPP Raises seizure threshold
Pharmacokinetics [6] Hepatically metabolised
0.5% renal excretion unchanged
Elimination t 1/2 11.6 h
First to zero order kinetics if plasma high
Significant accumulation
Advantages Rapid onset of action as induction agent
Favourable effects on CBF, CMRO2 and ICP
Inexpensive
Disadvantages and major side effects Accumulation with prolonged infusion
Hypotension
Gastroparesis
Loss of thermoregulation
Immunosuppression
Hypokalaemia during infusion
Hyperkalaemia on emergence
Life threatening arrhythmias on coma emergence
Dosage Induction of anaesthesia: 2–5 mg/kg
EEG burst suppression: 40 mg/kg followed by infusion at 4–8 mg/kg/h, titrated to EEG
Other significant facts May precipitate if given concurrently with IV muscle relaxants [7]
Appropriate uses in TBI Induction of anaesthesia, with caution regarding hypotension
Refractory elevated ICP
Refractory status epilepticus
Open in a separate window
High lipid solubility allows rapid transfer across the blood-brain barrier and exceptionally fast onset of action. The induction of anaesthesia sufficient for intubation within one arm-brain circulation time initially popularized the use of thiopentone as an induction agent in rapid-sequence intubation (RSI) [42]. The hypotensive effects caused by direct myocardial and central vasomotor depression should be anticipated and addressed by using only low doses and coadministering vasopressors such as metaraminol or phenylephrine if the blood pressure is suboptimal before RSI.
A recent Cochrane review concluded that barbiturates are not indicted as a maintenance sedative agent or for use prophylactically to prevent elevations in ICP [43], predominantly because the hypotension and other side effects offset any ICP lowering effect on CPP.
Significant accumulation will occur with repeated doses or infusions due to the long context-sensitive t 1/2 and the elimination kinetics changing from 1st to zero order at plasma levels >30 mg/L. To treat refractory elevated ICP or refractory status epilepticus, a clinical endpoint of burst suppression on EEG is targeted, which requires plasma levels >40 mg/L. Unfortunately, the high doses of thiopentone required to achieve this preclude neurological assessment for several days.
Therefore thiopentone may be used as an induction agent in TBI if hypotension is not already problematic and precautions are taken. It has a role in treatment of refractory elevated ICP and refractory status epilepticus, but not as a maintenance sedative in TBI.
Go to:
6. Etomidate
See Table 6. Etomidate is a carboxylated imidazole derivative predominantly used as an intravenous induction agent in the setting of haemodynamic instability. It causes less hypotension and cardiovascular depression than other sedatives in this context [44], with the exception of ketamine. Other advantages include a rapid onset of anaesthesia (10s) lasting 3–5 minutes following a dose of 0.3 mg/kg, and a short elimination t 1/2 of 2.6 h [45]. There is a reduction in CBF and ICP [46] and it can even achieve burst suppression on EEG [47].
Table 6
Etomidate
Group Caroboxylated imidazole derivative
Mechanism of Action/Pharmakodynamics GABAA receptor agonist
Neuroprotective effects Reduces CBF, CMRO2 and ICP
Maintains or increases CPP
Lowers seizure threshold
Pharmacokinetics 75% protein bound
Highly lipid soluble
High volume of distribution, three compartment model
Hepatic metabolism
Renal excretion (some bile)
Short context sensitive t 1/2 (4.8 h)
Advantages Rapid onset of action as induction agent
Only lasts 3–5 minutes after single bolus
Favourable effects on CBF, CMRO2 and ICP
Disadvantages and major side effects Adrenal suppression
Metabolic acidosis from propylene glycol vehicle
Pain on injection
Myoclonic movements
Nausea and vomiting
Dosage Induction: 0.2–0.4 mg/kg
Other significant facts Originally developed as an anti-fungal agent
Appropriate uses in TBI Induction of anaesthesia, with caution regarding adrenal suppression
Open in a separate window
However, the safety of etomidate has been questioned. Continuous infusions have been shown in a retrospective study to cause a significant increase in mortality [48]. Etomidate causes adrenal suppression by suppressing corticosteroid synthesis through the inhibition of the enzyme 11-β-hydroxylase, which converts 11-deoxycortisol to cortisol. This effect has been demonstrated with both infusions and with a single bolus. A single dose of etomidate reduces the synthesis of cortisol and aldosterone and increases the risk of relative adrenocortical insufficiency (RAI) for at least 24 hours [49]. Hypotension related to RAI has implications for CPP and neurological outcome. Etomidate may also lower seizure threshold [50]. Other adverse effects include pain on injection, myoclonic movements, and nausea and vomiting [51].
Therefore etomidate should be avoided as a continuous sedative agent in TBI but may be considered with caution as an induction agent, although ketamine offers many of the same advantages without the risks of adrenal suppression.
Go to:
7. Ketamine
See Table 7. Ketamine is an N-methyl-D-aspartate receptor antagonist. It has traditionally been avoided in the management of patients with traumatic brain injury owing to concerns that it may increase intracranial pressure. Furthermore, there are theoretical concerns regarding its epileptogenic potential. Indeed, it receives little attention in guidelines for the management of TBI [1]. Conversely, it has been argued that in comparison to most widely used sedative agents ketamine does not decrease blood pressure and therefore may preserve cerebral perfusion pressure. In particular, it has been argued that this haemodynamic stability enables ketamine to be used as a safe induction agent in patients with TBI [52].
Table 7
Ketamine
Group Phencyclidine derivative
Mechanism of Action/Pharmacodynamics Competitive NMDA receptor antagonist
Interaction with opioid and muscarinic receptors
Na+ Channel
Effect on ICP None or decrease
Neuroprotective effects Decreased glutamate
Pharmacokinetics 20% Bioavailability
40% protein bound
Distribution t 1/2 10 minutes
Hepatic metabolism
Elimination t 1/2 2.5 h
Advantages Preserves MAP and CPP
Disadvantages and major side effects Early studies ↑ICP, ?epileptogenic
Hallucinations/Emergence phenomena
Dosage Induction: 2 mg/kg
Maintenance: 50 mcg/kg/min
Other significant facts
Appropriate uses in TBI Haemodynamic instability
Open in a separate window
Concerns regarding the potential for ketamine to raise ICP stem from small case control series several decades ago in patients with abnormal CSF flow dynamics [53]. A rise in ICP was observed in spontaneously breathing patients, undergoing diagnostic pneumoventriculography, in whom ketamine was administered to. However, this rise in ICP only occurred in those patients with abnormal CSF pathways. In the remaining patients there was an overall rise in MAP, an increase in cerebral blood flow, and improved cerebral perfusion pressure [54, 55].
Several recent studies have refuted the original findings and showed no statistically significant rise in ICP in brain injured patients who are sedated with ketamine [56]. Bourgoin et al. randomised patients with TBI to receive either sufentanyl-midazolam or ketamine-midazolam sedation using target controlled infusions. The target concentrations of sufentanil and ketamine were doubled for 15 minutes, and the plasma concentrations of both were measured. There was no significant change in ICP or CPP with increased plasma concentrations. In an interesting editorial, the possibility that cerebral haemodynamics are better preserved through the use of target controlled infusion was discussed [57]. Whilst bolus doses of some commonly used sedatives may adversely affect haemodynamics and increase ICP, it is argued that a system relying on pharmacokinetic models alone is insufficient in managing patients with TBI.
Another study looked at the use of ketamine in 30 sedated and ventilated children with TBI and raised ICP resistant to first-tier therapies [58]. Variables examined included ICP, hemodynamic variables, and CPP. Ketamine was administered as a single dose of 1–1.5 mg/kg either to prevent further ICP increases during distressing procedures or as an additional measure to lower ICP. There was an overall decrease in ICP and increase in CPP in both situations. The authors conclude that ketamine is a safe and effective sedative agent to use in patients with TBI.
There is conflicting data as to whether ketamine induces epileptiform activity. The blocking of NMBA receptors and subsequent entry of calcium into neurons may limit seizure activity. Furthermore, the use of ketamine as an adjunct in the treatment of status epliepticus is well described in the literature [59]. The antagonism of NMDA receptors decreases the release of neurotoxic glutamate and may impart a protective effect in patients with traumatic brain injury [60].
Therefore ketamine is indicated particularly as an induction agent in patients with TBI and haemodynamic instability. It may have a role for refractory seizure activity.
Go to:
8. Dexmedetomidine
See Table 8. Dexmedetomidine is a highly selective alpha-2 receptor agonist that acts by a receptor distinct from the GABA receptor utilised by propofol and the benzodiazepines. A high selectivity for alpha-2 receptors, seven to eight times that of clonidine, explains its anxiolytic and sedative effects. A relatively short elimination t 1/2 of two hours enables intravenous titration to effect. Furthermore, dexmedetomidine does not appear to cause respiratory depression, with one study reporting no significant difference in respiratory rate and oxygen saturations between dexmedetomidine recipients and those that received placebo. This enables it to be continued after-extubation [61, 62]. Hypotension and bradycardia are among the most commonly reported side effects of dexmedetomidine, particularly when using a loading dose. For this reason, some commentators recommend an avoidance of a loading dose in patients with TBI.
Table 8
Dexmedetomidine
Group Selective α2 adrenergic agonist
Mechanism of Action/Pharmacodynamics Peripheral α2A, brain & spinal cord α2B, α2C adrenoreceptor subtypes
Neuroprotective effects Reduces CBF and ICP
Pharmacokinetics
Hepatic metabolism
Distribution t 1/2 6 minutes
Elimination t 1/2 2 hours
Advantages Minimal respiratory depression
Reduction in delerium
Disadvantages and major side effects Hypotension (28%)
Bradycardia
Arrhythmias including atrial fibrillation
Relatively high cost
Dosage Loading dose: 1 mcg/kg
Infusion: 0.42–1.0 mcg/kg/hour
Other significant facts Minimal effect on respiratory function
Appropriate uses in TBI Maintenance sedation agent pre & post extubation
Management of agitated delirium
Open in a separate window
Several trials have examined the use of dexmedetomidine sedation in ICU patients.
Riker et al. performed a prospective, double-blinded RCT in medical and surgical ICU patients comparing the efficacy and safety of dexmedetomidine with midazolam sedation [63]. Patients in the dexmedetomidine arm spent less time on the ventilator and experienced less hypertension and tachycardia. 42.2% of patients in the dexmedetomidine arm experienced bradycardia compared to 18.9% of patients who received midazolam sedation.
A potential advantage of dexmedetomidine may be in decreasing the incidence or severity of delirium. Many commonly used sedatives, including opioids and benzodiazepines, have been shown to increase the risk of delirium. In one prospective, double-blinded RCT, patients after cardiac surgery were randomised to receive either a dexmedetomidine or morphine-based sedative regimen [64]. Patients in the dexmedetomidine arm showed a significant reduction in the duration of delirium, although there was no statistically significant reduction in the incidence of delirium. A reduction in the incidence of delirium was also found in an a priori subgroup analysis of the MENDS trial. There was a reduced duration of brain dysfunction, particularly in septic patients [58].
There have been relatively few studies examining the role of dexmedetomidine in patients with TBI. Its use in neurosurgical patients was described in a retrospective study by Aryan et al. [65]. They describe a mean increase in cerebral perfusion pressure and a decrease in intracranial pressure in the 39 patients studied. The relatively small sample size and retrospective nature of this study limit its conclusions, and the authors argue for further studies to establish an optimal dosage regimen in neurosurgical patients. Grof et al. undertook a small, prospective, observational study, of patients receiving dexmedetomidine on a neurosurgical ICU [66]. The majority of these patients had traumatic brain injury. Dexmedetomidine was utilised in an attempt to wean patients off other sedative regimens. Relatively high doses of dexmedetomidine were required to achieve the desired level of sedation, up to a rate of 2.5 mcg/kg/hour. The authors postulate that significant changes in neurotransmitter systems in TBI might explain the need for higher doses of dexmedetomidine in this patient population.
There is a need for further high-quality RCTs to evaluate the use of dexmedetomidine as a sedative agent both in general ICU patients and in patients with TBI. The SPICE pilot study will examine the feasibility of conducting a large multi-centre trial, comparing current sedation practice with a dexmedetomidine-based sedation regimen. The DahLIA trial is currently recruiting patients and is a prospective, double-blinded RCT comparing dexmedetomidine to placebo in the treatment of delirium and agitation.
Therefore dexmedetomidine has a number of potential advantages as a sedative agent in TBI. There is evidence that it may reduce delirium and a lack of respiratory depressant effects enables it to be used in non-intubated patients.
Go to:
9. Conclusion
Sedation is a vital component of the management of patients with traumatic brain injuries. However, there is limited high-quality evidence examining outcomes in TBI to guide clinicians on their choice of agent. Consequently a wide variety of agents and dosages are used. Recent work has challenged traditional views on the best agents to use in TBI. For example, there is increasing evidence that ketamine may be safe to use in TBI as an induction agent and has advantages over traditional agents such as the barbiturates. There has also been increased interest in shorter acting, newer agents such as remifentanil and dexmedetomidine. These offer potential advantages in allowing faster recovery of consciousness and assessment of neurology in patients.
There is a need for further prospective, randomised controlled trials, examining both physiological and clinical outcomes, to assess these agents in the context of traumatic brain injury. Meanwhile, in the absence of extensive high-quality evidence to guide clinicians in their choice of agent, there is a need for a pragmatic approach, based on the clinical situation and knowledge of the potential advantages and disadvantages of each agent.
Go to:
10. Recommendations
See Table 9. For induction of anaesthesia in TBI, there is no single agent that is perfect and the way in which the medication is given, and the dose used, may be more important than the agents themselves. Considered preparation, experienced assistants and meticulous control and monitoring of blood pressure are essential. Thiopentone remains a reasonable choice, with the proviso that it is used judiciously in the haemodynamically unstable patient. Alternative induction agents include propofol (usually requiring a concomitant vasopressor bolus) or ketamine. There is little role for etomidate either as an agent for induction or continued sedation.
Table 9
Induction agents
(i) Haemodynamically unstable Ketamine (2 mg/kg) OR Midazolam (0.1 mg/kg) and fentanyl (1–3 mcg/kg)
(ii) Haemodynamically stable Thiopentone (1–3 mg/kg OR propofol (0.5–2.5 mg/kg), with fentanyl (1–3 mcg/kg)
Maintenance agents Propofol (1.5–4.5 mg/kg/h) and fentanyl (0.5–2 mcg/kg/h)
Open in a separate window
Propofol as an agent for continued sedation, usually administered with a short-acting narcotic, offers the advantage of a relatively rapid offset of sedation, facilitating neurological assessment. Remifentanil has many advantages over other narcotics in this setting as long as hyperalgesia on cessation is considered. In patients who require high doses of propofol, hypotensive patients, or for more prolonged sedation, midazolam is a suitable alternative. Thiopentone is not indicated as a maintenance sedative agent in TBI, and its use is primarily limited to the treatment of refractory intracranial hypertension. Dexmedetomidine shows promise as a sedative agent in TBI, particularly in the non-intubated patient.
Thanks to Professor Richard Lee for his helpful suggestions regarding this review.
Go to:
References
1. Guidelines for the management of severe traumatic brain injury. Journal of Neurotrauma. 2007;24(supplement 1):S1–S106. [PubMed] [Google Scholar]
2. Fearnside MR, Cook RJ, McDougall P, McNeil RJ. The Westmead Head Injury Project outcome in severe head injury. A comparative analysis of pre-hospital, clinical and CT variables. British Journal of Neurosurgery. 1993;7(3):267–279. [PubMed] [Google Scholar]
3. Chesnut RM, Marshall LF, Klauber MR, et al. The role of secondary brain injury in determining outcome from severe head injury. Journal of Trauma. 1993;34(2):216–222. [PubMed] [Google Scholar]
4. Roberts DJ, Hall RI, Kramer AH, et al. Sedation for critically ill adults with severe traumatic brain injury: a systematic review of randomized controlled trials. Critical Care Medicine. 2011;39(12):2743–2751. [PubMed] [Google Scholar]
5. Barash C, Stoelting RK, Cahalan M, Stock MC, editors. Clinical Anesthesia. Philadephia, PA, USA: Lippencott Williams & Wilkins; 2009. [Google Scholar]
6. Turcant A, Delhumeau A, Premel-Cabic A. Thiopental pharmacokinetics under conditions of long-term infusion. Anesthesiology. 1985;63(1):50–54. [PubMed] [Google Scholar]
7. Khan S, Stannard N, Greijn J. Precipitation of thiopental with muscle relaxants: a potential hazard. JRSM Short Reports. 2011;2(7, article 58) [PMC free article] [PubMed] [Google Scholar]
8. Pinaud M, Lelausque JN, Chetanneau A, Fauchoux N, Menegalli D, Souron R. Effects of propofol on cerebral hemodynamics and metabolism in patients with brain trauma. Anesthesiology. 1990;73(3):404–409. [PubMed] [Google Scholar]
9. Farling PA, Johnston JR, Coppel DL. Propofol infusion for sedation of patients with head injury in intensive care. A preliminary report. Anaesthesia. 1989;44(3):222–226. [PubMed] [Google Scholar]
10. Ronan KP, Gallagher TJ, George B, Hamby B. Comparison of propofol and midazolam for sedation in intensive care unit patients. Critical Care Medicine. 1995;23(2):286–293. [PubMed] [Google Scholar]
11. Mustafa AG, Singh IN, Wang J, Carrico KM, Hall ED. Mitochondrial protection after traumatic brain injury by scavenging lipid peroxyl radicals. Journal of Neurochemistry. 2010;114(1):271–280. [PMC free article] [PubMed] [Google Scholar]
12. Jayakumar AR, Rao KV, Panickar KS, et al. Trauma-induced cell swelling in cultured astrocytes. Journal of Neuropathology and Experimental Neurology. 2008;67(5):417–427. [PubMed] [Google Scholar]
13. Tanguy M, Seguin P, Laviolle B, et al. Cerebral microdialysis effects of propofol versus midazolam in severe traumatic brain injury. Journal of Neurotrauma. 2012;29(6):1105–1110. [PubMed] [Google Scholar]
14. Junttila MJ, Gonzalez M, Lizotte E, et al. Induced brugada-type electrocardiogram, a sign for imminent malignant arrhythmias. Circulation. 2008;117(14):1890–1893. [PubMed] [Google Scholar]
15. Cremer OL, Moons KGM, Bouman EAC, Kruijswijk JE, De Smet AMGA, Kalkman CJ. Long-term propofol infusion and cardiac failure in adult head-injured patients. The Lancet. 2001;357(9250):117–118. [PubMed] [Google Scholar]
16. Otterspoor LC, Kalkman CJ, Cremer OL. Update on the propofol infusion syndrome in ICU management of patients with head injury. Current Opinion in Anaesthesiology. 2008;21(5):544–551. [PubMed] [Google Scholar]
17. Donmez A, Arslan G, Pirat A, Demirhan B. Is pancreatitis a complication of propofol infusion? European Academy of Anaesthesiology. 1999;16(6):367–370. [PubMed] [Google Scholar]
18. Langevin PB, Gravenstein N, Doyle TJ, et al. Growth of Staphylococcus aureus in Diprivan and Intralipid: implications on the pathogenesis of infections. Anesthesiology. 1999;91(5):1394–1400. [PubMed] [Google Scholar]
19. Hodkinson BP, Frith RW, Mee EW. Proprofol and the electroencephalogram. The Lancet. 1987;2(8574):p. 1518. [PubMed] [Google Scholar]
20. Sutherland MJ, Burt P. Propofol and seizures. Anaesthesia and Intensive Care. 1994;22(6):733–737. [PubMed] [Google Scholar]
21. Carley S, Crawford I. Towards evidence based emergency medicine: best BETs from the Manchester Royal Infirmary. Propofol for resistant status epilepticus. Emergency medicine Journal. 2002;19(2):143–144. [PMC free article] [PubMed] [Google Scholar]
22. Illievich UM, Petricek W, Schramm W, Weindlmayr-Goettel M, Czech T, Spiss CK. Electroencephalographic burst suppression by propofol infusion in humans: hemodynamic consequences. Anesthesia and Analgesia. 1993;77(1):155–160. [PubMed] [Google Scholar]
23. Wong JM. Propofol infusion syndrome. American Journal of Therapeutics. 2010;17(5):487–491. [PubMed] [Google Scholar]
24. Matta B, Menon D. Severe head injury in the United Kingdom and Ireland: a survey of practice and implications for management. Critical Care Medicine. 1996;24(10):1743–1748. [PubMed] [Google Scholar]
25. Rhoney DH, Parker D. Use of sedative and analgesic agents in neurotrauma patients: effects on cerebral physiology. Neurological Research. 2001;23(2-3):237–259. [PubMed] [Google Scholar]
26. Urwin SC, Menon DK. Comparative tolerability of sedative agents in head-injured adults. Drug Safety. 2004;27(2):107–133. [PubMed] [Google Scholar]
27. Allonen H, Ziegler G, Klotz U. Midazolam kinetics. Clinical Pharmacology and Therapeutics. 1981;30(5):653–661. [PubMed] [Google Scholar]
28. Papazian L, Albanese J, Thirion X, Perrin G, Durbec O, Martin C. Effect of bolus doses of midazolam on intracranial pressure and cerebral perfusion pressure in patients with severe head injury. British Journal of Anaesthesia. 1993;71(2):267–271. [PubMed] [Google Scholar]
29. Fleischer JE, Milde JH, Moyer TP, Michenfelder JD. Cerebral effects of high-dose midazolam and subsequent reversal with Ro 15–1788 in dogs. Anesthesiology. 1988;68(2):234–242. [PubMed] [Google Scholar]
30. Mets B, Horsell A, Linton DM. Midazolam-induced benzodiazepine withdrawal syndrome. Anaesthesia. 1991;46(1):28–29. [PubMed] [Google Scholar]
31. Pandharipande P, Cotton BA, Shintani A, et al. Prevalence and risk factors for development of delirium in surgical and trauma intensive care unit patients. The Journal of Trauma. 2008;65(1):34–41. [PMC free article] [PubMed] [Google Scholar]
32. Morandi A, Jackson JC. Delirium in the intensive care unit: a review. Neurologic Clinics. 2011;29(4):749–763. [PubMed] [Google Scholar]
33. Camps AS, Sanchez-Izquierdo Riera JA, Vazquez DT, Sa Borges M, Rodriguez JP, Lopez EA. Midazolam and 2% propofol in long-term sedation of traumatized, critically ill patients: efficacy and safety comparison. Critical Care Medicine. 2000;28(11):3612–3619. [PubMed] [Google Scholar]
34. Ghori KA, Harmon DC, Elashaal A, et al. Effect of midazolam versus propofol sedation on markers of neurological injury and outcome after isolated severe head injury: a pilot study. Critical Care and Resuscitation. 2007;9(2):166–171. [PubMed] [Google Scholar]
35. Egerod I, Jensen MB, Herling SF, Welling KL. Effect of an analgo-sedation protocol for neurointensive patients: a two-phase interventional non-randomized pilot study. Critical Care. 2010;14(2, article R71) [PMC free article] [PubMed] [Google Scholar]
36. Osborne RJ, Joel SP, Slevin ML. Morphine intoxication in renal failure: the role of morphine-6-glucuronide. British Medical Journal. 1986;292(6535):1548–1549. [PMC free article] [PubMed] [Google Scholar]
37. Scholz J, Steinfath M, Schulz M. Clinical pharmacokinetics of alfentanil, fentanyl and sufentanil. An update. Clinical Pharmacokinetics. 1996;31(4):275–292. [PubMed] [Google Scholar]
38. Smith NT, Benthuysen JL, Bickford RG, et al. Seizures during opioid anesthetic induction—are they opioid-induced rigidity? Anesthesiology. 1989;71(6):852–862. [PubMed] [Google Scholar]
39. Egan TD, Lemmens HJM, Fiset P, et al. The pharmacokinetics of the new short-acting opioid remifentanil (GI87084B) in healthy adult male volunteers. Anesthesiology. 1993;79(5):881–892. [PubMed] [Google Scholar]
40. Karabinis A, Mandragos K, Stergiopoulos S, et al. Safety and efficacy of analgesia-based sedation with remifentanil versus standard hypnotic-based regimens in intensive care unit patients with brain injuries: a randomised, controlled trial [ISRCTN50308308] Critical Care. 2004;8(4):R268–280. [PMC free article] [PubMed] [Google Scholar]
41. Jeevaratnam DR, Menon DK. Survey of intensive care of severely head injured patients in the United Kingdom. British Medical Journal. 1996;312(7036):944–947. [PMC free article] [PubMed] [Google Scholar]
42. Stept WJ, Safar P. Rapid induction-intubation for prevention of gastric-content aspiration. Anesthesia and Analgesia. 1970;49(4):633–636. [PubMed] [Google Scholar]
43. Roberts I. Barbiturates for acute traumatic brain injury. Cochrane Database of Systematic Reviews. 2000;(2)CD000033 [PubMed] [Google Scholar]
44. Gooding JM, Weng JT, Smith RA. Cardiovascular and pulmonary responses following etomidate induction of anesthesia in patients with demonstrated cardiac disease. Anesthesia and Analgesia. 1979;58(1):40–41. [PubMed] [Google Scholar]
45. Van Hamme MJ, Ghoneim MM, Ambre JJ. Pharmacokinetics of etomidate, a new intravenous anesthetic. Anesthesiology. 1978;49(4):274–277. [PubMed] [Google Scholar]
46. Moss E, Powell D, Gibson RM, McDowall DG. Effect of etomidate on intracranial pressure and cerebral perfusion pressure. British Journal of Anaesthesia. 1979;51(4):347–352. [PubMed] [Google Scholar]
47. Modica PA, Tempelhoff R. Intracranial pressure during induction of anaesthesia and tracheal intubation with etomidate-induced EEG burst suppression. Canadian Journal of Anaesthesia. 1992;39(3):236–241. [PubMed] [Google Scholar]
48. Watt I, McA LI. Mortality amongst multiple trauma patients admitted to an intensive therapy unit. Anaesthesia. 1984;39(10):973–981. [PubMed] [Google Scholar]
49. Malerba G, Romano-Girard F, Cravoisy A, et al. Risk factors of relative adrenocortical deficiency in intensive care patients needing mechanical ventilation. Intensive Care Medicine. 2005;31(3):388–392. [PubMed] [Google Scholar]
50. Ding Z, White PF. Anesthesia for electroconvulsive therapy. Anesthesia and Analgesia. 2002;94(5):1351–1364. [PubMed] [Google Scholar]
51. Forman SA. Clinical and molecular pharmacology of etomidate. Anesthesiology. 2011;114(3):695–707. [PMC free article] [PubMed] [Google Scholar]
52. Filanovsky Y, Miller P, Kao J. Myth: ketamine should not be used as an induction agent for intubation in patients with head injury. Canadian Journal of Emergency Medicine. 2010;12(2):154–201. [PubMed] [Google Scholar]
53. Gibbs JM. The effect of intravenous ketamine on cerebrospinal fluid pressure. British Journal of Anaesthesia. 1972;44(12):1298–1302. [PubMed] [Google Scholar]
54. Evans J, Rosen M, Weeks RD, Wise C. Ketamine in neurosurgical procedures. The Lancet. 1971;1(7688):40–41. [PubMed] [Google Scholar]
55. Shapiro HM, Wyte SR, Harris AB. Ketamine anaesthesia in patients with intracranial pathology. British Journal of Anaesthesia. 1972;44(11):1200–1204. [PubMed] [Google Scholar]
56. Bourgoin A, Albanèse J, Léone M, Sampol-Manos E, Viviand X, Martin C. Effects of sufentanil or ketamine administered in target-controlled infusion on the cerebral hemodynamics of severely brain-injured patients. Critical Care Medicine. 2005;33(5):1109–1113. [PubMed] [Google Scholar]
57. Lele A, Ziai WC. Target-controlled infusion for sedation of traumatic brain-injured patients: role uncertain. Critical Care Medicine. 2005;33(5):1172–1174. [PubMed] [Google Scholar]
58. Pandharipande PP, Sanders RD, Girard TD, et al. Effect of dexmedetomidine versus lorazepam on outcome in patients with sepsis: an a priori-designed analysis of the MENDS randomized controlled trial. Critical Care. 2010;14(2, article R38) [PMC free article] [PubMed] [Google Scholar]
59. Sheth RD, Gidal BE. Refractory status epilepticus: response to ketamine. Neurology. 1998;51(6):1765–1766. [PubMed] [Google Scholar]
60. Bar-Joseph G, Guilburd Y, Tamir A, Guilburd JN. Effectiveness of ketamine in decreasing intracranial pressure in children with intracranial hypertension: clinical article. Journal of Neurosurgery. 2009;4(1):40–46. [PubMed] [Google Scholar]
61. Carollo DS, Nossaman BD, Ramadhyani U. Dexmedetomidine: a review of clinical applications. Current Opinion in Anaesthesiology. 2008;21(4):457–461. [PubMed] [Google Scholar]
62. Martin E, Ramsay G, Mantz J, Sum-Ping STJ. The role of the α2-adrenoceptor agonist dexmedetomidine in postsurgical sedation in the intensive care unit. Journal of Intensive Care Medicine. 2003;18(1):29–41. [PubMed] [Google Scholar]
63. Riker RR, Shehabi Y, Bokesch PM, et al. Dexmedetomidine vs midazolam for sedation of critically Ill patients A randomized trial. Journal of the American Medical Association. 2009;301(5):489–499. [PubMed] [Google Scholar]
64. Shehabi Y, Grant P, Wolfenden H, et al. Prevalence of delirium with dexmedetomidine compared with morphine based therapy after cardiac surgery: a randomized controlled trial (DEXmedetomidine compared to morphine-DEXCOM study) Anesthesiology. 2009;111(5):1075–1084. [PubMed] [Google Scholar]
65. Aryan HE, Box KW, Ibrahim D, Desiraju U, Ames CP. Safety and efficacy of dexmedetomidine in neurosurgical patients. Brain Injury. 2006;20(8):791–798. [PubMed] [Google Scholar]
66. Grof TM, Bledsoe KA. Evaluating the use of dexmedetomidine in neurocritical care patients. Neurocritical Care. 2010;12(3):356–361. [PubMed] [Google Scholar]
67. Allely P. What Is Brugada Syndrome? 2009, http://lifeinthefastlane.com/2009/09/what-is-brugada-syndrome/
Articles from Emergency Medicine International are provided here courtesy of Hindawi Limited
OTHER FORMATS
PDF (3.4M)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 1534226 | https://sv.wikipedia.org/wiki/Tianeptin | Tianeptin | Tianeptin (varumärkena Stablon, Coaxil, Tatinol, Tianeurax och Salymbra) är en medicin som används primärt i behandlingen av kronisk depression. Den har dock visat på potential som behandling av astma och IBS – känslig tarm. Kemiskt sett så är den nära besläktad med tricykliska antidepressiva (TCA), men dess farmakologiska egenskaper är närmast helt atypiska i jämförelse med TCA-preparaten – nyare forskning tyder på att Tianeptin verkar antidepressivt genom indirekt förändring av glutamaterg aktivitet (på AMPA och NMDA -receptorerna) och genom frigörelse av BDNF, som i sin tur påverkar neuroplasticitet.
Det fullständiga systematiska namnet för substansen är 7-[(3-kloro-6-metyl-5,5-dioxo-11H-benzo[c][2,1]benzotiazepin-11-yl)amino]heptanoisk syra.
Tianeptin har antidepressiva och anxiolytiska (ångestdämpande) egenskaper utan några framträdande sederande, antikolinerga eller kardiovaskulära bieffekter, vilket har gjort att den anses som särskilt lämplig i vården av äldre och alkoholister, då sådana personer kan vara mer känsliga för bieffekter från psykotropiska mediciner. Nyare resultat indikerar även potentiella antiepileptiska och smärtstillande egenskaper hos Tianeptin, via antingen direkt eller indirekt modulation av adenosin A1-receptorerna (eftersom dessa effekter har funnits kunna blockeras av experimentella antagonister av denna receptor).
Tianeptin är en lågeffekts men full agonist av de opioida μ- och δ- receptorerna, men inte på κ-receptorerna.
Opiata μ-agonister inducerar i regel eufori och så har även Tianeptin funnits göra, på doser långt utöver de som används inom vården av patienter.
Tianeptin upptäcktes och patenterades av Franska Nationalinstitutet för Medicin i Paris - Société de Médecine de Paris – på 1960-talet. För tillfället är Tianeptin godkänd i Frankrike där den tillverkas och marknadsförs av Laboratories Servier SA, men den marknadsförs under licens i ett flertal andra europeiska länder under varunamn som ”Coaxil”, såväl som i Asien (inklusive Singapore) och Latinamerika under namnen ”Stablon” och ”Tatinol” men den är inte tillgänglig på marknaden i Australien, Kanada, Nya Zeeland, Storbritannien eller USA.
Ej heller är Tianeptin godkänt av läkemedelsverket i Sverige.
Medicinsk användning
Tianeptin har effekt mot allvarliga depressiva episoder (kronisk depression), jämförbar med Amitryptylin, Imipramin och Fluoxetin, men med signifikant färre bieffekter.
Den har även visat sig vara mer effektiv än Maprotilin i en patientgrupp bestående av patienter med både depression och ångestsjukdom. Tianeptin uppvisar även betydande anxiolytiska (ångestdämpande) egenskaper och är användbar för att behandla ett spektrum av olika ångestsjukdomar, inklusive panikångest. Detta bevisades i en studie där deltagarna gavs en blandning av 65% syre och 35% koldioxid-gas (vilket inducerar panikångest) och där Tianeptin visade jämförbar förmåga med Paroxetin i att blockera paniksymptom.
Likt många andra antidepressiva (såsom bupropion, ssri, snri och moclobemid) så kan den ha fördelaktiga effekter på kognitionen hos människor med depressionsinducerad kognitiv nedsättning.
Tianeptin har visat sig vara effektiv i behandlingen av depression som en del av Parkinsons sjukdom samt posttraumatiskt stressyndrom, där den visade sig lika säker och effektiv som Fluoxetin och Moclobemid. .
En klinisk prövning har genomförts för att testa dess effektivitet och tolererbarhet i jämförelse med Amitryptylin vid behandling av IBS – känslig tarm. Resultaten av prövningen visade att Tianeptin var minst lika effektivt som Amitryptylin och producerade mindre bieffekter såsom muntorrhet och förstoppning.
Tianeptin har även rapporterats vara mycket effektiv för behandlingen av astma. I augusti 1998 publicerade Dr. Fuad Lechin och kollegor vid Central University of Venezuelas institut för experimentell medicin i Caracas resultaten från en 52-veckors dubbelblindad placebo-kontrollerad studie av behandlingen av astmatiska barn; barnen i gruppen som fick Tianeptin hade en markant minskning i uppmätta astmatiska symptom och ökad lungfunktion.
Två år tidigare hade samma grupp funnit en nära korrelation mellan nivåerna av fritt serotonin i blodplasma och allvarsgraden av symptomen hos deras patienter. Då Tianeptin var den enda kända förening som visat sig både reducera mängden fritt serotonin i blodplasma samt förstärka återupptaget av serotonin i blodplättar så bestämde de sig för att använda Tianeptin för att se om en reduktion av nivåerna av fritt serotonin i blodplasma skulle hjälpa med astmasymptomen.
Fram tills november 2004 har det genomförts två dubbelblindade placebo-kontrollerade studier och en öppen 25 000 personers studie som varade över 7 år, som alla påvisade effektivitet.
En egyptisk studie från 2005 visade på att Tianeptin hade effekt på män med depression relaterad till erektil dysfunktion.
Tianeptin har även antiepileptiska och smärtlindrande effekter och en klinisk prövning i Spanien med avslutning i januari 2007 visade att Tianeptin är effektiv i behandlingen av smärta relaterad till Fibromyalgi.
Tianeptin har även visat viss effekt, med minimala bieffekter, i behandlingen av ADHD.
Bieffekter
I jämförelse med de närbesläktade TCA-preparaten så orsakar Tianeptin betydligt färre kardiovaskulära, antikolinerga (såsom muntorrhet eller förstoppning), sederande eller aptitstimulerande bieffekter. En nyligen genomförd överblickande granskning fann att den var bland de antidepressiva som uppvisade mest tendens att orsaka leverskada – dock så är bevisen för detta svaga och har ifrågasatts. Även om bieffekten inte har uppvisats hos Tianeptin, så är de närbesläktade TCA-preparaten kända för att orsaka hjärtarytmier, varför Tianeptin även misstänks ha denna potentiella bieffekt.
Efter förekomst
Källor:
Vanliga (>1% förekomst)
Huvudvärk (upp till 18%)
Yrsel(upp till 10%)
Sömnlöshet/mardrömmar (upp till 20%)
Dåsighet (upp till 10%)
Muntorrhet (upp till 20%)
Förstoppning (upp till 15%)
Illamående
Buksmärta
Viktuppgång (~3%)
Oro
Ångest/lättretlighet
Ovanliga (0.1-1% förekomst)
Bitter smak
Gaser i magen
Gastralgi
Dimsyn
Muskelvärk
Ventrikulärt extraslag
Störningar vid urinering
Hjärtklappning
Blodtrycksfall
Värmevallning
Tremor
Sällsynta(<0,1% förekomst)
Hepatit
Hypomani
Eufori
ECG förändringar
Pruritus/allergiska hudreaktioner
Utdragen muskelvärk
Allmän trötthet
Rekreationsbruk och potentiellt beroendeskapande
Tianeptin har använts genom intravenös injektion av missbrukare i Ryssland.
Denna intagningsmetod orsakar enligt rapporter ett svagt opioidliknande rus och görs ibland i ett försök att minska symptomen från opioidabstinens.
Såsom många andra tabletter så innehåller dock Tianeptin silica vilket gör tabletterna svårupplösta. Ofta är den lösning som produceras inte filtrerad tillräckligt väl vilket innebär att större partiklar blir kvar och när dessa injiceras intravenöst så blockerar de kapillärerna, vilket leder till trombosis (blodpropp) med påföljande vävnadsdöd. Därför är försäljningen av Tianeptin (under varunamnet "Coaxil") i Ryssland hårt reglerad.
Kontraindikationer
Kända kontraindikationer inkluderar följande:
Behandling med Monoaminoxidashämmare 14 dagar eller mindre före behandling med tianeptin. P ga potentiella kardiovaskulära effekter (såsom Hypertoni och Cirkulatorisk kollaps), konvulsioner (krampanfall), hypertermi (hög kroppstemperatur) och plötslig död.
Hypersensitivitet mot Tianeptin eller något annat innehållsämne i tabletterna.
Om du är under 15 års ålder.
Verkningsmekanism
Under de initiala studierna så fann man först att den omedelbara och upprepade effekten av tianeptin var att det minskade de extracellulära nivåerna av serotonin i försöksdjurens hjärnor, utan att minska själva frisläppet av serotonin - därmed kallades den för världens första selektiva serotoninåterupptagsförstärkare.
Tianeptin har funnits vara en verksam μ-opioid receptor agonist(Ki (människa) med en affinitet på 383 ± 183 nM och ett EC50 (människa) på 194 ± 70 nM). Samma studie avslöjade även att den också är en funktionabel δ-opioid receptor agonist, men med mycket lägre potens.
Det är dessa effekter som anses vara ansvariga för preparatets missbrukspotential vid höga doser långt utöver den vanligt förskrivna.
Se även
Amineptin
Tricykliska antidepressiva
Antidepressiva läkemedel
Noter
Referenser
Externa länkar
Tianeptin informationsblad och bipacksedel från tillverkare
Tianeptine.com
Antidepressiva läkemedel | swedish | 0.469579 |
fainting_mechanism/PMC6597794.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List Physiol Rep v.7(13); 2019 Jul PMC6597794
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of physrep
Physiol Rep. 2019 Jul; 7(13): e14148.
Published online 2019 Jun 27. doi: 10.14814/phy2.14148
PMCID: PMC6597794
PMID: 31250563
Mechanisms of tilt‐induced vasovagal syncope in healthy volunteers and postural tachycardia syndrome patients without past history of syncope
Julian M. Stewart,corresponding author 1 Mohamed A. Shaban, 1 Tyler Fialkoff, 1 Brianna Tuma‐Marcella, 1 Paul Visintainer, 2 Courtney Terilli, 1 and Marvin S. Medow 1
Author information Article notes Copyright and License information PMC Disclaimer
Go to:
Abstract
Upright tilt table testing has been used to test for vasovagal syncope (VVS) but can result in “false positives” in which tilt‐induced fainting (tilt+) occurs in the absence of real‐world fainting. Tilt+ occurs in healthy volunteers and in patients with postural tachycardia syndrome (POTS) and show enhanced susceptibility to orthostatic hypotension. We hypothesized that the mechanisms for hypotensive susceptibility differs between tilt+ healthy volunteers (Control‐Faint (N = 12)), tilt+ POTS patients (POTS‐Faint (N = 12)) and a non‐fainter control group of (Control‐noFaint) (N = 10). Subjects were studied supine and during 70° upright tilt while blood pressure (BP), cardiac output (CO), and systemic vascular resistance (SVR), were measured continuously. Impedance plethysmography estimated regional blood volumes, flows, and vascular resistance. Heart rate was increased while central blood volume was decreased in both Faint groups. CO increased in Control‐Faint because of reduced splanchnic vascular resistance; splanchnic pooling was similar to Control‐noFaint. Splanchnic blood flow in POTS‐Faint decreased and resistance increased similar to Control‐noFaint but splanchnic blood volume was markedly increased. Decreased SVR and splanchnic arterial vasoconstriction is the mechanism for faint in Control‐Faint. Decreased CO caused by enhanced splanchnic pooling is the mechanism for faint in POTS‐Faint. We propose that intrahepatic resistance is increased in POTS‐Faint resulting in pooling and that both intrahepatic resistance and splanchnic arterial vasoconstriction are reduced in Control‐Faint resulting in increased splanchnic blood flow and reduced splanchnic resistance.
Keywords: Cardiac output, false positive Tilt, POTS, vasoconstriction, vasovagal syncope, young
Go to:
Introduction
Syncope is defined by rapid onset transient loss of consciousness and postural tone due to cerebral hypoperfusion with spontaneous recovery (Moya et al. 2009). Most of syncope in young patients is due to simple faint, denoted “vasovagal syncope” (VVS) (Sheldon et al. 2015), often triggered by upright posture. VVS is quite common in young people with an incidence of one or more events occurring in approximately 20–40% of the general population (van Steenwijk et al. 1995; Ganzeboom et al. 2003). The diagnosis of VVS can often be made by taking a detailed history, obtaining an electrocardiogram and performing a detailed physical exam to rule out potential contributory cardiac disease and orthostatic hypotension (Freeman et al. 2011; Wieling et al. 2015). VVS can occur in patients diagnosed with the postural tachycardia syndrome (POTS). The incidence of VVS in POTS patients is believed to be about 30%, similar to its incidence in the general population (Raj 2006; Kimpinski et al. 2012), which is consistent with our experience.
Upright tilt table testing has been used in the diagnosis of VVS. Caution is advised regarding interpretation because tilt testing can fail to provoke syncope in some young patients with clinically confirmed real‐world VVS, but can also result in VVS during tilt in many healthy young volunteers with no history of fainting in the real world (Fouad et al. 1993; van Steenwijk et al. 1995; Lewis et al. 1997). This latter group has been often denoted “false positive fainting” (Leonelli et al. 2000). We have observed similar percentages of “false positive” faints in POTS patients during 10‐min unmedicated upright tilt testing at 70°.
“False positive” tilt tests have been reevaluated as demonstrating “hypotensive susceptibility” to orthostatic stress which potentiated syncope of whatever origin (Sutton and Brignole 2014) via reduced cardiac venous return (Verheyden et al. 2008; Jardine et al. 2018) or reduced systemic vascular resistance in younger patients with VVS (van Steenwijk et al. 1995; Stewart et al. 2017a; Jardine et al. 2018).
POTS is related to absolute or redistributive central hypovolemia with reduced venous return (Stewart and Montgomery 2004). We recently showed that hypotensive susceptibility in young healthy volunteers is driven by reduced SVR, the predominant mechanism of VVS in the young (Stewart et al. 2017a). In the present study, we therefore hypothesize that mechanisms for hypotensive susceptibility in POTS is likely related to decreased venous return and cardiac output.
To test this hypothesis, we examined hemodynamics in POTS and healthy volunteer controls. Neither POTS patients nor controls had prior history of VVS and both groups fainted during upright tilt. We also compared these data to healthy volunteers who did not faint during tilt testing.
Go to:
Methods
Subjects
“False positive vasovagal syncope” enrollees comprised either POTS patients (designated POTS‐Faint) or healthy volunteer control subjects (designated Control‐Faint) free of past history of VVS but who developed vasovagal syncope during a 10‐min 70° upright tilt test. POTS patients had undergone an earlier upright 10‐min tilt test which confirmed the diagnosis of POTS including excessive tachycardia, symptoms of orthostatic intolerance (OI), and absence of hypotension.
VVS was identified by orthostatic prodromal features included pallor, lightheadedness, nausea with abdominal discomfort, diaphoresis, a feeling of warmth, visual scotomata or frank loss of vision, and loss of consciousness or impending loss of consciousness with hypotension and a relative bradycardia as defined below. Unconsciousness or impending loss of consciousness resolved in all participants within 30 sec when placed supine. All patients had normal resting electrocardiograms and resting physical examinations.
We prospectively enrolled 12 POTS‐Faint subjects aged 15–25 years old (mean age 20 ± 2 years, 10 females, 2 males) with POTS defined by standard criteria (Schondorf and Low 1993; Singer et al. 2011). Once POTS had been diagnosed previously by signs and symptoms of OI (Stewart et al. 2018) including an excessive increase in HR without hypotension within 10 min of a head‐up tilt during which they did not faint (Low et al. 1995; Raj 2006; Medow and Stewart 2007), patients were invited to enroll in this study. We only enrolled POTS patients categorized as “Normal Flow” to improve subject homogeneity, using our previously established techniques (Stewart et al. 2006). Symptoms of OI were present on a daily basis for >6 months and were relieved once supine. Excessive orthostatic tachycardia was defined in adolescents by an increase in heart rate (HR) by at least 40 bpm or to a HR > 130 bpm. Excessive orthostatic tachycardia was defined in patients over 19 years by an increase in HR by at least 30 bpm, or to a HR > 120 bpm, during a prior 10 min upright tilt table test (Freeman et al. 2011; Singer et al. 2011; Plash et al. 2013). Alternative medical or psychological problems that could explain these signs or symptoms had been ruled out. During participation in the present study, this subset of POTS subjects became syncopal during tilt table testing and is hence referred to as POTS‐Faint. We also prospectively enrolled 12 gender matched subjects aged 15–25 (mean age 21 ± 2 years, 10 females, 2 male) from a larger set of “false positive vasovagal syncope” healthy control subjects, designated Control‐Faint, from subjects who were evaluated for syncope.
We also prospectively recruited 10 healthy non‐fainting control subjects, with no past history of VVS or OI, aged 15 to 24 years (mean age 21 ± 1 years, 8 females, 2 males) for comparison with the two “faint” groups. These were designated Control‐noFaint. There were no differences in the ages, weight, and body mass index between groups. Since sex hormones influence cutaneous blood flow (Charkoudian and Johnson 1999), and systemic hemodynamics (Fu et al. 2010), all women in this study participated while in the early to midluteal phase, or high hormone phase of oral contraception, when autonomic tone is maximized (Minson et al. 2000).
Fainting patients did not meet the definition for orthostatic hypotension (Freeman et al. 2011) and were examined for cardiogenic causes of syncope; none were found. Control subjects reported no clinical illnesses, and had never previously fainted.
Exclusion criteria for participation in this study were any infectious or systemic disease (including other cardiovascular disease), recent long‐term bed rest, competitive athletic training, use of nicotine containing products or pregnancy within the last year. Medical therapy for POTS, if any, had been stopped for at least 2 weeks prior to participation in this study. Subjects refrained from caffeine for at least 72 h prior to testing. Subjects fasted for a minimum of 4 h prior to testing. This study was approved by the Institutional Review Board of New York Medical College. All subjects 18 or older signed an informed consent; those younger than 18 assented to participate and their parent or legal guardian signed an informed consent.
Protocol
Subjects arrived at our center at 9:30 am. Tests and instrumentation were explained. Subjects were instrumented while supine. Beat‐to‐beat blood pressure was measured by Finometer finger photoplethysmograph (FMS, Amsterdam, The Netherlands) on the right forefinger or middle finger. The Finometer estimates beat‐to‐beat CO by pulse‐contour analysis using the Modelflow algorithm® (Bogert and van Lieshout 2005). ModelFlow CO was calibrated against an Innocor inert gas rebreathing CO measurement (Innovision, Denmark) while supine before experiments began. We computed beat‐to‐beat systemic vascular resistance (SVR) by dividing the time average arterial pressure (mean arterial pressure, MAP) by the ModelFlow CO averaged over each cardiac cycle. We also computed the pulse pressure (PP) for each cardiac cycle by subtracting the diastolic from the systolic BP. Regional blood volumes, blood flows, and vascular resistance were measured by impedance plethysmography (Stewart et al. 2006). We placed paired electrodes using anatomic landmarks to estimate thoracic, splanchnic, pelvic, and calf segmental blood volumes. Respiratory plethysmography (Respitrace, NIMS Scientific, Miami Beach, FL) and capnography (Smith Medical PM, Waukesha, WI) measured changes in respiration and end tidal carbon dioxide (ETCO2). An electrocardiograph measured HR from the beat‐to‐beat cardiac electrical interval. Signals were acquired at 200 samples/s, multiplexed, and A/D converted using custom software.
Subjects remained awake and supine for 30 min to acclimate to instrumentation. Baseline data comprising averaged HR, BP, ETCO2, CO, SVR, and thoracic, splanchnic, pelvic, and calf segmental impedances and rate of change of impedances were collected. Baseline data over the 10 min immediately preceding tilt were used for comparison with tilted information.
Subjects were tilted upright to 70°. The duration of upright tilt was 10 min for Control‐noFaint. Fainting subjects remained upright until fainting was imminent which by design occurred within 10 min of tilt‐up. Tilt was performed without pharmacologic provocation. Continuous HR, BP, ETCO2, CO, SVR, and impedance data were recorded for off‐line analysis.
Fainting patients were tilted back to supine when syncope was imminent. Imminent vasovagal syncope was defined by a tilt‐induced decrease in mean arterial pressure (MAP) to <60 mmHg or a decrease in systolic BP (SBP) <70 mmHg associated with symptoms of impending loss of consciousness, severe lightheadedness, nausea, heat, or diaphoresis. Fainters developed classic vasovagal syncope with hypotension followed by bradycardia during tilt (Wieling et al. 2016; Stewart et al. 2017a; Jardine et al. 2018).
Detailed methods
Fiducial event markers We analyzed results at fiducial time points, rather than at specific time points in accordance with previous work (Taneja et al. 2008; Stewart et al. 2017a). Fiducial markers identified comparable times at which physiological events occurred allowing us to study events which corresponded to similar occurrences in each fainter. Six defining events points were determined from the BP trace of each VVS subject; these events are represented for one subject in Figure 1, and correspond to the phases of vasovagal syncope described by Jardine et al. (2018). The first fiducial point, was baseline, denoted “supine” on the graph. Following initial orthostatic hypotension (Wieling et al. 2007), BP stabilizes (Phase 1); this is denoted “1 min” and was chosen to be mid Phase 1 approximately 1 min after tilt. Thereafter, a gradual progressive early hypotension associated with reflexively increased HR occurred and was identified with the onset of Phase 2 and is designated “early.” BP falls off rapidly and abruptly in Phase 3; Late phase 2 was identified at the transition between Phase 2 and Phase 3 and is designated “late.” We defined a “mid” point as midway between “early” and “late.” The last fiducial point, designated “faint,” occurred at the time of imminent syncope, or at 10 min for Control‐noFaint subjects. The non‐fainting control subjects did not have a significant fall in BP. We defined equivalent fiducial time points for Control‐noFaint by taking the averaged time of occurrence of each fiducial marker in fainting subjects divided by the time from onset of tilt (the fractional time to marker) and multiplied by 10. Control‐noFaint and fainting subjects were thus compared at equivalent times to obtain a uniform evaluation of hemodynamic quantities across groups.
An external file that holds a picture, illustration, etc.
Object name is PHY2-7-e14148-g001.jpg
Figure 1
Head‐up tilt table test for a vasovagal syncope patient. Mean arterial pressure (MAP) is shown in the top panel and heart rate (HR) in the bottom panel. Fiducial points are marked at the top. MAP and HR follow a characteristic time course comprising four stages: Early blood pressure (BP) stabilization (Phase 1, fiducial point 2); “Progressive early hypotension” with BP slowly declining as HR increases (fiducial points 3–5); Syncope with hypotension and bradycardia (Phase 3, fiducial point 6). Phase 4 recovery of BP occurs within 30 sec once supine.
Measurement of changes in segmental blood volumes Changes in segmental or regional blood volumes and segmental blood flows employed impedance plethysmography (IPG) using a tetrapolar high‐resolution four‐channel digital impedance plethysmograph (UFI, Morro Bay, CA) to measure impedances and the rate of change of impedances as described previously (Montgomery 1993a,b). These quantities were obtained within four anatomic segments defined by electrode placement on anatomic landmarks that delimit respective regional circulations. These were designated the thoracic segment (supraclavicular area to xyphoid process), the splanchnic segment (xyphoid process to iliac crest), the pelvic segment including lower pelvis to the knee (iliac crest to knee), and the leg or calf segment (upper calf just below the knee to the ankle).
Measurement of changes in segmental blood flow IPG was also used to estimate segmental or regional blood flows (Montgomery et al. 1993). These methods have been validated in our laboratory against the reference standard indocyanine green dye techniques and calf venous occlusion plethysmography and were used to measure leg, thoracic, and splanchnic blood flow while subjects were supine and during incremental tilt‐table testing, as described previously (Stewart et al. 2006, 2007).
Data analysis and statistics
All data were digitized and stored and were analyzed off‐line with custom software. There were three subject groups for comparison: Control‐noFaint, Control‐Faint, and POTS‐Faint.
Baseline data for BP, HR, CI, SVR, and ETCO2 are shown in absolute units in Table 1. We also analyzed time to faint for both groups. These were compared among the groups by one‐way ANOVA. Significant interactions if any were further analyzed by converting the ratio of F values to a t‐distribution by use of a Scheffé test, and probabilities were determined thereafter. All tabular results are reported as mean ± SEM.
Table 1
Baseline hemodynamics
Control – No Faint Control – Faint POTS – Faint
Systolic BP (mmHg) 116 ± 4 120 ± 3 121 ± 4
Diastolic BP (mmHg) 61 ± 3 62 ± 3 65 ± 2
MAP (mmHg) 79 ± 2 83 ± 3 83 ± 3
HR (bpm) 64 ± 3 66 ± 2 73 ± 3a
CI (L/min/m2) 4.8 ± 0.5 5.3 ± 0.3 5.5 ± 0.4
SVR (mmHg/L/min) 18 ± 2 15 ± 2 17 ± 1
ETCO2 (Torr) 42 ± 2 42 ± 1 44 ± 1
Open in a separate window
a P < 0.05 compared to Control‐noFaint.
Graphic data are shown as absolute units for CI and SVR and as percent of baseline for segmental blood flows and resistance expressed as mean ± SEM. Data were obtained from original time series averaged over 15s intervals centered at the fiducial markers. Repeated‐measures ANOVA were used to compare groups at times before Faint. We assigned greatest importance to “group × time effects” representing the interaction of subject group with time dependent changes during upright tilt. We assumed a covariance structure of compound symmetry. Reported P‐values reflect the interaction term using the Greenhouse‐Geisser correction. Statistical significance was set at P ≤ 0.05. Results were calculated by using GraphPad Prism version 8. Significance (P‐values) appears in the figures as well as text.
Go to:
Results
Baseline supine data
Baseline data measured while subjects were supine are displayed in Table 1, which shows data for Control‐noFaint, Control‐Faint, and POTS‐Faint. There were no baseline differences in systolic, diastolic or mean BP, CI, SVR, or ETCO2 between Control‐noFaint and fainting groups. HR was significantly higher in POTS‐Faint compared with Control‐noFaint (P < 0.025).
Upright tilt data
Time to faint The time from tilt up to syncope for Control‐Faint averaged 375 ± 49s, and was not different from the time to syncope for POTS‐Faint which averaged 349 ± 58 sec. By definition, Control‐noFaint subjects did not experience symptoms of orthostatic intolerance nor did they faint.
Representative tracings during tilt, VVS groups Figure 2 shows BP, HR, CO, and SVR for representative subjects belonging to each group. HR increased in all groups during orthostatic stress imposed by an upright tilt to 70°, shown by the arrows. Following initial BP stabilization there was a gradual fall off in BP in both Control‐Faint and POTS‐Faint representing Phase 2. During Phase 2 CO increased above baseline for Control‐Faint while SVR decreased. During Phase 2 CO progressively decreased throughout tilt in POTS‐Faint while SVR increased. Phase 3 ensued in both Control‐Faint and POTS‐Faint with rapid hypotension followed by bradycardia associated with decreased SVR and CO. This did not occur in Control‐noFaint.
An external file that holds a picture, illustration, etc.
Object name is PHY2-7-e14148-g002.jpg
Figure 2
Blood pressure, heart rate, cardiac output, and systemic vascular resistance for representative subjects during head‐up tilt table testing. Figure 2 shows representative data from left to right for Control‐noFaint, Control‐Faint and POTS‐Faint subjects. Each data panel contains graphs of BP, HR, CO, and SVR in top to bottom order for representative subjects belonging to each group. HR increased in all groups during orthostatic stress. Following initial pressure stabilization there was a gradual fall off in BP in fainters representing Phase 2. During Phase 2, CO increased above baseline for Control‐Faint while SVR decreased and CO progressively decreased throughout tilt in POTS‐Faint while SVR increased. Phase 3 ensued with rapid hypotension followed by bradycardia associated with decreased SVR and CO in all fainters.
Averaged hemodynamic data are shown in Figures 3, ,4,4, ,5,5, ,66.
An external file that holds a picture, illustration, etc.
Object name is PHY2-7-e14148-g003.jpg
Figure 3
Averaged heart rate, mean arterial pressure, and pulse pressure during upright tilt. Heart rate (HR) appears in the upper panel, mean arterial pressure (MAP) in the middle panel and pulse pressure (PP) in the lower panel at designated fiducial time points. Control‐noFaint data are shown as open circles (Ο), Control‐Faint data are shown as black boxes (■), and POTS‐Faint are shown as black triangles (▲).HR was increased above control (P < 0.001) in all faint groups prior to faint. HR fell precipitously in all Fainters at the time of faint. MAP and PP decreased significantly throughout tilt in all VVS (P < 0.05) compared to Control‐noFaint subjects.
An external file that holds a picture, illustration, etc.
Object name is PHY2-7-e14148-g004.jpg
Figure 4
Regional blood flows during upright tilt. The top panel shows the cardiac output (CO) in L/min, the second shows percent change in splanchnic blood flow (%∆ Splanchnic Blood Flow), the third panel shows percent change in pelvic blood flow (%∆ Pelvic Blood Flow), and the bottom panel shows percent change in calf blood flow (%∆ Calf Blood Flow). Control‐noFaint data are shown as open circles (Ο), Control‐Faint data are shown as black boxes (■), POTS‐Faint are shown as black triangles (▲). CO decreased progressively in POTS‐Faint compared to Control‐noFaint (P < 0.001). Splanchnic blood flow was increased in Control‐Faint (P < 0.001).
An external file that holds a picture, illustration, etc.
Object name is PHY2-7-e14148-g005.jpg
Figure 5
Percent change in regional blood volumes during upright tilt. The top panel shows the percent change in central blood volume (%∆ Central Blood Volume), the second shows percent change in splanchnic blood volume (%∆ Splanchnic Blood Volume), the third panel shows percent change in pelvic blood volume (%∆ Pelvic Blood Volume) and the bottom panel shows percent change in calf blood volume (%∆ Calf Blood Volume). Control‐noFaint data are shown as open circles (Ο), Control‐Faint data are shown as black boxes (■), POTS‐Faint are shown as black triangles (▲). Central, splanchnic, pelvic, and calf blood volumes are expressed as percent change from supine, measured before subjects were tilted upright to 70°. The Percent Central Blood Volume (%CBV) decreased in all subjects with tilt. %CBV decreased significantly more in fainting subjects than Control‐noFaint (P < 0.01). The Percent Splanchnic Blood Volume (VolSpl) increased during tilt in all subjects, was similar to Control‐noFaint in Control‐Faint, but was significantly larger for POTS‐Faint compared to Control‐noFaint (P < 0.01). The Percent Pelvic Blood volume (VolPelv) increased similarly in all groups during tilt. %Calf Blood Volume increased with tilt for all subjects but was further increased in both fainting groups compared to Control‐noFaint (P < 0.0025).
An external file that holds a picture, illustration, etc.
Object name is PHY2-7-e14148-g006.jpg
Figure 6
Percent change in regional arterial vascular resistance during upright tilt. The top panel shows the percent change in systemic vascular resistance (SVR), the second shows percent change in splanchnic resistance (%ΔRSplanchnic), the third panel shows percent change in pelvic resistance (%ΔRPelvic) and the bottom panel shows percent change in calf resistance (%ΔRcalf). Control‐noFaint data are shown as open circles (Ο), Control‐Faint data are shown as black boxes (■), POTS‐Faint are shown as black triangles (▲). SVR and %ΔRSplanchnic were markedly reduced in Control‐Faint and slightly increased in POTS‐Faint. Splanchnic, pelvic, and calf blood vascular resistances are expressed as percent change from supine, measured before subjects were tilted upright to 70°. SVR increased by 1 min following tilt for all subjects. SVR then decreased progressively in Control‐Faint (P < 0.05). SVR remained similar to Control‐no‐Faint for POTS‐Faint SVR until syncope supervened. Splanchnic Resistance increased similarly for Control‐noFaint and POTS‐Faint until syncope occurred, while decreasing throughout tilt in Control‐Faint (P < 0.005).
HR, MAP, pulse pressure (PP) Following subjects being tilted upright to 70°, HR was increased above Control‐noFaint (P < 0.001) in Control‐Faint and POTS‐Faint before the “late” fiducial point when HR fell precipitously. MAP and PP were decreased significantly during tilt compared with Control‐noFaint (P < 0.05) (Fig. 3).
Regional blood flows Splanchnic, pelvic, and calf blood flows are expressed as percent change from supine, measured before subjects were tilted upright to 70°. Control‐noFaint had an initial decrease in CO by 1 min after tilt which was sustained. With tilt CO decreased progressively in POTS‐Faint compared with Control‐noFaint (P < 0.001). CO did not decrease with tilt in Control‐Faint but instead was sustained at supine values Group comparison with Control‐noFaint) until syncope supervened and CO fell precipitously (Fig. 4).
Splanchnic Blood Flow decreased similarly from supine in Control‐noFaint and in POTS‐Faint. Splanchnic blood flow increased from supine in Control‐Faint (P < 0.001), until syncope supervened and then fell precipitously. Pelvic Blood Flow decreased with tilt in all groups and was not different in Control‐Faint compared to Control‐noFaint until syncope supervened. Pelvic blood flow was reduced during tilt in POTS‐Faint compared to Control‐noFaint (P < 0.001). Calf Blood Flow decreased similarly with tilt for all subjects prior to faint but decreased for both fainting groups when syncope supervened.
Regional blood volumes Central, splanchnic, pelvic, and calf blood volumes are expressed as percent change from supine, measured before subjects were tilted upright to 70°. The Percent Central Blood Volume (%CBV) decreased in all subjects with tilt. %CBV decreased significantly more in fainting subjects than Control‐noFaint (P < 0.01). The Percent Splanchnic Blood Volume (VolSpl) increased during tilt in all subjects, was similar to Control‐noFaint in Control‐Faint, but was significantly larger for POTS‐Faint compared to Control‐noFaint (P < 0.01). The percent Pelvic Blood volume (VolPelv) increased similarly in all groups during tilt. The %Calf Blood Volume increased with tilt for all subjects but was further increased in both fainting groups compared to Control‐noFaint (P < 0.0025) (Fig. 5).
Regional arterial resistances Splanchnic, pelvic, and calf blood vascular resistances are expressed as percent change from supine, measured before subjects were tilted upright to 70°. SVR increased by 1 min following tilt for all subjects. SVR then decreased progressively in Control‐Faint (P < 0.05). SVR remained similar to Control‐no‐Faint for POTS‐Faint SVR until syncope supervened (Fig. 6).
Splanchnic Resistance increased similarly for Control‐noFaint and POTS‐Faint until syncope occurred, while decreasing throughout tilt in Control‐Faint (P < 0.005). Pelvic and calf Resistance increased in all groups and was larger than Control in POTS‐Faint until the time of syncope. Calf Resistance increased initially from supine in all subjects then decreased at the time of syncope in all fainters.
Go to:
Discussion
In this study, decreased SVR caused by impaired splanchnic arterial vasoconstriction in the absence of splanchnic pooling is the mechanism for faint in Control‐Faint. This is similar to the predominant mechanism for faint in young patients with recurrent VVS (van Steenwijk et al. 1995; Lautt 2009). Decreased CO caused by enhanced splanchnic pooling is the mechanism for faint in POTS‐Faint. This is similar to the predominant mechanism for faint in older recurrent VVS patients (Lautt 2009; Kimpinski et al. 2012).
We have previously demonstrated that decreased SVR, primarily due to splanchnic vasodilation rather than decreased cardiac output, was the main mechanism for VVS during orthostatic stress in young recurrent fainters. (Stewart et al. 2017a). We also showed that inhibiting NO synthesis increased adrenergic vasoconstriction, increased SVR and splanchnic vasodilation, and normalized orthostatic tolerance in young patients with VVS (Stewart et al. 2016, 2017b). Blood flow and vascular resistance responses to orthostatic stress in our current Control‐Faint group were similar to those found previously in recurrent VVS patients, findings caused exclusively by decreased SVR.
Our current observations contrast with those of Fu et al. (2012) who found that a minority of healthy volunteers who lacked real‐world recurrent syncope yet experienced VVS during upright tilt had decreased SVR as the mechanism for hypotension. The age range of their cohort was wider than ours, being both younger and older And studies have confirmed a progressively larger role of declining cardiac output with age in syncope in healthy volunteers (Hainsworth and Al‐Shamma 1988). Other studies of younger healthy volunteers experiencing vasovagal syncope or presyncope show maintained CO and falling SVR during orthostatic stress (Evans et al. 2001) in agreement with our present studies.
Our data also show that decreased CO, equivalent to decreased venous return, is the mechanism for fainting during upright tilt in POTS‐Faint patients, that is primarily due to venous pooling of blood within the splanchnic vasculature. Reduced venous return is consequent to excessive splanchnic blood pooling within the capacitance vessels of the liver and mesentery (Stewart et al. 2006) and to a lesser extent within the calf, but not within the lower abdominal and thigh vasculature (“pelvic segment”).
Splanchnic pooling in POTS‐Faint occurs despite sustained splanchnic vasoconstriction. One explanation for this finding would be an increase in splanchnic capacitance (decreased venoconstriction), but splanchnic venoconstriction and splanchnic arterial vasoconstriction typically occur in parallel (Pang 2001; Gelman and Mushlin 2004) and splanchnic vasoconstriction appears intact. The distributed hemodynamic properties of the splanchnic vasculature, arterial inflow resistance, venous capacitance, and outflow resistance (hepatic vascular resistance) occur at different locations. Arterial vasoconstriction occurs at the inlet of each splanchnic organ and is primarily mediated by sympathetic release of norepinephrine which causes adrenergic vasoconstriction of the vascular smooth muscle (Gelman and Mushlin 2004; Lautt 2009). In humans, splanchnic venous capacitance resides largely within the mesenteric venous system and the liver (Pang 2001; Gelman and Mushlin 2004; Lautt 2009) which are connected in series at similar pressures by the low resistance portal vein. The hepatic and mesenteric capacitance vessels have both α1‐ and α2‐adrenergic receptors contributing to active venoconstriction but lack β2 receptors (Patel et al. 1981; Rothe 1983; Gelman and Mushlin 2004). Splanchnic outflow resistance localizes mostly to the hepatic sinusoids in humans, referred to as “intrahepatic resistance” which is strongly nitric oxide (NO) dependent (Lautt 2009; Vollmar and Menger 2009). Ordinarily intrahepatic resistance is low and maintains hepatic and mesenteric capacitances at pressures only slightly higher than inferior vena cava pressure (Greenway and Lautt 1988). Small increments in intrahepatic resistance due to NO deficiency can exert large effects on hepatic and mesenteric venous pressures which may in part explain splanchnic pooling in POTS (Medow et al. 2005; Liao et al. 2010; Stewart et al. 2011).
Therefore, intrahepatic resistance is likely increased in POTS‐Faint, resulting in pooling despite sustained splanchnic arterial vasoconstriction. In addition, both intrahepatic resistance and splanchnic arterial vasoconstriction are likely reduced in Control‐Faint, resulting in increased splanchnic blood flow and reduced splanchnic resistance. Changes in NO may play a role which holds forth the potential for treating splanchnic pooling and specific disorders of orthostatic intolerance with agents that increase NO.
In summary, we found that false positive VVS in young healthy volunteers resulted from decreased systemic vascular resistance due primarily to vasoconstrictive deficits of splanchnic vasculature with sustained cardiac output. In contrast, we found that young POTS patients have increased systemic resistance and decreased venous return and cardiac output caused by excessive venous pooling in the splanchnic vasculature.
Limitations
POTS‐Faint and Control‐Faint were identified by their fainting within 10 min of tilt. We used standard 10‐min tilt as a design feature in all our studies of POTS patients. All of our Control‐Faint patients also had 10 min studies. Increasing the time of HUT could potentially yield different information. However, the mean time to VVS in past studies of recurrent fainters was 10–11 min, and our results inform primarily on phase 2 hypotension which started before 10 min in all VVS patients.
Fiducial time markers remove the absolute time dependence of the observed phenomena while facilitating inter‐group comparison.
Impedance plethysmography of the splanchnic vasculature cannot distinguish blood pooling among the splanchnic organs (e.g., liver vs. mesenteric) and only measures fractional change in regional blood volumes and blood flows. These were validated against reference standards in prior studies (Stewart et al. 2006, 2007).
Modelflow methods yield measures of relative cardiac output. These were standardized while supine CO against inert gas rebreathing CO. Inert gas rebreathing requires deep breathing was not performed during the tilt because it would alter hemodynamics.
Go to:
Conflict of Interest
None declared.
Go to:
Notes
Stewart J. M., Shaban M. A., Fialkoff T., Tuma‐Marcella B., Visintainer P., Terilli C., Medow M. S.. Mechanisms of tilt‐induced vasovagal syncope in healthy volunteers and postural tachycardia syndrome patients without past history of syncope, Physiol Rep, 7 (13), 2019, e14148, 10.14814/phy2.14148 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Funding Information
Funding for this project was provided by grants RO1 HL 112736 and RO1 HL 134674 from the National Heart Lung and Blood Institute (NHLBI), and R21 NS 094644 from the National Institute of Neurologic Disorders and Stroke.
Go to:
References
Bogert, L. W. , and van Lieshout J. J.. 2005. Non‐invasive pulsatile arterial pressure and stroke volume changes from the human finger. Exp. Physiol. 90:437–446. [PubMed] [Google Scholar]
Charkoudian, N. , and Johnson J. M.. 1999. Altered reflex control of cutaneous circulation by female sex steroids is independent of prostaglandins. Am. J. Physiol. 276:H1634–H1640. [PubMed] [Google Scholar]
Evans, J. M. , Leonelli F. M., Ziegler M. G., McIntosh C. M., Patwardhan A. R., Ertl A. C., et al. 2001. Epinephrine, vasodilation and hemoconcentration in syncopal, healthy men and women. Auton. Neurosci. 93:79–90. [PubMed] [Google Scholar]
Fouad, F. M. , Sitthisook S., Vanerio G., Maloney J. III, Okabe M., Jaeger F., et al. 1993. Sensitivity and specificity of the tilt table test in young patients with unexplained syncope. Pacing Clin. Electrophysiol. 16:394–400. [PubMed] [Google Scholar]
Freeman, R. , Wieling W., Axelrod F. B., Benditt D. G., Benarroch E., Biaggioni I., et al. 2011. Consensus statement on the definition of orthostatic hypotension, neurally mediated syncope and the postural tachycardia syndrome. Clin. Auton. Res. 21:69–72. [PubMed] [Google Scholar]
Fu, Q. , Vangundy T. B., Shibata S., Auchus R. J., Williams A. J., and Levine B. D.. 2010. Menstrual cycle affects renal‐adrenal and hemodynamic responses during prolonged standing in the postural orthostatic tachycardia syndrome. Hypertension 56:82–90. [PMC free article] [PubMed] [Google Scholar]
Fu, Q. , Verheyden B., Wieling W., and Levine B. D.. 2012. Cardiac output and sympathetic vasoconstrictor responses during upright tilt to presyncope in healthy humans. J. Physiol. 590:1839–1848. [PMC free article] [PubMed] [Google Scholar]
Ganzeboom, K. S. , Colman N., Reitsma J. B., Shen W. K., and Wieling W.. 2003. Prevalence and triggers of syncope in medical students. Am. J. Cardiol. 91:1006–1008. [PubMed] [Google Scholar]
Gelman, S. , and Mushlin P. S.. 2004. Catecholamine‐induced changes in the splanchnic circulation affecting systemic hemodynamics. Anesthesiology 100:434–439. [PubMed] [Google Scholar]
Greenway, C. V. , and Lautt W. W.. 1988. Distensibility of hepatic venous resistance sites and consequences on portal pressure. Am. J. Physiol. 254:H452–H458. [PubMed] [Google Scholar]
Hainsworth, R. , and Al‐Shamma Y. M.. 1988. Cardiovascular responses to upright tilting in healthy subjects. Clin. Sci. (Lond.) 74:17–22. [PubMed] [Google Scholar]
Jardine, D. L. , Wieling W., Brignole M., Lenders J. W. M., Sutton R., and Stewart J.. 2018. The pathophysiology of the vasovagal response. Heart Rhythm 15:921–929. [PMC free article] [PubMed] [Google Scholar]
Kimpinski, K. , Figueroa J. J., Singer W., Sletten D. M., Iodice V., Sandroni P., et al. 2012. A prospective, 1‐year follow‐up study of postural tachycardia syndrome. Mayo Clin. Proc. 87:746–752. [PMC free article] [PubMed] [Google Scholar]
Lautt, W. W. 2009. Hepatic Circulation: Physiology and Pathophysiology. Morgan & Claypool Life Sciences, San Rafael, CA. [PubMed] [Google Scholar]
Leonelli, F. M. , Wang K., Evans J. M., Patwardhan A. R., Ziegler M. G., Natale A., et al. 2000. False positive head‐up tilt: hemodynamic and neurohumoral profile. J. Am. Coll. Cardiol. 35:188–193. [PubMed] [Google Scholar]
Lewis, D. A. , Zlotocha J., Henke L., and Dhala A.. 1997. Specificity of head‐up tilt testing in adolescents: effect of various degrees of tilt challenge in normal control subjects. J. Am. Coll. Cardiol. 30:1057–1060. [PubMed] [Google Scholar]
Liao, Y. , Chen S., Liu X., Zhang Q., Ai Y., Wang Y., et al. 2010. Flow‐mediated vasodilation and endothelium function in children with postural orthostatic tachycardia syndrome. Am. J. Cardiol. 106:378–382. [PubMed] [Google Scholar]
Low, P. A. , Opfer‐Gehrking T. L., Textor S. C., Benarroch E. E., Shen W. K., Schondorf R., et al. 1995. Postural tachycardia syndrome (POTS). Neurology 45:S19–S25. [PubMed] [Google Scholar]
Medow, M. S. , and Stewart J. M.. 2007. The postural tachycardia syndrome. Cardiol. Rev. 15:67–75. [PubMed] [Google Scholar]
Medow, M. S. , Minson C. T., and Stewart J. M.. 2005. Decreased microvascular nitric oxide‐dependent vasodilation in postural tachycardia syndrome. Circulation 112:2611–2618. [PMC free article] [PubMed] [Google Scholar]
Minson, C. T. , Halliwill J. R., Young T. M., and Joyner M. J.. 2000. Influence of the menstrual cycle on sympathetic activity, baroreflex sensitivity, and vascular transduction in young women. Circulation 101:862–868. [PubMed] [Google Scholar]
Montgomery, L. D. 1993a. Body volume changes during simulated microgravity I: technique and comparison of men and women during horizontal bed rest. Aviat. Space Environ. Med. 64:893–898. [PubMed] [Google Scholar]
Montgomery, L. D. 1993b. Body volume changes during simulated microgravity. II: comparison of horizontal and head‐down bed rest. Aviat. Space Environ. Med. 64:899–904. [PubMed] [Google Scholar]
Montgomery, L. D. , Parmet A. J., and Booher C. R.. 1993. Body volume changes during simulated microgravity: auditory changes, segmental fluid redistribution, and regional hemodynamics. Ann. Biomed. Eng. 21:417–433. [PubMed] [Google Scholar]
Moya, A. , Sutton R., Ammirati F., Blanc J. J., Brignole M., Dahm J. B., et al. 2009. Guidelines for the diagnosis and management of syncope (version 2009). Eur. Heart J. 30:2631–2671. [PMC free article] [PubMed] [Google Scholar]
Pang, C. C. 2001. Autonomic control of the venous system in health and disease: effects of drugs. Pharmacol. Ther. 90:179–230. [PubMed] [Google Scholar]
Patel, P. , Bose D., and Greenway C.. 1981. Effects of prazosin and phenoxybenzamine on alpha‐ and beta‐receptor‐mediated responses in intestinal resistance and capacitance vessels. J. Cardiovasc. Pharmacol. 3:1050–1059. [PubMed] [Google Scholar]
Plash, W. B. , Diedrich A., Biaggioni I., Garland E. M., Paranjape S. Y., Black B. K., et al. 2013. Diagnosing postural tachycardia syndrome: comparison of tilt testing compared with standing haemodynamics. Clin. Sci. (Lond.) 124:109–114. [PMC free article] [PubMed] [Google Scholar]
Raj, S. R. 2006. The Postural Tachycardia Syndrome (POTS): pathophysiology, diagnosis & management. Indian Pacing Electrophysiol. J. 6:84–99. [PMC free article] [PubMed] [Google Scholar]
Rothe, C. F. 1983. Venous system: physiology of the capacitance vessels Pp. 397–452 in Shepherd J. T., ed. The Cardiovascular System: Peripheral Circulation and Organ Blood Flow. Handbook of Physiology. American Physiological Society, Bethesda, MD. [Google Scholar]
Schondorf, R. , and Low P. A.. 1993. Idiopathic postural orthostatic tachycardia syndrome: an attenuated form of acute pandysautonomia? Neurology 43:132–137. [PubMed] [Google Scholar]
Sheldon, R. S. , Grubb B. P., Olshansky B., Shen W. K., Calkins H., Brignole M., et al. 2015. 2015 heart rhythm society expert consensus statement on the diagnosis and treatment of postural tachycardia syndrome, inappropriate sinus tachycardia, and vasovagal syncope. Heart Rhythm 12:e41–e63. [PMC free article] [PubMed] [Google Scholar]
Singer, W. , Sletten D. M., Opfer‐Gehrking T. L., Brands C. K., Fischer P. R., and Low P. A.. 2011. Postural tachycardia in children and adolescents: what is abnormal? J. Pediatr. 160:222–226. [PMC free article] [PubMed] [Google Scholar]
van Steenwijk, C. J. , Wieling W., Johannes J. M., Harms M. P., Kuis W., and Wesseling K. H.. 1995. Incidence and hemodynamic characteristics of near‐fainting in healthy 6‐ to 16‐year old subjects. J. Am. Coll. Cardiol. 25:1615–1621. [PubMed] [Google Scholar]
Stewart, J. M. , and Montgomery L. D.. 2004. Regional blood volume and peripheral blood flow in postural tachycardia syndrome. Am. J. Physiol. Heart Circ. Physiol. 287:H1319–H1327. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Medow M. S., Glover J. L., and Montgomery L. D.. 2006. Persistent splanchnic hyperemia during upright tilt in postural tachycardia syndrome. Am. J. Physiol. Heart Circ. Physiol. 290:H665–H673. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Montgomery L. D., Glover J. L., and Medow M. S.. 2007. Changes in regional blood volume and blood flow during static handgrip. Am. J. Physiol. Heart Circ. Physiol. 292:H215–H223. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Nafday A., Ocon A. J., Terilli C., and Medow M. S.. 2011. Cutaneous constitutive nitric oxide synthase activation in postural tachycardia syndrome with splanchnic hyperemia. Am. J. Physiol. Heart Circ. Physiol. 301:H704–H711. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Suggs M., Merchant S., Sutton R., Terilli C., Visintainer P., et al. 2016. Postsynaptic alpha1‐adrenergic vasoconstriction is impaired in young patients with vasovagal syncope and is corrected by nitric oxide synthase inhibition. Circ. Arrhythm. Electrophysiol. 9(8):e003828. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Medow M. S., Sutton R., Visintainer P., Jardine D. L., and Wieling W.. 2017a. Mechanisms of vasovagal syncope in the young: reduced systemic vascular resistance versus reduced cardiac output. J. Am. Heart Assoc. 6:1–11. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Sutton R., Kothari M. L., Goetz A. M., Visintainer P., and Medow M. S.. 2017b. Nitric oxide synthase inhibition restores orthostatic tolerance in young vasovagal syncope patients. Heart 103:1711–1718. [PMC free article] [PubMed] [Google Scholar]
Stewart, J. M. , Boris J. R., Chelimsky G., Fischer P. R., Fortunato J. E., Grubb B. P., et al. 2018. Pediatric disorders of orthostatic intolerance. Pediatrics 141:1–13. [Google Scholar]
Sutton, R. , and Brignole M.. 2014. Twenty‐eight years of research permit reinterpretation of tilt‐testing: hypotensive susceptibility rather than diagnosis. Eur. Heart J. 35:2211–2212. [PubMed] [Google Scholar]
Taneja, I. , Medow M. S., Glover J. L., Raghunath N. K., and Stewart J. M.. 2008. Increased vasoconstriction predisposes to hyperpnea and postural faint. Am. J. Physiol. Heart Circ. Physiol. 295:H372–H381. [PMC free article] [PubMed] [Google Scholar]
Verheyden, B. , Liu J., van Dijk N., Westerhof B. E., Reybrouck T., Aubert A. E., et al. 2008. Steep fall in cardiac output is main determinant of hypotension during drug‐free and nitroglycerine‐induced orthostatic vasovagal syncope. Heart Rhythm 5:1695–1701. [PubMed] [Google Scholar]
Vollmar, B. , and Menger M. D.. 2009. The hepatic microcirculation: mechanistic contributions and therapeutic targets in liver injury and repair. Physiol. Rev. 89:1269–1339. [PubMed] [Google Scholar]
Wieling, W. , Krediet C. T., van Dijk N., Linzer M., and Tschakovsky M. E.. 2007. Initial orthostatic hypotension: review of a forgotten condition. Clin. Sci. (Lond.) 112:157–165. [PubMed] [Google Scholar]
Wieling, W. , Van Dijk N., De Lange F. J., Olde Nordkamp L. R., Thijs R. D., Van Dijk J. G., et al. 2015. History taking as a diagnostic test in patients with syncope: developing expertise in syncope. Eur. Heart J. 36:277–280. [PubMed] [Google Scholar]
Wieling, W. , Jardine D. L., de Lange F. J., Brignole M., Nielsen H. B., Stewart J., et al. 2016. Cardiac output and vasodilation in the vasovagal response: an analysis of the classic papers. Heart Rhythm 13:798–805. [PMC free article] [PubMed] [Google Scholar]
Articles from Physiological Reports are provided here courtesy of Wiley-Blackwell
OTHER FORMATS
PDF (1.2M)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 1517898 | https://sv.wikipedia.org/wiki/Propranolol | Propranolol | Propranolol är en antihypertensiv medicin som agerar på kroppens noradrenerga system. Den minskar aktiviteten hos kroppens sympatiska nervsystem genom att blockera adrenerga beta-receptorer, vilket bland annat leder till sänkt blodtryck.
Propranolol skrivs framförallt ut för att behandla sjukdomar i kardiovaskulära systemet, till exempel hjärtinfarkt, högt blodtryck, angina pectoris, men även för giftstruma, migrän, tumörer i binjuremärgen och tremor. Den har även använts experimentellt mot posttraumatiskt stressyndrom, där den verkar minska symptomen på tillståndet.
Eftersom betablockerare sänker blodsockret, kan propranolol ge hypoglykemi. Det kan också förvärra obstruktiva lungsjukdomar och sänka pH-värdet i blodet. Ökad drömaktivitet och mardrömmar förekommer vid behandling med propranolol. Dess påverkan på blodkärlen kan leda till vita fingrar. Agranulocytos är en allvarlig biverkning som kan uppkomma.
Medicinsk användning
Propranolol används för att behandla olika medicinska tillstånd, såsom:
Kardiovaskulär
Hypertoni
Kärlkramp
Hjärtinfarkt
Takykardi
Portahypertension
Förebygga blödningar och ascites hos Esofagusvaricer
Ångest
Tidigare var propanolol en av de bästa behandlingar för hypertoni. Eftersom de inte kunde ge lika bra effekt som andra mediciner, särskilt bland de äldre, blev beta-blockeraren nedgraderad till fjärde-klass i juni 2006 i Storbritannien. Fler och fler bevis hos de mest frekvent använda beta-blockerare vid normal dosering har även påvisat att öka risken för Typ 2-diabetes.
Psykiatri
Propranolol används ibland till för att behandla scenskräck. Dock finns inte lika bra bevis på att behandla andra ångeststörningar. De utförda experiment i andra psykiatriska områden:
Posttraumatiskt stressyndrom (PTSD) och specifika Fobier
Förbättringar av sociala förmågor hos personer med Autismspektrumstörning
Aggressivt beteende av patienter med Hjärnskada
Behandling för överdrivet drickande av vätskor som orsakas av Polydipsi
Syntes
Propranolol framställs ur 1-naftol och epiklorhydrin. I första steget öppnas epoxid-ringen upp av nukleofil-attack från OH-gruppen i 1-naftol, samtidigt sker 2 stegs protonöverföring. Vätet som sitter på -OH hamnar nu på epoxidsyret. I det andra steget sker en SN2-reaktion, i det här fallet, en nukleofil substitution av isopropylamin på det kolet som binder till den hyfsat bra lämnande grupp -Cl. Därav bildas den önskade slutprodukten - propranolol.
Man kan även använda sig av en stark bas som NaOH i det undre steget. Den starka basen NaOH hjälper till att deprotonera vätet som är bunden till OH-gruppen i 1-naftol, samtidigt hjälper den till att stabilisera den bildande alkoxid. Den nya natrium-1-naftolat intermediat som har bildats är en bra nukleofil. På samma sätt kommer den att attackera kolet som binder till -Cl på epiklorhydrin och en ny mellanprodukt bildas. Därefter görs en nukleofil-attack av isopropylamin på det elektrofila kolet som sitter på epoxidsyret. Vilket resulterar i en ringöppning och syret protoneras till en alkohol, därav fås slutprodukten - propranolol.
Propranolol är en racematisk blandning – det vill säga, två enantiomerer, R(+) och S(-). S(-)-enantiomeren är ungefär 100 gånger bättre än R(+)-enantiomeren på att blockera adrenerga beta-receptorer.
Syntesen ovan (1) visar ett generellt och billigt sätt att syntetisera propranolol. Men för att undvika racemat har forskare behövt hitta andra sätt. Som till exempel syntesen nedan (2) som är steoreoselektiv, vilket betyder att det mesta av produkten blir S(-)-propranolol.
(I) K2CO3, CH2=CHCH2Br, acetone, reflux, 12 h; 97-99%;
(II) cat-OsO4, (DHQD)2-PHAL, K3Fe(CN)6, K2CO3, t -BuOH:H2O, 0 °C, 12 h, 94-98%; 73-90% ee;
(III) SOCl2, Et3N, CH2Cl2, 0 °C, 40 min.; 96-99%;
(IV) cat. RuCl3.3H2O, NaIO4, CH3CN:H2O, 0 °C, 30 min., 94-98%;
(V) LiBr, THF 25 °C, 2-3 h;
(VI) 20% H2SO4, Et2O, 25 °C, 10 h;
(VII) K2CO3, MeOH, 0 °C, 2 h, 80-85%;
(VIII) iPr-NH2, H2O (cat.), reflux, 2 h, 99%.
Referenser
Noter
Källor
Inderal produktinformation på FASS
Data sources include Micromedex® (updated Jan 4th, 2017), Cerner Multum™ (updated Jan 10th, 2017), Wolters Kluwer™ (updated Jan 6th, 2017) and others
Betablockerare | swedish | 0.588656 |
fainting_mechanism/PMC4499525.txt | Skip to main content
U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now.
Search PMC Full-Text Archive
Search PMC Full-Text Archive
Search in PMC
Advanced Search User Guide
Journal List World J Diabetes v.6(7); 2015 Jul 10 PMC4499525
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with, the contents by NLM or the National Institutes of Health.
Learn more: PMC Disclaimer | PMC Copyright Notice
Logo of wjd
World J Diabetes. 2015 Jul 10; 6(7): 912–926.
Published online 2015 Jul 10. doi: 10.4239/wjd.v6.i7.912
PMCID: PMC4499525
PMID: 26185599
Mechanisms of hypoglycemia unawareness and implications in diabetic patients
Iciar Martín-Timón and Francisco Javier del Cañizo-Gómez
Author information Article notes Copyright and License information PMC Disclaimer
Go to:
Abstract
Hypoglycemia unawareness (HU) is defined at the onset of neuroglycopenia before the appearance of autonomic warning symptoms. It is a major limitation to achieving tight diabetes and reduced quality of life. HU occurs in approximately 40% of people with type 1 diabetes mellitus (T1DM) and with less frequency in T2DM. Though the aetiology of HU is multifactorial, possible mechanisms include chronic exposure to low blood glucose, antecedent hypoglycaemia, recurrent severe hypoglycaemia and the failure of counter-regulatory hormones. Clinically it manifests as the inability to recognise impeding hypoglycaemia by symptoms, but the mechanisms and mediators remain largely unknown. Prevention and management of HU is complex, and can only be achieved by a multifactorial intervention of clinical care and structured patient education by the diabetes team. Less know regarding the impact of medications on the development or recognition of this condition in patients with diabetes. Several medications are thought to worsen or promote HU, whereas others may have an attenuating effect on the problem. This article reviews recent advances in how the brain senses and responds to hypoglycaemia, novel mechanisms by which people with insulin-treated diabetes develop HU and impaired counter-regulatory responses. The consequences that HU has on the person with diabetes and their family are also described. Finally, it examines the evidence for prevention and treatment of HU, and summarizes the effects of medications that may influence it.
Keywords: Hypoglycemia unawareness, Impaired awareness of hypoglycemia, Hypoglycemia associated autonomic failure, Diabetes mellitus, Counter-regulation
Core tip: This review describes novel mechanisms by which people with insulin-treated diabetes develop hypoglycemia unawareness (HU), the consequences that HU has on the person with diabetes and their family, the evidence for prevention and treatment of HU, and the effects of medications that may influence it.
Go to:
INTRODUCTION
Hypoglycemia is usually defined as a plasma glucose level < 70 mg/dL (3.9 mmol/L)[1]. Since the brain is permanently dependent on glucose, strong counter-regulatory mechanisms exists to quickly increase glucose levels to protect the human body from the negative consequences of hypoglycemia. Counter-regulatory response to hypoglycemia (Figure (Figure1)1) includes inhibition of the endogenous insulin secretion and stimulation of glucagon, catecholamines (norepinephrine, epinephrine), cortisol and growth hormone secretion, which all together stimulate hepatic glucose production and cut down glucose utilization in peripheral tissues, increasing in this way plasma glucose levels. As glycaemia comes down, the activation of the autonomic nervous system leads to neurogenic symptoms (palpitations, sweating, hunger, anxiety, tremors, etc.), which allows the perception of hypoglycaemia (hypoglycaemia awareness) (Figure (Figure22).
An external file that holds a picture, illustration, etc.
Object name is WJD-6-912-g001.jpg
Figure 1
Counter-regulatory response to hypoglycemia.
An external file that holds a picture, illustration, etc.
Object name is WJD-6-912-g002.jpg
Figure 2
Symptoms and signs associated with progressive hypoglycemia. ACTH: Adrenocorticotropic hormone; GH: Growth hormone.
Hypoglycemia unawareness (HU) is defined as the onset of neuroglycopenia before the appearance of autonomic warning symptoms[2] or as the failure to sense a significant fall in blood glucose below normal levels[3]. In patients with type 1 (T1DM) or type 2 diabetes mellitus (T2DM), recurrent hypoglycemia has been shown to reduce the glucose level that precipitates the counter-regulatory response necessary to restore euglycemia during a subsequent episode of hypoglycemia[4,5].
HU was observed in 40% T1DM patients[6] and less frequently in T2DM patients with low C-peptide levels. The presence of HU increases the risk of severe hypoglycaemia (six-fold for T1DM[7] and 17-fold for T2DM[8]). HU is more common in individuals with longer duration of diabetes, history of recent and/or recurrent hypoglycaemic events, patients with intensive glycemic therapy and in advanced age[9].
Presently, the major risk factors for the development of HU are duration of the disease and improved metabolic control. The severity of HU was associated with longer diabetes duration and with a history of frequent low glycemic levels[6], whereas aging and the blood glucose decreasing rate using professional continuous glucose monitoring systems (CGMS), which falls from near blood glucose level, were risk of severe HU[10]. Data from Pittsburgh Epidemiology of Diabetes Complications[11] showed that diabetes duration, HbA1c and intensive insulin therapy predicted HU in men, whereas severity and frequency of hypoglycemia, QTc interval and hypertension predicted HU in women. Thus, women are more likely to have HU, which unlike in men, is also marginally related to hypertension, QTc interval and hypoglycemia. On the other hand, in patients with T1DM, HU was 3.4-fold more common among patients homozygous for Gly16 than among patients with other variants of the Arg16Gly polymorphism, so that T1DM patients who carry two alleles of the Gly16 variant of ADRB2 are at increased risk of developing HU[12]. Finally, in both T1 and T2DM patients with impaired HU, hypoglycemia-induced electroencephalogram changes, such increased theta band amplitude, were not affected by antecedent of hypoglycemia[13].
This article reviews recent advances in how the brain senses and responds to hypoglycemia, novel mechanisms by which people with insulin-treated diabetes develop HU and impaired counter-regulatory responses. The consequences that HU had on the person with diabetes and their family is also described. Finally, it examines the evidence for prevention and management of HU, and summarizes the effects of medications that may influence it.
Go to:
MECHANISMS OF HU
Aberrant glucose counter-regulation (as a result of a failure in the reduction of insulin production and an increase in glucagon release), and HU (as the result of an attenuated increase in sympathoadrenal activity) are the components of hypoglycemia-associated autonomic failure (HAAF) in diabetics patients. HAAF is most often caused by recent/recurrent iatrogenic hypoglycemia, and indeed HAAF is maintained by recurrent hypoglycemia[14,15] (Figure (Figure33).
An external file that holds a picture, illustration, etc.
Object name is WJD-6-912-g003.jpg
Figure 3
Recurrent hypoglycemia leads to cellular adaptation and hypoglycemia-associated autonomic failure. HAAF: Hypoglycemia-associated autonomic failure.
Diverse causes of HAAF and HU in diabetes[16]
Catecholamines: Previous hypoglycemia leads to a blunted catecholamine response to a following episode of hypoglycemia. These has been demonstrated in several studies; for example Ramanathan et al[17] showed that intravenous infusion of adrenergic blockers on one day of a hypoglycemia prevent the counter-regulatory failure in the response on the next day of hypoglycemia. This study implicates that HAAF needs a previous hypoglycemia (with its sympathoadrenal responses). If we use this hypothesis to think in a possible pharmacologic treatment, we can concluded that blocking the action of catecholamines we can limit the development of HAAF and protect against subsequent hypoglycemias; but unfortunately, blocking the action of catecholamines in periphery we would tend to an increase in the severity of hypoglycemia. We would need to develop a selective adrenergic receptor modulators that favourably change central nervous system response without modify the beneficial peripheral effects of the sympathoadrenal response.
Sleep: Sleep is a peripheral mediator of HAAF linked with catecholamine response. Patients with T1DM, while they are sleeping, they have a significantly decreased epinephrine response to hypoglycemia[18], and also a reduced awakening from sleep during hypoglycemia[19]. So, because of the HU and the impaired adrenomedullary response, we can explain some of the overnight deaths of healthy young people with T1DM.
Cortisol: Hypoglycemia is associated with an elevation in systemic corticosteroids, and this has been proposed to feedback to the hypothalamus contributing to HAAF[20-22]. However it remains controversial if the endogenous hypercortisolemia is of sufficient magnitude to blunt de counter-regulatory response to hypoglycemia[23,24]. It have been shown that corticotrophin releasing hormone agonist impair the counter-regulatory response to a subsequent hypoglycemia, suggesting a possible role in HAAF[25].
Opioids: Preclinical and clinical studies with opioids demonstrated a rise in endogenous opioids during hypoglycemia, for example naloxone (an opioid receptor blocker), increased the sympathoadrenal response to hypoglycemia, and when is infused during previous hypoglycemia, it prevent HAAF[26,27]. Hence there is a potential therapeutic function for opioid receptor blockade to protect against HAAF.
Exercise: The inability to reduced circulating insulin during exercise, lead T1DM patients, at an increased risk for hypoglycemia during or after exercise. In addition to, during exercise the opioid beta endorphin is released to activate the sympathoadrenal response. In a recent study, healthy individuals who exercised and elevated endorphin levels, they had reduced catecholamine response during hypoglycemia in the next day[28], suggesting that endogenous opioids, again, play a role in HAAF, and that blocking their action may protect against exercise-autonomic failure.
Recurrent hypoglycemia and HU
Clinically HAAF can be viewed as both, maladaptative or adaptative response[29]. At one end, patients with T1DM and HU make tests of cognitive function during hypoglycemia better than patients with HU. Additionally, the time necessary for complete cognitive recovery after restoration of normoglycemia is faster in patients who have HU[30]. HAAF in humans may be similar than in rats; rats with recurrent moderate hypoglycemia had less brain cell death[31] and less mortality during or following marked hypoglycemia than those without recurrent hypoglycemia. On the other hand, HAAF is without doubt a maladaptive response if we consider that defective glucose counter-regulation and HU rise the risk of severe hypoglycemia with its morbidity and potential mortality[32].
Although it is well established that recurrent hypoglycemia leads to HU, the mechanism responsible for this are unknown. Several current mechanistic hypotheses are discussed below.
The brain glucose transport or glucose metabolism hypothesis: Several studies have identified specific brain regions that exhibit decrease glucose uptake. In diabetic patients with and without HU, the effects of acute moderate hypoglycemia and the condition of HU on regional brain uptake of the labeled glucose analog [(18)F]fluorodeoxyglucose (FDG) using positron emission tomography were examined[33,34]. In the group with hypoglycemia awareness, there was an increase in the normalized FDG uptake in a subthalamic brain region[33], in left amygdale and in bilateral ventral striatum[34] in response to hypoglycemia; whereas in the group with HU the uptake in these brain regions fell significantly[33,34]. Reduced responses in these brain regions in HU, suggest habituation of higher behavioral responses to hypoglycemia as a basis for unawareness, and demonstrated a change in its metabolic function associated with the failure to trigger a counter-regulatory response. On the other hand, in subjects with T1DM and HU a positive correlation was observed between thalamic response and epinephrine response to hypoglycemia, suggesting that this brain region may be involved in the coordination of the counter-regulatory response to hypoglycemia[35]. During recurrent hypoglycemia, cerebral blood flow reduced significantly in the thalamus and hypothalamus of T1DM subjects, compared to healthy controls[36], suggesting that there is reduced neuronal activation in these brain regions that participate in glucose sensing and/or coordination of counter-regulation response in subjects with T1DM that likely contributes to the development of HU.
It has been hypothesized that recurrent hypoglycemia leads to HU through an alteration in the glucose transport or metabolism. Altered glucose transport or metabolism as a cause of HU is less substantiated in humans. Subjects with T1DM and HU had significantly higher brain glucose concentrations compared to that in controls under the same conditions[37]. These date suggest that changes in brain glucose transport or metabolism may occur as a result of recurrent hypoglycemia.
The brain glycogen supercompensation hypothesis: It has been hypothesized that increased brain glycogen contributes to the development of HU and impaired sympathoadrenal responses by providing energy for the brain during periods of systemic hypoglycemia. Experimental studies and in humans have shown that after one or more episodes of hypoglycemia, increased glycogen content in the brain[38,39]. Subsequent studies indicated lower glycogen content in brain of humans with T1DM, implying that glycogen supercompensation does not contribute to the development of HU[40]. The most important question to resolve is whether changes to brain glucose levels, physiologically or pharmacologically induced, may provide people who suffer from recurrent hypoglycemia a therapeutic benefit to preserve both the sympathoadrenal response and HU.
The brain fuel hypothesis: When there is a decrease in the supply of glucose from the periphery, the brain may be able to keep your metabolic processes by increasing uptake of alternative carbon fuels such as lactate or ketones. Plasma lactate concentrations are approximately tenfold higher than those of acetate, making it a primary candidate as an alternative brain fuel during hypoglycemia. On the other hand, increased of blood-brain barrier monocarboxylic acid (MCA) transport and metabolism among T1DM individuals with HU may be a mechanism to supply the brain with non-glucose fuels during episodes of acute hypoglycemia and may contribute to the maintenance of brain energetic during hypoglycemia and to the syndrome of HU, independent of diabetes[41]. Finally, in T1DM patients with HU, upregulation of the MCA transporter promotes increased brain lactate uptake[42].
The brain neuronal communication hypothesis: Neuronal communication relies on the release of classical neurotransmitters, such as Gamma-Aminobutyric Acid (GABA), a potent inhibitory neurotransmitter. GABA levels in ventromedial hypothalamus (VMH) interstitial fluid are decreased during acute hypoglycemia[43]. Recurrent hypoglycemia leads to a significant increase in VMH GABA concentrations[44], that fail to decrease normally during subsequent hypoglycemia, and which correlates with the reduced glucagon and epinephrine responses[45]. These data suggest that recurrent hypoglycemia results in increased VMH GABA inhibitory tone, and that altered GABA tone may be an important common mediator in the development of HAAF, especially in diabetic patients.
Go to:
CONSEQUENCES OF HU
Consequences of HU on morbidity, mortality, and cardiovascular outcomes
People who have HU have a much greater risk of severe hypoglycemia, up to six fold, with its attendant morbidity[46,47]. HU may result in many serious forms of morbidity including seizure, coma, fractures and joint dislocation and cardiac arrhythmias, and is occasionally fatal.
Severe episodes of hypoglycemia or HU requiring the assistance of another have been shown to be associated with an increased risk of mortality in both the Action to Control Cardiovascular Risk in Diabetes (ACCORD)[48] and the Action in Diabetes and Vascular Disease[49] studies. On the other hand, post hoc analysis of the ACCORD study cohort, to examine the relationship between frequent and unrecognized hypoglycemia and mortality, 10096 ACCORD study participants were included. In this study, recognized and unrecognized hypoglycemia was more common in the intensive group than in the standard group; and in the intensive group, a small but statistically significant inverse relationship was identified between the number of hypoglycemic episodes and the risk of death among participants[50]. This latter finding does not mean that we should change our clinical practice and include frequent episodes of hypoglycemia in the targets of T2DM patients and cardiovascular risk factors. Instead, we must strive to achieve optimal glycemic control in our patients, without episodes of hypoglycemia.
Consequences of HU on adults with T1DM
Several prospective studies as the Diabetes Control and Complications Trial[51] and the Stockholm Diabetes Intervention Study[52] suggests that cognitive function does not deteriorate in patients with T1DM who suffer recurrent hypoglycemia, at least less than 10 years of these studies.
Gold et al[53] to compare the degree of cognitive dysfunction experienced by T1DM patients who had normal awareness of the onset of hypoglycemia with patients who had history of impaired awareness of hypoglycemia, found that T1DM patients with HU exhibited more profound cognitive dysfunction during acute hypoglycemia which persisted for longer following blood glucose recovery. Intellectual activity is likely to be affected and cause sub-optimal performance during this recovery period. Recent investigations with advanced imaging techniques have demonstrated that adults with T1DM appear to call upon a greater volume of the brain to perform a working memory task during hypoglycemia[54]. These findings suggest that adults with T1DM must recruit more regions to preserve cognitive function during hypoglycemia than adults without the disease.
Evidence of clinical audit in T1DM patients with intensive insulin therapy with HU showed that these patients had less adhesion to changes in insulin regimens to compare them with patients with hypoglycemia awareness, despite the observed increase in clinical contacts[55]. Neuroimaging studies have shown that patients with HU showed a reduced activation in appetitive motivational networks associated with integrated behavioral responses to hypoglycemia[34]. This may suggest that in some patients with HU behavioral strategies are more important than educational strategies; however treatment of HU will require a combination of both strategies, behavioral and educational, along with the use of technology, such as therapy with continuous insulin pump and online glucose monitoring[56].
Consequences of HU in children and adolescents with T1DM
A significant proportion of children and adolescents with T1DM have HU. Screening for HU is an important component of routine diabetes care and can identify patients at increased risk of severe hypoglycemic events[57]. The youngest patients are most vulnerable to the adverse consequences of hypoglycemia. Ongoing maturation of the central nervous system puts these children at greater risk for cognitive deficits as a consequence of HU[58]. HU is a significant problem for children and adolescents with T1DM and the major risk factor for development of hypoglycemia[57]. Those children with T1DM diagnosed before age of 6, who suffer repeated and severe episodes of hypoglycemia may have more increased range of cognitive dysfunction, brain abnormalities[59], structural brain changes[60], lower mental abilities latter on in life, and behavior problems than those who do not have HU until latter[61,62].
Consequences of HU on subjects with T2DM
HU is less common in T2DM patients. Two retrospective surveys of subjects with insulin-treated T2DM showed that only 8% and 9.8% respectively had HU estimated by a validated scoring system[8,46]. However, in the patients with HU the incidence of severe hypoglycemia was nine-fold and 17-fold higher respectively than those with normal hypoglycemia awareness[8,46]. In several studies, using continuous monitoring system, asymptomatic hypoglycemia was detected in 47%[63] and 56%[64] of subjects with T2DM, treated with different treatment regimes. These findings suggest that HU may be more prevalent in T2DM than is appreciated.
Severe hypoglycemia, due to HU, was associated in T2DM patients with cardiovascular and neurological complications[1,48]. In patients with T2DM and coronary artery disease, severe hypoglycemia was associated with ischemic electrocardiogram changes and chest pain, and may account for sudden mortality[65,66]. In a retrospective study in T2DM subjects, the patients who experienced outpatient severe hypoglycemia were also shown to have a 79% higher odds ratio of experiencing acute cardiovascular events than patients without severe hypoglycemia[67]; and a case-control study in patients with T2DM showed a 65% increase in the odds of myocardial infarction with severe hypoglycemia within the previous two weeks; the risk of myocardial infarction persisted elevated for up to six months following a hypoglycemic event[68].
Behavioral changes, cognitive impairment, seizures, coma and a mortality rate estimated at between 4.9% and 9% are well-known neurological complications of severe and prolonged hypoglycemia secondary to HU[69-71]. Severe hypoglycemia secondary to HU can cause neuronal cell death and may damage regions of the brain that oversea memory, especially in older people with T2DM[72].
Finally, a frequently problem in T2DM is nocturnal hypoglycemia. Undetected nocturnal hypoglycemia often contributes to HU. Nocturnal hypoglycemia has been associated with cardiac arrhythmias resulting in sudden death[73].
Consequences of HU on the elderly
Patients in the older age-groups are especially vulnerable to HU. Aging modifies the cognitive, symptomatic, and counter-regulatory hormonal responses to hypoglycemia[74]. Older adults with diabetes are at much higher risk for the geriatric syndrome, which includes falls, incontinence, frailty, cognitive impairment and depressive symptoms[75]. In the elderly subjects, episodes of severe hypoglycemia are more likely to be followed by changes in the blood brain circulation which may further increase the risk of neurological damage in this population[76,77]. In older patients with T2DM, Whitmer et al[72] found a significant association between the number of severe hypoglycemic episodes and dementia; with ≥ 3 episodes almost doubling the risk more episodes of severe hypoglycemia secondary to HU had increasing likelihoods of being subsequently diagnosed with dementia. Another authors also found an association between severe hypoglycemia and cognitive impairment in these patients[78]. These reports suggest that severe hypoglycemia and HU in older people with diabetes may be associated with cognitive decline[79].
Consequences of HU during pregnancy
Pregnancy is associated with a high risk of severe hypoglycemia in diabetic subjects. History of HU has been documented as risk factors of severe hypoglycemia during pregnancy[80-82]. Reduced sympathoadrenal responses during hypoglycemia may contribute to defective glucose counter-regulation and HU[83,84]. In pregnant woman severe hypoglycemia episodes and HU occur three to five times more frequently in first trimester than third trimester when compared with the incidence in the year preceding the pregnancy[80,81,85] and may lead to severe morbidity and even death[86].
Consequences of HU on quality of life and social impact
Hypoglycemia and HU are associated with significant reductions in quality of life measures in both T1DM and T2DM patients[87-89]. The wellbeing of patients may be affected both from the effects of hypoglycemia and from fear of recurrence[89,90]. A positive association was found between severity and/or frequency of hypoglycemic events and greater fear of hypoglycemic episodes[71]. As a result fear of hypoglycemia makes the patients to promote compensatory behaviors in a way to have less episodes of hypoglycemia such as decreased insulin doses resulting in negative glycemic control, and an increased risk of serious health consequences[91]. Patients with recurrent hypoglycemia and HU were more likely to have a lower quality of life in several parameters including depression and anxiety[89,92,93], increased pain and limitations in mobility and usual activities[89], and decline in the quantity and quality of sleep[94]. On the other hand, young adults with T1DM reported the presence of interpersonal conflict, and difficulty talking about issues related to hypoglycemia with significant others[95], that may carry over to their work life, where hypoglycemia has been linked to reduced productivity[88].
Despite that many countries require documentation that severe hypoglycemia and HU is not occurring before persons with diabetes are permitted to have a license to operate a motor vehicle; HU has not consistently been associated with an increased risk of car collisions[96-98].
Consequences of HU on family members
In the subjects with diabetes, HU can have a profound impact on the lives of their family members, and are often reliant on immediate relatives or partners to detect and treat hypoglycemia episodes. A recent study based in-depth interviews with 24 adult family members of persons with T1DM and HU, showed that family members restricted their own lives in order to help the person with HU to detect and treat hypoglycemia[99]. In this study, some family members of people with HU, report that they are afraid of their partners, during episodes of hypoglycemia because of their aggressive behavior and their personality changes, making it difficult managing their treatment. The study showed that family members of patients with HU restricted their own lives in order to help the person with HU to detect and treat hypoglycemia, and felt anxious about the safety of the person with HU; which sometimes leads family members to neglect their own health, leading to resentment over time[100]. On the other hand, personality changes during hypoglycemia events of the person with diabetes, such as aggression, also caused, in some family members, physical fear of your partner or relative, and made treatment difficult. Family members emphasized that there is an unmet need for information and emotional support for caregivers, and the researchers suggest that proactive support for the families of patients with diabetes and HU should be considered and provided by healthcare professionals[99].
Psychological consequences of HU
The psychological consequences of HU include subsequent fear to hypoglycemia, and secondary poor treatment compliance, increased anxiety and decreased levels of satisfaction and happiness. Fear of hypoglycemia will be a barrier to achieving good glycemic control. The hypoglycemia fear survey (HFS) used to measure behaviors (HFS-B) and worries (HFS-W) related to hypoglycemia in adults with T1DM, such as maintaining higher blood glucose levels than recommended, and limiting exercise or physical activity, or concerns may have about hypoglycemic episodes, such as nocturnal episodes; have been shown to be significantly higher in women than in men and among patients who have experienced severe hypoglycemia in the past compared with those that have not[100]. If patients experience repeated severe hypoglycemic events, both the patient’s and the physician’s subsequent treatment policy are affected. In one study that reviewed hospital records and examined daily insulin doses and HbA1c levels before and after and episode of severe hypoglycemia in patients with insulin-dependent diabetes, it was found that, in 69% of these cases, either the physician or patient or both decreased the daily insulin dose. Furthermore, physicians decreased the insulin dose in a third of patients in whom the cause of hypoglycemia was preventable and due to a cause other than erroneous administration of excess insulin[101].
Economic consequences of HU
The economic consequences of severe hypoglycemia events and HU in patients with diabetes are higher than that of a mild episode and have been examined in a number of studies in Europe and United States[102-105]. Reported costs of a severe hypoglycemic event varied from approximately $80 to $5000, depending on the requirement for resources including hospitalization, emergency services, healthcare professionals and diagnostic test.
A United Kingdom study estimated the total cost of emergency treatments of 244 episodes of severe hypoglycemia in 160 patients with T1DM and T2DM over the course of one year. The total cost was approximately £92078 (£400 per episode)[102]. On the other hand, in a Swedish study the total cost (direct and indirect) of severe hypoglycemia in T2DM patients was between $12.90 and $14.10 for one month period[90].
An analysis of several United States studies, the estimated annual total cost attributable to severe hypoglycemia was between $1400 and $1500[106]. In this analysis the estimated work days lost per hypoglycemic event was between 0.22 and 6.60 d[103]. A recent study estimated that in patients with diabetes who experienced severe hypoglycemia, the lost of productivity ranged from $15.26 to $93.47 per severe hypoglycemic event, representing 8.3-15.9 h of lost work time per month[106]. Among the patients who experimented a severe hypoglycemic event at work, 18.3% missed work for a mean duration of 9.9 h, whereas the patients who had severe hypoglycemic event outside working hours, 22.7% arrived late for work or missed a full day[104]. If the hypoglycemia has occurred during the night, the number of working hours lost increased to 14.7 h[104].
Go to:
PREVENTION AND MANAGEMENT OF HU
Prevention of HU
Prevention of HU is an important part of modern day intensive diabetes therapy. To prevent HU, the goal is the complete avoidance of hypoglycemia, which is very difficult to achieve[105]. Blood glucose monitoring, individualized targets and educational programs are important in the bid to prevent and manage HU.
Blood glucose monitoring: CGMS, that can detect hypoglycemia, represents an important technological advance on the methods used for self-monitoring of blood glucose, and they are welcome to both patients and clinicians[106]. The ability of CGMS systems is to advise patients when glucose levels fall too low or rise too high, and has the potential to reduce de duration of hypoglycemia and hyperglycemia events[107,108]. Also, CGMS can be used for objective detection of patients with HU[109]. In adult patients with long-standing T1DM, a fasting level of C peptide of ≤ 0.6 ng/mL, and a HbA1c ≤ 9%, hypoglycemic episodes with a duration more than 90 minutes detected by CGMS, identified patients who had HU with an 88% specificity and 75% sensitivity[109]. On the other hand, the epinephrine response to hypoglycemia in adolescents patients with T1DM with HU was greater after the use of real-time CGMS with low glucose alarms than with standard medical therapy alone[110]. This suggests that real-time CGMS is a useful clinical tool to improve HU in adolescents with T1DM[110]. Choudhary et al[111] assessed the effect of CGMS on the frequency of severe hypoglycemia episodes, using the Gold scoring method[46] in 35 people with T1DM who have HU, via retrospective audit. A significant decline was observed in the mean rate of severe hypoglycemia (8.1 to 0.6 events per year) and also in HbA1c level (8.1% to 7.6%), between its initiation and the end of the 1-year follow-up period; while the mean Gold score did not change significantly[111]. These results support previous reports that CGMS can lower the incidence of severe hypoglycemia in patients with T1DM and HU, with no impact on the severity of HU over a 1-year period. A randomized cross-over study to assess the effects of CGMS use on glycemic levels and quality of life in patients with T1DM and HU, using the change in the Gold scoring as one of the secondary endpoints, is currently in progress and the results will not be available until 2015[112].
The impact of closed-loop CGMS, which link CGM technology with insulin pumps, whereby insulin infusion is programmed to stop automatically when glucose levels drop below a pre-determined glycemic threshold, on reducing the incidence of hypoglycemia events appears to be limited and so their usefulness in improving HU is debatable[16].
Individualized targets: In diabetic patients with HU blood glucose targets should be relaxed but not abandoned. Appropriate targeting of plasma glucose may help patients and practitioners achieve HbA1c goals, reduce excessive self-testing and minimize the occurrence of severe hypoglycemic events[113]. Glycemic goals should be individualized with some degree of safety particularly for patients with long duration of diabetes, patients who have a high risk of HU and severe hypoglycemia development, and/or subjects with multiple co-morbidities[114,115]. Basically, an HbA1c goal of less than 7% remains recommended, but is there a safe range for HbA1c? In patients with T1DM undertaking insulin therapy, the rates of severe hypoglycemia were increased among those with HbA1c < 6% and therefore it was suggested that using current therapy, an HbA1c of between 6%-7% represents the best compromise between the risk of severe hypoglycemia and that of developing microvascular complications[116].
Educational programs: The central objective of a hypoglycemia-reversal program is to prevent any period of hypoglycemia for at least four weeks. In diabetic patients with HU an appropriate educational program includes an emphasis on regular snacks at right times, warnings to take special care at periods of greater risk such as before lunch, moderation in alcohol intake and about the danger of delayed hypoglycemia after heavy alcohol intake or prolonged exercise. Diabetes self-management education can have physical and psychosocial benefits, and results in behavior changes with positive influence in outcome. A self-awareness intervention of 8 sessions, each lasting 3 h, was designed to determine whether there are psychosocial and physical benefits of self-awareness intervention in 29 adults with T1DM and HU. Post-intervention the participants detected more cues of euglycemia and hypoglycemia and experienced significant increases in integration and metabolic control[117].
In a randomized, prospective multi-centre trial, the effect of a specific training program for patients with hypoglycemia problem was compared with a control group receiving a standardized education program aiming of at avoidance of hypoglycemia by optimization of insulin therapy[118]. Compared to control group, the specific training program demonstrates additional benefits in terms of improving HU, reducing mild hypoglycemia, and detecting ant treating low blood glucose[118]. In the Dose Adjustment for Normal Eating-Hypoglycemia Awareness Restoration study, a 6-wk pilot intervention using motivational interviews and cognitive behavioral techniques around hypoglycemia, in 23 people with HU; support the importance of educational programs to improve HU. One year after the intervention HU had improved, mean rates of severe hypoglycemia fell from 3 to 0 per person per year, and worry and behavior around hypoglycemia improved[119]. In a sub-study of HypoCOMPaSS trial aimed to assess the restoration of impaired hypoglycemia awareness and defective hypoglycemia counter-regulation by an educational strategy targeted at hypoglycemia avoidance, in 18 adults patients with T1DM; following the 6-mo intervention the mean glucose concentration at which participants first experienced symptoms of hypoglycemia significantly increased from baseline (from 2.6 to 3.1 mg/dL), and counter-regulatory responses to hypoglycemia were also enhanced[120].
Jointly, the results of these three studies suggest that interventions that include education around hypoglycemia avoidance may help to decrease HU.
Treatment of HU
The treatment options for the management of HU are listed in Table Table11.
Table 1
Treatment options for the management of hypoglycemia unawareness and mechanisms of action
Treatments options Mechanism of action
Optimizing insulin treatment Avoidance of hypoglycemia
Pharmacological therapy
β2-adrenergic agents Enhancement of adrenaline effect
Methylxanthine derivates (caffeine, theophylline) Central nervous system stimulation
Serotonin reuptake inhibitors (fluoxetine, sertraline, paroxetine) Unknown. It has been hypothesized that the effect could be mediated by an atypical presentation of serotonin syndrome that will lead to autonomic dysfunction
KATP channel modulators Modulation of hypoglycemia sensing
Other treatments
Islet cell transplantation Improving metabolic control
Fructose Modulation of hypoglycemia sensing
Open in a separate window
Optimizing insulin treatment: It is important that in patients with a history of recurrent hypoglycemia and HU, the time of episodes be identified and the treatment regimen be adjusted accordingly[121]. Compared with regular insulin, rapid-acting insulin analogs have a more rapid onset of action, higher peak action, and shorter duration of action, which more closely approximates endogenous mealtime insulin response, allowing more flexibility in the time of meals and exercise, and, consequently, a lower risk of severe hypoglycemic events[122]. Similarly, long-acting insulin analogs exhibit a more consistent, longer, and flatter action profile than NPH insulin, and demonstrate a lower risk of hypoglycemia, particularly nocturnal[123,124]. In diabetic patients with HU substitution of regular insulin with rapid-acting insulin analogs (aspart, lispro or glulisine) reduces frequency of daytime hypoglycemia; and substitution of long-acting insulin analogues (detemir or glargine) for intermediate-acting insulin (NPH or premix) reduces the frequency of nocturnal and day time hypoglycemia[121,125]. Compared with insulin glargine, the newest basal analog insulin degludec offers a more constant time-action profile, a long duration of action, and a lower risk of hypoglycemia[126,127]. While clinical experience with insulin degludec is limited, a meta-analysis evaluating 5 clinical trials of 3372 subjects with T2DM demonstrated a 17% lower rate of overall hypoglycemia and a 32% lower rate of nocturnal hypoglycemia with insulin degludec, compared with insulin glargine[128]. These characteristics may facilitate the achievement of glycemic control with insulin degludec with fewer hypoglycemic events in patients with HU.
An alternative approach is to use continuous subcutaneous insulin infusion (CSII). A study was designed by Giménez et al[129] to evaluate the effect of CSII on hypoglycemia awareness and on glucose profile in a cohort of T1DM subjects in which 95% had established HU and had experienced two or more episodes of severe hypoglycemia in the preceding two years, for a 24-mo period. Severe hypoglycemic episodes fell from 1.25 per subject-year to 0.05 after 24 mo, an improvement in all the aspects of quality of life, and an improved symptomatic response to experimentally-induced hypoglycemia was observed[130]. Previous studies[130-132] have also shown a reduction in hypoglycemia with CSII, particularly when a short-acting insulin analogue is used[2,133]. The decrease is partly due to better pharmacokinetic delivery of insulin and a 15%-20% reduction in insulin requirements compared with multiple doses of insulin[134]. Substitution of CSII for NPH insulin in patients with T1DM, especially at bedtime, resulted in a lower frequency of hypoglycemic episodes, and improved counter-regulatory and symptomatic responses during subsequent acute hypoglycemia[135]. On the other hand, administration of bolus doses of glucagon at times of impeding hypoglycemia during CSII lowered the frequency of hypoglycemia[136].
Pharmacological therapy: β-adrenergic antagonists or β-blockers alter the effects of epinephrine and could have potential effects on glucose homeostasis and the hypoglycemic counter-regulatory system. The more troubling concern regarding β-blockers is their potential effect on HU and blunting of the return to euglycemic levels after hypoglycemia has occurred, through the suppression of all adrenergically mediated symptoms of hypoglycemia. In patients with T1DM without HU, adrenergic symptoms did occur at lower glucose levels when subjects were treated with β-blockers[137]. Cardioselective β-blockers cause less alteration in the perception of hypoglycemia and may have an effect on correction of hypoglycemia than do their noncardioselective counterparts[138]. These agents should not be avoided in patients with diabetes but should be used with the same caution as when any new medication is added to a patient’s therapeutic regime.
It has been suggested that people with HU may have reduced β-adrenergic sensitivity, and this can be reversed by strict avoidance of hypoglycemia[139]. In T1DM patients, the use of β-adrenergic agonist terbutaline was associated with statistically significant higher glucose levels compared to control subjects during the first half and second half of the night, and with reduction of nocturnal hypoglycemic episodes (22 in the control group vs 1 in the group of terbutaline). β-adrenergic agonist had therefore been suggested as possible therapeutic options for HU, at the cost of inducing morning hyperglycemia. One of the concerns about using β-adrenergic agonist for the treatment of HU was associated with reduced β2 sensitivity observed in vitro. A recent study from De Galan et al[140] showed that sensitivity to β2-adrenergic receptor agonist stimulation is preserved in T1DM patients with HU. No long-term clinical trials to evaluate the usefulness of β-adrenergic agonist in the prevention of HU have been reported.
Several studies have evaluated the effects of the methylxantines derivatives caffeine and theophylline on HU and the counter-regulatory response to hypoglycemia. Both have been shown to augment symptom intensity and improve counter-regulatory responses in patients with T1DM with and without HU[2,141]. Using functional magnetic imaging, caffeine can restore regional brain activation normally lost during acute hypoglycemia[142]. In another trial designed to assess the impact of caffeine on the frequency and perception of hypoglycemia over a 3-mo period; patients receiving caffeine (200 mg/twice-daily) had statistically significant more symptomatic hypoglycemia episodes and more intense warning symptoms than patients receiving placebo[143]. These results suggest that modest amounts of caffeine enhance the sensitivity of hypoglycemia warning symptoms in patients with T1DM without increasing the incidence of severe hypoglycemia. de Galan et al[144] planned one study to evaluate the impact of theophylline on the response to hypoglycemia in 15 patients with T1DM who had a history of HU and 15 matched healthy control subjects. When compared with placebo, theophylline (2.8 mg/kg) improves de counter-regulatory response to a perception of hypoglycemia in the group with T1DM with HU[144]. Although modest doses of caffeine and theophylline may be effective at reducing HU in patients with T1DM at a low cost and without significant toxicity, larger doses may carry risk, and large trials are needed to determine efficacy, toxicity and dose-response curves.
The development of HU was associated with the use of selective serotonin reuptake inhibitors (SSRIs) in three patients with T1DM treated with different SSRIs (fluoxetine, sertraline and paroxetine) for depression and who were previously able to recognize and treat hypoglycemia symptoms[145]. HU occurred in all three patients within weeks of starting SSRI therapy. HU reversed after discontinuation of SSRI therapy[145]. The mechanism by which SSRIs might be associated with HU is unknown, but it has been hypothesized that the effect could be mediated by an atypical presentation of serotonin syndrome that will lead to autonomic dysfunction[146]. These observations suggest that in some patients, treatment with SSRIs may alter the perception of hypoglycemia, and should be used with caution in diabetic subjects with HU.
Infusion of the opioid-receptor antagonist naloxone increases the plasma epinephrine response to hypoglycemia and, when administered during hypoglycemia prevents attenuation of the plasma epinephrine response to subsequent hypoglycemia in humans[26,27].
Administration of a selective Kir6.2/SUR-1 KATP-channel agonist increases the epinephrine response to hypoglycemia in rats[147]. However, systemic administration of the nonselective KATP-channel agonist diazoxide suppresses the glucagon response and has no effect on the epinephrine response to hypoglycemia in nondiabetic humans[148]. These results suggest that KATP-channel modulators are not effective in humans, possibly due to inability to cross blood-brain barrier.
Other treatments: Islet cell transplantation (ICTx) prevents severe hypoglycemia[149], and restores some counter-regulatory hormone secretion[150]. In a retrospective study conducted in 31 T1DM recipients of ICTx, HU was assessed using the Clark hypoglycemic score (minimum = 0; maximum = 7; no hypoglycemia = 0; HU ≥ 4)[151] twice. A reduction in the proportion of patients with HU was observed post-ICTx (pre vs post-ICTx: 87% vs 13%) and a significant increase in glucose threshold that resulted in symptoms (pre vs post-ICTx: 41.4 mg/dL vs 58.4 mg/dL)[152]. These results were sustained even after the patient’s stratification based in islet function, graft dysfunction and graft failure[152]. These results suggests that improved metabolic control achieved with ICTx can restore hypoglycemia awareness in patients with T1DM, persisting even after islet graft failure.
Fructose infusion amplifies epinephrine and glucagon responses and increases glucose production during hypoglycemia in humans[153]. Fructose is a promising treatment but has not been tested in clinical trials.
Go to:
CONCLUSION
HU is a complex, difficult-to-study phenomenon that carries with it great risk to patients. HU is common in people with T1DM and is observed with less frequency in insulin-treated T2DM. Exposure to antecedent hypoglycemia, especially repeated episodes, is an important factor in the pathogenesis of HU. Although enormous advances have been made in our knowledge of the mechanisms of HU, further research is needed to elucidate the pathophysiology of counter-regulatory impairment and HU, and enable the development of more targeted strategies that support glucose counter-regulation and consequently reduce hypoglycemia. Numerous research studies have begun to uncover the mechanisms by which the central nervous system responds and adapts to hypoglycemia. Understanding these mechanisms will lead to better management and therapies that reduce the risk for hypoglycemia. Studies aiming to improve or even reverse HU have met with variable success and a number of research groups are considering new candidate pathways to develop a therapy. Therefore, until effective measures are developed to reverse HU, part of the role of the healthcare professional should be to educate people with diabetes on the risks associated with HU and should discuss hypoglycemia prevention strategies with their patients, so that they can have a better chance of achieving their glucose controls goals while avoiding the morbidity and mortality associated with hypoglycemia.
Go to:
Footnotes
P- Reviewer: Das UN, Osian G, Skok P S- Editor: Tian YL L- Editor: A E- Editor: Liu SQ
Conflict-of-interest statement: None.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: August 2, 2014
First decision: December 17, 2014
Article in press: April 2, 2015
Go to:
References
1. Desouza CV, Bolli GB, Fonseca V. Hypoglycemia, diabetes, and cardiovascular events. Diabetes Care. 2010;33:1389–1394. [PMC free article] [PubMed] [Google Scholar]
2. de Galan BE, Schouwenberg BJ, Tack CJ, Smits P. Pathophysiology and management of recurrent hypoglycaemia and hypoglycaemia unawareness in diabetes. Neth J Med. 2006;64:269–279. [PubMed] [Google Scholar]
3. Moghissi E, Ismail-Beigi F, Devine RC. Hypoglycemia: minimizing its impact in type 2 diabetes. Endocr Pract. 2013;19:526–535. [PubMed] [Google Scholar]
4. Briscoe VJ, Davis SN. Hypoglycemia in type 1 and type 2 diabetes: physiology, pathophysiology, and management. Clinical Diabetes. 2006;24:115–121. [Google Scholar]
5. Vignesh JP, Mohan V. Hypoglycaemia unawareness. J Assoc Physicians India. 2004;52:727–732. [PubMed] [Google Scholar]
6. Czyzewska K, Czerniawska E, Szadkowska A. Prevalence of hypoglycemia unawareness in patients with type 1 diabetes. Pediatr Diabet. 2012;13 Suppl 17:77. [Google Scholar]
7. Geddes J, Schopman JE, Zammitt NN, Frier BM. Prevalence of impaired awareness of hypoglycaemia in adults with Type 1 diabetes. Diabet Med. 2008;25:501–504. [PubMed] [Google Scholar]
8. Schopman JE, Geddes J, Frier BM. Prevalence of impaired awareness of hypoglycaemia and frequency of hypoglycaemia in insulin-treated type 2 diabetes. Diabetes Res Clin Pract. 2010;87:64–68. [PubMed] [Google Scholar]
9. Cryer PE. The barrier of hypoglycemia in diabetes. Diabetes. 2008;57:3169–3176. [PMC free article] [PubMed] [Google Scholar]
10. Miura J, Kajiura M, Hoshina S, Kobayashi H, Uchigata Y. The investigation of risk factor for the hypoglycemia unawareness in patients with type 1 diabetes using CGMS. Diabetes. 2012;61:A554. [Google Scholar]
11. Pambianco GL, Costacou T, Orchard TJ. Does hypoglycemia unawareness (HU) differ by gender in type 1 diabetes (T1D)? Diabetes. 2009;58 Suppl 1:A544. [Google Scholar]
12. Schouwenberg BJ, Veldman BA, Spiering W, Coenen MJ, Franke B, Tack CJ, de Galan BE, Smits P. The Arg16Gly variant of the beta2-adrenergic receptor predisposes to hypoglycemia unawareness in type 1 diabetes mellitus. Pharmacogenet Genomics. 2008;18:369–372. [PubMed] [Google Scholar]
13. Sejling AS, Kjaer TW, Pedersen-Bjergaard U, Remvig LS, Larsen A, Nielsen MN, Tarnow L, Thorsteinsson B, Juhl CB. The effect of recurrent hypoglycaemia on cerebral electrical activity in patients with type 1 diabetes and hypoglycaemia unawareness. Diabetes. 2013;62:A104. [Google Scholar]
14. Dagogo-Jack S, Rattarasarn C, Cryer PE. Reversal of hypoglycemia unawareness, but not defective glucose counterregulation, in IDDM. Diabetes. 1994;43:1426–1434. [PubMed] [Google Scholar]
15. Fanelli C, Pampanelli S, Epifano L, Rambotti AM, Di Vincenzo A, Modarelli F, Ciofetta M, Lepore M, Annibale B, Torlone E. Long-term recovery from unawareness, deficient counterregulation and lack of cognitive dysfunction during hypoglycaemia, following institution of rational, intensive insulin therapy in IDDM. Diabetologia. 1994;37:1265–1276. [PubMed] [Google Scholar]
16. Reno CM, Litvin M, Clark AL, Fisher SJ. Defective counterregulation and hypoglycemia unawareness in diabetes: mechanisms and emerging treatments. Endocrinol Metab Clin North Am. 2013;42:15–38. [PMC free article] [PubMed] [Google Scholar]
17. Ramanathan R, Cryer PE. Adrenergic mediation of hypoglycemia-associated autonomic failure. Diabetes. 2011;60:602–606. [PMC free article] [PubMed] [Google Scholar]
18. Jones TW, Porter P, Sherwin RS, Davis EA, O’Leary P, Frazer F, Byrne G, Stick S, Tamborlane WV. Decreased epinephrine responses to hypoglycemia during sleep. N Engl J Med. 1998;338:1657–1662. [PubMed] [Google Scholar]
19. Banarer S, Cryer PE. Sleep-related hypoglycemia-associated autonomic failure in type 1 diabetes: reduced awakening from sleep during hypoglycemia. Diabetes. 2003;52:1195–1203. [PubMed] [Google Scholar]
20. McGregor VP, Banarer S, Cryer PE. Elevated endogenous cortisol reduces autonomic neuroendocrine and symptom responses to subsequent hypoglycemia. Am J Physiol Endocrinol Metab. 2002;282:E770–E777. [PubMed] [Google Scholar]
21. Davis SN, Shavers C, Davis B, Costa F. Prevention of an increase in plasma cortisol during hypoglycemia preserves subsequent counterregulatory responses. J Clin Invest. 1997;100:429–438. [PMC free article] [PubMed] [Google Scholar]
22. Davis SN, Shavers C, Costa F, Mosqueda-Garcia R. Role of cortisol in the pathogenesis of deficient counterregulation after antecedent hypoglycemia in normal humans. J Clin Invest. 1996;98:680–691. [PMC free article] [PubMed] [Google Scholar]
23. Raju B, McGregor VP, Cryer PE. Cortisol elevations comparable to those that occur during hypoglycemia do not cause hypoglycemia-associated autonomic failure. Diabetes. 2003;52:2083–2089. [PubMed] [Google Scholar]
24. Goldberg PA, Weiss R, McCrimmon RJ, Hintz EV, Dziura JD, Sherwin RS. Antecedent hypercortisolemia is not primarily responsible for generating hypoglycemia-associated autonomic failure. Diabetes. 2006;55:1121–1126. [PubMed] [Google Scholar]
25. McCrimmon RJ, Song Z, Cheng H, McNay EC, Weikart-Yeckel C, Fan X, Routh VH, Sherwin RS. Corticotrophin-releasing factor receptors within the ventromedial hypothalamus regulate hypoglycemia-induced hormonal counterregulation. J Clin Invest. 2006;116:1723–1730. [PMC free article] [PubMed] [Google Scholar]
26. Caprio S, Gerety G, Tamborlane WV, Jones T, Diamond M, Jacob R, Sherwin RS. Opiate blockade enhances hypoglycemic counterregulation in normal and insulin-dependent diabetic subjects. Am J Physiol. 1991;260:E852–E858. [PubMed] [Google Scholar]
27. Vele S, Milman S, Shamoon H, Gabriely I. Opioid receptor blockade improves hypoglycemia-associated autonomic failure in type 1 diabetes mellitus. J Clin Endocrinol Metab. 2011;96:3424–3431. [PMC free article] [PubMed] [Google Scholar]
28. Milman S, Leu J, Shamoon H, Vele S, Gabriely I. Magnitude of exercise-induced β-endorphin response is associated with subsequent development of altered hypoglycemia counterregulation. J Clin Endocrinol Metab. 2012;97:623–631. [PMC free article] [PubMed] [Google Scholar]
29. Seaquist ER, Anderson J, Childs B, Cryer P, Dagogo-Jack S, Fish L, Heller SR, Rodriguez H, Rosenzweig J, Vigersky R. Hypoglycemia and diabetes: a report of a workgroup of the American Diabetes Association and the Endocrine Society. Diabetes Care. 2013;36:1384–1395. [PMC free article] [PubMed] [Google Scholar]
30. Zammitt NN, Warren RE, Deary IJ, Frier BM. Delayed recovery of cognitive function following hypoglycemia in adults with type 1 diabetes: effect of impaired awareness of hypoglycemia. Diabetes. 2008;57:732–736. [PubMed] [Google Scholar]
31. Puente EC, Silverstein J, Bree AJ, Musikantow DR, Wozniak DF, Maloney S, Daphna-Iken D, Fisher SJ. Recurrent moderate hypoglycemia ameliorates brain damage and cognitive dysfunction induced by severe hypoglycemia. Diabetes. 2010;59:1055–1062. [PMC free article] [PubMed] [Google Scholar]
32. Cryer PE. Death during intensive glycemic therapy of diabetes: mechanisms and implications. Am J Med. 2011;124:993–996. [PMC free article] [PubMed] [Google Scholar]
33. Cranston I, Reed LJ, Marsden PK, Amiel SA. Changes in regional brain (18)F-fluorodeoxyglucose uptake at hypoglycemia in type 1 diabetic men associated with hypoglycemia unawareness and counter-regulatory failure. Diabetes. 2001;50:2329–2336. [PubMed] [Google Scholar]
34. Dunn JT, Cranston I, Marsden PK, Amiel SA, Reed LJ. Attenuation of amydgala and frontal cortical responses to low blood glucose concentration in asymptomatic hypoglycemia in type 1 diabetes: a new player in hypoglycemia unawareness? Diabetes. 2007;56:2766–2773. [PubMed] [Google Scholar]
35. Mangia S, Tesfaye N, De Martino F, Kumar AF, Kollasch P, Moheet AA, Eberly LE, Seaquist ER. Hypoglycemia-induced increases in thalamic cerebral blood flow are blunted in subjects with type 1 diabetes and hypoglycemia unawareness. J Cereb Blood Flow Metab. 2012;32:2084–2090. [PMC free article] [PubMed] [Google Scholar]
36. Tesfaye N, Nangia S, De Martino F, Kumar A, Moheet A, Iverson E, Eberly LE, Seaquist ER. Hypoglycemia-induced increases in cerebral blood flow (CBF) are blunted in subjects with type 1 diabetes (TID) and hypoglycemia unawareness (HU) Diabetes. 2011;60:A79–A80. [PMC free article] [PubMed] [Google Scholar]
37. Criego AB, Tkac I, Kumar A, Thomas W, Gruetter R, Seaquist ER. Brain glucose concentrations in patients with type 1 diabetes and hypoglycemia unawareness. J Neurosci Res. 2005;79:42–47. [PubMed] [Google Scholar]
38. Oz G, Kumar A, Rao JP, Kodl CT, Chow L, Eberly LE, Seaquist ER. Human brain glycogen metabolism during and after hypoglycemia. Diabetes. 2009;58:1978–1985. [PMC free article] [PubMed] [Google Scholar]
39. Canada SE, Weaver SA, Sharpe SN, Pederson BA. Brain glycogen supercompensation in the mouse after recovery from insulin-induced hypoglycemia. J Neurosci Res. 2011;89:585–591. [PMC free article] [PubMed] [Google Scholar]
40. Öz G, Tesfaye N, Kumar A, Deelchand DK, Eberly LE, Seaquist ER. Brain glycogen content and metabolism in subjects with type 1 diabetes and hypoglycemia unawareness. J Cereb Blood Flow Metab. 2012;32:256–263. [PMC free article] [PubMed] [Google Scholar]
41. Gulanski BI, De Feyter HM, Page KA, Belfort-DeAguiar R, Mason GF, Rothman DL, Sherwin RS. Increased brain transport and metabolism of acetate in hypoglycemia unawareness. J Clin Endocrinol Metab. 2013;98:3811–3820. [PMC free article] [PubMed] [Google Scholar]
42. De Feyter HM, Mason GF, Shulman GI, Rothman DL, Petersen KF. Increased brain lactate concentrations without increased lactate oxidation during hypoglycemia in type 1 diabetic individuals. Diabetes. 2013;62:3075–3080. [PMC free article] [PubMed] [Google Scholar]
43. Moheet A, Emir UE, Terpstra M, Kumar A, Eberly LE, Seaquist ER, Öz G. Initial experience with seven tesla magnetic resonance spectroscopy of hypothalamic GABA during hyperinsulinemic euglycemia and hypoglycemia in healthy humans. Magn Reson Med. 2014;71:12–18. [PMC free article] [PubMed] [Google Scholar]
44. Chan O, Cheng H, Herzog R, Czyzyk D, Zhu W, Wang A, McCrimmon RJ, Seashore MR, Sherwin RS. Increased GABAergic tone in the ventromedial hypothalamus contributes to suppression of counterregulatory responses after antecedent hypoglycemia. Diabetes. 2008;57:1363–1370. [PMC free article] [PubMed] [Google Scholar]
45. Chan O, Paranjape S, Czyzyk D, Horblitt A, Zhu W, Ding Y, Fan X, Seashore M, Sherwin R. Increased GABAergic output in the ventromedial hypothalamus contributes to impaired hypoglycemic counterregulation in diabetic rats. Diabetes. 2011;60:1582–1589. [PMC free article] [PubMed] [Google Scholar]
46. Gold AE, MacLeod KM, Frier BM. Frequency of severe hypoglycemia in patients with type I diabetes with impaired awareness of hypoglycemia. Diabetes Care. 1994;17:697–703. [PubMed] [Google Scholar]
47. Choudhary P, Geddes J, Freeman JV, Emery CJ, Heller SR, Frier BM. Frequency of biochemical hypoglycaemia in adults with Type 1 diabetes with and without impaired awareness of hypoglycaemia: no identifiable differences using continuous glucose monitoring. Diabet Med. 2010;27:666–672. [PubMed] [Google Scholar]
48. Gerstein HC, Miller ME, Byington RP, Goff DC, Bigger JT, Buse JB, Cushman WC, Genuth S, Ismail-Beigi F, Grimm RH, et al. Effects of intensive glucose lowering in type 2 diabetes. N Engl J Med. 2008;358:2545–2559. [PMC free article] [PubMed] [Google Scholar]
49. Zoungas S, Patel A, Chalmers J, de Galan BE, Li Q, Billot L, Woodward M, Ninomiya T, Neal B, MacMahon S, et al. Severe hypoglycemia and risks of vascular events and death. N Engl J Med. 2010;363:1410–1418. [PubMed] [Google Scholar]
50. Seaquist ER, Miller ME, Bonds DE, Feinglos M, Goff DC, Peterson K, Senior P. The impact of frequent and unrecognized hypoglycemia on mortality in the ACCORD study. Diabetes Care. 2012;35:409–414. [PMC free article] [PubMed] [Google Scholar]
51. Jacobson AM, Musen G, Ryan CM, Silvers N, Cleary P, Waberski B, Burwood A, Weinger K, Bayless M, Dahms W, et al. Long-term effect of diabetes and its treatment on cognitive function. N Engl J Med. 2007;356:1842–1852. [PMC free article] [PubMed] [Google Scholar]
52. Reichard P, Pihl M. Mortality and treatment side-effects during long-term intensified conventional insulin treatment in the Stockholm Diabetes Intervention Study. Diabetes. 1994;43:313–317. [PubMed] [Google Scholar]
53. Gold AE, MacLeod KM, Deary IJ, Frier BM. Hypoglycemia-induced cognitive dysfunction in diabetes mellitus: effect of hypoglycemia unawareness. Physiol Behav. 1995;58:501–511. [PubMed] [Google Scholar]
54. Bolo NR, Musen G, Jacobson AM, Weinger K, McCartney RL, Flores V, Renshaw PF, Simonson DC. Brain activation during working memory is altered in patients with type 1 diabetes during hypoglycemia. Diabetes. 2011;60:3256–3264. [PMC free article] [PubMed] [Google Scholar]
55. Smith CB, Choudhary P, Pernet A, Hopkins D, Amiel SA. Hypoglycemia unawareness is associated with reduced adherence to therapeutic decisions in patients with type 1 diabetes: evidence from a clinical audit. Diabetes Care. 2009;32:1196–1198. [PMC free article] [PubMed] [Google Scholar]
56. Graveling AJ, Frier BM. Hypoglycemia unawareness is associated with reduced adherence to therapeutic decisions in patients with type 1 diabetes: evidence from a clinical audit: response to Smith et al. Diabetes Care. 2010;33:e15; author reply e16. [PubMed] [Google Scholar]
57. Ly TT, Gallego PH, Davis EA, Jones TW. Impaired awareness of hypoglycemia in a population-based sample of children and adolescents with type 1 diabetes. Diabetes Care. 2009;32:1802–1806. [PMC free article] [PubMed] [Google Scholar]
58. Hannonen R, Tupola S, Ahonen T, Riikonen R. Neurocognitive functioning in children with type-1 diabetes with and without episodes of severe hypoglycaemia. Dev Med Child Neurol. 2003;45:262–268. [PubMed] [Google Scholar]
59. Northam EA, Anderson PJ, Jacobs R, Hughes M, Warne GL, Werther GA. Neuropsychological profiles of children with type 1 diabetes 6 years after disease onset. Diabetes Care. 2001;24:1541–1546. [PubMed] [Google Scholar]
60. Ho MS, Weller NJ, Ives FJ, Carne CL, Murray K, Vanden Driesen RI, Nguyen TP, Robins PD, Bulsara M, Davis EA, et al. Prevalence of structural central nervous system abnormalities in early-onset type 1 diabetes mellitus. J Pediatr. 2008;153:385–390. [PubMed] [Google Scholar]
61. Golden MP, Ingersoll GM, Brack CJ, Russell BA, Wright JC, Huberty TJ. Longitudinal relationship of asymptomatic hypoglycemia to cognitive function in IDDM. Diabetes Care. 1989;12:89–93. [PubMed] [Google Scholar]
62. Perantie DC, Lim A, Wu J, Weaver P, Warren SL, Sadler M, White NH, Hershey T. Effects of prior hypoglycemia and hyperglycemia on cognition in children with type 1 diabetes mellitus. Pediatr Diabetes. 2008;9:87–95. [PubMed] [Google Scholar]
63. Chico A, Vidal-Ríos P, Subirà M, Novials A. The continuous glucose monitoring system is useful for detecting unrecognized hypoglycemias in patients with type 1 and type 2 diabetes but is not better than frequent capillary glucose measurements for improving metabolic control. Diabetes Care. 2003;26:1153–1157. [PubMed] [Google Scholar]
64. Hay LC, Wilmshurst EG, Fulcher G. Unrecognized hypo- and hyperglycemia in well-controlled patients with type 2 diabetes mellitus: the results of continuous glucose monitoring. Diabetes Technol Ther. 2003;5:19–26. [PubMed] [Google Scholar]
65. Desouza C, Salazar H, Cheong B, Murgo J, Fonseca V. Association of hypoglycemia and cardiac ischemia: a study based on continuous monitoring. Diabetes Care. 2003;26:1485–1489. [PubMed] [Google Scholar]
66. Tanenberg RJ, Newton CA, Drake AJ. Confirmation of hypoglycemia in the “dead-in-bed” syndrome, as captured by a retrospective continuous glucose monitoring system. Endocr Pract. 2010;16:244–248. [PubMed] [Google Scholar]
67. Johnston SS, Conner C, Aagren M, Smith DM, Bouchard J, Brett J. Evidence linking hypoglycemic events to an increased risk of acute cardiovascular events in patients with type 2 diabetes. Diabetes Care. 2011;34:1164–1170. [PMC free article] [PubMed] [Google Scholar]
68. Miller DR, Fincke G, Lafrance JP, Palnati M, Shao Q, Zhang Q, Fonseca V, Riddle M, Vijan S, Christiansen CI. Hypoglycaemia and risk of myocardial infarction in US veterans with diabetes. Diabetologia. 2009;52(suppl 1):S63. [Google Scholar]
69. Holstein A, Egberts EH. Risk of hypoglycaemia with oral antidiabetic agents in patients with Type 2 diabetes. Exp Clin Endocrinol Diabetes. 2003;111:405–414. [PubMed] [Google Scholar]
70. Amiel SA, Dixon T, Mann R, Jameson K. Hypoglycaemia in Type 2 diabetes. Diabet Med. 2008;25:245–254. [PMC free article] [PubMed] [Google Scholar]
71. Marrett E, Radican L, Davies MJ, Zhang Q. Assessment of severity and frequency of self-reported hypoglycemia on quality of life in patients with type 2 diabetes treated with oral antihyperglycemic agents: A survey study. BMC Res Notes. 2011;4:251. [PMC free article] [PubMed] [Google Scholar]
72. Whitmer RA, Karter AJ, Yaffe K, Quesenberry CP, Selby JV. Hypoglycemic episodes and risk of dementia in older patients with type 2 diabetes mellitus. JAMA. 2009;301:1565–1572. [PMC free article] [PubMed] [Google Scholar]
73. Allen KV, Frier BM. Nocturnal hypoglycemia: clinical manifestations and therapeutic strategies toward prevention. Endocr Pract. 2003;9:530–543. [PubMed] [Google Scholar]
74. Alagiakrishnan K, Mereu L. Approach to managing hypoglycemia in elderly patients with diabetes. Postgrad Med. 2010;122:129–137. [PubMed] [Google Scholar]
75. Bruce DG, Casey GP, Grange V, Clarnette RC, Almeida OP, Foster JK, Ives FJ, Davis TM. Cognitive impairment, physical disability and depressive symptoms in older diabetic patients: the Fremantle Cognition in Diabetes Study. Diabetes Res Clin Pract. 2003;61:59–67. [PubMed] [Google Scholar]
76. Bree AJ, Puente EC, Daphna-Iken D, Fisher SJ. Diabetes increases brain damage caused by severe hypoglycemia. Am J Physiol Endocrinol Metab. 2009;297:E194–E201. [PMC free article] [PubMed] [Google Scholar]
77. Abbaszadeh Ahranjani S, Tabatabaei-Malazy O, Pajouhi M. Diabetes in old age, a review. Iranian J Diabetes and Lipid Disorders. 2009;8:113–128. [Google Scholar]
78. Aung PP, Strachan MW, Frier BM, Butcher I, Deary IJ, Price JF. Severe hypoglycaemia and late-life cognitive ability in older people with Type 2 diabetes: the Edinburgh Type 2 Diabetes Study. Diabet Med. 2012;29:328–336. [PubMed] [Google Scholar]
79. Budnitz DS, Lovegrove MC, Shehab N, Richards CL. Emergency hospitalizations for adverse drug events in older Americans. N Engl J Med. 2011;365:2002–2012. [PubMed] [Google Scholar]
80. Evers IM, ter Braak EW, de Valk HW, van Der Schoot B, Janssen N, Visser GH. Risk indicators predictive for severe hypoglycemia during the first trimester of type 1 diabetic pregnancy. Diabetes Care. 2002;25:554–559. [PubMed] [Google Scholar]
81. Nielsen LR, Pedersen-Bjergaard U, Thorsteinsson B, Johansen M, Damm P, Mathiesen ER. Hypoglycemia in pregnant women with type 1 diabetes: predictors and role of metabolic control. Diabetes Care. 2008;31:9–14. [PubMed] [Google Scholar]
82. Robertson H, Pearson DW, Gold AE. Severe hypoglycaemia during pregnancy in women with Type 1 diabetes is common and planning pregnancy does not decrease the risk. Diabet Med. 2009;26:824–826. [PubMed] [Google Scholar]
83. Rossi G, Lapaczewski P, Diamond MP, Jacob RJ, Shulman GI, Sherwin RS. Inhibitory effect of pregnancy on counterregulatory hormone responses to hypoglycemia in awake rat. Diabetes. 1993;42:1440–1445. [PubMed] [Google Scholar]
84. Rosenn BM, Miodovnik M, Khoury JC, Siddiqi TA. Counterregulatory hormonal responses to hypoglycemia during pregnancy. Obstet Gynecol. 1996;87:568–574. [PubMed] [Google Scholar]
85. Ringholm L, Pedersen-Bjergaard U, Thorsteinsson B, Damm P, Mathiesen ER. Hypoglycaemia during pregnancy in women with Type 1 diabetes. Diabet Med. 2012;29:558–566. [PubMed] [Google Scholar]
86. Heller S, Damm P, Mersebach H, Skjøth TV, Kaaja R, Hod M, Durán-García S, McCance D, Mathiesen ER. Hypoglycemia in type 1 diabetic pregnancy: role of preconception insulin aspart treatment in a randomized study. Diabetes Care. 2010;33:473–477. [PMC free article] [PubMed] [Google Scholar]
87. Barendse S, Singh H, Frier BM, Speight J. The impact of hypoglycaemia on quality of life and related patient-reported outcomes in Type 2 diabetes: a narrative review. Diabet Med. 2012;29:293–302. [PubMed] [Google Scholar]
88. Davis RE, Morrissey M, Peters JR, Wittrup-Jensen K, Kennedy-Martin T, Currie CJ. Impact of hypoglycaemia on quality of life and productivity in type 1 and type 2 diabetes. Curr Med Res Opin. 2005;21:1477–1483. [PubMed] [Google Scholar]
89. Williams SA, Pollack MF, Dibonaventura M. Effects of hypoglycemia on health-related quality of life, treatment satisfaction and healthcare resource utilization in patients with type 2 diabetes mellitus. Diabetes Res Clin Pract. 2011;91:363–370. [PubMed] [Google Scholar]
90. Lundkvist J, Berne C, Bolinder B, Jönsson L. The economic and quality of life impact of hypoglycemia. Eur J Health Econ. 2005;6:197–202. [PubMed] [Google Scholar]
91. Fidler C, Elmelund Christensen T, Gillard S. Hypoglycemia: an overview of fear of hypoglycemia, quality-of-life, and impact on costs. J Med Econ. 2011;14:646–655. [PubMed] [Google Scholar]
92. Gold AE, Deary IJ, Frier BM. Hypoglycaemia and non-cognitive aspects of psychological function in insulin-dependent (type 1) diabetes mellitus (IDDM) Diabet Med. 1997;14:111–118. [PubMed] [Google Scholar]
93. Strachan MW, Deary IJ, Ewing FM, Frier BM. Recovery of cognitive function and mood after severe hypoglycemia in adults with insulin-treated diabetes. Diabetes Care. 2000;23:305–312. [PubMed] [Google Scholar]
94. King P, Kong MF, Parkin H, Macdonald IA, Tattersall RB. Well-being, cerebral function, and physical fatigue after nocturnal hypoglycemia in IDDM. Diabetes Care. 1998;21:341–345. [PubMed] [Google Scholar]
95. Ritholz MD, Jacobson AM. Living with hypoglycemia. J Gen Intern Med. 1998;13:799–804. [PMC free article] [PubMed] [Google Scholar]
96. Eadington DW, Frier BM. Type 1 diabetes and driving experience: an eight-year cohort study. Diabet Med. 1989;6:137–141. [PubMed] [Google Scholar]
97. Lave LB, Songer TJ, LaPorte RE. Should persons with diabetes be licensed to drive trucks?--Risk management. Risk Anal. 1993;13:327–334. [PubMed] [Google Scholar]
98. Cox DJ, Kovatchev B, Vandecar K, Gonder-Frederick L, Ritterband L, Clarke W. Hypoglycemia preceding fatal car collisions. Diabetes Care. 2006;29:467–468. [PubMed] [Google Scholar]
99. Lawton J, Rankin D, Elliott J, Heller SR, Rogers HA, De Zoysa N, Amiel S. Experiences, views, and support needs of family members of people with hypoglycemia unawareness: interview study. Diabetes Care. 2014;37:109–115. [PubMed] [Google Scholar]
100. Gonder-Frederick LA, Schmidt KM, Vajda KA, Greear ML, Singh H, Shepard JA, Cox DJ. Psychometric properties of the hypoglycemia fear survey-ii for adults with type 1 diabetes. Diabetes Care. 2011;34:801–806. [PMC free article] [PubMed] [Google Scholar]
101. Tupola S, Rajantie J, Akerblom HK. Experience of severe hypoglycaemia may influence both patient’s and physician’s subsequent treatment policy of insulin-dependent diabetes mellitus. Eur J Pediatr. 1998;157:625–627. [PubMed] [Google Scholar]
102. Leese GP, Wang J, Broomhall J, Kelly P, Marsden A, Morrison W, Frier BM, Morris AD. Frequency of severe hypoglycemia requiring emergency treatment in type 1 and type 2 diabetes: a population-based study of health service resource use. Diabetes Care. 2003;26:1176–1180. [PubMed] [Google Scholar]
103. Zhang Y, Wieffer H, Modha R, Balar B, Pollack M, Krishnarajah G. The burden of hypoglycemia in type 2 diabetes: a systematic review of patient and economic perspectives. J Clin Outcomes Manage. 2010;17:547–557. [Google Scholar]
104. Brod M, Christensen T, Thomsen TL, Bushnell DM. The impact of non-severe hypoglycemic events on work productivity and diabetes management. Value Health. 2011;14:665–671. [PubMed] [Google Scholar]
105. Fanelli CG, Epifano L, Rambotti AM, Pampanelli S, Di Vincenzo A, Modarelli F, Lepore M, Annibale B, Ciofetta M, Bottini P. Meticulous prevention of hypoglycemia normalizes the glycemic thresholds and magnitude of most of neuroendocrine responses to, symptoms of, and cognitive function during hypoglycemia in intensively treated patients with short-term IDDM. Diabetes. 1993;42:1683–1689. [PubMed] [Google Scholar]
106. Klonoff DC. Continuous glucose monitoring: roadmap for 21st century diabetes therapy. Diabetes Care. 2005;28:1231–1239. [PubMed] [Google Scholar]
107. Bode B, Gross K, Rikalo N, Schwartz S, Wahl T, Page C, Gross T, Mastrototaro J. Alarms based on real-time sensor glucose values alert patients to hypo- and hyperglycemia: the guardian continuous monitoring system. Diabetes Technol Ther. 2004;6:105–113. [PubMed] [Google Scholar]
108. Garg S, Zisser H, Schwartz S, Bailey T, Kaplan R, Ellis S, Jovanovic L. Improvement in glycemic excursions with a transcutaneous, real-time continuous glucose sensor: a randomized controlled trial. Diabetes Care. 2006;29:44–50. [PubMed] [Google Scholar]
109. Streja D. Can continuous glucose monitoring provide objective documentation of hypoglycemia unawareness? Endocr Pract. 2005;11:83–90. [PubMed] [Google Scholar]
110. Ly TT, Hewitt J, Davey RJ, Lim EM, Davis EA, Jones TW. Improving epinephrine responses in hypoglycemia unawareness with real-time continuous glucose monitoring in adolescents with type 1 diabetes. Diabetes Care. 2011;34:50–52. [PMC free article] [PubMed] [Google Scholar]
111. Choudhary P, Ramasamy S, Green L, Gallen G, Pender S, Brackenridge A, Amiel SA, Pickup JC. Real-time continuous glucose monitoring significantly reduces severe hypoglycemia in hypoglycemia-unaware patients with type 1 diabetes. Diabetes Care. 2013;36:4160–4162. [PMC free article] [PubMed] [Google Scholar]
112. The Clinical Trials gov. The effect of RT-CGM on glycemia and Qol in patients with T1DM and IHA (IN CONTROL). [Updated 2013 Mar 6] Available from: https://clinicaltrials.gov/ct2/show/NCT01787903. [Google Scholar]
113. Schrot RJ. Targeting plasma glucose: preprandial versus postprandial. Clin Diabetes. 2004;22:169–172. [Google Scholar]
114. Cryer PE, Axelrod L, Grossman AB, Heller SR, Montori VM, Seaquist ER, Service FJ. Evaluation and management of adult hypoglycemic disorders: an Endocrine Society Clinical Practice Guideline. J Clin Endocrinol Metab. 2009;94:709–728. [PubMed] [Google Scholar]
115. Qaseem A, Vijan S, Snow V, Cross JT, Weiss KB, Owens DK. Glycemic control and type 2 diabetes mellitus: the optimal hemoglobin A1c targets. A guidance statement from the American College of Physicians. Ann Intern Med. 2007;147:417–422. [PubMed] [Google Scholar]
116. Pampanelli S, Fanelli C, Lalli C, Ciofetta M, Sindaco PD, Lepore M, Modarelli F, Rambotti AM, Epifano L, Di Vincenzo A, et al. Long-term intensive insulin therapy in IDDM: effects on HbA1c, risk for severe and mild hypoglycaemia, status of counterregulation and awareness of hypoglycaemia. Diabetologia. 1996;39:677–686. [PubMed] [Google Scholar]
117. Hamdan M, Brunet P, Wagner J. [Metastatic jejunal perforation of cancer of the larynx] J Chir (Paris) 2008;128:332. [PubMed] [Google Scholar]
118. Hermanns N, Kulzer B, Kubiak T, Krichbaum M, Haak T. The effect of an education programme (HyPOS) to treat hypoglycaemia problems in patients with type 1 diabetes. Diabetes Metab Res Rev. 2007;23:528–538. [PubMed] [Google Scholar]
119. de Zoysa N, Rogers H, Stadler M, Gianfrancesco C, Beveridge S, Britneff E, Choudhary P, Elliott J, Heller S, Amiel SA. A psychoeducational program to restore hypoglycemia awareness: the DAFNE-HART pilot study. Diabetes Care. 2014;37:863–866. [PubMed] [Google Scholar]
120. Leelarathna L, Little SA, Walkinshaw E, Tan HK, Lubina-Solomon A, Kumareswaran K, Lane AP, Chadwick T, Marshall SM, Speight J, et al. Restoration of self-awareness of hypoglycemia in adults with long-standing type 1 diabetes: hyperinsulinemic-hypoglycemic clamp substudy results from the HypoCOMPaSS trial. Diabetes Care. 2013;36:4063–4070. [PMC free article] [PubMed] [Google Scholar]
121. Cryer PE, Davis SN, Shamoon H. Hypoglycemia in diabetes. Diabetes Care. 2003;26:1902–1912. [PubMed] [Google Scholar]
122. Hirsch IB. Insulin analogues. N Engl J Med. 2005;352:174–183. [PubMed] [Google Scholar]
123. Yki-Järvinen H, Dressler A, Ziemen M. Less nocturnal hypoglycemia and better post-dinner glucose control with bedtime insulin glargine compared with bedtime NPH insulin during insulin combination therapy in type 2 diabetes. HOE 901/3002 Study Group. Diabetes Care. 2000;23:1130–1136. [PubMed] [Google Scholar]
124. Rosenstock J, Schwartz SL, Clark CM, Park GD, Donley DW, Edwards MB. Basal insulin therapy in type 2 diabetes: 28-week comparison of insulin glargine (HOE 901) and NPH insulin. Diabetes Care. 2001;24:631–636. [PubMed] [Google Scholar]
125. Mensing C, Boucher J, Cypress M, Weinger K, Mulcahy K, Barta P, Hosey G, Kopher W, Lasichak A, Lamb B, et al. National standards for diabetes self-management education. Diabetes Care. 2006;29 Suppl 1:S78–S85. [PubMed] [Google Scholar]
126. Garber AJ, King AB, Del Prato S, Sreenan S, Balci MK, Muñoz-Torres M, Rosenstock J, Endahl LA, Francisco AM, Hollander P. Insulin degludec, an ultra-longacting basal insulin, versus insulin glargine in basal-bolus treatment with mealtime insulin aspart in type 2 diabetes (BEGIN Basal-Bolus Type 2): a phase 3, randomised, open-label, treat-to-target non-inferiority trial. Lancet. 2012;379:1498–1507. [PubMed] [Google Scholar]
127. Heise T, Nosek L, Bøttcher SG, Hastrup H, Haahr H. Ultra-long-acting insulin degludec has a flat and stable glucose-lowering effect in type 2 diabetes. Diabetes Obes Metab. 2012;14:944–950. [PubMed] [Google Scholar]
128. Ratner RE, Gough SC, Mathieu C, Del Prato S, Bode B, Mersebach H, Endahl L, Zinman B. Hypoglycaemia risk with insulin degludec compared with insulin glargine in type 2 and type 1 diabetes: a pre-planned meta-analysis of phase 3 trials. Diabetes Obes Metab. 2013;15:175–184. [PMC free article] [PubMed] [Google Scholar]
129. Giménez M, Lara M, Conget I. Sustained efficacy of continuous subcutaneous insulin infusion in type 1 diabetes subjects with recurrent non-severe and severe hypoglycemia and hypoglycemia unawareness: a pilot study. Diabetes Technol Ther. 2010;12:517–521. [PubMed] [Google Scholar]
130. Chantelau E, Spraul M, Mühlhauser I, Gause R, Berger M. Long-term safety, efficacy and side-effects of continuous subcutaneous insulin infusion treatment for type 1 (insulin-dependent) diabetes mellitus: a one centre experience. Diabetologia. 1989;32:421–426. [PubMed] [Google Scholar]
131. Bendtson I, Kverneland A, Pramming S, Binderboland C. Incidence of nocturnal hypoglycaemia in insulin-dependent diabetic patients on intensive therapy. Acta Med Scand. 1988;223:543–548. [PubMed] [Google Scholar]
132. Fanelli CG, Porcellati F, Pampanelli S, Bolli GB. Insulin therapy and hypoglycaemia: the size of the problem. Diabetes Metab Res Rev. 2004;20 Suppl 2:S32–S42. [PubMed] [Google Scholar]
133. Skyler JS, Ponder S, Kruger DF, Matheson D, Parkin CG. Is there a place for insulin pump therapy in your practice? Clinical Diabetes. 2007;25:50–56. [Google Scholar]
134. Bode BW, Steed RD, Davidson PC. Reduction in severe hypoglycemia with long-term continuous subcutaneous insulin infusion in type I diabetes. Diabetes Care. 1996;19:324–327. [PubMed] [Google Scholar]
135. Kanc K, Janssen MM, Keulen ET, Jacobs MA, Popp-Snijders C, Snoek FJ, Heine RJ. Substitution of night-time continuous subcutaneous insulin infusion therapy for bedtime NPH insulin in a multiple injection regimen improves counterregulatory hormonal responses and warning symptoms of hypoglycaemia in IDDM. Diabetologia. 1998;41:322–329. [PubMed] [Google Scholar]
136. Castle JR, Engle JM, El Youssef J, Massoud RG, Yuen KC, Kagan R, Ward WK. Novel use of glucagon in a closed-loop system for prevention of hypoglycemia in type 1 diabetes. Diabetes Care. 2010;33:1282–1287. [PMC free article] [PubMed] [Google Scholar]
137. Hirsch IB, Boyle PJ, Craft S, Cryer PE. Higher glycemic thresholds for symptoms during beta-adrenergic blockade in IDDM. Diabetes. 1991;40:1177–1186. [PubMed] [Google Scholar]
138. White JR, Campbell RK. Dangerous and common drug interactions in patients with diabetes mellitus. Endocrinol Metab Clin North Am. 2000;29:789–802. [PubMed] [Google Scholar]
139. Fritsche A, Stumvoll M, Häring HU, Gerich JE. Reversal of hypoglycemia unawareness in a long-term type 1 diabetic patient by improvement of beta-adrenergic sensitivity after prevention of hypoglycemia. J Clin Endocrinol Metab. 2000;85:523–525. [PubMed] [Google Scholar]
140. De Galan BE, De Mol P, Wennekes L, Schouwenberg BJ, Smits P. Preserved sensitivity to beta2-adrenergic receptor agonists in patients with type 1 diabetes mellitus and hypoglycemia unawareness. J Clin Endocrinol Metab. 2006;91:2878–2881. [PubMed] [Google Scholar]
141. Kerr D, Sherwin RS, Pavalkis F, Fayad PB, Sikorski L, Rife F, Tamborlane WV, During MJ. Effect of caffeine on the recognition of and responses to hypoglycemia in humans. Ann Intern Med. 1993;119:799–804. [PubMed] [Google Scholar]
142. Rosenthal MJ, Smith D, Yaguez L, Giampietro V, Kerr D, Bullmore E, Brammer M, Williams SC, Amiel SA. Caffeine restores regional brain activation in acute hypoglycaemia in healthy volunteers. Diabet Med. 2007;24:720–727. [PubMed] [Google Scholar]
143. Watson JM, Jenkins EJ, Hamilton P, Lunt MJ, Kerr D. Influence of caffeine on the frequency and perception of hypoglycemia in free-living patients with type 1 diabetes. Diabetes Care. 2000;23:455–459. [PubMed] [Google Scholar]
144. de Galan BE, Tack CJ, Lenders JW, Pasman JW, Elving LD, Russel FG, Lutterman JA, Smits P. Theophylline improves hypoglycemia unawareness in type 1 diabetes. Diabetes. 2002;51:790–796. [PubMed] [Google Scholar]
145. Kerr D, Everett J. Coffee, diabetes and insulin sensitivity. Diabetologia. 2005;48:1418. [PubMed] [Google Scholar]
146. Sawka AM, Burgart V, Zimmerman D. Loss of awareness of hypoglycemia temporally associated with selective serotonin reuptake inhibitors. Diabetes Care. 2001;24:1845–1846. [PubMed] [Google Scholar]
147. Fan X, Ding Y, Cheng H, Gram DX, Sherwin RS, McCrimmon RJ. Amplified hormonal counterregulatory responses to hypoglycemia in rats after systemic delivery of a SUR-1-selective K(+) channel opener? Diabetes. 2008;57:3327–3334. [PMC free article] [PubMed] [Google Scholar]
148. Raju B, Cryer PE. Loss of the decrement in intraislet insulin plausibly explains loss of the glucagon response to hypoglycemia in insulin-deficient diabetes: documentation of the intraislet insulin hypothesis in humans. Diabetes. 2005;54:757–764. [PubMed] [Google Scholar]
149. Froud T, Ricordi C, Baidal DA, Hafiz MM, Ponte G, Cure P, Pileggi A, Poggioli R, Ichii H, Khan A, et al. Islet transplantation in type 1 diabetes mellitus using cultured islets and steroid-free immunosuppression: Miami experience. Am J Transplant. 2005;5:2037–2046. [PubMed] [Google Scholar]
150. Rickels MR, Schutta MH, Mueller R, Kapoor S, Markmann JF, Naji A, Teff KL. Glycemic thresholds for activation of counterregulatory hormone and symptom responses in islet transplant recipients. J Clin Endocrinol Metab. 2007;92:873–879. [PubMed] [Google Scholar]
151. Clarke WL, Cox DJ, Gonder-Frederick LA, Julian D, Schlundt D, Polonsky W. Reduced awareness of hypoglycemia in adults with IDDM. A prospective study of hypoglycemic frequency and associated symptoms. Diabetes Care. 1995;18:517–522. [PubMed] [Google Scholar]
152. Leitão CB, Tharavanij T, Cure P, Pileggi A, Baidal DA, Ricordi C, Alejandro R. Restoration of hypoglycemia awareness after islet transplantation. Diabetes Care. 2008;31:2113–2115. [PMC free article] [PubMed] [Google Scholar]
153. Gabriely I, Hawkins M, Vilcu C, Rossetti L, Shamoon H. Fructose amplifies counterregulatory responses to hypoglycemia in humans. Diabetes. 2002;51:893–900. [PubMed] [Google Scholar]
Articles from World Journal of Diabetes are provided here courtesy of Baishideng Publishing Group Inc
OTHER FORMATS
PDF (1.4M)
ACTIONS
Cite
Collections
SHARE
RESOURCES
Similar articles
Cited by other articles
Links to NCBI Databases
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov
Tell us what you think!Close
| biology | 36173 | https://no.wikipedia.org/wiki/Hypoglykemi | Hypoglykemi | Hypoglykemi (gr. hypo + glyk + emi, for lite søtt i blodet), ofte kalt føling, er en tilstand hvor blodsukkeret er for lavt. Blodsukker under 4 mmol/liter (70 mg/dl) anses for hypoglykemi. Dette kan skyldes sykdom eller bruk av legemidler og doping. Sykdommer og tilstander som kan medføre hypoglykemi er feilbehandling av diabetes (ved for høye doser av insulin eller antidiabetika, ofte forbundet med at det ikke tas høyde for endret insulinbehov ved økt fysisk aktivitet, forandring i karbohydratinntak eller alkoholinntak), hyperinsulinemi, og faste. Legemidler som kan medføre hypoglykemi er insulin og antidiabetika.
Det skilles ofte mellom mild, moderat og alvorlig hypoglykemi. Ved mild hypoglykemi, klarer diabetikeren selv å øke blodsukkeret gjennom tilførsel av mat, før det synker til nivåer som kan forårsake bevisstløshet. Mild hypoglykemi forekommer hyppig blant diabetes type-1 pasienter, og er ansett som nesten umulig å unngå når man bruker insulin og har et velregulert blodsukker. Symptomer på mild hypoglykemi er uro, skjelving, svimmelhet, svette, rask puls, hjertebank, humørsvingninger og sult. Noen opplever uklart syn, kvalme eller oppkast. Det er også vanlig å få hodepine, bli forvirret, aggressiv eller få konsentrasjonsvansker og uklar tale. Hypoglykemi kan misforstås av andre som å være alkoholpåvirket
Symptomer på hypoglykemi varierer mellom ulike personer, men hos hver person inntrer hvert symptom ofte ved bestemte blodsukkernivåer og kan være en pekepinn på hvor alvorlig hypoglykemien er.
Alvorlig hypoglykemi oppstår med blodsukker lavere enn 2,8 mmol/l. Dersom hypoglykemien ikke korrigeres med inntak av raske karbohydrater, og blodsukkeret synker lavere enn 2 mmol/l, er det vanlig å oppleve kramper, bevissthetstap, bevisstløshet og lammelser. Hypoglykemi kan blant annet oppstå av at diabetikeren feilberegner insulindosen, og injiserer for mye insulin i forhold til mengden karbohydrater, det går for lang tid mellom- eller man hopper over måltid, eller på grunn av aktivitet som senker blodsukkeret. Ved velregulert diabetes type 1, er det vanlig med rundt to ukentlige hypoglykemier som krever resolutt handling.
Alvorlig hypoglykemi kan medføre hjerneskade og må derfor behandles øyeblikkelig. En diabetiker med anfall må alltid antas å kunne ha hypoglykemi og behandles deretter. Behandling av hypoglykemi er sukkerholdig mat og drikke så lenge personen er i stand til å svelge.
Dersom en person blir bevisstløs på grunn av hypoglykemi, omtales tilstanden som insulinsjokk. Ved insulinsjokk er ikke diabetikeren i stand til å svelge, og må få et seigt, sukkerholdig stoff som honning, sjokoladepålegg, sjokoladesaus/karamellsaus eller lignende smurt på innsiden av kinn/lepper imellom kinnet/leppene og tennene. Dersom diabetikeren har glukagonsprøyte tilgjengelig, settes denne i lårmuskelen og personen legges i stabilt sideleie. På sykehus kan glukagon injiseres intramuskulært eller glukose gis som intravenøs infusjon.
Personer uten diabetes kan tro de har lavt blodsukker når de er sultne. Noen merker symtomene på hypoglykemi, for eksempel sult, ørhet og konsentrasjonsvansker. Disse følelsen kommer på grunn av sult, ikke på grunn av hypoglykemi. Personer uten diabetes kan ha et helt normalt blodsukker i flere dager uten inntak av mat.
Det motsatte av hypoglykemi, høyt blodsukker, er hyperglykemi.
Se også
Hemoglobin A1c
Referanser
Litteratur
Eksterne lenker
Akutte medisinske lidelser
Endokrine sykdommer, ernæringssykdommer og metabolske forstyrrelser
Diabetes | norwegian_bokmål | 0.305315 |
fainting_mechanism/HypoglycemiainType1a.txt | Skip to Main Content
Umbrella Alt Text
Umbrella Alt TextClose
JOURNALS
PROFESSIONAL BOOKS
ADA STANDARDS OF CARE
ADA MEETING ABSTRACTS
CLINICAL COMPENDIA
PODCASTS
PROFESSIONAL NEWS
header search
search input
Search...
Search input auto suggest
filter your search
Clinical Diabetes
Search
Advanced Search
Sign In
Clinical Diabetes
Current
Browse
Info & About
Podcasts
Subscriptions
Alerts
Submit
Skip Nav Destination
Volume 24, Issue 3
1 July 2006
Issue Cover
Previous Article
Next Article
Prevalence of Hypoglycemia in Diabetes
Clinical Impact of Iatrogenic Hypoglycemia
Symptoms of Hypoglycemia
Hypoglycemia and Glycemic Thresholds
Counterregulatory Hormone Responses to Hypoglycemia in Older Adults
Counterregulatory Hormone Responses to Hypoglycemia in Women
Exercise-Related Hypoglycemia
Mechanisms of Counterregulatory Responses to Hypoglycemia in Type 1 Diabetes
Mechanisms of Counterregulatory Responses to Hypoglycemia in Type 2 Diabetes
Reducing the Risk of Iatrogenic Hypoglycemia
Conclusions
References
FEATURES| JULY 01 2006
Hypoglycemia in Type 1 and Type 2 Diabetes: Physiology, Pathophysiology,and Management
Vanessa J. Briscoe, PhD; Stephen N. Davis, MD
Crossmark: Check for Updates
Clin Diabetes 2006;24(3):115–121
https://doi.org/10.2337/diaclin.24.3.115
Connected Content
A reference has been published: Clinical Diabetes: Imagination and Innovation
Split-Screen
Views Icon
Views
Open the
PDFfor in another window
Share Icon
Share
Cite Icon
Cite
Get Permissions
IN BRIEF
The threat and incidence of hypoglycemia is the major limiting factor in intensive glycemic control for both type 1 and type 2 diabetes. This article reviews the physiology of the normal counterregulatory responses to hypoglycemia and the deficient counterregulatory defenses that occur in patients with diabetes. Treatment paradigms for establishing good glycemic control while limiting hypoglycemia are also discussed.
Hypoglycemia is one of the most feared complications of diabetes treatment. Unfortunately, the threat and incidence of iatrogenic hypoglycemia is increased in attempts to achieve euglycemia as recommended by current treatment guidelines. These recommendations are based on results from two landmark studies, the Diabetes Control and Complications Trial (DCCT) and U.K. Prospective Diabetes Study (UKPDS), which demonstrated the benefits of intensive glycemic control in type 1 and type 2 diabetes,respectively.1,2 These studies proved that microvascular and some macrovascular complications could be reduced by rigorous metabolic control. However, the associated increased frequency of hypoglycemia has limited the clinical implementation of such intensive therapy because of the pharmacokinetic imperfections of available treatment regimens.3-5
Hypoglycemia commonly occurs in clinical practice. Approximately 90% of all patients who receive insulin have experienced hypoglycemic episodes.6 Nonetheless, the combination of understanding the physiological counterregulatory responses induced by hypoglycemia and monitoring glycemic therapy can help reduce the prevalence of iatrogenic hypoglycemia.3-10
Prevalence of Hypoglycemia in Diabetes
Surveys investigating the prevalence of hypoglycemia have provided some alarming results. The DCCT reported a threefold increase in severe hypoglycemia and coma in intensively treated patients versus conventionally treated patients.11 An intensively treated individual with type 1 diabetes can experience up to 10 episodes of symptomatic hypoglycemia per week and severe temporarily disabling hypoglycemia at least once a year.1,11,12 An estimated 2-4% of deaths of people with type 1 diabetes have been attributed to hypoglycemia.4 Hypoglycemia is also relatively common in type 2 diabetes, with prevalence rates of 70-80% in clinical trials using insulin to achieve good metabolic control.2
Donnelly et al.13 randomly surveyed individuals (n = 267) with type 1 diabetes and insulin-treated type 2 diabetes to prospectively record hypoglycemic events encountered over a 4-week period. Of the 267 subjects, 155 reported 572 incidents of hypoglycemia. The type 1 diabetic subjects reported a rate of 43 events per patient per year, whereas subjects with type 2 diabetes reported a rate of 16 events per patient per year. For individuals with type 1 diabetes,predictors of hypoglycemia included a history of previous hypoglycemia(P = 0.006). Predictors for hypoglycemia for the insulin-treated type 2 diabetic subjects included a history of previous hypoglycemia (P< 0.0001) and duration of insulin treatment (P = 0.014). Self-reports of severe hypoglycemia in type 2 diabetic subjects were lower than in type 1 diabetic subjects. The authors also concluded that hypoglycemia occurs more often than previously reported2 in insulin-treated type 2 diabetes and with sufficient frequency to cause significant morbidity.
Clinical Impact of Iatrogenic Hypoglycemia
The brain depends on a continual supply of glucose and is vulnerable to any glucose deprivation.5,8,9 Unable to synthesize or store this primary source of energy, the brain is one of the first organs affected by lowered blood glucose levels.5,8,9 Once plasma glucose concentrations fall below the physiological range at a glycemic threshold of ∼ 70 mg/dl,5 a sequence of responses is activated that includes release of neuroendocrine hormones(also called counterregulatory or anti-insulin hormones), stimulation of the autonomic nervous system (ANS), and production of neurogenic and neuroglycopenic symptoms to protect the brain and limit systematic effects of hypoglycemia.
The normal physiological counterregulatory response to hypoglycemia consists of suppression of insulin release and secretion of glucagon and pancreatic polypeptide from the pancreas, epinephrine from the adrenal medullae, norepinephrine from sympathetic postganglionic nerve terminals and adrenal medulla, cortisol from the adrenal cortex, and growth hormone from the anterior pituitary gland.7 In humans,inhibition of insulin secretion is the initial defense against falling glucose and occurs at a plasma glucose concentration of ∼ 80 mg/dl.
Glucagon and epinephrine are the primary fast-acting hormones in the defense against acute hypoglycemia. Glucagon acts to increase endogenous glucose production and does so via increases in hepatic glycogenolysis and gluconeogenesis, providing three carbon glucose substrates (lactate, pyruvate,alanine, and glycerol). Epinephrine can also acutely increase endogenous glucose production. Epinephrine has effects similar to glucagon on hepatic glucose production but can also stimulate net renal glucose production. Additionally, epinephrine has an important physiological function in reducing insulin-stimulated glucose uptake.
During the prolonged hypoglycemia that is usually observed in clinical practice, it is the reduced glucose uptake in peripheral tissues that contributes most to the preservation of circulating glucose levels and hence the defense against hypoglycemia. Activation of the sympathetic nervous system(via both circulating catecholamines and direct innervation) results in increased lipolysis in adipocytes. The increased release of free fatty acids(FFAs) results in significant glucose sparing (because tissues can oxidize FFAs instead of glucose). In fact, the contribution of FFAs has been estimated to be 25% of the total defense against hypoglycemia.3-7
Growth hormone and cortisol play a modest role in the metabolic defense against acute hypoglycemia but become more important during prolonged hypoglycemia.7 In fact, the counterregulatory actions of growth hormone and cortisol on increasing glucose production and restraining glucose disposal do not become evident until 4 hours after the onset of hypoglycemia. Even so, their counterregulatory actions are only ∼ 20% compared to that of epinephrine.7
Symptoms of Hypoglycemia
For people with type 1 diabetes and many with advanced type 2 diabetes,hypoglycemia is a fact of life.1-5 Those attempting to achieve better glycemic control suffer many episodes of mild to moderate hypoglycemia. Although the level of plasma glucose that indicates hypoglycemia is sometimes debated, it may be best defined in a physiological context as a plasma glucose of < 70 mg/dl (< 60 mg/dl whole blood). This is because the glycemic threshold for activation of the anti-insulin neuroendocrine counterregulatory response occurs at a plasma glucose of 70 mg/dl. Additionally, antecedent hypoglycemia of 70 mg/dl has been demonstrated to reduce counterregulatory responses to subsequent hypoglycemia.3-7
Symptoms of hypoglycemia are divided into two categories. Neurogenic(autonomic) symptoms are triggered by a falling glucose level and cause patients to recognize that they are experiencing a hypoglycemic episode.5,7,8 These symptoms are activated by the ANS and are mediated in part by sympathoadrenal release of catecholamines (norepinephrine and epinephrine)from the adrenal medullae and acetylcholine from postsynaptic sympathetic nerve endings.5,8 Neurogenic symptoms and signs associated with elevated epinephrine levels include shakiness, anxiety, nervousness, palpitations, sweating, dry mouth,pallor, and pupil dilation.4,5,9 The cholinergic-mediated symptoms include diaphoresis, hunger, and paresthesias.3-5 However, it should be noted that epinephrine infusion in the presence of euglycemia to achieve levels commonly seen during hypoglycemia only produces 20% of the total neurogenic symptom scores found during hypoglycemia. This indicates that the genesis of hypoglycemic symptoms is multifocal and is probably mainly generated from central nervous system (CNS) efferent pathways.14
Neuroglycopenic symptoms occur as a result of brain neuronal glucose deprivation.3-5,9,10 Evidence of neuroglycopenia can be the signal most often recognized by patients' family and friends. These symptoms include abnormal mentation,irritability, confusion, difficulty speaking, ataxia, paresthesias, headaches,stupor, and eventually (if untreated) seizures, coma, and even death.3-5,9 Neuroglycopenic symptoms can also include transient focal neurological deficits (e.g., diplopia,hemiparesis)3-5 (Table 1).
Table 1.
Symptoms of Hypoglycemia
Symptoms of Hypoglycemia
View large
Hypoglycemia and Glycemic Thresholds
The glycemic thresholds responsible for the activation of the physiological defenses against hypoglycemia are dynamic rather than static.7 Thus,patients with a higher hemoglobin A1c (A1C) may perceive symptoms of hypoglycemia at a higher plasma glucose level than those with more intensive control.8 In fact, these patients may generate hypoglycemic symptoms even when their plasma glucose is above the normal range. This phenomenon is called“relative hypoglycemia” and is associated with release of counterregulatory hormones. This phenomenon commonly occurs when patients undergo intensification of their glucose control. This syndrome is self-limiting and usually takes 2-4 weeks for the brain to readjust to the improved and thus relatively reduced circulating glucose levels.8-10,15,16
The opposite is true in intensively controlled individuals with diabetes. They may not recognize hypoglycemia until their plasma glucose is considerably lower than the normal physiological glycemic thresholds.5,10 The changes in glycemic thresholds can be caused acutely by antecedent hypoglycemia and chronically by persistent hyperglycemia.1-5
To test the hypothesis that hypoglycemia itself causes reduced neuroendocrine and symptomatic responses to subsequent hypoglycemia, Heller and Cryer17 measured counterregulatory responses during repeated hypoglycemic clamp studies. These seminal experiments determined that two episodes of antecedent moderate hypoglycemia (50 mg/dl) resulted in significant reductions of plasma epinephrine, glucagon, pancreatic polypeptide, and cortisol responses to next-day hypoglycemia. Neurogenic and neuroglycopenic symptom responses were also reduced after antecedent hypoglycemia. A later study18 investigated the effects of morning hypoglycemia on neuroendocrine and metabolic responses to subsequent afternoon hypoglycemia. These experiments demonstrated that only one episode of prolonged, moderate hypoglycemia can also produce substantial blunting of counterregulatory hormones and the symptomatic response to subsequent hypoglycemia.18
The above studies combined with conceptually similar results from differing laboratories allowed the term “hypoglycemia-associated autonomic failure” to be coined. This syndrome includes reduced neuroendocrine counterregulatory responses to hypoglycemia and lowered glycemic thresholds for activation of physiological defenses against hypoglycemia, which together lead to a condition of hypoglycemic unawareness. Glycemic thresholds are shifted to lower plasma glucose levels in intensively treated type 1 and type 2 diabetic individuals,10,18-21 which further limits efforts to attain euglycemia.3,4
Counterregulatory Hormone Responses to Hypoglycemia in Older Adults
The risk of severe or fatal hypoglycemia associated with the use of oral hypoglycemic agents and insulin increases exponentially with age.22,23 Also, older adults with comorbidities, those using multiple medications, and those who are frequently hospitalized are at greater risk for iatrogenic hypoglycemia.24 Most people with type 2 diabetes are > 60 years of age.8 Therefore, it is important to appreciate the idiosyncratic and agespecific manifestations of hypoglycemic symptoms.9
Meneilly et al.22 have investigated the effects of age on counterregulatory responses during clamped hypoglycemia. Older adults with type 2 diabetes demonstrated reduced glucagon and growth hormone responses but increased epinephrine and cortisol responses when compared to age-matched nondiabetic control subjects. However,hypoglycemic symptom scores were similar in both groups at all levels of glycemia.
Matyka et al.,25 on the other hand, found differences in hypoglycemic symptom responses when comparing healthy older men aged 60-70 years with younger men aged 22-26 years. During hyperinsulinemic-hypoglycemic clamp studies, neuroendocrine responses for the two groups were similar. However, symptoms began earlier in the younger men and were more intense. Measures of psychomotor coordination deteriorated earlier in the older subjects and to a greater degree. The usual 10-20 mg/dl plasma glucose difference between the subjective awareness of hypoglycemia and the onset of cognitive dysfunction was lost in the older men.
This altered counterregulatory effect may contribute to the altered cognitive response to reductions in blood glucose. Thus, the lower glycemic threshold to hypoglycemia in older people may limit the time available to self-treat and thereby increase the risk of developing incapacitating neuroglycopenia.22,25 Additionally, these neurological symptoms of hypoglycemia may be misinterpreted in older patients because of coexisting illnesses, such as cerebrovascular diseases or dementia.22,25
Counterregulatory Hormone Responses to Hypoglycemia in Women
There is a large sexual dimorphism in counterregulatory responses to hypoglycemia. It has been clearly demonstrated that both healthy young women and women with type 1 diabetes have reduced neuroendocrine, ANS, and metabolic(endogenous glucose production) counterregulatory responses compared to age-and BMI-matched men.26-30
This is not because of differences in glycemic thresholds for activation of countrerregulatory responses. In a series of separate glucose clamps at glycemic targets of 90, 70, 60, and 50 mg/dl, Davis et al.29 demonstrated that reduced CNS drive is responsible for the sexual dimorphic responses to hypoglycemia occurring in women. In a subsequent study, Sandoval et al.30 determined that estrogen is the mechanism responsible for this sexual dimorphism.
Despite this, the prevalence of hypoglycemic episodes in type 1 diabetes is similar for men and women.1 This apparent paradox may be explained by the fact that women may be more resistant than men to the blunting effects of antecedent hypoglycemia on the ANS.30 Thus, two episodes of antecedent hypoglycemia in men will cause a twofold greater blunting of counterregulatory responses to subsequent hypoglycemia compared to women, with the result being that the usual sexual dimorphic response to hypoglycemia is eliminated.
Exercise-Related Hypoglycemia
Hypoglycemia can occur during, 1-2 hours after, or up to 17 hours after exercise. The mechanisms responsible for this phenomenon have been the subject of recent work.7 Aerobic exercise results in an increase in both insulin- and non-insulin-medicated glucose uptake.
During moderate-intensity exercise in nondiabetic individuals, endogenous insulin secretion is reduced by 40-60%. Thus, reductions are recommended in replacement insulin doses during exercise (basal and/or preprandial insulin). This can be supplemented with oral intake of 10-20 g of carbohydrate every 30-60 minutes depending on the intensity of exercise. Insulin sensitivity increases ∼ 2 hours after moderate-intensity exercise. Thus, consideration should be given to reducing basal and/or prandial insulin doses for 24 hours after exercise.
Additionally, recent studies have demonstrated that there is a vicious cycle of counterregulatory failure between exercise and hypoglycemia.7 Thus,two episodes of prolonged, moderate-intensity exercise can reduce ANS and neuroendocrine responses by 50% during subsequent hypoglycemia. Similarly, two episodes of antecedent hypoglycemia can reduce counterregulatory responses during subsequent exercise by 40-50%.7 Therefore,individuals who have had a previous episode of hypoglycemia are at greater risk of hypoglycemia during exercise. This may be countered by temporarily increasing glycemic targets, reducing preexercise insulin, and consuming appropriate amounts of carbohydrate.
Mechanisms of Counterregulatory Responses to Hypoglycemia in Type 1 Diabetes
Epinephrine (not glucagon) is the main defense against hypoglycemia in patients with type 1 diabetes of > 5 years' duration. This is because the pancreatic α-cell glucagon secretory response to hypoglycemia is irreversibly lost.3-6
Unfortunately, epinephrine responses to hypoglycemia also become impaired in type 1 diabetic patients undergoing intensive insulin treatment. This places intensively treated type 1 diabetic patients at a significant risk for recurrent hypoglycemia.31,32 These frequent bouts of hypoglycemia further reduce the counterregulatory responses to future hypoglycemia by ≥ 50%. This creates a vicious cycle of iatrogenic hypoglycemia-associated autonomic failure, whereby hypoglycemia induces further hypoglycemia.3-5,31
Davis et al.33 demonstrated that the magnitude of antecedent hypoglycemia produced proportional blunting of counterregulatory responses to subsequent hypoglycemia. In other words, the greater the depth of antecedent hypoglycemia, the greater the magnitude of subsequent counterregulatory failure.
The ANS is exquisitely sensitive to the effects of antecedent hypoglycemia. Two episodes of hypoglycemia of only 70 mg/dl can blunt subsequent counterregulatory responses by ∼ 30% in men. Similarly, short durations(20 minutes) of antecedent hypoglycemia also produce significant blunting of subsequent counterrregulatory responses.33 The reduction in ANS counterregulatory responses has significant clinical consequences because type 1 diabetic patients with deficient glucagon and epinephrine responses to hypoglycemia have a ≥ 25-fold risk of hypoglycemia during intensive insulin therapy.1
Hypoglycemia-associated autonomic failure is an acutely acquired syndrome that should be differentiated from classical diabetic autonomic neuropathy.3-6,19,31 It is also possible that patients with hypoglycemia-associated autonomic failure also have reduced adrenergic sensitivity (i.e., tissue responsiveness to circulating epinephrine). Korytkowski et al.34 demonstrated that type 1 diabetic subjects with blunted counterregulatory responses to hypoglycemia had reduced β-adrenergic sensitivity compared to patients with normal counterregulatory responses to hypoglycemia and healthy control subjects. Aftab-Guy et al.14 also demonstrated that patients with type diabetes had reduced whole-body tissue sensitivity to epinephrine, which was exacerbated by intensive glycemic control. This reduced tissue sensitivity to epinephrine resulted in lower endogenous glucose production and less inhibition of insulin-stimulated glucose uptake. The above data may be interpreted to indicate that reduced tissue responsiveness to epinephrine is an additional contributor to the syndrome of hypoglycemia-associated autonomic failure
Fritsche et al.35 demonstrated that if hypoglycemic episodes are avoided for 4 months, β-adrenergic sensitivity and hypoglycemic symptom responses increase, despite a persistently blunted epinephrine response to hypoglycemia. This may indicate that increases in β- adrenergic sensitivity are a prelude to restoration of endocrine and autonomic function when hypoglycemic episodes are avoided.7 Although controversial, other studies have also reported that some or all of the features of hypoglycemia-associated autonomic failure (i.e., blunted neuroendocrine counterregulatory responses) can be reversed with strict avoidance of antecedent hypoglycemia.3-5,36-38
Mechanisms of Counterregulatory Responses to Hypoglycemia in Type 2 Diabetes
Type 2 diabetes is a heterogeneous disease affecting a range of individuals from children to older adults. Therapies include diet, oral medications,glucagonlike peptide-1 analogs, insulin, or combination therapies and vary depending on patients' progressive β-cell failure.39 Hence,the clinical effect of hypoglycemia-associated autonomic failure in type 2 diabetes is less well established,3-5 and results differ considerably with respect to age, comorbidity, treatment modality (diet versus oral hypoglycemic agents versus insulin), metabolic control, body fat composition, and the presence of diabetic neuropathies.39,40
However, Segel et al.40 tested the hypothesis that there are neuroendocrine changes in glycemic responses to hypoglycemia in individuals with advanced type 2 diabetes. They reported that the glucagon response to falling plasma glucose was virtually absent in advanced insulin-treated type 2 diabetes. Glycemic thresholds for autonomic and symptomatic responses to hypoglycemia were also shifted to lower glucose concentrations by recent antecedent hypoglycemia.
Hence, patients with advanced type 2 diabetes, like those with type 1 diabetes, are at risk for hypoglycemia-associated autonomic failure and the resultant vicious cycle of recurrent iatrogenic hypoglycemia.31,40
Reducing the Risk of Iatrogenic Hypoglycemia
Fear of hypoglycemia is the major concern of patients receiving endogenous or exogenous insulin replacement therapy. Furthermore, patients receiving intensive insulin therapy have about a threefold greater incidence of severe disabling hypoglycemia than those receiving conventional insulin therapy.1,41
Education regarding all aspects of diabetes care is important in the prevention and treatment of hypoglycemia. Carbohydrate counting, insulin and oral medication dosing, concomitant medications, alcohol intake, exercise, and even driving should be included in the discussion. Education will help alleviate fear of hypoglycemia that may impede ideal glycemic control.42 Reducing iatrogenic hypoglycemia will involve patient empowerment and anticipatory guidance by both patients and health care providers. Providers will also take on the role of facilitator as they help patients navigate through the maze of diabetes self-care.
The topic of hypoglycemia has priority and demands to be routinely addressed with patients receiving medications that may themselves or in combination cause hypoglycemia. Lack of understanding of the diabetes-related therapeutic regimen will contribute to repeated incidents of hypoglycemia.42-47 Patients must understand time action profiles of their diabetes medications and realize that excessive treatment can be harmful. Providers should urge patients to wear potentially lifesaving diabetes alert identification.
Blood glucose monitoring is fundamentally important for people who experience hypoglycemic episodes, especially before they perform critical tasks such as driving.41,42 Also, in older individuals with diabetes who have comorbidities such as dementia, cerebral vascular accident, or depression, consideration should be given to these confounding factors.48 Factors that may predispose such patients to hypoglycemia include increased polypharmacy or medication nonadherence, impaired renal or hepatic metabolism,and poor or erratic nutrition.22,23,41,42 Hence, the American Geriatrics Society has recommended an A1C of ≤ 7% for healthy older adults and an A1C of ≤ 8% for the frail elderly.48
If patients report a history of hypoglycemia, details regarding the time of episodes need to be identified and the treatment regimen adjusted accordingly(Table 2). If these events go without intervention, the risk of recurrent severe hypoglycemia is high.45
Table 2.
Hypoglycemia Risk Factors
Hypoglycemia Risk Factors
View large
Injected insulin can produce absolute or relative insulin excess largely because of dosing and pharmacokinetics.42 With a basal-bolus insulin regimen, morning fasting hypoglycemia implicates the long- or intermediate-acting insulin. Daytime hypoglycemia may be caused by the rapid-, short-, or longer-acting insulins, depending on the regimen. Nocturnal hypoglycemia may also be caused by regular and longer-acting insulin. Substitution of preprandial regular insulin with rapid-acting insulin(e.g., glulisine, lispro, or aspart) reduces the frequency of daytime hypoglycemia.43,44 Similarly, substitution of a long-acting insulin analog (e.g., glargine or detemir) for intermediate-acting insulins such as NPH, lente, or premix 70/30 or 50/50 also reduces the frequency of nocturnal and daytime hypoglycemia.44
Insulin pump therapy (continuous subcutaneous insulin infusion) that uses rapid-acting insulin analogs can cause both nocturnal and morning fasting hypoglycemia. With nocturnal hypoglycemia, the basal insulin infusion rate may be problematic, whereas with fasting or daytime hypoglycemia, the preprandial insulin bolus doses, the basal insulin infusion rate, or both may be causing the problem.44
Insulin secretagogues—sulfonylureas, repaglinide, and nateglinide—can also produce hypoglycemia related to absolute or relative insulin excess. However, the sulfonylureas may pose the greatest risk of hypoglycemia in patients with altered renal or hepatic function and in older adults.42 Hence, agents such as glimepiride, glipizide XL, or nateglinide that are shorter-acting and have glucose-dependent insulin secretion would be preferable to reduce hypoglycemic risks.42
Hypoglycemia unawareness (loss of warning symptoms of hypoglycemia) implies recurrent hypoglycemia.42 Assessment of frequency and severity of hypoglycemia is required at each clinic visit. Additionally, inquiring at what blood glucose level patients can first sense low plasma glucose will provide an assessment of hypoglycemia unawareness.
If there is still no apparent cause from the history or blood glucose log,patients may be experiencing hypoglycemia during the night.41,42 Indeed, nighttime hypoglycemia can be a common occurrence in people with type 1diabetes.1,45 Sleep can preclude detection of symptoms warning of impending hypoglycemia.41,42 Approaches to the problem of nocturnal hypoglycemia include insulin regimen adjustments, such as the use of rapid-acting rather than regular insulin during the day and a long-acting basal insulin. Administration of bedtime snacks may be also appropriate.41,42,45
If a diagnosis of hypoglycemic unawareness is made, the solution will involve the acceptance of somewhat higher glucose levels in the short term. At least a 3-week period of meticulous avoidance of hypoglycemia could be attempted with the goal of encouraging a return to awareness of hypoglycemia.1-5 With the return of symptomatic hypoglycemia, patients can once more work toward achieving better glycemic control.
Review of patients' self-monitoring of blood glucose log will help interpret blood glucose patterns. Patients should always have a rapidly available source of glucose with them to treat hypoglycemia at the first sign of a low glucose (Table 3). Hypoglycemia (plasma glucose < 70 mg/dl), including asymptomatic hypoglycemia and most episodes of mild to moderate symptomatic hypoglycemia,is effectively self-treated by ingestion of some form of glucose. Pure glucose is preferred, although any form of carbohydrate that contains glucose will raise plasma glucose.
Table 3.
Tips for Preventing Hypoglycemia
Tips for Preventing Hypoglycemia
View large
The “rule of 15” is a helpful treatment regimen when patients are able to self-treat. Typically, 15 g of carbohydrate (rapidly absorbing forms of glucose such as glucose gel, sugar-containing soda, or glucose tablets) should raise the blood glucose by 50 mg/dl in ∼ 15 minutes. The glycemic response to oral glucose is transient; therefore, ingestion of a small complex carbohydrate snack shortly after the plasma glucose concentration rises is generally advisable, especially if the next meal is longer than 1 hour away.
Hypoglycemic patients who are unconscious or unable because of neuroglycopenia to take in oral carbohydrates can be treated with a parental glucagon injection. Glucagon kits require a prescription. Glucagon acts by mobilizing glucose stores from the liver via glycogenolysis. Thus, it is less effective in glycogen-depleted states (e.g., prolonged starvation or alcohol ingestion).
It is important that a glucagon kit be available for use and that patients'family members or caregivers are knowledgeable in its use. One does not need to be a health care professional to administer glucagon. Instruction regarding the potential side effects of glucagon (i.e., vomiting) is important. This will prevent any surprise and subsequent hesitancy to use it in the future. Also, care should be taken to ensure that the kit has not expired.
Intravenous glucose is the preferable treatment of severe iatrogenic hypoglycemia, particularly that caused by a sulfonylurea. These reactions are more likely to occur in elderly patients and are often prolonged and require continuous glucose infusion and frequent feedings.41,42
Conclusions
The threat and incidence of iatrogenic hypoglycemia is a major limiting factor in intensive glycemic management of diabetes. Nonetheless, it is possible to both improve glycemic control and minimize hypoglycemic risks by understanding the physiological counterregulatory responses and aggressively monitoring glycemic therapy.
Hypoglycemia is problematic in type 1 diabetes during aggressive glycemic therapy and in advanced type 2 diabetes because of compromised glucose counterregulatory systems. Therefore, education concerning self-monitoring of blood glucose, diet, physiological insulin replacement, medication, and lifestyle are important to maintain good glycemic control, avoid hypoglycemia,and prevent long-term complications.
Vanessa J. Briscoe, PhD, is a clinical research nurse practitioner, and Stephen N. Davis, MD, is chief of the Division of Diabetes, Endocrinology and Metabolism and Rudolph Kampmeier Professor of Medicine and Molecular Physiology and Biophysics at the Vanderbilt School of Medicine in Nashville,Tenn.
References
1The DCCT Research Group: The effect of intensive treatment of diabetes on the development and progression of long term complication in insulin-dependent diabetes mellitus. N Engl J Med 329: 977-986,1993
2The U.K. Prospective Diabetes Study Group: Intensive blood-glucose control with sulfonylureas or insulin compared with conventional treatment and risk of complication in patients with type 2 diabetes. Lancet 352:837-853, 1998
3Cryer PE:Hypoglycemia risk reduction in type 1 diabetes. Exp Clin Endocrinol Diabetes 109:S412-S423, 2001
4Cryer PE: Current concepts: Diverse causes of hypoglycemia-associated autonomic failure in diabetes. N Engl J Med 350:2272-2279, 2004
5Cryer PE, Davis SN, Shamoon H: Hypoglycemia in diabetes. Diabetes Care 26: 1902-1912,2003
6Cryer PE: Hypoglycemia: Pathophysiology, Diagnosis, and Treatment. New York, Oxford University Press,1997
7Diedrich L,Sandoval D, Davis SN: Hypoglycemia associated autonomic failure. Clin Auton Res 12:358-365, 2002
8Zammitt NN, Frier BM: Hypoglycemia in type 2 diabetes. Diabetes Care 28: 2948-2961,2005
9McAuley V, Deary IJ, Freier BM: Symptoms of hypoglycemia in people with diabetes. Diabet Med 18:690-705, 2001
10Amiel SA, Sherwin RS, Simonson DC, Tamborlane WV: Effect of intensive insulin therapy on glycemic thresholds for counterregulatory hormone release. Diabetes 37:901-907, 1988
11The DCCT Research Group: Hypoglycemia in the Diabetes Control and Complications Trial. Diabetes 46: 271-286,1997
12Hepburn DA,MacLeod KM, Pell AC, Scougal IJ, Frier BM: Frequency and symptoms of hypoglycaemia experienced by patients with type 2 diabetes treated with insulin. Diabet Med 10:231-237, 1993
13Donnelly LA,Morris AD, Frier BM, Ellis JD, Donnan PT, Durrant R, Band MM, Reekie G, Leese P: Frequency and predictors of hypoglycaemia in type 1 and insulin-treated type 2 diabetes: a population-based study. Diabet Med 22: 749-755,2005
14Aftab-Guy D,Sandoval D, Richardson MA, Tate D, Davis SN: Effects of glycemic control on target organ responses to epinephrine in type 1 diabetes. Am J Physiol Endocrinol Metab 289:E258-E265, 2005
15Korzon-BurakowskaA, Hopkins D, Matyka K, Lomas J, Pernet A, Macdonald I, Amiel S: Effects of glycemic control on protective responses against hypoglycemia in type 2 diabetes. Diabetes Care 21: 283-290,1998
16Boyle PJ, Schwartz NS, Shah SD, Clutter WE, Cryer PE: Plasma glucose concentrations at the onset of hypoglycemic symptoms in patients with poorly controlled diabetes and in nondiabetics. N Engl J Med 318:1487-1492, 1988
17Heller SR, Cryer PE: Reduced neuroendocrine and symptomatic responses to subsequent hypoglycemia after 1 episode of hypoglycemia in nondiabetic humans. Diabetes 4:223-226, 1991
18Davis SN, Tate D:Effects of morning hypoglycemia on neuroendocrine and metabolic responses to subsequent afternoon hypoglycemia in normal man. J Clin Endocrinol Metab 86:2043-2050, 2001
19Dagogo-Jack SE,Craft S, Cryer PE: Hypoglycemia-associated autonomic failure in insulin-dependent diabetes mellitus. J Clin Invest 91: 819-828,1993
20Segel SA, Paramore DS, Cryer PE: Defective glucose counterregulation in type 2 diabetes(Abstract). Diabetes 49:A131, 2000
21Spyer G,Hattersley AT, MacDonald IA, Amiel S, MacLeod KM: Hypoglycaemic counterregulation at normal blood glucose concentrations in patients with well controlled type 2 diabetes. Lancet 356: 1970-1974,2000
22Meneilly GS,Cheung E, Tuokko H: Altered responses to hypoglycemia of healthy elderly people. Diabetes 43:403-410, 1994
23Jaap AJ, Jones GC,McCrimmon RJ, Deary IJ, Frier BM: Perceived symptoms of hypoglycaemia in elderly type 2 diabetic patients treated with insulin. Diabet Med 15: 398-401,1998
24Shorr RI, Ray WA,Daugherty JR, Griffin MR: Incidence and risk factors for serious hypoglycemia in older persons using insulin or sulfonylureas. Arch Intern Med 157:1681-1686, 1997
25Matyka K, Evans M,Lomas J, Cranston I, MacDonald I, Amiel SA: Altered hierarchy of protective responses against severe hypoglycemia in normal aging in healthy men. Diabetes Care 20:135-141, 1997
26Amiel SA, Maran A,Powrie JK, Umpleby AM, MacDonald IA: Gender differences in counterregulation to hypoglycemia. Diabetologia 36: 460-464,1993
27Davis SN,Goldstein RE, Price L, Jacobs J, Cherringon AD: The effects of insulin on the counterregulatory response to equivalent hypoglycemia in patients with insulin-dependent diabetes mellitus. J Clin Endocrinol Metab 77:1300-1307, 1993
28Diamond MP, Jones T, Caprio S, Hallarman L, Diamond MC, Addabbo M, Tamborlane WV, Sherwin RS:Gender influences counterregulatory hormone responses to hypoglycemia. Metabolism 42:1568-1572, 1993
29Davis SN, Fowler S, Costa F: Hypoglycemic counterregulatory responses differ between men and women with type 1 diabetes. Diabetes 49: 65-72,2000
30Sandoval DA, Ertl AC, Richardson MA, Tate DB, Davis SN: Estrogen blunts neuroendocrine and metabolic responses to hypoglycemia. Diabetes 52: 1749-1755,2003
31Cryer PE:Mechanisms of hypoglycemia-associated autonomic failure and its component syndromes in diabetes. Diabetes 54: 3592-3598,2005
32White NH, Skor A,Cryer PE, Levandoski L, Dier DM, Santiago JV: Identification of type 1 diabetic patients at increased risk for hypoglycemia during intensive therapy. N Engl J Med 308:485-491, 1993
33Davis SN, Shavers C, Mosqueda-Garcia R, Costa F: Effects of differing antecedent hypoglycemia on subsequent counterregulation in normal humans. Diabetes 46:1328-1335, 1997
34Korytkowski MT,Mokan M, Veneman TE, Mitrakou A, Cryer PE, Gerich JE: Reduced betaadrenergic sensitivity in patients with type 1 diabetes and hypoglycemia unawareness. Diabetes Care 21:1939-1943, 1998
35Fritsche A, Stefan N, Haring H, Gerich J, Stumvoll M: Avoidance of hypoglycemia restores hypoglycemia awareness by increasing betaadrenergic sensitivity in type 1 diabetes. Ann Intern Med 134:729-736, 2001
36Cranston I, Lomas J, Maran A, Macdonald I, Amiel SA: Restoration of hypoglycemia awareness in patients with long-duration insulin-dependent diabetes. Lancet 344:283-287, 1994
37Fanelli C,Pampanelli S, Epifano L, Rambotti AM, Ciofetta M, Modarelli F, DiVincenzo A,Annibale B, Lepore M, Lalli C, Sindaco P, Brunetti P, Bolli G: Relative roles of insulin and hypoglycemia on induction of neuroendocrine responses to,symptoms of, and deterioration of cognitive function in hypoglycemia in male and female humans. Diabetologia 37: 797-807,1994
38Dagogo-Jack S,Rattarasarn C, Cryer PE: Reversal of hypoglycemia unawareness, but not defective glucose counterregulation, in IDDM. Diabetes 43: 1426-1434,1994
39de Galan BE,Hoekstra JBL: Glucose counterregulation in type 2 diabetes mellitus. Diabet Med 18:519-527, 2001
40Segel SA, Paramore DS, Cryer PE: Hypoglycemia-associated autonomic failure in advanced type 2 diabetes. Diabetes 51:724-732, 2002
41The DCCT Research Group: Epidemiology of severe hypoglycemia in the diabetes control and complication trial. Am J Med 90: 450-459,1991
42Gabriely I,Shamoon H: Hypoglycemia in diabetes: common, often unrecognized. Cleveland Clin J Med 71:335-342, 2004
43Cryer PE, Childs BP: Negotiating the barrier of hypoglycemia in diabetes. Diabetes Spectrum 15:20-27, 2002
44Heller SR, Amiel SA, Mansell P: Effect of the fast-acting insulin analog lispro on the risk of nocturnal hypoglycemia during intensified insulin therapy. Diabetes Care 22:1607-1611, 1999
45Ratner RE, Hirsch IB, Neifing JL, Garg SK, Mecca TE, Wilson CA: Less hypoglycemia with insulin glargine in intensive insulin therapy for type 1 diabetes. Diabetes Care 23: 639-643,2000
46Muhlhause I,Overmann H, Bender R, Bott U, Berger M: Risk factors of severe hypoglycemia in adult patients with type 1 diabetes—a prospective population based study. Diabetologia 41:1274-1282, 1998
47Perriello G, DeFeo P, Torlone E, Fanelli C, Santeusanio F, Brunetti P, Bolli GB: The dawn phenomenon in type 1 (insulin-dependent) diabetes mellitus: magnitude,frequency, variability, and dependency on glucose counterregulation and insulin sensitivity. Diabetologia 34: 21-28,1991
48Wright EE: Treat to target: ABCs for the elderly. DOC News 3: 4,2006
American Diabetes Association
Article has an altmetric score of 22
View Metrics
Email Alerts
Article Activity Alert
Online Ahead of Print Alert
Latest Issue Alert
See Also
Clinical Diabetes: Imagination and Innovation
We Recommend
Attenuated sympathoadrenal responses, but not severe hypoglycemia, during aggressive glycemic therapy of early type 2 diabetes.
Stephanie A Amiel et al., Diabetes, 2009
Hypoglycemia: An Excuse for Poor Glycemic Control?
Mohammad Saleh et al., Clin Diabetes, 2001
Antecedent hypoglycemia impairs autonomic cardiovascular function: implications for rigorous glycemic control.
Gail K Adler et al., Diabetes, 2008
Hypoglycemia-Associated Autonomic Failure in Advanced Type 2 Diabetes
Scott A. Segel et al., Diabetes, 2002
Negotiating the Barrier of Hypoglycemia in Diabetes
Philip E. Cryer et al., Diabetes Spectr, 2002
Inhibiting the inhibitors: Development of the IAP inhibitor xevinapant for the treatment of locally advanced squamous cell carcinoma of the head and neck
Robert L. Ferris et al., Cancer Treatment Reviews, 2023
Powered by
Latest
Most Read
Misconceptions and Beliefs Related to Diabetes Care and Their Effect on A1c Values in People With Type 2 Diabetes
From Surviving to Thriving: A Qualitative Study of Adapting Telehealth Systems for Specialty Diabetes Care Across Four California Medical Centers
Beneficial Effects of Glucagon-Like Peptide 1 (GLP-1) Receptor Agonist Use With Control-IQ Technology in Type 2 Diabetes
Unleashing the Potential of Blood Glucose Monitoring Data With the Ambulatory Glucose Profile Report
Section 14: Children and Adolescents
Online ISSN 1945-4953 Print ISSN 0891-8929
Journals
Diabetes
Diabetes Care
Clinical Diabetes
Diabetes Spectrum
Standards of Medical Care in Diabetes
Scientific Sessions Abstracts
BMJ Open Diabetes Research & Care
Books
ShopDiabetes.org
ADA Professional Books
Clinical Compendia
Clinical Compendia Home
News
Latest News
DiabetesPro SmartBrief
Other
Special Collections
DiabetesPro®
Diabetes Food Hub™
Insulin Affordability
Know Diabetes By Heart™
About
About the ADA
Journal Policies
For Reviewers
Advertising in ADA Journals
Reprints and Permission for Reuse
Copyright Notice/Public Access Policy
Resources
ADA Professional Membership
ADA Member Directory
Diabetes.org
X (Twitter)
Facebook
LinkedIn
Contact Us Cookie Policy Accessibility Terms & Conditions Get Adobe Acrobat Reader© Copyright American Diabetes Association
This site uses cookies. By continuing to use our website, you are agreeing to our privacy policy.
Accept | biology | 36173 | https://no.wikipedia.org/wiki/Hypoglykemi | Hypoglykemi | Hypoglykemi (gr. hypo + glyk + emi, for lite søtt i blodet), ofte kalt føling, er en tilstand hvor blodsukkeret er for lavt. Blodsukker under 4 mmol/liter (70 mg/dl) anses for hypoglykemi. Dette kan skyldes sykdom eller bruk av legemidler og doping. Sykdommer og tilstander som kan medføre hypoglykemi er feilbehandling av diabetes (ved for høye doser av insulin eller antidiabetika, ofte forbundet med at det ikke tas høyde for endret insulinbehov ved økt fysisk aktivitet, forandring i karbohydratinntak eller alkoholinntak), hyperinsulinemi, og faste. Legemidler som kan medføre hypoglykemi er insulin og antidiabetika.
Det skilles ofte mellom mild, moderat og alvorlig hypoglykemi. Ved mild hypoglykemi, klarer diabetikeren selv å øke blodsukkeret gjennom tilførsel av mat, før det synker til nivåer som kan forårsake bevisstløshet. Mild hypoglykemi forekommer hyppig blant diabetes type-1 pasienter, og er ansett som nesten umulig å unngå når man bruker insulin og har et velregulert blodsukker. Symptomer på mild hypoglykemi er uro, skjelving, svimmelhet, svette, rask puls, hjertebank, humørsvingninger og sult. Noen opplever uklart syn, kvalme eller oppkast. Det er også vanlig å få hodepine, bli forvirret, aggressiv eller få konsentrasjonsvansker og uklar tale. Hypoglykemi kan misforstås av andre som å være alkoholpåvirket
Symptomer på hypoglykemi varierer mellom ulike personer, men hos hver person inntrer hvert symptom ofte ved bestemte blodsukkernivåer og kan være en pekepinn på hvor alvorlig hypoglykemien er.
Alvorlig hypoglykemi oppstår med blodsukker lavere enn 2,8 mmol/l. Dersom hypoglykemien ikke korrigeres med inntak av raske karbohydrater, og blodsukkeret synker lavere enn 2 mmol/l, er det vanlig å oppleve kramper, bevissthetstap, bevisstløshet og lammelser. Hypoglykemi kan blant annet oppstå av at diabetikeren feilberegner insulindosen, og injiserer for mye insulin i forhold til mengden karbohydrater, det går for lang tid mellom- eller man hopper over måltid, eller på grunn av aktivitet som senker blodsukkeret. Ved velregulert diabetes type 1, er det vanlig med rundt to ukentlige hypoglykemier som krever resolutt handling.
Alvorlig hypoglykemi kan medføre hjerneskade og må derfor behandles øyeblikkelig. En diabetiker med anfall må alltid antas å kunne ha hypoglykemi og behandles deretter. Behandling av hypoglykemi er sukkerholdig mat og drikke så lenge personen er i stand til å svelge.
Dersom en person blir bevisstløs på grunn av hypoglykemi, omtales tilstanden som insulinsjokk. Ved insulinsjokk er ikke diabetikeren i stand til å svelge, og må få et seigt, sukkerholdig stoff som honning, sjokoladepålegg, sjokoladesaus/karamellsaus eller lignende smurt på innsiden av kinn/lepper imellom kinnet/leppene og tennene. Dersom diabetikeren har glukagonsprøyte tilgjengelig, settes denne i lårmuskelen og personen legges i stabilt sideleie. På sykehus kan glukagon injiseres intramuskulært eller glukose gis som intravenøs infusjon.
Personer uten diabetes kan tro de har lavt blodsukker når de er sultne. Noen merker symtomene på hypoglykemi, for eksempel sult, ørhet og konsentrasjonsvansker. Disse følelsen kommer på grunn av sult, ikke på grunn av hypoglykemi. Personer uten diabetes kan ha et helt normalt blodsukker i flere dager uten inntak av mat.
Det motsatte av hypoglykemi, høyt blodsukker, er hyperglykemi.
Se også
Hemoglobin A1c
Referanser
Litteratur
Eksterne lenker
Akutte medisinske lidelser
Endokrine sykdommer, ernæringssykdommer og metabolske forstyrrelser
Diabetes | norwegian_bokmål | 0.305315 |
fainting_mechanism/Reflex_syncope.txt |
Reflex syncope is a brief loss of consciousness due to a neurologically induced drop in blood pressure and/or a decrease in heart rate. Before an affected person passes out, there may be sweating, a decreased ability to see, or ringing in the ears. Occasionally, the person may twitch while unconscious. Complications of reflex syncope include injury due to a fall.
Reflex syncope is divided into three types: vasovagal, situational, and carotid sinus. Vasovagal syncope is typically triggered by seeing blood, pain, emotional stress, or prolonged standing. Situational syncope is often triggered by urination, swallowing, or coughing. Carotid sinus syncope is due to pressure on the carotid sinus in the neck. The underlying mechanism involves the nervous system slowing the heart rate and dilating blood vessels, resulting in low blood pressure and thus not enough blood flow to the brain. Diagnosis is based on the symptoms after ruling out other possible causes.
Recovery from a reflex syncope episode happens without specific treatment. Prevention of episodes involves avoiding a person's triggers. Drinking sufficient fluids, salt, and exercise may also be useful. If this is insufficient for treating vasovagal syncope, medications such as midodrine or fludrocortisone may be tried. Occasionally, a cardiac pacemaker may be used as treatment. Reflex syncope affects at least 1 in 1,000 people per year. It is the most common type of syncope, making up more than 50% of all cases.
Signs and symptoms[edit]
Episodes of vasovagal syncope are typically recurrent and usually occur when the predisposed person is exposed to a specific trigger. Before losing consciousness, the individual frequently experiences early signs or symptoms such as lightheadedness, nausea, the feeling of being extremely hot or cold (accompanied by sweating), ringing in the ears, an uncomfortable feeling in the heart, fuzzy thoughts, confusion, a slight inability to speak or form words (sometimes combined with mild stuttering), weakness and visual disturbances such as lights seeming too bright, fuzzy or tunnel vision, black cloud-like spots in vision, and a feeling of nervousness can occur as well. The symptoms may become more intense over several seconds to several minutes before the loss of consciousness (if it is lost). Onset usually occurs when a person is sitting up or standing.
When people lose consciousness, they fall down (unless prevented from doing so) and, when in this position, effective blood flow to the brain is immediately restored, allowing the person to regain consciousness. If the person does not fall into a fully flat, supine position, and the head remains elevated above the trunk, a state similar to a seizure may result from the blood's inability to return quickly to the brain, and the neurons in the body will fire off and generally cause muscles to twitch very slightly but mostly remain very tense.
The autonomic nervous system's physiological state (see below) leading to loss of consciousness may persist for several minutes, so
If patients try to sit or stand when they wake up, they may pass out again
The person may be nauseated, pale, and sweaty for several minutes or hours
Causes[edit]
Reflex syncope occurs in response to a trigger due to dysfunction of the heart rate and blood pressure regulating mechanism. When heart rate slows or blood pressure drops, the resulting lack of blood to the brain causes fainting.
Vasovagal[edit]
Typical triggers include:
Prolonged standing
Emotional stress
Pain
The sight of blood
Fear of needles
Time varying magnetic field (i.e. transcranial magnetic stimulation)
Situational[edit]
After or during urination (micturition syncope)
Straining, such as to have a bowel movement
Coughing
Swallowing
Lifting a heavy weight
Carotid sinus[edit]
Pressing upon a certain spot in the neck. This may happen when wearing a tight collar, shaving, or turning the head.
Pathophysiology[edit]
Regardless of the trigger, the mechanism of syncope is similar in the various vasovagal syncope syndromes. The nucleus tractus solitarii of the brainstem is activated directly or indirectly by the triggering stimulus, resulting in simultaneous enhancement of parasympathetic nervous system (vagal) tone and withdrawal of sympathetic nervous system tone.
This results in a spectrum of hemodynamic responses:
On one end of the spectrum is the cardioinhibitory response, characterized by a drop in heart rate (negative chronotropic effect) and in contractility (negative inotropic effect) leading to a decrease in cardiac output that is significant enough to result in a loss of consciousness. It is thought that this response results primarily from enhancement in parasympathetic tone.
On the other end of the spectrum is the vasodepressor response, caused by a drop in blood pressure (to as low as 80/20) without much change in heart rate. This phenomenon occurs due to dilation of the blood vessels, probably as a result of withdrawal of sympathetic nervous system tone.
The majority of people with vasovagal syncope have a mixed response somewhere between these two ends of the spectrum.
One account for these physiological responses is the Bezold-Jarisch reflex.
Vasovagal syncope may be part of an evolved response, specifically, the fight-or-flight response.
Diagnosis[edit]
In addition to the mechanism described above, a number of other medical conditions may cause syncope. Making the correct diagnosis for loss of consciousness is difficult. The core of the diagnosis of vasovagal syncope rests upon a clear description of a typical pattern of triggers, symptoms, and time course.
It is pertinent to differentiate lightheadedness, seizures, vertigo, and low blood sugar as other causes.
In people with recurrent vasovagal syncope, diagnostic accuracy can often be improved with one of the following diagnostic tests:
A tilt table test (results should be interpreted in the context of patients' clinical presentations and with an understanding of the sensitivity and specificity of the test)
Implantation of an insertable loop recorder
A Holter monitor or event monitor
An echocardiogram
An electrophysiology study
Treatment[edit]
Treatment for reflex syncope focuses on avoidance of triggers, restoring blood flow to the brain during an impending episode, and measures that interrupt or prevent the pathophysiologic mechanism described above.
Lifestyle changes[edit]
The cornerstone of treatment is avoidance of triggers known to cause syncope in that person. However, research has shown that people show great reductions in vasovagal syncope through exposure-based exercises with therapists if the trigger is mental or emotional, e.g., sight of blood. However, if the trigger is a specific drug, then avoidance is the only treatment.
A technique known as "applied tension" may be additionally useful in those who have syncope with exposure to blood. The technique is done by tightening the skeletal muscles for about 15 seconds when the exposure occurs and then slowing releasing them. This is then repeated every 30 seconds for a few minutes.
Because vasovagal syncope causes a decrease in blood pressure, relaxing the entire body as a mode of avoidance is not favorable. A person can move or cross their legs and tighten leg muscles to keep blood pressure from dropping so significantly before an injection.
Before known triggering events, the affected person may increase consumption of salt and fluids to increase blood volume. Sports drinks or drinks with electrolytes may be helpful.
People should be educated on how to respond to further episodes of syncope, especially if they experience prodromal warning signs: they should lie down and raise their legs, or at least lower their head to increase blood flow to the brain. At the very least, upon the onset of initial symptoms the patient should try to relocate to a 'safe', perhaps cushioned, location in case of losing consciousness. Positioning themselves in a way where the impact from falling or collapsing would be minimized is ideal. The 'safe' area should be within close proximity, since, time is of the essence and these symptoms usually climax to loss of consciousness within a matter of minutes. If the individual has lost consciousness, he or she should be laid down in the recovery position. Tight clothing should be loosened. If the inciting factor is known, it should be removed if possible (for instance, the cause of pain).
Wearing graded compression stockings may be helpful.
Medications[edit]
Certain medications may also be helpful:
Beta blockers (β-adrenergic antagonists) were once the most common medication given; however, they have been shown to be ineffective in a variety of studies and are thus no longer prescribed. In addition, they may cause the syncope by lowering the blood pressure and heart rate.
Medications which may be effective include: CNS stimulants fludrocortisone, midodrine, SSRIs such as paroxetine or sertraline, disopyramide, and, in health-care settings where a syncope is anticipated, atropine or epinephrine (adrenaline).
For people with the cardioinhibitory form of vasovagal syncope, implantation of a permanent pacemaker may be beneficial or even curative.
Types of long-term therapy for vasovagal syncope include
Preload agents
Vasoconstrictors
Anticholinergic agents
Negative cardiac inotropes
Central agents
Mechanical device
Discontinuation of medications known to lower blood pressure may be helpful, but stopping antihypertensive drugs can also be dangerous in some people. Taking antihypertensive drugs may worsen the syncope, as the hypertension may have been the body's way to compensate for the low blood pressure.
Prognosis[edit]
Brief periods of unconsciousness usually cause no lasting harm to health. Reflex syncope can occur in otherwise healthy individuals, and has many possible causes, often trivial ones such as prolonged standing with the legs locked.
The main danger of vasovagal syncope (or dizzy spells from vertigo) is the risk of injury by falling while unconscious. Medication therapy could possibly prevent future vasovagal responses; however, for some individuals medication is ineffective and they will continue to have fainting episodes.
See also[edit]
Cardioneuroablation
Orthostatic hypotension
Orthostatic intolerance
Postural orthostatic tachycardia syndrome
Roemheld Syndrome | biology | 91826 | https://da.wikipedia.org/wiki/Apopleksi | Apopleksi | Apopleksi (apoplexia cerebri, også kendt som slagtilfælde, hjerneblødning, blodprop i hjernen og stroke) er en pludseligt opstået neurologisk skade eller udfald på baggrund af iskæmi (nedsat blodforsyning) i hjernen. Den kan enten skyldes en blodprop i et forsynende blodkar (85%) eller en bristning af et blodkar, der medfører en blødning i hjernens væv (15%). Ved apopleksi er hurtig reaktion og behandling vigtig for at undgå død og skadelige følger.
Symptomerne i den akutte fase afhænger af hvilket hjerneområde, der er ramt. Hyppigt drejer det sig om nedsat bevidsthed, nedsat kraft og følelse i den ene side af kroppen og kognitive forstyrrelser, der påvirker sprog, rumfornemmelse og hverdagsfunktioner. Psykiske forandringer og depressioner er ligeledes almindelige.
Diagnosen stilles ud fra en klinisk undersøgelse, men det er ikke muligt at skelne mellem en blodprop og en blødning på baggrund af denne. Som supplement til den kliniske undersøgelse vil der næsten altid udføres en CT-scanning eller MR-scanning. Formålet med disse scanninger er at understøtte diagnosen, vurdere prognosen og udelukke differentialdiagnoser, f.eks. tumor/metastaser, subaraknoidal blødning (SAH) og subduralt hæmatom (SDH).I praksis skelnes der mellem apopleksi, hvor symptomerne har en varighed på over 24 timer og TCI (transitorisk cerebral iskæmi), hvor symptomerne forsvinder inden for 24 timer.
Der ses årligt mellem 10.000-12.000 nye tilfælde af apopleksi i Danmark og det er derved en af de største folkesygdomme. Op imod 10 % af patienterne dør inden for den første måned, og hos de overlevende medfører sygdommen ofte en betydelig invalidering.
Symptomer
De forskellige dele af hjernen har forskellige arbejdsopgaver og symptomerne vil derfor også være forskellige alt efter hvor i hjernen skaden er sket. Er der f.eks. et infarkt i det område der styrer venstre ben, vil patienten have en lammelse i venstre ben.
Dertil kan der være nogle mere generelle symptomer, fx hovedpine, træthed og epileptiske kramper. Symptomerne opstår enten fordi det område, som styrer den specifikke funktion, bliver ødelagt, eller fordi projektionsbanerne mellem det styrende område og kroppen bliver afskåret.
Lammelser, føleforstyrrelser og bevægeforstyrrelser
En stor del af vores hjerne beskæftiger sig med at bevæge forskellige dele af kroppen. Bevægelserne blive igangsat i det motoriske cortex, mens tilstødende områder har "programmer" for hvordan bevægelserne skal laves. Dertil er der en række områder dybere i hjerne, der, sammen med cerebellum, er vigtige for koordination og balance. Alle disse områder er forbundet med hinanden og samarbejder om at lave præcise og glidende bevægelser.
De motoriske nervebaner, der løber fra cortex og ned igennem ryggen, krydser nede i hjernestammen. Det betyder, at bevægelser i venstre side af kroppen styres af højre hjernehalvdel.
Der kan ses en række forskellige motoriske symptomer:
Parese: er nedsat kraft i en kropsdel. Der er bevægelse, men med mindre styrke end normalt. Hvis det er hele den ene side af kroppen kaldes det hemiparese.
Paralyse: betyder at al bevægelse er væk. Kropsdelen er altså hel lammet.
Styringsbesvær/ataksi: Betyder, at patienten har problemer med at styre en kropsdel eller hele kroppen. Beder man f.eks. patienten om at sætte en finger på næsen, kan man se, at armen ikke bevæges i en blød bue, men i en hakket og ujævn bue og ofte vil fingeren ramme forbi næsen.
Facialisparese
En facialisparese er en lammelse af ansigtets muskler (de muskler der enerveres af nervus facialis). Ved apopleksi er det næsten altid en central facialisparese hvor patienten har hængende mundvig og ikke kan puste kinden op. Derimod vil de ikke have hængende øjenlåg eller problemer med at rynke panden. Dette er til forskel for en perifer facialis parese, som f.eks. ses ved borrelia.
Dysfagi
Hvis musklerne i munden og svælget er blevet lammet, kan det påvirke patientens evne til at synke normalt. I svære tilfælde udebliver synkebevægelsen helt, mens den i lettere tilfælde kommer med sekunders forsinkelse.
Ved dysfagi lukkes der ikke ordentligt af til luftvejene og der er derfor risiko for, at der kommer vand, mad og sekreter ned i luftvejene (aspiration), hvilket kan medføre lungebetændelse.
Føleforstyrrelser
Ligesom der er motoriske områder, der styre bevægelser, er der sensoriske områder, der modtager beskeder fra kroppen. Beskederne kan groft deles i følesansen og proprioception.
Følesansen modtager besked om berøring, smerte, temperatur og tryk, og proprioception modtager besked fra muskler, sener og led om hvor de forskellige dele af kroppen befinder sig og om bevægelse.
Hvis følesansen bliver påvirket, bliver patientens evne til at føle berøring, smerte etc. nedsat. Mange beskriver det også som en summende eller stikkende fornemmelse. Det kan blive et problem, fordi patienten ikke kan mærke hvis de fx støder foden ind i noget, en hånd sidder i klemme eller de er ved at få trykssår.
Hvis proprioceptionen er påvirket, får hjernen ikke be-sked om hvor kropsdelene befinder sig i forhold til krop-pen. Det gør det meget svært at styre og koordinere bevægelserne rigtigt og gøre at patienten fremstår klodset og kluntet.
De sensoriske og motoriske baner hænger tæt sammen og derfor vil patienter med parese næsten altid også have føleforstyrrelser, og omvendt.
Taleproblemer
Det at tale er en kompliceret opgave, der involverer flere dele af hjernen. Der er dels en række ’talecentre’ der hos de fleste mennesker sidder i venstre hjernehalvdel, dels de motoriske områder, der laver bevægelserne i munden, tungen og svælget.
Der findes flere måder at opdele taleproblemer, men den opdeling vi bruger i hverdagen er:
Afasi: Omfatter både problemer med forståelsen og produktionen af sprog. Det kan altså både være et problem med at finde de rigtige ord, udtale ordene rigtigt og opbygge sætninger (ekspressiv afasi) og med at forstå det der bliver sagt til dem (impressiv afasi). De fleste patienter med afasi har dog både ekspressive og impressive problemer, og de kan samtidig have problemer med at læse og skrive.
Dysartri: Ved dysartri er der udelukkende et problem med at udtale ordene pga. lammelse i de muskler i mund, tunge og svælg man bruger til at tale. Der er ikke problemer med sætningsopbygning og forståelse. (Dog kan man godt have afasi og dysartri samtidig).
Patienter der har taleproblemer vurderes af en logopæd (talepædagog).
Kognitive problemer
Dette er en bred samlebetegnelse for problemer, der har med tankevirksomhed at gøre, herunder hukommelse, indlæring, indsigt, koncentration og hverdagsfærdigheder at gøre. Disse problemer er mindre synlige end f.eks. en lammelse og det kan være svært for patienten at beskrive dem. Det kræver derfor tæt observation og undersøgelse, før man finder ud af det præcise problem.
Nedsat sygdomsindsigt (Anosognosi): Betyder, at patienten ikke oplever eller erkender sin sygdom. Der er et område i parietallappen, som har med kropsopfattelse og sygdomsfornemmelse at gøre. Hvis dette område skades, får resten af hjernen ikke besked om at der er noget galt med kroppen. Patienten er derfor ikke bevidst om, at han/hun er syg og kan i nogle tilfælde benægte, at der skulle være noget galt.
Den manglende sygdomserkendelse kan både give problemer i forhold til genoptræningen, fordi patienten ikke er motiveret til træning, og i forhold til patientens sikkerhed, idet patienten ikke kan se sine egne begrænsninger.
Neglect & nedsat opmærksomhed: Betyder, at patienten retter det meste eller al sin opmærksomhed væk fra den syge side. Det kan både være i forhold til selve kroppen, at patienten ’glemmer’ den lammede arm eller ben. Det kan også være i forhold til omgivelserne, at patienten slet ikke oplever hvad der foregår på den lammede side og f.eks. ikke høre når der bliver talt fra den side og ikke ser hvad der foregår.
Pushing: Betyder, at patientens opfattelse af kroppens midtlinje er forstyrret. Derfor vil patienten spontant læne sig over mod den lammede side og bruge de raske ekstremiteter til aktivt at skubbe sig væk fra den raske side (deraf betegnelsen pushing). Patienten vil ofte insistere på at kroppen er lige, og ved forsøg på at rette den op, vil patienten arbejde imod.
Apraksi: Betyder nedsat evne til at planlægge og udføre hverdagsaktiviteter. Det kan være fra at patienten ikke kan finde ud af at bruge redskaberne rigtigt (prøver at rede hår med tandbørsten) eller at udføre de enkelte delaktiviteter i en forkert rækkefølge (begynder at tørre hår inden de har skyllet sæben ud).
Agnosi: betyder problemer med genkendelse, f.eks. af genstande eller ansigter. Hvis man f.eks. viser patienten en tandbørste, kan han ikke se hvad det er (visuel agnosi), men når patienten får den i hånden er han ikke i tvivl. Der kan også være problemer med genkendelse ved berøring af genstande (taktil agnosi).
Et andet typisk fænomen er ansigtsagnosi, hvor patienten ikke kan genkende ansigter. Når patienten ser sine pårørende virker de ikke bekendte, men lige så snart de pårørende snakker, kan patienten genkende stemmerne.
Hukommelsesproblemer: Det kan f.eks. være problemer med at huske sin fødselsdato eller huske informationerne fra personalet.
Gråd- og latterlabilitet: Det kaldes også for tvangs-gråd eller tvangslatter, hvor patienten umotiveret og ude af sammenhæng enten græder eller griner vold-somt, uden at patienten kan fortælle hvorfor. Det kan være meget ubehageligt for patienten, fordi deres opførsel virker upassende overfor andre.
Synsproblemer
Synet kan blive påvirket på flere måder. Hvis synscortex bagerst i occipitallappen eller nervebanerne mellem synscortex og øjnene bliver skadet, kan dele af synet forsvinde. Ofte sker det ved, at enten halvdelen eller det kvarte af synet på begge øjne forsvinder, det kaldes henholdsvis for hemianopsi og kvadrantanopsi. Det er ikke altid, at patienten selv oplever det manglende synsfelt. Her kan det f.eks. observeres ved, at patienten støder ind i ting på den ene side. Dette adskiller sig fra neglect ved at patienten ellers reagere normalt på stimuli fra den påvirkede side.
Dobbeltsyn er også et hyppigt symptom. Det kan opstå hvis koordinationen af øjnene bliver forstyrret eller hvis øjnenes muskler bliver lammet.
Andre symptomer ved apopleksi
Nedsat bevidsthed
Ved store skader i hjernen kan bevidstheden være påvirket på forskellige måder. Man kan tale om enten bevidsthedsindhold eller bevidsthedsniveau.
Bevidsthedsindhold beskriver om patienten er orienteret og relevant. Kan patienten fortælle sit navn og cpr-nummer, hvor han/hun er og hvilken dag det f.eks. er? Ved patienten om det er dag eller nat?
Bevidsthedsniveau beskriver hvor vågen patienten er. Ligger patienten med åbne øjne og følger med i hvad der foregår? Ligger patienten med lukkede øjne, men åbner dem ved tiltale eller smertestimuli? Er patienten ukontaktbar og uden reaktion?
Somnolens betyder at patienten er sovende, men kan vækkes til vågen tilstand
Ukontaktbar bruges om patienter, som er bevidstløse og ikke kan vækkes ved smertestimuli. Her bør dog uddybes med en beskrivelse af patienten og en Glasgow Coma Scale.
Hovedpine
Mange patienter, selv ved mindre apopleksier, klager over hovedpine. Den kan både være konstant, trykkende eller dunkende. Hos nogle er den værst om morgenen og hos andre er den lige kraftig igennem døgnet.
Patienter, der er bevidsthedssvækkede eller har afasi, kan ikke altid give udtryk for smerter. Her kan smerterne komme til udtryk som motorisk uro, grimasser eller øget puls og respirationsfrekvens.
Feber
Som en del af kroppens reaktion på den skade der er sket i hjernen, kan patienten godt få forhøjet temperatur. Det betyder ikke nødvendigvis, at patienten har en infektion, men kan bare være et tegn på at kroppen reagere. Temperatur forhøjelsen er imidlertid skadelig for hjernen og derfor forsøger man altid at sænke en temperatur over 37,5 °C ved at give Paracetamol.
Svimmelhed og balanceproblemer
Særligt hos patienter med skader i hjernestammen og cerebellum kan der være problemer med svimmelhed og med at holde balancen.
Kramper
Det skadede område i hjernen kan nogle gange være så irriteret, at det udløser et krampeanfald. I den akutte fase er det oftest patienter med blødning, der får krampeanfald. Men i det senere forløb kan både patienter med blødning og infarkt udvikle regelret epilepsi.
Træthed
Træthed (eller fatigue) er en af de hyppigste følger af apopleksi. Nogle patienter oplever, at de er trætte hele tiden eller hurtigt udtrættes ved aktivitet og måske sover meget. Andre oplever at de mangler energi, er uoplagte eller har svært ved at komme i gang. Mens nogle oplever, at de bliver meget følsomme over for for mange mennesker, støj og lignende.
Vigtigste symptomer og kliniske fund efter lokalitet
Ætiologi
De hyppigste årsager til apopleksi er:
Aterosclerose, som forårsager tilstopning af arterier inde i hjernen eller forkalkninger i større arterier uden for hjernen.
Kardielle embolier, blodpropper kan dannes andre steder i kroppen og føres til hjernen via blodstrømmen. Emboliske blodpropper kan stamme fra hjertet, hvor atrieflimmer eller skader på hjertevæggen, kan få blodet til at størkne og danne blodpropper, eller fra halspulsårene.
Intracranielle blødninger, ofte pga. kronisk forhøjet blodtryk, der har været ubehandlet eller dårligt reguleret, men kan også skyldes andre årsager, som blodfortyndende medicin, cancer, leversygdomme eller blodsygdomme.
Småkarstilstopning, hos patienter med kronisk forhøjet blodtryk kommer der små skader på arterierne, der får karvæggen til at hæve op. I de små kar kan hævelse få karet til at lukke helt af så blodstrømmen stopper. Dette giver nogle karakteristiske tynde langstrakte infarkter (lakunære infarkter) dybt i hjernen
Karsygdomme, som både kan være medfødte (f.eks. CADASIL) eller på grund af inflammation af kar-væggen, som kan komme af infektioner eller reumatiske sygdomme.
Risikofaktorer
Der findes over hundrede risikofaktorer for apopleksi. De mest betydningsfulde er disse:
Høj alder.
Forhøjet blodtryk, er en vigtig risikofaktor for både iskæmiske og hæmorrhagiske apopleksi.
Diabetes, fordobler risikoen for iskæmisk apopleksi. Specielt patienter med meget svingende blodsukker er i risiko. Mange ved ikke selv, at de har type-2 diabetes, og derfor skal alle patienter, som indlægges for apopleksi screenes for diabetes med faste blodsukker.
Hjertesygdomme, en hyppig årsag til apopleksi er atrieflimmer. Også andre hjertesygdomme, som fx myocardiopati, arytmi og hjerteklapsygdomme er risikofaktorer.
Dyslipidæmi, for meget kolesterol og triglycerid i blodet øger risikoen for blodpropper.
Rygning.
Alkoholoverforbrug, mere end henholdsvis 7 og 14 genstande for kvinder og mænd om ugen.
Familiær disponering, altså hvis andre i familien har haft blodpropper i hjernen eller hjertet.
Tidligere apopleksi eller TCI.
Forsnævring af halspulsårerne.
Overvægt og nedsat fysisk aktivitet.
Behandling
Inden man begynder behandlingen, skal man have afklaret om apopleksien skyldes en blodprop eller en hjerneblødning. Dette kan gøres ved en CT-scanning eller en MR-scanning. Ved en CT-scanning kan man i begyndelsen kun se forandringer, hvis der er tale om en blødning, mens man ved en MR-scanning kan se forandringerne i alle tilfælde.
Det evidensbaserede grundlag for behandlingen af patienter med apopleksi i Danmark er samlet i Referenceprogrammet for behandling af patienter med apopleksi og TCI. De fleste større sygehuse i Danmark har i løbet af de sidste 20 år oprettet specialiserede apopleksiafdelinger, til modtagelse og udredning af apopleksipatienter, i den akutte del af forløbet. I Referenceprogrammet bliver en apopleksiafdeling defineret som: en sygehusafdeling, der udelukkende eller næsten udelukkende beskæftiger sig med udredning og behandling af patienter med apopleksi, og som er karakteriseret ved tværfaglige team, et personale med særlig interesse for apopleksi, medinddragelse af pårørende og stadig kompetenceudvikling af personalet. Patienter, som bliver indlagt på disse afdelinger, bliver undersøgt og udredt efter standardiserede forløbsprogrammer. Disse programmer har til formål at sikre ensartede patientforløb af en høj kvalitet og med høj patienttilfredshed. Standardprogrammet vil på de fleste apopleksiafdelinger indbefatte:
Klinisk undersøgelse af en reservelæge ved modtagelsen og en speciallæge på andendagen
CT-scanning inden for 24 timer
Monitorering af vitalværdier: blodtryk, puls, saturation (iltmætning), respirationsfrekvens og temperatur
Undersøgelse for hjertesygdomme med EKG og telemetri
Blodprøver: Blodbillede, infektionsparametre, blodsukker, lever-/galde-/pancreastal, blodlipider
Antitrombotisk behandling: Ved modtagelsen gives 300 mg Acetylsalicylsyre og herefter tablet clopidogrel 75 mg dagligt
Vurdering af ergoterapeut, fysioterapeut og evt. logopæd
Patienten vil forblive indlagt til observation de første 48 timer efter symptomdebut, fordi risikoen for nye blodpropper er størst i denne periode.
Trombolyse
Hvis apopleksien skyldes en blodprop i hjernen har enkelte afdelinger i Danmark de senere år haft mulighed for at give blodpropsopløsende medicin, trombolysebehandling. Ved trombolyse gives der en kraftig blodfortyndende medicin intravenøst. Medicinen kan i en del tilfælde opløse blodproppen og derved forhindre yderligere skade på nervevævet. Behandlingen kan derfor kun bruges hvis der er tale om en blodprop, og derfor skal der først laves en CT-scanning for at udelukke en blødning. Hvis man giver trombolyse til en patient med en hjerneblødning, vil det få katastrofale følger, idet trombolysen vil modvirke blodets naturlige evne til at koagulere, og dermed vil blødningen være meget længere om at standse og dermed forrette større skade.
Behandling med trombolyse skal, af hensyn til effekt og sikkerhed, gives så hurtigt som muligt. Dels er sandsynligheden for at behandlingen har god effekt større, jo hurtigere den startes. Patienter der behandles efter 4½ time har ingen effekt af behandlingen. Derudover stiger risikoen for bivirkninger, fx blødning, jo længere tid der går fra symptomerne starter. Indenfor 3 timer er der ikke forskel i risikoen for blødning mellem behandling og placebo. Efter 3 timer er risikoen marginalt større i behandlingsgruppen, mens risikoen stiger betydeligt efter 4½ time. Derfor gives trombolysebehandling normalt ikke til patienter med apopleksi efter 4½ time. Behandlingen er stadig ny og noget kontroversiel. Diskussionen bygger på, at behandlingen er meget omkostningsfuld (10-15.000 kr.) og at der har været tvivl om den egentlige effekt af den. Dertil kommer at behandlingen skal iværksættes meget hurtigt; inden for 4½ time efter symptomerne er startet, hvilket i mange tilfælde kan være svært at efterkomme.
Trombolyse tilbydes i Aalborg, Holstebro, Århus, Esbjerg, Odense, Roskilde, Glostrup (på ulige datoer) og Bisbebjerg (lige datoer). Hvis det ikke lykkedes at fjerne blodproppen med trombolyse har man mulighed for at fjerne blodproppen med et kateter gennem lysken (thrombektomi). Dette foregår i Århus, Odense og på Rigshospitalet. Det skal foregå hurtigst muligt.
Hvis apopleksien skyldes en blødning, vil man kun i sjældne tilfælde tilbyde en operation. Hvis patienten får følger efter apopleksien, vil der tilbydes genoptræning, enten under indlæggelse eller ambulant.
Kørselsforbud
Efter en blodprop i hjernen er der øget risiko for at udvikle en ny blodprop, også selv om der behandles forebyggende. Tidligere fik alle patienter kørselsforbud efter en blodprop i hjernen i minimum tre måneder, men i dag er det kun patienter med særligt forhøjet risiko, som ikke må køre bil. Man vurderer denne risiko ud fra en standardiseret skala, hvor hver risikofaktor gives point. Det er derved den samlede pointscore, som er afgørende for, om der nedlægges kørselsforbud eller ej.
Se også
Hjerneskadeforeningen
Referencer
Neurologiske sygdomme
Dødsårsager | danish | 0.596077 |
fainting_mechanism/NBK534841.txt | U.S. flagAn official website of the United States government Here's how you know
NIH NLM LogoLog in
Access keysNCBI HomepageMyNCBI HomepageMain ContentMain Navigation
Bookshelf
Search database
Books
Search term
Search
Browse Titles AdvancedHelpDisclaimer
Cover of StatPearls
StatPearls [Internet].
Show details
Search term
Hypoglycemia
Philip Mathew; Deepu Thoppil.
Author Information and Affiliations
Last Update: December 26, 2022.
Go to:
Continuing Education Activity
Hypoglycemia is often defined by a plasma glucose concentration below 70 mg/dL; however, signs and symptoms may not occur until plasma glucose concentrations drop below 55 mg/dL. The symptoms of Whipple's triad have been used to describe hypoglycemia since 1938. For the Whipple triad, the practitioner must first recognize hypoglycemia symptoms, obtain low blood glucose, and finally, demonstrate immediate relief of symptoms by the correction of the low blood glucose after treatment. This activity reviews the evaluation and management of hypoglycemia and highlights the interprofessional team's role in educating patients with this condition about follow up care.
Objectives:
Describe the pathophysiology of hypoglycemia.
Explain the diagnostic approach on how to determine that a patient has hypoglycemia.
Review the management options available for hypoglycemia treatment.
Summarize the importance of interprofessional team strategies for improving care coordination and communication to aid in prompt diagnosis of hypoglycemia and improving outcomes in patients diagnosed with the condition.
Access free multiple choice questions on this topic.
Go to:
Introduction
Hypoglycemia is often defined by a plasma glucose concentration below 70 mg/dL; however, signs and symptoms may not occur until plasma glucose concentrations drop below 55 mg/dL. The symptoms of Whipple's triad have been used to describe hypoglycemia since 1938. For Whipple's triad, the practitioner must first recognize symptoms of hypoglycemia, then obtain low blood glucose, and finally, demonstrate immediate relief of symptoms by the correction of the low blood glucose with glucose treatment. Glucose is the primary metabolic fuel for the brain under physiologic conditions. Unlike other tissues of the body, the brain is very limited in supplying its glucose. Expectedly, the brain requires a steady supply of arterial glucose for adequate metabolic function. Potential complications can arise from an interruption in the glucose supply. As such, protective mechanisms to guard against low serum blood glucose (hypoglycemia) have evolved in the body.
During fasting states, serum glucose levels are maintained via gluconeogenesis and glycogenolysis in the liver. Gluconeogenesis is the pathway in which glucose is generated from non-carbohydrate sources. These non-carbohydrate sources could be protein, lipids, pyruvate, or lactate. In contrast, glycogenolysis is the breakdown of glycogen stored into glucose products. Much of glycogenolysis occurs in hepatocytes (liver) and myocytes (muscle).
Hypoglycemia is most often seen in patients who have diabetes that undergoing pharmacologic intervention. Among this group, patients with type 1 diabetes are three times as likely to experience hypoglycemia than patients with type 2 diabetes when receiving treatment.
Go to:
Etiology
In patients who do not have diabetes, hypoglycemia is uncommon, but when it occurs, there are a few major causes of hypoglycemia: pharmacologic, alcohol, critical illness, counter-regulatory hormone deficiencies, and non-islet cell tumors[1].
Most hypoglycemia cases occur in patients with diabetes who are undergoing therapeutic intervention with meglitinides, sulfonylureas, or insulin. Drugs are the most common cause of hypoglycemia.[2] Metformin, glucagon-like peptide-1 (GLP-1) receptor agonists, sodium-glucose co-transporter 2 inhibitors (SGLT-2), and dipeptidyl peptidase-4 (DPP-4) inhibitor use very infrequently will lead to hypoglycemia.Non-diabetic patients with intact hepatic function will rarely experience fasting hypoglycemia because of preventative counter-regulatory measures. An episode of true hypoglycemia in a non-diabetic patient might be due to iatrogenic causes such as surreptitious insulin use. Other potential causes of hypoglycemia are critical illness, alcohol, cortisol deficiency, or malnourishment.
Alcohol inhibits gluconeogenesis in the body but does not affect glycogenolysis. Thus, hypoglycemia occurs after several days of alcohol consumption, and after glycogen stores are depleted.
For example, in critical illness states, end-stage liver disease, sepsis, starvation, or renal failure, glucose utilization exceeds glucose intake, glycogenolysis, and/or gluconeogenesis. The result of this imbalance is potentially the reason why hypoglycemia may occur. Counter-regulatory hormone deficiencies can occur as in states of adrenal insufficiency. Hypoglycemia associated with such deficiencies is rare. Non-islet cell tumors may also cause hypoglycemia through increased secretion of insulin-like growth factor 2 (IGF-2).[3] IGF-2 increases glucose utilization, which can lead to hypoglycemia.
Insulinomas are hyperfunctioning islet cell tumors associated with increased insulin secretion. They can be life-threatening and primarily manifest with fasting morning hypoglycemia. Although these tumors are rare, they should be a consideration in the workup of suspected cases.[4]
Go to:
Epidemiology
Hypoglycemia is common with type 1 diabetes, particularly in those patients receiving intensive insulin therapy. Severe hypoglycemic events have reportedly been anywhere between 62 to 320 episodes per 100 patient-years in type 1 diabetes.[5] As opposed to patients who have type 1 diabetes and require insulin therapy exclusively, patients with type 2 diabetes experience hypoglycemia relatively less frequently compared to patients with type 1 diabetes. This can be, in part, due to pharmacotherapies that do not induce hypoglycemia like metformin. The incidence of hypoglycemia in patients with type 2 diabetes has been reportedly approximately 35 episodes for 100 patient-years.[6] There are no reported disparities in incidents based on gender.
Go to:
Pathophysiology
The body has inherent counter-regulatory mechanisms to prevent hypoglycemic episodes. All of these counter-regulatory mechanisms include an interplay of hormones and neural signals to regulate the release of endogenous insulin, to increase hepatic glucose output, and to alter peripheral glucose utilization. Among the counter-regulatory mechanisms, the regulation of insulin production plays a major role. Decrease in insulin production as a response to low serum glucose isn't the body's first line of defense against hypoglycemia. For endogenous glucose production to take place, particularly hepatic glycogenolysis, low insulin levels are necessary. As plasma glucose levels decline, beta-cell secretion of insulin also decreases, leading to increased hepatic/renal gluconeogenesis and hepatic glycogenolysis. Glycogenolysis maintains serum glucose levels over 8 to 12 hours until glycogen stores are depleted. Over time, hepatic gluconeogenesis contributes more to maintaining euglycemia when required.
The decrease in insulin production occurs while the glucose level is in the low-normal range. This serves as a distinctive feature compared to other counter-regulatory measures. Additional counter-regulatory measures typically occur once the serum glucose levels decrease beyond the physiologic range. Among the additional counter-regulatory mechanisms, pancreatic alpha cell secretion of glucagon is the next line of defense against hypoglycemia. Should increased glucagon fail to achieve euglycemia, adrenomedullary epinephrine is secreted.[7] All three counter-regulatory measures occur in the acute stage of hypoglycemia.
On occasions the previously mentioned counter-regulatory mechanisms may fail to resolve the hypoglycemia. At this point, further counter-regulatory measures are employed in the form of growth hormone and cortisol. Both the release of growth hormone and cortisol are seen in prolonged hypoglycemic state.
Go to:
History and Physical
The clinical manifestations of hypoglycemia can be classified as either neuroglycopenic or neurogenic. Neuroglycopenic signs and symptoms are signs and symptoms that result from direct central nervous system (CNS) deprivation of glucose. These include behavioral changes, confusion, fatigue, seizure, coma, and potential death if not immediately corrected. Neurogenic signs and symptoms can either be adrenergic (including tremors, palpitations, anxiety) or cholinergic (including hunger, diaphoresis, paresthesias). Neurogenic symptoms and signs arise from sympathoadrenal involvement (either norepinephrine or acetylcholine release) in response to perceived hypoglycemia.
A detailed history is essential in evaluating hypoglycemia. Pertinent issues that should be addressed while taking a patient's history include:
A detailed medication history
History of alcohol and/or drug use
History of psychiatric disorders
Personal or family history of diabetes mellitus or multiple endocrine neoplasia syndromes (MEN)
Unintentional weight changes
Changes in medication
Consideration of acute kidney injury or renal failure
Symptoms of different hormone deficiencies.
Timing of the hypoglycemic episode relative to meals or exercise.
There is no agreed-upon lab value that defines hypoglycemia. Hypoglycemia is said to be present when the patient has symptoms consistent with hypoglycemia in addition to a low serum glucose measurement (less than 70 mg/dL). This perspective reflects the idea that hypoglycemia is a clinical presentation coupled with a lab finding of low serum glucose rather than a pure chemistry finding. Typically neurogenic and neuroglycopenic symptoms of hypoglycemia occur at a glucose level of or below 50 to 55 mg/dL, but this threshold can vary from individual to individual.
Patients who have diabetes can present with symptoms of hypoglycemia at relatively higher serum glucose levels. The chronic hyperglycemia alters the "set point" in which neuroglycopenic/neurogenic symptoms become apparent. This phenomenon is referred to as "pseudohypoglycemia"[8] because the serum glucose may be within normal range despite symptom presentation.
Go to:
Evaluation
As previously mentioned, documentation of Whipple’s triad is a potential indicator of hypoglycemia, and any initial laboratory evaluation should confirm hypoglycemia. Other pertinent labs to consider include insulin, proinsulin, and C-peptide levels during any episode of suspected hypoglycemia. If C-peptide levels are low in the presence of high insulin levels, the patient has received exogenous insulin. The pro-form of insulin created within the body is attached to C-peptide. The body cleaves C-peptide from the pro-form of the molecule to create active insulin. Elevated C-peptide levels together with insulin levels can be seen with secretagogue agents such as sulfonylureas or insulin secretagogues since both classes of agents stimulate endogenous insulin secretion.
Once the use of exogenous insulin administration is ruled out, sources of endogenous hyperinsulinemia need to be considered. Localization is usually performed via abdominal computed tomography (CT) or with MRI.
Go to:
Treatment / Management
Identification of a hypoglycemic patient is critical due to potential adverse effects, including coma and/or death. Severe hypoglycemia can be treated with intravenous (IV) dextrose followed by infusion of glucose. For conscious patients able to take oral (PO) medications, readily absorbable carbohydrate sources (such as fruit juice) should be given. For patients unable to take oral agents, glucagon should be administered. Glucagon can be given intramusularly, or intranasally with the newest available formulations.[9][10] Once the patient is more awake, a complex carbohydrate food source should be given to achieve sustained euglycemia. More frequent blood glucose monitoring should occur to rule out further drops in blood sugar.
Nonpharmacological management of recurrent hypoglycemia involves patient education and lifestyle changes. Some patients are unaware of the serious ramifications of persistent hypoglycemia. As such, patients should be educated on the importance of routine blood glucose monitoring as well as on the identification of the individual's symptoms of hypoglycemia. If lifestyle changes are not effective in preventing further episodes, then pharmacologic intervention should be modified. Patients should be advised to wear a medical alert bracelet or necklace and to also carry a glucose source like gel, candy, or tablets on their purse in case symptoms arise. In the outpatient setting, reviewing blood sugar logs and food logs may help identify problem areas for the patient.
Glycemic control has been an important aspect of medical management due to the association between glycated hemoglobin levels and cardiovascular events in diabetes mellitus type 2 patients. In the 2008 ACCORD trial, it was determined that intensive therapy (defined as a goal hemoglobin A1C less than 6.0%) did not significantly reduce major cardiovascular events and was associated with increased mortality and risk for hypoglycemia.[11] However, it should be noted that the intensive therapy group had proportionally more participants using rosiglitazone compared to the standard therapy group (91.2% versus 57.5%), thus possibly contributing to an increased incidence of cardiovascular events in the intensive therapy group.
The 2009 VADT study additionally studied the effect of intensive blood glucose control in a sample of 1791 veterans with poorly controlled diabetes mellitus type 2. More rigid glycemic control did not appear to have a significant effect on cardiovascular outcomes, although it did improve microalbuminuria compared to the standard therapy arm.[12] However, the results cannot be extrapolated to females since 97% of the study participants were male. Besides, there was a significant dropout (approximately 15%), limiting statistical power.
Regarding endogenous sources of insulin, insulinomas are often managed surgically. Evidence of an insulinoma should prompt workup or investigative effort into potential multiple endocrine neoplasia (MEN) disorders.
Go to:
Differential Diagnosis
If hypoglycemia is confirmed, the focus should be on correcting the hypoglycemia and identifying the underlying cause. In the workup of hypoglycemia, history should include medication and dietary adherence, medication changes, suspicion for acute kidney injury, or intentional/unintentional weight changes, especially weight loss.
Go to:
Prognosis
Severe insulin reactions may be proven fatal, so it is essential to contact EMS immediately when an individual is disoriented or unconscious. The patients with diabetes should also reach out to their clinicians if they begin to experience frequent hypoglycemic episodes, as they may need to have adjustments made to their medication regimen, meal plans, or even their exercise/activity regimen. Non-diabetic individuals who show signs or symptoms of hypoglycemia should contact their clinician to further evaluate their situation. Severe or prolonged hypoglycemia can be life-threatening, and in patients with diabetes, there is a correlation with increased mortality. In non-diabetic individuals experiencing reactive hypoglycemia, the prognosis is good.
Go to:
Complications
Complications of untreated hypoglycemia can lead to serious neurologic consequences, like seizures, brain damage and even death.
Go to:
Consultations
Most cases of hypoglycemia can be managed conservatively. Recurrent episodes of hypoglycemia with no apparent or obvious cause may warrant specialty consultation with an endocrinologist. Consultation with a diabetic educator may also be beneficial for the long-term management of diabetes and hypoglycemia.
Go to:
Deterrence and Patient Education
Patient education remains a pivotal component in the prevention of hypoglycemic episodes. Focus on preventing hypoglycemia should include patient education on signs and symptoms that constitute hypoglycemia and early recognition of these signs and symptoms. Patients may also need counseling on meal plans and exercise to manage their condition better.
Go to:
Pearls and Other Issues
Hypoglycemia in non-diabetic patients is uncommon. When it does occur, a critical illness[13], sustained alcohol use[14], malnutrition, and exogenous medications should be considered. Tumors may be a cause of hypoglycemia but are rare.[15]
Hypoglycemia is relatively common in neonates, particularly in mothers with uncontrolled diabetes. A 2017 study performed in Israel showed that 559 neonates out of 3595 live births were observed to have a glucose level of less than 47 mg/dL.[16] Gestational glucose intolerance in the mother is usually attributed to the presence of human placental lactogen. Pregnant women with impaired glucose tolerance not responsive to diet or exercise should be started on insulin. Insulin does not cross the placenta, although the fetus will be exposed to maternal hyperglycemia. Since the fetal pancreatic islet cells produce insulin starting at 10 weeks gestation, the fetal pancreas is capable of responding to hyperglycemia. Upon delivery, the newborn pancreas continues to secrete insulin, although maternal hyperglycemia is withdrawn. Subsequently, the neonate’s glucose will decrease, resulting in an insulin-glucose imbalance and hypoglycemia.
Go to:
Enhancing Healthcare Team Outcomes
An interprofessional approach to hypoglycemia is recommended. Adequate interprofessional measures to minimize hypoglycemic events involve participation and effective communication between the primary care physicians, physician assistants, nurse practitioners, endocrinologists, diabetes educators, pharmacists, specialty-trained diabetes nurses, the patient's family, nutritionists and/or dieticians, and the patient. The cornerstone of this management is the patient.
Patient education should address the importance of relatively detailed documentation regarding blood glucose levels, timing, units of insulin administered, and any pertinent notes such as increased/decreased food intake or exercise level relative to blood glucose measurements. Such documentation allows a primary care physician or an endocrinologist to make appropriate adjustments to diabetic medication therapy for optimizing blood glucose levels. Stability of blood glucose levels can be obtained with consistent dietary and exercise habits in addition to the appropriate timing of insulin therapy to avoid drastic spikes or dips in blood glucose levels. Hospitalized patients with newly diagnosed diabetes can be taught by the nurse diabetic educator how to do the insulin administration and self-injections while in the hospital. The educator should collaborate with the clinician managing the case so that appropriate patient education is achieved before discharge. Discussions with the patient and pharmacist will help decide the best medications to achieve safe glycemic control. Group education classes and local event planning can help patients with diabetes to learn and grow their knowledge among themselves and other members of the household.
Non-adherence to medication or diet is the most common cause of treatment failure. Patients should monitor themselves for signs or symptoms of hypoglycemia and always have sources of glucose (for example, hard candy, fruit juice) immediately available. Developing programs to educate healthcare staff has also been shown to provide better outcomes. Teaming up with local pharmacies or grocery stores can help overcome some of the barriers often encountered by patients with diabetes.
Patients should be advised to have fairly consistent exercise and dietary habits to avoid drastic changes in hour-to-hour blood glucose measurements.
With an interprofessional approach to hypoglycemia, outcomes will be optimized, leading to better patient quality of life. [Level 5]
Go to:
Review Questions
Access free multiple choice questions on this topic.
Comment on this article.
Go to:
References
1.
Koch CA, Petersenn S. Black swans - neuroendocrine tumors of rare locations. Rev Endocr Metab Disord. 2018 Jun;19(2):111-121. [PubMed]
2.
Marks V, Teale JD. Drug-induced hypoglycemia. Endocrinol Metab Clin North Am. 1999 Sep;28(3):555-77. [PubMed]
3.
Daughaday WH. Hypoglycemia due to paraneoplastic secretion of insulin-like growth factor-I. J Clin Endocrinol Metab. 2007 May;92(5):1616. [PubMed]
4.
Dardano A, Daniele G, Lupi R, Napoli N, Campani D, Boggi U, Del Prato S, Miccoli R. Nesidioblastosis and Insulinoma: A Rare Coexistence and a Therapeutic Challenge. Front Endocrinol (Lausanne). 2020;11:10. [PMC free article] [PubMed]
5.
Cryer PE. Hypoglycemia in type 1 diabetes mellitus. Endocrinol Metab Clin North Am. 2010 Sep;39(3):641-54. [PMC free article] [PubMed]
6.
Donnelly LA, Morris AD, Frier BM, Ellis JD, Donnan PT, Durrant R, Band MM, Reekie G, Leese GP., DARTS/MEMO Collaboration. Frequency and predictors of hypoglycaemia in Type 1 and insulin-treated Type 2 diabetes: a population-based study. Diabet Med. 2005 Jun;22(6):749-55. [PubMed]
7.
Cryer PE. Hypoglycemia in diabetes: pathophysiological mechanisms and diurnal variation. Prog Brain Res. 2006;153:361-5. [PubMed]
8.
Lee KT, Abadir PM. Failure of Glucose Monitoring in an Individual with Pseudohypoglycemia. J Am Geriatr Soc. 2015 Aug;63(8):1706-8. [PMC free article] [PubMed]
9.
La Sala L, Pontiroli AE. New Fast Acting Glucagon for Recovery from Hypoglycemia, a Life-Threatening Situation: Nasal Powder and Injected Stable Solutions. Int J Mol Sci. 2021 Sep 30;22(19) [PMC free article] [PubMed]
10.
Isaacs D, Clements J, Turco N, Hartman R. Glucagon: Its evolving role in the management of hypoglycemia. Pharmacotherapy. 2021 Jul;41(7):623-633. [PubMed]
11.
Action to Control Cardiovascular Risk in Diabetes Study Group. Gerstein HC, Miller ME, Byington RP, Goff DC, Bigger JT, Buse JB, Cushman WC, Genuth S, Ismail-Beigi F, Grimm RH, Probstfield JL, Simons-Morton DG, Friedewald WT. Effects of intensive glucose lowering in type 2 diabetes. N Engl J Med. 2008 Jun 12;358(24):2545-59. [PMC free article] [PubMed]
12.
Duckworth W, Abraira C, Moritz T, Reda D, Emanuele N, Reaven PD, Zieve FJ, Marks J, Davis SN, Hayward R, Warren SR, Goldman S, McCarren M, Vitek ME, Henderson WG, Huang GD., VADT Investigators. Glucose control and vascular complications in veterans with type 2 diabetes. N Engl J Med. 2009 Jan 08;360(2):129-39. [PubMed]
13.
Plečko D, Bennett N, Mårtensson J, Bellomo R. The obesity paradox and hypoglycemia in critically ill patients. Crit Care. 2021 Nov 01;25(1):378. [PMC free article] [PubMed]
14.
Shukla L, Reddy S, Kulkarni G, Chand PK, Murthy P. Alcohol Dependence, Hypoglycemia, and Transient Movement Disorders. Prim Care Companion CNS Disord. 2019 Jan 03;21(1) [PubMed]
15.
Yadav RS, Pokharel A, Gaire D, Shrestha S, Pokharel A, Pradhan S, Kansakar PBS. Multiple Endocrine Neoplasia Type 1 with Concomitant Existence of Malignant Insulinoma: A Rare Finding. Case Rep Endocrinol. 2021;2021:8842667. [PMC free article] [PubMed]
16.
Bromiker R, Perry A, Kasirer Y, Einav S, Klinger G, Levy-Khademi F. Early neonatal hypoglycemia: incidence of and risk factors. A cohort study using universal point of care screening. J Matern Fetal Neonatal Med. 2019 Mar;32(5):786-792. [PubMed]
Disclosure: Philip Mathew declares no relevant financial relationships with ineligible companies.
Disclosure: Deepu Thoppil declares no relevant financial relationships with ineligible companies.
Copyright © 2024, StatPearls Publishing LLC.
This book is distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0) ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ), which permits others to distribute the work, provided that the article is not altered or used commercially. You are not required to obtain permission to distribute this article, provided that you credit the author and journal.
Bookshelf ID: NBK534841PMID: 30521262
Share on FacebookShare on Twitter
Views
PubReader
Print View
Cite this Page
In this Page
Continuing Education Activity
Introduction
Etiology
Epidemiology
Pathophysiology
History and Physical
Evaluation
Treatment / Management
Differential Diagnosis
Prognosis
Complications
Consultations
Deterrence and Patient Education
Pearls and Other Issues
Enhancing Healthcare Team Outcomes
Review Questions
References
Bulk Download
Bulk download StatPearls data from FTP
Related information
PMC
PubMed
Similar articles in PubMed
Hypoglycemia (Nursing).
[StatPearls. 2024]
EMS Diabetic Protocols For Treat and Release.
[StatPearls. 2024]
Review Hypoglycemia.
[Obstet Gynecol Clin North Am. ...]
Recurrent hypoglycemia increases hepatic gluconeogenesis without affecting glycogen metabolism or systemic lipolysis in rat.
[Metabolism. 2022]
Review Hypoglycemia in Neonates, Infants, and Children.
[Endotext. 2000]
See reviews...
See all...
Recent Activity
ClearTurn Off
Hypoglycemia - StatPearls
Physiology, Synapse - StatPearls
Types of Eye Movements and Their Functions - Neuroscience
Physiology, Baroreceptors - StatPearls
Anatomy, Head and Neck: Eye Orbicularis Oculi Muscle - StatPearls
See more...
FOLLOW NCBI
Connect with NLM
National Library of Medicine
8600 Rockville Pike
Bethesda, MD 20894
Web Policies
FOIA
HHS Vulnerability Disclosure
Help
Accessibility
Careers
NLM
NIH
HHS
USA.gov | biology | 12304 | https://da.wikipedia.org/wiki/Sukkersyge | Sukkersyge | Sukkersyge, med den medicinske betegnelse diabetes mellitus, er en gruppe sygdomme, hvor blodets indhold af sukker (glukose) er øget ud over det normale, blodsukkeret. Dette skyldes enten, at bugspytkirtlen ikke producerer nok af hormonet insulin, eller at kroppens celler ikke responderer på det producerede insulin. Det høje blodsukker medfører de klassiske symptomer: Hyppig vandladning samt øget følelse af tørst og sult.
Der er tre hovedgrupper af sukkersyge:
Type 1-diabetes mellitus. Skyldes, at kroppen ikke producerer insulin og kræver injektioner af insulin eller anvendelse af en insulinpumpe. Denne type blev tidligere kaldt insulin-afhængig diabetes og ungdomssukkersyge. I Danmark har omkring 35.000 type 1-diabetes.
Type 2-diabetes mellitus. Skyldes insulinresistens, hvor kroppens celler ikke kan udnytte insulin, nogle gange kombineret med nedsat produktion af insulin. Blev tidligere kaldt ikke-insulin afhængig diabetes eller gammelmandssukkersyge.
Type 3 diabetes, Alzheimers sygdom eller Alzheimers syndrom. Diabetes type 1 og 2 kan udvikle sig til diabetes type 3.
Gestationel diabetes. Denne type opstår, når gravide kvinder uden tidligere sukkersyge udvikler højt blodsukker. Er nogle gange forgænger til udvikling af type 2-diabetes.
Svangerskabsdiabetes (Gestationel diabetes). Denne type diabetes er en reaktion på de hormonelle ændringer, der sker under graviditeten. De hormoner, der dannes i moderkagen, kan sænke din krops følsomhed over for insulin. Dette kan resultere i højt blodsukker under graviditeten.
Type 1, type 2 og svangerskabsdiabetes er meget almindelige, men de er ikke de eneste typer diabetes. Der er flere andre typer diabetes, og nogle få, der ikke officielt er klassificeret som typer.
Det har været muligt at behandle alle former af sukkersyge, siden insulin blev tilgængeligt i 1921, og type 2-diabetes kan behandles med perorale lægemidler. Både type 1 og 2 er kroniske sygdomme, da der ikke findes nogen kur. Gestationel diabetes går som oftest over efter fødslen. Ubehandlet sukkersyge medfører mange komplikationer. Akutte komplikationer omfatter hypoglykæmi og diabetisk ketoacidose. Kroniske komplikationer omfatter hjerte-kar-sygdom, kronisk nyresvigt og diabetisk retinopati (nethindeskade). Tilstrækkelig behandling af sukkersyge er derfor vigtig, ligesom kontrol med blodtryk og livsstilsfaktorer såsom rygestop og opretholdelse af en sund kropsvægt.
Klassificering
Diabetes mellitus klassificeres i fire brede kategorier: Type 1, type 2, gestationel diabetes samt "andre specifikke typer". Disse "andre specifikke typer" er en samling af flere individuelle sjældne årsager. Udtrykket "sukkersyge" eller "diabetes" henviser normalt til diabetes mellitus. Den mere sjældne diabetes insipidus har tilsvarende symptomer som diabetes mellitus, men skyldes ikke forstyrrelser i sukkerstofskiftet.
Type 1-diabetes
Type 1-diabetes mellitus er karakteriseret ved et tab af de insulin-producerende betaceller i bugspytkirtelen, som fører til insulinmangel. Denne type kan yderligere opdeles i immun-medieret og idiopatisk (ukendt årsag til betacelletab). Hovedparten af type 1-tilfælde er immun-medieret, hvor tabet af betaceller skyldes et T-celle-medieret autoimmunt angreb. Der er ingen kendt forebyggelse imod type 1-diabetes, som forårsager 5-10 % af diabetes mellitus-tilfælde i Danmark. De fleste mennesker er raske med en sund kropsvægt, når sygdommen opstår. Følsomheden over for insulin er som oftest normal, særligt i de tidlige stadier af sygdommen. Type 1-diabetes kan ramme både børn og voksne, men blev traditionelt kaldt "ungdomssukkersyge" fordi sygdommen som oftest opstår hos børn.
Type 2-diabetes
Type 2-diabetes mellitus er karakteriseret ved nedsat insulinfølsomhed, som kan være kombineret med nedsat insulinproduktion. Den nedsatte følsomhed over for insulin skyldes sandsynligvis insulinreceptoren. De nærmere mekanismer er dog stadig ukendte. En anden mulig årsag til insulinresistens er det insulinnedbrydende enzym, IDE (efter engelsk: insulin-degrading enzyme). Type 2-diabetes er den mest hyppigt forekommende type og står for 85 % af diabetes mellitus-tilfælde hos danske patienter.
I de tidlige stadier af type 2 ses primært nedsat insulinfølsomhed. I dette stadium kan det øgede niveau af sukker i blodet (hyperglykæmien) afhjælpes ved livsstilsændringer og/eller perorale lægemidler, som øger insulinfølsomheden eller sænker produktionen af glukose i leveren.
Gestationel diabetes
Gestationel diabetes ligner type 2-diabetes på mange måder og skyldes en nedsat insulinfølsomhed eventuelt kombineret med nedsat insulinsekretion. Denne type opstår ved 2-3 % af alle graviditeter og forsvinder som oftest efter fødslen. Gestationel diabetes kan behandles, men kræver forsigtig overvågning under hele graviditeten. Omkring 40 % af de ramte kvinder udvikler type 2-diabetes senere i livet.
Andre specifikke typer
Der findes over 30 typer i denne kategori. Forskellige genetiske mutationer kan føre til defekter i funktionen af betacellerne. Enhver sygdom, som skader bugspytkirtlen, kan føre til diabetes (fx betændelse i bugspytkirtlen eller cystisk fibrose). Mange lægemidler hæmmer produktionen af insulin, og visse giftstoffer kan skade betacellerne.
Symptomer og komplikationer
De klassiske symptomer på ubehandlet diabetes er vægttab, øget vandladning (polyuri) samt øget følelse af tørst (polydipsi) og sult (polyfagi). Symptomerne udvikler sig hurtigt (på uger eller måneder) i type 1-diabetes, mens de typisk udvikler sig over meget længere tid i type 2-diabetes.
Højt indhold af glukose (hyperglykæmi) i blodet over længere tid kan forårsage absorption af glukose ind i linsen i øjet, hvilket medfører ændringer i formen af linsen og dermed også synsændringer. Sløret syn er et hyppigt symptom, som medfører en diagnose for diabetes. Type 1-diabetes bør altid mistænkes i tilfælde af pludseligt opståede synsændringer, mens ændringerne i type 2 opstår mere gradvist. Forskellige type hududslæt kan opstå ved diabetes og kaldes samlet diabetiske dermadromer.
Diabetiske akuttilfælde
Mennesker med diabetes (oftest type 1) kan også opleve diabetisk ketoacidose, en tilstand med forstyrrelser i regulering af stofskiftet, som karakteriseres ved lugten af acetone i udåndingsluften, hurtig, dyb vejrtrækning, kvalme, opkast, abdominalsmerter og ændret bevidsthedstilstand.
En sjælden, men lige så alvorlig tilstand kaldes hyperosmolær ikke-ketotisk tilstand og er mere almindelig hos type 2-diabetikere. Denne tilstand skyldes hovedsageligt dehydrering.
Komplikationer
Alle former for diabetes øger risikoen for komplikationer på lang sigt. Disse udvikler sig typisk over mange år (10-20), men kan være de første symptomer hos personer, der ikke er blevet diagnosticeret tidligere. De primære langtidskomplikationer er relateret til blodkar. Diabetes fordobler risikoen for hjerte-kar-sygdom. De primære makrovaskulære (åreforkalkning i de store blodkar) komplikationer er iskæmisk hjertesygdom (angina pectoris og myokardieinfarkt), slagtilfælde og perifær vaskulær sygdom.
Diabetes medfører også mikrovaskulære komplikationer, som er skader i de små blodkar (kapillærer). Diabetisk retinopati (skader i øjets nethinde) fører til synsændringer, nedsat syn og muligvis blindhed. Diabetisk nefropati (skader i nyrerne) fører til arvævsdannelse i nyrerne, tab af proteiner igennem urinen og til sidst kronisk nyresvigt, som kræver dialyse. Diabetisk neuropati (skader i nerver) fører til følelsesløshed, paræstesier (følelsen af nåle i huden, som når en kropsdel "sover") og smerter i fødderne. Sammen med den perifære vaskulære sygdom i fødderne øger neuropatien risikoen for diabetes-relaterede fodproblemer (sårdannelse og smerter), som kan være svære at behandle og kan kræve amputation.
Overdødelighed
Diabetikere har en højere overdødelighed end resten af befolkningen. Det betyder, at diabetikere gennemsnitligt dør tidligere end personer, der ikke har diabetes. I 1997 var overdødeligheden på 1,91, men i 2012 var overdødeligheden nedbragt til 1,49. Det betyder, at diabetesbefolkningen i 2012 i gennemsnit har 49 % flere dødsfald end resten af befolkningen i samme aldersinterval.
Årsager
Årsagen til sukkersyge afhænger af typen.
Type 1-diabetes er delvist arveligt, men forekommer desuden at blive udløst af visse infektioner. En del bevis peger på virussen Coxsackie B4 som en årsag. Den individuelle følsomhed over for disse udløsere kan spores til HLA-genotyper (genotypen i histokompatibilitetskomplekset). Det er dog ikke nok at arve den "forkerte" genotype. Udvikling af type 1-diabetes kræver, at man også bliver udsat for en udløser i omgivelserne. Udvikling af type 1-diabetes er ikke relateret til livsstil.
Type 2-diabetes skyldes primært livsstil og arv. Overvægt og en fysisk inaktiv livsstil øger betydeligt risikoen for udvikling af sygdommen.
Listen nedenfor viser forskellige specifikke årsager til diabetes:
Genetiske defekter i betacelle funktion:
Mutationer i mitokondrie DNA
Genetiske defekter i funktionen af insulin:
Defekter i omdannelsen af proinsulin
Mutationer i genet for insulin
Mutationer i genet for insulinreceptoren
Eksokrine defekter i bugspytkirtlen:
Kronisk pankreatitis (betændelse i bugspytkirtlen)
Pankreatektomi (fjernelse af bugspytkirtlen)
Kræft i bugspytkirtlen
Cystisk fibrose
Hæmokromatose (forhøjede jern niveauer)
Forstyrrelser i det endokrine system:
For meget væksthormon
Cushings syndrom
Hyperthyroidisme (forhøjet stofskifte)
Fæokromocytom (katekolamin-udskillende tumor)
Glukagonom (glukagon-udskillende tumor)
Infektioner:
Cytomegalovirus
Coxsackievirus B
Lægemidler:
Binyrebarkhormon
Thyroidhormon
Beta-adrenerge agonister
Statiner
Sygdomsmekanisme
Insulin er det primære hormon der regulerer optagelsen af glukose fra blodet og ind i cellerne. Insulinmangel eller manglende følsomhed overfor insulin spiller derfor en central rolle i alle former for sukkersyge.
Mennesker kan fordøje visse kulhydrater. Stivelse og visse disakkarider som sukrose, bliver omdannet hurtigt til simple monosakkarider, oftest glukose, som er den primære energikilde i kroppen. Resten bliver videreført til tyktarmen hvor de bliver bearbejdet af tarmfloraen. Insulin frigives til blodet fra betacellerne, som findes i et specielt kirtelvæv i bugspytkirtlen der kaldes de Langerhanske øer, som et respons på stigende glukose i blodet der typisk sker efter indtagelse af et måltid. Insulin signalerer til kroppens celler at de skal optage den cirkulerende glukose, til anvendelse som brændstof, omdannelse til andre molekyler eller til lagring.
Insulin er også det primære signal til omdannelse af glukose til glykogen til lagring i leveren og musklerne. Lave niveauer af glukose i blodet nedsætter udskillelsen af insulin fra betacellerne og i nedbrydningen af glykogen til glukose. Dette kontrolleres af hormonet glukagon, som virker modsat insulin. Glukose produceret i leveren fra glykogen bliver således frigivet til blodet.
Høje insulinniveauer øger anaboliske (opbyggende) processer, som cellevækst, proteinsyntese og fedtlagring. Insulin er det primære signal der omdanner mange stofskifteprocesser fra en katabolisk (nedbrydende) til en anabolisk tilstand.
Hvis mængden af insulin er utilstrækkelig, hvis cellerne responderer dårligt på insulinen eller hvis insulinmolekylerne er defekte, vil glukosen i blodet ikke medføre disse normale virkninger, så glukosen bliver ikke optaget i cellerne, og den bliver ikke lagret i lever og muskler. Slutresultatet bliver vedvarende høje niveauer af glukose i blodet, nedsat proteinsyntese, som fører til vægttab, og andre forstyrrelser i stofskiftet som acidose.
Den forhøjede glukosekoncentrationen i blodet når for høje niveauer til at genoptagelsen af glukose i nyrerne bliver ufuldstændig, og noget af glukosen går tabt igennem urinen. Dette forårsager en osmotisk diurese og således en øget urindannelse (polyuri). På grund af tabet af væske vil der ske en dehydrering som fører til øget tørst (polydipsi).
Diagnose
Diabetes mellitus er karakteriseret ved vedvarende forhøjet blodsukker. Sygdommen diagnosticeres ved en af følgende metoder:
Glykosyleret hæmoglobin (HbA1c) ≥ 6,5 %
Fastende blodsukker (plasma glukose) ≥ 7,0 mmol/L
Ikke-fastende blodsukker (plasma glukose) ≥ 11,1 mmol/L
Et positivt resultat, ved fravær af tydelig hyperglykæmi, bør bekræftes ved at gentage en af metoderne på en anden dag. Ikke-fastende blodsukker måles ved en såkaldt oral glukose tolerance test (OGTT), hvor personen indtager 75 g glukose peroralt og man efter to timer måler blodsukkeret.
Verdenssundhedsorganisationen (WHO) indførte i 2011 glykolyseret hæmoglobin (HbA1c) som diagnostisk kriterium. Sundhedsstyrelsen har på basis heraf anbefalet at HbA1c bør anvendes som primært diagnostisk redskab i Danmark.
Personer med fastende blodsukker fra 6,1 til 6,9 mmol/L betragtes som havende forhøjet fastende glukose (IFG, fra engelsk: Impaired fasting glucose). Personer med ikke-fastende blodsukker over 7,8 mmol/L (men ikke over 11,1 mmol/L) betragtes som havende nedsat glukose tolerance (IGT, fra engelsk: Impaired glucose tolerance). Disse to præ-diabetiske tilstande, særligt den sidstnævnte, er betydelige risikofaktorer for udvikling af diabetes mellitus, såvel som hjerte-kar-sygdom.
Behandling
Sukkersyge er en kronisk sygdom som ikke kan kureres undtagen i få specielle tilfælde. Behandlingen fokuserer på at holde blodsukkerniveauerne så tæt på normalt som muligt, uden at forårsage hypoglykæmi. Dette kan normalt opnås igennem kost, motion, og brug af passende lægemidler (insulin ved type 1-diabetes, perorale antidiabetika, og i visse tilfælde insulin, ved type 2-diabetes). Semaglutid, et hormonlignende lægemiddel (Ozempic, Wegovy eller Rybelsus) til behandling af type 2-diabetes.
Vejledning af patienten, forståelse og deltagelse er afgørende, da komplikationer ved sukkersyge er sjældne og mindre alvorlige hos personer med et velreguleret blodsukker. Målet for behandlingen er i startfaserne af sygdommen et HbA1c under 6,5 %. Dette mål er dog svært at opnå senere i sygdomsforløbet og det kan derfor blive relevant at sigte efter et lidt højere, mere realistisk niveaue. Der lægges også vægt på andre sundhedsproblemer som kan forstærke komplikationerne ved sukkersyge. Disse omfatter rygning, forhøjet kolesterol, overvægt, forhøjet blodtryk og mangel på regelmæssig motion. Specialiseret fodtøj anvendes ofte for at mindske risikoen for sårdannelse i foden hos personer med risiko herfor.
Kost
Den kost der anbefales til diabetikere er generelt den samme sunde kost som anbefales til andre mennesker. Dette vil sige varieret, fiberrig, fedt- og kaloriefattig kost, desuden lavt indtag af fruktose og dermed også almindeligt sukker (sukrose). Hos personer der ikke behandles med insulin, er antallet af måltider om dagen ikke vigtig for reguleringen af blodsukker. Hos personer i behandling med insulin, anbefales regelmæssige måltider med konstante mængder kulhydrat, da vores indtag af især kulhydrater får blodsukkeret til at stige.
Indtil 2008 kunne diabetikere i Danmark få et økonomisk tilskud til kost, så de havde råd til at købe de rette fødevarer. En undersøgelse foretaget af Servicestyrelsen i 2007-08 viste dog, at dansk normalkost med tiden var blevet lige så dyr som diabeteskosten, så derfor ydes der ikke længere et sådant tilskud.
Fysisk aktivitet
Regelmæssig motion øger følsomheden overfor insulin i vævene og kan reducere HbA1c med omkring 0,6 %. Ny forskning fra Syddansk Universitet og Harvard School of Public Health viser, at styrketræning kan bruges til at forebygge type 2-diabetes.
Lægemidler
Perorale antidiabetika
Metformin (oprindeligt handelsnavn Glucophage) anbefales ofte som førstevalg til behandling af type 2-diabetes, da det har vist sig at sænke risikoen for hjerte-kar-sygdom og død. Metformin bevirker også vægttab, som er en fordel ved behandling af overvægtige. Er virkningen af metformin ikke tilstrækkelig, kan behandlingen kombineres med et sulfonylurinstof (glibenclamid, gliclazid, glimepirid, glipizid eller tolbutamid).
Andre nyere perorale antidiabetika der kan anvendes som tillæg eller alternativ omfatter glitazoner, DPP-4-hæmmere (gliptiner), α-glucosidasehæmmere, SGLT-2-hæmmere og meglitinider.
Insulin
Type 1-diabetes behandles typisk med en kombination af kort- og langtidsvirkende insulin, eller syntetiske insulin analoger. Hos de personer med type 2-diabetes der har behov for insulin, anvendes ofte et langtidsvirkende præparat i starten og et korttidsvirkende præparat kan derefter lægges til ved utilstrækkelig effekt. Behandling med metformin fortsættes som regel, mens andre perorale antidiabetika stoppes.
Epidemiologi
Type 2-diabetes er en hyppigt forekommende sygdom i Danmark. Omkring 240.000 personer har diagnosticeret type 2-diabetes i Danmark, mens der er yderligere omkring 100.000-200.000 udiagnosticerede tilfælde.
Noter
Eksterne henvisninger
Netdoktor.dk om diabetes
Diabetesforeningen i Danmark
Se også
Insulinkalkulator
Endokrine sygdomme | danish | 0.332111 |
bacterium_infect_another/Bdellovibrio.txt |
Bdellovibrio is a genus of Gram-negative, obligate aerobic bacteria. One of the more notable characteristics of this genus is that members can prey upon other Gram-negative bacteria and feed on the biopolymers, e.g. proteins and nucleic acids, of their hosts. They have two lifestyles: a host-dependent, highly mobile phase, the "attack phase", in which they form "bdelloplasts" in their host bacteria; and a slow-growing, irregularly shaped, host-independent form.
Bdellovibrio bacteriovorus[edit]
The most well studied of these is Bdellovibrio bacteriovorus, which is found almost exclusively in host dependent growth in nature. In this free swimming attack form after searching for prey using its pili, it burrows through the host outer membrane/ peptidoglycan cell wall and enters the periplasmic space. The Bdellovibrio bacterium then forms a structure called a bdelloplast. This bdelloplast is created as the host cell is modified to become spherical in shape. Inside the bdelloplast, the singular large flagellum of the predatory Bdellovibrio is lost. The host cell is then rapidly killed allowing the passage of molecules from the interior of the host cytoplasm through to the periplasm freely, and the periplasm dwelling Bdellovibrio to feed. Using some of these molecules the Bdellovibrio creates a protective environment by reinforcing the peptidoglycan cell wall of the host in which it now dwells using amidases and transpeptidases. After around 4hrs, depending on ambient temperature, the Bdellovibrio has increased in size dramatically through this nourishment. It divides to replicate and then leaves via a final lysis of the host's cell wall and membranes. The newly emerging Bdellovibrio use their newly grown powerful flagella to swim away and find the next suitable host. Because of this intermittent bdelloplast stage, and momentary parasitic phase (15-20 mins), Bdellovibrio could be considered bacterial predators or parasites.
Bdellovibrio bacteriovorus was first described by Stolp and Petzold in 1962. In 2012 another member of the Bdellovibrio species was identified "Bdellovibrio tiberius" of the River tiber. This species is more capable of host-independent growth.
Little is known of Bdellovibrio exovorus, an extra-parasitic bdellovibrio, which cannot enter its prey, and does not form Bdelloplasts.
Appearance[edit]
Under a light microscope, host-dependent Bdellovibrio appears to be a comma-shaped motile rod that is about 0.3–0.5 by 0.5–1.4 µm in size with a barely discernible flagellum. Bdellovibrio show up as a growing clear plaque in an E. coli “lawn”. Notably, Bdellovibrio has a sheath that covers its flagellum – a rare feature for bacteria. Flagellar motion stops once Bdellovibrio has penetrated its prey, and the flagella is then shed.
Host-independent Bdellovibrio appear amorphous, and larger than the predatory phase.
Culture conditions[edit]
B. bacteriovorus appears to be ubiquitous in nature and manmade habitats. They have been found in soil samples, rhizosphere of plant roots, rivers, oceans, sewage, intestines and feces of birds and mammals, and even in oyster shells and the gills of crabs. B. bacteriovorus are able to thrive in almost any habitat, the general requirements are that there needs to be oxygen and some other Gram-negative bacteria present in its environment. Its optimal temperature is between 28-30°C, making B. bacteriovorus a mesophile. Bdellovibrio is grown in the laboratory in its stationary HI (host-independent) phase at 29°C on yeast peptone broth agar. Host-dependent (predatory) cultures are grown with a population of E. coli S-17 at 29°C for 16 hrs. They may also be cultured using YPSC (yeast extract, peptone, sodium acetate, calcium chloride) overlays or prey lysates.
Life cycle and parasitism[edit]
Bdellovibrio life cycle. The Bdellovibrio attaches to a Gram-negative bacterium after contact, and penetrates into the prey's periplasmic space. Once inside, elongation occurs and progeny cells are released within 4 hours.
Bdellovibrio cells can swim as fast as 160 µm/s, or over 100 times their body-length per second. It swims using a single sheathed polar flagellum with a characteristic dampened filament waveform. Bdellovibrio attacks other Gram-negative bacteria by attaching itself to the prey cell's outer membrane and peptidoglycan layer, after which it creates a small hole in the outer membrane. The Bdellovibrio cell then enters the host periplasmic space. It remains reversibly attached to it for a short "recognition" period.
After the recognition period, it becomes irreversibly attached via the pole opposite the flagellum. Once inside the periplasm, the Bdellovibrio cell seals the membrane hole and converts the host cell to a spherical morphology, this is due to secretion of L,D transpeptidases which breaks the peptidoglycan apart, and therefore causes the cell to become amorphous. The two-cell complex formed is called a bdelloplast. The Bdellovibrio cell uses hydrolytic enzymes to break down the host cell molecules, which it uses to grow filamentously. When the host cell nutrients are exhausted, the filament septates to form progeny Bdellovibrios. The progeny become motile before they lyse the host cell and are released into the environment. The entire life cycle takes three to four hours, and produces an average of 3–6 progeny cells from a single E. coli, or up to 90 from larger prey such as filamentous E. coli.
Targets of Bdellovibrio species, including Vibrio vulnificus, may undergo co-infection by Bdellovibrio and bacteriophage. Although the Bdellovibrio rounding of prey is thought to be evolved to reduce co-infection of multiple Bdellovibrio, larger prey that do not round may be infected by multiple Bdello's.
Genomics[edit]
The genome of Bdellovibrio bacteriovorus HD100 was sequenced in 2004. The HD100 genome is 3782950 nucleotides long, larger than expected given its small size.
See also[edit]
List of bacterial orders
List of bacteria genera | biology | 2572804 | https://sv.wikipedia.org/wiki/Heteracris%20brevipennis | Heteracris brevipennis | Heteracris brevipennis är en insektsart som först beskrevs av Bolívar, I. 1914. Heteracris brevipennis ingår i släktet Heteracris och familjen gräshoppor.
Underarter
Arten delas in i följande underarter:
H. b. brevipennis
H. b. laticercus
H. b. nyambeniensis
Källor
Gräshoppor
brevipennis | swedish | 1.036443 |
bacterium_infect_another/Bacteria.txt |
Bacteria (/bækˈtɪəriə/ ; sg.: bacterium) are ubiquitous, mostly free-living organisms often consisting of one biological cell. They constitute a large domain of prokaryotic microorganisms. Typically a few micrometres in length, bacteria were among the first life forms to appear on Earth, and are present in most of its habitats. Bacteria inhabit soil, water, acidic hot springs, radioactive waste, and the deep biosphere of Earth's crust. Bacteria play a vital role in many stages of the nutrient cycle by recycling nutrients and the fixation of nitrogen from the atmosphere. The nutrient cycle includes the decomposition of dead bodies; bacteria are responsible for the putrefaction stage in this process. In the biological communities surrounding hydrothermal vents and cold seeps, extremophile bacteria provide the nutrients needed to sustain life by converting dissolved compounds, such as hydrogen sulphide and methane, to energy. Bacteria also live in mutualistic, commensal and parasitic relationships with plants and animals. Most bacteria have not been characterised and there are many species that cannot be grown in the laboratory. The study of bacteria is known as bacteriology, a branch of microbiology.
Like all animals, humans carry vast numbers (approximately 10 to 10) of bacteria. Most are in the gut, though there are many on the skin. Most of the bacteria in and on the body are harmless or rendered so by the protective effects of the immune system, and many are beneficial, particularly the ones in the gut. However, several species of bacteria are pathogenic and cause infectious diseases, including cholera, syphilis, anthrax, leprosy, tuberculosis, tetanus and bubonic plague. The most common fatal bacterial diseases are respiratory infections. Antibiotics are used to treat bacterial infections and are also used in farming, making antibiotic resistance a growing problem. Bacteria are important in sewage treatment and the breakdown of oil spills, the production of cheese and yogurt through fermentation, the recovery of gold, palladium, copper and other metals in the mining sector, as well as in biotechnology, and the manufacture of antibiotics and other chemicals.
Once regarded as plants constituting the class Schizomycetes ("fission fungi"), bacteria are now classified as prokaryotes. Unlike cells of animals and other eukaryotes, bacterial cells do not contain a nucleus and rarely harbour membrane-bound organelles. Although the term bacteria traditionally included all prokaryotes, the scientific classification changed after the discovery in the 1990s that prokaryotes consist of two very different groups of organisms that evolved from an ancient common ancestor. These evolutionary domains are called Bacteria and Archaea.
Etymology
Rod-shaped Bacillus subtilis
The word bacteria is the plural of the Neo-Latin bacterium, which is the Latinisation of the Ancient Greek βακτήριον (baktḗrion), the diminutive of βακτηρία (baktēría), meaning "staff, cane", because the first ones to be discovered were rod-shaped.
Origin and early evolution
Main article: Evolution of bacteria
Further information: Earliest known life forms, Evolutionary history of life, and Timeline of evolution
Phylogenetic tree of Bacteria, Archaea and Eucarya, with the last universal common ancestor (LUCA) at the root.
The ancestors of bacteria were unicellular microorganisms that were the first forms of life to appear on Earth, about 4 billion years ago. For about 3 billion years, most organisms were microscopic, and bacteria and archaea were the dominant forms of life. Although bacterial fossils exist, such as stromatolites, their lack of distinctive morphology prevents them from being used to examine the history of bacterial evolution, or to date the time of origin of a particular bacterial species. However, gene sequences can be used to reconstruct the bacterial phylogeny, and these studies indicate that bacteria diverged first from the archaeal/eukaryotic lineage. The most recent common ancestor (MRCA) of bacteria and archaea was probably a hyperthermophile that lived about 2.5 billion–3.2 billion years ago. The earliest life on land may have been bacteria some 3.22 billion years ago.
Bacteria were also involved in the second great evolutionary divergence, that of the archaea and eukaryotes. Here, eukaryotes resulted from the entering of ancient bacteria into endosymbiotic associations with the ancestors of eukaryotic cells, which were themselves possibly related to the Archaea. This involved the engulfment by proto-eukaryotic cells of alphaproteobacterial symbionts to form either mitochondria or hydrogenosomes, which are still found in all known Eukarya (sometimes in highly reduced form, e.g. in ancient "amitochondrial" protozoa). Later, some eukaryotes that already contained mitochondria also engulfed cyanobacteria-like organisms, leading to the formation of chloroplasts in algae and plants. This is known as primary endosymbiosis.
Habitat
Bacteria are ubiquitous, living in every possible habitat on the planet including soil, underwater, deep in Earth's crust and even such extreme environments as acidic hot springs and radioactive waste. There are thought to be approximately 2×10 bacteria on Earth, forming a biomass that is only exceeded by plants. They are abundant in lakes and oceans, in arctic ice, and geothermal springs where they provide the nutrients needed to sustain life by converting dissolved compounds, such as hydrogen sulphide and methane, to energy. They live on and in plants and animals. Most do not cause diseases, are beneficial to their environments, and are essential for life. The soil is a rich source of bacteria and a few grams contain around a thousand million of them. They are all essential to soil ecology, breaking down toxic waste and recycling nutrients. They are even found in the atmosphere and one cubic metre of air holds around one hundred million bacterial cells. The oceans and seas harbour around 3 x 10 bacteria which provide up to 50% of the oxygen humans breathe. Only around 2% of bacterial species have been fully studied.
Extremophile bacteria
Habitat
Species
Reference
Cold (minus 15 °C Antarctica)
Cryptoendoliths
Hot (70–100 °C geysers)
Thermus aquaticus
Radiation, 5MRad
Deinococcus radiodurans
Saline, 47% salt (Dead Sea, Great Salt Lake)
several species
Acid pH 3
several species
Alkaline pH 12.8
betaproteobacteria
Space (6 years on a NASA satellite)
Bacillus subtilis
3.2 km underground
several species
High pressure (Mariana Trench – 1200 atm)
Moritella, Shewanella and others
Morphology
Further information: Bacterial cell structure § Cell morphology
Bacteria display many cell morphologies and arrangements
Size. Bacteria display a wide diversity of shapes and sizes. Bacterial cells are about one-tenth the size of eukaryotic cells and are typically 0.5–5.0 micrometres in length. However, a few species are visible to the unaided eye—for example, Thiomargarita namibiensis is up to half a millimetre long, Epulopiscium fishelsoni reaches 0.7 mm, and Thiomargarita magnifica can reach even 2 cm in length, which is 50 times larger than other known bacteria. Among the smallest bacteria are members of the genus Mycoplasma, which measure only 0.3 micrometres, as small as the largest viruses. Some bacteria may be even smaller, but these ultramicrobacteria are not well-studied.
Shape. Most bacterial species are either spherical, called cocci (singular coccus, from Greek kókkos, grain, seed), or rod-shaped, called bacilli (sing. bacillus, from Latin baculus, stick). Some bacteria, called vibrio, are shaped like slightly curved rods or comma-shaped; others can be spiral-shaped, called spirilla, or tightly coiled, called spirochaetes. A small number of other unusual shapes have been described, such as star-shaped bacteria. This wide variety of shapes is determined by the bacterial cell wall and cytoskeleton and is important because it can influence the ability of bacteria to acquire nutrients, attach to surfaces, swim through liquids and escape predators.
The range of sizes shown by prokaryotes (Bacteria), relative to those of other organisms and biomolecules.
Multicellularity. Most bacterial species exist as single cells; others associate in characteristic patterns: Neisseria forms diploids (pairs), streptococci form chains, and staphylococci group together in "bunch of grapes" clusters. Bacteria can also group to form larger multicellular structures, such as the elongated filaments of Actinomycetota species, the aggregates of Myxobacteria species, and the complex hyphae of Streptomyces species. These multicellular structures are often only seen in certain conditions. For example, when starved of amino acids, myxobacteria detect surrounding cells in a process known as quorum sensing, migrate towards each other, and aggregate to form fruiting bodies up to 500 micrometres long and containing approximately 100,000 bacterial cells. In these fruiting bodies, the bacteria perform separate tasks; for example, about one in ten cells migrate to the top of a fruiting body and differentiate into a specialised dormant state called a myxospore, which is more resistant to drying and other adverse environmental conditions.
Biofilms. Bacteria often attach to surfaces and form dense aggregations called biofilms, and larger formations known as microbial mats. These biofilms and mats can range from a few micrometres in thickness to up to half a metre in depth, and may contain multiple species of bacteria, protists and archaea. Bacteria living in biofilms display a complex arrangement of cells and extracellular components, forming secondary structures, such as microcolonies, through which there are networks of channels to enable better diffusion of nutrients. In natural environments, such as soil or the surfaces of plants, the majority of bacteria are bound to surfaces in biofilms. Biofilms are also important in medicine, as these structures are often present during chronic bacterial infections or in infections of implanted medical devices, and bacteria protected within biofilms are much harder to kill than individual isolated bacteria.
Cellular structure
Further information: Bacterial cell structure
Structure and contents of a typical Gram-positive bacterial cell (seen by the fact that only one cell membrane is present).
Intracellular structures
The bacterial cell is surrounded by a cell membrane, which is made primarily of phospholipids. This membrane encloses the contents of the cell and acts as a barrier to hold nutrients, proteins and other essential components of the cytoplasm within the cell. Unlike eukaryotic cells, bacteria usually lack large membrane-bound structures in their cytoplasm such as a nucleus, mitochondria, chloroplasts and the other organelles present in eukaryotic cells. However, some bacteria have protein-bound organelles in the cytoplasm which compartmentalize aspects of bacterial metabolism, such as the carboxysome. Additionally, bacteria have a multi-component cytoskeleton to control the localisation of proteins and nucleic acids within the cell, and to manage the process of cell division.
Many important biochemical reactions, such as energy generation, occur due to concentration gradients across membranes, creating a potential difference analogous to a battery. The general lack of internal membranes in bacteria means these reactions, such as electron transport, occur across the cell membrane between the cytoplasm and the outside of the cell or periplasm. However, in many photosynthetic bacteria, the plasma membrane is highly folded and fills most of the cell with layers of light-gathering membrane. These light-gathering complexes may even form lipid-enclosed structures called chlorosomes in green sulfur bacteria.
An electron micrograph of Halothiobacillus neapolitanus cells with carboxysomes inside, with arrows highlighting visible carboxysomes. Scale bars indicate 100 nm.
Bacteria do not have a membrane-bound nucleus, and their genetic material is typically a single circular bacterial chromosome of DNA located in the cytoplasm in an irregularly shaped body called the nucleoid. The nucleoid contains the chromosome with its associated proteins and RNA. Like all other organisms, bacteria contain ribosomes for the production of proteins, but the structure of the bacterial ribosome is different from that of eukaryotes and archaea.
Some bacteria produce intracellular nutrient storage granules, such as glycogen, polyphosphate, sulfur or polyhydroxyalkanoates. Bacteria such as the photosynthetic cyanobacteria, produce internal gas vacuoles, which they use to regulate their buoyancy, allowing them to move up or down into water layers with different light intensities and nutrient levels.
Extracellular structures
Further information: Cell envelope
Around the outside of the cell membrane is the cell wall. Bacterial cell walls are made of peptidoglycan (also called murein), which is made from polysaccharide chains cross-linked by peptides containing D-amino acids. Bacterial cell walls are different from the cell walls of plants and fungi, which are made of cellulose and chitin, respectively. The cell wall of bacteria is also distinct from that of achaea, which do not contain peptidoglycan. The cell wall is essential to the survival of many bacteria, and the antibiotic penicillin (produced by a fungus called Penicillium) is able to kill bacteria by inhibiting a step in the synthesis of peptidoglycan.
There are broadly speaking two different types of cell wall in bacteria, that classify bacteria into Gram-positive bacteria and Gram-negative bacteria. The names originate from the reaction of cells to the Gram stain, a long-standing test for the classification of bacterial species.
Gram-positive bacteria possess a thick cell wall containing many layers of peptidoglycan and teichoic acids. In contrast, Gram-negative bacteria have a relatively thin cell wall consisting of a few layers of peptidoglycan surrounded by a second lipid membrane containing lipopolysaccharides and lipoproteins. Most bacteria have the Gram-negative cell wall, and only members of the Bacillota group and actinomycetota (previously known as the low G+C and high G+C Gram-positive bacteria, respectively) have the alternative Gram-positive arrangement. These differences in structure can produce differences in antibiotic susceptibility; for instance, vancomycin can kill only Gram-positive bacteria and is ineffective against Gram-negative pathogens, such as Haemophilus influenzae or Pseudomonas aeruginosa. Some bacteria have cell wall structures that are neither classically Gram-positive or Gram-negative. This includes clinically important bacteria such as mycobacteria which have a thick peptidoglycan cell wall like a Gram-positive bacterium, but also a second outer layer of lipids.
In many bacteria, an S-layer of rigidly arrayed protein molecules covers the outside of the cell. This layer provides chemical and physical protection for the cell surface and can act as a macromolecular diffusion barrier. S-layers have diverse functions and are known to act as virulence factors in Campylobacter species and contain surface enzymes in Bacillus stearothermophilus.
Helicobacter pylori electron micrograph, showing multiple flagella on the cell surface
Flagella are rigid protein structures, about 20 nanometres in diameter and up to 20 micrometres in length, that are used for motility. Flagella are driven by the energy released by the transfer of ions down an electrochemical gradient across the cell membrane.
Fimbriae (sometimes called "attachment pili") are fine filaments of protein, usually 2–10 nanometres in diameter and up to several micrometres in length. They are distributed over the surface of the cell, and resemble fine hairs when seen under the electron microscope. Fimbriae are believed to be involved in attachment to solid surfaces or to other cells, and are essential for the virulence of some bacterial pathogens. Pili (sing. pilus) are cellular appendages, slightly larger than fimbriae, that can transfer genetic material between bacterial cells in a process called conjugation where they are called conjugation pili or sex pili (see bacterial genetics, below). They can also generate movement where they are called type IV pili.
Glycocalyx is produced by many bacteria to surround their cells, and varies in structural complexity: ranging from a disorganised slime layer of extracellular polymeric substances to a highly structured capsule. These structures can protect cells from engulfment by eukaryotic cells such as macrophages (part of the human immune system). They can also act as antigens and be involved in cell recognition, as well as aiding attachment to surfaces and the formation of biofilms.
The assembly of these extracellular structures is dependent on bacterial secretion systems. These transfer proteins from the cytoplasm into the periplasm or into the environment around the cell. Many types of secretion systems are known and these structures are often essential for the virulence of pathogens, so are intensively studied.
Endospores
Further information: Endospore
Bacillus anthracis (stained purple) growing in cerebrospinal fluid
Some genera of Gram-positive bacteria, such as Bacillus, Clostridium, Sporohalobacter, Anaerobacter, and Heliobacterium, can form highly resistant, dormant structures called endospores. Endospores develop within the cytoplasm of the cell; generally, a single endospore develops in each cell. Each endospore contains a core of DNA and ribosomes surrounded by a cortex layer and protected by a multilayer rigid coat composed of peptidoglycan and a variety of proteins.
Endospores show no detectable metabolism and can survive extreme physical and chemical stresses, such as high levels of UV light, gamma radiation, detergents, disinfectants, heat, freezing, pressure, and desiccation. In this dormant state, these organisms may remain viable for millions of years. Endospores even allow bacteria to survive exposure to the vacuum and radiation of outer space, leading to the possibility that bacteria could be distributed throughout the Universe by space dust, meteoroids, asteroids, comets, planetoids, or directed panspermia.
Endospore-forming bacteria can cause disease; for example, anthrax can be contracted by the inhalation of Bacillus anthracis endospores, and contamination of deep puncture wounds with Clostridium tetani endospores causes tetanus, which, like botulism, is caused by a toxin released by the bacteria that grow from the spores. Clostridioides difficile infection, a common problem in healthcare settings, is caused by spore-forming bacteria.
Metabolism
Further information: Microbial metabolism
Bacteria exhibit an extremely wide variety of metabolic types. The distribution of metabolic traits within a group of bacteria has traditionally been used to define their taxonomy, but these traits often do not correspond with modern genetic classifications. Bacterial metabolism is classified into nutritional groups on the basis of three major criteria: the source of energy, the electron donors used, and the source of carbon used for growth.
Phototrophic bacteria derive energy from light using photosynthesis, while chemotrophic bacteria breaking down chemical compounds through oxidation, driving metabolism by transferring electrons from a given electron donor to a terminal electron acceptor in a redox reaction. Chemotrophs are further divided by the types of compounds they use to transfer electrons. Bacteria that derive electrons from inorganic compounds such as hydrogen, carbon monoxide, or ammonia are called lithotrophs, while those that use organic compounds are called organotrophs. Still, more specifically, aerobic organisms use oxygen as the terminal electron acceptor, while anaerobic organisms use other compounds such as nitrate, sulfate, or carbon dioxide.
Many bacteria, called heterotrophs, derive their carbon from other organic carbon. Others, such as cyanobacteria and some purple bacteria, are autotrophic, meaning they obtain cellular carbon by fixing carbon dioxide. In unusual circumstances, the gas methane can be used by methanotrophic bacteria as both a source of electrons and a substrate for carbon anabolism.
Nutritional types in bacterial metabolism
Nutritional type
Source of energy
Source of carbon
Examples
Phototrophs
Sunlight
Organic compounds (photoheterotrophs) or carbon fixation (photoautotrophs)
Cyanobacteria, Green sulfur bacteria, Chloroflexota, or Purple bacteria
Lithotrophs
Inorganic compounds
Organic compounds (lithoheterotrophs) or carbon fixation (lithoautotrophs)
Thermodesulfobacteriota, Hydrogenophilaceae, or Nitrospirota
Organotrophs
Organic compounds
Organic compounds (chemoheterotrophs) or carbon fixation (chemoautotrophs)
Bacillus, Clostridium, or Enterobacteriaceae
In many ways, bacterial metabolism provides traits that are useful for ecological stability and for human society. For example, diazotrophs have the ability to fix nitrogen gas using the enzyme nitrogenase. This trait, which can be found in bacteria of most metabolic types listed above, leads to the ecologically important processes of denitrification, sulfate reduction, and acetogenesis, respectively. Bacterial metabolic processes are important drivers in biological responses to pollution; for example, sulfate-reducing bacteria are largely responsible for the production of the highly toxic forms of mercury (methyl- and dimethylmercury) in the environment. Nonrespiratory anaerobes use fermentation to generate energy and reducing power, secreting metabolic by-products (such as ethanol in brewing) as waste. Facultative anaerobes can switch between fermentation and different terminal electron acceptors depending on the environmental conditions in which they find themselves.
Growth and reproduction
Further information: Bacterial growth
Many bacteria reproduce through binary fission, which is compared to mitosis and meiosis in this image.
A culture of Salmonella
A colony of Escherichia coli
Unlike in multicellular organisms, increases in cell size (cell growth) and reproduction by cell division are tightly linked in unicellular organisms. Bacteria grow to a fixed size and then reproduce through binary fission, a form of asexual reproduction. Under optimal conditions, bacteria can grow and divide extremely rapidly, and some bacterial populations can double as quickly as every 17 minutes. In cell division, two identical clone daughter cells are produced. Some bacteria, while still reproducing asexually, form more complex reproductive structures that help disperse the newly formed daughter cells. Examples include fruiting body formation by myxobacteria and aerial hyphae formation by Streptomyces species, or budding. Budding involves a cell forming a protrusion that breaks away and produces a daughter cell.
In the laboratory, bacteria are usually grown using solid or liquid media. Solid growth media, such as agar plates, are used to isolate pure cultures of a bacterial strain. However, liquid growth media are used when the measurement of growth or large volumes of cells are required. Growth in stirred liquid media occurs as an even cell suspension, making the cultures easy to divide and transfer, although isolating single bacteria from liquid media is difficult. The use of selective media (media with specific nutrients added or deficient, or with antibiotics added) can help identify specific organisms.
Most laboratory techniques for growing bacteria use high levels of nutrients to produce large amounts of cells cheaply and quickly. However, in natural environments, nutrients are limited, meaning that bacteria cannot continue to reproduce indefinitely. This nutrient limitation has led the evolution of different growth strategies (see r/K selection theory). Some organisms can grow extremely rapidly when nutrients become available, such as the formation of algal and cyanobacterial blooms that often occur in lakes during the summer. Other organisms have adaptations to harsh environments, such as the production of multiple antibiotics by Streptomyces that inhibit the growth of competing microorganisms. In nature, many organisms live in communities (e.g., biofilms) that may allow for increased supply of nutrients and protection from environmental stresses. These relationships can be essential for growth of a particular organism or group of organisms (syntrophy).
Bacterial growth follows four phases. When a population of bacteria first enter a high-nutrient environment that allows growth, the cells need to adapt to their new environment. The first phase of growth is the lag phase, a period of slow growth when the cells are adapting to the high-nutrient environment and preparing for fast growth. The lag phase has high biosynthesis rates, as proteins necessary for rapid growth are produced. The second phase of growth is the logarithmic phase, also known as the exponential phase. The log phase is marked by rapid exponential growth. The rate at which cells grow during this phase is known as the growth rate (k), and the time it takes the cells to double is known as the generation time (g). During log phase, nutrients are metabolised at maximum speed until one of the nutrients is depleted and starts limiting growth. The third phase of growth is the stationary phase and is caused by depleted nutrients. The cells reduce their metabolic activity and consume non-essential cellular proteins. The stationary phase is a transition from rapid growth to a stress response state and there is increased expression of genes involved in DNA repair, antioxidant metabolism and nutrient transport. The final phase is the death phase where the bacteria run out of nutrients and die.
Genetics
Main article: Bacterial genetics
Helium ion microscopy image showing T4 phage infecting E. coli. Some of the attached phage have contracted tails indicating that they have injected their DNA into the host. The bacterial cells are ~ 0.5 µm wide.
Most bacteria have a single circular chromosome that can range in size from only 160,000 base pairs in the endosymbiotic bacteria Carsonella ruddii, to 12,200,000 base pairs (12.2 Mbp) in the soil-dwelling bacteria Sorangium cellulosum. There are many exceptions to this; for example, some Streptomyces and Borrelia species contain a single linear chromosome, while some Vibrio species contain more than one chromosome. Some bacteria contain plasmids, small extra-chromosomal molecules of DNA that may contain genes for various useful functions such as antibiotic resistance, metabolic capabilities, or various virulence factors.
Bacteria genomes usually encode a few hundred to a few thousand genes. The genes in bacterial genomes are usually a single continuous stretch of DNA. Although several different types of introns do exist in bacteria, these are much rarer than in eukaryotes.
Bacteria, as asexual organisms, inherit an identical copy of the parent's genome and are clonal. However, all bacteria can evolve by selection on changes to their genetic material DNA caused by genetic recombination or mutations. Mutations arise from errors made during the replication of DNA or from exposure to mutagens. Mutation rates vary widely among different species of bacteria and even among different clones of a single species of bacteria. Genetic changes in bacterial genomes emerge from either random mutation during replication or "stress-directed mutation", where genes involved in a particular growth-limiting process have an increased mutation rate.
Some bacteria transfer genetic material between cells. This can occur in three main ways. First, bacteria can take up exogenous DNA from their environment in a process called transformation. Many bacteria can naturally take up DNA from the environment, while others must be chemically altered in order to induce them to take up DNA. The development of competence in nature is usually associated with stressful environmental conditions and seems to be an adaptation for facilitating repair of DNA damage in recipient cells. Second, bacteriophages can integrate into the bacterial chromosome, introducing foreign DNA in a process known as transduction. Many types of bacteriophage exist; some infect and lyse their host bacteria, while others insert into the bacterial chromosome. Bacteria resist phage infection through restriction modification systems that degrade foreign DNA, and a system that uses CRISPR sequences to retain fragments of the genomes of phage that the bacteria have come into contact with in the past, which allows them to block virus replication through a form of RNA interference. Third, bacteria can transfer genetic material through direct cell contact via conjugation.
In ordinary circumstances, transduction, conjugation, and transformation involve transfer of DNA between individual bacteria of the same species, but occasionally transfer may occur between individuals of different bacterial species, and this may have significant consequences, such as the transfer of antibiotic resistance. In such cases, gene acquisition from other bacteria or the environment is called horizontal gene transfer and may be common under natural conditions.
Behaviour
Movement
Main article: Bacterial motility
Transmission electron micrograph of Desulfovibrio vulgaris showing a single flagellum at one end of the cell. Scale bar is 0.5 micrometers long.
Many bacteria are motile (able to move themselves) and do so using a variety of mechanisms. The best studied of these are flagella, long filaments that are turned by a motor at the base to generate propeller-like movement. The bacterial flagellum is made of about 20 proteins, with approximately another 30 proteins required for its regulation and assembly. The flagellum is a rotating structure driven by a reversible motor at the base that uses the electrochemical gradient across the membrane for power.
The different arrangements of bacterial flagella: A-Monotrichous; B-Lophotrichous; C-Amphitrichous; D-Peritrichous
Bacteria can use flagella in different ways to generate different kinds of movement. Many bacteria (such as E. coli) have two distinct modes of movement: forward movement (swimming) and tumbling. The tumbling allows them to reorient and makes their movement a three-dimensional random walk. Bacterial species differ in the number and arrangement of flagella on their surface; some have a single flagellum (monotrichous), a flagellum at each end (amphitrichous), clusters of flagella at the poles of the cell (lophotrichous), while others have flagella distributed over the entire surface of the cell (peritrichous). The flagella of a unique group of bacteria, the spirochaetes, are found between two membranes in the periplasmic space. They have a distinctive helical body that twists about as it moves.
Two other types of bacterial motion are called twitching motility that relies on a structure called the type IV pilus, and gliding motility, that uses other mechanisms. In twitching motility, the rod-like pilus extends out from the cell, binds some substrate, and then retracts, pulling the cell forward.
Motile bacteria are attracted or repelled by certain stimuli in behaviours called taxes: these include chemotaxis, phototaxis, energy taxis, and magnetotaxis. In one peculiar group, the myxobacteria, individual bacteria move together to form waves of cells that then differentiate to form fruiting bodies containing spores. The myxobacteria move only when on solid surfaces, unlike E. coli, which is motile in liquid or solid media.
Several Listeria and Shigella species move inside host cells by usurping the cytoskeleton, which is normally used to move organelles inside the cell. By promoting actin polymerisation at one pole of their cells, they can form a kind of tail that pushes them through the host cell's cytoplasm.
Communication
See also: Prokaryote § Sociality
A few bacteria have chemical systems that generate light. This bioluminescence often occurs in bacteria that live in association with fish, and the light probably serves to attract fish or other large animals.
Bacteria often function as multicellular aggregates known as biofilms, exchanging a variety of molecular signals for intercell communication and engaging in coordinated multicellular behaviour.
The communal benefits of multicellular cooperation include a cellular division of labour, accessing resources that cannot effectively be used by single cells, collectively defending against antagonists, and optimising population survival by differentiating into distinct cell types. For example, bacteria in biofilms can have more than five hundred times increased resistance to antibacterial agents than individual "planktonic" bacteria of the same species.
One type of intercellular communication by a molecular signal is called quorum sensing, which serves the purpose of determining whether the local population density is sufficient to support investment in processes that are only successful if large numbers of similar organisms behave similarly, such as excreting digestive enzymes or emitting light. Quorum sensing enables bacteria to coordinate gene expression and to produce, release, and detect autoinducers or pheromones that accumulate with the growth in cell population.
Classification and identification
Main article: Bacterial taxonomy
Further information: Scientific classification, Systematics, Bacterial phyla, and Clinical pathology
Streptococcus mutans visualised with a Gram stain.
Phylogenetic tree showing the diversity of bacteria, compared to other organisms. Here bacteria are represented by three main supergroups: the CPR ultramicrobacterias, Terrabacteria and Gracilicutes according to recent genomic analyzes (2019).
Classification seeks to describe the diversity of bacterial species by naming and grouping organisms based on similarities. Bacteria can be classified on the basis of cell structure, cellular metabolism or on differences in cell components, such as DNA, fatty acids, pigments, antigens and quinones. While these schemes allowed the identification and classification of bacterial strains, it was unclear whether these differences represented variation between distinct species or between strains of the same species. This uncertainty was due to the lack of distinctive structures in most bacteria, as well as lateral gene transfer between unrelated species. Due to lateral gene transfer, some closely related bacteria can have very different morphologies and metabolisms. To overcome this uncertainty, modern bacterial classification emphasises molecular systematics, using genetic techniques such as guanine cytosine ratio determination, genome-genome hybridisation, as well as sequencing genes that have not undergone extensive lateral gene transfer, such as the rRNA gene. Classification of bacteria is determined by publication in the International Journal of Systematic Bacteriology, and Bergey's Manual of Systematic Bacteriology. The International Committee on Systematic Bacteriology (ICSB) maintains international rules for the naming of bacteria and taxonomic categories and for the ranking of them in the International Code of Nomenclature of Bacteria.
Historically, bacteria were considered a part of the Plantae, the Plant kingdom, and were called "Schizomycetes" (fission-fungi). For this reason, collective bacteria and other microorganisms in a host are often called "flora".
The term "bacteria" was traditionally applied to all microscopic, single-cell prokaryotes. However, molecular systematics showed prokaryotic life to consist of two separate domains, originally called Eubacteria and Archaebacteria, but now called Bacteria and Archaea that evolved independently from an ancient common ancestor. The archaea and eukaryotes are more closely related to each other than either is to the bacteria. These two domains, along with Eukarya, are the basis of the three-domain system, which is currently the most widely used classification system in microbiology. However, due to the relatively recent introduction of molecular systematics and a rapid increase in the number of genome sequences that are available, bacterial classification remains a changing and expanding field. For example, Cavalier-Smith argued that the Archaea and Eukaryotes evolved from Gram-positive bacteria.
The identification of bacteria in the laboratory is particularly relevant in medicine, where the correct treatment is determined by the bacterial species causing an infection. Consequently, the need to identify human pathogens was a major impetus for the development of techniques to identify bacteria.
The Gram stain, developed in 1884 by Hans Christian Gram, characterises bacteria based on the structural characteristics of their cell walls. The thick layers of peptidoglycan in the "Gram-positive" cell wall stain purple, while the thin "Gram-negative" cell wall appears pink. By combining morphology and Gram-staining, most bacteria can be classified as belonging to one of four groups (Gram-positive cocci, Gram-positive bacilli, Gram-negative cocci and Gram-negative bacilli). Some organisms are best identified by stains other than the Gram stain, particularly mycobacteria or Nocardia, which show acid fastness on Ziehl–Neelsen or similar stains. Other organisms may need to be identified by their growth in special media, or by other techniques, such as serology.
Culture techniques are designed to promote the growth and identify particular bacteria while restricting the growth of the other bacteria in the sample. Often these techniques are designed for specific specimens; for example, a sputum sample will be treated to identify organisms that cause pneumonia, while stool specimens are cultured on selective media to identify organisms that cause diarrhea while preventing growth of non-pathogenic bacteria. Specimens that are normally sterile, such as blood, urine or spinal fluid, are cultured under conditions designed to grow all possible organisms. Once a pathogenic organism has been isolated, it can be further characterised by its morphology, growth patterns (such as aerobic or anaerobic growth), patterns of hemolysis, and staining.
As with bacterial classification, identification of bacteria is increasingly using molecular methods, and mass spectroscopy. Most bacteria have not been characterised and there are many species that cannot be grown in the laboratory. Diagnostics using DNA-based tools, such as polymerase chain reaction, are increasingly popular due to their specificity and speed, compared to culture-based methods. These methods also allow the detection and identification of "viable but nonculturable" cells that are metabolically active but non-dividing. However, even using these improved methods, the total number of bacterial species is not known and cannot even be estimated with any certainty. Following present classification, there are a little less than 9,300 known species of prokaryotes, which includes bacteria and archaea; but attempts to estimate the true number of bacterial diversity have ranged from 10 to 10 total species—and even these diverse estimates may be off by many orders of magnitude.
Phyla
Main article: Bacterial phyla
The following phyla have been validly published according to the Bacteriological Code:
Acidobacteriota
Actinomycetota
Aquificota
Armatimonadota
Atribacterota
Bacillota
Bacteroidota
Balneolota
Bdellovibrionota
Caldisericota
Calditrichota
Campylobacterota
Chlamydiota
Chlorobiota
Chloroflexota
Chrysiogenota
Coprothermobacterota
Deferribacterota
Deinococcota
Dictyoglomota
Elusimicrobiota
Fibrobacterota
Fusobacteriota
Gemmatimonadota
Ignavibacteriota
Lentisphaerota
Mycoplasmatota
Myxococcota
Nitrospinota
Nitrospirota
Planctomycetota
Pseudomonadota
Rhodothermota
Spirochaetota
Synergistota
Thermodesulfobacteriota
Thermomicrobiota
Thermotogota
Verrucomicrobiota
Interactions with other organisms
Further information: Microbes in human culture
Overview of bacterial infections and main species involved.
Despite their apparent simplicity, bacteria can form complex associations with other organisms. These symbiotic associations can be divided into parasitism, mutualism and commensalism.
Commensals
The word "commensalism" is derived from the word "commensal", meaning "eating at the same table" and all plants and animals are colonised by commensal bacteria. In humans and other animals, millions of them live on the skin, the airways, the gut and other orifices.
Referred to as "normal flora", or "commensals", these bacteria usually cause no harm but may occasionally invade other sites of the body and cause infection. Escherichia coli is a commensal in the human gut but can cause urinary tract infections. Similarly, streptococci, which are part of the normal flora of the human mouth, can cause heart disease.
Predators
Some species of bacteria kill and then consume other microorganisms; these species are called predatory bacteria. These include organisms such as Myxococcus xanthus, which forms swarms of cells that kill and digest any bacteria they encounter. Other bacterial predators either attach to their prey in order to digest them and absorb nutrients or invade another cell and multiply inside the cytosol. These predatory bacteria are thought to have evolved from saprophages that consumed dead microorganisms, through adaptations that allowed them to entrap and kill other organisms.
Mutualists
Certain bacteria form close spatial associations that are essential for their survival. One such mutualistic association, called interspecies hydrogen transfer, occurs between clusters of anaerobic bacteria that consume organic acids, such as butyric acid or propionic acid, and produce hydrogen, and methanogenic archaea that consume hydrogen. The bacteria in this association are unable to consume the organic acids as this reaction produces hydrogen that accumulates in their surroundings. Only the intimate association with the hydrogen-consuming archaea keeps the hydrogen concentration low enough to allow the bacteria to grow.
In soil, microorganisms that reside in the rhizosphere (a zone that includes the root surface and the soil that adheres to the root after gentle shaking) carry out nitrogen fixation, converting nitrogen gas to nitrogenous compounds. This serves to provide an easily absorbable form of nitrogen for many plants, which cannot fix nitrogen themselves. Many other bacteria are found as symbionts in humans and other organisms. For example, the presence of over 1,000 bacterial species in the normal human gut flora of the intestines can contribute to gut immunity, synthesise vitamins, such as folic acid, vitamin K and biotin, convert sugars to lactic acid (see Lactobacillus), as well as fermenting complex undigestible carbohydrates. The presence of this gut flora also inhibits the growth of potentially pathogenic bacteria (usually through competitive exclusion) and these beneficial bacteria are consequently sold as probiotic dietary supplements.
Nearly all animal life is dependent on bacteria for survival as only bacteria and some archaea possess the genes and enzymes necessary to synthesize vitamin B12, also known as cobalamin, and provide it through the food chain. Vitamin B12 is a water-soluble vitamin that is involved in the metabolism of every cell of the human body. It is a cofactor in DNA synthesis and in both fatty acid and amino acid metabolism. It is particularly important in the normal functioning of the nervous system via its role in the synthesis of myelin.
Pathogens
Main article: Pathogenic bacteria
Neisseria gonorrhoeae and pus cells from a penile discharge (Gram stain)
Colour-enhanced scanning electron micrograph showing Salmonella typhimurium (red) invading cultured human cells
The body is continually exposed to many species of bacteria, including beneficial commensals, which grow on the skin and mucous membranes, and saprophytes, which grow mainly in the soil and in decaying matter. The blood and tissue fluids contain nutrients sufficient to sustain the growth of many bacteria. The body has defence mechanisms that enable it to resist microbial invasion of its tissues and give it a natural immunity or innate resistance against many microorganisms. Unlike some viruses, bacteria evolve relatively slowly so many bacterial diseases also occur in other animals.
If bacteria form a parasitic association with other organisms, they are classed as pathogens. Pathogenic bacteria are a major cause of human death and disease and cause infections such as tetanus (caused by Clostridium tetani), typhoid fever, diphtheria, syphilis, cholera, foodborne illness, leprosy (caused by Mycobacterium leprae) and tuberculosis (caused by Mycobacterium tuberculosis). A pathogenic cause for a known medical disease may only be discovered many years later, as was the case with Helicobacter pylori and peptic ulcer disease. Bacterial diseases are also important in agriculture, and bacteria cause leaf spot, fire blight and wilts in plants, as well as Johne's disease, mastitis, salmonella and anthrax in farm animals.
In bacterial vaginosis, beneficial bacteria in the vagina (top) are displaced by pathogens (bottom). Gram stain.
Each species of pathogen has a characteristic spectrum of interactions with its human hosts. Some organisms, such as Staphylococcus or Streptococcus, can cause skin infections, pneumonia, meningitis and sepsis, a systemic inflammatory response producing shock, massive vasodilation and death. Yet these organisms are also part of the normal human flora and usually exist on the skin or in the nose without causing any disease at all. Other organisms invariably cause disease in humans, such as Rickettsia, which are obligate intracellular parasites able to grow and reproduce only within the cells of other organisms. One species of Rickettsia causes typhus, while another causes Rocky Mountain spotted fever. Chlamydia, another phylum of obligate intracellular parasites, contains species that can cause pneumonia or urinary tract infection and may be involved in coronary heart disease. Some species, such as Pseudomonas aeruginosa, Burkholderia cenocepacia, and Mycobacterium avium, are opportunistic pathogens and cause disease mainly in people who are immunosuppressed or have cystic fibrosis. Some bacteria produce toxins, which cause diseases. These are endotoxins, which come from broken bacterial cells, and exotoxins, which are produced by bacteria and released into the environment. The bacterium Clostridium botulinum for example, produces a powerful exotoxin that cause respiratory paralysis, and Salmonellae produce an endotoxin that causes gastroenteritis. Some exotoxins can be converted to toxoids, which are used as vaccines to prevent the disease.
Bacterial infections may be treated with antibiotics, which are classified as bacteriocidal if they kill bacteria or bacteriostatic if they just prevent bacterial growth. There are many types of antibiotics, and each class inhibits a process that is different in the pathogen from that found in the host. An example of how antibiotics produce selective toxicity are chloramphenicol and puromycin, which inhibit the bacterial ribosome, but not the structurally different eukaryotic ribosome. Antibiotics are used both in treating human disease and in intensive farming to promote animal growth, where they may be contributing to the rapid development of antibiotic resistance in bacterial populations. Infections can be prevented by antiseptic measures such as sterilising the skin prior to piercing it with the needle of a syringe, and by proper care of indwelling catheters. Surgical and dental instruments are also sterilised to prevent contamination by bacteria. Disinfectants such as bleach are used to kill bacteria or other pathogens on surfaces to prevent contamination and further reduce the risk of infection.
Significance in technology and industry
Bacteria, often lactic acid bacteria, such as Lactobacillus species and Lactococcus species, in combination with yeasts and moulds, have been used for thousands of years in the preparation of fermented foods, such as cheese, pickles, soy sauce, sauerkraut, vinegar, wine, and yogurt.
The ability of bacteria to degrade a variety of organic compounds is remarkable and has been used in waste processing and bioremediation. Bacteria capable of digesting the hydrocarbons in petroleum are often used to clean up oil spills. Fertiliser was added to some of the beaches in Prince William Sound in an attempt to promote the growth of these naturally occurring bacteria after the 1989 Exxon Valdez oil spill. These efforts were effective on beaches that were not too thickly covered in oil. Bacteria are also used for the bioremediation of industrial toxic wastes. In the chemical industry, bacteria are most important in the production of enantiomerically pure chemicals for use as pharmaceuticals or agrichemicals.
Bacteria can also be used in place of pesticides in biological pest control. This commonly involves Bacillus thuringiensis (also called BT), a Gram-positive, soil-dwelling bacterium. Subspecies of this bacteria are used as Lepidopteran-specific insecticides under trade names such as Dipel and Thuricide. Because of their specificity, these pesticides are regarded as environmentally friendly, with little or no effect on humans, wildlife, pollinators, and most other beneficial insects.
Because of their ability to quickly grow and the relative ease with which they can be manipulated, bacteria are the workhorses for the fields of molecular biology, genetics, and biochemistry. By making mutations in bacterial DNA and examining the resulting phenotypes, scientists can determine the function of genes, enzymes, and metabolic pathways in bacteria, then apply this knowledge to more complex organisms. This aim of understanding the biochemistry of a cell reaches its most complex expression in the synthesis of huge amounts of enzyme kinetic and gene expression data into mathematical models of entire organisms. This is achievable in some well-studied bacteria, with models of Escherichia coli metabolism now being produced and tested. This understanding of bacterial metabolism and genetics allows the use of biotechnology to bioengineer bacteria for the production of therapeutic proteins, such as insulin, growth factors, or antibodies.
Because of their importance for research in general, samples of bacterial strains are isolated and preserved in Biological Resource Centers. This ensures the availability of the strain to scientists worldwide.
History of bacteriology
For the history of microbiology, see Microbiology. For the history of bacterial classification, see Bacterial taxonomy. For the natural history of Bacteria, see Last universal common ancestor.
Antonie van Leeuwenhoek, the first microbiologist and the first person to observe bacteria using a microscope.
Bacteria were first observed by the Dutch microscopist Antonie van Leeuwenhoek in 1676, using a single-lens microscope of his own design. He then published his observations in a series of letters to the Royal Society of London. Bacteria were Leeuwenhoek's most remarkable microscopic discovery. Their size was just at the limit of what his simple lenses could resolve, and, in one of the most striking hiatuses in the history of science, no one else would see them again for over a century. His observations also included protozoans which he called animalcules, and his findings were looked at again in the light of the more recent findings of cell theory.
Christian Gottfried Ehrenberg introduced the word "bacterium" in 1828. In fact, his Bacterium was a genus that contained non-spore-forming rod-shaped bacteria, as opposed to Bacillus, a genus of spore-forming rod-shaped bacteria defined by Ehrenberg in 1835.
Louis Pasteur demonstrated in 1859 that the growth of microorganisms causes the fermentation process and that this growth is not due to spontaneous generation (yeasts and molds, commonly associated with fermentation, are not bacteria, but rather fungi). Along with his contemporary Robert Koch, Pasteur was an early advocate of the germ theory of disease. Before them, Ignaz Semmelweis and Joseph Lister had realised the importance of sanitized hands in medical work. Semmelweis, who in the 1840s formulated his rules for handwashing in the hospital, prior to the advent of germ theory, attributed disease to "decomposing animal organic matter." His ideas were rejected and his book on the topic condemned by the medical community. After Lister, however, doctors started sanitizing their hands in the 1870s.
Robert Koch, a pioneer in medical microbiology, worked on cholera, anthrax and tuberculosis. In his research into tuberculosis, Koch finally proved the germ theory, for which he received a Nobel Prize in 1905. In Koch's postulates, he set out criteria to test if an organism is the cause of a disease, and these postulates are still used today.
Ferdinand Cohn is said to be a founder of bacteriology, studying bacteria from 1870. Cohn was the first to classify bacteria based on their morphology.
Though it was known in the nineteenth century that bacteria are the cause of many diseases, no effective antibacterial treatments were available. In 1910, Paul Ehrlich developed the first antibiotic, by changing dyes that selectively stained Treponema pallidum—the spirochaete that causes syphilis—into compounds that selectively killed the pathogen. Ehrlich, who had been awarded a 1908 Nobel Prize for his work on immunology, pioneered the use of stains to detect and identify bacteria, with his work being the basis of the Gram stain and the Ziehl–Neelsen stain.
A major step forward in the study of bacteria came in 1977 when Carl Woese recognised that archaea have a separate line of evolutionary descent from bacteria. This new phylogenetic taxonomy depended on the sequencing of 16S ribosomal RNA and divided prokaryotes into two evolutionary domains, as part of the three-domain system.
See also
Bacteriohopanepolyol
Genetically modified bacteria
Marine prokaryotes | biology | 164678 | https://no.wikipedia.org/wiki/Mutualisme%20%28biologi%29 | Mutualisme (biologi) | Mutualisme i biologi er en interaksjon mellom to arter der begge arter drar fordel av interaksjonen. Mutualisme en form for symbiose.
Velkjente eksempler på mutualisme er: lav, der en sopp og en alge eller blågrønnbakterie lever sammen og soppen sørger for bl.a. vann, næringssalter og beskyttelse og algen/blågrønnbakterien sørger for karbohydrater via fotosyntese. Pussefisk som leppefisker som spiser parasitter fra større fisk. Dyrepollinering der pollinatoren skaffer seg en ressurs, ofte mat, og planta oppnår befruktning. Nitrogenfikserende bakterier som finnes i røttene til flere plantegrupper og som tilfører planta nitrogenoksider samtidig som bakteriene er beskytta mot luftas oksygen.
Hvor tett koblingen er mellom de to organismene er varierer, i noen tilfelle er en eller begge totalt avhengig av mutualismen for å overleve eller reprodusere. Dette kalles obligat mutualisme. I andre tilfeller er avhengigheten ikke livsnødvendig, den er fakultativ. Eksempler på begge typer finnes inne pollinering. I de fleste tilfeller vil både planta og dyret kunne nyttiggjøre seg flere ulike arter og mutualismen mellom to bestemte arter er fakultativ. Imidlertid er det noen tilfeller der en planteart er helt avhengig av en insektart. Slik obligat mutalisme finnes mellom fiken og fikenveps og trolig mellom Madaskars stjerneorkide (Anaegraceum sesquipedale) og dens pollinator, Xanthopan morgani praedicta.
Økologi | norwegian_bokmål | 0.687766 |
bacterium_infect_another/Coliform_bacteria.txt | Coliform bacteria are defined as either motile or non-motile Gram-negative non-spore forming bacilli that possess β-galactosidase to produce acids and gases under their optimal growth temperature of 35–37 °C. They can be aerobes or facultative aerobes, and are a commonly used indicator of low sanitary quality of foods, milk, and water. Coliforms can be found in the aquatic environment, in soil and on vegetation; they are universally present in large numbers in the feces of warm-blooded animals as they are known to inhabit the gastrointestinal system. While coliform bacteria are not normally causes of serious illness, they are easy to culture, and their presence is used to infer that other pathogenic organisms of fecal origin may be present in a sample, or that said sample is not safe to consume. Such pathogens include disease-causing bacteria, viruses, or protozoa and many multicellular parasites.
Every drinking water source must be tested for the presence of these total coliform bacteria.
Genera[edit]
Typical genera include:
Citrobacter are peritrichous facultative anaerobic bacilli between 0.6–6 μm in length. Citrobacter species inhabit intestinal flora without causing harm, but can lead to urinary tract infections, bacteremia, brain abscesses, pneumonia, intra abdominal sepsis, meningitis, and joint infections if they are given the opportunity. Infections of a Citrobacter species has a mortality rate between 33–48%, with infants and immunocompromised individuals being more susceptible.
Enterobacter are motile, flagellated bacilli known for causing infections such as bacteremia, respiratory tract infections, urinary tract infections, infections of areas where surgery occurred, and in extreme cases meningitis, sinusitis and osteomyelitis. To determine the presence of Enterobacter in a sample, they are first grown on MacConkey agar to confirm they are lactose fermenting. An indole test will differentiate Enterobacter from Escherichia, as Enterobacter are indole negative and Escherichia is positive. Enterobacter are distinguished from Klebsiella because of their differences in motility.
Klebsiella are non-motile, Gram-negative bacilli ranging from 1–2 μm in length. They are facultative anaerobes with a capsule composed of complex acid polysaccharides that allows them to withstand drying for several months. Klebsiella pneumoniae is the most common Klebsiella species found in humans, the gastrointestinal tracts of animals, in sewage and in soil. On carbohydrate-rich media, Klebsiella colonies appear greyish-white in colour with a mucosal outer surface. The media used for selecting for Klebsiella species in a mixed sample is an agar including ornithine, raffinose, and Koser citrate, where members of this genus will form yellow, wet-looking colonies.
Escherichia species normally inhabit the human intestine and those of other warm-blooded animals, and are the most commonly responsible for causing disease in humans. E. coli specifically is the most common organism seen in the human intestine and are known to cause a variety of diseases in humans. Most E. coli strains are motile and have obtained many of their virulence features from horizontal gene transfer. There are several different pathotypes of E. coli causing gastrointestinal syndromes: diarrheagenic E. coli (DEC), enterotoxigenic E. coli (ETEC); EPEC; Shiga toxin–producingE. coli (STEC), which includes EHEC; enteroaggregative E. coli (EAEC); and enteroinvasive E. coli (EIEC). There are different ways to identify E. coli based on variation of their O, H and K polysaccharides on their cell surface or by using selective medias.
Escherichia coli (E. coli) can be distinguished from most other coliforms by its ability to ferment lactose at 44 °C in the fecal coliform test, and by its growth and color reaction on certain types of culture media. When cultured on an eosin methylene blue (EMB) plate, a positive result for E. coli is metallic green colonies on a dark purple medium. Also can be cultured on Tryptone Bile X-Glucuronide (TBX) to appear as blue or green colonies after incubation period of 24 hours. Escherichia coli have an incubation period of 12–72 hours with the optimal growth temperature being 37 °C. Unlike the general coliform group, E. coli are almost exclusively of fecal origin and their presence is thus an effective confirmation of fecal contamination. Most strains of E. coli are harmless, but some can cause serious illness in humans. Infection symptoms and signs include bloody diarrhea, stomach cramps, vomiting and occasionally, fever. The bacteria can also cause pneumonia, other respiratory illnesses and urinary tract infections.
An easy way to differentiate between different types of coliform bacteria is by using an eosin methylene blue agar plate. This plate is partially inhibitory to Gram (+) bacteria, and will produce a color change in the Gram (-) bacterial colonies based on lactose fermentation abilities. Strong lactose fermenters will appear as dark blue/purple/black, and E.coli (which also ferments lactose) colonies will be dark colored, but will also appear to have a metallic green sheen. Other coliform bacteria will appear as thick, slimy colonies, with non-fermenters being colorless, and weak fermenters being pink.
E. coli on EMB agar
Incidence of coliform outbreaks[edit]
Escherichia coli O157[edit]
As of November 15, 2021, seven states in the USA declared ten cases of illnesses from an E. coli O157:H7 strain. These cases were reported from October 15, 2021 through October 27, 2021 and an investigation was carried out by the Minnesota Department of Agriculture and FDA. It was concluded that packages of spinach collected from homes of infected people were contaminated with a strain of E. coli that matched the strain causing illness. This was determined by performing whole genome sequencing on the strain extracted from the spinach and comparing it to the strain taken from infected individuals.
As of February 7, 2022, the provinces of Alberta and Saskatchewan in Canada reported a collective fourteen confirmed cases of E. coli O157 strain illnesses. These were reported between December 2021 and January 2022, and the Public Health Agency of Canada (PHAC), the Canadian Food Inspection Agency (CFIA), and Health Canada were able to determine a specific brand of Original Kimchi to be the source of the organism. On January 28, 2022 and February 6, 2022, the CFIA issued a recall on Hankook Original Kimchi.
Detection of coliform bacteria in drinking water[edit]
PCR[edit]
Amplification of the beta-galactosidase gene is used to detect coliforms in general, because all coliform organisms produce this compound. The amplification of the beta-D glucuronidase is used to detect E. coli, or the amplification of their verotoxin gene(s) to detect verotoxin-producing E. coli.
Chemiluminescent in-situ hybridization[edit]
Specific areas of the 16S rRNA in the Enterobacteriaceae genus are bound by oligonucleotide probes, which aids in monitoring the quality of drinking water. Specifically, E. coli is labelled with a soybean peroxidase-labeled peptide nucleic acid (PNA) probes that bind to a specific sequence in their 16S rRNA. When used in conjunction with a chemiluminescent substrate, light is produced where each colony of E. coli is located, indicating that they are present in the sample.
Violet red bile agar[edit]
The solid medium is used to grow lactose-fermenting coliforms and utilizes a neutral red pH indicator. Pink colonies appear when lactose is fermented and are surrounded by bile that has precipitated out. To confirm if these colonies are coliforms, they are transferred to brilliant green lactose bile (BGLB) and incubated. If gas is visible after incubation, it can be confirmed that the sample had coliforms present.
Membrane filter method[edit]
Test samples are filtered through standard filter paper and then transferred to M-endo or LES Endo Agar mediums. Colonies appear pinkish-red with green metallic sheen after 22–24 hours of incubation. These colonies can be confirmed as coliforms if they are inoculated in lauryl tryptose (LST), produce gas, and then inoculated in BGLB. If there is gas production in the BGLB tubes, the test is positive for the presence of coliform bacteria.
See also[edit]
Medicine portal
Bacteriological water analysis
Coliform index
Fecal coliform
Indicator bacteria
Pathogenic Escherichia coli
| biology | 429630 | https://sv.wikipedia.org/wiki/Morula | Morula | En morula är en cellklump bestående av cirka 16 celler eller fler, som bildas till följd av snabb celldelning efter att ett ägg genomgått en lyckad befruktning av en spermie. Den är inte ihålig. När morulan uppnått ett par hundra celler utvecklas ett hålrum inuti klumpen och kallas därefter blastocyst. Morulan har samma storlek som den ursprungliga storleken hos zygoten, tills den fäster sig i livmodern (det vill säga om den gör det, den kan bland annat följa med ut ur livmodern under menstruationen, eller stötas bort från livmodern och dö) och får näring från moderkakan. Anledningen till att den inte blir större är att zona pellucida, membranet som skyddar mot att morulan skall fastna i äggledaren och bli ett utomkvedshavandeskap, inte utvidgas och släpper in vätska. Därför innehåller alla celler i morulan en del av den ursprungliga äggcellens cytoplasma.
Morula betyder litet mullbär ett namn den har fått av att den liknar ett mullbär.
Referenser
Noter
Cellbiologi
Embryologi | swedish | 0.896942 |
bacterium_infect_another/Gram-negative_bacteria.txt | Gram-negative bacteria are bacteria that do not retain the crystal violet stain used in the Gram staining method of bacterial differentiation. Their defining characteristic is their cell envelope, which consists of a thin peptidoglycan cell wall sandwiched between an inner (cytoplasmic) membrane and an outer membrane. These bacteria are found in all environments that support life on Earth.
Within this category, notable species include the model organism Escherichia coli, along with various pathogenic bacteria, such as Pseudomonas aeruginosa, Chlamydia trachomatis, and Yersinia pestis. They pose significant challenges in the medical field due to their outer membrane, which acts as a protective barrier against numerous antibiotics (including penicillin), detergents that would normally damage the inner cell membrane, and the antimicrobial enzyme lysozyme produced by animals as part of their innate immune system. Furthermore, the outer leaflet of this membrane contains a complex lipopolysaccharide (LPS) whose lipid A component can trigger a toxic reaction when the bacteria are lysed by immune cells. This reaction may lead to septic shock, resulting in low blood pressure, respiratory failure, reduced oxygen delivery, and lactic acidosis.
Several classes of antibiotics have been developed to target gram-negative bacteria, including aminopenicillins, ureidopenicillins, cephalosporins, beta-lactam-betalactamase inhibitor combinations (such as piperacillin-tazobactam), folate antagonists, quinolones, and carbapenems. Many of these antibiotics also cover gram-positive bacteria. The antibiotics that specifically target gram-negative organisms include aminoglycosides, monobactams (such as aztreonam), and ciprofloxacin.
Characteristics[edit]
Gram-negative cell wall structure
Gram-positive and -negative bacteria are differentiated chiefly by their cell wall structure
Conventional gram-negative (LPS-diderm) bacteria display these characteristics:
An inner cell membrane is present (cytoplasmic)
A thin peptidoglycan layer is present (this is much thicker in gram-positive bacteria)
Has outer membrane containing lipopolysaccharides (LPS, which consists of lipid A, core polysaccharide, and O antigen) in its outer leaflet and phospholipids in the inner leaflet
Porins exist in the outer membrane, which act like pores for particular molecules
Between the outer membrane and the cytoplasmic membrane there is a space filled with a concentrated gel-like substance called periplasm
The S-layer is directly attached to the outer membrane rather than to the peptidoglycan
If present, flagella have four supporting rings instead of two
Teichoic acids or lipoteichoic acids are absent
Lipoproteins are attached to the polysaccharide backbone
Some contain Braun's lipoprotein, which serves as a link between the outer membrane and the peptidoglycan chain by a covalent bond
Most, with few exceptions, do not form spores
Classification[edit]
Along with cell shape, Gram staining is a rapid diagnostic tool and once was used to group species at the subdivision of Bacteria.
Historically, the kingdom Monera was divided into four divisions based on Gram staining: Firmacutes (+), Gracillicutes (−), Mollicutes (0) and Mendocutes (var.).
Since 1987, the monophyly of the gram-negative bacteria has been disproven with molecular studies. However some authors, such as Cavalier-Smith still treat them as a monophyletic taxon (though not a clade; his definition of monophyly requires a single common ancestor but does not require holophyly, the property that all descendants be encompassed by the taxon) and refer to the group as a subkingdom "Negibacteria".
Taxonomy[edit]
Main article: Bacterial taxonomy
Further information: Bacteria § Classification and identification
This section may be too technical for most readers to understand. Please help improve it to make it understandable to non-experts, without removing the technical details. (March 2014) (Learn how and when to remove this template message)
It has been suggested that this section be split out into another article titled Gram stain. (Discuss) (November 2023)
Bacteria are traditionally classified based on their Gram-staining response into the gram-positive and gram-negative bacteria. Having just one membrane, the gram-positive bacteria are also known as monoderm bacteria, while gram-negative bacteria, having two membranes, are also known as diderm bacteria. It was traditionally thought that the groups represent lineages, i.e., the extra membrane only evolved once, such that gram-negative bacteria are more closely related to one another than to any gram-positive bacteria. While this is often true, the classification system breaks down in some cases, with lineage groupings not matching the staining result. Thus, Gram staining cannot be reliably used to assess familial relationships of bacteria. Nevertheless, staining often gives reliable information about the composition of the cell membrane, distinguishing between the presence or absence of an outer lipid membrane.
Of these two structurally distinct groups of prokaryotic organisms, monoderm prokaryotes are thought to be ancestral. Based upon a number of different observations, including that the gram-positive bacteria are the most sensitive to antibiotics and that the gram-negative bacteria are, in general, resistant to antibiotics, it has been proposed that the outer cell membrane in gram-negative bacteria (diderms) evolved as a protective mechanism against antibiotic selection pressure. Some bacteria such as Deinococcus, which stain gram-positive due to the presence of a thick peptidoglycan layer, but also possess an outer cell membrane are suggested as intermediates in the transition between monoderm (gram-positive) and diderm (gram-negative) bacteria. The diderm bacteria can also be further differentiated between simple diderms lacking lipopolysaccharide (LPS); the archetypical diderm bacteria, in which the outer cell membrane contains lipopolysaccharide; and the diderm bacteria, in which the outer cell membrane is made up of mycolic acid (e. g. Mycobacterium).
The conventional LPS-diderm group of gram-negative bacteria (e.g., Pseudomonadota, Aquificota, Chlamydiota, Bacteroidota, Chlorobiota, "Cyanobacteria", Fibrobacterota, Verrucomicrobiota, Planctomycetota, Spirochaetota, Acidobacteriota; "Hydrobacteria") are uniquely identified by a few conserved signature indel (CSI) in the HSP60 (GroEL) protein. In addition, a number of bacterial taxa (including Negativicutes, Fusobacteriota, Synergistota, and Elusimicrobiota) that are either part of the phylum Bacillota (a monoderm group) or branches in its proximity are also found to possess a diderm cell structure. They lack the GroEL signature. The presence of this CSI in all sequenced species of conventional lipopolysaccharide-containing gram-negative bacterial phyla provides evidence that these phyla of bacteria form a monophyletic clade and that no loss of the outer membrane from any species from this group has occurred.
Example species[edit]
The proteobacteria are a major superphylum of gram-negative bacteria, including E. coli, Salmonella, Shigella, and other Enterobacteriaceae, Pseudomonas, Moraxella, Helicobacter, Stenotrophomonas, Bdellovibrio, acetic acid bacteria, Legionella etc. Other notable groups of gram-negative bacteria include the cyanobacteria, spirochaetes, green sulfur, and green non-sulfur bacteria.
Medically-relevant gram-negative cocci include the four types that cause a sexually transmitted disease (Neisseria gonorrhoeae), a meningitis (Neisseria meningitidis), and respiratory symptoms (Moraxella catarrhalis, Haemophilus influenzae).
Medically relevant gram-negative bacilli include a multitude of species. Some of them cause primarily respiratory problems (Klebsiella pneumoniae, Legionella pneumophila, Pseudomonas aeruginosa), primarily urinary problems (Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Serratia marcescens), and primarily gastrointestinal problems (Helicobacter pylori, Salmonella enteritidis, Salmonella typhi).
Gram-negative bacteria associated with hospital-acquired infections include Acinetobacter baumannii, which cause bacteremia, secondary meningitis, and ventilator-associated pneumonia in hospital intensive-care units.
Bacterial transformation[edit]
Transformation is one of three processes for horizontal gene transfer, in which exogenous genetic material passes from one bacterium to another, the other two being conjugation (transfer of genetic material between two bacterial cells in direct contact) and transduction (injection of foreign DNA by a bacteriophage virus into the host bacterium). In transformation, the genetic material passes through the intervening medium, and uptake is completely dependent on the recipient bacterium.
As of 2014 about 80 species of bacteria were known to be capable of transformation, about evenly divided between gram-positive and gram-negative bacteria; the number might be an overestimate since several of the reports are supported by single papers. Transformation has been studied in medically important gram-negative bacteria species such as Helicobacter pylori, Legionella pneumophila, Neisseria meningitidis, Neisseria gonorrhoeae, Haemophilus influenzae and Vibrio cholerae. It has also been studied in gram-negative species found in soil such as Pseudomonas stutzeri, Acinetobacter baylyi, and gram-negative plant pathogens such as Ralstonia solanacearum and Xylella fastidiosa.
Role in disease[edit]
Example of a workup algorithm of possible bacterial infection in cases with no specifically requested targets (non-bacteria, mycobacteria etc.), with most common situations and agents seen in a New England setting. Clinically significant Gram-negative bacteria are usually rods, as shown near bottom right. Although some gram-negative bacteria can be recognized by "bench tests", diagnosis in the modern microbiology lab usually involves MALDI-TOF and/or multitarget assay.
One of the several unique characteristics of gram-negative bacteria is the structure of the bacterial outer membrane. The outer leaflet of this membrane contains lipopolysaccharide (LPS), whose lipid A portion acts as an endotoxin. If gram-negative bacteria enter the circulatory system, LPS can trigger an innate immune response, activating the immune system and producing cytokines (hormonal regulators). This leads to inflammation and can cause a toxic reaction, resulting in fever, an increased respiratory rate, and low blood pressure. That is why some infections with gram-negative bacteria can lead to life-threatening septic shock.
The outer membrane protects the bacteria from several antibiotics, dyes, and detergents that would normally damage either the inner membrane or the cell wall (made of peptidoglycan). The outer membrane provides these bacteria with resistance to lysozyme and penicillin. The periplasmic space (space between the two cell membranes) also contains enzymes which break down or modify antibiotics. Drugs commonly used to treat gram negative infections include amino, carboxy and ureido penicillins (ampicillin, amoxicillin, pipercillin, ticarcillin) these drugs may be combined with beta-lactamase inhibitors to combat the presence of enzymes that can digest these drugs (known as beta-lactamases) in the peri-plasmic space. Other classes of drugs that have gram negative spectrum include cephalosporins, monobactams (aztreonam), aminoglycosides, quinolones, macrolides, chloramphenicol, folate antagonists, and carbapenems.
Orthographic note[edit]
The adjectives gram-positive and gram-negative derive from the surname of Hans Christian Gram, a Danish bacteriologist; as eponymous adjectives, their initial letter can be either capital G or lower-case g, depending on which style guide (e.g., that of the CDC), if any, governs the document being written. This is further explained at Gram staining § Orthographic note.
See also[edit]
Autochaperone
Gram-variable and gram-indeterminate bacteria
OMPdb (2011)
Outer membrane receptor | biology | 25322 | https://no.wikipedia.org/wiki/Gramfarging | Gramfarging | Gramfarging er en mikrobiologisk fargeteknikk som ble utviklet av dansken Hans Christian Gram (1853 – 1938) i 1884, med senere modifikasjoner. Metoden brukes til å skille de to viktigste hovedgrupper av bakterier: gram-positive og gram-negative. Forskjellen skyldes en vesentlig forskjell i celleveggstrukturen. Gram-negative bakterier har en tynn cellevegg av peptidoglykan og en ytre lipopolysakkaridmembran, mens de gram-positive har en mer porøs og mye tykkere cellevegg av peptidoglykan og mangler den ytre membranen. Gram-positive bakterier farges lettere da alkoholen dehydrerer det tykke peptidoglykanlaget slik at krystallfiolett-jodid-komplekset ikke kan vaskes ut med alkohol. De gram-positive bakteriene er alle i forholdsvis nær slekt, mens de gram-negative er svært variert.
Gramfarging - trinn for trinn
Bakteriekulturen spres på et objektglass og tørkes inn.
Bakteriene fikseres med varme fra en flamme.
Fargestoffet krystallfiolett påføres.
Grams Jod-løsning tilsettes for å danne et vann-uløselig kompleks med krystallviolett.
Glasset vaskes med alkohol for å fjerne fargen fra de gram-negative.
Fargestoffet safranin påføres som motfarging.
Resultatet kan observeres i mikroskop.
Gram-negative bakterier fremstår da rosa eller røde på grunn av safranin, mens gram-positive er lilla på grunn av krystallfiolett-jodid-komplekset.
Eksterne lenker
Microbiologybites: Gramfarging. Infoside med video om gramfarging fra Microbiologybites.com
Mikrobiologi
Danske oppfinnelser | norwegian_bokmål | 0.546248 |
bacterium_infect_another/Infection.txt | An infection is the invasion of tissues by pathogens, their multiplication, and the reaction of host tissues to the infectious agent and the toxins they produce. An infectious disease, also known as a transmissible disease or communicable disease, is an illness resulting from an infection.
Infections can be caused by a wide range of pathogens, most prominently bacteria and viruses. Hosts can fight infections using their immune systems. Mammalian hosts react to infections with an innate response, often involving inflammation, followed by an adaptive response.
Specific medications used to treat infections include antibiotics, antivirals, antifungals, antiprotozoals, and antihelminthics. Infectious diseases resulted in 9.2 million deaths in 2013 (about 17% of all deaths). The branch of medicine that focuses on infections is referred to as infectious diseases.
Infections are caused by infectious agents (pathogens) including:
The signs and symptoms of an infection depend on the type of disease. Some signs of infection affect the whole body generally, such as fatigue, loss of appetite, weight loss, fevers, night sweats, chills, aches and pains. Others are specific to individual body parts, such as skin rashes, coughing, or a runny nose.
In certain cases, infectious diseases may be asymptomatic for much or even all of their course in a given host. In the latter case, the disease may only be defined as a "disease" (which by definition means an illness) in hosts who secondarily become ill after contact with an asymptomatic carrier. An infection is not synonymous with an infectious disease, as some infections do not cause illness in a host.
As bacterial and viral infections can both cause the same kinds of symptoms, it can be difficult to distinguish which is the cause of a specific infection. Distinguishing the two is important, since viral infections cannot be cured by antibiotics whereas bacterial infections can.
There is a general chain of events that applies to infections, sometimes called the chain of infection or transmission chain. The chain of events involves several steps – which include the infectious agent, reservoir, entering a susceptible host, exit and transmission to new hosts. Each of the links must be present in a chronological order for an infection to develop. Understanding these steps helps health care workers target the infection and prevent it from occurring in the first place.
Infection begins when an organism successfully enters the body, grows and multiplies. This is referred to as colonization. Most humans are not easily infected. Those with compromised or weakened immune systems have an increased susceptibility to chronic or persistent infections. Individuals who have a suppressed immune system are particularly susceptible to opportunistic infections. Entrance to the host at host–pathogen interface, generally occurs through the mucosa in orifices like the oral cavity, nose, eyes, genitalia, anus, or the microbe can enter through open wounds. While a few organisms can grow at the initial site of entry, many migrate and cause systemic infection in different organs. Some pathogens grow within the host cells (intracellular) whereas others grow freely in bodily fluids.
Wound colonization refers to non-replicating microorganisms within the wound, while in infected wounds, replicating organisms exist and tissue is injured. All multicellular organisms are colonized to some degree by extrinsic organisms, and the vast majority of these exist in either a mutualistic or commensal relationship with the host. An example of the former is the anaerobic bacteria species, which colonizes the mammalian colon, and an example of the latter are the various species of staphylococcus that exist on human skin. Neither of these colonizations are considered infections. The difference between an infection and a colonization is often only a matter of circumstance. Non-pathogenic organisms can become pathogenic given specific conditions, and even the most virulent organism requires certain circumstances to cause a compromising infection. Some colonizing bacteria, such as Corynebacteria sp. and Viridans streptococci, prevent the adhesion and colonization of pathogenic bacteria and thus have a symbiotic relationship with the host, preventing infection and speeding wound healing.
The variables involved in the outcome of a host becoming inoculated by a pathogen and the ultimate outcome include:
As an example, several staphylococcal species remain harmless on the skin, but, when present in a normally sterile space, such as in the capsule of a joint or the peritoneum, multiply without resistance and cause harm.
An interesting fact that gas chromatography–mass spectrometry, 16S ribosomal RNA analysis, omics, and other advanced technologies have made more apparent to humans in recent decades is that microbial colonization is very common even in environments that humans think of as being nearly sterile. Because it is normal to have bacterial colonization, it is difficult to know which chronic wounds can be classified as infected and how much risk of progression exists. Despite the huge number of wounds seen in clinical practice, there are limited quality data for evaluated symptoms and signs. A review of chronic wounds in the Journal of the American Medical Association's "Rational Clinical Examination Series" quantified the importance of increased pain as an indicator of infection. The review showed that the most useful finding is an increase in the level of pain [likelihood ratio (LR) range, 11–20] makes infection much more likely, but the absence of pain (negative likelihood ratio range, 0.64–0.88) does not rule out infection (summary LR 0.64–0.88).
Disease can arise if the host's protective immune mechanisms are compromised and the organism inflicts damage on the host. Microorganisms can cause tissue damage by releasing a variety of toxins or destructive enzymes. For example, Clostridium tetani releases a toxin that paralyzes muscles, and staphylococcus releases toxins that produce shock and sepsis. Not all infectious agents cause disease in all hosts. For example, less than 5% of individuals infected with polio develop disease. On the other hand, some infectious agents are highly virulent. The prion causing mad cow disease and Creutzfeldt–Jakob disease invariably kills all animals and people that are infected.
Persistent infections occur because the body is unable to clear the organism after the initial infection. Persistent infections are characterized by the continual presence of the infectious organism, often as latent infection with occasional recurrent relapses of active infection. There are some viruses that can maintain a persistent infection by infecting different cells of the body. Some viruses once acquired never leave the body. A typical example is the herpes virus, which tends to hide in nerves and become reactivated when specific circumstances arise.
Persistent infections cause millions of deaths globally each year. Chronic infections by parasites account for a high morbidity and mortality in many underdeveloped countries.
For infecting organisms to survive and repeat the infection cycle in other hosts, they (or their progeny) must leave an existing reservoir and cause infection elsewhere. Infection transmission can take place via many potential routes:
The relationship between virulence versus transmissibility is complex; with studies have shown that there were no clear relationship between the two. There is still a small number of evidence that partially suggests a link between virulence and transmissibility.
Diagnosis of infectious disease sometimes involves identifying an infectious agent either directly or indirectly. In practice most minor infectious diseases such as warts, cutaneous abscesses, respiratory system infections and diarrheal diseases are diagnosed by their clinical presentation and treated without knowledge of the specific causative agent. Conclusions about the cause of the disease are based upon the likelihood that a patient came in contact with a particular agent, the presence of a microbe in a community, and other epidemiological considerations. Given sufficient effort, all known infectious agents can be specifically identified.
Diagnosis of infectious disease is nearly always initiated by medical history and physical examination. More detailed identification techniques involve the culture of infectious agents isolated from a patient. Culture allows identification of infectious organisms by examining their microscopic features, by detecting the presence of substances produced by pathogens, and by directly identifying an organism by its genotype.
Many infectious organisms are identified without culture and microscopy. This is especially true for viruses, which cannot grow in culture. For some suspected pathogens, doctors may conduct tests that examine a patient's blood or other body fluids for antigens or antibodies that indicate presence of a specific pathogen that the doctor suspects.
Other techniques (such as X-rays, CAT scans, PET scans or NMR) are used to produce images of internal abnormalities resulting from the growth of an infectious agent. The images are useful in detection of, for example, a bone abscess or a spongiform encephalopathy produced by a prion.
The benefits of identification, however, are often greatly outweighed by the cost, as often there is no specific treatment, the cause is obvious, or the outcome of an infection is likely to be benign.
The diagnosis is aided by the presenting symptoms in any individual with an infectious disease, yet it usually needs additional diagnostic techniques to confirm the suspicion. Some signs are specifically characteristic and indicative of a disease and are called pathognomonic signs; but these are rare. Not all infections are symptomatic.
In children the presence of cyanosis, rapid breathing, poor peripheral perfusion, or a petechial rash increases the risk of a serious infection by greater than 5 fold. Other important indicators include parental concern, clinical instinct, and temperature greater than 40 °C.
Many diagnostic approaches depend on microbiological culture to isolate a pathogen from the appropriate clinical specimen. In a microbial culture, a growth medium is provided for a specific agent. A sample taken from potentially diseased tissue or fluid is then tested for the presence of an infectious agent able to grow within that medium. Many pathogenic bacteria are easily grown on nutrient agar, a form of solid medium that supplies carbohydrates and proteins necessary for growth, along with copious amounts of water. A single bacterium will grow into a visible mound on the surface of the plate called a colony, which may be separated from other colonies or melded together into a "lawn". The size, color, shape and form of a colony is characteristic of the bacterial species, its specific genetic makeup (its strain), and the environment that supports its growth. Other ingredients are often added to the plate to aid in identification. Plates may contain substances that permit the growth of some bacteria and not others, or that change color in response to certain bacteria and not others. Bacteriological plates such as these are commonly used in the clinical identification of infectious bacterium. Microbial culture may also be used in the identification of viruses: the medium, in this case, being cells grown in culture that the virus can infect, and then alter or kill. In the case of viral identification, a region of dead cells results from viral growth, and is called a "plaque". Eukaryotic parasites may also be grown in culture as a means of identifying a particular agent.
In the absence of suitable plate culture techniques, some microbes require culture within live animals. Bacteria such as Mycobacterium leprae and Treponema pallidum can be grown in animals, although serological and microscopic techniques make the use of live animals unnecessary. Viruses are also usually identified using alternatives to growth in culture or animals. Some viruses may be grown in embryonated eggs. Another useful identification method is Xenodiagnosis, or the use of a vector to support the growth of an infectious agent. Chagas disease is the most significant example, because it is difficult to directly demonstrate the presence of the causative agent, Trypanosoma cruzi in a patient, which therefore makes it difficult to definitively make a diagnosis. In this case, xenodiagnosis involves the use of the vector of the Chagas agent T. cruzi, an uninfected triatomine bug, which takes a blood meal from a person suspected of having been infected. The bug is later inspected for growth of T. cruzi within its gut.
Another principal tool in the diagnosis of infectious disease is microscopy. Virtually all of the culture techniques discussed above rely, at some point, on microscopic examination for definitive identification of the infectious agent. Microscopy may be carried out with simple instruments, such as the compound light microscope, or with instruments as complex as an electron microscope. Samples obtained from patients may be viewed directly under the light microscope, and can often rapidly lead to identification. Microscopy is often also used in conjunction with biochemical staining techniques, and can be made exquisitely specific when used in combination with antibody based techniques. For example, the use of antibodies made artificially fluorescent (fluorescently labeled antibodies) can be directed to bind to and identify a specific antigens present on a pathogen. A fluorescence microscope is then used to detect fluorescently labeled antibodies bound to internalized antigens within clinical samples or cultured cells. This technique is especially useful in the diagnosis of viral diseases, where the light microscope is incapable of identifying a virus directly.
Other microscopic procedures may also aid in identifying infectious agents. Almost all cells readily stain with a number of basic dyes due to the electrostatic attraction between negatively charged cellular molecules and the positive charge on the dye. A cell is normally transparent under a microscope, and using a stain increases the contrast of a cell with its background. Staining a cell with a dye such as Giemsa stain or crystal violet allows a microscopist to describe its size, shape, internal and external components and its associations with other cells. The response of bacteria to different staining procedures is used in the taxonomic classification of microbes as well. Two methods, the Gram stain and the acid-fast stain, are the standard approaches used to classify bacteria and to diagnosis of disease. The Gram stain identifies the bacterial groups Bacillota and Actinomycetota, both of which contain many significant human pathogens. The acid-fast staining procedure identifies the Actinomycetota genera Mycobacterium and Nocardia.
Biochemical tests used in the identification of infectious agents include the detection of metabolic or enzymatic products characteristic of a particular infectious agent. Since bacteria ferment carbohydrates in patterns characteristic of their genus and species, the detection of fermentation products is commonly used in bacterial identification. Acids, alcohols and gases are usually detected in these tests when bacteria are grown in selective liquid or solid media.
The isolation of enzymes from infected tissue can also provide the basis of a biochemical diagnosis of an infectious disease. For example, humans can make neither RNA replicases nor reverse transcriptase, and the presence of these enzymes are characteristic., of specific types of viral infections. The ability of the viral protein hemagglutinin to bind red blood cells together into a detectable matrix may also be characterized as a biochemical test for viral infection, although strictly speaking hemagglutinin is not an enzyme and has no metabolic function.
Serological methods are highly sensitive, specific and often extremely rapid tests used to identify microorganisms. These tests are based upon the ability of an antibody to bind specifically to an antigen. The antigen, usually a protein or carbohydrate made by an infectious agent, is bound by the antibody. This binding then sets off a chain of events that can be visibly obvious in various ways, dependent upon the test. For example, "Strep throat" is often diagnosed within minutes, and is based on the appearance of antigens made by the causative agent, S. pyogenes, that is retrieved from a patient's throat with a cotton swab. Serological tests, if available, are usually the preferred route of identification, however the tests are costly to develop and the reagents used in the test often require refrigeration. Some serological methods are extremely costly, although when commonly used, such as with the "strep test", they can be inexpensive.
Complex serological techniques have been developed into what are known as immunoassays. Immunoassays can use the basic antibody – antigen binding as the basis to produce an electro-magnetic or particle radiation signal, which can be detected by some form of instrumentation. Signal of unknowns can be compared to that of standards allowing quantitation of the target antigen. To aid in the diagnosis of infectious diseases, immunoassays can detect or measure antigens from either infectious agents or proteins generated by an infected organism in response to a foreign agent. For example, immunoassay A may detect the presence of a surface protein from a virus particle. Immunoassay B on the other hand may detect or measure antibodies produced by an organism's immune system that are made to neutralize and allow the destruction of the virus.
Instrumentation can be used to read extremely small signals created by secondary reactions linked to the antibody – antigen binding. Instrumentation can control sampling, reagent use, reaction times, signal detection, calculation of results, and data management to yield a cost-effective automated process for diagnosis of infectious disease.
Technologies based upon the polymerase chain reaction (PCR) method will become nearly ubiquitous gold standards of diagnostics of the near future, for several reasons. First, the catalog of infectious agents has grown to the point that virtually all of the significant infectious agents of the human population have been identified. Second, an infectious agent must grow within the human body to cause disease; essentially it must amplify its own nucleic acids in order to cause a disease. This amplification of nucleic acid in infected tissue offers an opportunity to detect the infectious agent by using PCR. Third, the essential tools for directing PCR, primers, are derived from the genomes of infectious agents, and with time those genomes will be known, if they are not already.
Thus, the technological ability to detect any infectious agent rapidly and specifically are currently available. The only remaining blockades to the use of PCR as a standard tool of diagnosis are in its cost and application, neither of which is insurmountable. The diagnosis of a few diseases will not benefit from the development of PCR methods, such as some of the clostridial diseases (tetanus and botulism). These diseases are fundamentally biological poisonings by relatively small numbers of infectious bacteria that produce extremely potent neurotoxins. A significant proliferation of the infectious agent does not occur, this limits the ability of PCR to detect the presence of any bacteria.
Given the wide range of bacterial, viral, fungal, protozoal, and helminthic pathogens that cause debilitating and life-threatening illnesses, the ability to quickly identify the cause of infection is important yet often challenging. For example, more than half of cases of encephalitis, a severe illness affecting the brain, remain undiagnosed, despite extensive testing using the standard of care (microbiological culture) and state-of-the-art clinical laboratory methods. Metagenomic sequencing-based diagnostic tests are currently being developed for clinical use and show promise as a sensitive, specific, and rapid way to diagnose infection using a single all-encompassing test. This test is similar to current PCR tests; however, an untargeted whole genome amplification is used rather than primers for a specific infectious agent. This amplification step is followed by next-generation sequencing or third-generation sequencing, alignment comparisons, and taxonomic classification using large databases of thousands of pathogen and commensal reference genomes. Simultaneously, antimicrobial resistance genes within pathogen and plasmid genomes are sequenced and aligned to the taxonomically classified pathogen genomes to generate an antimicrobial resistance profile – analogous to antibiotic sensitivity testing – to facilitate antimicrobial stewardship and allow for the optimization of treatment using the most effective drugs for a patient's infection.
Metagenomic sequencing could prove especially useful for diagnosis when the patient is immunocompromised. An ever-wider array of infectious agents can cause serious harm to individuals with immunosuppression, so clinical screening must often be broader. Additionally, the expression of symptoms is often atypical, making a clinical diagnosis based on presentation more difficult. Thirdly, diagnostic methods that rely on the detection of antibodies are more likely to fail. A rapid, sensitive, specific, and untargeted test for all known human pathogens that detects the presence of the organism's DNA rather than antibodies is therefore highly desirable.
There is usually an indication for a specific identification of an infectious agent only when such identification can aid in the treatment or prevention of the disease, or to advance knowledge of the course of an illness prior to the development of effective therapeutic or preventative measures. For example, in the early 1980s, prior to the appearance of AZT for the treatment of AIDS, the course of the disease was closely followed by monitoring the composition of patient blood samples, even though the outcome would not offer the patient any further treatment options. In part, these studies on the appearance of HIV in specific communities permitted the advancement of hypotheses as to the route of transmission of the virus. By understanding how the disease was transmitted, resources could be targeted to the communities at greatest risk in campaigns aimed at reducing the number of new infections. The specific serological diagnostic identification, and later genotypic or molecular identification, of HIV also enabled the development of hypotheses as to the temporal and geographical origins of the virus, as well as a myriad of other hypothesis. The development of molecular diagnostic tools have enabled physicians and researchers to monitor the efficacy of treatment with anti-retroviral drugs. Molecular diagnostics are now commonly used to identify HIV in healthy people long before the onset of illness and have been used to demonstrate the existence of people who are genetically resistant to HIV infection. Thus, while there still is no cure for AIDS, there is great therapeutic and predictive benefit to identifying the virus and monitoring the virus levels within the blood of infected individuals, both for the patient and for the community at large.
Symptomatic infections are apparent and clinical, whereas an infection that is active but does not produce noticeable symptoms may be called inapparent, silent, subclinical, or occult. An infection that is inactive or dormant is called a latent infection. An example of a latent bacterial infection is latent tuberculosis. Some viral infections can also be latent, examples of latent viral infections are any of those from the Herpesviridae family.
The word infection can denote any presence of a particular pathogen at all (no matter how little) but also is often used in a sense implying a clinically apparent infection (in other words, a case of infectious disease). This fact occasionally creates some ambiguity or prompts some usage discussion; to get around this it is common for health professionals to speak of colonization (rather than infection) when they mean that some of the pathogens are present but that no clinically apparent infection (no disease) is present.
Different terms are used to describe how and where infections present over time. In an acute infection, symptoms develop rapidly; its course can either be rapid or protracted. In chronic infection, symptoms usually develop gradually over weeks or months and are slow to resolve. In subacute infections, symptoms take longer to develop than in acute infections but arise more quickly than those of chronic infections. A focal infection is an initial site of infection from which organisms travel via the bloodstream to another area of the body.
Among the many varieties of microorganisms, relatively few cause disease in otherwise healthy individuals. Infectious disease results from the interplay between those few pathogens and the defenses of the hosts they infect. The appearance and severity of disease resulting from any pathogen depend upon the ability of that pathogen to damage the host as well as the ability of the host to resist the pathogen. However, a host's immune system can also cause damage to the host itself in an attempt to control the infection. Clinicians, therefore, classify infectious microorganisms or microbes according to the status of host defenses – either as primary pathogens or as opportunistic pathogens.
Primary pathogens cause disease as a result of their presence or activity within the normal, healthy host, and their intrinsic virulence (the severity of the disease they cause) is, in part, a necessary consequence of their need to reproduce and spread. Many of the most common primary pathogens of humans only infect humans, however, many serious diseases are caused by organisms acquired from the environment or that infect non-human hosts.
Opportunistic pathogens can cause an infectious disease in a host with depressed resistance (immunodeficiency) or if they have unusual access to the inside of the body (for example, via trauma). Opportunistic infection may be caused by microbes ordinarily in contact with the host, such as pathogenic bacteria or fungi in the gastrointestinal or the upper respiratory tract, and they may also result from (otherwise innocuous) microbes acquired from other hosts (as in Clostridium difficile colitis) or from the environment as a result of traumatic introduction (as in surgical wound infections or compound fractures). An opportunistic disease requires impairment of host defenses, which may occur as a result of genetic defects (such as chronic granulomatous disease), exposure to antimicrobial drugs or immunosuppressive chemicals (as might occur following poisoning or cancer chemotherapy), exposure to ionizing radiation, or as a result of an infectious disease with immunosuppressive activity (such as with measles, malaria or HIV disease). Primary pathogens may also cause more severe disease in a host with depressed resistance than would normally occur in an immunosufficient host.
While a primary infection can practically be viewed as the root cause of an individual's current health problem, a secondary infection is a sequela or complication of that root cause. For example, an infection due to a burn or penetrating trauma (the root cause) is a secondary infection. Primary pathogens often cause primary infection and often cause secondary infection. Usually, opportunistic infections are viewed as secondary infections (because immunodeficiency or injury was the predisposing factor).
Other types of infection consist of mixed, iatrogenic, nosocomial, and community-acquired infection. A mixed infection is an infection that is caused by two or more pathogens. An example of this is appendicitis, which is caused by Bacteroides fragilis and Escherichia coli. The second is an iatrogenic infection. This type of infection is one that is transmitted from a health care worker to a patient. A nosocomial infection is also one that occurs in a health care setting. Nosocomial infections are those that are acquired during a hospital stay. Lastly, a community-acquired infection is one in which the infection is acquired from a whole community.
One manner of proving that a given disease is infectious, is to satisfy Koch's postulates (first proposed by Robert Koch), which require that first, the infectious agent be identifiable only in patients who have the disease, and not in healthy controls, and second, that patients who contract the infectious agent also develop the disease. These postulates were first used in the discovery that Mycobacteria species cause tuberculosis.
However, Koch's postulates cannot usually be tested in modern practice for ethical reasons. Proving them would require experimental infection of a healthy individual with a pathogen produced as a pure culture. Conversely, even clearly infectious diseases do not always meet the infectious criteria; for example, Treponema pallidum, the causative spirochete of syphilis, cannot be cultured in vitro – however the organism can be cultured in rabbit testes. It is less clear that a pure culture comes from an animal source serving as host than it is when derived from microbes derived from plate culture.
Epidemiology, or the study and analysis of who, why and where disease occurs, and what determines whether various populations have a disease, is another important tool used to understand infectious disease. Epidemiologists may determine differences among groups within a population, such as whether certain age groups have a greater or lesser rate of infection; whether groups living in different neighborhoods are more likely to be infected; and by other factors, such as gender and race. Researchers also may assess whether a disease outbreak is sporadic, or just an occasional occurrence; endemic, with a steady level of regular cases occurring in a region; epidemic, with a fast arising, and unusually high number of cases in a region; or pandemic, which is a global epidemic. If the cause of the infectious disease is unknown, epidemiology can be used to assist with tracking down the sources of infection.
Infectious diseases are sometimes called contagious diseases when they are easily transmitted by contact with an ill person or their secretions (e.g., influenza). Thus, a contagious disease is a subset of infectious disease that is especially infective or easily transmitted. Other types of infectious, transmissible, or communicable diseases with more specialized routes of infection, such as vector transmission or sexual transmission, are usually not regarded as "contagious", and often do not require medical isolation (sometimes loosely called quarantine) of those affected. However, this specialized connotation of the word "contagious" and "contagious disease" (easy transmissibility) is not always respected in popular use.
Infectious diseases are commonly transmitted from person to person through direct contact. The types of contact are through person to person and droplet spread. Indirect contact such as airborne transmission, contaminated objects, food and drinking water, animal person contact, animal reservoirs, insect bites, and environmental reservoirs are another way infectious diseases are transmitted.
Infections can be classified by the anatomic location or organ system infected, including:
In addition, locations of inflammation where infection is the most common cause include pneumonia, meningitis and salpingitis.
Techniques like hand washing, wearing gowns, and wearing face masks can help prevent infections from being passed from one person to another. Aseptic technique was introduced in medicine and surgery in the late 19th century and greatly reduced the incidence of infections caused by surgery. Frequent hand washing remains the most important defense against the spread of unwanted organisms. There are other forms of prevention such as avoiding the use of illicit drugs, using a condom, wearing gloves, and having a healthy lifestyle with a balanced diet and regular exercise. Cooking foods well and avoiding foods that have been left outside for a long time is also important.
Antimicrobial substances used to prevent transmission of infections include:
One of the ways to prevent or slow down the transmission of infectious diseases is to recognize the different characteristics of various diseases. Some critical disease characteristics that should be evaluated include virulence, distance traveled by those affected, and level of contagiousness. The human strains of Ebola virus, for example, incapacitate those infected extremely quickly and kill them soon after. As a result, those affected by this disease do not have the opportunity to travel very far from the initial infection zone. Also, this virus must spread through skin lesions or permeable membranes such as the eye. Thus, the initial stage of Ebola is not very contagious since its victims experience only internal hemorrhaging. As a result of the above features, the spread of Ebola is very rapid and usually stays within a relatively confined geographical area. In contrast, the human immunodeficiency virus (HIV) kills its victims very slowly by attacking their immune system. As a result, many of its victims transmit the virus to other individuals before even realizing that they are carrying the disease. Also, the relatively low virulence allows its victims to travel long distances, increasing the likelihood of an epidemic.
Another effective way to decrease the transmission rate of infectious diseases is to recognize the effects of small-world networks. In epidemics, there are often extensive interactions within hubs or groups of infected individuals and other interactions within discrete hubs of susceptible individuals. Despite the low interaction between discrete hubs, the disease can jump and spread in a susceptible hub via a single or few interactions with an infected hub. Thus, infection rates in small-world networks can be reduced somewhat if interactions between individuals within infected hubs are eliminated (Figure 1). However, infection rates can be drastically reduced if the main focus is on the prevention of transmission jumps between hubs. The use of needle exchange programs in areas with a high density of drug users with HIV is an example of the successful implementation of this treatment method. Another example is the use of ring culling or vaccination of potentially susceptible livestock in adjacent farms to prevent the spread of the foot-and-mouth virus in 2001.
A general method to prevent transmission of vector-borne pathogens is pest control.
In cases where infection is merely suspected, individuals may be quarantined until the incubation period has passed and the disease manifests itself or the person remains healthy. Groups may undergo quarantine, or in the case of communities, a cordon sanitaire may be imposed to prevent infection from spreading beyond the community, or in the case of protective sequestration, into a community. Public health authorities may implement other forms of social distancing, such as school closings, lockdowns or temporary restrictions (e.g. circuit breakers) to control an epidemic.
Infection with most pathogens does not result in death of the host and the offending organism is ultimately cleared after the symptoms of the disease have waned. This process requires immune mechanisms to kill or inactivate the inoculum of the pathogen. Specific acquired immunity against infectious diseases may be mediated by antibodies and/or T lymphocytes. Immunity mediated by these two factors may be manifested by:
The immune system response to a microorganism often causes symptoms such as a high fever and inflammation, and has the potential to be more devastating than direct damage caused by a microbe.
Resistance to infection (immunity) may be acquired following a disease, by asymptomatic carriage of the pathogen, by harboring an organism with a similar structure (crossreacting), or by vaccination. Knowledge of the protective antigens and specific acquired host immune factors is more complete for primary pathogens than for opportunistic pathogens. There is also the phenomenon of herd immunity which offers a measure of protection to those otherwise vulnerable people when a large enough proportion of the population has acquired immunity from certain infections.
Immune resistance to an infectious disease requires a critical level of either antigen-specific antibodies and/or T cells when the host encounters the pathogen. Some individuals develop natural serum antibodies to the surface polysaccharides of some agents although they have had little or no contact with the agent, these natural antibodies confer specific protection to adults and are passively transmitted to newborns.
The organism that is the target of an infecting action of a specific infectious agent is called the host. The host harbouring an agent that is in a mature or sexually active stage phase is called the definitive host. The intermediate host comes in contact during the larvae stage. A host can be anything living and can attain to asexual and sexual reproduction.
The clearance of the pathogens, either treatment-induced or spontaneous, it can be influenced by the genetic variants carried by the individual patients. For instance, for genotype 1 hepatitis C treated with Pegylated interferon-alpha-2a or Pegylated interferon-alpha-2b (brand names Pegasys or PEG-Intron) combined with ribavirin, it has been shown that genetic polymorphisms near the human IL28B gene, encoding interferon lambda 3, are associated with significant differences in the treatment-induced clearance of the virus. This finding, originally reported in Nature, showed that genotype 1 hepatitis C patients carrying certain genetic variant alleles near the IL28B gene are more possibly to achieve sustained virological response after the treatment than others. Later report from Nature demonstrated that the same genetic variants are also associated with the natural clearance of the genotype 1 hepatitis C virus.
When infection attacks the body, anti-infective drugs can suppress the infection. Several broad types of anti-infective drugs exist, depending on the type of organism targeted; they include antibacterial (antibiotic; including antitubercular), antiviral, antifungal and antiparasitic (including antiprotozoal and antihelminthic) agents. Depending on the severity and the type of infection, the antibiotic may be given by mouth or by injection, or may be applied topically. Severe infections of the brain are usually treated with intravenous antibiotics. Sometimes, multiple antibiotics are used in case there is resistance to one antibiotic. Antibiotics only work for bacteria and do not affect viruses. Antibiotics work by slowing down the multiplication of bacteria or killing the bacteria. The most common classes of antibiotics used in medicine include penicillin, cephalosporins, aminoglycosides, macrolides, quinolones and tetracyclines.
Not all infections require treatment, and for many self-limiting infections the treatment may cause more side-effects than benefits. Antimicrobial stewardship is the concept that healthcare providers should treat an infection with an antimicrobial that specifically works well for the target pathogen for the shortest amount of time and to only treat when there is a known or highly suspected pathogen that will respond to the medication.
Pandemics such as COVID-19 show that people dramatically differ in their susceptibility to infection. This may be because of general health, age, or their immune status, e.g. when they have been infected previously. However, it also has become clear that there are genetic factor which determine susceptibility to infection. For instance, up to 40% of SARS-CoV-2 infections may be asymptomatic, suggesting that many people are naturally protected from disease. Large genetic studies have defined risk factors for severe SARS-CoV-2 infections, and genome sequences from 659 patients with severe COVID-19 revealed genetic variants that appear to be associated with life-threatening disease. One gene identified in these studies is type I interferon (IFN). Autoantibodies against type I IFNs were found in up to 13.7% of patients with life-threatening COVID-19, indicating that a complex interaction between genetics and the immune system is important for natural resistance to Covid.
Similarly, mutations in the ERAP2 gene, encoding endoplasmic reticulum aminopeptidase 2, seem to increase the susceptibility to the plague, the disease caused by an infection with the bacteria Yersinia pestis. People who inherited two copies of a complete variant of the gene were twice as likely to have survived the plague as those who inherited two copies of a truncated variant.
Susceptibility also determined the epidemiology of infection, given that different populations have different genetic and environmental conditions that affect infections.
In 2010, about 10 million people died of infectious diseases.
The World Health Organization collects information on global deaths by International Classification of Disease (ICD) code categories. The following table lists the top infectious disease by number of deaths in 2002. 1993 data is included for comparison.
The top three single agent/disease killers are HIV/AIDS, TB and malaria. While the number of deaths due to nearly every disease have decreased, deaths due to HIV/AIDS have increased fourfold. Childhood diseases include pertussis, poliomyelitis, diphtheria, measles and tetanus. Children also make up a large percentage of lower respiratory and diarrheal deaths. In 2012, approximately 3.1 million people have died due to lower respiratory infections, making it the number 4 leading cause of death in the world.
With their potential for unpredictable and explosive impacts, infectious diseases have been major actors in human history. A pandemic (or global epidemic) is a disease that affects people over an extensive geographical area. For example:
In most cases, microorganisms live in harmony with their hosts via mutual or commensal interactions. Diseases can emerge when existing parasites become pathogenic or when new pathogenic parasites enter a new host.
Several human activities have led to the emergence of zoonotic human pathogens, including viruses, bacteria, protozoa, and rickettsia, and spread of vector-borne diseases, see also globalization and disease and wildlife disease:
In Antiquity, the Greek historian Thucydides (c. 460 – c. 400 BCE) was the first person to write, in his account of the plague of Athens, that diseases could spread from an infected person to others. In his On the Different Types of Fever (c. 175 AD), the Greco-Roman physician Galen speculated that plagues were spread by "certain seeds of plague", which were present in the air. In the Sushruta Samhita, the ancient Indian physician Sushruta theorized: "Leprosy, fever, consumption, diseases of the eye, and other infectious diseases spread from one person to another by sexual union, physical contact, eating together, sleeping together, sitting together, and the use of same clothes, garlands and pastes." This book has been dated to about the sixth century BC.
A basic form of contagion theory was proposed by Persian physician Ibn Sina (known as Avicenna in Europe) in The Canon of Medicine (1025), which later became the most authoritative medical textbook in Europe up until the 16th century. In Book IV of the Canon, Ibn Sina discussed epidemics, outlining the classical miasma theory and attempting to blend it with his own early contagion theory. He mentioned that people can transmit disease to others by breath, noted contagion with tuberculosis, and discussed the transmission of disease through water and dirt. The concept of invisible contagion was later discussed by several Islamic scholars in the Ayyubid Sultanate who referred to them as najasat ("impure substances"). The fiqh scholar Ibn al-Haj al-Abdari (c. 1250–1336), while discussing Islamic diet and hygiene, gave warnings about how contagion can contaminate water, food, and garments, and could spread through the water supply, and may have implied contagion to be unseen particles.
When the Black Death bubonic plague reached Al-Andalus in the 14th century, the Arab physicians Ibn Khatima (c. 1369) and Ibn al-Khatib (1313–1374) hypothesised that infectious diseases were caused by "minute bodies" and described how they can be transmitted through garments, vessels and earrings. Ideas of contagion became more popular in Europe during the Renaissance, particularly through the writing of the Italian physician Girolamo Fracastoro. Anton van Leeuwenhoek (1632–1723) advanced the science of microscopy by being the first to observe microorganisms, allowing for easy visualization of bacteria.
In the mid-19th century John Snow and William Budd did important work demonstrating the contagiousness of typhoid and cholera through contaminated water. Both are credited with decreasing epidemics of cholera in their towns by implementing measures to prevent contamination of water. Louis Pasteur proved beyond doubt that certain diseases are caused by infectious agents, and developed a vaccine for rabies. Robert Koch provided the study of infectious diseases with a scientific basis known as Koch's postulates. Edward Jenner, Jonas Salk and Albert Sabin developed effective vaccines for smallpox and polio, which would later result in the eradication and near-eradication of these diseases, respectively. Alexander Fleming discovered the world's first antibiotic, penicillin, which Florey and Chain then developed. Gerhard Domagk developed sulphonamides, the first broad spectrum synthetic antibacterial drugs.
The medical treatment of infectious diseases falls into the medical field of Infectious Disease and in some cases the study of propagation pertains to the field of Epidemiology. Generally, infections are initially diagnosed by primary care physicians or internal medicine specialists. For example, an "uncomplicated" pneumonia will generally be treated by the internist or the pulmonologist (lung physician). The work of the infectious diseases specialist therefore entails working with both patients and general practitioners, as well as laboratory scientists, immunologists, bacteriologists and other specialists.
An infectious disease team may be alerted when:
Several studies have reported associations between pathogen load in an area and human behavior. Higher pathogen load is associated with decreased size of ethnic and religious groups in an area. This may be due high pathogen load favoring avoidance of other groups, which may reduce pathogen transmission, or a high pathogen load preventing the creation of large settlements and armies that enforce a common culture. Higher pathogen load is also associated with more restricted sexual behavior, which may reduce pathogen transmission. It also associated with higher preferences for health and attractiveness in mates. Higher fertility rates and shorter or less parental care per child is another association that may be a compensation for the higher mortality rate. There is also an association with polygyny which may be due to higher pathogen load, making selecting males with a high genetic resistance increasingly important. Higher pathogen load is also associated with more collectivism and less individualism, which may limit contacts with outside groups and infections. There are alternative explanations for at least some of the associations although some of these explanations may in turn ultimately be due to pathogen load. Thus, polygyny may also be due to a lower male: female ratio in these areas but this may ultimately be due to male infants having increased mortality from infectious diseases. Another example is that poor socioeconomic factors may ultimately in part be due to high pathogen load preventing economic development.
Evidence of infection in fossil remains is a subject of interest for paleopathologists, scientists who study occurrences of injuries and illness in extinct life forms. Signs of infection have been discovered in the bones of carnivorous dinosaurs. When present, however, these infections seem to tend to be confined to only small regions of the body. A skull attributed to the early carnivorous dinosaur Herrerasaurus ischigualastensis exhibits pit-like wounds surrounded by swollen and porous bone. The unusual texture of the bone around the wounds suggests they were affected by a short-lived, non-lethal infection. Scientists who studied the skull speculated that the bite marks were received in a fight with another Herrerasaurus. Other carnivorous dinosaurs with documented evidence of infection include Acrocanthosaurus, Allosaurus, Tyrannosaurus and a tyrannosaur from the Kirtland Formation. The infections from both tyrannosaurs were received by being bitten during a fight, like the Herrerasaurus specimen.
A 2006 Space Shuttle experiment found that Salmonella typhimurium, a bacterium that can cause food poisoning, became more virulent when cultivated in space. On April 29, 2013, scientists in Rensselaer Polytechnic Institute, funded by NASA, reported that, during spaceflight on the International Space Station, microbes seem to adapt to the space environment in ways "not observed on Earth" and in ways that "can lead to increases in growth and virulence". More recently, in 2017, bacteria were found to be more resistant to antibiotics and to thrive in the near-weightlessness of space. Microorganisms have been observed to survive the vacuum of outer space.
Types[edit]
Infections are caused by infectious agents (pathogens) including:
Bacteria (e.g. Mycobacterium tuberculosis, Staphylococcus aureus, Escherichia coli, Clostridium botulinum, and Salmonella spp.)
Viruses and related agents such as viroids. (E.g. HIV, Rhinovirus, Lyssaviruses such as Rabies virus, Ebolavirus and Severe acute respiratory syndrome coronavirus 2)
Fungi, further subclassified into:
Ascomycota, including yeasts such as Candida (the most common fungal infection); filamentous fungi such as Aspergillus; Pneumocystis species; and dermatophytes, a group of organisms causing infection of skin and other superficial structures in humans.
Basidiomycota, including the human-pathogenic genus Cryptococcus.
Parasites, which are usually divided into:
Unicellular organisms (e.g. malaria, Toxoplasma, Babesia)
Macroparasites (worms or helminths) including nematodes such as parasitic roundworms and pinworms, tapeworms (cestodes), and flukes (trematodes, such as schistosomes). Diseases caused by helminths are sometimes termed infestations, but are sometimes called infections.
Arthropods such as ticks, mites, fleas, and lice, can also cause human disease, which conceptually are similar to infections, but invasion of a human or animal body by these macroparasites is usually termed infestation.
Prions (although they do not secrete toxins)
Signs and symptoms[edit]
The signs and symptoms of an infection depend on the type of disease. Some signs of infection affect the whole body generally, such as fatigue, loss of appetite, weight loss, fevers, night sweats, chills, aches and pains. Others are specific to individual body parts, such as skin rashes, coughing, or a runny nose.
In certain cases, infectious diseases may be asymptomatic for much or even all of their course in a given host. In the latter case, the disease may only be defined as a "disease" (which by definition means an illness) in hosts who secondarily become ill after contact with an asymptomatic carrier. An infection is not synonymous with an infectious disease, as some infections do not cause illness in a host.
Bacterial or viral[edit]
As bacterial and viral infections can both cause the same kinds of symptoms, it can be difficult to distinguish which is the cause of a specific infection. Distinguishing the two is important, since viral infections cannot be cured by antibiotics whereas bacterial infections can.
Comparison of viral and bacterial infection
Characteristic
Viral infection
Bacterial infection
Typical symptoms
In general, viral infections are systemic. This means they involve many different parts of the body or more than one body system at the same time; i.e. a runny nose, sinus congestion, cough, body aches etc. They can be local at times as in viral conjunctivitis or "pink eye" and herpes. Only a few viral infections are painful, like herpes. The pain of viral infections is often described as itchy or burning.
The classic symptoms of a bacterial infection are localized redness, heat, swelling and pain. One of the hallmarks of a bacterial infection is local pain, pain that is in a specific part of the body. For example, if a cut occurs and is infected with bacteria, pain occurs at the site of the infection. Bacterial throat pain is often characterized by more pain on one side of the throat. An ear infection is more likely to be diagnosed as bacterial if the pain occurs in only one ear. A cut that produces pus and milky-colored liquid is most likely infected.
Cause
Pathogenic viruses
Pathogenic bacteria
Pathophysiology[edit]
Chain of infection; the chain of events that lead to infection
There is a general chain of events that applies to infections, sometimes called the chain of infection or transmission chain. The chain of events involves several steps – which include the infectious agent, reservoir, entering a susceptible host, exit and transmission to new hosts. Each of the links must be present in a chronological order for an infection to develop. Understanding these steps helps health care workers target the infection and prevent it from occurring in the first place.
Colonization[edit]
Infection of an ingrown toenail; there is pus (yellow) and resultant inflammation (redness and swelling around the nail).
Infection begins when an organism successfully enters the body, grows and multiplies. This is referred to as colonization. Most humans are not easily infected. Those with compromised or weakened immune systems have an increased susceptibility to chronic or persistent infections. Individuals who have a suppressed immune system are particularly susceptible to opportunistic infections. Entrance to the host at host–pathogen interface, generally occurs through the mucosa in orifices like the oral cavity, nose, eyes, genitalia, anus, or the microbe can enter through open wounds. While a few organisms can grow at the initial site of entry, many migrate and cause systemic infection in different organs. Some pathogens grow within the host cells (intracellular) whereas others grow freely in bodily fluids.
Wound colonization refers to non-replicating microorganisms within the wound, while in infected wounds, replicating organisms exist and tissue is injured. All multicellular organisms are colonized to some degree by extrinsic organisms, and the vast majority of these exist in either a mutualistic or commensal relationship with the host. An example of the former is the anaerobic bacteria species, which colonizes the mammalian colon, and an example of the latter are the various species of staphylococcus that exist on human skin. Neither of these colonizations are considered infections. The difference between an infection and a colonization is often only a matter of circumstance. Non-pathogenic organisms can become pathogenic given specific conditions, and even the most virulent organism requires certain circumstances to cause a compromising infection. Some colonizing bacteria, such as Corynebacteria sp. and Viridans streptococci, prevent the adhesion and colonization of pathogenic bacteria and thus have a symbiotic relationship with the host, preventing infection and speeding wound healing.
This image depicts the steps of pathogenic infection.
The variables involved in the outcome of a host becoming inoculated by a pathogen and the ultimate outcome include:
the route of entry of the pathogen and the access to host regions that it gains
the intrinsic virulence of the particular organism
the quantity or load of the initial inoculant
the immune status of the host being colonized
As an example, several staphylococcal species remain harmless on the skin, but, when present in a normally sterile space, such as in the capsule of a joint or the peritoneum, multiply without resistance and cause harm.
An interesting fact that gas chromatography–mass spectrometry, 16S ribosomal RNA analysis, omics, and other advanced technologies have made more apparent to humans in recent decades is that microbial colonization is very common even in environments that humans think of as being nearly sterile. Because it is normal to have bacterial colonization, it is difficult to know which chronic wounds can be classified as infected and how much risk of progression exists. Despite the huge number of wounds seen in clinical practice, there are limited quality data for evaluated symptoms and signs. A review of chronic wounds in the Journal of the American Medical Association's "Rational Clinical Examination Series" quantified the importance of increased pain as an indicator of infection. The review showed that the most useful finding is an increase in the level of pain [likelihood ratio (LR) range, 11–20] makes infection much more likely, but the absence of pain (negative likelihood ratio range, 0.64–0.88) does not rule out infection (summary LR 0.64–0.88).
Disease[edit]
Disease can arise if the host's protective immune mechanisms are compromised and the organism inflicts damage on the host. Microorganisms can cause tissue damage by releasing a variety of toxins or destructive enzymes. For example, Clostridium tetani releases a toxin that paralyzes muscles, and staphylococcus releases toxins that produce shock and sepsis. Not all infectious agents cause disease in all hosts. For example, less than 5% of individuals infected with polio develop disease. On the other hand, some infectious agents are highly virulent. The prion causing mad cow disease and Creutzfeldt–Jakob disease invariably kills all animals and people that are infected.
Persistent infections occur because the body is unable to clear the organism after the initial infection. Persistent infections are characterized by the continual presence of the infectious organism, often as latent infection with occasional recurrent relapses of active infection. There are some viruses that can maintain a persistent infection by infecting different cells of the body. Some viruses once acquired never leave the body. A typical example is the herpes virus, which tends to hide in nerves and become reactivated when specific circumstances arise.
Persistent infections cause millions of deaths globally each year. Chronic infections by parasites account for a high morbidity and mortality in many underdeveloped countries.
Transmission[edit]
Main article: Transmission (medicine)
A southern house mosquito (Culex quinquefasciatus) is a vector that transmits the pathogens that cause West Nile fever and avian malaria among others.
For infecting organisms to survive and repeat the infection cycle in other hosts, they (or their progeny) must leave an existing reservoir and cause infection elsewhere. Infection transmission can take place via many potential routes:
Droplet contact, also known as the respiratory route, and the resultant infection can be termed airborne disease. If an infected person coughs or sneezes on another person the microorganisms, suspended in warm, moist droplets, may enter the body through the nose, mouth or eye surfaces.
Fecal-oral transmission, wherein foodstuffs or water become contaminated (by people not washing their hands before preparing food, or untreated sewage being released into a drinking water supply) and the people who eat and drink them become infected. Common fecal-oral transmitted pathogens include Vibrio cholerae, Giardia species, rotaviruses, Entamoeba histolytica, Escherichia coli, and tape worms. Most of these pathogens cause gastroenteritis.
Sexual transmission, with the result being called sexually transmitted infection.
Oral transmission, diseases that are transmitted primarily by oral means may be caught through direct oral contact such as kissing, or by indirect contact such as by sharing a drinking glass or a cigarette.
Transmission by direct contact, Some diseases that are transmissible by direct contact include athlete's foot, impetigo and warts.
Vehicle transmission, transmission by an inanimate reservoir (food, water, soil).
Vertical transmission, directly from the mother to an embryo, fetus or baby during pregnancy or childbirth. It can occur as a result of a pre-existing infection or one acquired during pregnancy.
Iatrogenic transmission, due to medical procedures such as injection or transplantation of infected material.
Vector-borne transmission, transmitted by a vector, which is an organism that does not cause disease itself but that transmits infection by conveying pathogens from one host to another.
The relationship between virulence versus transmissibility is complex; with studies have shown that there were no clear relationship between the two. There is still a small number of evidence that partially suggests a link between virulence and transmissibility.
Diagnosis[edit]
Diagnosis of infectious disease sometimes involves identifying an infectious agent either directly or indirectly. In practice most minor infectious diseases such as warts, cutaneous abscesses, respiratory system infections and diarrheal diseases are diagnosed by their clinical presentation and treated without knowledge of the specific causative agent. Conclusions about the cause of the disease are based upon the likelihood that a patient came in contact with a particular agent, the presence of a microbe in a community, and other epidemiological considerations. Given sufficient effort, all known infectious agents can be specifically identified.
Diagnosis of infectious disease is nearly always initiated by medical history and physical examination. More detailed identification techniques involve the culture of infectious agents isolated from a patient. Culture allows identification of infectious organisms by examining their microscopic features, by detecting the presence of substances produced by pathogens, and by directly identifying an organism by its genotype.
Many infectious organisms are identified without culture and microscopy. This is especially true for viruses, which cannot grow in culture. For some suspected pathogens, doctors may conduct tests that examine a patient's blood or other body fluids for antigens or antibodies that indicate presence of a specific pathogen that the doctor suspects.
Other techniques (such as X-rays, CAT scans, PET scans or NMR) are used to produce images of internal abnormalities resulting from the growth of an infectious agent. The images are useful in detection of, for example, a bone abscess or a spongiform encephalopathy produced by a prion.
The benefits of identification, however, are often greatly outweighed by the cost, as often there is no specific treatment, the cause is obvious, or the outcome of an infection is likely to be benign.
Symptomatic diagnostics[edit]
The diagnosis is aided by the presenting symptoms in any individual with an infectious disease, yet it usually needs additional diagnostic techniques to confirm the suspicion. Some signs are specifically characteristic and indicative of a disease and are called pathognomonic signs; but these are rare. Not all infections are symptomatic.
In children the presence of cyanosis, rapid breathing, poor peripheral perfusion, or a petechial rash increases the risk of a serious infection by greater than 5 fold. Other important indicators include parental concern, clinical instinct, and temperature greater than 40 °C.
Microbial culture[edit]
Four nutrient agar plates growing colonies of common Gram negative bacteria
Many diagnostic approaches depend on microbiological culture to isolate a pathogen from the appropriate clinical specimen. In a microbial culture, a growth medium is provided for a specific agent. A sample taken from potentially diseased tissue or fluid is then tested for the presence of an infectious agent able to grow within that medium. Many pathogenic bacteria are easily grown on nutrient agar, a form of solid medium that supplies carbohydrates and proteins necessary for growth, along with copious amounts of water. A single bacterium will grow into a visible mound on the surface of the plate called a colony, which may be separated from other colonies or melded together into a "lawn". The size, color, shape and form of a colony is characteristic of the bacterial species, its specific genetic makeup (its strain), and the environment that supports its growth. Other ingredients are often added to the plate to aid in identification. Plates may contain substances that permit the growth of some bacteria and not others, or that change color in response to certain bacteria and not others. Bacteriological plates such as these are commonly used in the clinical identification of infectious bacterium. Microbial culture may also be used in the identification of viruses: the medium, in this case, being cells grown in culture that the virus can infect, and then alter or kill. In the case of viral identification, a region of dead cells results from viral growth, and is called a "plaque". Eukaryotic parasites may also be grown in culture as a means of identifying a particular agent.
In the absence of suitable plate culture techniques, some microbes require culture within live animals. Bacteria such as Mycobacterium leprae and Treponema pallidum can be grown in animals, although serological and microscopic techniques make the use of live animals unnecessary. Viruses are also usually identified using alternatives to growth in culture or animals. Some viruses may be grown in embryonated eggs. Another useful identification method is Xenodiagnosis, or the use of a vector to support the growth of an infectious agent. Chagas disease is the most significant example, because it is difficult to directly demonstrate the presence of the causative agent, Trypanosoma cruzi in a patient, which therefore makes it difficult to definitively make a diagnosis. In this case, xenodiagnosis involves the use of the vector of the Chagas agent T. cruzi, an uninfected triatomine bug, which takes a blood meal from a person suspected of having been infected. The bug is later inspected for growth of T. cruzi within its gut.
Microscopy[edit]
Another principal tool in the diagnosis of infectious disease is microscopy. Virtually all of the culture techniques discussed above rely, at some point, on microscopic examination for definitive identification of the infectious agent. Microscopy may be carried out with simple instruments, such as the compound light microscope, or with instruments as complex as an electron microscope. Samples obtained from patients may be viewed directly under the light microscope, and can often rapidly lead to identification. Microscopy is often also used in conjunction with biochemical staining techniques, and can be made exquisitely specific when used in combination with antibody based techniques. For example, the use of antibodies made artificially fluorescent (fluorescently labeled antibodies) can be directed to bind to and identify a specific antigens present on a pathogen. A fluorescence microscope is then used to detect fluorescently labeled antibodies bound to internalized antigens within clinical samples or cultured cells. This technique is especially useful in the diagnosis of viral diseases, where the light microscope is incapable of identifying a virus directly.
Other microscopic procedures may also aid in identifying infectious agents. Almost all cells readily stain with a number of basic dyes due to the electrostatic attraction between negatively charged cellular molecules and the positive charge on the dye. A cell is normally transparent under a microscope, and using a stain increases the contrast of a cell with its background. Staining a cell with a dye such as Giemsa stain or crystal violet allows a microscopist to describe its size, shape, internal and external components and its associations with other cells. The response of bacteria to different staining procedures is used in the taxonomic classification of microbes as well. Two methods, the Gram stain and the acid-fast stain, are the standard approaches used to classify bacteria and to diagnosis of disease. The Gram stain identifies the bacterial groups Bacillota and Actinomycetota, both of which contain many significant human pathogens. The acid-fast staining procedure identifies the Actinomycetota genera Mycobacterium and Nocardia.
Biochemical tests[edit]
Biochemical tests used in the identification of infectious agents include the detection of metabolic or enzymatic products characteristic of a particular infectious agent. Since bacteria ferment carbohydrates in patterns characteristic of their genus and species, the detection of fermentation products is commonly used in bacterial identification. Acids, alcohols and gases are usually detected in these tests when bacteria are grown in selective liquid or solid media.
The isolation of enzymes from infected tissue can also provide the basis of a biochemical diagnosis of an infectious disease. For example, humans can make neither RNA replicases nor reverse transcriptase, and the presence of these enzymes are characteristic., of specific types of viral infections. The ability of the viral protein hemagglutinin to bind red blood cells together into a detectable matrix may also be characterized as a biochemical test for viral infection, although strictly speaking hemagglutinin is not an enzyme and has no metabolic function.
Serological methods are highly sensitive, specific and often extremely rapid tests used to identify microorganisms. These tests are based upon the ability of an antibody to bind specifically to an antigen. The antigen, usually a protein or carbohydrate made by an infectious agent, is bound by the antibody. This binding then sets off a chain of events that can be visibly obvious in various ways, dependent upon the test. For example, "Strep throat" is often diagnosed within minutes, and is based on the appearance of antigens made by the causative agent, S. pyogenes, that is retrieved from a patient's throat with a cotton swab. Serological tests, if available, are usually the preferred route of identification, however the tests are costly to develop and the reagents used in the test often require refrigeration. Some serological methods are extremely costly, although when commonly used, such as with the "strep test", they can be inexpensive.
Complex serological techniques have been developed into what are known as immunoassays. Immunoassays can use the basic antibody – antigen binding as the basis to produce an electro-magnetic or particle radiation signal, which can be detected by some form of instrumentation. Signal of unknowns can be compared to that of standards allowing quantitation of the target antigen. To aid in the diagnosis of infectious diseases, immunoassays can detect or measure antigens from either infectious agents or proteins generated by an infected organism in response to a foreign agent. For example, immunoassay A may detect the presence of a surface protein from a virus particle. Immunoassay B on the other hand may detect or measure antibodies produced by an organism's immune system that are made to neutralize and allow the destruction of the virus.
Instrumentation can be used to read extremely small signals created by secondary reactions linked to the antibody – antigen binding. Instrumentation can control sampling, reagent use, reaction times, signal detection, calculation of results, and data management to yield a cost-effective automated process for diagnosis of infectious disease.
PCR-based diagnostics[edit]
Nucleic acid testing conducted using an Abbott Laboratories ID Now device
Technologies based upon the polymerase chain reaction (PCR) method will become nearly ubiquitous gold standards of diagnostics of the near future, for several reasons. First, the catalog of infectious agents has grown to the point that virtually all of the significant infectious agents of the human population have been identified. Second, an infectious agent must grow within the human body to cause disease; essentially it must amplify its own nucleic acids in order to cause a disease. This amplification of nucleic acid in infected tissue offers an opportunity to detect the infectious agent by using PCR. Third, the essential tools for directing PCR, primers, are derived from the genomes of infectious agents, and with time those genomes will be known, if they are not already.
Thus, the technological ability to detect any infectious agent rapidly and specifically are currently available. The only remaining blockades to the use of PCR as a standard tool of diagnosis are in its cost and application, neither of which is insurmountable. The diagnosis of a few diseases will not benefit from the development of PCR methods, such as some of the clostridial diseases (tetanus and botulism). These diseases are fundamentally biological poisonings by relatively small numbers of infectious bacteria that produce extremely potent neurotoxins. A significant proliferation of the infectious agent does not occur, this limits the ability of PCR to detect the presence of any bacteria.
Metagenomic sequencing[edit]
This section does not cite any sources. Please help improve this section by adding citations to reliable sources. Unsourced material may be challenged and removed. (November 2023) (Learn how and when to remove this template message)
Given the wide range of bacterial, viral, fungal, protozoal, and helminthic pathogens that cause debilitating and life-threatening illnesses, the ability to quickly identify the cause of infection is important yet often challenging. For example, more than half of cases of encephalitis, a severe illness affecting the brain, remain undiagnosed, despite extensive testing using the standard of care (microbiological culture) and state-of-the-art clinical laboratory methods. Metagenomic sequencing-based diagnostic tests are currently being developed for clinical use and show promise as a sensitive, specific, and rapid way to diagnose infection using a single all-encompassing test. This test is similar to current PCR tests; however, an untargeted whole genome amplification is used rather than primers for a specific infectious agent. This amplification step is followed by next-generation sequencing or third-generation sequencing, alignment comparisons, and taxonomic classification using large databases of thousands of pathogen and commensal reference genomes. Simultaneously, antimicrobial resistance genes within pathogen and plasmid genomes are sequenced and aligned to the taxonomically classified pathogen genomes to generate an antimicrobial resistance profile – analogous to antibiotic sensitivity testing – to facilitate antimicrobial stewardship and allow for the optimization of treatment using the most effective drugs for a patient's infection.
Metagenomic sequencing could prove especially useful for diagnosis when the patient is immunocompromised. An ever-wider array of infectious agents can cause serious harm to individuals with immunosuppression, so clinical screening must often be broader. Additionally, the expression of symptoms is often atypical, making a clinical diagnosis based on presentation more difficult. Thirdly, diagnostic methods that rely on the detection of antibodies are more likely to fail. A rapid, sensitive, specific, and untargeted test for all known human pathogens that detects the presence of the organism's DNA rather than antibodies is therefore highly desirable.
Indication of tests[edit]
A temporary drive-in testing site for COVID-19 set up with tents in a parking lot
There is usually an indication for a specific identification of an infectious agent only when such identification can aid in the treatment or prevention of the disease, or to advance knowledge of the course of an illness prior to the development of effective therapeutic or preventative measures. For example, in the early 1980s, prior to the appearance of AZT for the treatment of AIDS, the course of the disease was closely followed by monitoring the composition of patient blood samples, even though the outcome would not offer the patient any further treatment options. In part, these studies on the appearance of HIV in specific communities permitted the advancement of hypotheses as to the route of transmission of the virus. By understanding how the disease was transmitted, resources could be targeted to the communities at greatest risk in campaigns aimed at reducing the number of new infections. The specific serological diagnostic identification, and later genotypic or molecular identification, of HIV also enabled the development of hypotheses as to the temporal and geographical origins of the virus, as well as a myriad of other hypothesis. The development of molecular diagnostic tools have enabled physicians and researchers to monitor the efficacy of treatment with anti-retroviral drugs. Molecular diagnostics are now commonly used to identify HIV in healthy people long before the onset of illness and have been used to demonstrate the existence of people who are genetically resistant to HIV infection. Thus, while there still is no cure for AIDS, there is great therapeutic and predictive benefit to identifying the virus and monitoring the virus levels within the blood of infected individuals, both for the patient and for the community at large.
Classification[edit]
Subclinical versus clinical (latent versus apparent)[edit]
Symptomatic infections are apparent and clinical, whereas an infection that is active but does not produce noticeable symptoms may be called inapparent, silent, subclinical, or occult. An infection that is inactive or dormant is called a latent infection. An example of a latent bacterial infection is latent tuberculosis. Some viral infections can also be latent, examples of latent viral infections are any of those from the Herpesviridae family.
The word infection can denote any presence of a particular pathogen at all (no matter how little) but also is often used in a sense implying a clinically apparent infection (in other words, a case of infectious disease). This fact occasionally creates some ambiguity or prompts some usage discussion; to get around this it is common for health professionals to speak of colonization (rather than infection) when they mean that some of the pathogens are present but that no clinically apparent infection (no disease) is present.
Course of infection[edit]
Different terms are used to describe how and where infections present over time. In an acute infection, symptoms develop rapidly; its course can either be rapid or protracted. In chronic infection, symptoms usually develop gradually over weeks or months and are slow to resolve. In subacute infections, symptoms take longer to develop than in acute infections but arise more quickly than those of chronic infections. A focal infection is an initial site of infection from which organisms travel via the bloodstream to another area of the body.
Primary versus opportunistic[edit]
See also: Coinfection
Among the many varieties of microorganisms, relatively few cause disease in otherwise healthy individuals. Infectious disease results from the interplay between those few pathogens and the defenses of the hosts they infect. The appearance and severity of disease resulting from any pathogen depend upon the ability of that pathogen to damage the host as well as the ability of the host to resist the pathogen. However, a host's immune system can also cause damage to the host itself in an attempt to control the infection. Clinicians, therefore, classify infectious microorganisms or microbes according to the status of host defenses – either as primary pathogens or as opportunistic pathogens.
Primary pathogens[edit]
Primary pathogens cause disease as a result of their presence or activity within the normal, healthy host, and their intrinsic virulence (the severity of the disease they cause) is, in part, a necessary consequence of their need to reproduce and spread. Many of the most common primary pathogens of humans only infect humans, however, many serious diseases are caused by organisms acquired from the environment or that infect non-human hosts.
Opportunistic pathogens[edit]
Main article: Opportunistic infection
Opportunistic pathogens can cause an infectious disease in a host with depressed resistance (immunodeficiency) or if they have unusual access to the inside of the body (for example, via trauma). Opportunistic infection may be caused by microbes ordinarily in contact with the host, such as pathogenic bacteria or fungi in the gastrointestinal or the upper respiratory tract, and they may also result from (otherwise innocuous) microbes acquired from other hosts (as in Clostridium difficile colitis) or from the environment as a result of traumatic introduction (as in surgical wound infections or compound fractures). An opportunistic disease requires impairment of host defenses, which may occur as a result of genetic defects (such as chronic granulomatous disease), exposure to antimicrobial drugs or immunosuppressive chemicals (as might occur following poisoning or cancer chemotherapy), exposure to ionizing radiation, or as a result of an infectious disease with immunosuppressive activity (such as with measles, malaria or HIV disease). Primary pathogens may also cause more severe disease in a host with depressed resistance than would normally occur in an immunosufficient host.
Secondary infection[edit]
While a primary infection can practically be viewed as the root cause of an individual's current health problem, a secondary infection is a sequela or complication of that root cause. For example, an infection due to a burn or penetrating trauma (the root cause) is a secondary infection. Primary pathogens often cause primary infection and often cause secondary infection. Usually, opportunistic infections are viewed as secondary infections (because immunodeficiency or injury was the predisposing factor).
Other types of infection[edit]
Other types of infection consist of mixed, iatrogenic, nosocomial, and community-acquired infection. A mixed infection is an infection that is caused by two or more pathogens. An example of this is appendicitis, which is caused by Bacteroides fragilis and Escherichia coli. The second is an iatrogenic infection. This type of infection is one that is transmitted from a health care worker to a patient. A nosocomial infection is also one that occurs in a health care setting. Nosocomial infections are those that are acquired during a hospital stay. Lastly, a community-acquired infection is one in which the infection is acquired from a whole community.
Infectious or not[edit]
One manner of proving that a given disease is infectious, is to satisfy Koch's postulates (first proposed by Robert Koch), which require that first, the infectious agent be identifiable only in patients who have the disease, and not in healthy controls, and second, that patients who contract the infectious agent also develop the disease. These postulates were first used in the discovery that Mycobacteria species cause tuberculosis.
However, Koch's postulates cannot usually be tested in modern practice for ethical reasons. Proving them would require experimental infection of a healthy individual with a pathogen produced as a pure culture. Conversely, even clearly infectious diseases do not always meet the infectious criteria; for example, Treponema pallidum, the causative spirochete of syphilis, cannot be cultured in vitro – however the organism can be cultured in rabbit testes. It is less clear that a pure culture comes from an animal source serving as host than it is when derived from microbes derived from plate culture.
Epidemiology, or the study and analysis of who, why and where disease occurs, and what determines whether various populations have a disease, is another important tool used to understand infectious disease. Epidemiologists may determine differences among groups within a population, such as whether certain age groups have a greater or lesser rate of infection; whether groups living in different neighborhoods are more likely to be infected; and by other factors, such as gender and race. Researchers also may assess whether a disease outbreak is sporadic, or just an occasional occurrence; endemic, with a steady level of regular cases occurring in a region; epidemic, with a fast arising, and unusually high number of cases in a region; or pandemic, which is a global epidemic. If the cause of the infectious disease is unknown, epidemiology can be used to assist with tracking down the sources of infection.
Contagiousness[edit]
Infectious diseases are sometimes called contagious diseases when they are easily transmitted by contact with an ill person or their secretions (e.g., influenza). Thus, a contagious disease is a subset of infectious disease that is especially infective or easily transmitted. Other types of infectious, transmissible, or communicable diseases with more specialized routes of infection, such as vector transmission or sexual transmission, are usually not regarded as "contagious", and often do not require medical isolation (sometimes loosely called quarantine) of those affected. However, this specialized connotation of the word "contagious" and "contagious disease" (easy transmissibility) is not always respected in popular use.
Infectious diseases are commonly transmitted from person to person through direct contact. The types of contact are through person to person and droplet spread. Indirect contact such as airborne transmission, contaminated objects, food and drinking water, animal person contact, animal reservoirs, insect bites, and environmental reservoirs are another way infectious diseases are transmitted.
By anatomic location[edit]
Infections can be classified by the anatomic location or organ system infected, including:
Urinary tract infection
Skin infection
Respiratory tract infection
Odontogenic infection (an infection that originates within a tooth or in the closely surrounding tissues)
Vaginal infections
Intra-amniotic infection
In addition, locations of inflammation where infection is the most common cause include pneumonia, meningitis and salpingitis.
Prevention[edit]
Main articles: Public health and Infection control
Washing one's hands, a form of hygiene, is an effective way to prevent the spread of infectious disease.
Techniques like hand washing, wearing gowns, and wearing face masks can help prevent infections from being passed from one person to another. Aseptic technique was introduced in medicine and surgery in the late 19th century and greatly reduced the incidence of infections caused by surgery. Frequent hand washing remains the most important defense against the spread of unwanted organisms. There are other forms of prevention such as avoiding the use of illicit drugs, using a condom, wearing gloves, and having a healthy lifestyle with a balanced diet and regular exercise. Cooking foods well and avoiding foods that have been left outside for a long time is also important.
Antimicrobial substances used to prevent transmission of infections include:
antiseptics, which are applied to living tissue/skin
disinfectants, which destroy microorganisms found on non-living objects.
antibiotics, called prophylactic when given as prevention rather as treatment of infection. However, long term use of antibiotics leads to resistance of bacteria. While humans do not become immune to antibiotics, the bacteria does. Thus, avoiding using antibiotics longer than necessary helps preventing bacteria from forming mutations that aide in antibiotic resistance.
One of the ways to prevent or slow down the transmission of infectious diseases is to recognize the different characteristics of various diseases. Some critical disease characteristics that should be evaluated include virulence, distance traveled by those affected, and level of contagiousness. The human strains of Ebola virus, for example, incapacitate those infected extremely quickly and kill them soon after. As a result, those affected by this disease do not have the opportunity to travel very far from the initial infection zone. Also, this virus must spread through skin lesions or permeable membranes such as the eye. Thus, the initial stage of Ebola is not very contagious since its victims experience only internal hemorrhaging. As a result of the above features, the spread of Ebola is very rapid and usually stays within a relatively confined geographical area. In contrast, the human immunodeficiency virus (HIV) kills its victims very slowly by attacking their immune system. As a result, many of its victims transmit the virus to other individuals before even realizing that they are carrying the disease. Also, the relatively low virulence allows its victims to travel long distances, increasing the likelihood of an epidemic.
Another effective way to decrease the transmission rate of infectious diseases is to recognize the effects of small-world networks. In epidemics, there are often extensive interactions within hubs or groups of infected individuals and other interactions within discrete hubs of susceptible individuals. Despite the low interaction between discrete hubs, the disease can jump and spread in a susceptible hub via a single or few interactions with an infected hub. Thus, infection rates in small-world networks can be reduced somewhat if interactions between individuals within infected hubs are eliminated (Figure 1). However, infection rates can be drastically reduced if the main focus is on the prevention of transmission jumps between hubs. The use of needle exchange programs in areas with a high density of drug users with HIV is an example of the successful implementation of this treatment method. Another example is the use of ring culling or vaccination of potentially susceptible livestock in adjacent farms to prevent the spread of the foot-and-mouth virus in 2001.
A general method to prevent transmission of vector-borne pathogens is pest control.
In cases where infection is merely suspected, individuals may be quarantined until the incubation period has passed and the disease manifests itself or the person remains healthy. Groups may undergo quarantine, or in the case of communities, a cordon sanitaire may be imposed to prevent infection from spreading beyond the community, or in the case of protective sequestration, into a community. Public health authorities may implement other forms of social distancing, such as school closings, lockdowns or temporary restrictions (e.g. circuit breakers) to control an epidemic.
Immunity[edit]
Mary Mallon (a.k.a. Typhoid Mary) was an asymptomatic carrier of typhoid fever. Over the course of her career as a cook, she infected 53 people, three of whom died.
Infection with most pathogens does not result in death of the host and the offending organism is ultimately cleared after the symptoms of the disease have waned. This process requires immune mechanisms to kill or inactivate the inoculum of the pathogen. Specific acquired immunity against infectious diseases may be mediated by antibodies and/or T lymphocytes. Immunity mediated by these two factors may be manifested by:
a direct effect upon a pathogen, such as antibody-initiated complement-dependent bacteriolysis, opsonoization, phagocytosis and killing, as occurs for some bacteria,
neutralization of viruses so that these organisms cannot enter cells,
or by T lymphocytes, which will kill a cell parasitized by a microorganism.
The immune system response to a microorganism often causes symptoms such as a high fever and inflammation, and has the potential to be more devastating than direct damage caused by a microbe.
Resistance to infection (immunity) may be acquired following a disease, by asymptomatic carriage of the pathogen, by harboring an organism with a similar structure (crossreacting), or by vaccination. Knowledge of the protective antigens and specific acquired host immune factors is more complete for primary pathogens than for opportunistic pathogens. There is also the phenomenon of herd immunity which offers a measure of protection to those otherwise vulnerable people when a large enough proportion of the population has acquired immunity from certain infections.
Immune resistance to an infectious disease requires a critical level of either antigen-specific antibodies and/or T cells when the host encounters the pathogen. Some individuals develop natural serum antibodies to the surface polysaccharides of some agents although they have had little or no contact with the agent, these natural antibodies confer specific protection to adults and are passively transmitted to newborns.
Host genetic factors[edit]
The organism that is the target of an infecting action of a specific infectious agent is called the host. The host harbouring an agent that is in a mature or sexually active stage phase is called the definitive host. The intermediate host comes in contact during the larvae stage. A host can be anything living and can attain to asexual and sexual reproduction.
The clearance of the pathogens, either treatment-induced or spontaneous, it can be influenced by the genetic variants carried by the individual patients. For instance, for genotype 1 hepatitis C treated with Pegylated interferon-alpha-2a or Pegylated interferon-alpha-2b (brand names Pegasys or PEG-Intron) combined with ribavirin, it has been shown that genetic polymorphisms near the human IL28B gene, encoding interferon lambda 3, are associated with significant differences in the treatment-induced clearance of the virus. This finding, originally reported in Nature, showed that genotype 1 hepatitis C patients carrying certain genetic variant alleles near the IL28B gene are more possibly to achieve sustained virological response after the treatment than others. Later report from Nature demonstrated that the same genetic variants are also associated with the natural clearance of the genotype 1 hepatitis C virus.
Treatments[edit]
When infection attacks the body, anti-infective drugs can suppress the infection. Several broad types of anti-infective drugs exist, depending on the type of organism targeted; they include antibacterial (antibiotic; including antitubercular), antiviral, antifungal and antiparasitic (including antiprotozoal and antihelminthic) agents. Depending on the severity and the type of infection, the antibiotic may be given by mouth or by injection, or may be applied topically. Severe infections of the brain are usually treated with intravenous antibiotics. Sometimes, multiple antibiotics are used in case there is resistance to one antibiotic. Antibiotics only work for bacteria and do not affect viruses. Antibiotics work by slowing down the multiplication of bacteria or killing the bacteria. The most common classes of antibiotics used in medicine include penicillin, cephalosporins, aminoglycosides, macrolides, quinolones and tetracyclines.
Not all infections require treatment, and for many self-limiting infections the treatment may cause more side-effects than benefits. Antimicrobial stewardship is the concept that healthcare providers should treat an infection with an antimicrobial that specifically works well for the target pathogen for the shortest amount of time and to only treat when there is a known or highly suspected pathogen that will respond to the medication.
Susceptibility to infection[edit]
Pandemics such as COVID-19 show that people dramatically differ in their susceptibility to infection. This may be because of general health, age, or their immune status, e.g. when they have been infected previously. However, it also has become clear that there are genetic factor which determine susceptibility to infection. For instance, up to 40% of SARS-CoV-2 infections may be asymptomatic, suggesting that many people are naturally protected from disease. Large genetic studies have defined risk factors for severe SARS-CoV-2 infections, and genome sequences from 659 patients with severe COVID-19 revealed genetic variants that appear to be associated with life-threatening disease. One gene identified in these studies is type I interferon (IFN). Autoantibodies against type I IFNs were found in up to 13.7% of patients with life-threatening COVID-19, indicating that a complex interaction between genetics and the immune system is important for natural resistance to Covid.
Similarly, mutations in the ERAP2 gene, encoding endoplasmic reticulum aminopeptidase 2, seem to increase the susceptibility to the plague, the disease caused by an infection with the bacteria Yersinia pestis. People who inherited two copies of a complete variant of the gene were twice as likely to have survived the plague as those who inherited two copies of a truncated variant.
Susceptibility also determined the epidemiology of infection, given that different populations have different genetic and environmental conditions that affect infections.
Epidemiology[edit]
Deaths due to infectious and parasitic diseases per million persons in 2012: 28–81 82–114 115–171 172–212 213–283 284–516 517–1,193 1,194–2,476 2,477–3,954 3,955–6,812
See also: Epidemic and Pandemic
Disability-adjusted life year for infectious and parasitic diseases per 100,000 inhabitants in 2004: no data ≤250 250–500 500–1000 1000–2000 2000–3000 3000–4000 4000–5000 5000–6250 6250–12,500 12,500–25,000 25,000–50,000 ≥50,000
In 2010, about 10 million people died of infectious diseases.
The World Health Organization collects information on global deaths by International Classification of Disease (ICD) code categories. The following table lists the top infectious disease by number of deaths in 2002. 1993 data is included for comparison.
Worldwide mortality due to infectious diseases
Rank
Cause of death
Deaths 2002 (in millions)
Percentage of all deaths
Deaths 1993 (in millions)
1993 Rank
N/A
All infectious diseases
14.7
25.9%
16.4
32.2%
1
Lower respiratory infections
3.9
6.9%
4.1
1
2
HIV/AIDS
2.8
4.9%
0.7
7
3
Diarrheal diseases
1.8
3.2%
3.0
2
4
Tuberculosis (TB)
1.6
2.7%
2.7
3
5
Malaria
1.3
2.2%
2.0
4
6
Measles
0.6
1.1%
1.1
5
7
Pertussis
0.29
0.5%
0.36
7
8
Tetanus
0.21
0.4%
0.15
12
9
Meningitis
0.17
0.3%
0.25
8
10
Syphilis
0.16
0.3%
0.19
11
11
Hepatitis B
0.10
0.2%
0.93
6
12–17
Tropical diseases (6)
0.13
0.2%
0.53
9, 10, 16–18
Note: Other causes of death include maternal and perinatal conditions (5.2%), nutritional deficiencies (0.9%), noncommunicable conditions (58.8%), and injuries (9.1%).
The top three single agent/disease killers are HIV/AIDS, TB and malaria. While the number of deaths due to nearly every disease have decreased, deaths due to HIV/AIDS have increased fourfold. Childhood diseases include pertussis, poliomyelitis, diphtheria, measles and tetanus. Children also make up a large percentage of lower respiratory and diarrheal deaths. In 2012, approximately 3.1 million people have died due to lower respiratory infections, making it the number 4 leading cause of death in the world.
Historic pandemics[edit]
See also: List of epidemics
The Great Plague of Marseille in 1720 killed 100,000 people in the city and the surrounding provinces.
With their potential for unpredictable and explosive impacts, infectious diseases have been major actors in human history. A pandemic (or global epidemic) is a disease that affects people over an extensive geographical area. For example:
Plague of Justinian, from 541 to 542, killed between 50% and 60% of Europe's population.
The Black Death of 1347 to 1352 killed 25 million in Europe over five years. The plague reduced the old world population from an estimated 450 million to between 350 and 375 million in the 14th century.
The introduction of smallpox, measles, and typhus to the areas of Central and South America by European explorers during the 15th and 16th centuries caused pandemics among the native inhabitants. Between 1518 and 1568 disease pandemics are said to have caused the population of Mexico to fall from 20 million to 3 million.
The first European influenza epidemic occurred between 1556 and 1560, with an estimated mortality rate of 20%.
Smallpox killed an estimated 60 million Europeans during the 18th century (approximately 400,000 per year). Up to 30% of those infected, including 80% of the children under 5 years of age, died from the disease, and one-third of the survivors went blind.
In the 19th century, tuberculosis killed an estimated one-quarter of the adult population of Europe; by 1918 one in six deaths in France were still caused by TB.
The Influenza Pandemic of 1918 (or the Spanish flu) killed 25–50 million people (about 2% of world population of 1.7 billion). Today Influenza kills about 250,000 to 500,000 worldwide each year.
Emerging diseases[edit]
In most cases, microorganisms live in harmony with their hosts via mutual or commensal interactions. Diseases can emerge when existing parasites become pathogenic or when new pathogenic parasites enter a new host.
Coevolution between parasite and host can lead to hosts becoming resistant to the parasites or the parasites may evolve greater virulence, leading to immunopathological disease.
Human activity is involved with many emerging infectious diseases, such as environmental change enabling a parasite to occupy new niches. When that happens, a pathogen that had been confined to a remote habitat has a wider distribution and possibly a new host organism. Parasites jumping from nonhuman to human hosts are known as zoonoses. Under disease invasion, when a parasite invades a new host species, it may become pathogenic in the new host.
Several human activities have led to the emergence of zoonotic human pathogens, including viruses, bacteria, protozoa, and rickettsia, and spread of vector-borne diseases, see also globalization and disease and wildlife disease:
Encroachment on wildlife habitats. The construction of new villages and housing developments in rural areas force animals to live in dense populations, creating opportunities for microbes to mutate and emerge.
Changes in agriculture. The introduction of new crops attracts new crop pests and the microbes they carry to farming communities, exposing people to unfamiliar diseases.
The destruction of rain forests. As countries make use of their rain forests, by building roads through forests and clearing areas for settlement or commercial ventures, people encounter insects and other animals harboring previously unknown microorganisms.
Uncontrolled urbanization. The rapid growth of cities in many developing countries tends to concentrate large numbers of people into crowded areas with poor sanitation. These conditions foster transmission of contagious diseases.
Modern transport. Ships and other cargo carriers often harbor unintended "passengers", that can spread diseases to faraway destinations. While with international jet-airplane travel, people infected with a disease can carry it to distant lands, or home to their families, before their first symptoms appear.
Germ theory of disease[edit]
Main article: Germ theory of disease
In Antiquity, the Greek historian Thucydides (c. 460 – c. 400 BCE) was the first person to write, in his account of the plague of Athens, that diseases could spread from an infected person to others. In his On the Different Types of Fever (c. 175 AD), the Greco-Roman physician Galen speculated that plagues were spread by "certain seeds of plague", which were present in the air. In the Sushruta Samhita, the ancient Indian physician Sushruta theorized: "Leprosy, fever, consumption, diseases of the eye, and other infectious diseases spread from one person to another by sexual union, physical contact, eating together, sleeping together, sitting together, and the use of same clothes, garlands and pastes." This book has been dated to about the sixth century BC.
A basic form of contagion theory was proposed by Persian physician Ibn Sina (known as Avicenna in Europe) in The Canon of Medicine (1025), which later became the most authoritative medical textbook in Europe up until the 16th century. In Book IV of the Canon, Ibn Sina discussed epidemics, outlining the classical miasma theory and attempting to blend it with his own early contagion theory. He mentioned that people can transmit disease to others by breath, noted contagion with tuberculosis, and discussed the transmission of disease through water and dirt. The concept of invisible contagion was later discussed by several Islamic scholars in the Ayyubid Sultanate who referred to them as najasat ("impure substances"). The fiqh scholar Ibn al-Haj al-Abdari (c. 1250–1336), while discussing Islamic diet and hygiene, gave warnings about how contagion can contaminate water, food, and garments, and could spread through the water supply, and may have implied contagion to be unseen particles.
When the Black Death bubonic plague reached Al-Andalus in the 14th century, the Arab physicians Ibn Khatima (c. 1369) and Ibn al-Khatib (1313–1374) hypothesised that infectious diseases were caused by "minute bodies" and described how they can be transmitted through garments, vessels and earrings. Ideas of contagion became more popular in Europe during the Renaissance, particularly through the writing of the Italian physician Girolamo Fracastoro. Anton van Leeuwenhoek (1632–1723) advanced the science of microscopy by being the first to observe microorganisms, allowing for easy visualization of bacteria.
In the mid-19th century John Snow and William Budd did important work demonstrating the contagiousness of typhoid and cholera through contaminated water. Both are credited with decreasing epidemics of cholera in their towns by implementing measures to prevent contamination of water. Louis Pasteur proved beyond doubt that certain diseases are caused by infectious agents, and developed a vaccine for rabies. Robert Koch provided the study of infectious diseases with a scientific basis known as Koch's postulates. Edward Jenner, Jonas Salk and Albert Sabin developed effective vaccines for smallpox and polio, which would later result in the eradication and near-eradication of these diseases, respectively. Alexander Fleming discovered the world's first antibiotic, penicillin, which Florey and Chain then developed. Gerhard Domagk developed sulphonamides, the first broad spectrum synthetic antibacterial drugs.
Medical specialists[edit]
The medical treatment of infectious diseases falls into the medical field of Infectious Disease and in some cases the study of propagation pertains to the field of Epidemiology. Generally, infections are initially diagnosed by primary care physicians or internal medicine specialists. For example, an "uncomplicated" pneumonia will generally be treated by the internist or the pulmonologist (lung physician). The work of the infectious diseases specialist therefore entails working with both patients and general practitioners, as well as laboratory scientists, immunologists, bacteriologists and other specialists.
An infectious disease team may be alerted when:
The disease has not been definitively diagnosed after an initial workup
The patient is immunocompromised (for example, in AIDS or after chemotherapy);
The infectious agent is of an uncommon nature (e.g. tropical diseases);
The disease has not responded to first line antibiotics;
The disease might be dangerous to other patients, and the patient might have to be isolated
Society and culture[edit]
Several studies have reported associations between pathogen load in an area and human behavior. Higher pathogen load is associated with decreased size of ethnic and religious groups in an area. This may be due high pathogen load favoring avoidance of other groups, which may reduce pathogen transmission, or a high pathogen load preventing the creation of large settlements and armies that enforce a common culture. Higher pathogen load is also associated with more restricted sexual behavior, which may reduce pathogen transmission. It also associated with higher preferences for health and attractiveness in mates. Higher fertility rates and shorter or less parental care per child is another association that may be a compensation for the higher mortality rate. There is also an association with polygyny which may be due to higher pathogen load, making selecting males with a high genetic resistance increasingly important. Higher pathogen load is also associated with more collectivism and less individualism, which may limit contacts with outside groups and infections. There are alternative explanations for at least some of the associations although some of these explanations may in turn ultimately be due to pathogen load. Thus, polygyny may also be due to a lower male: female ratio in these areas but this may ultimately be due to male infants having increased mortality from infectious diseases. Another example is that poor socioeconomic factors may ultimately in part be due to high pathogen load preventing economic development.
Fossil record[edit]
Main article: Paleopathology
Herrerasaurus skull
Evidence of infection in fossil remains is a subject of interest for paleopathologists, scientists who study occurrences of injuries and illness in extinct life forms. Signs of infection have been discovered in the bones of carnivorous dinosaurs. When present, however, these infections seem to tend to be confined to only small regions of the body. A skull attributed to the early carnivorous dinosaur Herrerasaurus ischigualastensis exhibits pit-like wounds surrounded by swollen and porous bone. The unusual texture of the bone around the wounds suggests they were affected by a short-lived, non-lethal infection. Scientists who studied the skull speculated that the bite marks were received in a fight with another Herrerasaurus. Other carnivorous dinosaurs with documented evidence of infection include Acrocanthosaurus, Allosaurus, Tyrannosaurus and a tyrannosaur from the Kirtland Formation. The infections from both tyrannosaurs were received by being bitten during a fight, like the Herrerasaurus specimen.
Outer space[edit]
Main articles: Effect of spaceflight on the human body, Medical treatment during spaceflight, and Space medicine
A 2006 Space Shuttle experiment found that Salmonella typhimurium, a bacterium that can cause food poisoning, became more virulent when cultivated in space. On April 29, 2013, scientists in Rensselaer Polytechnic Institute, funded by NASA, reported that, during spaceflight on the International Space Station, microbes seem to adapt to the space environment in ways "not observed on Earth" and in ways that "can lead to increases in growth and virulence". More recently, in 2017, bacteria were found to be more resistant to antibiotics and to thrive in the near-weightlessness of space. Microorganisms have been observed to survive the vacuum of outer space.
See also[edit]
Biological hazard
Blood-borne disease
Coinfection
Copenhagen Consensus
Cordon sanitaire
Epidemiological transition
Foodborne illness
Hospital-acquired infection
Eradication of infectious diseases
Infection control
Isolation (health care)
List of causes of death by rate
List of diseases caused by insects
List of infectious diseases
Mathematical modelling of infectious disease
Multiplicity of infection
Neglected tropical diseases
Sentinel surveillance
Spillover infection
Threshold host density
Transmission (medicine)
Vaccine-preventable diseases
Waterborne diseases | biology | 430918 | https://sv.wikipedia.org/wiki/Infektionssjukdom | Infektionssjukdom | En infektionssjukdom eller smittsam sjukdom är en sjukdom orsakad av ett smittämne. Smittämnen kan bestå av mikroorganismer i form av bakterier, amöbor, mikroskopiska svampar eller maskar. Även virus och prioner, som är icke levande smittämnen, orsakar infektionssjukdomar.
Infektionssjukdomar var under början av 2000-talet orsak till en fjärdedel av alla dödsfall globalt enligt WHO, men orsakar en minimal mängd av dödsfallen i Sverige. Mikroorganismer har stor förmåga att anpassa och förändra sig, vilket kan leda till att de utvecklar resistens som gör dem svåra att bekämpa. Mellan 180 och 260 personer avlider varje år i Sverige på grund av antibiotikaresistens (uppgifter från 2011). Mikroorganismernas förändringsförmågan kan också leda till att en mikroorganism som tidigare endast kunde angripa ett visst organ eller en viss art utvecklar en förmåga att angripa andra organsystem eller värdarter.
Normalt harmlösa mikroorganismer som till exempel lever i munhålan eller tarmkanalen kan orsaka allvarliga infektioner, så kallade opportunistiska infektioner om bäraren utsätts för speciella förhållanden. Oftast sker detta då bärarens immunsystem är försvagat. Mikroorganismer som orsakar sådana infektioner kallas opportunister.
Smitta
När smittämnena sprids mellan olika individer kallas det smitta. Spridningen kan ske på många olika sätt till exempel genom beröring, luft, vatten eller mat. Zoonoser kallas de infektioner som naturligt förs över mellan djur och människor. Infektionssjukdomen malaria sprids med myggor. Organismer som på detta sätt för över ett smittämne till en annan varelse kallas vektorer.
Individer som är smittbärare riskerar att föra över en infektionssjukdom till andra i sin omgivning. En smittbärare kan ha en symtomfri (asymtomatisk) infektion och ändå riskera att smitta andra. Infektionssjukdomen humant immunbristvirus (HIV) är ett exempel på en sjukdom som i början är symtomfri.
För att förhindra spridning av infektionssjukdomar används smittskydd. Att se till att vattensystem och avloppssystem fungerar på ett sätt som förhindrar spridning av smitta är viktigt. Hygien är också viktigt. Isolering av smittbärande individer är mer eller mindre effektiv beroende på hur smittan sprids. I Sverige har vi en myndighet kallad smittskyddsinstitutet som har till ansvar att se till att landets smittskydd fungerar effektivt i Sverige samt till att sprida information om smittsamma sjukdomar.
Antimikrobiell behandling
Infektionssjukdomar kan behandlas med antimikrobiell behandling. De läkemedel man använder mot bakterier kallas antibiotika. Mot virus använder man så kallade antivirala medel. Antimykotika används mot svampar. Mot maskar och encelliga organismer används antiparasitära preparat.
Antibiotika
Bakterier som orsakar infektioner kan behandlas med antibiotika. Verkan av antibiotika kan vara att den dödar bakterierna, vilket kallas bakteriecid. Om verkan av antibiotikan är att den hindrar bakteriers växt utan att döda kallas den bakteriostatisk. Antibiotika består av kemiska ämnen. Många av dessa ämnen härstammar från naturen, ett exempel på detta är penicillin som upptäcktes redan 1928. Pencillinerna är de vanligast använda antibiotiska läkemedlen än i dag. På grund av bakteriers förändringsförmåga behöver man vara försiktig med att använda antibiotika så att inte bakterierna utvecklar resistens mot läkemedlet. Till exempel försöker man ibland låta bli att behandla milda infektioner med antibiotika trots att patienten med en sådan behandling snabbare skulle tillfriskna. Vanligast är att man använder ordet antibiotika endast om läkemedel mot bakterier som orsakar infektionssjukdomar, men ibland används ordet även om läkemedel mot sjukdomsframkallande svampar, virus eller till och med cancerceller.
Antivirala medel
Det är ofta svårare att behandla virus än bakterier. Detta har dels sin förklaring i att virus förökar sig inuti värdorganismens celler och använder sig av värdcellens komponenter (organeller) för att reproducera sig. Således är det svårare att utveckla läkemedel som påverkar virusets reproduktionsförmåga, utan att påverka värdcellen i någon högre grad.
Antimykotika
Antimykotika används ofta vid ytliga infektioner som en lokalbehandling på det infekterade området. Infektioner förekommer ofta i hår, hud och naglar som behandlas med krämer, nagellack och schampon och liknande. Många sådana produkter är receptfria.
Hos personer med försvagat immunförsvar kan djupare svampinfektioner utvecklas. Exempel på orsaker till försvagat immunförsvar är cancer och cancerbehandling, obehandlad hivinfektion, och vid immunhämmande behandling efter organtransplantation.
Antiparasitära medel
Det finns många sorters antiparasitära medel. De som verkar mot just maskar kallas antihelmintika. Ett preparat kallat metronidazol motverkar infektioner orsakade av amöbor.
Exempel på infektionssjukdomar
spetälska
mjältbrand
borrelios
mul- och klövsjuka
tuberkulos
syfilis
pest
skabb
scharlakansfeber
Chagas sjukdom
denguefeber
dysenteri
kala-azar
kolera
schistosomiasis
tyfus
afrikansk sömnsjuka
Se även
Epidemi
Infektion
Kochs postulat
Nya infektionssjukdomar
Smitta
Organiska psykiska störningar
Referenser
http://www.ne.se
Noter | swedish | 0.476044 |
mechanism_mentally_tired/Mental_disorder.txt |
A mental disorder, also referred to as a mental illness, a mental health condition, or a psychiatric disorder, is a behavioral or mental pattern that causes significant distress or impairment of personal functioning. A mental disorder is also characterized by a clinically significant disturbance in an individual's cognition, emotional regulation, or behavior, often in a social context. Such disturbances may occur as single episodes, may be persistent, or may be relapsing–remitting. There are many different types of mental disorders, with signs and symptoms that vary widely between specific disorders. A mental disorder is one aspect of mental health.
The causes of mental disorders are often unclear. Theories incorporate findings from a range of fields. Disorders may be associated with particular regions or functions of the brain. Disorders are usually diagnosed or assessed by a mental health professional, such as a clinical psychologist, psychiatrist, psychiatric nurse, or clinical social worker, using various methods such as psychometric tests, but often relying on observation and questioning. Cultural and religious beliefs, as well as social norms, should be taken into account when making a diagnosis.
Services for mental disorders are usually based in psychiatric hospitals, outpatient clinics, or in the community, Treatments are provided by mental health professionals. Common treatment options are psychotherapy or psychiatric medication, while lifestyle changes, social interventions, peer support, and self-help are also options. In a minority of cases, there may be involuntary detention or treatment. Prevention programs have been shown to reduce depression.
In 2019, common mental disorders around the globe include: depression, which affects about 264 million people; dementia, which affects about 50 million; bipolar disorder, which affects about 45 million; and schizophrenia and other psychoses, which affect about 20 million people. Neurodevelopmental disorders include attention deficit hyperactivity disorder (ADHD), autism spectrum disorder (ASD), and intellectual disability, of which onset occurs early in the developmental period. Stigma and discrimination can add to the suffering and disability associated with mental disorders, leading to various social movements attempting to increase understanding and challenge social exclusion.
The definition and classification of mental disorders are key issues for researchers as well as service providers and those who may be diagnosed. For a mental state to be classified as a disorder, it generally needs to cause dysfunction. Most international clinical documents use the term mental "disorder", while "illness" is also common. It has been noted that using the term "mental" (i.e., of the mind) is not necessarily meant to imply separateness from the brain or body.
According to the fourth edition of the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV), published in 1994, a mental disorder is a psychological syndrome or pattern that is associated with distress (e.g., via a painful symptom), disability (impairment in one or more important areas of functioning), increased risk of death, or causes a significant loss of autonomy; however, it excludes normal responses such as the grief from loss of a loved one and also excludes deviant behavior for political, religious, or societal reasons not arising from a dysfunction in the individual.
DSM-IV predicates the definition with caveats, stating that, as in the case with many medical terms, mental disorder "lacks a consistent operational definition that covers all situations", noting that different levels of abstraction can be used for medical definitions, including pathology, symptomology, deviance from a normal range, or etiology, and that the same is true for mental disorders, so that sometimes one type of definition is appropriate and sometimes another, depending on the situation.
In 2013, the American Psychiatric Association (APA) redefined mental disorders in the DSM-5 as "a syndrome characterized by clinically significant disturbance in an individual's cognition, emotion regulation, or behavior that reflects a dysfunction in the psychological, biological, or developmental processes underlying mental functioning." The final draft of ICD-11 contains a very similar definition.
The terms "mental breakdown" or "nervous breakdown" may be used by the general population to mean a mental disorder. The terms "nervous breakdown" and "mental breakdown" have not been formally defined through a medical diagnostic system such as the DSM-5 or ICD-10 and are nearly absent from scientific literature regarding mental illness. Although "nervous breakdown" is not rigorously defined, surveys of laypersons suggest that the term refers to a specific acute time-limited reactive disorder involving symptoms such as anxiety or depression, usually precipitated by external stressors. Many health experts today refer to a nervous breakdown as a mental health crisis.
In addition to the concept of mental disorder, some people have argued for a return to the old-fashioned concept of nervous illness. In How Everyone Became Depressed: The Rise and Fall of the Nervous Breakdown (2013), Edward Shorter, a professor of psychiatry and the history of medicine, says:
There are currently two widely established systems that classify mental disorders:
Both of these list categories of disorder and provide standardized criteria for diagnosis. They have deliberately converged their codes in recent revisions so that the manuals are often broadly comparable, although significant differences remain. Other classification schemes may be used in non-western cultures, for example, the Chinese Classification of Mental Disorders, and other manuals may be used by those of alternative theoretical persuasions, such as the Psychodynamic Diagnostic Manual. In general, mental disorders are classified separately from neurological disorders, learning disabilities or intellectual disability.
Unlike the DSM and ICD, some approaches are not based on identifying distinct categories of disorder using dichotomous symptom profiles intended to separate the abnormal from the normal. There is significant scientific debate about the relative merits of categorical versus such non-categorical (or hybrid) schemes, also known as continuum or dimensional models. A spectrum approach may incorporate elements of both.
In the scientific and academic literature on the definition or classification of mental disorder, one extreme argues that it is entirely a matter of value judgements (including of what is normal) while another proposes that it is or could be entirely objective and scientific (including by reference to statistical norms). Common hybrid views argue that the concept of mental disorder is objective even if only a "fuzzy prototype" that can never be precisely defined, or conversely that the concept always involves a mixture of scientific facts and subjective value judgments. Although the diagnostic categories are referred to as 'disorders', they are presented as medical diseases, but are not validated in the same way as most medical diagnoses. Some neurologists argue that classification will only be reliable and valid when based on neurobiological features rather than clinical interview, while others suggest that the differing ideological and practical perspectives need to be better integrated.
The DSM and ICD approach remains under attack both because of the implied causality model and because some researchers believe it better to aim at underlying brain differences which can precede symptoms by many years.
The high degree of comorbidity between disorders in categorical models such as the DSM and ICD have led some to propose dimensional models. Studying comorbidity between disorders have demonstrated two latent (unobserved) factors or dimensions in the structure of mental disorders that are thought to possibly reflect etiological processes. These two dimensions reflect a distinction between internalizing disorders, such as mood or anxiety symptoms, and externalizing disorders such as behavioral or substance use symptoms. A single general factor of psychopathology, similar to the g factor for intelligence, has been empirically supported. The p factor model supports the internalizing-externalizing distinction, but also supports the formation of a third dimension of thought disorders such as schizophrenia. Biological evidence also supports the validity of the internalizing-externalizing structure of mental disorders, with twin and adoption studies supporting heritable factors for externalizing and internalizing disorders. A leading dimensional model is the Hierarchical Taxonomy of Psychopathology.
There are many different categories of mental disorder, and many different facets of human behavior and personality that can become disordered.
An anxiety disorder is anxiety or fear that interferes with normal functioning may be classified as an anxiety disorder. Commonly recognized categories include specific phobias, generalized anxiety disorder, social anxiety disorder, panic disorder, agoraphobia, obsessive–compulsive disorder and post-traumatic stress disorder.
Other affective (emotion/mood) processes can also become disordered. Mood disorder involving unusually intense and sustained sadness, melancholia, or despair is known as major depression (also known as unipolar or clinical depression). Milder, but still prolonged depression, can be diagnosed as dysthymia. Bipolar disorder (also known as manic depression) involves abnormally "high" or pressured mood states, known as mania or hypomania, alternating with normal or depressed moods. The extent to which unipolar and bipolar mood phenomena represent distinct categories of disorder, or mix and merge along a dimension or spectrum of mood, is subject to some scientific debate.
Patterns of belief, language use and perception of reality can become dysregulated (e.g., delusions, thought disorder, hallucinations). Psychotic disorders in this domain include schizophrenia, and delusional disorder. Schizoaffective disorder is a category used for individuals showing aspects of both schizophrenia and affective disorders. Schizotypy is a category used for individuals showing some of the characteristics associated with schizophrenia, but without meeting cutoff criteria.
Personality—the fundamental characteristics of a person that influence thoughts and behaviors across situations and time—may be considered disordered if judged to be abnormally rigid and maladaptive. Although treated separately by some, the commonly used categorical schemes include them as mental disorders, albeit on a separate axis II in the case of the DSM-IV. A number of different personality disorders are listed, including those sometimes classed as eccentric, such as paranoid, schizoid and schizotypal personality disorders; types that have described as dramatic or emotional, such as antisocial, borderline, histrionic or narcissistic personality disorders; and those sometimes classed as fear-related, such as anxious-avoidant, dependent, or obsessive–compulsive personality disorders. Personality disorders, in general, are defined as emerging in childhood, or at least by adolescence or early adulthood. The ICD also has a category for enduring personality change after a catastrophic experience or psychiatric illness. If an inability to sufficiently adjust to life circumstances begins within three months of a particular event or situation, and ends within six months after the stressor stops or is eliminated, it may instead be classed as an adjustment disorder. There is an emerging consensus that personality disorders, similar to personality traits in general, incorporate a mixture of acute dysfunctional behaviors that may resolve in short periods, and maladaptive temperamental traits that are more enduring. Furthermore, there are also non-categorical schemes that rate all individuals via a profile of different dimensions of personality without a symptom-based cutoff from normal personality variation, for example through schemes based on dimensional models.
An eating disorder is a serious mental health condition that involves an unhealthy relationship with food and body image. They can cause severe physical and psychological problems. Eating disorders involve disproportionate concern in matters of food and weight. Categories of disorder in this area include anorexia nervosa, bulimia nervosa, exercise bulimia or binge eating disorder.
Sleep disorders are associated with disruption to normal sleep patterns. A common sleep disorder is insomnia, which is described as difficulty falling and/or staying asleep. Other sleep disorders include narcolepsy, sleep apnea, REM sleep behavior disorder, chronic sleep deprivation, and restless leg syndrome.
Narcolepsy is a condition of extreme tendencies to fall asleep whenever and wherever. People with narcolepsy feel refreshed after their random sleep, but eventually get sleepy again. Narcolepsy diagnosis requires an overnight stay at a sleep center for analysis, during which doctors ask for a detailed sleep history and sleep records. Doctors also use actigraphs and polysomnography. Doctors will do a multiple sleep latency test, which measures how long it takes a person to fall asleep.
Sleep apnea, when breathing repeatedly stops and starts during sleep, can be a serious sleep disorder. Three types of sleep apnea include obstructive sleep apnea, central sleep apnea, and complex sleep apnea. Sleep apnea can be diagnosed at home or with polysomnography at a sleep center. An ear, nose, and throat doctor may further help with the sleeping habits.
Sexual disorders include dyspareunia and various kinds of paraphilia (sexual arousal to objects, situations, or individuals that are considered abnormal or harmful to the person or others).
Impulse control disorder: People who are abnormally unable to resist certain urges or impulses that could be harmful to themselves or others, may be classified as having an impulse control disorder, and disorders such as kleptomania (stealing) or pyromania (fire-setting). Various behavioral addictions, such as gambling addiction, may be classed as a disorder. Obsessive–compulsive disorder can sometimes involve an inability to resist certain acts but is classed separately as being primarily an anxiety disorder.
Substance use disorder: This disorder refers to the use of drugs (legal or illegal, including alcohol) that persists despite significant problems or harm related to its use. Substance dependence and substance abuse fall under this umbrella category in the DSM. Substance use disorder may be due to a pattern of compulsive and repetitive use of a drug that results in tolerance to its effects and withdrawal symptoms when use is reduced or stopped.
Dissociative disorder: People with severe disturbances of their self-identity, memory, and general awareness of themselves and their surroundings may be classified as having these types of disorders, including depersonalization disorder or dissociative identity disorder (which was previously referred to as multiple personality disorder or "split personality").
Cognitive disorder: These affect cognitive abilities, including learning and memory. This category includes delirium and mild and major neurocognitive disorder (previously termed dementia).
Developmental disorder: These disorders initially occur in childhood. Some examples include autism spectrum disorder, oppositional defiant disorder and conduct disorder, and attention deficit hyperactivity disorder (ADHD), which may continue into adulthood. Conduct disorder, if continuing into adulthood, may be diagnosed as antisocial personality disorder (dissocial personality disorder in the ICD). Popular labels such as psychopath (or sociopath) do not appear in the DSM or ICD but are linked by some to these diagnoses.
Somatoform disorders may be diagnosed when there are problems that appear to originate in the body that are thought to be manifestations of a mental disorder. This includes somatization disorder and conversion disorder. There are also disorders of how a person perceives their body, such as body dysmorphic disorder. Neurasthenia is an old diagnosis involving somatic complaints as well as fatigue and low spirits/depression, which is officially recognized by the ICD-10 but no longer by the DSM-IV.
Factitious disorders are diagnosed where symptoms are thought to be reported for personal gain. Symptoms are often deliberately produced or feigned, and may relate to either symptoms in the individual or in someone close to them, particularly people they care for.
There are attempts to introduce a category of relational disorder, where the diagnosis is of a relationship rather than on any one individual in that relationship. The relationship may be between children and their parents, between couples, or others. There already exists, under the category of psychosis, a diagnosis of shared psychotic disorder where two or more individuals share a particular delusion because of their close relationship with each other.
There are a number of uncommon psychiatric syndromes, which are often named after the person who first described them, such as Capgras syndrome, De Clerambault syndrome, Othello syndrome, Ganser syndrome, Cotard delusion, and Ekbom syndrome, and additional disorders such as the Couvade syndrome and Geschwind syndrome.
The onset of psychiatric disorders usually occurs from childhood to early adulthood. Impulse-control disorders and a few anxiety disorders tend to appear in childhood. Some other anxiety disorders, substance disorders, and mood disorders emerge later in the mid-teens. Symptoms of schizophrenia typically manifest from late adolescence to early twenties.
The likely course and outcome of mental disorders vary and are dependent on numerous factors related to the disorder itself, the individual as a whole, and the social environment. Some disorders may last a brief period of time, while others may be long-term in nature.
All disorders can have a varied course. Long-term international studies of schizophrenia have found that over a half of individuals recover in terms of symptoms, and around a fifth to a third in terms of symptoms and functioning, with many requiring no medication. While some have serious difficulties and support needs for many years, "late" recovery is still plausible. The World Health Organization (WHO) concluded that the long-term studies' findings converged with others in "relieving patients, carers and clinicians of the chronicity paradigm which dominated thinking throughout much of the 20th century."
A follow-up study by Tohen and coworkers revealed that around half of people initially diagnosed with bipolar disorder achieve symptomatic recovery (no longer meeting criteria for the diagnosis) within six weeks, and nearly all achieve it within two years, with nearly half regaining their prior occupational and residential status in that period. Less than half go on to experience a new episode of mania or major depression within the next two years.
Some disorders may be very limited in their functional effects, while others may involve substantial disability and support needs. In this context, the terms psychiatric disability and psychological disability are sometimes used instead of mental disorder. The degree of ability or disability may vary over time and across different life domains. Furthermore, psychiatric disability has been linked to institutionalization, discrimination and social exclusion as well as to the inherent effects of disorders. Alternatively, functioning may be affected by the stress of having to hide a condition in work or school, etc., by adverse effects of medications or other substances, or by mismatches between illness-related variations and demands for regularity.
It is also the case that, while often being characterized in purely negative terms, some mental traits or states labeled as psychiatric disabilities can also involve above-average creativity, non-conformity, goal-striving, meticulousness, or empathy. In addition, the public perception of the level of disability associated with mental disorders can change.
Nevertheless, internationally, people report equal or greater disability from commonly occurring mental conditions than from commonly occurring physical conditions, particularly in their social roles and personal relationships. The proportion with access to professional help for mental disorders is far lower, however, even among those assessed as having a severe psychiatric disability. Disability in this context may or may not involve such things as:
In terms of total disability-adjusted life years (DALYs), which is an estimate of how many years of life are lost due to premature death or to being in a state of poor health and disability, psychiatric disabilities rank amongst the most disabling conditions. Unipolar (also known as Major) depressive disorder is the third leading cause of disability worldwide, of any condition mental or physical, accounting for 65.5 million years lost. The first systematic description of global disability arising in youth, in 2011, found that among 10- to 24-year-olds nearly half of all disability (current and as estimated to continue) was due to psychiatric disabilities, including substance use disorders and conditions involving self-harm. Second to this were accidental injuries (mainly traffic collisions) accounting for 12 percent of disability, followed by communicable diseases at 10 percent. The psychiatric disabilities associated with most disabilities in high-income countries were unipolar major depression (20%) and alcohol use disorder (11%). In the eastern Mediterranean region, it was unipolar major depression (12%) and schizophrenia (7%), and in Africa it was unipolar major depression (7%) and bipolar disorder (5%).
Suicide, which is often attributed to some underlying mental disorder, is a leading cause of death among teenagers and adults under 35. There are an estimated 10 to 20 million non-fatal attempted suicides every year worldwide.
The predominant view as of 2018 is that genetic, psychological, and environmental factors all contribute to the development or progression of mental disorders. Different risk factors may be present at different ages, with risk occurring as early as during prenatal period.
A number of psychiatric disorders are linked to a family history (including depression, narcissistic personality disorder and anxiety). Twin studies have also revealed a very high heritability for many mental disorders (especially autism and schizophrenia). Although researchers have been looking for decades for clear linkages between genetics and mental disorders, that work has not yielded specific genetic biomarkers yet that might lead to better diagnosis and better treatments.
Statistical research looking at eleven disorders found widespread assortative mating between people with mental illness. That means that individuals with one of these disorders were two to three times more likely than the general population to have a partner with a mental disorder. Sometimes people seemed to have preferred partners with the same mental illness. Thus, people with schizophrenia or ADHD are seven times more likely to have affected partners with the same disorder. This is even more pronounced for people with Autism spectrum disorders who are 10 times more likely to have a spouse with the same disorder.
During the prenatal stage, factors like unwanted pregnancy, lack of adaptation to pregnancy or substance use during pregnancy increases the risk of developing a mental disorder. Maternal stress and birth complications including prematurity and infections have also been implicated in increasing susceptibility for mental illness. Infants neglected or not provided optimal nutrition have a higher risk of developing cognitive impairment.
Social influences have also been found to be important, including abuse, neglect, bullying, social stress, traumatic events, and other negative or overwhelming life experiences. Aspects of the wider community have also been implicated, including employment problems, socioeconomic inequality, lack of social cohesion, problems linked to migration, and features of particular societies and cultures. The specific risks and pathways to particular disorders are less clear, however.
Nutrition also plays a role in mental disorders.
In schizophrenia and psychosis, risk factors include migration and discrimination, childhood trauma, bereavement or separation in families, recreational use of drugs, and urbanicity.
In anxiety, risk factors may include parenting factors including parental rejection, lack of parental warmth, high hostility, harsh discipline, high maternal negative affect, anxious childrearing, modelling of dysfunctional and drug-abusing behavior, and child abuse (emotional, physical and sexual). Adults with imbalance work to life are at higher risk for developing anxiety.
For bipolar disorder, stress (such as childhood adversity) is not a specific cause, but does place genetically and biologically vulnerable individuals at risk for a more severe course of illness.
Mental disorders are associated with drug use including: cannabis, alcohol and caffeine, use of which appears to promote anxiety. For psychosis and schizophrenia, usage of a number of drugs has been associated with development of the disorder, including cannabis, cocaine, and amphetamines. There has been debate regarding the relationship between usage of cannabis and bipolar disorder. Cannabis has also been associated with depression. Adolescents are at increased risk for tobacco, alcohol and drug use; Peer pressure is the main reason why adolescents start using substances. At this age, the use of substances could be detrimental to the development of the brain and place them at higher risk of developing a mental disorder.
People living with chronic conditions like HIV and diabetes are at higher risk of developing a mental disorder. People living with diabetes experience significant stress from the biological impact of the disease, which places them at risk for developing anxiety and depression. Diabetic patients also have to deal with emotional stress trying to manage the disease. Conditions like heart disease, stroke, respiratory conditions, cancer, and arthritis increase the risk of developing a mental disorder when compared to the general population.
Risk factors for mental illness include a propensity for high neuroticism or "emotional instability". In anxiety, risk factors may include temperament and attitudes (e.g. pessimism).
Mental disorders can arise from multiple sources, and in many cases there is no single accepted or consistent cause currently established. An eclectic or pluralistic mix of models may be used to explain particular disorders. The primary paradigm of contemporary mainstream Western psychiatry is said to be the biopsychosocial model which incorporates biological, psychological and social factors, although this may not always be applied in practice.
Biological psychiatry follows a biomedical model where many mental disorders are conceptualized as disorders of brain circuits likely caused by developmental processes shaped by a complex interplay of genetics and experience. A common assumption is that disorders may have resulted from genetic and developmental vulnerabilities, exposed by stress in life (for example in a diathesis–stress model), although there are various views on what causes differences between individuals. Some types of mental disorders may be viewed as primarily neurodevelopmental disorders.
Evolutionary psychology may be used as an overall explanatory theory, while attachment theory is another kind of evolutionary-psychological approach sometimes applied in the context of mental disorders. Psychoanalytic theories have continued to evolve alongside and cognitive-behavioral and systemic-family approaches. A distinction is sometimes made between a "medical model" or a "social model" of psychiatric disability.
Psychiatrists seek to provide a medical diagnosis of individuals by an assessment of symptoms, signs and impairment associated with particular types of mental disorder. Other mental health professionals, such as clinical psychologists, may or may not apply the same diagnostic categories to their clinical formulation of a client's difficulties and circumstances. The majority of mental health problems are, at least initially, assessed and treated by family physicians (in the UK general practitioners) during consultations, who may refer a patient on for more specialist diagnosis in acute or chronic cases.
Routine diagnostic practice in mental health services typically involves an interview known as a mental status examination, where evaluations are made of appearance and behavior, self-reported symptoms, mental health history, and current life circumstances. The views of other professionals, relatives, or other third parties may be taken into account. A physical examination to check for ill health or the effects of medications or other drugs may be conducted. Psychological testing is sometimes used via paper-and-pen or computerized questionnaires, which may include algorithms based on ticking off standardized diagnostic criteria, and in rare specialist cases neuroimaging tests may be requested, but such methods are more commonly found in research studies than routine clinical practice.
Time and budgetary constraints often limit practicing psychiatrists from conducting more thorough diagnostic evaluations. It has been found that most clinicians evaluate patients using an unstructured, open-ended approach, with limited training in evidence-based assessment methods, and that inaccurate diagnosis may be common in routine practice. In addition, comorbidity is very common in psychiatric diagnosis, where the same person meets the criteria for more than one disorder. On the other hand, a person may have several different difficulties only some of which meet the criteria for being diagnosed. There may be specific problems with accurate diagnosis in developing countries.
More structured approaches are being increasingly used to measure levels of mental illness.
Since the 1980s, Paula Caplan has been concerned about the subjectivity of psychiatric diagnosis, and people being arbitrarily "slapped with a psychiatric label." Caplan says because psychiatric diagnosis is unregulated, doctors are not required to spend much time interviewing patients or to seek a second opinion. The Diagnostic and Statistical Manual of Mental Disorders can lead a psychiatrist to focus on narrow checklists of symptoms, with little consideration of what is actually causing the person's problems. So, according to Caplan, getting a psychiatric diagnosis and label often stands in the way of recovery.
In 2013, psychiatrist Allen Frances wrote a paper entitled "The New Crisis of Confidence in Psychiatric Diagnosis", which said that "psychiatric diagnosis... still relies exclusively on fallible subjective judgments rather than objective biological tests." Frances was also concerned about "unpredictable overdiagnosis." For many years, marginalized psychiatrists (such as Peter Breggin, Thomas Szasz) and outside critics (such as Stuart A. Kirk) have "been accusing psychiatry of engaging in the systematic medicalization of normality." More recently these concerns have come from insiders who have worked for and promoted the American Psychiatric Association (e.g., Robert Spitzer, Allen Frances). A 2002 editorial in the British Medical Journal warned of inappropriate medicalization leading to disease mongering, where the boundaries of the definition of illnesses are expanded to include personal problems as medical problems or risks of diseases are emphasized to broaden the market for medications.
Gary Greenberg, a psychoanalyst, in his book "the Book of Woe", argues that mental illness is really about suffering and how the DSM creates diagnostic labels to categorize people's suffering. Indeed, the psychiatrist Thomas Szasz, in his book "the Medicalization of Everyday Life", also argues that what is psychiatric illness, is not always biological in nature (i.e. social problems, poverty, etc.), and may even be a part of the human condition.
in 2018 the American Psychological Association commissioned a review to reach a consensus on whether modern clinical MRI/fMRI will be able to be used in the diagnosis of mental health disorders. The criteria presented by the APA stated that the biomarkers used in diagnosis should:
The review concluded that although neuroimaging diagnosis may technically be feasible, very large studies are needed to evaluate specific biomarkers which were not available.
The 2004 WHO report "Prevention of Mental Disorders" stated that "Prevention of these disorders is obviously one of the most effective ways to reduce the [disease] burden."
The 2011 European Psychiatric Association (EPA) guidance on prevention of mental disorders states "There is considerable evidence that various psychiatric conditions can be prevented through the implementation of effective evidence-based interventions."
A 2011 UK Department of Health report on the economic case for mental health promotion and mental illness prevention found that "many interventions are outstandingly good value for money, low in cost and often become self-financing over time, saving public expenditure".
In 2016, the National Institute of Mental Health re-affirmed prevention as a research priority area.
Parenting may affect the child's mental health, and evidence suggests that helping parents to be more effective with their children can address mental health needs.
Universal prevention (aimed at a population that has no increased risk for developing a mental disorder, such as school programs or mass media campaigns) need very high numbers of people to show effect (sometimes known as the "power" problem). Approaches to overcome this are (1) focus on high-incidence groups (e.g. by targeting groups with high risk factors), (2) use multiple interventions to achieve greater, and thus more statistically valid, effects, (3) use cumulative meta-analyses of many trials, and (4) run very large trials.
Treatment and support for mental disorders are provided in psychiatric hospitals, clinics or a range of community mental health services. In some countries services are increasingly based on a recovery approach, intended to support individual's personal journey to gain the kind of life they want.
There is a range of different types of treatment and what is most suitable depends on the disorder and the individual. Many things have been found to help at least some people, and a placebo effect may play a role in any intervention or medication. In a minority of cases, individuals may be treated against their will, which can cause particular difficulties depending on how it is carried out and perceived. Compulsory treatment while in the community versus non-compulsory treatment does not appear to make much of a difference except by maybe decreasing victimization.
Lifestyle strategies, including dietary changes, exercise and quitting smoking may be of benefit.
There is also a wide range of psychotherapists (including family therapy), counselors, and public health professionals. In addition, there are peer support roles where personal experience of similar issues is the primary source of expertise.
A major option for many mental disorders is psychotherapy. There are several main types. Cognitive behavioral therapy (CBT) is widely used and is based on modifying the patterns of thought and behavior associated with a particular disorder. Other psychotherapies include dialectic behavioral therapy (DBT) and interpersonal psychotherapy (IPT). Psychoanalysis, addressing underlying psychic conflicts and defenses, has been a dominant school of psychotherapy and is still in use. Systemic therapy or family therapy is sometimes used, addressing a network of significant others as well as an individual.
Some psychotherapies are based on a humanistic approach. There are many specific therapies used for particular disorders, which may be offshoots or hybrids of the above types. Mental health professionals often employ an eclectic or integrative approach. Much may depend on the therapeutic relationship, and there may be problems with trust, confidentiality and engagement.
A major option for many mental disorders is psychiatric medication and there are several main groups. Antidepressants are used for the treatment of clinical depression, as well as often for anxiety and a range of other disorders. Anxiolytics (including sedatives) are used for anxiety disorders and related problems such as insomnia. Mood stabilizers are used primarily in bipolar disorder. Antipsychotics are used for psychotic disorders, notably for positive symptoms in schizophrenia, and also increasingly for a range of other disorders. Stimulants are commonly used, notably for ADHD.
Despite the different conventional names of the drug groups, there may be considerable overlap in the disorders for which they are actually indicated, and there may also be off-label use of medications. There can be problems with adverse effects of medication and adherence to them, and there is also criticism of pharmaceutical marketing and professional conflicts of interest. However, these medications in combination with non-pharmacological methods, such as cognitive-behavioral therapy (CBT) are seen to be most effective in treating mental disorders.
Electroconvulsive therapy (ECT) is sometimes used in severe cases when other interventions for severe intractable depression have failed. ECT is usually indicated for treatment resistant depression, severe vegetative symptoms, psychotic depression, intense suicidal ideation, depression during pregnancy, and catatonia. Psychosurgery is considered experimental but is advocated by some neurologists in certain rare cases.
Counseling (professional) and co-counseling (between peers) may be used. Psychoeducation programs may provide people with the information to understand and manage their problems. Creative therapies are sometimes used, including music therapy, art therapy or drama therapy. Lifestyle adjustments and supportive measures are often used, including peer support, self-help groups for mental health and supported housing or supported employment (including social firms). Some advocate dietary supplements.
Reasonable accommodations (adjustments and supports) might be put in place to help an individual cope and succeed in environments despite potential disability related to mental health problems. This could include an emotional support animal or specifically trained psychiatric service dog. As of 2019 cannabis is specifically not recommended as a treatment.
Mental disorders are common. Worldwide, more than one in three people in most countries report sufficient criteria for at least one at some point in their life. In the United States, 46% qualify for a mental illness at some point. An ongoing survey indicates that anxiety disorders are the most common in all but one country, followed by mood disorders in all but two countries, while substance disorders and impulse-control disorders were consistently less prevalent. Rates varied by region.
A review of anxiety disorder surveys in different countries found average lifetime prevalence estimates of 16.6%, with women having higher rates on average. A review of mood disorder surveys in different countries found lifetime rates of 6.7% for major depressive disorder (higher in some studies, and in women) and 0.8% for Bipolar I disorder.
In the United States the frequency of disorder is: anxiety disorder (28.8%), mood disorder (20.8%), impulse-control disorder (24.8%) or substance use disorder (14.6%).
A 2004 cross-Europe study found that approximately one in four people reported meeting criteria at some point in their life for at least one of the DSM-IV disorders assessed, which included mood disorders (13.9%), anxiety disorders (13.6%), or alcohol disorder (5.2%). Approximately one in ten met the criteria within a 12-month period. Women and younger people of either gender showed more cases of the disorder. A 2005 review of surveys in 16 European countries found that 27% of adult Europeans are affected by at least one mental disorder in a 12-month period.
An international review of studies on the prevalence of schizophrenia found an average (median) figure of 0.4% for lifetime prevalence; it was consistently lower in poorer countries.
Studies of the prevalence of personality disorders (PDs) have been fewer and smaller-scale, but one broad Norwegian survey found a five-year prevalence of almost 1 in 7 (13.4%). Rates for specific disorders ranged from 0.8% to 2.8%, differing across countries, and by gender, educational level and other factors. A US survey that incidentally screened for personality disorder found a rate of 14.79%.
Approximately 7% of a preschool pediatric sample were given a psychiatric diagnosis in one clinical study, and approximately 10% of 1- and 2-year-olds receiving developmental screening have been assessed as having significant emotional/behavioral problems based on parent and pediatrician reports.
While rates of psychological disorders are often the same for men and women, women tend to have a higher rate of depression. Each year 73 million women are affected by major depression, and suicide is ranked 7th as the cause of death for women between the ages of 20–59. Depressive disorders account for close to 41.9% of the psychiatric disabilities among women compared to 29.3% among men.
Ancient civilizations described and treated a number of mental disorders. Mental illnesses were well known in ancient Mesopotamia, where diseases and mental disorders were believed to be caused by specific deities. Because hands symbolized control over a person, mental illnesses were known as "hands" of certain deities. One psychological illness was known as Qāt Ištar, meaning "Hand of Ishtar". Others were known as "Hand of Shamash", "Hand of the Ghost", and "Hand of the God". Descriptions of these illnesses, however, are so vague that it is usually impossible to determine which illnesses they correspond to in modern terminology. Mesopotamian doctors kept detailed record of their patients' hallucinations and assigned spiritual meanings to them. The royal family of Elam was notorious for its members often being insane. The Greeks coined terms for melancholy, hysteria and phobia and developed the humorism theory. Mental disorders were described, and treatments developed, in Persia, Arabia and in the medieval Islamic world.
Conceptions of madness in the Middle Ages in Christian Europe were a mixture of the divine, diabolical, magical and humoral, and transcendental. In the early modern period, some people with mental disorders may have been victims of the witch-hunts. While not every witch and sorcerer accused were mentally ill, all mentally ill were considered to be witches or sorcerers. Many terms for mental disorders that found their way into everyday use first became popular in the 16th and 17th centuries.
By the end of the 17th century and into the Enlightenment, madness was increasingly seen as an organic physical phenomenon with no connection to the soul or moral responsibility. Asylum care was often harsh and treated people like wild animals, but towards the end of the 18th century a moral treatment movement gradually developed. Clear descriptions of some syndromes may be rare before the 19th century.
Industrialization and population growth led to a massive expansion of the number and size of insane asylums in every Western country in the 19th century. Numerous different classification schemes and diagnostic terms were developed by different authorities, and the term psychiatry was coined (1808), though medical superintendents were still known as alienists.
The turn of the 20th century saw the development of psychoanalysis, which would later come to the fore, along with Kraepelin's classification scheme. Asylum "inmates" were increasingly referred to as "patients", and asylums were renamed as hospitals.
Early in the 20th century in the United States, a mental hygiene movement developed, aiming to prevent mental disorders. Clinical psychology and social work developed as professions. World War I saw a massive increase of conditions that came to be termed "shell shock".
World War II saw the development in the U.S. of a new psychiatric manual for categorizing mental disorders, which along with existing systems for collecting census and hospital statistics led to the first Diagnostic and Statistical Manual of Mental Disorders. The International Classification of Diseases (ICD) also developed a section on mental disorders. The term stress, having emerged from endocrinology work in the 1930s, was increasingly applied to mental disorders.
Electroconvulsive therapy, insulin shock therapy, lobotomies and the neuroleptic chlorpromazine came to be used by mid-century. In the 1960s there were many challenges to the concept of mental illness itself. These challenges came from psychiatrists like Thomas Szasz who argued that mental illness was a myth used to disguise moral conflicts; from sociologists such as Erving Goffman who said that mental illness was merely another example of how society labels and controls non-conformists; from behavioral psychologists who challenged psychiatry's fundamental reliance on unobservable phenomena; and from gay rights activists who criticised the APA's listing of homosexuality as a mental disorder. A study published in Science by Rosenhan received much publicity and was viewed as an attack on the efficacy of psychiatric diagnosis.
Deinstitutionalization gradually occurred in the West, with isolated psychiatric hospitals being closed down in favor of community mental health services. A consumer/survivor movement gained momentum. Other kinds of psychiatric medication gradually came into use, such as "psychic energizers" (later antidepressants) and lithium. Benzodiazepines gained widespread use in the 1970s for anxiety and depression, until dependency problems curtailed their popularity.
Advances in neuroscience, genetics, and psychology led to new research agendas. Cognitive behavioral therapy and other psychotherapies developed. The DSM and then ICD adopted new criteria-based classifications, and the number of "official" diagnoses saw a large expansion. Through the 1990s, new SSRI-type antidepressants became some of the most widely prescribed drugs in the world, as later did antipsychotics. Also during the 1990s, a recovery approach developed.
Most Africans view mental disturbances as external spiritual attack on the person. Those who have a mental illness are thought to be under a spell or bewitched. Often than usual, People view a mentally ill person as possessed of an evil spirit and is seen as more of sociological perspective than a psychological order.
The WHO estimated that fewer than 10% of mentally ill Nigerians have access to a psychiatrist or health worker, because there is a low ratio of mental-health specialists available in a country of 200 million people. WHO estimates that the number of mentally ill Nigerians ranges from 40 million to 60 million. Disorders such as depression, anxiety, schizophrenia, personality disorder, old age-related disorder, and substance-abuse disorder are common in Nigeria, as in other countries in Africa.
Nigeria is still nowhere near being equipped to solve prevailing mental health challenges. With little scientific research carried out, coupled with insufficient mental-health hospitals in the country, traditional healers provide specialized psychotherapy care to those that require their services and pharmacotherapy
Different societies or cultures, even different individuals in a subculture, can disagree as to what constitutes optimal versus pathological biological and psychological functioning. Research has demonstrated that cultures vary in the relative importance placed on, for example, happiness, autonomy, or social relationships for pleasure. Likewise, the fact that a behavior pattern is valued, accepted, encouraged, or even statistically normative in a culture does not necessarily mean that it is conducive to optimal psychological functioning.
People in all cultures find some behaviors bizarre or even incomprehensible. But just what they feel is bizarre or incomprehensible is ambiguous and subjective. These differences in determination can become highly contentious. The process by which conditions and difficulties come to be defined and treated as medical conditions and problems, and thus come under the authority of doctors and other health professionals, is known as medicalization or pathologization.
There is a perception in Latin American communities, especially among older people, that discussing problems with mental health can create embarrassment and shame for the family. This results in fewer people seeking treatment.
Latin Americans from the US are slightly more likely to have a mental health disorder than first-generation Latin American immigrants, although differences between ethnic groups were found to disappear after adjustment for place of birth.
From 2015 to 2018, rates of serious mental illness in young adult Latin Americans increased by 60%, from 4% to 6.4%. The prevalence of major depressive episodes in young and adult Latin Americans increased from 8.4% to 11.3%. More than a third of Latin Americans reported more than one bad mental health day in the last three months. The rate of suicide among Latin Americans was about half the rate of non-Latin American white Americans in 2018, and this was the second-leading cause of death among Latin Americans ages 15 to 34. However, Latin American suicide rates rose steadily after 2020 in relation to the COVID-19 pandemic, even as the national rate declined.
Family relations are an integral part of the Latin American community. Some research has shown that Latin Americans are more likely rely on family bonds, or familismo, as a source of therapy while struggling with mental health issues. Because Latin Americans have a high rate of religiosity, and because there is less stigma associated with religion than with psychiatric services, religion may play a more important therapeutic role for the mentally ill in Latin American communities. However, research has also suggested that religion may also play a role in stigmatizing mental illness in Latin American communities, which can discourage community members from seeking professional help.
Religious, spiritual, or transpersonal experiences and beliefs meet many criteria of delusional or psychotic disorders. A belief or experience can sometimes be shown to produce distress or disability—the ordinary standard for judging mental disorders. There is a link between religion and schizophrenia, a complex mental disorder characterized by a difficulty in recognizing reality, regulating emotional responses, and thinking in a clear and logical manner. Those with schizophrenia commonly report some type of religious delusion, and religion itself may be a trigger for schizophrenia.
Controversy has often surrounded psychiatry, and the term anti-psychiatry was coined by the psychiatrist David Cooper in 1967. The anti-psychiatry message is that psychiatric treatments are ultimately more damaging than helpful to patients, and psychiatry's history involves what may now be seen as dangerous treatments. Electroconvulsive therapy was one of these, which was used widely between the 1930s and 1960s. Lobotomy was another practice that was ultimately seen as too invasive and brutal. Diazepam and other sedatives were sometimes over-prescribed, which led to an epidemic of dependence. There was also concern about the large increase in prescribing psychiatric drugs for children. Some charismatic psychiatrists came to personify the movement against psychiatry. The most influential of these was R.D. Laing who wrote a series of best-selling books, including The Divided Self. Thomas Szasz wrote The Myth of Mental Illness. Some ex-patient groups have become militantly anti-psychiatric, often referring to themselves as survivors. Giorgio Antonucci has questioned the basis of psychiatry through his work on the dismantling of two psychiatric hospitals (in the city of Imola), carried out from 1973 to 1996.
The consumer/survivor movement (also known as user/survivor movement) is made up of individuals (and organizations representing them) who are clients of mental health services or who consider themselves survivors of psychiatric interventions. Activists campaign for improved mental health services and for more involvement and empowerment within mental health services, policies and wider society. Patient advocacy organizations have expanded with increasing deinstitutionalization in developed countries, working to challenge the stereotypes, stigma and exclusion associated with psychiatric conditions. There is also a carers rights movement of people who help and support people with mental health conditions, who may be relatives, and who often work in difficult and time-consuming circumstances with little acknowledgement and without pay. An anti-psychiatry movement fundamentally challenges mainstream psychiatric theory and practice, including in some cases asserting that psychiatric concepts and diagnoses of 'mental illness' are neither real nor useful.
Alternatively, a movement for global mental health has emerged, defined as 'the area of study, research and practice that places a priority on improving mental health and achieving equity in mental health for all people worldwide'.
Diagnostic guidelines of the 2000s, namely the DSM and to some extent the ICD, have been criticized as having a fundamentally Euro-American outlook. Opponents argue that even when diagnostic criteria are used across different cultures, it does not mean that the underlying constructs have validity within those cultures, as even reliable application can prove only consistency, not legitimacy. Advocating a more culturally sensitive approach, critics such as Carl Bell and Marcello Maviglia contend that the cultural and ethnic diversity of individuals is often discounted by researchers and service providers.
Cross-cultural psychiatrist Arthur Kleinman contends that the Western bias is ironically illustrated in the introduction of cultural factors to the DSM-IV. Disorders or concepts from non-Western or non-mainstream cultures are described as "culture-bound", whereas standard psychiatric diagnoses are given no cultural qualification whatsoever, revealing to Kleinman an underlying assumption that Western cultural phenomena are universal. Kleinman's negative view towards the culture-bound syndrome is largely shared by other cross-cultural critics. Common responses included both disappointment over the large number of documented non-Western mental disorders still left out and frustration that even those included are often misinterpreted or misrepresented.
Many mainstream psychiatrists are dissatisfied with the new culture-bound diagnoses, although for partly different reasons. Robert Spitzer, a lead architect of the DSM-III, has argued that adding cultural formulations was an attempt to appease cultural critics, and has stated that they lack any scientific rationale or support. Spitzer also posits that the new culture-bound diagnoses are rarely used, maintaining that the standard diagnoses apply regardless of the culture involved. In general, mainstream psychiatric opinion remains that if a diagnostic category is valid, cross-cultural factors are either irrelevant or are significant only to specific symptom presentations.
Clinical conceptions of mental illness also overlap with personal and cultural values in the domain of morality, so much so that it is sometimes argued that separating the two is impossible without fundamentally redefining the essence of being a particular person in a society. In clinical psychiatry, persistent distress and disability indicate an internal disorder requiring treatment; but in another context, that same distress and disability can be seen as an indicator of emotional struggle and the need to address social and structural problems. This dichotomy has led some academics and clinicians to advocate a postmodernist conceptualization of mental distress and well-being.
Such approaches, along with cross-cultural and "heretical" psychologies centered on alternative cultural and ethnic and race-based identities and experiences, stand in contrast to the mainstream psychiatric community's alleged avoidance of any explicit involvement with either morality or culture. In many countries there are attempts to challenge perceived prejudice against minority groups, including alleged institutional racism within psychiatric services. There are also ongoing attempts to improve professional cross cultural sensitivity.
Three-quarters of countries around the world have mental health legislation. Compulsory admission to mental health facilities (also known as involuntary commitment) is a controversial topic. It can impinge on personal liberty and the right to choose, and carry the risk of abuse for political, social, and other reasons; yet it can potentially prevent harm to self and others, and assist some people in attaining their right to healthcare when they may be unable to decide in their own interests. Because of this it is a concern of medical ethics.
All human rights oriented mental health laws require proof of the presence of a mental disorder as defined by internationally accepted standards, but the type and severity of disorder that counts can vary in different jurisdictions. The two most often used grounds for involuntary admission are said to be serious likelihood of immediate or imminent danger to self or others, and the need for treatment. Applications for someone to be involuntarily admitted usually come from a mental health practitioner, a family member, a close relative, or a guardian. Human-rights-oriented laws usually stipulate that independent medical practitioners or other accredited mental health practitioners must examine the patient separately and that there should be regular, time-bound review by an independent review body. The individual should also have personal access to independent advocacy.
For involuntary treatment to be administered (by force if necessary), it should be shown that an individual lacks the mental capacity for informed consent (i.e. to understand treatment information and its implications, and therefore be able to make an informed choice to either accept or refuse). Legal challenges in some areas have resulted in supreme court decisions that a person does not have to agree with a psychiatrist's characterization of the issues as constituting an "illness", nor agree with a psychiatrist's conviction in medication, but only recognize the issues and the information about treatment options.
Proxy consent (also known as surrogate or substituted decision-making) may be transferred to a personal representative, a family member, or a legally appointed guardian. Moreover, patients may be able to make, when they are considered well, an advance directive stipulating how they wish to be treated should they be deemed to lack mental capacity in the future. The right to supported decision-making, where a person is helped to understand and choose treatment options before they can be declared to lack capacity, may also be included in the legislation. There should at the very least be shared decision-making as far as possible. Involuntary treatment laws are increasingly extended to those living in the community, for example outpatient commitment laws (known by different names) are used in New Zealand, Australia, the United Kingdom, and most of the United States.
The World Health Organization reports that in many instances national mental health legislation takes away the rights of persons with mental disorders rather than protecting rights, and is often outdated. In 1991, the United Nations adopted the Principles for the Protection of Persons with Mental Illness and the Improvement of Mental Health Care, which established minimum human rights standards of practice in the mental health field. In 2006, the UN formally agreed the Convention on the Rights of Persons with Disabilities to protect and enhance the rights and opportunities of disabled people, including those with psychiatric disabilities.
The term insanity, sometimes used colloquially as a synonym for mental illness, is often used technically as a legal term. The insanity defense may be used in a legal trial (known as the mental disorder defence in some countries).
The social stigma associated with mental disorders is a widespread problem. The US Surgeon General stated in 1999 that: "Powerful and pervasive, stigma prevents people from acknowledging their own mental health problems, much less disclosing them to others." Additionally, researcher Wulf Rössler in 2016, in his article, "The Stigma of Mental Disorders" stated
In the United States, racial and ethnic minorities are more likely to experience mental health disorders often due to low socioeconomic status, and discrimination. In Taiwan, those with mental disorders are subject to general public's misperception that the root causes of the mental disorders are "over-thinking", "having a lot of time and nothing better to do", "stagnant", "not serious in life", "not paying enough attention to the real life affairs", "mentally weak", "refusing to be resilient", "turning back to perfectionistic strivings", "not bravery" and so forth.
Employment discrimination is reported to play a significant part in the high rate of unemployment among those with a diagnosis of mental illness. An Australian study found that having a psychiatric disability is a bigger barrier to employment than a physical disability. The mentally ill are stigmatized in Chinese society and can not legally marry.
Efforts are being undertaken worldwide to eliminate the stigma of mental illness, although the methods and outcomes used have sometimes been criticized.
Media coverage of mental illness comprises predominantly negative and pejorative depictions, for example, of incompetence, violence or criminality, with far less coverage of positive issues such as accomplishments or human rights issues. Such negative depictions, including in children's cartoons, are thought to contribute to stigma and negative attitudes in the public and in those with mental health problems themselves, although more sensitive or serious cinematic portrayals have increased in prevalence.
In the United States, the Carter Center has created fellowships for journalists in South Africa, the U.S., and Romania, to enable reporters to research and write stories on mental health topics. Former US First Lady Rosalynn Carter began the fellowships not only to train reporters in how to sensitively and accurately discuss mental health and mental illness, but also to increase the number of stories on these topics in the news media. There is also a World Mental Health Day, which in the United States and Canada falls within a Mental Illness Awareness Week.
The general public have been found to hold a strong stereotype of dangerousness and desire for social distance from individuals described as mentally ill. A US national survey found that a higher percentage of people rate individuals described as displaying the characteristics of a mental disorder as "likely to do something violent to others", compared to the percentage of people who are rating individuals described as being troubled. In the article, "Discrimination Against People with a Mental Health Diagnosis: Qualitative Analysis of Reported Experiences," an individual who has a mental disorder, revealed that, "If people don't know me and don't know about the problems, they'll talk to me quite happily. Once they've seen the problems or someone's told them about me, they tend to be a bit more wary." In addition, in the article,"Stigma and its Impact on Help-Seeking for Mental Disorders: What Do We Know?" by George Schomerus and Matthias Angermeyer, it is affirmed that "Family doctors and psychiatrists have more pessimistic views about the outcomes for mental illnesses than the general public (Jorm et al.,1999), and mental health professionals hold more negative stereotypes about mentally ill patients, but, reassuringly, they are less accepting of restrictions towards them."
Recent depictions in media have included leading characters successfully living with and managing a mental illness, including in bipolar disorder in Homeland (2011) and post-traumatic stress disorder in Iron Man 3 (2013).
Despite public or media opinion, national studies have indicated that severe mental illness does not independently predict future violent behavior, on average, and is not a leading cause of violence in society. There is a statistical association with various factors that do relate to violence (in anyone), such as substance use and various personal, social, and economic factors. A 2015 review found that in the United States, about 4% of violence is attributable to people diagnosed with mental illness, and a 2014 study found that 7.5% of crimes committed by mentally ill people were directly related to the symptoms of their mental illness. The majority of people with serious mental illness are never violent.
In fact, findings consistently indicate that it is many times more likely that people diagnosed with a serious mental illness living in the community will be the victims rather than the perpetrators of violence. In a study of individuals diagnosed with "severe mental illness" living in a US inner-city area, a quarter were found to have been victims of at least one violent crime over the course of a year, a proportion eleven times higher than the inner-city average, and higher in every category of crime including violent assaults and theft. People with a diagnosis may find it more difficult to secure prosecutions, however, due in part to prejudice and being seen as less credible.
However, there are some specific diagnoses, such as childhood conduct disorder or adult antisocial personality disorder or psychopathy, which are defined by, or are inherently associated with, conduct problems and violence. There are conflicting findings about the extent to which certain specific symptoms, notably some kinds of psychosis (hallucinations or delusions) that can occur in disorders such as schizophrenia, delusional disorder or mood disorder, are linked to an increased risk of serious violence on average. The mediating factors of violent acts, however, are most consistently found to be mainly socio-demographic and socio-economic factors such as being young, male, of lower socioeconomic status and, in particular, substance use (including alcohol use) to which some people may be particularly vulnerable.
High-profile cases have led to fears that serious crimes, such as homicide, have increased due to deinstitutionalization, but the evidence does not support this conclusion. Violence that does occur in relation to mental disorder (against the mentally ill or by the mentally ill) typically occurs in the context of complex social interactions, often in a family setting rather than between strangers. It is also an issue in health care settings and the wider community.
The recognition and understanding of mental health conditions have changed over time and across cultures and there are still variations in definition, assessment, and classification, although standard guideline criteria are widely used. In many cases, there appears to be a continuum between mental health and mental illness, making diagnosis complex. According to the World Health Organization, over a third of people in most countries report problems at some time in their life which meet the criteria for diagnosis of one or more of the common types of mental disorder. Corey M Keyes has created a two continua model of mental illness and health which holds that both are related, but distinct dimensions: one continuum indicates the presence or absence of mental health, the other the presence or absence of mental illness. For example, people with optimal mental health can also have a mental illness, and people who have no mental illness can also have poor mental health.
Psychopathology in non-human primates has been studied since the mid-20th century. Over 20 behavioral patterns in captive chimpanzees have been documented as (statistically) abnormal for frequency, severity or oddness—some of which have also been observed in the wild. Captive great apes show gross behavioral abnormalities such as stereotypy of movements, self-mutilation, disturbed emotional reactions (mainly fear or aggression) towards companions, lack of species-typical communications, and generalized learned helplessness. In some cases such behaviors are hypothesized to be equivalent to symptoms associated with psychiatric disorders in humans such as depression, anxiety disorders, eating disorders and post-traumatic stress disorder. Concepts of antisocial, borderline and schizoid personality disorders have also been applied to non-human great apes.
The risk of anthropomorphism is often raised concerning such comparisons, and assessment of non-human animals cannot incorporate evidence from linguistic communication. However, available evidence may range from nonverbal behaviors—including physiological responses and homologous facial displays and acoustic utterances—to neurochemical studies. It is pointed out that human psychiatric classification is often based on statistical description and judgment of behaviors (especially when speech or language is impaired) and that the use of verbal self-report is itself problematic and unreliable.
Psychopathology has generally been traced, at least in captivity, to adverse rearing conditions such as early separation of infants from mothers; early sensory deprivation; and extended periods of social isolation. Studies have also indicated individual variation in temperament, such as sociability or impulsiveness. Particular causes of problems in captivity have included integration of strangers into existing groups and a lack of individual space, in which context some pathological behaviors have also been seen as coping mechanisms. Remedial interventions have included careful individually tailored re-socialization programs, behavior therapy, environment enrichment, and on rare occasions psychiatric drugs. Socialization has been found to work 90% of the time in disturbed chimpanzees, although restoration of functional sexuality and caregiving is often not achieved.
Laboratory researchers sometimes try to develop animal models of human mental disorders, including by inducing or treating symptoms in animals through genetic, neurological, chemical or behavioral manipulation, but this has been criticized on empirical grounds and opposed on animal rights grounds.
Definition
"Nervous breakdown" redirects here. For other uses, see Nervous breakdown (disambiguation).
The definition and classification of mental disorders are key issues for researchers as well as service providers and those who may be diagnosed. For a mental state to be classified as a disorder, it generally needs to cause dysfunction. Most international clinical documents use the term mental "disorder", while "illness" is also common. It has been noted that using the term "mental" (i.e., of the mind) is not necessarily meant to imply separateness from the brain or body.
According to the fourth edition of the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV), published in 1994, a mental disorder is a psychological syndrome or pattern that is associated with distress (e.g., via a painful symptom), disability (impairment in one or more important areas of functioning), increased risk of death, or causes a significant loss of autonomy; however, it excludes normal responses such as the grief from loss of a loved one and also excludes deviant behavior for political, religious, or societal reasons not arising from a dysfunction in the individual.
DSM-IV predicates the definition with caveats, stating that, as in the case with many medical terms, mental disorder "lacks a consistent operational definition that covers all situations", noting that different levels of abstraction can be used for medical definitions, including pathology, symptomology, deviance from a normal range, or etiology, and that the same is true for mental disorders, so that sometimes one type of definition is appropriate and sometimes another, depending on the situation.
In 2013, the American Psychiatric Association (APA) redefined mental disorders in the DSM-5 as "a syndrome characterized by clinically significant disturbance in an individual's cognition, emotion regulation, or behavior that reflects a dysfunction in the psychological, biological, or developmental processes underlying mental functioning." The final draft of ICD-11 contains a very similar definition.
The terms "mental breakdown" or "nervous breakdown" may be used by the general population to mean a mental disorder. The terms "nervous breakdown" and "mental breakdown" have not been formally defined through a medical diagnostic system such as the DSM-5 or ICD-10 and are nearly absent from scientific literature regarding mental illness. Although "nervous breakdown" is not rigorously defined, surveys of laypersons suggest that the term refers to a specific acute time-limited reactive disorder involving symptoms such as anxiety or depression, usually precipitated by external stressors. Many health experts today refer to a nervous breakdown as a mental health crisis.
Nervous illness
In addition to the concept of mental disorder, some people have argued for a return to the old-fashioned concept of nervous illness. In How Everyone Became Depressed: The Rise and Fall of the Nervous Breakdown (2013), Edward Shorter, a professor of psychiatry and the history of medicine, says:
About half of them are depressed. Or at least that is the diagnosis that they got when they were put on antidepressants. ... They go to work but they are unhappy and uncomfortable; they are somewhat anxious; they are tired; they have various physical pains—and they tend to obsess about the whole business. There is a term for what they have, and it is a good old-fashioned term that has gone out of use. They have nerves or a nervous illness. It is an illness not just of mind or brain, but a disorder of the entire body. ... We have a package here of five symptoms—mild depression, some anxiety, fatigue, somatic pains, and obsessive thinking. ... We have had nervous illness for centuries. When you are too nervous to function ... it is a nervous breakdown. But that term has vanished from medicine, although not from the way we speak.... The nervous patients of yesteryear are the depressives of today. That is the bad news.... There is a deeper illness that drives depression and the symptoms of mood. We can call this deeper illness something else, or invent a neologism, but we need to get the discussion off depression and onto this deeper disorder in the brain and body. That is the point.— Edward Shorter, Faculty of Medicine, the University of Toronto
In eliminating the nervous breakdown, psychiatry has come close to having its own nervous breakdown.— David Healy, MD, FRCPsych, Professor of Psychiatry, University of Cardiff, Wales
Nerves stand at the core of common mental illness, no matter how much we try to forget them.— Peter J. Tyrer, FMedSci, Professor of Community Psychiatry, Imperial College, London
"Nervous breakdown" is a pseudo-medical term to describe a wealth of stress-related feelings and they are often made worse by the belief that there is a real phenomenon called "nervous breakdown". — Richard E. Vatz, co-author of explication of views of Thomas Szasz in "Thomas Szasz: Primary Values and Major Contentions"
Classifications
Main article: Classification of mental disorders
There are currently two widely established systems that classify mental disorders:
ICD-11 Chapter 06: Mental, behavioural or neurodevelopmental disorders, part of the International Classification of Diseases produced by the WHO (in effect since 1 January 2022).
Diagnostic and Statistical Manual of Mental Disorders (DSM-5) produced by the APA since 1952.
Both of these list categories of disorder and provide standardized criteria for diagnosis. They have deliberately converged their codes in recent revisions so that the manuals are often broadly comparable, although significant differences remain. Other classification schemes may be used in non-western cultures, for example, the Chinese Classification of Mental Disorders, and other manuals may be used by those of alternative theoretical persuasions, such as the Psychodynamic Diagnostic Manual. In general, mental disorders are classified separately from neurological disorders, learning disabilities or intellectual disability.
Unlike the DSM and ICD, some approaches are not based on identifying distinct categories of disorder using dichotomous symptom profiles intended to separate the abnormal from the normal. There is significant scientific debate about the relative merits of categorical versus such non-categorical (or hybrid) schemes, also known as continuum or dimensional models. A spectrum approach may incorporate elements of both.
In the scientific and academic literature on the definition or classification of mental disorder, one extreme argues that it is entirely a matter of value judgements (including of what is normal) while another proposes that it is or could be entirely objective and scientific (including by reference to statistical norms). Common hybrid views argue that the concept of mental disorder is objective even if only a "fuzzy prototype" that can never be precisely defined, or conversely that the concept always involves a mixture of scientific facts and subjective value judgments. Although the diagnostic categories are referred to as 'disorders', they are presented as medical diseases, but are not validated in the same way as most medical diagnoses. Some neurologists argue that classification will only be reliable and valid when based on neurobiological features rather than clinical interview, while others suggest that the differing ideological and practical perspectives need to be better integrated.
The DSM and ICD approach remains under attack both because of the implied causality model and because some researchers believe it better to aim at underlying brain differences which can precede symptoms by many years.
Dimensional models
The high degree of comorbidity between disorders in categorical models such as the DSM and ICD have led some to propose dimensional models. Studying comorbidity between disorders have demonstrated two latent (unobserved) factors or dimensions in the structure of mental disorders that are thought to possibly reflect etiological processes. These two dimensions reflect a distinction between internalizing disorders, such as mood or anxiety symptoms, and externalizing disorders such as behavioral or substance use symptoms. A single general factor of psychopathology, similar to the g factor for intelligence, has been empirically supported. The p factor model supports the internalizing-externalizing distinction, but also supports the formation of a third dimension of thought disorders such as schizophrenia. Biological evidence also supports the validity of the internalizing-externalizing structure of mental disorders, with twin and adoption studies supporting heritable factors for externalizing and internalizing disorders. A leading dimensional model is the Hierarchical Taxonomy of Psychopathology.
Disorders
See also: List of mental disorders as defined by the DSM and ICD
There are many different categories of mental disorder, and many different facets of human behavior and personality that can become disordered.
Anxiety disorder
Main article: Anxiety disorder
An anxiety disorder is anxiety or fear that interferes with normal functioning may be classified as an anxiety disorder. Commonly recognized categories include specific phobias, generalized anxiety disorder, social anxiety disorder, panic disorder, agoraphobia, obsessive–compulsive disorder and post-traumatic stress disorder.
Mood disorder
Main article: Mood disorder
Other affective (emotion/mood) processes can also become disordered. Mood disorder involving unusually intense and sustained sadness, melancholia, or despair is known as major depression (also known as unipolar or clinical depression). Milder, but still prolonged depression, can be diagnosed as dysthymia. Bipolar disorder (also known as manic depression) involves abnormally "high" or pressured mood states, known as mania or hypomania, alternating with normal or depressed moods. The extent to which unipolar and bipolar mood phenomena represent distinct categories of disorder, or mix and merge along a dimension or spectrum of mood, is subject to some scientific debate.
Psychotic disorder
Main article: Psychotic disorder
Patterns of belief, language use and perception of reality can become dysregulated (e.g., delusions, thought disorder, hallucinations). Psychotic disorders in this domain include schizophrenia, and delusional disorder. Schizoaffective disorder is a category used for individuals showing aspects of both schizophrenia and affective disorders. Schizotypy is a category used for individuals showing some of the characteristics associated with schizophrenia, but without meeting cutoff criteria.
Personality disorder
Main article: Personality disorder
Personality—the fundamental characteristics of a person that influence thoughts and behaviors across situations and time—may be considered disordered if judged to be abnormally rigid and maladaptive. Although treated separately by some, the commonly used categorical schemes include them as mental disorders, albeit on a separate axis II in the case of the DSM-IV. A number of different personality disorders are listed, including those sometimes classed as eccentric, such as paranoid, schizoid and schizotypal personality disorders; types that have described as dramatic or emotional, such as antisocial, borderline, histrionic or narcissistic personality disorders; and those sometimes classed as fear-related, such as anxious-avoidant, dependent, or obsessive–compulsive personality disorders. Personality disorders, in general, are defined as emerging in childhood, or at least by adolescence or early adulthood. The ICD also has a category for enduring personality change after a catastrophic experience or psychiatric illness. If an inability to sufficiently adjust to life circumstances begins within three months of a particular event or situation, and ends within six months after the stressor stops or is eliminated, it may instead be classed as an adjustment disorder. There is an emerging consensus that personality disorders, similar to personality traits in general, incorporate a mixture of acute dysfunctional behaviors that may resolve in short periods, and maladaptive temperamental traits that are more enduring. Furthermore, there are also non-categorical schemes that rate all individuals via a profile of different dimensions of personality without a symptom-based cutoff from normal personality variation, for example through schemes based on dimensional models.
Eating disorder
Main article: Eating disorder
An eating disorder is a serious mental health condition that involves an unhealthy relationship with food and body image. They can cause severe physical and psychological problems. Eating disorders involve disproportionate concern in matters of food and weight. Categories of disorder in this area include anorexia nervosa, bulimia nervosa, exercise bulimia or binge eating disorder.
Sleep disorder
Main article: Sleep disorder
Sleep disorders are associated with disruption to normal sleep patterns. A common sleep disorder is insomnia, which is described as difficulty falling and/or staying asleep. Other sleep disorders include narcolepsy, sleep apnea, REM sleep behavior disorder, chronic sleep deprivation, and restless leg syndrome.
Narcolepsy is a condition of extreme tendencies to fall asleep whenever and wherever. People with narcolepsy feel refreshed after their random sleep, but eventually get sleepy again. Narcolepsy diagnosis requires an overnight stay at a sleep center for analysis, during which doctors ask for a detailed sleep history and sleep records. Doctors also use actigraphs and polysomnography. Doctors will do a multiple sleep latency test, which measures how long it takes a person to fall asleep.
Sleep apnea, when breathing repeatedly stops and starts during sleep, can be a serious sleep disorder. Three types of sleep apnea include obstructive sleep apnea, central sleep apnea, and complex sleep apnea. Sleep apnea can be diagnosed at home or with polysomnography at a sleep center. An ear, nose, and throat doctor may further help with the sleeping habits.
Sexuality related
Sexual disorders include dyspareunia and various kinds of paraphilia (sexual arousal to objects, situations, or individuals that are considered abnormal or harmful to the person or others).
Other
Impulse control disorder: People who are abnormally unable to resist certain urges or impulses that could be harmful to themselves or others, may be classified as having an impulse control disorder, and disorders such as kleptomania (stealing) or pyromania (fire-setting). Various behavioral addictions, such as gambling addiction, may be classed as a disorder. Obsessive–compulsive disorder can sometimes involve an inability to resist certain acts but is classed separately as being primarily an anxiety disorder.
Substance use disorder: This disorder refers to the use of drugs (legal or illegal, including alcohol) that persists despite significant problems or harm related to its use. Substance dependence and substance abuse fall under this umbrella category in the DSM. Substance use disorder may be due to a pattern of compulsive and repetitive use of a drug that results in tolerance to its effects and withdrawal symptoms when use is reduced or stopped.
Dissociative disorder: People with severe disturbances of their self-identity, memory, and general awareness of themselves and their surroundings may be classified as having these types of disorders, including depersonalization disorder or dissociative identity disorder (which was previously referred to as multiple personality disorder or "split personality").
Cognitive disorder: These affect cognitive abilities, including learning and memory. This category includes delirium and mild and major neurocognitive disorder (previously termed dementia).
Developmental disorder: These disorders initially occur in childhood. Some examples include autism spectrum disorder, oppositional defiant disorder and conduct disorder, and attention deficit hyperactivity disorder (ADHD), which may continue into adulthood. Conduct disorder, if continuing into adulthood, may be diagnosed as antisocial personality disorder (dissocial personality disorder in the ICD). Popular labels such as psychopath (or sociopath) do not appear in the DSM or ICD but are linked by some to these diagnoses.
Somatoform disorders may be diagnosed when there are problems that appear to originate in the body that are thought to be manifestations of a mental disorder. This includes somatization disorder and conversion disorder. There are also disorders of how a person perceives their body, such as body dysmorphic disorder. Neurasthenia is an old diagnosis involving somatic complaints as well as fatigue and low spirits/depression, which is officially recognized by the ICD-10 but no longer by the DSM-IV.
Factitious disorders are diagnosed where symptoms are thought to be reported for personal gain. Symptoms are often deliberately produced or feigned, and may relate to either symptoms in the individual or in someone close to them, particularly people they care for.
There are attempts to introduce a category of relational disorder, where the diagnosis is of a relationship rather than on any one individual in that relationship. The relationship may be between children and their parents, between couples, or others. There already exists, under the category of psychosis, a diagnosis of shared psychotic disorder where two or more individuals share a particular delusion because of their close relationship with each other.
There are a number of uncommon psychiatric syndromes, which are often named after the person who first described them, such as Capgras syndrome, De Clerambault syndrome, Othello syndrome, Ganser syndrome, Cotard delusion, and Ekbom syndrome, and additional disorders such as the Couvade syndrome and Geschwind syndrome.
Signs and symptoms
Course
The onset of psychiatric disorders usually occurs from childhood to early adulthood. Impulse-control disorders and a few anxiety disorders tend to appear in childhood. Some other anxiety disorders, substance disorders, and mood disorders emerge later in the mid-teens. Symptoms of schizophrenia typically manifest from late adolescence to early twenties.
The likely course and outcome of mental disorders vary and are dependent on numerous factors related to the disorder itself, the individual as a whole, and the social environment. Some disorders may last a brief period of time, while others may be long-term in nature.
All disorders can have a varied course. Long-term international studies of schizophrenia have found that over a half of individuals recover in terms of symptoms, and around a fifth to a third in terms of symptoms and functioning, with many requiring no medication. While some have serious difficulties and support needs for many years, "late" recovery is still plausible. The World Health Organization (WHO) concluded that the long-term studies' findings converged with others in "relieving patients, carers and clinicians of the chronicity paradigm which dominated thinking throughout much of the 20th century."
A follow-up study by Tohen and coworkers revealed that around half of people initially diagnosed with bipolar disorder achieve symptomatic recovery (no longer meeting criteria for the diagnosis) within six weeks, and nearly all achieve it within two years, with nearly half regaining their prior occupational and residential status in that period. Less than half go on to experience a new episode of mania or major depression within the next two years.
Disability
Disorder
Disability-adjusted life years
Major depressive disorder
65.5 million
Alcohol-use disorder
23.7 million
Schizophrenia
16.8 million
Bipolar disorder
14.4 million
Other drug-use disorders
8.4 million
Panic disorder
7.0 million
Obsessive–compulsive disorder
5.1 million
Primary insomnia
3.6 million
Post-traumatic stress disorder
3.5 million
Some disorders may be very limited in their functional effects, while others may involve substantial disability and support needs. In this context, the terms psychiatric disability and psychological disability are sometimes used instead of mental disorder. The degree of ability or disability may vary over time and across different life domains. Furthermore, psychiatric disability has been linked to institutionalization, discrimination and social exclusion as well as to the inherent effects of disorders. Alternatively, functioning may be affected by the stress of having to hide a condition in work or school, etc., by adverse effects of medications or other substances, or by mismatches between illness-related variations and demands for regularity.
It is also the case that, while often being characterized in purely negative terms, some mental traits or states labeled as psychiatric disabilities can also involve above-average creativity, non-conformity, goal-striving, meticulousness, or empathy. In addition, the public perception of the level of disability associated with mental disorders can change.
Nevertheless, internationally, people report equal or greater disability from commonly occurring mental conditions than from commonly occurring physical conditions, particularly in their social roles and personal relationships. The proportion with access to professional help for mental disorders is far lower, however, even among those assessed as having a severe psychiatric disability. Disability in this context may or may not involve such things as:
Basic activities of daily living. Including looking after the self (health care, grooming, dressing, shopping, cooking etc.) or looking after accommodation (chores, DIY tasks, etc.)
Interpersonal relationships. Including communication skills, ability to form relationships and sustain them, ability to leave the home or mix in crowds or particular settings
Occupational functioning. Ability to acquire an employment and hold it, cognitive and social skills required for the job, dealing with workplace culture, or studying as a student.
In terms of total disability-adjusted life years (DALYs), which is an estimate of how many years of life are lost due to premature death or to being in a state of poor health and disability, psychiatric disabilities rank amongst the most disabling conditions. Unipolar (also known as Major) depressive disorder is the third leading cause of disability worldwide, of any condition mental or physical, accounting for 65.5 million years lost. The first systematic description of global disability arising in youth, in 2011, found that among 10- to 24-year-olds nearly half of all disability (current and as estimated to continue) was due to psychiatric disabilities, including substance use disorders and conditions involving self-harm. Second to this were accidental injuries (mainly traffic collisions) accounting for 12 percent of disability, followed by communicable diseases at 10 percent. The psychiatric disabilities associated with most disabilities in high-income countries were unipolar major depression (20%) and alcohol use disorder (11%). In the eastern Mediterranean region, it was unipolar major depression (12%) and schizophrenia (7%), and in Africa it was unipolar major depression (7%) and bipolar disorder (5%).
Suicide, which is often attributed to some underlying mental disorder, is a leading cause of death among teenagers and adults under 35. There are an estimated 10 to 20 million non-fatal attempted suicides every year worldwide.
Risk factors
Main article: Causes of mental disorders
The predominant view as of 2018 is that genetic, psychological, and environmental factors all contribute to the development or progression of mental disorders. Different risk factors may be present at different ages, with risk occurring as early as during prenatal period.
Genetics
Main article: Psychiatric genetics
A number of psychiatric disorders are linked to a family history (including depression, narcissistic personality disorder and anxiety). Twin studies have also revealed a very high heritability for many mental disorders (especially autism and schizophrenia). Although researchers have been looking for decades for clear linkages between genetics and mental disorders, that work has not yielded specific genetic biomarkers yet that might lead to better diagnosis and better treatments.
Statistical research looking at eleven disorders found widespread assortative mating between people with mental illness. That means that individuals with one of these disorders were two to three times more likely than the general population to have a partner with a mental disorder. Sometimes people seemed to have preferred partners with the same mental illness. Thus, people with schizophrenia or ADHD are seven times more likely to have affected partners with the same disorder. This is even more pronounced for people with Autism spectrum disorders who are 10 times more likely to have a spouse with the same disorder.
Environment
The prevalence of mental illness is higher in more economically unequal countries.
During the prenatal stage, factors like unwanted pregnancy, lack of adaptation to pregnancy or substance use during pregnancy increases the risk of developing a mental disorder. Maternal stress and birth complications including prematurity and infections have also been implicated in increasing susceptibility for mental illness. Infants neglected or not provided optimal nutrition have a higher risk of developing cognitive impairment.
Social influences have also been found to be important, including abuse, neglect, bullying, social stress, traumatic events, and other negative or overwhelming life experiences. Aspects of the wider community have also been implicated, including employment problems, socioeconomic inequality, lack of social cohesion, problems linked to migration, and features of particular societies and cultures. The specific risks and pathways to particular disorders are less clear, however.
Nutrition also plays a role in mental disorders.
In schizophrenia and psychosis, risk factors include migration and discrimination, childhood trauma, bereavement or separation in families, recreational use of drugs, and urbanicity.
In anxiety, risk factors may include parenting factors including parental rejection, lack of parental warmth, high hostility, harsh discipline, high maternal negative affect, anxious childrearing, modelling of dysfunctional and drug-abusing behavior, and child abuse (emotional, physical and sexual). Adults with imbalance work to life are at higher risk for developing anxiety.
For bipolar disorder, stress (such as childhood adversity) is not a specific cause, but does place genetically and biologically vulnerable individuals at risk for a more severe course of illness.
Drug use
Mental disorders are associated with drug use including: cannabis, alcohol and caffeine, use of which appears to promote anxiety. For psychosis and schizophrenia, usage of a number of drugs has been associated with development of the disorder, including cannabis, cocaine, and amphetamines. There has been debate regarding the relationship between usage of cannabis and bipolar disorder. Cannabis has also been associated with depression. Adolescents are at increased risk for tobacco, alcohol and drug use; Peer pressure is the main reason why adolescents start using substances. At this age, the use of substances could be detrimental to the development of the brain and place them at higher risk of developing a mental disorder.
Chronic disease
People living with chronic conditions like HIV and diabetes are at higher risk of developing a mental disorder. People living with diabetes experience significant stress from the biological impact of the disease, which places them at risk for developing anxiety and depression. Diabetic patients also have to deal with emotional stress trying to manage the disease. Conditions like heart disease, stroke, respiratory conditions, cancer, and arthritis increase the risk of developing a mental disorder when compared to the general population.
Personality traits
Risk factors for mental illness include a propensity for high neuroticism or "emotional instability". In anxiety, risk factors may include temperament and attitudes (e.g. pessimism).
Causal models
Mental disorders can arise from multiple sources, and in many cases there is no single accepted or consistent cause currently established. An eclectic or pluralistic mix of models may be used to explain particular disorders. The primary paradigm of contemporary mainstream Western psychiatry is said to be the biopsychosocial model which incorporates biological, psychological and social factors, although this may not always be applied in practice.
Biological psychiatry follows a biomedical model where many mental disorders are conceptualized as disorders of brain circuits likely caused by developmental processes shaped by a complex interplay of genetics and experience. A common assumption is that disorders may have resulted from genetic and developmental vulnerabilities, exposed by stress in life (for example in a diathesis–stress model), although there are various views on what causes differences between individuals. Some types of mental disorders may be viewed as primarily neurodevelopmental disorders.
Evolutionary psychology may be used as an overall explanatory theory, while attachment theory is another kind of evolutionary-psychological approach sometimes applied in the context of mental disorders. Psychoanalytic theories have continued to evolve alongside and cognitive-behavioral and systemic-family approaches. A distinction is sometimes made between a "medical model" or a "social model" of psychiatric disability.
Diagnosis
Psychiatrists seek to provide a medical diagnosis of individuals by an assessment of symptoms, signs and impairment associated with particular types of mental disorder. Other mental health professionals, such as clinical psychologists, may or may not apply the same diagnostic categories to their clinical formulation of a client's difficulties and circumstances. The majority of mental health problems are, at least initially, assessed and treated by family physicians (in the UK general practitioners) during consultations, who may refer a patient on for more specialist diagnosis in acute or chronic cases.
Routine diagnostic practice in mental health services typically involves an interview known as a mental status examination, where evaluations are made of appearance and behavior, self-reported symptoms, mental health history, and current life circumstances. The views of other professionals, relatives, or other third parties may be taken into account. A physical examination to check for ill health or the effects of medications or other drugs may be conducted. Psychological testing is sometimes used via paper-and-pen or computerized questionnaires, which may include algorithms based on ticking off standardized diagnostic criteria, and in rare specialist cases neuroimaging tests may be requested, but such methods are more commonly found in research studies than routine clinical practice.
Time and budgetary constraints often limit practicing psychiatrists from conducting more thorough diagnostic evaluations. It has been found that most clinicians evaluate patients using an unstructured, open-ended approach, with limited training in evidence-based assessment methods, and that inaccurate diagnosis may be common in routine practice. In addition, comorbidity is very common in psychiatric diagnosis, where the same person meets the criteria for more than one disorder. On the other hand, a person may have several different difficulties only some of which meet the criteria for being diagnosed. There may be specific problems with accurate diagnosis in developing countries.
More structured approaches are being increasingly used to measure levels of mental illness.
HoNOS is the most widely used measure in English mental health services, being used by at least 61 trusts. In HoNOS a score of 0–4 is given for each of 12 factors, based on functional living capacity. Research has been supportive of HoNOS, although some questions have been asked about whether it provides adequate coverage of the range and complexity of mental illness problems, and whether the fact that often only 3 of the 12 scales vary over time gives enough subtlety to accurately measure outcomes of treatment.
Criticism
This section relies excessively on references to primary sources. Please improve this section by adding secondary or tertiary sources. Find sources: "criticism" psychiatric diagnosis – news · newspapers · books · scholar · JSTOR (July 2021) (Learn how and when to remove this template message)
Since the 1980s, Paula Caplan has been concerned about the subjectivity of psychiatric diagnosis, and people being arbitrarily "slapped with a psychiatric label." Caplan says because psychiatric diagnosis is unregulated, doctors are not required to spend much time interviewing patients or to seek a second opinion. The Diagnostic and Statistical Manual of Mental Disorders can lead a psychiatrist to focus on narrow checklists of symptoms, with little consideration of what is actually causing the person's problems. So, according to Caplan, getting a psychiatric diagnosis and label often stands in the way of recovery.
In 2013, psychiatrist Allen Frances wrote a paper entitled "The New Crisis of Confidence in Psychiatric Diagnosis", which said that "psychiatric diagnosis... still relies exclusively on fallible subjective judgments rather than objective biological tests." Frances was also concerned about "unpredictable overdiagnosis." For many years, marginalized psychiatrists (such as Peter Breggin, Thomas Szasz) and outside critics (such as Stuart A. Kirk) have "been accusing psychiatry of engaging in the systematic medicalization of normality." More recently these concerns have come from insiders who have worked for and promoted the American Psychiatric Association (e.g., Robert Spitzer, Allen Frances). A 2002 editorial in the British Medical Journal warned of inappropriate medicalization leading to disease mongering, where the boundaries of the definition of illnesses are expanded to include personal problems as medical problems or risks of diseases are emphasized to broaden the market for medications.
Gary Greenberg, a psychoanalyst, in his book "the Book of Woe", argues that mental illness is really about suffering and how the DSM creates diagnostic labels to categorize people's suffering. Indeed, the psychiatrist Thomas Szasz, in his book "the Medicalization of Everyday Life", also argues that what is psychiatric illness, is not always biological in nature (i.e. social problems, poverty, etc.), and may even be a part of the human condition.
Potential routine use of MRI/fMRI in diagnosis
in 2018 the American Psychological Association commissioned a review to reach a consensus on whether modern clinical MRI/fMRI will be able to be used in the diagnosis of mental health disorders. The criteria presented by the APA stated that the biomarkers used in diagnosis should:
"have a sensitivity of at least 80% for detecting a particular psychiatric disorder"
should "have a specificity of at least 80% for distinguishing this disorder from other psychiatric or medical disorders"
"should be reliable, reproducible, and ideally be noninvasive, simple to perform, and inexpensive"
proposed biomarkers should be verified by 2 independent studies each by a different investigator and different population samples and published in a peer-reviewed journal.
The review concluded that although neuroimaging diagnosis may technically be feasible, very large studies are needed to evaluate specific biomarkers which were not available.
Prevention
Main article: Prevention of mental disorders
The 2004 WHO report "Prevention of Mental Disorders" stated that "Prevention of these disorders is obviously one of the most effective ways to reduce the [disease] burden."
The 2011 European Psychiatric Association (EPA) guidance on prevention of mental disorders states "There is considerable evidence that various psychiatric conditions can be prevented through the implementation of effective evidence-based interventions."
A 2011 UK Department of Health report on the economic case for mental health promotion and mental illness prevention found that "many interventions are outstandingly good value for money, low in cost and often become self-financing over time, saving public expenditure".
In 2016, the National Institute of Mental Health re-affirmed prevention as a research priority area.
Parenting may affect the child's mental health, and evidence suggests that helping parents to be more effective with their children can address mental health needs.
Universal prevention (aimed at a population that has no increased risk for developing a mental disorder, such as school programs or mass media campaigns) need very high numbers of people to show effect (sometimes known as the "power" problem). Approaches to overcome this are (1) focus on high-incidence groups (e.g. by targeting groups with high risk factors), (2) use multiple interventions to achieve greater, and thus more statistically valid, effects, (3) use cumulative meta-analyses of many trials, and (4) run very large trials.
Management
Main articles: Treatment of mental disorders, Services for mental disorders, and Mental health professional
"Haus Tornow am See" (former manor house), Germany from 1912 is today separated into a special education school and a hotel with integrated work/job- and rehabilitation-training for people with mental disorders.
Treatment and support for mental disorders are provided in psychiatric hospitals, clinics or a range of community mental health services. In some countries services are increasingly based on a recovery approach, intended to support individual's personal journey to gain the kind of life they want.
There is a range of different types of treatment and what is most suitable depends on the disorder and the individual. Many things have been found to help at least some people, and a placebo effect may play a role in any intervention or medication. In a minority of cases, individuals may be treated against their will, which can cause particular difficulties depending on how it is carried out and perceived. Compulsory treatment while in the community versus non-compulsory treatment does not appear to make much of a difference except by maybe decreasing victimization.
Lifestyle
Lifestyle strategies, including dietary changes, exercise and quitting smoking may be of benefit.
Therapy
There is also a wide range of psychotherapists (including family therapy), counselors, and public health professionals. In addition, there are peer support roles where personal experience of similar issues is the primary source of expertise.
A major option for many mental disorders is psychotherapy. There are several main types. Cognitive behavioral therapy (CBT) is widely used and is based on modifying the patterns of thought and behavior associated with a particular disorder. Other psychotherapies include dialectic behavioral therapy (DBT) and interpersonal psychotherapy (IPT). Psychoanalysis, addressing underlying psychic conflicts and defenses, has been a dominant school of psychotherapy and is still in use. Systemic therapy or family therapy is sometimes used, addressing a network of significant others as well as an individual.
Some psychotherapies are based on a humanistic approach. There are many specific therapies used for particular disorders, which may be offshoots or hybrids of the above types. Mental health professionals often employ an eclectic or integrative approach. Much may depend on the therapeutic relationship, and there may be problems with trust, confidentiality and engagement.
Medication
A major option for many mental disorders is psychiatric medication and there are several main groups. Antidepressants are used for the treatment of clinical depression, as well as often for anxiety and a range of other disorders. Anxiolytics (including sedatives) are used for anxiety disorders and related problems such as insomnia. Mood stabilizers are used primarily in bipolar disorder. Antipsychotics are used for psychotic disorders, notably for positive symptoms in schizophrenia, and also increasingly for a range of other disorders. Stimulants are commonly used, notably for ADHD.
Despite the different conventional names of the drug groups, there may be considerable overlap in the disorders for which they are actually indicated, and there may also be off-label use of medications. There can be problems with adverse effects of medication and adherence to them, and there is also criticism of pharmaceutical marketing and professional conflicts of interest. However, these medications in combination with non-pharmacological methods, such as cognitive-behavioral therapy (CBT) are seen to be most effective in treating mental disorders.
Other
Electroconvulsive therapy (ECT) is sometimes used in severe cases when other interventions for severe intractable depression have failed. ECT is usually indicated for treatment resistant depression, severe vegetative symptoms, psychotic depression, intense suicidal ideation, depression during pregnancy, and catatonia. Psychosurgery is considered experimental but is advocated by some neurologists in certain rare cases.
Counseling (professional) and co-counseling (between peers) may be used. Psychoeducation programs may provide people with the information to understand and manage their problems. Creative therapies are sometimes used, including music therapy, art therapy or drama therapy. Lifestyle adjustments and supportive measures are often used, including peer support, self-help groups for mental health and supported housing or supported employment (including social firms). Some advocate dietary supplements.
Reasonable accommodations (adjustments and supports) might be put in place to help an individual cope and succeed in environments despite potential disability related to mental health problems. This could include an emotional support animal or specifically trained psychiatric service dog. As of 2019 cannabis is specifically not recommended as a treatment.
Epidemiology
Main article: Prevalence of mental disorders
Deaths from mental and behavioral disorders per million persons in 2012 0–6 7–9 10–15 16–24 25–31 32–39 40–53 54–70 71–99 100–356
Disability-adjusted life year for neuropsychiatric conditions per 100,000 inhabitants in 2004 <2,200 2,200–2,400 2,400–2,600 2,600–2,800 2,800–3,000 3,000–3,200 3,200–3,400 3,400–3,600 3,600–3,800 3,800–4,000 4,000–4,200 >4,200
Mental disorders are common. Worldwide, more than one in three people in most countries report sufficient criteria for at least one at some point in their life. In the United States, 46% qualify for a mental illness at some point. An ongoing survey indicates that anxiety disorders are the most common in all but one country, followed by mood disorders in all but two countries, while substance disorders and impulse-control disorders were consistently less prevalent. Rates varied by region.
A review of anxiety disorder surveys in different countries found average lifetime prevalence estimates of 16.6%, with women having higher rates on average. A review of mood disorder surveys in different countries found lifetime rates of 6.7% for major depressive disorder (higher in some studies, and in women) and 0.8% for Bipolar I disorder.
In the United States the frequency of disorder is: anxiety disorder (28.8%), mood disorder (20.8%), impulse-control disorder (24.8%) or substance use disorder (14.6%).
A 2004 cross-Europe study found that approximately one in four people reported meeting criteria at some point in their life for at least one of the DSM-IV disorders assessed, which included mood disorders (13.9%), anxiety disorders (13.6%), or alcohol disorder (5.2%). Approximately one in ten met the criteria within a 12-month period. Women and younger people of either gender showed more cases of the disorder. A 2005 review of surveys in 16 European countries found that 27% of adult Europeans are affected by at least one mental disorder in a 12-month period.
An international review of studies on the prevalence of schizophrenia found an average (median) figure of 0.4% for lifetime prevalence; it was consistently lower in poorer countries.
Studies of the prevalence of personality disorders (PDs) have been fewer and smaller-scale, but one broad Norwegian survey found a five-year prevalence of almost 1 in 7 (13.4%). Rates for specific disorders ranged from 0.8% to 2.8%, differing across countries, and by gender, educational level and other factors. A US survey that incidentally screened for personality disorder found a rate of 14.79%.
Approximately 7% of a preschool pediatric sample were given a psychiatric diagnosis in one clinical study, and approximately 10% of 1- and 2-year-olds receiving developmental screening have been assessed as having significant emotional/behavioral problems based on parent and pediatrician reports.
While rates of psychological disorders are often the same for men and women, women tend to have a higher rate of depression. Each year 73 million women are affected by major depression, and suicide is ranked 7th as the cause of death for women between the ages of 20–59. Depressive disorders account for close to 41.9% of the psychiatric disabilities among women compared to 29.3% among men.
History
Main article: History of mental disorders
Ancient civilizations
Ancient civilizations described and treated a number of mental disorders. Mental illnesses were well known in ancient Mesopotamia, where diseases and mental disorders were believed to be caused by specific deities. Because hands symbolized control over a person, mental illnesses were known as "hands" of certain deities. One psychological illness was known as Qāt Ištar, meaning "Hand of Ishtar". Others were known as "Hand of Shamash", "Hand of the Ghost", and "Hand of the God". Descriptions of these illnesses, however, are so vague that it is usually impossible to determine which illnesses they correspond to in modern terminology. Mesopotamian doctors kept detailed record of their patients' hallucinations and assigned spiritual meanings to them. The royal family of Elam was notorious for its members often being insane. The Greeks coined terms for melancholy, hysteria and phobia and developed the humorism theory. Mental disorders were described, and treatments developed, in Persia, Arabia and in the medieval Islamic world.
Europe
Middle Ages
Conceptions of madness in the Middle Ages in Christian Europe were a mixture of the divine, diabolical, magical and humoral, and transcendental. In the early modern period, some people with mental disorders may have been victims of the witch-hunts. While not every witch and sorcerer accused were mentally ill, all mentally ill were considered to be witches or sorcerers. Many terms for mental disorders that found their way into everyday use first became popular in the 16th and 17th centuries.
Eighteenth century
Eight patients representing mental diagnoses as of the 19th century at the Salpêtrière, Paris
By the end of the 17th century and into the Enlightenment, madness was increasingly seen as an organic physical phenomenon with no connection to the soul or moral responsibility. Asylum care was often harsh and treated people like wild animals, but towards the end of the 18th century a moral treatment movement gradually developed. Clear descriptions of some syndromes may be rare before the 19th century.
Nineteenth century
Industrialization and population growth led to a massive expansion of the number and size of insane asylums in every Western country in the 19th century. Numerous different classification schemes and diagnostic terms were developed by different authorities, and the term psychiatry was coined (1808), though medical superintendents were still known as alienists.
Twentieth century
A patient in a strait-jacket and barrel contraption, 1908
The turn of the 20th century saw the development of psychoanalysis, which would later come to the fore, along with Kraepelin's classification scheme. Asylum "inmates" were increasingly referred to as "patients", and asylums were renamed as hospitals.
Europe and the United States
Insulin shock procedure, 1950s
Early in the 20th century in the United States, a mental hygiene movement developed, aiming to prevent mental disorders. Clinical psychology and social work developed as professions. World War I saw a massive increase of conditions that came to be termed "shell shock".
World War II saw the development in the U.S. of a new psychiatric manual for categorizing mental disorders, which along with existing systems for collecting census and hospital statistics led to the first Diagnostic and Statistical Manual of Mental Disorders. The International Classification of Diseases (ICD) also developed a section on mental disorders. The term stress, having emerged from endocrinology work in the 1930s, was increasingly applied to mental disorders.
Electroconvulsive therapy, insulin shock therapy, lobotomies and the neuroleptic chlorpromazine came to be used by mid-century. In the 1960s there were many challenges to the concept of mental illness itself. These challenges came from psychiatrists like Thomas Szasz who argued that mental illness was a myth used to disguise moral conflicts; from sociologists such as Erving Goffman who said that mental illness was merely another example of how society labels and controls non-conformists; from behavioral psychologists who challenged psychiatry's fundamental reliance on unobservable phenomena; and from gay rights activists who criticised the APA's listing of homosexuality as a mental disorder. A study published in Science by Rosenhan received much publicity and was viewed as an attack on the efficacy of psychiatric diagnosis.
Deinstitutionalization gradually occurred in the West, with isolated psychiatric hospitals being closed down in favor of community mental health services. A consumer/survivor movement gained momentum. Other kinds of psychiatric medication gradually came into use, such as "psychic energizers" (later antidepressants) and lithium. Benzodiazepines gained widespread use in the 1970s for anxiety and depression, until dependency problems curtailed their popularity.
Advances in neuroscience, genetics, and psychology led to new research agendas. Cognitive behavioral therapy and other psychotherapies developed. The DSM and then ICD adopted new criteria-based classifications, and the number of "official" diagnoses saw a large expansion. Through the 1990s, new SSRI-type antidepressants became some of the most widely prescribed drugs in the world, as later did antipsychotics. Also during the 1990s, a recovery approach developed.
Africa and Nigeria
Most Africans view mental disturbances as external spiritual attack on the person. Those who have a mental illness are thought to be under a spell or bewitched. Often than usual, People view a mentally ill person as possessed of an evil spirit and is seen as more of sociological perspective than a psychological order.
The WHO estimated that fewer than 10% of mentally ill Nigerians have access to a psychiatrist or health worker, because there is a low ratio of mental-health specialists available in a country of 200 million people. WHO estimates that the number of mentally ill Nigerians ranges from 40 million to 60 million. Disorders such as depression, anxiety, schizophrenia, personality disorder, old age-related disorder, and substance-abuse disorder are common in Nigeria, as in other countries in Africa.
Nigeria is still nowhere near being equipped to solve prevailing mental health challenges. With little scientific research carried out, coupled with insufficient mental-health hospitals in the country, traditional healers provide specialized psychotherapy care to those that require their services and pharmacotherapy
Society and culture
Different societies or cultures, even different individuals in a subculture, can disagree as to what constitutes optimal versus pathological biological and psychological functioning. Research has demonstrated that cultures vary in the relative importance placed on, for example, happiness, autonomy, or social relationships for pleasure. Likewise, the fact that a behavior pattern is valued, accepted, encouraged, or even statistically normative in a culture does not necessarily mean that it is conducive to optimal psychological functioning.
People in all cultures find some behaviors bizarre or even incomprehensible. But just what they feel is bizarre or incomprehensible is ambiguous and subjective. These differences in determination can become highly contentious. The process by which conditions and difficulties come to be defined and treated as medical conditions and problems, and thus come under the authority of doctors and other health professionals, is known as medicalization or pathologization.
Mental illness in the Latin American community
There is a perception in Latin American communities, especially among older people, that discussing problems with mental health can create embarrassment and shame for the family. This results in fewer people seeking treatment.
Latin Americans from the US are slightly more likely to have a mental health disorder than first-generation Latin American immigrants, although differences between ethnic groups were found to disappear after adjustment for place of birth.
From 2015 to 2018, rates of serious mental illness in young adult Latin Americans increased by 60%, from 4% to 6.4%. The prevalence of major depressive episodes in young and adult Latin Americans increased from 8.4% to 11.3%. More than a third of Latin Americans reported more than one bad mental health day in the last three months. The rate of suicide among Latin Americans was about half the rate of non-Latin American white Americans in 2018, and this was the second-leading cause of death among Latin Americans ages 15 to 34. However, Latin American suicide rates rose steadily after 2020 in relation to the COVID-19 pandemic, even as the national rate declined.
Family relations are an integral part of the Latin American community. Some research has shown that Latin Americans are more likely rely on family bonds, or familismo, as a source of therapy while struggling with mental health issues. Because Latin Americans have a high rate of religiosity, and because there is less stigma associated with religion than with psychiatric services, religion may play a more important therapeutic role for the mentally ill in Latin American communities. However, research has also suggested that religion may also play a role in stigmatizing mental illness in Latin American communities, which can discourage community members from seeking professional help.
Religion
See also: Psychology of religion
Religious, spiritual, or transpersonal experiences and beliefs meet many criteria of delusional or psychotic disorders. A belief or experience can sometimes be shown to produce distress or disability—the ordinary standard for judging mental disorders. There is a link between religion and schizophrenia, a complex mental disorder characterized by a difficulty in recognizing reality, regulating emotional responses, and thinking in a clear and logical manner. Those with schizophrenia commonly report some type of religious delusion, and religion itself may be a trigger for schizophrenia.
Movements
Giorgio Antonucci
Thomas Szasz
Controversy has often surrounded psychiatry, and the term anti-psychiatry was coined by the psychiatrist David Cooper in 1967. The anti-psychiatry message is that psychiatric treatments are ultimately more damaging than helpful to patients, and psychiatry's history involves what may now be seen as dangerous treatments. Electroconvulsive therapy was one of these, which was used widely between the 1930s and 1960s. Lobotomy was another practice that was ultimately seen as too invasive and brutal. Diazepam and other sedatives were sometimes over-prescribed, which led to an epidemic of dependence. There was also concern about the large increase in prescribing psychiatric drugs for children. Some charismatic psychiatrists came to personify the movement against psychiatry. The most influential of these was R.D. Laing who wrote a series of best-selling books, including The Divided Self. Thomas Szasz wrote The Myth of Mental Illness. Some ex-patient groups have become militantly anti-psychiatric, often referring to themselves as survivors. Giorgio Antonucci has questioned the basis of psychiatry through his work on the dismantling of two psychiatric hospitals (in the city of Imola), carried out from 1973 to 1996.
The consumer/survivor movement (also known as user/survivor movement) is made up of individuals (and organizations representing them) who are clients of mental health services or who consider themselves survivors of psychiatric interventions. Activists campaign for improved mental health services and for more involvement and empowerment within mental health services, policies and wider society. Patient advocacy organizations have expanded with increasing deinstitutionalization in developed countries, working to challenge the stereotypes, stigma and exclusion associated with psychiatric conditions. There is also a carers rights movement of people who help and support people with mental health conditions, who may be relatives, and who often work in difficult and time-consuming circumstances with little acknowledgement and without pay. An anti-psychiatry movement fundamentally challenges mainstream psychiatric theory and practice, including in some cases asserting that psychiatric concepts and diagnoses of 'mental illness' are neither real nor useful.
Alternatively, a movement for global mental health has emerged, defined as 'the area of study, research and practice that places a priority on improving mental health and achieving equity in mental health for all people worldwide'.
Cultural bias
See also: Depression and culture and Cultural competence in healthcare
Diagnostic guidelines of the 2000s, namely the DSM and to some extent the ICD, have been criticized as having a fundamentally Euro-American outlook. Opponents argue that even when diagnostic criteria are used across different cultures, it does not mean that the underlying constructs have validity within those cultures, as even reliable application can prove only consistency, not legitimacy. Advocating a more culturally sensitive approach, critics such as Carl Bell and Marcello Maviglia contend that the cultural and ethnic diversity of individuals is often discounted by researchers and service providers.
Cross-cultural psychiatrist Arthur Kleinman contends that the Western bias is ironically illustrated in the introduction of cultural factors to the DSM-IV. Disorders or concepts from non-Western or non-mainstream cultures are described as "culture-bound", whereas standard psychiatric diagnoses are given no cultural qualification whatsoever, revealing to Kleinman an underlying assumption that Western cultural phenomena are universal. Kleinman's negative view towards the culture-bound syndrome is largely shared by other cross-cultural critics. Common responses included both disappointment over the large number of documented non-Western mental disorders still left out and frustration that even those included are often misinterpreted or misrepresented.
Many mainstream psychiatrists are dissatisfied with the new culture-bound diagnoses, although for partly different reasons. Robert Spitzer, a lead architect of the DSM-III, has argued that adding cultural formulations was an attempt to appease cultural critics, and has stated that they lack any scientific rationale or support. Spitzer also posits that the new culture-bound diagnoses are rarely used, maintaining that the standard diagnoses apply regardless of the culture involved. In general, mainstream psychiatric opinion remains that if a diagnostic category is valid, cross-cultural factors are either irrelevant or are significant only to specific symptom presentations.
Clinical conceptions of mental illness also overlap with personal and cultural values in the domain of morality, so much so that it is sometimes argued that separating the two is impossible without fundamentally redefining the essence of being a particular person in a society. In clinical psychiatry, persistent distress and disability indicate an internal disorder requiring treatment; but in another context, that same distress and disability can be seen as an indicator of emotional struggle and the need to address social and structural problems. This dichotomy has led some academics and clinicians to advocate a postmodernist conceptualization of mental distress and well-being.
Such approaches, along with cross-cultural and "heretical" psychologies centered on alternative cultural and ethnic and race-based identities and experiences, stand in contrast to the mainstream psychiatric community's alleged avoidance of any explicit involvement with either morality or culture. In many countries there are attempts to challenge perceived prejudice against minority groups, including alleged institutional racism within psychiatric services. There are also ongoing attempts to improve professional cross cultural sensitivity.
Laws and policies
See also: Mental health law
Three-quarters of countries around the world have mental health legislation. Compulsory admission to mental health facilities (also known as involuntary commitment) is a controversial topic. It can impinge on personal liberty and the right to choose, and carry the risk of abuse for political, social, and other reasons; yet it can potentially prevent harm to self and others, and assist some people in attaining their right to healthcare when they may be unable to decide in their own interests. Because of this it is a concern of medical ethics.
All human rights oriented mental health laws require proof of the presence of a mental disorder as defined by internationally accepted standards, but the type and severity of disorder that counts can vary in different jurisdictions. The two most often used grounds for involuntary admission are said to be serious likelihood of immediate or imminent danger to self or others, and the need for treatment. Applications for someone to be involuntarily admitted usually come from a mental health practitioner, a family member, a close relative, or a guardian. Human-rights-oriented laws usually stipulate that independent medical practitioners or other accredited mental health practitioners must examine the patient separately and that there should be regular, time-bound review by an independent review body. The individual should also have personal access to independent advocacy.
For involuntary treatment to be administered (by force if necessary), it should be shown that an individual lacks the mental capacity for informed consent (i.e. to understand treatment information and its implications, and therefore be able to make an informed choice to either accept or refuse). Legal challenges in some areas have resulted in supreme court decisions that a person does not have to agree with a psychiatrist's characterization of the issues as constituting an "illness", nor agree with a psychiatrist's conviction in medication, but only recognize the issues and the information about treatment options.
Proxy consent (also known as surrogate or substituted decision-making) may be transferred to a personal representative, a family member, or a legally appointed guardian. Moreover, patients may be able to make, when they are considered well, an advance directive stipulating how they wish to be treated should they be deemed to lack mental capacity in the future. The right to supported decision-making, where a person is helped to understand and choose treatment options before they can be declared to lack capacity, may also be included in the legislation. There should at the very least be shared decision-making as far as possible. Involuntary treatment laws are increasingly extended to those living in the community, for example outpatient commitment laws (known by different names) are used in New Zealand, Australia, the United Kingdom, and most of the United States.
The World Health Organization reports that in many instances national mental health legislation takes away the rights of persons with mental disorders rather than protecting rights, and is often outdated. In 1991, the United Nations adopted the Principles for the Protection of Persons with Mental Illness and the Improvement of Mental Health Care, which established minimum human rights standards of practice in the mental health field. In 2006, the UN formally agreed the Convention on the Rights of Persons with Disabilities to protect and enhance the rights and opportunities of disabled people, including those with psychiatric disabilities.
The term insanity, sometimes used colloquially as a synonym for mental illness, is often used technically as a legal term. The insanity defense may be used in a legal trial (known as the mental disorder defence in some countries).
Perception and discrimination
Further information: Schizophrenogenic parents, Refrigerator mother, and Mentalism (discrimination)
Stigma
The social stigma associated with mental disorders is a widespread problem. The US Surgeon General stated in 1999 that: "Powerful and pervasive, stigma prevents people from acknowledging their own mental health problems, much less disclosing them to others." Additionally, researcher Wulf Rössler in 2016, in his article, "The Stigma of Mental Disorders" stated
"For millennia, society did not treat persons suffering from depression, autism, schizophrenia and other mental illnesses much better than slaves or criminals: they were imprisoned, tortured or killed".
In the United States, racial and ethnic minorities are more likely to experience mental health disorders often due to low socioeconomic status, and discrimination. In Taiwan, those with mental disorders are subject to general public's misperception that the root causes of the mental disorders are "over-thinking", "having a lot of time and nothing better to do", "stagnant", "not serious in life", "not paying enough attention to the real life affairs", "mentally weak", "refusing to be resilient", "turning back to perfectionistic strivings", "not bravery" and so forth.
Employment discrimination is reported to play a significant part in the high rate of unemployment among those with a diagnosis of mental illness. An Australian study found that having a psychiatric disability is a bigger barrier to employment than a physical disability. The mentally ill are stigmatized in Chinese society and can not legally marry.
Efforts are being undertaken worldwide to eliminate the stigma of mental illness, although the methods and outcomes used have sometimes been criticized.
Media and general public
Main article: Mental disorders in art and literature
Media coverage of mental illness comprises predominantly negative and pejorative depictions, for example, of incompetence, violence or criminality, with far less coverage of positive issues such as accomplishments or human rights issues. Such negative depictions, including in children's cartoons, are thought to contribute to stigma and negative attitudes in the public and in those with mental health problems themselves, although more sensitive or serious cinematic portrayals have increased in prevalence.
In the United States, the Carter Center has created fellowships for journalists in South Africa, the U.S., and Romania, to enable reporters to research and write stories on mental health topics. Former US First Lady Rosalynn Carter began the fellowships not only to train reporters in how to sensitively and accurately discuss mental health and mental illness, but also to increase the number of stories on these topics in the news media. There is also a World Mental Health Day, which in the United States and Canada falls within a Mental Illness Awareness Week.
The general public have been found to hold a strong stereotype of dangerousness and desire for social distance from individuals described as mentally ill. A US national survey found that a higher percentage of people rate individuals described as displaying the characteristics of a mental disorder as "likely to do something violent to others", compared to the percentage of people who are rating individuals described as being troubled. In the article, "Discrimination Against People with a Mental Health Diagnosis: Qualitative Analysis of Reported Experiences," an individual who has a mental disorder, revealed that, "If people don't know me and don't know about the problems, they'll talk to me quite happily. Once they've seen the problems or someone's told them about me, they tend to be a bit more wary." In addition, in the article,"Stigma and its Impact on Help-Seeking for Mental Disorders: What Do We Know?" by George Schomerus and Matthias Angermeyer, it is affirmed that "Family doctors and psychiatrists have more pessimistic views about the outcomes for mental illnesses than the general public (Jorm et al.,1999), and mental health professionals hold more negative stereotypes about mentally ill patients, but, reassuringly, they are less accepting of restrictions towards them."
Recent depictions in media have included leading characters successfully living with and managing a mental illness, including in bipolar disorder in Homeland (2011) and post-traumatic stress disorder in Iron Man 3 (2013).
Violence
Despite public or media opinion, national studies have indicated that severe mental illness does not independently predict future violent behavior, on average, and is not a leading cause of violence in society. There is a statistical association with various factors that do relate to violence (in anyone), such as substance use and various personal, social, and economic factors. A 2015 review found that in the United States, about 4% of violence is attributable to people diagnosed with mental illness, and a 2014 study found that 7.5% of crimes committed by mentally ill people were directly related to the symptoms of their mental illness. The majority of people with serious mental illness are never violent.
In fact, findings consistently indicate that it is many times more likely that people diagnosed with a serious mental illness living in the community will be the victims rather than the perpetrators of violence. In a study of individuals diagnosed with "severe mental illness" living in a US inner-city area, a quarter were found to have been victims of at least one violent crime over the course of a year, a proportion eleven times higher than the inner-city average, and higher in every category of crime including violent assaults and theft. People with a diagnosis may find it more difficult to secure prosecutions, however, due in part to prejudice and being seen as less credible.
However, there are some specific diagnoses, such as childhood conduct disorder or adult antisocial personality disorder or psychopathy, which are defined by, or are inherently associated with, conduct problems and violence. There are conflicting findings about the extent to which certain specific symptoms, notably some kinds of psychosis (hallucinations or delusions) that can occur in disorders such as schizophrenia, delusional disorder or mood disorder, are linked to an increased risk of serious violence on average. The mediating factors of violent acts, however, are most consistently found to be mainly socio-demographic and socio-economic factors such as being young, male, of lower socioeconomic status and, in particular, substance use (including alcohol use) to which some people may be particularly vulnerable.
High-profile cases have led to fears that serious crimes, such as homicide, have increased due to deinstitutionalization, but the evidence does not support this conclusion. Violence that does occur in relation to mental disorder (against the mentally ill or by the mentally ill) typically occurs in the context of complex social interactions, often in a family setting rather than between strangers. It is also an issue in health care settings and the wider community.
Mental health
Main article: Mental health
The recognition and understanding of mental health conditions have changed over time and across cultures and there are still variations in definition, assessment, and classification, although standard guideline criteria are widely used. In many cases, there appears to be a continuum between mental health and mental illness, making diagnosis complex. According to the World Health Organization, over a third of people in most countries report problems at some time in their life which meet the criteria for diagnosis of one or more of the common types of mental disorder. Corey M Keyes has created a two continua model of mental illness and health which holds that both are related, but distinct dimensions: one continuum indicates the presence or absence of mental health, the other the presence or absence of mental illness. For example, people with optimal mental health can also have a mental illness, and people who have no mental illness can also have poor mental health.
Other animals
Main article: Animal psychopathology
Psychopathology in non-human primates has been studied since the mid-20th century. Over 20 behavioral patterns in captive chimpanzees have been documented as (statistically) abnormal for frequency, severity or oddness—some of which have also been observed in the wild. Captive great apes show gross behavioral abnormalities such as stereotypy of movements, self-mutilation, disturbed emotional reactions (mainly fear or aggression) towards companions, lack of species-typical communications, and generalized learned helplessness. In some cases such behaviors are hypothesized to be equivalent to symptoms associated with psychiatric disorders in humans such as depression, anxiety disorders, eating disorders and post-traumatic stress disorder. Concepts of antisocial, borderline and schizoid personality disorders have also been applied to non-human great apes.
The risk of anthropomorphism is often raised concerning such comparisons, and assessment of non-human animals cannot incorporate evidence from linguistic communication. However, available evidence may range from nonverbal behaviors—including physiological responses and homologous facial displays and acoustic utterances—to neurochemical studies. It is pointed out that human psychiatric classification is often based on statistical description and judgment of behaviors (especially when speech or language is impaired) and that the use of verbal self-report is itself problematic and unreliable.
Psychopathology has generally been traced, at least in captivity, to adverse rearing conditions such as early separation of infants from mothers; early sensory deprivation; and extended periods of social isolation. Studies have also indicated individual variation in temperament, such as sociability or impulsiveness. Particular causes of problems in captivity have included integration of strangers into existing groups and a lack of individual space, in which context some pathological behaviors have also been seen as coping mechanisms. Remedial interventions have included careful individually tailored re-socialization programs, behavior therapy, environment enrichment, and on rare occasions psychiatric drugs. Socialization has been found to work 90% of the time in disturbed chimpanzees, although restoration of functional sexuality and caregiving is often not achieved.
Laboratory researchers sometimes try to develop animal models of human mental disorders, including by inducing or treating symptoms in animals through genetic, neurological, chemical or behavioral manipulation, but this has been criticized on empirical grounds and opposed on animal rights grounds.
See also
Philosophy portalPsychiatry portalPsychology portalSociety portal
50 Signs of Mental Illness
List of mental disorders
Mental illness portrayed in media
Mental disorders in film
Mental illness in fiction
Mental illness in American prisons
Parity of esteem
Psychological evaluation
| biology | 3713 | https://sv.wikipedia.org/wiki/Psykisk%20st%C3%B6rning | Psykisk störning | Psykiska störningar (även psykiska sjukdomar, förr mentalsjukdomar) är ohälsa i psyket, det vill säga i personligheten, emotionerna eller kognitionen, vilka primärt har psykiska orsaker. WHO:s diagnosmanual ICD definierar psykiska sjukdomar och syndrom samt beteendestörningar (F00-F99) som störningar i den psykologiska utvecklingen. Psykiska symtom kan också uppträda till följd av missbruk eller andra kroppsliga sjukdomar. Den allvarligaste graden av psykisk störning, oavsett sjukdom eller orsak, är psykos, där jaget och kognitionen brutit samman; allvarlig psykisk störning innefattar typiskt psykotiska tillstånd.
Begrepp för psykisk störning finns i alla kulturer, och betecknar som regel att någon avviker från vad som anses vara normalt. I FN:s regi arbetar WHO för att förbättra den psykiska hälsan globalt, och därför har deras diagnosmanual ICD-10 en särskild avdelning för psykiska störningar. Med störning menar ICD förekomst av kliniskt igenkänningsbara grupper av symtom eller beteenden som i de flesta fall är förknippade med distress och som inverkar på funktioner i personligheten.
En psykisk störning innebär att en människa mår så dåligt inombords att det orsakar ett signifikant lidande för personen själv, psykiskt, socialt eller ekonomiskt. Störningen klassificeras av ICD-10 utifrån personen själv, huruvida denne uppfattar det som ett lidande, samt hur själv- eller verklighetsuppfattningen förhåller sig till verkligheten. De flesta diagnoser i ICD kräver att om lidandet är befogat så ska inte lidandet innefattas i de psykiska störningarna; så klassificeras därför inte till exempel rädsla i en misshandelrelation som ångest, jagsvaghet efter psykisk misshandel som psykisk störning, sorg över en förlorad anhörig som depression, eller problem till följd av livsstil som tecken på psykisk störning (jämför allostas).
För omgivningen kan en psykisk störning yttra sig på många sätt (vanligen genom att personen beter sig annorlunda), bara märkas ibland (till exempel vid berusning) eller inte märkas alls beroende på arten av lidande och på personens bemästringsförmåga. En del psykiska störningar är av den arten att de alltid är socialt handikappande, påtagliga i beteende eller tal, eller nedsätter funktioner i personligheten. Dock behöver inte sociala funktionsnedsättningar vara psykiska störningar. Utbrändhet, högriskbeteenden ifråga om spel eller sex, tendens till konflikter eller accentuerade personlighetsdrag och dylikt kan visserligen vara orsaker till att personer har problem med livssituationen, men är inte psykiska störningar eftersom orsaken inte är en dysfunktionell personlighet.
Begreppsutredning
Begrepp för ohälsa i psyket har förändrats under tid och det finns flera divergerande synsätt på fenomenet. I direkt anslutning till detta finns också flera benämningar på sådan ohälsa. I äldre utgåvor av ICD talades om "vansinne", vilket ersattes av "störning" och "psykos" med ICD-7 (1955). Från 1975 infördes beteckningen "mentalsjukdom". I den engelska originalversionen av WHO:s ICD-10 (1990), har ordet "sjukdom" (disease, illness) slopats till förmån för "störning" (disorder). Huruvida störningen bör betraktas som en funktionsnedsättning eller handikapp regleras av WHO:s International classification of impairments, disabilities and handicaps från 1980. Socialstyrelsen står för den officiella översättningen av ICD, och har behållit begreppet 'sjukdom' samt ibland översatt disorder med "syndrom".
Ohälsa som motsats till välbefinnande
'Psykisk ohälsa' brukar definieras som frånvaron av psykiskt välbefinnande, och definieras i termer av anhedoni eller funktionsbortfall. Därmed ställs hälsa och ohälsa som två dikotomier mot varandra, ett förhållningssätt som blivit allt mera ifrågasatt; en som saknar psykiskt välbefinnande är inte nödvändigtvis psykiskt sjuk.
U.S. Public Health Service definierade 'psykisk hälsa' 1999 som ett tillstånd av framgångsrik användning av psykiska funktioner, vilket leder till produktiva aktiviteter, givande mellanmänskliga relationer, samt förmågan att anpassa sig till förändringar och hantera motgångar. Den definitionen utgår från DSM:s begrepp för psykisk hälsa respektive ohälsa.
I anslutning till den definitionen av psykisk störning, har den amerikanska forskaren Corey L. M. Keyes uppställt tretton symtom på psykisk hälsa. De första två symtomen kallar han 'hedoni' och de övriga 'positiv funktionalitet'.
Positiv affekt - vid gott humör, lugn och full av liv.
Upplevd lycka och nöjdhet - känner sig nöjd och tillfreds med livet.
Självacceptans - har en positiv bild av sig själv, sitt förflutna och accepterar olika aspekter av självet.
Social acceptans - har en positiv bild av andra samtidigt som personen accepterar olikheter och människors komplexitiet.
Personlighetsutveckling - visar insikt i sin potential och utveckling och öppenhet inför utmaningar.
Social aktualisation - anser att människor och samhället kan utvecklas positivt och växa.
Mening med livet - har målsättningar som ger livet en mening.
Social kontribution - anser sig ha något att tillföra samhället, och att vad man gör är av värde för andra.
Behärskning av omgivningen - klarar av att behärska komplicerade miljöer och kan välja miljö som passar sina behov.
Social koherens - intresserad av samhället eller omgivningen, och anser att samhället är meningsfullt och begripligt.
Autonomi - är självgående med av samhället accepterade eller konventionella uppfattningar, och kan stå emot oönskat grupptryck.
Goda mellanmänskliga relationer - har varma, tillitsfulla och tillfredsställande relationer, och har förmåga till initimitet och empati.
Social integration - har en känsla av att vara en del av samhället, och att samhället är ett stöd.
Ett liv innehåller psykiskt lidande och smärta, och att reagera på negativa erfarenheter psykiskt är normalt. Gränsen mellan sådant normalt lidande och psykisk ohälsa är diffus, men brukar erkännas som signifikant för när ett psykiskt tillstånd har övergått från att vara normalt till att ha blivit patologiskt och därför behöver vård. Ett vanligt kriterium på skillnaden, är att tillståndet är patologiskt när lidandet självt innebär ett lidande.
Sjukdom kontra störning
Att benämna ett psykiskt tillstånd "sjukdom", innebär enligt Jan-Otto Ottoson och Hans Ottosson att härleda tillståndet till organiska tillstånd, till exempel i kroppskonstitution, genetiska anlag, hjärnan eller endokrina systemet. Det psykiska tillståndet och dess behandling faller på psykiatrins bord, eftersom det därmed enligt Ottosson och Ottosson betraktas medicinskt. Många psykiska symtom kan förstås och förklaras utifrån sådana organiska tillstånd, dock inte alla. De förändringar som inte kan förklaras organiskt, kan som regel förklaras psykosocialt eller utifrån den drabbades personlighet. Att de senare symtomen inte kan förklaras organiskt, innebär inte enligt Ottosson och Ottosson att en organisk förklaring saknas - de menar snarare att den kan ännu vara okänd. I stället för att konstruera en dikotomi mellan psykiska tillstånd efter organisk genes, psykogenes och endogenes, anser Ottosson och Ottosson att patologiska psykiska tillstånd kollektivt bör kallas "störningar", eftersom de menar att den beteckningen innefattar både rena medicinska sjukdomstillstånd och psykosocialt betingade tillstånd.
DSM-5 och psykobiologisk dysfunktion
Med anledning av revideringen av DSM har utkast inför DSM-5 antagits, där definitionen av psykisk störning (mental disorder) utgår från att dessa orsakas av psykobiologiska dysfunktioner som yttrar sig i ett beteendemässigt eller psykologiskt syndrom eller mönster, samt att detta orsakar betydande lidande för personen eller en funktionsnedsättning. DSM-5 använder uttrycket "psykobiologisk funktion" för att betona sambandet mellan biologi och beteende och psyke. Till psykisk störning förs därför inte ett förväntat lidande som kommer av vanliga livserfarenheter, och heller inte lidande som beror på att personen är på kant med samhället eller har ett avvikande beteende.
DSM-5 ställer också kravet att den psykiska diagnosen ska prövas mot hur personen svarar på behandling och mot andra sätt att säkerställa att rätt bedömning gjorts, samt att diagnos bara bör ställas om det underlättar förståelsen och behandlingen. Diagnoser ska inte ses som absoluta eller klart avgränsade begrepp, eller som fullständiga beskrivningar över alla tänkbara tillstånd.
P-faktorn
Genom faktoranalys har Caspi et al. kommit fram till att psykopatologiska symtom bäst kan förklaras av en dimension - mer eller mindre psykisk ohälsa. Detta skiljer sig från antaganden som gör det möjligt att dela in psykiska störningar i olika diagnoser. Högre p-faktor är associerat till högre livsnedsättning, värre utvecklingshistoria och mer utsatt hjärnfunktion tidigare i livet. P-faktorn förklarar varför det är så svårt att hitta orsaker, biomarkörer och behandlingar med specificitet till enskilda psykiska störningar.
Orsaker och bot
Psykiska störningar, som inte beror på missbruk eller kroppslig sjukdom, brukar ofta förklaras som uppstå i samspelet mellan personen och dess omgivning. Exkludering från sin grupp, isolering, kärlekslösa familjerelationer, och stress är faktorer som ofta ligger bakom psykisk ohälsa. Ofta finns en ärftlig predisposition, att somliga är mer sårbara än andra. Psykisk ohälsa brukar förklaras med stress - sårbarhetsmodellen, som tar fasta på både medfödda och förvärvade sårbarheter som orsak till utbrott av störningar.
Det finns ett samband mellan psykisk ohälsa och fattigdom, något som har förklarats olika av olika forskare. Antingen har ohälsan förklarats orsaka fattigdom, eller så har fattigdomen setts som en etiologi till ohälsan. En brittisk studie har visat att människor med ekonomiska skulder ofta har psykisk ohälsa. Under årtionden har studier gjorts angående förhållandet mellan kroppsvikt (BMI) och psykisk ohälsa. För de flesta psykiska störningarna finns ett samband med övervikt och fetma (affektiva störningar, drogmissbruk, ångeststörningar, personlighetsstörningar). Diagnoser som generaliserat ångestsyndrom och vissa fobier förekommer lika ofta i alla viktkategorier. Undervikt hänger samman med ett fåtal fobier och maniska episoder, och har negativt samband med flera ångeststörningar och personlighetsstörningar. Omedicinerade personer med schizofreni eller schizofrenilikkande sjukdomar har dock inte annorlunda BMI än befolkningen i övrigt.
Medicinering av psykiska störningar utgår från hur störningarna yttrar sig i hjärnan, där till exempel antipsykotisk medicin blockerar dopamin, antidepressiv medicin ökar serotonin, och lugnande medicin kan innehålla ämnen som påminner om opioidpeptider eller påverkar GABA.
Vid behandling av psykiska störningar ingår ofta psykoterapi. Varianterna innefattar bl.a. psykoanalys, som försöker härleda störningarna till trauman och därefter finna metoder att befria personen från traumat, samt kognitiv beteendeterapi, som i stället försöker förändra beteendet på den sjuke. Det finns flera andra terapier, som chockterapi och desensibilisering.
Juridiskt ramverk
Termen "psykisk störning" infördes i och med nuvarande tvångslagstiftning, Lag om psykiatrisk tvångsvård (LPT) 1991/92. Psykiska störningar inom specialiteten psykiatri avser personlighetsstörningar som skiljs från psykiska sjukdomstillstånd. Intresseföreningarna RSMH och Schizofreniförbundet förespråkar dock termen psykisk funktionsnedsättning som de uppfattar som mindre stigmatiserande. Denna term användes bland annat i samband med Psykiatrireformen och i den svenska lagstiftningen, se LSS.
Vid brott skiljs psykiska störningar från psykiska störningar i juridisk bemärkelse, allvarlig psykisk störning. För att en brottsling ska vara psykiskt sjuk i juridisk bemärkelse måste en psykos eller liknande vara så allvarlig att personen inte kan anses ha rättshandlingsförmåga.
Typer av psykiska störningar
De psykiska sjukdomarna och störningarna klassificeras och avgränsas i vedertagna diagnossystem som DSM-IV (Diagnostic Statistical Manual of Mental Disorders, Fourth Revision) och ICD-10 (International Classification of Diseases and Related Health problems, Tenth Revision). ICD-10 ersatte 1990 föregångaren ICD 9 som användes från 1975. ICD utges av WHO och är avsedd som internationell basklassifikation för allmänt medicinskt-statistiskt bruk. I länder utanför USA används ofta DSM vid det diagnostiska arbetet, men den internationella diagnosmanualen ICD-10 är den som officiellt används. I till exempel Sverige får inte DSM användas för diagnos, utan psykiska störningar ska enligt beslut från Socialstyrelsen anges enligt ICD-10.
Psykiska symtom kan yttra sig kognitivt, neurologiskt och beteendemässigt (se vidare neurologiska manifestationer, kognitiva symtom och beteendesymtom). Psykiatriska manualer som ICD-10 och DSM-IV sammanställer grupper av sådana symtom till psykiatriska diagnoser. Det förekommer kulturspecifika psykiska störningar, vilka förmodas bero på kulturspecifika förutsättningar. Sådana brukar dock diagnosticeras enligt vedertagna, internationella standarder.
Funktionella, organiska och drogutlösta störningar
Huvudartiklar: Organiska psykiska störningar och Drogutlösta psykiska störningar
Med psykisk störning förstås i första hand störningar som har sin grund i psyket. Psykiska störningar kan dock uppkomma till följd av droger eller fysiska sjukdomar. Beroende på huvudorsak, kan den psykiska störningen ses som primär, vilket kallas funktionell, eller som ett symtom på droger eller sjukdomar. Psykisk ohälsa förklaras med stress - sårbarhetsmodellen, vilket innebär att psykiska symtom kan ha flera orsaker (vara multifaktorella).
I ICD-10 klassificeras organiska psykiska störningar i F00-F09. För att klassificeras som en organisk psykisk störning, krävs att personen är diagnosticerad med en kroppslig sjukdom som man vet påverkar psyket. Dit hör endokrina sjukdomar, hjärnskador, åldersbetingad neurodegenation samt vissa infektionssjukdomar (till exempel HIV). Organiska psykiska störningar innefattar sådana där framför allt minnet är påverkat (demens), som ger vanföreställningar (delirium), psykoser (organiska psykoser), affektiva störningar och personlighetsförändringar (organisk personlighetsstörning). Neurodegeneration ger ofta demens, vilket sällan kräver någon större utredning för att konstatera. Andra organiska psykiska störningar kan dock vara svåra att skilja från funktionella utan blod- eller urinprov eller undersökning av hjärnan. Praktiskt taget alla psykiska störningar kan bero på organiska sjukdomar, och finns i ICD-10:s FO6 (vanföreställningssyndrom, schizofreni, bipolär sjukdom, ångeststörningar, depressioner, personlighetsstörningar, dissociativa störningar etc).
Vissa normala kroppsliga förändringar kan också ge psykiska symtom. Premenstruellt syndrom och premenstruell dysforisk störning är tämligen vanliga psykiska besvär. Vid förlossning kan kvinnor drabbas av postpartumdepression och postpartumpsykos.
Drogutlösta psykiska störningar klassificeras särskilt i ICD-10 under F10-F19. Dit hör substansmissbruk, oönskad psykisk påverkan efter alkohol, narkotika, tobak och koffein, liksom beroende av sådana psykoaktiva substanser. Akut drogförgiftning ger ofta psykisk påverkan, liksom långvarigt missbruk, avvänjning efter ett drogberoende, med mera. Drogutlösta psykiska störningar kan ofta uppkomma i kombination med en smärre sjukdom eller en psykisk svaghet (dubbeldiagnos).
Faser av sjukdomarna
Innan en psykisk störning bryter ut drabbas personen som regel av en fas av prodrom, vilket vid psykoser kallas prepsykos. Första tiden under sjukdomen kallas akut fas. Om störningen blir bestående kan personen vänja sig vid sina symtom, men under den akuta fasen, vid första insjuknandet, förekommer ofta ångest för de nya symtomen.
Störningens sjukdomsförlopp kan ofta antyda vad störningen har för orsak, om den är funktionell, drogutlöst eller beror på en fysisk sjukdom eller skada.
Kroniska psykiska störningar kan uppträda på olika sätt. En del störningar påverkar personen ständigt, i varierande grad. Andra störningar, i synnerhet bipolär sjukdom, uppkommer i skov. Störningar som botats kan återkomma, vilket kallas recidiv. En tillfällig förbättring vid kroniska sjukdomar, remission, kan vara fullständig eller ofullständig när vissa symtom kvarstår.
Typer av diagnoser
Standardiserade diagnosmanualer som ICD, brukar gruppera störningar först och främst efter vad i psyket som är mest påverkat av ohälsan. Vid affektiva störningar är det framför allt känslorna och stämningsläget som är påverkade. Tillståndet kan vara konstant som vid depressioner eller förändras i cykler, som vid bipolär sjukdom. Medvetandet och perceptionen är ur funktion vid dissociativa störningar, dock utan att nödvändigtvis fullständigt ha brutit samman som vid psykos. I gruppen somatoforma störningar är självuppfattningen, och i synnerhet kroppsuppfattningen störd, något som också antas orsaka ätstörningar.
En personlighetsstörning kan ses som en psykologisk störning av utvecklingen där ogynnsamma miljöfaktorer, ofta i hemmet, haft betydelse för störningens uppkomst. Enligt nedanstående klassifikationssystem får personer under 18 års ålder inte erhålla personlighetsstörningsdiagnos. Personlighetsstörningarna är ofta livslånga, liksom många psykossjukdomar är.
Till psykiska störningar hör också mer eller mindre tillfälliga störningar, såsom sömnproblem. Kvarstående men efter trauman kännetecknar diagnoser som fobier, posttraumatiskt stressyndrom och ångeststörningar, med kvarvarande, obefogad rädsla och förhöjd vaksamhet. Ångest är också en viktig komponent i tvångssyndrom. Tvångssyndrom kännetecknas av tvångsmässigt beteende, vilket också kännetecknar impulskontrollstörningar som kleptomani och spelberoende. Vid de senare fallen sammanhänger handlingarna dock med lyckokänslor, "kickar".
Diagnoser som utvecklas i ung ålder brukar skiljas från de som utvecklas i vuxen ålder, eftersom förutsättningarna är annorlunda. Till denna grupp hör psykisk utvecklingsstörning, autismspektrumtillstånd samt ADHD.
Klassifikation av psykiska störningar
Källa: Nationalencyklopedin
Se även
Stress-sårbarhetsmodellen
Psykopatologi
Psykoterapi
Funktionsnedsättning
Koder i DSM-IV
Stress
Autismspektrumstörning
Referenser
Noter
Källförteckning
ICD-10
DSM-IV
The ICD-10 Classification of Mental and Behavioural Disorders, Diagnostic criteria for research, World Health Organization Geneva 1993
The ICD-10 Classification of Mental and Behavioural Disorders, Clinical descriptions and diagnostic guidelines
DSM-5
Externa länkar
NIMH.NIH.gov - National Institute of Mental Health (USA)
NIMHE.org.uk - National Institute for Mental Health (Storbritannien)
Wikipedia:Basartiklar | swedish | 0.462234 |
mechanism_mentally_tired/Mentalization.txt | In psychology, mentalization is the ability to understand the mental state – of oneself or others – that underlies overt behaviour.
Mentalization can be seen as a form of imaginative mental activity that lets us perceive and interpret human behaviour in terms of intentional mental states (e.g., needs, desires, feelings, beliefs, goals, purposes, and reasons). It is sometimes described as "understanding misunderstanding." Another term that David Wallin has used for mentalization is "Thinking about thinking". Mentalization can occur either automatically or consciously. Mentalization ability, or mentalizing, is weakened by intense emotion.
While the broader concept of theory of mind has been explored at least since Descartes, the specific term 'mentalization' emerged in psychoanalytic literature in the late 1960s, and became empirically tested in 1983 when Heinz Wimmer and Josef Perner ran the first experiment to investigate when children can understand false belief, inspired by Daniel Dennett's interpretation of a Punch and Judy scene.
The field diversified in the early 1990s when Simon Baron-Cohen and Uta Frith, building on the Wimmer and Perner study, and others merged it with research on the psychological and biological mechanisms underlying autism and schizophrenia. Concomitantly, Peter Fonagy and colleagues applied it to developmental psychopathology in the context of attachment relationships gone awry. More recently, several child mental health researchers such as Arietta Slade, John Grienenberger, Alicia Lieberman, Daniel Schechter, and Susan Coates have applied mentalization both to research on parenting and to clinical interventions with parents, infants, and young children.
Mentalization has implications for attachment theory and self-development. According to Peter Fonagy, individuals with disorganized attachment style (e.g., due to physical, psychological, or sexual abuse) can have greater difficulty developing the ability to mentalize. Attachment history partially determines the strength of mentalizing capacity of individuals. Securely attached individuals tend to have had a primary caregiver that has more complex and sophisticated mentalizing abilities. As a consequence, these children possess more robust capacities to represent the states of their own and other people's minds. Early childhood exposure to mentalization can protect the individual from psychosocial adversity. This early childhood exposure to genuine parental mentalization fosters development of mentalizing capabilities in the child themselves. There is also suggestion that genuine parental mentalization is beneficial to child learning; when a child feels they are being viewed as an intentional agent, they feel contingently responded to, which promotes epistemic trust and triggers learning in the form of natural pedagogy - this increases the quality of learning in the child. This theory needs further empirical support.
Mentalization or better mentalizing, has a number of different facets which can be measured with various methods. A prominent method of assessment of Parental Mentalization is the Parental Development Interview (PDI), a 45-question semi-structured interview, investigating parents’ representations of their children, themselves as parents, and their relationships with their children. An efficient self-report measure of Parental Mentalization is the Parental Reflective Functioning Questionnaire (PRFQ) created by Patrick Luyten and colleagues. The PRFQ is a brief, multidimensional assessment of parental reflective functioning (mentalization), aimed to be easy to administer to parents in a wide range of socioeconomic populations. The PRFQ is recommended for use as a screening tool for studies with large populations and does not aim to replace more comprehensive measures, such as the PDI or observer-based measures.
According to the American Psychiatric Association's Handbook of Mentalizing in Mental Health Practice, mentalization takes place along a series of four parameters or dimensions: Automatic/Controlled, Self/Other, Inner/Outer, and Cognitive/Affective.
Each dimension can be exercised in either a balanced or unbalanced way, while effective mentalization also requires a balanced perspective across all four dimensions.
Background[edit]
While the broader concept of theory of mind has been explored at least since Descartes, the specific term 'mentalization' emerged in psychoanalytic literature in the late 1960s, and became empirically tested in 1983 when Heinz Wimmer and Josef Perner ran the first experiment to investigate when children can understand false belief, inspired by Daniel Dennett's interpretation of a Punch and Judy scene.
The field diversified in the early 1990s when Simon Baron-Cohen and Uta Frith, building on the Wimmer and Perner study, and others merged it with research on the psychological and biological mechanisms underlying autism and schizophrenia. Concomitantly, Peter Fonagy and colleagues applied it to developmental psychopathology in the context of attachment relationships gone awry. More recently, several child mental health researchers such as Arietta Slade, John Grienenberger, Alicia Lieberman, Daniel Schechter, and Susan Coates have applied mentalization both to research on parenting and to clinical interventions with parents, infants, and young children.
Implications[edit]
Mentalization has implications for attachment theory and self-development. According to Peter Fonagy, individuals with disorganized attachment style (e.g., due to physical, psychological, or sexual abuse) can have greater difficulty developing the ability to mentalize. Attachment history partially determines the strength of mentalizing capacity of individuals. Securely attached individuals tend to have had a primary caregiver that has more complex and sophisticated mentalizing abilities. As a consequence, these children possess more robust capacities to represent the states of their own and other people's minds. Early childhood exposure to mentalization can protect the individual from psychosocial adversity. This early childhood exposure to genuine parental mentalization fosters development of mentalizing capabilities in the child themselves. There is also suggestion that genuine parental mentalization is beneficial to child learning; when a child feels they are being viewed as an intentional agent, they feel contingently responded to, which promotes epistemic trust and triggers learning in the form of natural pedagogy - this increases the quality of learning in the child. This theory needs further empirical support.
Research[edit]
Mentalization or better mentalizing, has a number of different facets which can be measured with various methods. A prominent method of assessment of Parental Mentalization is the Parental Development Interview (PDI), a 45-question semi-structured interview, investigating parents’ representations of their children, themselves as parents, and their relationships with their children. An efficient self-report measure of Parental Mentalization is the Parental Reflective Functioning Questionnaire (PRFQ) created by Patrick Luyten and colleagues. The PRFQ is a brief, multidimensional assessment of parental reflective functioning (mentalization), aimed to be easy to administer to parents in a wide range of socioeconomic populations. The PRFQ is recommended for use as a screening tool for studies with large populations and does not aim to replace more comprehensive measures, such as the PDI or observer-based measures.
Fourfold dimensions[edit]
According to the American Psychiatric Association's Handbook of Mentalizing in Mental Health Practice, mentalization takes place along a series of four parameters or dimensions: Automatic/Controlled, Self/Other, Inner/Outer, and Cognitive/Affective.
Each dimension can be exercised in either a balanced or unbalanced way, while effective mentalization also requires a balanced perspective across all four dimensions.
Automatic/Controlled. Automatic (or implicit) mentalizing is a fast-processing unreflective process, calling for little conscious effort or input; whereas controlled mentalization (explicit) is slow, effortful, and demanding of full awareness. In a balanced personality, shifts from automatic to controlled smoothly occur when misunderstandings arise in a conversation or social setting, to put things right. Inability to shift from automatic mentalization can lead to a simplistic, one-sided view of the world, especially when emotions run high; while conversely inability to leave controlled mentalization leaves one trapped in a 'heavy', endlessly ruminative thought-mode.
Self/Other involves the ability to mentalize about one's own state of mind, as well as about that of another. Lack of balance means an overemphasis on either self or other.
Inner/Outer: Here problems can arise from an over-emphasis on external conditions, and a neglect of one's own feelings and experience.
Cognitive/Affective are in balance when both dimensions are engaged, as opposed to either an excessive certainty about one's own one-sided ideas, or an overwhelming of thought by floods of emotion.
See also[edit]
AMBIT (Adolescent Mentalization-Based Integrative Treatment)
Mentalization based treatment
Metacognition
Psychic equivalence
Social cognition | biology | 1205924 | https://sv.wikipedia.org/wiki/Mentalisering | Mentalisering | Mentalisering är ett psykologiskt begrepp som skapats av de engelska psykiatrikerna och forskarna Anthony Bateman och Peter Fonagy. Mentalisering handlar om att ha mentala tillstånd i sitt medvetande eller på engelska "holding mind in mind". Mestadels kretsar mentalisering kring att tänka och känna om sig själv och om andra. Mentalisering handlar också om att förstå mentala tillstånds natur, till exempel att de är "flyktiga" och att de kan ändras snabbt eller att det aldrig går att veta exakt vad en annan person känner och tänker.
Begreppet mentalisering är nära förknippat med diagnosen borderline personlighetsstörning, då teorin uppstått under arbetet att ta fram en terapeutisk metod för diagnosen. Teorin går ut på att alla ibland tappar mentaliseringsförmågan och hamnar i mentaliseringssammanbrott. Problemet vid borderline personlighetssyndrom är att mentaliseringsförmågan förloras snabbare samt att det tar längre tid att återhämta sig från ett mentaliseringssammanbrott. Förmågan att mentalisera, i trygga förhållanden, är dock inte nedsatt hos personer med borderline personlighetssyndrom, enligt teorin. Terapimetoden som arbetet mynnade ut i kallas för mentaliseringsbaserad terapi (MBT).
Mentalisering delas upp i fyra olika dimensioner. Dessa dimensioner ska ses som spektrum, där mentalisering uppstår om man kan röra sig förhållandevis flexibelt i dimensionen. De fyra dimensionerna är:
Automatisk – kontrollerad
Själv – andra
Internt fokus – externt fokus
Kognition – affekt
Tre andra viktiga begrepp inom mentaliseringsteorin är tre förmentaliserande lägen, i vilka man kan hamna i vid mentaliseringssammanbrott. Dessa tre lägen är:
Teleologisk hållning
Psykisk ekvivalens
Låtsas-läge
Vid teleologisk hållning upplevs erfarenheter endast giltiga när konsekvenser är synliga. Psykisk ekvivalens handlar om att inre verklighet likställs med den yttre. I låtsas-läge är istället den yttre verkligheten helt skild från den inre.
Se även
Psykoterapi
Referenser
Termer inom psykologi | swedish | 0.677195 |
mechanism_mentally_tired/Fatigue.txt | Fatigue describes a state of tiredness (which is not sleepiness) or exhaustion. In general usage, fatigue often follows prolonged physical or mental activity. When fatigue occurs independently of physical or mental exertion, or does not resolve after rest or sleep, it may have other causes, such as a medical condition.
Fatigue (in a medical context) is complex and its cause is often unknown. Fatigue is associated with a wide variety of conditions including autoimmune disease, organ failure, chronic pain conditions, mood disorders, heart disease, infectious diseases, and post-infectious-disease states.
Fatigue (in the general usage sense of normal tiredness) can include both physical and mental fatigue. Physical fatigue results from muscle fatigue brought about by intense physical activity. Mental fatigue results from prolonged periods of cognitive activity which impairs cognitive ability. Mental fatigue can manifest as sleepiness, lethargy, or directed attention fatigue. Mental fatigue can also impair physical performance.
Definition[edit]
Fatigue in a medical context is used to cover experiences of low energy that are not caused by normal life.
Distinguishing features of such fatigue include
not linking fatigue to an obvious cause,
lack of improvement with rest,
variability in severity,
unpredictability, and
fatigue being more profound/overwhelming, and having extensive impact on daily living.
Correlation of the quantum of fatigue with the severity of an underlying disease is often not present.
Fatigue is multi-faceted and broadly defined, which makes understanding the cause of its manifestations especially difficult in conditions with diverse pathology including autoimmune diseases.
A 2021 review proposed a definition for fatigue as a starting point for discussion: "A multi-dimensional phenomenon in which the biophysiological, cognitive, motivational and emotional state of the body is affected resulting in significant impairment of the individual's ability to function in their normal capacity".
Another definition is that fatigue is "a significant subjective sensation of weariness, increasing sense of effort, mismatch between effort expended and actual performance, or exhaustion independent from medications, chronic pain, physical deconditioning, anaemia, respiratory dysfunction, depression, and sleep disorders.".
Terminology[edit]
The use of the term "fatigue" in medical contexts may carry inaccurate connotations from the more general usage of the same word. More accurate terminology may also be needed to cover variants within the umbrella term of fatigue.
Comparison with other terms[edit]
Tiredness[edit]
Tiredness which is a normal result of work, mental stress, anxiety, overstimulation and understimulation, jet lag, active recreation, boredom, or lack of sleep is not considered medical fatigue. This is the tiredness described in MeSH Descriptor Data.
Sleepiness[edit]
Fatigue is generally considered a longer-term condition than sleepiness (somnolence). Although sleepiness can be a symptom of a medical condition, it usually results from lack of restful sleep, or lack of stimulation. Fatigue is often described as an uncomfortable tiredness, whereas sleepiness can be comfortable and inviting.
Classification[edit]
See also: Central nervous system fatigue and Muscle fatigue
By type[edit]
Uni- or multi-dimensional[edit]
It is disputed whether there are different dimensions of fatigue, such as peripheral (muscle) and central (mental) fatigue, or whether fatigue is a uni-dimensional phenomenon that influences different aspects of human life.
A 2021 review considered that different 'types/subsets' of fatigue may exist and that patients normally present with more than one such 'type/subset". These different "types/subsets" of fatigue may be different dimensions of the same symptom, and the relative manifestations of each may depend on the relative contribution of different mechanisms. Inflammation may be the root causal mechanism in many cases.
Physical[edit]
Physical fatigue, or muscle fatigue, is the temporary physical inability of muscles to perform optimally. The onset of muscle fatigue during physical activity is gradual, and depends upon an individual's level of physical fitness – other factors include sleep deprivation and overall health. Physical fatigue can be caused by a lack of energy in the muscle, by a decrease of the efficiency of the neuromuscular junction or by a reduction of the drive originating from the central nervous system, and can be reversed by rest. The central component of fatigue is triggered by an increase of the level of serotonin in the central nervous system. During motor activity, serotonin released in synapses that contact motor neurons promotes muscle contraction. During high level of motor activity, the amount of serotonin released increases and a spillover occurs. Serotonin binds to extrasynaptic receptors located on the axonal initial segment of motor neurons with the result that nerve impulse initiation and thereby muscle contraction are inhibited.
Muscle strength testing can be used to determine the presence of a neuromuscular disease, but cannot determine its cause. Additional testing, such as electromyography, can provide diagnostic information, but information gained from muscle strength testing alone is not enough to diagnose most neuromuscular disorders.
Mental[edit]
Mental fatigue is a temporary inability to maintain optimal cognitive performance. The onset of mental fatigue during any cognitive activity is gradual, and depends upon an individual's cognitive ability, and also upon other factors, such as sleep deprivation and overall health.
Mental fatigue has also been shown to decrease physical performance. It can manifest as somnolence, lethargy, directed attention fatigue, or disengagement. Research also suggests that mental fatigue is closely linked to the concept of ego depletion, though the validity of the concept is disputed. For example, one pre-registered study of 686 participants found that after exerting mental effort, people are likely to disengage and become less interested in exerting further effort.
Decreased attention can also be described as a more or less decreased level of consciousness. In any case, this can be dangerous when performing tasks that require constant concentration, such as operating large vehicles. For instance, a person who is sufficiently somnolent may experience microsleep. However, objective cognitive testing can be used to differentiate the neurocognitive deficits of brain disease from those attributable to tiredness.
The perception of mental fatigue is believed to be modulated by the brain's reticular activating system (RAS).
Fatigue impacts a driver's reaction time, awareness of hazards around them and their attention. Drowsy drivers are three times more likely to be involved in a car crash, and being awake over 20 hours is the equivalent of driving with a blood-alcohol concentration level of 0.08%.
Neurological fatigue[edit]
People with multiple sclerosis experience a form of overwhelming tiredness that can occur at any time of the day, for any duration, and that does not necessarily recur in a recognizable pattern for any given patient, referred to as "neurological fatigue", and often as "multiple sclerosis fatigue" or "lassitude".
People with autoimmune diseases including inflammatory rheumatic diseases such as rheumatoid arthritis, psoriatic arthritis and primary Sjögren's syndrome, experience similar fatigue.
By timescale[edit]
Acute[edit]
Acute fatigue is that which is temporary and self-limited. Acute fatigue is most often caused by an infection such as the common cold and can be cognized as one part of the sickness behavior response occurring when the immune system fights an infection.
Other common causes of acute fatigue include depression and chemical causes, such as dehydration, poisoning, low blood sugar, or mineral or vitamin deficiencies.
Prolonged[edit]
Prolonged fatigue is a self-reported, persistent (constant) fatigue lasting at least one month.
Chronic[edit]
Chronic fatigue is a self-reported fatigue lasting at least 6 consecutive months. Chronic fatigue may be either persistent or relapsing. Chronic fatigue is a symptom of many chronic illnesses and of idiopathic chronic fatigue.
By effect[edit]
Fatigue can have significant negative impacts on quality of life. Profound and debilitating fatigue is the most common complaint reported among individuals with autoimmune disease, such as systemic lupus erythematosus, multiple sclerosis, type 1 diabetes, celiac disease, chronic fatigue syndrome, and rheumatoid arthritis.
Fatigue that dissociates by quantum with disease activity represents a large health economic burden and unmet need to patients and to society.
Measurement[edit]
Fatigue is currently measured by many different self-measurement surveys. There is no consensus on best practice, and the existing surveys do not capture the intermittent nature of some forms of fatigue.
Nintendo announced plans for a device to possibly quantitatively measure fatigue in 2014, but the project was stopped in 2016.
Causes[edit]
Undiagnosed[edit]
The cause of fatigue is often undiagnosed.
Idiopathic chronic fatigue[edit]
Main article: Idiopathic chronic fatigue
Idiopathic chronic fatigue is chronic fatigue not caused by a known proximate cause such as a discrete medical condition, drug or alcohol use.
Medications[edit]
Fatigue may be a side effect of certain medications (e.g., lithium salts, ciprofloxacin); beta blockers, which can induce exercise intolerance; and many cancer treatments, particularly chemotherapy and radiotherapy.
Drug use[edit]
Caffeine and alcohol can cause fatigue.
Sleep deprivation[edit]
Sleep deprivation and disruption is associated with subsequent fatigue.
Obesity[edit]
Obesity appears to correlate with greater fatigue incidence.
Psychological stress and conditions[edit]
Depression and adverse life events have been associated with fatigue.
Association with diseases[edit]
Fatigue is often associated with diseases and conditions. Some major categories of conditions that often list fatigue as a symptom include:
Autoimmune diseases, such as celiac disease, lupus, multiple sclerosis, myasthenia gravis, NMOSD, Sjögren's syndrome, spondyloarthropathy and UCTD. This population's primary concern is fatigue.
Anxiety disorders, such as generalized anxiety disorder
Blood disorders, such as anemia and hemochromatosis
Brain injury
Cancer, in which case it is called cancer fatigue
Covid-19; long Covid.
Substance use disorders including alcohol use disorder
Depression and other mental disorders that feature depressed mood
Developmental disorders such as autism spectrum disorder
Eating disorders, which can produce fatigue due to inadequate nutrition
Endocrine diseases or metabolic disorders: diabetes mellitus, hypothyroidism and Addison's disease
Fibromyalgia
Gulf War syndrome
Heart failure
HIV
Inborn errors of metabolism such as fructose malabsorption.
Infectious diseases such as infectious mononucleosis or tuberculosis
Irritable bowel syndrome
Kidney diseases e.g. acute renal failure, chronic renal failure
Leukemia or lymphoma
Liver failure or liver diseases e.g. Hepatitis
Lyme disease
Myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS)
Neurological disorders such as narcolepsy, Parkinson's disease, Postural Orthostatic Tachycardia Syndrome and post-concussion syndrome
Physical trauma and other pain-causing conditions, such as arthritis
Sleep deprivation or sleep disorders, e.g. sleep apnea
Stroke
Thyroid disease such as hypothyroidism
Primary vs. secondary[edit]
In some areas it has been proposed that fatigue be separated into primary fatigue, caused directly by a disease process, and ordinary or secondary fatigue, caused by a range of causes including exertion and also secondary impacts on a person of having a disease (such as disrupted sleep). The ICD definition of fatigue
captures both types of fatigue; it includes fatigue that "occur[s] in the absence of... exertion... as a symptom of health conditions."
Mechanisms[edit]
The mechanisms that cause fatigue are not well understood. Several mechanisms may be in operation within a patient, with the relative contribution of each mechanism differing over time.
Inflammation[edit]
Inflammation distorts neural chemistry, brain function and functional connectivity across a broad range of brain networks, and has been linked to many types of fatigue. Findings implicate neuroinflammation in the etiology of fatigue in autoimmune and related disorders.
Low-grade inflammation may cause an imbalance between energy availability and expenditure. Cytokines are small protein molecules that modulate immune responses and inflammation (as well as other functions) and may have causal roles in fatigue. The inflammation model may have difficulty in explaining the "unpredictability" and "variability" (i.e. appearing intermittently during the day, and not on all days) of the fatigue associated with inflammatory rheumatic diseases and autoimmune diseases (such as multiple sclerosis).
Heat shock proteins[edit]
A small 2016 study found that primary Sjögren's syndrome patients with high fatigue, when compared with those with low fatigue, had significantly higher plasma concentrations of HSP90α, and a tendency to higher concentrations of HSP72.
Reduced brain connectivity[edit]
Fatigue has been correlated with reductions in structural and functional connectivity in the brain. This has included in post-stroke, MS, NMOSD and MOG, and ME/CFS. This was also found for fatigue after brain injury, including a significant linear correlation between self-reported fatigue and brain functional connectivity. (Areas of the brain for which there is evidence of relation to fatigue are the thalamus and middle frontal cortex, fronto-parietal and cingulo-opercular, and default mode network, salience network, and thalamocortical loop areas.)
Prevalence[edit]
Minor dark circles, in addition to a hint of eye bags, a combination which is suggestive of sleep deprivation and/or mental fatigue
2023 guidance stated fatigue prevalence is between 4.3% and 21.9%. Prevalence is higher in women than men.
2021 German study found that fatigue was the main or secondary reason for 10–20% of all consultations with a primary care physician.
Diagnosis[edit]
The overall goal in diagnosis is to identify and rule out any treatable conditions. This is done by considering
the person's medical history, any other symptoms present.
evaluation of the qualities of the fatigue itself. Identifiable patterns to the fatigue can include being more tired at certain times of day, whether fatigue increases throughout the day, and whether fatigue is reduced after taking a nap.
sleep patterns. These can be examined by enquiring about quality of sleep, the emotional state of the person, sleep pattern, and stress level. The amount of sleep, the hours that are set aside for sleep, and the number of times that a person awakens during the night are important. A sleep study may be ordered to rule out a sleep disorder.
depression and other psychological conditions.
substance use disorders, and use of caffeine and alcohol.
poor diet.
lack of physical exercise (which paradoxically increases fatigue).
Further tests may include
blood tests to check for infection or anemia.
urinalysis to look for signs of liver disease or diabetes mellitus.
other tests to check for kidney and liver function, such as a comprehensive metabolic panel. Other tests may be chosen depending on the patient's social history, such as an HIV test or pregnancy test.
A 2009 study concluded about 50% of people who have fatigue receive a diagnosis that could explain the fatigue after a year with the condition. In those people who have a possible diagnosis, musculoskeletal (19.4%) and psychological problems (16.5%) are the most common. Definitive physical conditions were only found in 8.2% of cases.
2023 guidance stated
In the primary care setting, a medical or psychiatric diagnosis is found in at least two-thirds of patients.
The most common diagnoses are viral illness, upper respiratory infection, iron-deficiency anaemia, acute bronchitis, adverse effects of a medical agent in the proper dose, and depression or other mental disorder (such as panic disorder, and somatisation disorder).
The origin of fatigue may be central (brain-derived) or peripheral (usually a neuromuscular origin). It may be attributed to Physical illness, Psychological (e.g., psychiatric disorder), social (e.g., family problems), and physiological factors (e.g., old age), Occupational illness (e.g., workplace stress).
When unexplained, clinically evaluated chronic fatigue can be separated into ME/CFS and idiopathic chronic fatigue.
Treatment and Management[edit]
Management may include the following;
Review of existing medications[edit]
Medications may be evaluated for side effects that contribute to fatigue and the interactions of medications are complex.
Lifestyle changes[edit]
Fatigue may be reduced by reducing obesity, caffeine and alcohol intake, pain and sleep disturbance, and by improving mental well-being.
Medications used to treat fatigue[edit]
The UK NICE recommends consideration of amantadine, modafinil and SSRIs for MS fatigue. Psychostimulants such as methylphenidate, amphetamines, and modafinil have been used in the treatment of fatigue related to depression, and medical illness such as chronic fatigue syndrome and cancer. They have also been used to counteract fatigue in sleep loss and in aviation.
Intermittent fasting[edit]
A small 2022 study found both physical and mental fatigue were significantly reduced after three months of 16:8 intermittent fasting.
Vagus nerve stimulation[edit]
A small study showed possible efficacy of vagus nerve stimulation for fatigue reduction in Sjogren's patients.
Mental health tools[edit]
CBT has been found useful.
Avoidance of body heat[edit]
Fatigue in MS has been linked to relatively high endogenous body temperature.
Possible purposes of fatigue[edit]
Evolutionary purposes[edit]
It has been posited that fatigue had evolutionary benefits in making more of the body's resources available for healing processes, such as immune responses, and in limiting disease spread by tending to reduce social interactions.
Body resource management purposes[edit]
Fatigue has been posited as a bio-psycho-physiological state reflecting the body's overall strategy in resource (energy) management. Fatigue may occur when the body wants to limit resource utilisation ("rationing") in order to conserve energy for a particular need, including a threat. This need may be a current or future anticipated need.
See also[edit]
Acquiescence
Affect
Cancer-related fatigue
Central governor
Chronic stress
Clouding of consciousness
Combat stress reaction
Directed attention fatigue
Effects of fatigue on safety
Feeling
Gaucher's disease
Heat illness
Malaise
Microsleep
Museum fatigue
Presenteeism
Sleep-deprived driving
Pacing (activity management)
Zoom fatigue | biology | 827480 | https://da.wikipedia.org/wiki/Tr%C3%A6thed | Træthed | Træthed (også kaldet udmattelse, sløvhed eller mathed) er en subjektiv følelse, som er adskilt fra svaghed, og som har variende omfang. I modsætning til svaghed så kan træthed mindskes ved perioder med søvn. Træthed kan have fysiske eller psykiske årsager. Fysisk træthed er en forbigående tilstand, hvor en muskel ikke kan fastholde en optimal fysisk præstation, hvilket oftere sker ved intens fysisk aktivitet. Mental træthed er en forbigående nedgang i maksimale kognitive præstationer, som er forårsaget af længerevarende perioder med kognitiv aktivitet. Det kan vise sig ved døsighed, letargi eller direkte opmærksomhedstræthed.
Medicinsk udgør træthed et ikke-specifikt symptom, som der kan være mange forskellige årsager til. Træthed betragtes som et symptom, snarere end et sygdomstegn, fordi det er en subjektiv følelse der reporteres af patienten, snarere end et et objektivt tegn, der kan observeres af andre. Træthed og træthedsfølelse sammenblandes ofte. Mange handicappede har i løbet af en dag en begrænset mængde energi til rådighed, der, når den er udtømt, leder til stor træthed, som beskrevet i sketeorien.
Se også
Søvnighed
Noter
Videre læsning
Byung-Chul Han: Müdigkeitsgesellschaft. Matthes & Seitz, Berlin 2010, . (Philosophical essay about fatigue as a sociological problem and symptom).
Dansk udgave: Træthedssamfundet. Møller, 2012, .
Eksterne henvisninger
Fatigue — Information for Patients, U.S. National Cancer Institute
Information leaflet from mental health charity The Royal College of Psychiatrists
Is exhaustion an actual medical diagnosis? from Slate.com
Symptomer
Mentale processer | danish | 0.526839 |
mechanism_mentally_tired/Directed_attention_fatigue.txt | Directed attention fatigue (DAF) is a neuro-psychological phenomenon that results from overuse of the brain's inhibitory attention mechanisms, which handle incoming distractions while maintaining focus on a specific task. The greatest threat to a given focus of attention is competition from other stimuli that can cause a shift in focus. This is because one maintains focus on a particular thought by inhibiting all potential distractions and not by strengthening that central mental activity. Directed attention fatigue occurs when a particular part of the brain's global inhibitory system is overworked due to the suppression of increasing numbers of stimuli. This temporary condition is not a clinical illness or a personality disorder. It is rather a temporary fatigue of the inhibitory mechanisms in the brain. According to inhibition theory, it is natural for one to alternate between periods of attention and distraction. Although one's efforts may involve very different tasks, each incoming stimulus calls upon the same directed attention mechanism.
Symptoms and signs[edit]
Extreme levels of mental fatigue lead to an inability to perform regular tasks and irritability. Studies that require participants to carry out attention-demanding tasks under conditions of high distraction reveal how unpleasant a mentally fatigued person can be. After exposure to such an experience, individuals are less likely to help someone in need. They are also more aggressive, less tolerant, and less sensitive to socially important cues. Fatigue that is experienced by participants of these kinds of studies is induced by attention-intensive tasks, and the observed effects of such fatigue are correlated with decline in inhibitory control. Signs of Directed Attention Fatigue include temporarily feeling unusually distractible, impatient, forgetful, or cranky when there is no associated illness. In more severe forms, it can lead to bad judgment, apathy, or accidents, and can contribute to increased stress levels.
There are 6 major areas of mental processing that are affected during onset of DAF, which are as follows:
Input — One may experience misperception and miss social cues.
Thinking — One may experience feelings of restlessness, confusion, forgetfulness and/or decreased metacognition.
Behavior — One may experience feelings of impulsiveness and recklessness, and may find that they have a diminished level of threshold between thoughts and actions. One may also act out-of-character.
Executive Functioning — One may experience an inability to plan and make appropriate decisions and may experience impaired judgment ability.
Emotions — One may experience being short-tempered and feelings of unpleasantness.
Social Interactions — One may experience heightened irritability and increased frequency of antisocial feelings.
Overlap of symptoms from the six above categories is highly indicative of Directed Attention Fatigue.
Link to ADHD[edit]
The symptoms of attention deficit hyperactivity disorder closely mirror the symptoms of Directed Attention Fatigue. Like Directed Attention Fatigue, ADHD involves the prefrontal cortex. Specifically, the right prefrontal cortex is less active among children with ADHD. Experimentation has shown that the severity of ADHD symptoms can be correlated to the degree of asymmetry between blood flow in the left and right prefrontal cortex. It is possible that DAF and ADHD involve disruption of the same underlying mechanism, and recent clinical evidence has found that the same treatments used for Directed Attention Fatigue may reduce the symptoms of ADHD in children. However, it is important to note that unlike ADHD, DAF is a temporary condition rather than a clinical disorder.
Cause[edit]
This section does not cite any sources. Please help improve this section by adding citations to reliable sources. Unsourced material may be challenged and removed. (November 2012) (Learn how and when to remove this template message)
The onset of directed attention fatigue can be triggered by a number of activities, all of which involve use of the brain's inhibitory system. Activities that engage this system include multitasking, working in an environment with disruptive background noise or after a lack of sleep, and rapidly changing focus during a prolonged period of attention. DAF can also be brought about by performing concentration-intensive tasks such as puzzle-solving or learning unfamiliar ideas. External factors such as stress resulting from emergencies, exams or work deadlines can also induce DAF. Any illness or injury to the brain that interrupts the brain circuits involved in maintaining attention and inhibiting external stimuli may also contribute to the development of directed attention fatigue.
Anatomy[edit]
Directed attention, or voluntary attention, requires a great deal of concentration and focus, and is employed in tasks such as problem solving. This type of attention employs the inhibitory mechanisms of the brain, which help block incoming stimuli that are unrelated to the task at hand. Several parts of the brain are involved in maintaining directed attention, primarily those located in the frontal lobe and the parietal lobe of the brain. Specifically, the mechanism of directed attention employs the prefrontal cortex (PFC), the anterior cingulate cortex (ACC) and the brain stem's basal ganglia. Some fMRI studies have shown that directed attention involves changes in the anterior cingulate cortex and the lateral prefrontal cortex, perhaps as a consequence of increased connectivity between these two areas. Evidence also suggests that the right inferior frontal cortex (IFC) plays a specialized role in response inhibition. It seems that this region plays a key role in the integration of bottom-up response-related information and facilitates goal-directed behavior.
While these areas of the brain are known to be involved in DAF, their specific molecular mechanisms in the perpetuation of DAF symptoms are not yet known.
Diagnosis[edit]
Differential diagnosis[edit]
The concept of stress is used in many situations that would be described as mental fatigue, but the two concepts are distinctly different. Stress involves preparation for an anticipated event that has been evaluated as being threatening or harmful. Though mental fatigue may well result from stressful circumstances, it also arises out of hard work on a project one enjoys. In such cases, there is no anticipation of threat or harm present but still the result is fatigue. Characteristic of mental fatigue is difficulty focusing. For a mentally fatigued person, paying attention to something uninteresting is burdensome, even though focusing on something of great interest poses no particular challenge. Hence, there are two types of attention, distinguished in terms of the effort involved in their use and their changes in attentional shift:
Involuntary attention refers to attention that requires no effort at all, as when something exciting or interesting happens.
Voluntary attention, or directed attention, refers to attention that requires a great deal of effort, as when something is monotonous or boring.
Treatment[edit]
There are measures that can be taken in order to reduce the impact of DAF. These include reducing the number of distractions present in one's external environment, trying to clear one's mind of any internal distractions and taking short breaks during prolonged periods of focus. Directed attention fatigue can be reduced by getting a sufficient amount of sleep each night, because inhibitory attention chemicals are replenished during sleep.
An aesthetic environment may also serve a restorative function in fostering recovery from mental fatigue. Research has shown that restorative experiences, such as clearing one's head and reflecting on one's life and priorities, may help combat Directed Attention Fatigue. As investigated by attention restoration theory, natural environments, such as forests, mountain landscapes or beaches, appear to be particularly effective for restoring attention, perhaps because they contain a vast amount of diverse, relatively weak stimuli, thus inciting the mind to wander freely while relaxing its strict focus.
Research[edit]
Ongoing research is examining ways in which the incidence of DAF can be decreased, and suggests that exposure to the natural environment may aid in the reduction of DAF symptoms. A number of researchers have investigated Directed Attention Fatigue recently. Leading contributors include Rachel and Stephen Kaplan, environmental psychologists at the University of Michigan. Rachel and Stephen Kaplan are credited with much of the research relating to Directed Attention Fatigue and were the first to discover that extended periods of focused attention can lead to DAF. Their research suggests that Directed Attention Fatigue can be alleviated with exposure to nature. Together, the Kaplans devised the Attention restoration theory (ART), which states that a person is better able to maintain focused directed attention after spending time in the natural environment.
A number of studies have been performed that specifically focus on Directed Attention Fatigue experienced by cancer patients. Such studies suggests that the DAF experienced by cancer patients following surgery improves significantly through outdoor restorative activities for 20 minutes per day.
Similarly, it has been discovered that even brief amounts of time spent on a busy metropolitan street can affect one's ability to maintain focus on a given task. Experimental findings suggest that spending time in a natural environment or even looking at pictures of nature can improve maintenance of directed attention.
The Landscape and Human Health Laboratory (LHHL) has completed studies examining the complex relationship between people and their environment. Researchers here discovered the correlation between Directed Attention Fatigue, irritability and aggression. Their findings suggest that people deprived of the restorative effects of nature display more aggressive qualities. Results also demonstrate that communities with several trees and other natural areas tend to be stronger, safer and house residents with a higher attention level. More recent experimentation done at the LHHL suggests that children possessing attention deficits increase their attention level after walking outdoors.
According to this and much other research that has been performed regarding directed attention fatigue, the brain's ability to maintain directed attention is heightened after exposure to nature.
See also[edit]
Attention
Attention span
Environmental psychology
Ego depletion
Human multitasking | biology | 1047381 | https://da.wikipedia.org/wiki/Endogen%20og%20eksogen%20opm%C3%A6rksomhed | Endogen og eksogen opmærksomhed | Endogen opmærksomhed er en målrettet opmærksomhed, mens eksogen opmærksomhed er en stimulusdreven opmærksomhed.
Endogen eller top-down opmærksomhed er bevidst rettes mod noget. Det ses den ved, at et individ fokuserer på et objekt eller en handling på baggrund af individets mål og intentioner. Et eksempel er, at man skal følge med i en film. Her er man opmærksom på, hvad der foregår i filmen i egen interesse. Man vælger altså frivilligt at kontrollere, hvad man er opmærksom på.
Eksogen (eller bottom-up) opmærksomhed rettes mod noget i det øjeblik man modtager en stimulus. Da eksogen opmærksomhed er en stimulusdreven, ses den ved at et individs opmærksomhed ufrivilligt bliver rettet mod den pludselige stimulus. Et eksempel er, at det ofte er den type opmærksomhed, der rettes mod en pludselig høj lyd, eller et lysglimt.
Det er det dorsale fronto-parietale system i hjernen, der er aktivt ved begge typer opmærksomhed. Forskellen ligger i aktiviteten:. Ved endogen opmærksomhed er venstre hjernehalvdels forreste del mere aktiv end den parietale del. Det modsatte mønster ses ved eksogen opmærksomhed.
Referencer
Kognitionspsykologi | danish | 0.629718 |
mechanism_mentally_tired/Mental_health.txt |
Mental health encompasses emotional, psychological, and social well-being, influencing cognition, perception, and behavior. According to World Health Organization (WHO), it is a "state of well-being in which the individual realizes his or her abilities, can cope with the normal stresses of life, can work productively and fruitfully, and can contribute to his or her community". It likewise determines how an individual handles stress, interpersonal relationships, and decision-making. Mental health includes subjective well-being, perceived self-efficacy, autonomy, competence, intergenerational dependence, and self-actualization of one's intellectual and emotional potential, among others. From the perspectives of positive psychology or holism, mental health may include an individual's ability to enjoy life and to create a balance between life activities and efforts to achieve psychological resilience. Cultural differences, personal philosophy, subjective assessments, and competing professional theories all affect how one defines "mental health". Some early signs related to mental health difficulties are sleep irritation, lack of energy, lack of appetite, thinking of harming oneself or others, self-isolating (though introversion and isolation aren't necessarily unhealthy), and frequently zoning out.
Mental disorders[edit]
See also: Mental disorder
Mental health, as defined by the Public Health Agency of Canada, is an individual's capacity to feel, think, and act in ways to achieve a better quality of life while respecting personal, social, and cultural boundaries. Impairment of any of these are risk factor for mental disorders, or mental illnesses, which are a component of mental health. In 2019, about 970 million people worldwide suffered from a mental disorder, with anxiety and depression being the most common. The number of people suffering from mental disorders has risen significantly throughout the years. Mental disorders are defined as health conditions that affect and alter cognitive functioning, emotional responses, and behavior associated with distress and/or impaired functioning. The ICD-11 is the global standard used to diagnose, treat, research, and report various mental disorders. In the United States, the DSM-5 is used as the classification system of mental disorders.
Mental health is associated with a number of lifestyle factors such as diet, exercise, stress, drug abuse, social connections and interactions. Psychiatrists, psychologists, licensed professional clinical counselors, social workers, nurse practitioners, and family physicians can help manage mental illness with treatments such as therapy, counseling, and medication.
History[edit]
See also: Well-being, Eudaimonia, and History of mental disorders
Early history[edit]
In the mid-19th century, William Sweetser was the first to coin the term mental hygiene, which can be seen as the precursor to contemporary approaches to work on promoting positive mental health. Isaac Ray, the fourth president of the American Psychiatric Association and one of its founders, further defined mental hygiene as "the art of preserving the mind against all incidents and influences calculated to deteriorate its qualities, impair its energies, or derange its movements".
In American history, mentally ill patients were thought to be religiously punished. This response persisted through the 1700s, along with the inhumane confinement and stigmatization of such individuals. Dorothea Dix (1802–1887) was an important figure in the development of the "mental hygiene" movement. Dix was a school teacher who endeavored to help people with mental disorders and to expose the sub-standard conditions into which they were put. This became known as the "mental hygiene movement". Before this movement, it was not uncommon that people affected by mental illness would be considerably neglected, often left alone in deplorable conditions without sufficient clothing. From 1840 to 1880, she won the support of the federal government to set up over 30 state psychiatric hospitals; however, they were understaffed, under-resourced, and were accused of violating human rights.
Emil Kraepelin in 1896 developed the taxonomy of mental disorders which has dominated the field for nearly 80 years. Later, the proposed disease model of abnormality was subjected to analysis and considered normality to be relative to the physical, geographical and cultural aspects of the defining group.
At the beginning of the 20th century, Clifford Beers founded "Mental Health America – National Committee for Mental Hygiene", after publication of his accounts as a patient in several lunatic asylums, A Mind That Found Itself, in 1908 and opened the first outpatient mental health clinic in the United States.
The mental hygiene movement, similar to the social hygiene movement, had at times been associated with advocating eugenics and sterilization of those considered too mentally deficient to be assisted into productive work and contented family life. In the post-WWII years, references to mental hygiene were gradually replaced by the term 'mental health' due to its positive aspect that evolves from the treatment of illness to preventive and promotive areas of healthcare.
Deinstitutionalization and transinstitutionalization[edit]
When US government-run hospitals were accused of violating human rights, advocates pushed for deinstitutionalization: the replacement of federal mental hospitals for community mental health services. The closure of state-provisioned psychiatric hospitals was enforced by the Community Mental Health Centers Act in 1963 that laid out terms in which only patients who posed an imminent danger to others or themselves could be admitted into state facilities. This was seen as an improvement from previous conditions. However, there remains a debate on the conditions of these community resources.
It has been proven that this transition was beneficial for many patients: there was an increase in overall satisfaction, a better quality of life, and more friendships between patients all at an affordable cost. This proved to be true only in the circumstance that treatment facilities that had enough funding for staff and equipment as well as proper management. However, this idea is a polarizing issue. Critics of deinstitutionalization argue that poor living conditions prevailed, patients were lonely, and they did not acquire proper medical care in these treatment homes. Additionally, patients that were moved from state psychiatric care to nursing and residential homes had deficits in crucial aspects of their treatment. Some cases result in the shift of care from health workers to patients' families, where they do not have the proper funding or medical expertise to give proper care. On the other hand, patients that are treated in community mental health centers lack sufficient cancer testing, vaccinations, or otherwise regular medical check-ups.
Other critics of state deinstitutionalization argue that this was simply a transition to "transinstitutionalization", or the idea that prisons and state-provisioned hospitals are interdependent. In other words, patients become inmates. This draws on the Penrose Hypothesis of 1939, which theorized that there was an inverse relationship between prisons' population size and the number of psychiatric hospital beds. This means that populations that require psychiatric mental care will transition between institutions, which in this case, includes state psychiatric hospitals and criminal justice systems. Thus, a decrease in available psychiatric hospital beds occurred at the same time as an increase in inmates. Although some are skeptical that this is due to other external factors, others will reason this conclusion to a lack of empathy for the mentally ill. There is no argument in the social stigmatization of those with mental illnesses, they have been widely marginalized and discriminated against in society. In this source, researchers analyze how most compensation prisoners (detainees who are unable or unwilling to pay a fine for petty crimes) are unemployed, homeless, and with an extraordinarily high degree of mental illnesses and substance use disorders. Compensation prisoners then lose prospective job opportunities, face social marginalization, and lack access to resocialization programs, which ultimately facilitate reoffending. The research sheds light on how the mentally ill—and in this case, the poor—are further punished for certain circumstances that are beyond their control, and that this is a vicious cycle that repeats itself. Thus, prisons embody another state-provisioned mental hospital.
Families of patients, advocates, and mental health professionals still call for the increase in more well-structured community facilities and treatment programs with a higher quality of long-term inpatient resources and care. With this more structured environment, the United States will continue with more access to mental health care and an increase in the overall treatment of the mentally ill.
However, there is still a lack of studies for mental health conditions (MHCs) to raise awareness, knowledge development, and attitude of seeking medical treatment for MHCs in Bangladesh. People in rural areas often seek treatment from the traditional healers and MHCs are sometimes considered a spiritual matter.
Epidemiology[edit]
See also: Prevalence of mental disorders
Mental illnesses are more common than cancer, diabetes, or heart disease. As of 2021, over 22 percent of all Americans over the age of 18 meet the criteria for having a mental illness. Evidence suggests that 970 million people worldwide have a mental disorder. Major depression ranks third among the top 10 leading causes of disease worldwide. By 2030, it is predicted to become the leading cause of disease worldwide. Over 700 000 thousand people commit suicide every year and around 14 million attempt it. A World Health Organization (WHO) report estimates the global cost of mental illness at nearly $2.5 trillion (two-thirds in indirect costs) in 2010, with a projected increase to over $6 trillion by 2030.
Evidence from the WHO suggests that nearly half of the world's population is affected by mental illness with an impact on their self-esteem, relationships and ability to function in everyday life. An individual's emotional health can impact their physical health. Poor mental health can lead to problems such as the inability to make adequate decisions and substance use disorders.
Good mental health can improve life quality whereas poor mental health can worsen it. According to Richards, Campania, & Muse-Burke, "There is growing evidence that is showing emotional abilities are associated with pro-social behaviors such as stress management and physical health." Their research also concluded that people who lack emotional expression are inclined to anti-social behaviors (e.g., substance use disorder and alcohol use disorder, physical fights, vandalism), which reflects one's mental health and suppressed emotions. Adults and children who face mental illness may experience social stigma, which can exacerbate the issues.
Global prevalence[edit]
See also: Global mental health and Category:Mental health by country
The Two Continua Model of Mental Health and Mental Illness
Mental health can be seen as a continuum, where an individual's mental health may have many different possible values. Mental wellness is viewed as a positive attribute; this definition of mental health highlights emotional well-being, the capacity to live a full and creative life, and the flexibility to deal with life's inevitable challenges. Some discussions are formulated in terms of contentment or happiness. Many therapeutic systems and self-help books offer methods and philosophies espousing strategies and techniques vaunted as effective for further improving the mental wellness. Positive psychology is increasingly prominent in mental health.
A holistic model of mental health generally includes concepts based upon anthropological, educational, psychological, religious, and sociological perspectives. There are also models as theoretical perspectives from personality, social, clinical, health and developmental psychology.
The tripartite model of mental well-being views mental well-being as encompassing three components of emotional well-being, social well-being, and psychological well-being. Emotional well-being is defined as having high levels of positive emotions, whereas social and psychological well-being are defined as the presence of psychological and social skills and abilities that contribute to optimal functioning in daily life. The model has received empirical support across cultures. The Mental Health Continuum-Short Form (MHC-SF) is the most widely used scale to measure the tripartite model of mental well-being.
Demographics[edit]
Children and young adults[edit]
Further information: Infant mental health, Mental disorders diagnosed in childhood, and Developmental psychopathology
See also: Depression in childhood and adolescence and Adverse childhood experiences
Mental health conditions are 16% of the global burden of disease and injury in people aged 10–19 years. 8.9 million young adults are affected by mental illness. 42% of those young adults went untreated as of 2018. In the United States alone in 2021, at least roughly 17.5% of the population (ages 18 and older) were recorded as having a mental illness. The comparison between reports and statistics of mental health issues in newer generations (18–25 years old to 26–49 years old) and the older generation (50 years or older) signifies an increase in mental health issues as only 15% of the older generation reported a mental health issue whereas the newer generations reported 33.7% (18-25) and 28.1% (26-49).></ref> Half of all mental health conditions start by 14 years of age but most cases go undetected and untreated. The role of caregivers for youth with mental health needs is valuable, and caregivers benefit most when they have sufficient psychoeducation and peer support. Depression is one of the leading causes of illness and disability among adolescents. Suicide is the fourth leading cause of death in 15-19-year-olds. Exposure to childhood trauma can cause mental health disorders and poor academic achievement. Ignoring mental health conditions in adolescents can impact adulthood. 50% of preschool children show a natural reduction in behavioral problems. The remaining experience long-term consequences. It impairs physical and mental health and limits opportunities to live fulfilling lives. A result of depression during adolescence and adulthood may be substance abuse. The average age of onset is between 11 and 14 years for depressive disorders. Only approximately 25% of children with behavioral problems refer to medical services. The majority of children go untreated.
Homeless population[edit]
Further information: Homelessness and mental health
Mental illness is thought to be highly prevalent among homeless populations, though access to proper diagnoses is limited. An article written by Lisa Goodman and her colleagues summarized Smith's research into PTSD in homeless single women and mothers in St. Louis, Missouri, which found that 53% of the respondents met diagnostic criteria, and which describes homelessness as a risk factor for mental illness. At least two commonly reported symptoms of psychological trauma, social disaffiliation and learned helplessness are highly prevalent among homeless individuals and families.
While mental illness is prevalent, people infrequently receive appropriate care. Case management linked to other services is an effective care approach for improving symptoms in people experiencing homelessness. Case management reduced admission to hospitals, and it reduced substance use by those with substance abuse problems more than typical care.
Immigrants and refugees[edit]
See also: Mental health of refugees
States that produce refugees are sites of social upheaval, civil war, even genocide. Most refugees experience trauma. It can be in the form of torture, sexual assault, family fragmentation, and death of loved ones.
Refugees and immigrants experience psychosocial stressors after resettlement. These include discrimination, lack of economic stability, and social isolation causing emotional distress. For example, Not far into the 1900s, campaigns targeting Japanese immigrants were being formed that inhibited their ability to participate in U.S life, painting them as a threat to the American working-class. They were subject to prejudice and slandered by American media as well as anti-Japanese legislation being implemented. For refugees family reunification can be one of the primary needs to improve quality of life. Post-migration trauma is a cause of depressive disorders and psychological distress for immigrants.
Cultural and religious considerations[edit]
Mental health is a socially constructed concept; different societies, groups, cultures (both ethnic and national/regional), institutions, and professions have very different ways of conceptualizing its nature and causes, determining what is mentally healthy, and deciding what interventions, if any, are appropriate. Thus, different professionals will have different cultural, class, political and religious backgrounds, which will impact the methodology applied during treatment. In the context of deaf mental health care, it is necessary for professionals to have cultural competency of deaf and hard of hearing people and to understand how to properly rely on trained, qualified, and certified interpreters when working with culturally Deaf clients.
Research has shown that there is stigma attached to mental illness. Due to such stigma, individuals may resist labeling and may be driven to respond to mental health diagnoses with denialism. Family caregivers of individuals with mental disorders may also suffer discrimination or face stigma.
Addressing and eliminating the social stigma and perceived stigma attached to mental illness has been recognized as crucial to education and awareness surrounding mental health issues. In the United Kingdom, the Royal College of Psychiatrists organized the campaign Changing Minds (1998–2003) to help reduce stigma, while in the United States, efforts by entities such as the Born This Way Foundation and The Manic Monologues specifically focus on removing the stigma surrounding mental illness. The National Alliance on Mental Illness (NAMI) is a U.S. institution founded in 1979 to represent and advocate for those struggling with mental health issues. NAMI helps to educate about mental illnesses and health issues, while also working to eliminate stigma attached to these disorders.
Many mental health professionals are beginning to, or already understand, the importance of competency in religious diversity and spirituality, or the lack thereof. They are also partaking in cultural training to better understand which interventions work best for these different groups of people. The American Psychological Association explicitly states that religion must be respected. Education in spiritual and religious matters is also required by the American Psychiatric Association, however, far less attention is paid to the damage that more rigid, fundamentalist faiths commonly practiced in the United States can cause. This theme has been widely politicized in 2018 such as with the creation of the Religious Liberty Task Force in July of that year. Also, many providers and practitioners in the United States are only beginning to realize that the institution of mental healthcare lacks knowledge and competence of many non-Western cultures, leaving providers in the United States ill-equipped to treat patients from different cultures.
Occupations[edit]
See also: Mental health in aviation
Occupational therapy[edit]
Occupational therapy practitioners aim to improve and enable a client or group's participation in meaningful, everyday occupations. In this sense, occupation is defined as any activity that “occupies one's time". Examples of those activities include daily tasks (dressing, bathing, eating, house chores, driving, etc.), sleep and rest, education, work, play, leisure (hobbies), and social interactions. The OT profession offers a vast range of services for all stages of life in a myriad of practice settings, though the foundations of OT come from mental health. Community support for mental health through expert-moderated support groups can aid those who want to recover from mental illness or otherwise improve their emotional well-being.
OT services focused on mental health can be provided to persons, groups, and populations across the lifespan and experiencing varying levels of mental health performance. For example, occupational therapy practitioners provide mental health services in school systems, military environments, hospitals, outpatient clinics, and inpatient mental health rehabilitation settings. Interventions or support can be provided directly through specific treatment interventions or indirectly by providing consultation to businesses, schools, or other larger groups to incorporate mental health strategies on a programmatic level. Even people who are mentally healthy can benefit from the health promotion and additional prevention strategies to reduce the impact of difficult situations.
The interventions focus on positive functioning, sensory strategies, managing emotions, interpersonal relationships, sleep, community engagement, and other cognitive skills (i.e. visual-perceptual skills, attention, memory, arousal/energy management, etc.).
Mental health in social work[edit]
Further information: Social work
See also: Clinical social work
Social work in mental health, also called psychiatric social work, is a process where an individual in a setting is helped to attain freedom from overlapping internal and external problems (social and economic situations, family and other relationships, the physical and organizational environment, psychiatric symptoms, etc.). It aims for harmony, quality of life, self-actualization and personal adaptation across all systems. Psychiatric social workers are mental health professionals that can assist patients and their family members in coping with both mental health issues and various economic or social problems caused by mental illness or psychiatric dysfunctions and to attain improved mental health and well-being. They are vital members of the treatment teams in Departments of Psychiatry and Behavioral Sciences in hospitals. They are employed in both outpatient and inpatient settings of a hospital, nursing homes, state and local governments, substance use clinics, correctional facilities, health care services, private practice, etc.
In the United States, social workers provide most of the mental health services. According to government sources, 60 percent of mental health professionals are clinically trained social workers, 10 percent are psychiatrists, 23 percent are psychologists, and 5 percent are psychiatric nurses.
Mental health social workers in Japan have professional knowledge of health and welfare and skills essential for person's well-being. Their social work training enables them as a professional to carry out Consultation assistance for mental disabilities and their social reintegration; Consultation regarding the rehabilitation of the victims; Advice and guidance for post-discharge residence and re-employment after hospitalized care, for major life events in regular life, money and self-management and other relevant matters to equip them to adapt in daily life. Social workers provide individual home visits for mentally ill and do welfare services available, with specialized training a range of procedural services are coordinated for home, workplace and school. In an administrative relationship, Psychiatric social workers provides consultation, leadership, conflict management and work direction. Psychiatric social workers who provides assessment and psychosocial interventions function as a clinician, counselor and municipal staff of the health centers.
Risk factors and causes of mental health problems[edit]
Main article: Causes of mental disorders
There are many things that can contribute to mental health problems, including biological factors, genetic factors, life experiences (such as psychological trauma or abuse), and a family history of mental health problems.
Biological factors[edit]
According to the National Institute of Health Curriculum Supplement Series book, most scientists believe that changes in neurotransmitters can cause mental illnesses. In the section "The Biology of Mental Illnesses" the issue is explained in detail, "…there may be disruptions in the neurotransmitters dopamine, glutamate, and norepinephrine in individuals who have schizophrenia".
Demographic factors[edit]
Gender, age, ethnicity, life expectancy, longevity, population density, and community diversity are all demographic characteristics that can increase the risk and severity of mental disorders. Existing evidence demonstrates that the female gender is connected with an elevated risk of depression at differerent phases of life, commencing in adolescence in different contexts. Females, for example, have a higher risk of anxiety and eating disorders, whereas males have a higher chance of substance abuse and behavioural and developmental issues. This does not imply that women are less likely to suffer from developmental disorders such autism spectrum disorder, attention deficit hyperactivity disorder, Tourette syndrome, or early-onset schizophrenia. factors. Ethnicity and ethnic heterogeneity have also been identified as risk factors for the prevalence of mental disorders, with minority groups being at a higher risk due to discrimination and exclusion.
Unemployment has been shown to hurt an individual's emotional well-being, self-esteem, and more broadly their mental health. Increasing unemployment has been shown to have a significant impact on mental health, predominantly depressive disorders. This is an important consideration when reviewing the triggers for mental health disorders in any population survey. According to a 2009 meta-analysis by Paul and Moser, countries with high income inequality and poor unemployment protections experience worse mental health outcomes among the unemployed.
The prevalence of mental illness is higher in more economically unequal countries.
Emotional mental disorders are a leading cause of disabilities worldwide. Investigating the degree and severity of untreated emotional mental disorders throughout the world is a top priority of the World Mental Health (WMH) survey initiative, which was created in 1998 by the World Health Organization (WHO). "Neuropsychiatric disorders are the leading causes of disability worldwide, accounting for 37% of all healthy life years lost through disease. These disorders are most destructive to low and middle-income countries due to their inability to provide their citizens with proper aid. Despite modern treatment and rehabilitation for emotional mental health disorders, "even economically advantaged societies have competing priorities and budgetary constraints".
Unhappily married couples suffer 3–25 times the risk of developing clinical depression.
The World Mental Health survey initiative has suggested a plan for countries to redesign their mental health care systems to best allocate resources.
"A first step is documentation of services being used and the extent and nature of unmet treatment needs. A second step could be to do a cross-national comparison of service use and unmet needs in countries with different mental health care systems. Such comparisons can help to uncover optimum financing, national policies, and delivery systems for mental health care."
Knowledge of how to provide effective emotional mental health care has become imperative worldwide. Unfortunately, most countries have insufficient data to guide decisions, absent or competing visions for resources, and near-constant pressures to cut insurance and entitlements. WMH surveys were done in Africa (Nigeria, South Africa), the Americas (Colombia, Mexico, United States), Asia and the Pacific (Japan, New Zealand, Beijing and Shanghai in the People's Republic of China), Europe (Belgium, France, Germany, Italy, Netherlands, Spain, Ukraine), and the Middle East (Israel, Lebanon). Countries were classified with World Bank criteria as low-income (Nigeria), lower-middle-income (China, Colombia, South Africa, Ukraine), higher middle-income (Lebanon, Mexico), and high-income.
The coordinated surveys on emotional mental health disorders, their severity, and treatments were implemented in the aforementioned countries. These surveys assessed the frequency, types, and adequacy of mental health service use in 17 countries in which WMH surveys are complete. The WMH also examined unmet needs for treatment in strata defined by the seriousness of mental disorders. Their research showed that "the number of respondents using any 12-month mental health service was generally lower in developing than in developed countries, and the proportion receiving services tended to correspond to countries' percentages of gross domestic product spent on health care".
"High levels of unmet need worldwide are not surprising, since WHO Project ATLAS' findings of much lower mental health expenditures than was suggested by the magnitude of burdens from mental illnesses. Generally, unmet needs in low-income and middle-income countries might be attributable to these nations spending reduced amounts (usually <1%) of already diminished health budgets on mental health care, and they rely heavily on out-of-pocket spending by citizens who are ill-equipped for it".
Stress[edit]
Dementia Friends training
The Centre for Addiction and Mental Health discuss how a certain amount of stress is a normal part of daily life. Small doses of stress help people meet deadlines, be prepared for presentations, be productive and arrive on time for important events. However, long-term stress can become harmful. When stress becomes overwhelming and prolonged, the risks for mental health problems and medical problems increase." Also on that note, some studies have found language to deteriorate mental health and even harm humans.
Poverty[edit]
This section is an excerpt from Poverty § Mental health.[edit]
A Venezuelan eating from garbage during the crisis in Bolivarian Venezuela
A psychological study has been conducted by four scientists during inaugural Convention of Psychological Science. The results find that people who thrive with financial stability or fall under low socioeconomic status (SES) tend to perform worse cognitively due to external pressure imposed upon them. The research found that stressors such as low income, inadequate health care, discrimination, and exposure to criminal activities all contribute to mental disorders. This study also found that children exposed to poverty-stricken environments have slower cognitive thinking. It is seen that children perform better under the care of their parents and that children tend to adopt speaking language at a younger age. Since being in poverty from childhood is more harmful than it is for an adult, it is seen that children in poor households tend to fall behind in certain cognitive abilities compared to other average families.
For a child to grow up emotionally healthy, the children under three need "A strong, reliable primary caregiver who provides consistent and unconditional love, guidance, and support. Safe, predictable, stable environments. Ten to 20 hours each week of harmonious, reciprocal interactions. This process, known as attunement, is most crucial during the first 6–24 months of infants' lives and helps them develop a wider range of healthy emotions, including gratitude, forgiveness, and empathy. Enrichment through personalized, increasingly complex activities". In one survey, 67% of children from disadvantaged inner cities said they had witnessed a serious assault, and 33% reported witnessing a homicide. 51% of fifth graders from New Orleans (median income for a household: $27,133) have been found to be victims of violence, compared to 32% in Washington, DC (mean income for a household: $40,127). Studies have shown that poverty changes the personalities of children who live in it. The Great Smoky Mountains Study was a ten-year study that was able to demonstrate this. During the study, about one-quarter of the families saw a dramatic and unexpected increase in income. The study showed that among these children, instances of behavioral and emotional disorders decreased, and conscientiousness and agreeableness increased.
Environmental factors[edit]
This section is an excerpt from Effects of climate change on mental health.[edit]
Smoke in Sydney (Australia) from large bushfires (in 2019), affected some people's mental health in a direct way. The likelihood of wildfires is increased by climate change.
The effects of climate change on mental health and wellbeing are documented. This is especially the case for vulnerable populations and those with pre-existing serious mental illness. There are three broad pathways by which these effects can take place: directly, indirectly or via awareness. The direct pathway includes stress-related conditions caused by exposure to extreme weather events. These include post-traumatic stress disorder (PTSD). Scientific studies have linked mental health to several climate-related exposures. These include heat, humidity, rainfall, drought, wildfires and floods. The indirect pathway can be disruption to economic and social activities. An example is when an area of farmland is less able to produce food. The third pathway can be of mere awareness of the climate change threat, even by individuals who are not otherwise affected by it.
An additional aspect to consider is the detrimental impact climate change can have on green or blue natural spaces, which in themselves have been proven to have beneficial impact on mental health. Impacts of anthropogenic climate change, such as freshwater pollution or deforestation, degrade these landscapes and reduce public access. Even when the green and blue spaces are intact, access to them is not equal across society, which is an issue of environmental justice and economic inequality.
Mental health outcomes have been measured in several studies. These use indicators such as psychiatric hospital admissions, mortality, self-harm and suicide rates. People with pre-existing mental illness, Indigenous peoples, migrants and refugees, and children and adolescents are all vulnerable. The emotional responses to the threat of climate change can include eco-anxiety, ecological grief and eco-anger. Such emotions can be rational responses to the degradation of the natural world and lead to adaptive action.
Assessing the exact mental health effects of climate change is difficult; increases in heat extremes pose risks to mental health which can manifest themselves in increased mental health-related hospital admissions and suicidality.
Prevention and promotion[edit]
See also: Prevention of mental disorders
"The terms mental health promotion and prevention have often been confused. Promotion is defined as intervening to optimize positive mental health by addressing determinants of positive mental health (i.e. protective factors) before a specific mental health problem has been identified, with the ultimate goal of improving the positive mental health of the population. Mental health prevention is defined as intervening to minimize mental health problems (i.e. risk factors) by addressing determinants of mental health problems before a specific mental health problem has been identified in the individual, group, or population of focus with the ultimate goal of reducing the number of future mental health problems in the population."
In order to improve mental health, the root of the issue has to be resolved. "Prevention emphasizes the avoidance of risk factors; promotion aims to enhance an individual's ability to achieve a positive sense of self-esteem, mastery, well-being, and social inclusion." Mental health promotion attempts to increase protective factors and healthy behaviors that can help prevent the onset of a diagnosable mental disorder and reduce risk factors that can lead to the development of a mental disorder. Yoga is an example of an activity that calms one's entire body and nerves. According to a study on well-being by Richards, Campania, and Muse-Burke, "mindfulness is considered to be a purposeful state, it may be that those who practice it belief in its importance and value being mindful, so that valuing of self-care activities may influence the intentional component of mindfulness." Akin to surgery, sometimes the body must be further damaged, before it can properly heal
Mental health is conventionally defined as a hybrid of the absence of a mental disorder and the presence of well-being. Focus is increasing on preventing mental disorders.
Prevention is beginning to appear in mental health strategies, including the 2004 WHO report "Prevention of Mental Disorders", the 2008 EU "Pact for Mental Health" and the 2011 US National Prevention Strategy. Some commentators have argued that a pragmatic and practical approach to mental disorder prevention at work would be to treat it the same way as physical injury prevention.
Prevention of a disorder at a young age may significantly decrease the chances that a child will have a disorder later in life, and shall be the most efficient and effective measure from a public health perspective. Prevention may require the regular consultation of a physician for at least twice a year to detect any signs that reveal any mental health concerns.
Additionally, social media is becoming a resource for prevention. In 2004, the Mental Health Services Act began to fund marketing initiatives to educate the public on mental health. This California-based project is working to combat the negative perception with mental health and reduce the stigma associated with it. While social media can benefit mental health, it can also lead to deterioration if not managed properly. Limiting social media intake is beneficial.
Studies report that patients in mental health care who can access and read their Electronic Health Records (EHR) or Open Notes online experience increased understanding of their mental health, feeling in control of their care, and enhanced trust in their clinicians. Patients' also reported feelings of greater validation, engagement, remembering their care plan, and acquiring a better awareness of potential side effects of their medications, when reading their mental health notes. Other common experiences were that shared mental health notes enhance patient empowerment and augment patient autonomy.
Furthermore, recent studies have shown that social media is an effective way to draw attention to mental health issues. By collecting data from Twitter, researchers found that social media presence is heightened after an event relating to behavioral health occurs. Researchers continue to find effective ways to use social media to bring more awareness to mental health issues through online campaigns in other sites such as Facebook and Instagram.
Care navigation[edit]
Main article: Mental health care navigator
Mental health care navigation helps to guide patients and families through the fragmented, often confusing mental health industries. Care navigators work closely with patients and families through discussion and collaboration to provide information on best therapies as well as referrals to practitioners and facilities specializing in particular forms of emotional improvement. The difference between therapy and care navigation is that the care navigation process provides information and directs patients to therapy rather than providing therapy. Still, care navigators may offer diagnosis and treatment planning. Though many care navigators are also trained therapists and doctors. Care navigation is the link between the patient and the below therapies. A clear recognition that mental health requires medical intervention was demonstrated in a study by Kessler et al. of the prevalence and treatment of mental disorders from 1990 to 2003 in the United States. Despite the prevalence of mental health disorders remaining unchanged during this period, the number of patients seeking treatment for mental disorders increased threefold.
Methods[edit]
Pharmacotherapy[edit]
Pharmacotherapy is a therapy that uses pharmaceutical drugs. Pharmacotherapy is used in the treatment of mental illness through the use of antidepressants, benzodiazepines, and the use of elements such as lithium. It can only be prescribed by a medical professional trained in the field of Psychiatry.
Physical activity[edit]
Physical exercise can improve mental and physical health. Playing sports, walking, cycling, or doing any form of physical activity trigger the production of various hormones, sometimes including endorphins, which can elevate a person's mood.
Studies have shown that in some cases, physical activity can have the same impact as antidepressants when treating depression and anxiety.
Moreover, cessation of physical exercise may have adverse effects on some mental health conditions, such as depression and anxiety. This could lead to different negative outcomes such as obesity, skewed body image and many health risks associated with mental illnesses. Exercise can improve mental health but it should not be used as an alternative to therapy.
Activity therapies[edit]
Activity therapies also called recreation therapy and occupational therapy, promote healing through active engagement. An example of occupational therapy would be promoting an activity that improves daily life, such as self-care or improving hobbies.
Each of these therapies have proven to improve mental health and have resulted in healthier, happier individuals. In recent years, for example, coloring has been recognized as an activity that has been proven to significantly lower the levels of depressive symptoms and anxiety in many studies.
Expressive therapies[edit]
Expressive therapies or creative arts therapies are a form of psychotherapy that involves the arts or art-making. These therapies include art therapy, music therapy, drama therapy, dance therapy, and poetry therapy. It has been proven that music therapy is an effective way of helping people with a mental health disorder. Drama therapy is approved by NICE for the treatment of psychosis.
Psychotherapy[edit]
Main article: Psychotherapy
Psychotherapy is the general term for the scientific based treatment of mental health issues based on modern medicine. It includes a number of schools, such as gestalt therapy, psychoanalysis, cognitive behavioral therapy, psychedelic therapy, transpersonal psychology/psychotherapy, and dialectical behavioral therapy.
Group therapy involves any type of therapy that takes place in a setting involving multiple people. It can include psychodynamic groups, expressive therapy groups, support groups (including the Twelve-step program), problem-solving and psychoeducation groups.
Self-compassion[edit]
Main article: Self-compassion
According to Neff, self-compassion consists of three main positive components and their negative counterparts: Self-Kindness versus Self-Judgment, Common Humanity versus Isolation and Mindfulness versus Over-Identification. Furthermore, there is evidence from a study by Shin & Lin suggesting specific components of self-compassion can predict specific dimensions of positive mental health (emotional, social, and psychological well-being).
Social-emotional learning[edit]
Further information: Social emotional development § Social emotional learning & development in schools
The Collaborative for academic, social, emotional learning (CASEL) addresses five broad and interrelated areas of competence and highlights examples for each: self-awareness, self-management, social awareness, relationship skills, and responsible decision-making. A meta-analysis was done by Alexendru Boncu, Iuliana Costeau, & Mihaela Minulescu (2017) looking at social-emotional learning (SEL) studies and the effects on emotional and behavior outcomes. They found a small but significant effect size (across the studies looked into) for externalized problems and social-emotional skills.
Meditation[edit]
Main articles: Meditation and Mindfulness-based cognitive therapy
The practice of mindfulness meditation has several potential mental health benefits, such as bringing about reductions in depression, anxiety and stress. Mindfulness meditation may also be effective in treating substance use disorders.
Lucid Dreaming[edit]
Lucid dreaming has been found to be associated with greater mental well-being. It also was not associated with poorer sleep quality nor with cognitive dissociation. There is also some evidence lucid dreaming therapy can help with nightmare reduction.
Mental fitness[edit]
Mental fitness is a mental health movement that encourages people to intentionally regulate and maintain their emotional wellbeing through friendship, regular human contact, and activities that include meditation, calming exercises, aerobic exercise, mindfulness, having a routine and maintaining adequate sleep. Mental fitness is intended to build resilience against every-day mental and potentially physical health challenges to prevent an escalation of anxiety, depression, and suicidal ideation. This can help people, including older adults with health challenges, to more effectively cope with the escalation of those feelings if they occur.
Spiritual counseling[edit]
Spiritual counsellors meet with people in need to offer comfort and support and to help them gain a better understanding of their issues and develop a problem-solving relation with spirituality. These types of counselors deliver care based on spiritual, psychological and theological principles.
Laws and public health policies[edit]
See also: Mental health law and Public health
There are many factors that influence mental health including:
Mental illness, disability, and suicide are ultimately the result of a combination of biology, environment, and access to and utilization of mental health treatment.
Public health policies can influence access and utilization, which subsequently may improve mental health and help to progress the negative consequences of depression and its associated disability.
United States[edit]
Emotional mental illnesses should be a particular concern in the United States since the U.S. has the highest annual prevalence rates (26 percent) for mental illnesses among a comparison of 14 developing and developed countries. While approximately 80 percent of all people in the United States with a mental disorder eventually receive some form of treatment, on average persons do not access care until nearly a decade following the development of their illness, and less than one-third of people who seek help receive minimally adequate care. The government offers everyone programs and services, but veterans receive the most help, there is certain eligibility criteria that has to be met.
Policies[edit]
Mental health policies in the United States have experienced four major reforms: the American asylum movement led by Dorothea Dix in 1843; the mental hygiene movement inspired by Clifford Beers in 1908; the deinstitutionalization started by Action for Mental Health in 1961; and the community support movement called for by The CMCH Act Amendments of 1975.
In 1843, Dorothea Dix submitted a Memorial to the Legislature of Massachusetts, describing the abusive treatment and horrible conditions received by the mentally ill patients in jails, cages, and almshouses. She revealed in her Memorial: "I proceed, gentlemen, briefly to call your attention to the present state of insane persons confined within this Commonwealth, in cages, closets, cellars, stalls, pens! Chained, naked, beaten with rods, and lashed into obedience...." Many asylums were built in that period, with high fences or walls separating the patients from other community members and strict rules regarding the entrance and exit. In 1866, a recommendation came to the New York State Legislature to establish a separate asylum for chronic mentally ill patients. Some hospitals placed the chronic patients into separate wings or wards, or different buildings.
In A Mind That Found Itself (1908) Clifford Whittingham Beers described the humiliating treatment he received and the deplorable conditions in the mental hospital. One year later, the National Committee for Mental Hygiene (NCMH) was founded by a small group of reform-minded scholars and scientists—including Beers himself—which marked the beginning of the "mental hygiene" movement. The movement emphasized the importance of childhood prevention. World War I catalyzed this idea with an additional emphasis on the impact of maladjustment, which convinced the hygienists that prevention was the only practical approach to handle mental health issues. However, prevention was not successful, especially for chronic illness; the condemnable conditions in the hospitals were even more prevalent, especially under the pressure of the increasing number of chronically ill and the influence of the depression.
In 1961, the Joint Commission on Mental Health published a report called Action for Mental Health, whose goal was for community clinic care to take on the burden of prevention and early intervention of the mental illness, therefore to leave space in the hospitals for severe and chronic patients. The court started to rule in favor of the patients' will on whether they should be forced to treatment. By 1977, 650 community mental health centers were built to cover 43 percent of the population and serve 1.9 million individuals a year, and the lengths of treatment decreased from 6 months to only 23 days. However, issues still existed. Due to inflation, especially in the 1970s, the community nursing homes received less money to support the care and treatment provided. Fewer than half of the planned centers were created, and new methods did not fully replace the old approaches to carry out its full capacity of treating power. Besides, the community helping system was not fully established to support the patients' housing, vocational opportunities, income supports, and other benefits. Many patients returned to welfare and criminal justice institutions, and more became homeless. The movement of deinstitutionalization was facing great challenges.
After realizing that simply changing the location of mental health care from the state hospitals to nursing houses was insufficient to implement the idea of deinstitutionalization, the National Institute of Mental Health (NIMH) in 1975 created the Community Support Program (CSP) to provide funds for communities to set up a comprehensive mental health service and supports to help the mentally ill patients integrate successfully in the society. The program stressed the importance of other supports in addition to medical care, including housing, living expenses, employment, transportation, and education; and set up new national priority for people with serious mental disorders. In addition, the Congress enacted the Mental Health Systems Act of 1980 to prioritize the service to the mentally ill and emphasize the expansion of services beyond just clinical care alone. Later in the 1980s, under the influence from the Congress and the Supreme Court, many programs started to help the patients regain their benefits. A new Medicaid service was also established to serve people who were diagnosed with a "chronic mental illness". People who were temporally hospitalized were also provided aid and care and a pre-release program was created to enable people to apply for reinstatement prior to discharge. Not until 1990, around 35 years after the start of the deinstitutionalization, did the first state hospital begin to close. The number of hospitals dropped from around 300 by over 40 in the 1990s, and finally a Report on Mental Health showed the efficacy of mental health treatment, giving a range of treatments available for patients to choose.
However, several critics maintain that deinstitutionalization has, from a mental health point of view, been a thoroughgoing failure. The seriously mentally ill are either homeless, or in prison; in either case (especially the latter), they are getting little or no mental health care. This failure is attributed to a number of reasons over which there is some degree of contention, although there is general agreement that community support programs have been ineffective at best, due to a lack of funding.
The 2011 National Prevention Strategy included mental and emotional well-being, with recommendations including better parenting and early intervention programs, which increase the likelihood of prevention programs being included in future US mental health policies. The NIMH is researching only suicide and HIV/AIDS prevention, but the National Prevention Strategy could lead to it focusing more broadly on longitudinal prevention studies.
In 2013, United States Representative Tim Murphy introduced the Helping Families in Mental Health Crisis Act, HR2646. The bipartisan bill went through substantial revision and was reintroduced in 2015 by Murphy and Congresswoman Eddie Bernice Johnson. In November 2015, it passed the Health Subcommittee by an 18–12 vote.
See also[edit]
Psychology portal
Abnormal psychology
Emotional resilience
Ethnopsychopharmacology
Mental environment
Mental health day
Mental health during the COVID-19 pandemic
Mental health first aid
Mental health in education
Mental health in the workplace
Mental health of Asian Americans
Self-help groups for mental health
Social determinants of mental health
Social stigma
Suicide awareness
Telemental health
World Mental Health Day
Well-being | biology | 1591722 | https://sv.wikipedia.org/wiki/Psykisk%20h%C3%A4lsa | Psykisk hälsa | Psykisk hälsa ses numer som ett inre tillstånd av mentalt välbefinnande som är mer än enbart motsatsen till psykisk sjukdom eller störning. Definitionen av psykisk hälsa har under perioder pendlat mellan att utgå från normalitetsbegreppet, mellan att utgå från verklighetsuppfattning och andra jagfunktioner, mellan bedömningar av den inre upplevelsen (psykoanalys), bedömningar av beteendet (behaviorism) och medicinska studier av hjärnans och centrala nervsystemets funktioner (personlighetsgenetik, neurologi och neuroendokrinologi). Psykisk hälsa studeras särskilt inom den så kallade positiva psykologin.Så säger läraren Hans.
WHO definierar psykisk hälsa som "ett tillstånd av mentalt välbefinnande när individen kan förverkliga sina egna möjligheter, hantera normala påfrestningar, arbeta produktivt och bidra till samhället personen lever i".
WHO:s definitionen av (positiv) psykisk hälsa utgår från att positiv psykisk hälsa består av emotionellt välbefinnande och optimal psykologisk och social funktion. Att ha psykologisk funktion innebär till exempel att kunna hanterar vardagen med dess problem, till exempel stress, livsförändringar, samt de förluster och lidanden som hör till ett vanligt liv.
Symtom på positiv psykisk hälsa (subjektivt psykiskt välbefinnande)
Den amerikanska forskaren Corey L. M. Keyes uppställt tretton symtom på positiv psykisk hälsa som består av symtom på emotinellt välbefinnande och psykologisk och social funktion.
Positiv affekt - vid gott humör, lugn och full av liv.
Upplevd lycka och nöjdhet - känner sig nöjd och tillfreds med livet.
Självacceptans - har en positiv bild av sig själv, sitt förflutna och accepterar olika aspekter av självet.
Social acceptans - har en positiv bild av andra samtidigt som personen accepterar olikheter och människors komplexitiet.
Personlighetsutveckling - visar insikt i sin potential och utveckling och öppenhet inför utmaningar.
Social aktualisation - anser att människor och samhället kan utvecklas positivt och växa.
Mening med livet - har målsättningar som ger livet en mening.
Social kontribution - anser sig ha något att tillföra samhället, och att vad man gör är av värde för andra.
Behärskning av omgivningen - klarar av att behärska komplicerade miljöer och kan välja miljö som passar sina behov.
Social koherens - intresserad av samhället eller omgivningen, och anser att samhället är meningsfullt och begripligt.
Autonomi - är självgående med av samhället accepterade eller konventionella uppfattningar, och kan stå emot oönskat grupptryck.
Goda mellanmänskliga relationer - har varma, tillitsfulla och tillfredsställande relationer, och har förmåga till initimitet och empati.
Social integration - har en känsla av att vara en del av samhället, och att samhället är ett stöd.
Total psykisk hälsa
Eftersom positiv psykisk hälsa är ett eget tillstånd och inte motsatsen till psykisk ohälsa behöver hänsyn tas både till symtom på positiv psykisk hälsa och symtom på psykisk ohälsa när man pratar om en persons totala psykiska hälsa. Det innebär att man tillämpar en tvåfaktor modell för psykisk hälsa. En person med full psykisk hälsa har symtom på positiv psykisk hälsa (se ovan) och samtidigt frånvaro av symtom på psykisk ohälsa.
Se även
Klinisk psykologi
Metakognitiv terapi
Psykisk ohälsa
Psykoterapi
Källor
Marie Jahoda, Current Conpepts of Mental Health, North Stratford 1999
http://www.akademiska.se/upload/45747/infoblad%20-%20vad%20%C3%A4r%20psykisk%20h%C3%A4lsa.pdf
Noter
Psykologi | swedish | 0.736456 |
extinction_fungus/Prototaxites.txt |
Prototaxites /ˌproʊtoʊˈtæksɪtiːz/ is an extinct genus of terrestrial fossil fungi dating from the Late Silurian until the Late Devonian periods. Prototaxites formed large trunk-like structures up to 1 metre (3 ft) wide, reaching 8 metres (26 ft) in length, made up of interwoven tubes around 50 micrometres (0.0020 in) in diameter, making it by far the largest land-dwelling organism of its time.
The taxonomy of Prototaxites has long been the subject of debate. Currently, it is widely considered a fungus, but the debate is ongoing. Its exact relationship with extant fungus lineages is uncertain. It was almost certainly a perennial organism that grew over multiple years. Several ecologies have been proposed, including that it was saprotrophic like many modern fungi, or that it was a lichenised autotroph.
Morphology[edit]
Dawson's 1888 reconstruction of a conifer-like Prototaxites
The microstructure of Prototaxites under a light microscope
P. milwaukeensis from Wisconsin
Scanning electron microscope view of spherical phycobiont and elongate mycobiont of P. loganii
With a diameter of up to 1 metre (3 ft 3 in), and a height reaching 8.8 metres (29 ft), Prototaxites fossils are remnants of by far the largest organism discovered from the period of its existence. Viewed from afar, the fossils take the form of tree-trunks, spreading slightly near their base in a fashion that suggests a connection to unpreserved root-like structures. Infilled casts which may represent the spaces formerly occupied by "roots" of Prototaxites are common in early Devonian strata. Concentric growth rings, sometimes containing embedded plant material, suggest that the organism grew sporadically by the addition of external layers. It is probable that the preserved "trunks" represent the fruiting body, or "sporophore", of a fungus, which would have been fuelled by a mycelium, a net of dispersed filaments ("hyphae"). On a microscopic scale, the fossils consist of narrow tube-like structures, which weave around one another. These come in two types: skeletal "tubes", 20–50 μm across, have thick (2–6 μm) walls and are undivided for their length, and generative "filaments", which are thinner (5–10 μm diameter) and branch frequently; these mesh together to form the organism's matrix. These thinner filaments are septate—that is to say, they bear internal walls. These septa are perforate—i.e. they contain a pore, a trait only present in the modern red algae and fungi.
The similarity of these tubes to structures in the early plant Nematothallus has led to suggestions that the latter may represent leaves of Prototaxites. Unfortunately for this hypothesis, the two have never been found in connection, although this may be a consequence of their detachment after the organisms' death.
History of research[edit]
First collected in 1843, it was not until 14 years later that John William Dawson, a Canadian scientist, studied Prototaxites fossils, which he described as partially rotten giant conifers, containing the remains of the fungi which had been decomposing them. This concept was not disputed until 1872, when the rival scientist William Carruthers poured ridicule on the idea. Such was his fervour that he rebuked the name Prototaxites (loosely translated as "first yew") and insisted that the name Nematophycus ("stringy alga") be adopted, a move strongly against scientific convention. Dawson fought adamantly to defend his original interpretation until studies of the microstructure made it clear that his position was untenable, whence he promptly attempted to rename the genus himself (to Nematophyton, "stringy plant"), denying with great vehemence that he'd ever considered it to be a tree. Despite these political attempts to rename the genus, the rules of botanical nomenclature mean that the name "Prototaxites", however inappropriate in meaning, remains in use today.
Despite the overwhelming evidence that the organism grew on land, Carruthers's interpretation that it was a giant marine alga was challenged just the once, in 1919, when Church suggested that Carruthers had been too quick to rule out the possibility of the fungi. The lack of any characters diagnostic of any extant group made the presentation of a firm hypothesis difficult; the fossil remained an enigmatic mystery and subject of debate. It was not until 2001, after 20 years of research, that Francis Hueber, of Washington's National Museum of Natural History, published a long-awaited paper which attempted to put Prototaxites in its place. The paper deduced, based on its morphology, that Prototaxites was a fungus.
This idea was received with disbelief, denial and strong scepticism, but further evidence is emerging to support it. In 2007, isotopic analyses by a team including Hueber and Kevin Boyce of the University of Chicago concluded that Prototaxites was a giant fungus. They detected a highly variable range of values of carbon isotope ratios in a range of Prototaxites specimens; autotrophs (organisms such as plants and algae, that make a living via photosynthesis) living at the same time draw on the same (atmospheric) source of carbon; as organisms of the same type share the same chemical machinery, they reflect this atmospheric composition with a constant carbon isotope trace. The inconsistent ratio observed in Prototaxites appears to show that the organism did not survive by photosynthesis, and Boyce's team deduced that the organism fed on a range of substrates, such as the remains of whichever other organisms were nearby. Nevertheless, the large size of the organism would necessitate an extensive network of subterranean mycelia in order to obtain enough organic carbon to accumulate the necessary biomass. Root-like structures have circumstantially been interpreted as Prototaxites's rhizomorphs, and could support the possibility of the organism transporting nutrients large distances to support its above-ground body.
Other recent research has suggested that Prototaxites represents a rolled-up bundle of liverworts, but this interpretation has substantial difficulties.
A similar genus, Nematasketum, also consists of banded and branching tubes in axial bundles; this seems to be a fungus.
In 2021, Gregory Retallack described new species Prototaxites honeggeri from the Darriwilian (Middle Ordovician) age Douglas Lake Member of the Lenoir Limestone, at Douglas Dam, Tennessee, which makes the earliest appearance of this genus. While Ordovician origin of this genus is mentioned by some study, paleobotanist Dianne Edwards referred this study and commented "When diagnostic features are absent, such fragmentary organic materials can be misinterpreted, leading to implausible attributions". Nelson and Boyce (2022) referred this study by Retallack, but considered appearance of genus is Late Silurian.
A 2022 paper suggested that Prototaxites was a fungal rhizomorph that grew on its side and likely at least partially underground, as opposed to the traditional view that it grew upright.
Ecological context[edit]
Prototaxites would have been the tallest living organism in its day by far. In comparison, the plant Cooksonia only reached 6 centimetres (2.4 in) and itself towered over the "moss forests". Invertebrates were the only other land-dwelling multi-cellular life. Prototaxites became extinct as vascular plants rose to prominence. The organism could have used its tall columnar structure for spore dispersal. Alternatively, if Prototaxites contained photosynthetic structures, the height would have increased light capture. The University of Chicago research team has it reconstructed as a branchless, columnar structure. The presence of bio-molecules often associated with algae may suggest that the organism was covered by symbiotic (or parasitic) algae (making it in essence a huge lichen), or even that it was an alga itself. However, the variability in the ratios of δC between specimens of Prototaxites suggest that it was heterotrophic.
Prototaxites mycelia (strands) have been fossilised invading the tissue of vascular plants; in turn, there is evidence of animals inhabiting Prototaxites: mazes of tubes have been found within some specimens, with the fungus re-growing into the voids, leading to speculation that the organisms' extinction may have been caused by such activity; however, evidence of arthropod borings in Prototaxites has been found from the early and late Devonian, suggesting the organism survived the stress of boring for many millions of years. Intriguingly, Prototaxites was bored long before plants developed a structurally equivalent woody stem, and it is possible that the borers transferred to plants when these evolved.
| biology | 998 | https://da.wikipedia.org/wiki/Art | Art | Arten (species, forkortet sp., flertal: spp.) er den grundlæggende systematiske enhed inden for biologien. Arten defineres ofte som en naturlig gruppe af populationer, hvor udveksling af gener finder sted (eller kan finde sted) og som i forhold til forplantning er isoleret fra andre grupper. Det vil sige at kun individer inden for samme art kan parre sig og få forplantningsdygtigt afkom. Dette kaldes det biologiske artsbegreb. For organismer, der formerer sig ukønnet eller ved selvbestøvning, må arter afgrænses ud fra ligheder og forskelle mellem forskellige individer. Nogle dyrearter kan i fangenskab hybridisere og få fertilt afkom, men da dette ikke vil ske i naturen, selv om de mødes her, betragtes de som forskellige arter.
Eksempel
To heste kan parre sig og få et føl, der igen kan få føl med andre heste – hestene tilhører derfor samme art. En hest og et æsel kan også parre sig og deres unger kaldes enten muldyr eller mulæsel, afhængig af hvem der er moren, men muldyret eller mulæselet kan (normalt) ikke få unger, da de oftest er sterile. Af den grund regnes hest og æsel som to forskellige arter. Det samme princip gælder også for planterne. Denne naturskabte afgrænsning mellem to arter kaldes en artsbarriere. Den kan af og til gennembrydes, når ellers sterile krydsninger spontant eller kunstigt får gennemført en kromosomfordobling. Se f.eks. Vadegræs (Spartina pectinata).
Arter over for hybrider
Man kan dog godt komme ud for, at arter kan krydses og får blandet afkom, men hybriden vil kun kunne bestå på steder, hvor ingen af forældrearterne kan klare sig. Dette er et særligt udpræget problem med Rododendron (Rhododendron) og Tjørn (Crataegus), fordi disse slægter breder sig voldsomt efter skovbrand eller stormfald. Da hybriderne bliver frugtbare i en yngre alder end arterne, kan de dominere i en periode, men når skoven lukker sig, så fortrænges hybriderne og kun de specialiserede arter kan overleve i skovens dybe skygge eller ude i lyset i sumpe, på ur og i kalksten, m.m.
Flere artsbegreber
Fordi det biologiske artsbegreb kan være besværligt at anvende i praksis, er der efterhånden skabt en række andre artsbegreber:
Morfologisk artsbegreb Arterne adskiller sig fra hinanden ved deres bygning. Dette begreb er blevet meget anvendt gennem tiden.
Økologisk artsbegreb Definerer en art som en gruppe af organismer, der udfylder samme niche. Krydsninger mellem to nærtstående arter vil ikke være optimalt tilpasset til forældrearternes nicher og vil ikke klare sig i konkurrencen.
Evolutionære artsbegreb Også kaldet det kladistiske eller fylogenetiske artsbegreb. Naturen er dynamisk, ikke statisk - alle arter ændrer sig med tiden og bliver, hvis de ikke uddør som følge af konkurrence, naturkatastrofer m.v., til én eller flere nye arter. Det evolutionære artsbegreb minder om det biologiske, men inddrager tidsdimensionen, det vil sige at en art udvikler sig over tid og at nye arter opstår ved artsdannelse. Individer der fylogenetisk har samme stamfader tilhører samme art.
Pluralistisk artsbegreb En art er et samfund af populationer, der formerer sig og lever inden for en bestemt niche i naturen.
Se også
Systematik
Evolutionsteori
Kilder
Lars Skipper: Hvad er en art? Citat: "...Arten er den eneste [klassifikations-kategori] der eksisterer i virkeligheden, alle andre (slægter, familier, ordener m.v.) er indført for overskuelighedens skyld..."
Eksterne henvisninger
2003-12-31, ScienceDaily: Working On The 'Porsche Of Its Time': New Model For Species Determination Offered Citat: "...two species of dinosaur that are members of the same genera varied from each other by just 2.2 percent. Translation of the percentage into an actual number results in an average of just three skeletal differences out of the total 338 bones in the body. Amazingly, 58 percent of these differences occurred in the skull alone. "This is a lot less variation than I'd expected," said Novak..."
2003-08-08, ScienceDaily: Cross-species Mating May Be Evolutionarily Important And Lead To Rapid Change, Say Indiana University Researchers Citat: "...the sudden mixing of closely related species may occasionally provide the energy to impel rapid evolutionary change..."
2004-01-09 ScienceDaily: Mayo Researchers Observe Genetic Fusion Of Human, Animal Cells; May Help Explain Origin Of AIDS Citat: "...The researchers have discovered conditions in which pig cells and human cells can fuse together in the body to yield hybrid cells that contain genetic material from both species..."What we found was completely unexpected," says Jeffrey Platt, M.D..."
2000-09-18, ScienceDaily: Scientists Unravel Ancient Evolutionary History Of Photosynthesis Citat: "...gene-swapping was common among ancient bacteria early in evolution..."
2004-06-07, Sciencedaily: Parting Genomes: University Of Arizona Biologists Discover Seeds Of Speciation Citat: "...There's a huge amount of biodiversity out there, and we don't know where it comes from. Evolutionary biologists are excited to figure out what causes what we see out there--the relative forces of selection and drift--whether things are adapting to their environment or variation is random..."
2005-07-05, Sciencedaily: Trees, Vines And Nets -- Microbial Evolution Changes Its Face Citat: "... EBI researchers have changed our view of 4 billion years of microbial evolution...In all, more than 600,000 vertical transfers are observed, coupled with 90,000 gene loss events and approximately 40,000 horizontal gene transfers...A few species, including beneficial nitrogen-fixing soil bacteria, appear to be 'champions'of horizontal gene transfer; "it's entirely possible that apparently harmless organisms are quietly spreading antibiotic resistance under our feet," concludes Christos Ouzounis..."
2005-11-11, Sciencedaily: Lateral Thinking Produces First Map Of Gene Transmission Citat: "...Their results clearly show genetic modification of organisms by lateral transfer is a widespread natural phenomenon, and it can occur even between distantly related organisms... it was assumed that transfer of genes could only be vertical, i.e. from parents to offspring..."
Økologi
Biologi | danish | 0.791178 |
extinction_fungus/1216931.txt | Skip to Article Content
Skip to Article Information
The Ecological Society of America
Search
Search within
This Journal
Search term
Search
Login / Register
JOURNALS
Become a Member
ESA.org
Ecological Monographs
Ecological Monographs
Article
Free Access
A first comprehensive census of fungi in soil reveals both hyperdiversity and fine-scale niche partitioning
D. Lee Taylor, Teresa N. Hollingsworth, Jack W. McFarland, Niall J. Lennon, Chad Nusbaum, Roger W. Ruess
First published: 01 February 2014 https://doi.org/10.1890/12-1693.1Citations: 269
SECTIONSPDFPDFTOOLS SHARE
Abstract
Fungi play key roles in ecosystems as mutualists, pathogens, and decomposers. Current estimates of global species richness are highly uncertain, and the importance of stochastic vs. deterministic forces in the assembly of fungal communities is unknown. Molecular studies have so far failed to reach saturated, comprehensive estimates of fungal diversity. To obtain a more accurate estimate of global fungal diversity, we used a direct molecular approach to census diversity in a boreal ecosystem with precisely known plant diversity, and we carefully evaluated adequacy of sampling and accuracy of species delineation. We achieved the first exhaustive enumeration of fungi in soil, recording 1002 taxa in this system. We show that the fungus : plant ratio in Picea mariana forest soils from interior Alaska is at least 17:1 and is regionally stable. A global extrapolation of this ratio would suggest 6 million species of fungi, as opposed to leading estimates ranging from 616 000 to 1.5 million. We also find that closely related fungi often occupy divergent niches. This pattern is seen in fungi spanning all major functional guilds and four phyla, suggesting a major role of deterministic niche partitioning in community assembly. Extinctions and range shifts are reorganizing biodiversity on Earth, yet our results suggest that 98% of fungi remain undescribed and that many of these species occupy unique niches.
Introduction
The extent of species diversity on Earth is both highly uncertain and controversial (May 1988, 1991, Hawksworth 1991, Mora et al. 2011), yet few questions could be more fundamental given ongoing environmental change. The diversity of large eukaryotes is reasonably well known, whereas the diversities of smaller eukaryotes and prokaryotes are poorly known. Among small organisms such as fungi, only the largest and showiest species (charismatic megamycota) are well known. Fungi provide the global foundation for plant growth as mutualists (e.g., mycorrhizae, endophytes) and decomposers (e.g., soil hyphomycetes, wood-decaying basidiomycetes), and thereby function as key drivers in the carbon cycle. Earth has entered a period of unprecedented extinction (Barnosky et al. 2011), involving species range shifts and overall reorganization of biodiversity. An accurate estimate of the diversity of fungi and improved knowledge of the forces that control the distribution of species and assembly of communities are increasingly critical in light of global climate and land-use changes, loss of biodiversity, and the need to predict microbial functional feedbacks to climate.
The taxonomic diversity of fungi is known mostly from their macroscopic reproductive structures, i.e., sporocarps such as mushrooms. Though there are only roughly 100 000 described species in the Eumycota (Kirk et al. 2008), a true diversity of 1.5 million (Hawksworth 1991), or even 5 million (O'Brien et al. 2005) species has been suggested. Hence, it is clear that much of global fungal diversity remains to be documented. The majority of fungi depend in some way on living or dead plants for their energy supply, suggesting important linkages between aboveground and belowground diversity (Hooper et al. 2000). Hence, there has long been an interest in comparing and relating plant and fungal diversity. Hawksworth (1991) synthesized data from multiple sites in the United Kingdom with known plant diversities and extensive fungal collection records, and found that the ratio of species of fungi to vascular plants was remarkably consistent across sites, and averaged roughly 6 to 1. Based on a global count of 270 000 vascular plants, the UK ratio gave rise to a global extrapolation of 1.5 million species of fungi. While this extrapolation has been controversial (May 1991), it is also the most widely cited estimate of global fungal species richness. Several other studies have also reported statistically significant correlations between diversity within particular fungal functional or taxonomic groups and plant diversity at local to regional scales (Schmit et al. 2005, Sugiyama et al. 2008). However, these compilations have often involved data sets collected by different investigators using differing methods, and have not included molecular data. More systematic and exhaustive evaluations of fungal to plant (F:P) ratios and the mechanistic linkages between aboveground and belowground diversity are needed.
To date, compilations of F:P ratios have been entirely morphological, and thus, have relied upon identification of macroscopic fungal fruiting structures or identification of strains that could be isolated in culture. Previous F:P ratio analyses likely underestimate the true ratio due to the difficulty in culturing many fungi, our inability to distinguish morphological features that might differentiate species in simple organisms, the seasonality of sporocarp production, and the absence of records from non-fruiting stages or taxa (Gardes and Bruns 1996). Rapidly advancing molecular methods offer the prospect of a more thorough documentation of diversity. However, while numerous recent studies (O'Brien et al. 2005, Buée et al. 2009, Jumpponen and Jones 2009) have utilized high-throughput sequencing to examine various aspects of fungal biodiversity, these studies have failed to saturate a sampling (rarefaction) curve. Therefore, these studies have not enumerated total fungal diversity in their samples. Furthermore, most of these studies have analyzed modest numbers of samples over small spatial scales, and have not reported detailed characterization of the associated plant communities, thus leaving F:P ratios unaddressed. While they have not directly addressed F:P ratios, recent molecular surveys in a variety of habitats have suggested that fungal diversity is much higher than previously reported. For example, Bueé et al. (2009) recorded an average of 830 fungal operational taxonomic units (OTUs) per site in temperate forest plantations planted with single tree species, while Jumpponen and colleagues found nearly 700 OTUs on Quercus leaf surfaces (Jumpponen and Jones 2009). These observations invite a closer examination of the factors that underlie fungal hyperdiversity.
Fundamental theories to explain community assembly and species coexistence can be divided into those that emphasize deterministic forces (e.g., Lotka-Volterra competition [Gause 1932, Diamond 1978], colonization–recruitment trade-offs [Skellam 1951], resource-ratio theory [Tilman 1982], plant–soil feedback [Bever 1994]) and those that emphasize stochastic factors (e.g., historical contingency [Losos 1994], lottery hypothesis [Sale 1977]). The “unified neutral theory” of Hubbell (Hubbell 2001) has particularly stimulated rigorous comparisons of deterministic vs. stochastic explanations for observed patterns. The greater the numbers of species that coexist without obvious differences in the niche they occupy, the more urgent is the need to search for explanations (Silvertown 2004). Fungal hyperdiversity is thus ripe for careful dissection of coexistence, and the logical starting point is an examination of resource use and evidence for niche partitioning. We recognize that the term niche has implied a wide range of meanings among different authors, contributing to a confused and controversial scientific legacy. We use the term in the sense of the range of biotic and abiotic conditions (axes) for which individuals of a species have fitness greater than zero, as conceptualized by Grinnell (1917), Hutchinson (1957), and others. This may be contrasted with definitions of niche that focus more on the impact of a species on its environment, particularly through resource consumption (Elton 1927, MacArthur and Levins 1967) as discussed by Leibold (1995). However, the “requirement” and “role” aspects of niche can be tightly intertwined in fungi, as we discuss later. With respect to niche partitioning, we are interested in both alpha and beta niches (Silvertown 2004), the alpha niche encompassing environmental axes that vary at a scale smaller than the extent of the focal community, and the beta niche describing responses to habitat gradients over wider spatial scales within a regional context.
Many fungal species are known to have unique environmental requirements and functional roles, examples including the numerous host-specific plant pathogens. A smaller number of studies have also demonstrated niche partitioning among species that comprise complex communities (Dickie et al. 2002). However, due to their invisibility belowground and challenges in measuring appropriate environmental variables at appropriate temporal and spatial scales, we know relatively little about which niche axes may be important for fungal communities in soil. Moreover, the evolutionary timescales over which fungi may diverge in resource acquisition strategies or other traits relevant to coexistence is little known. To address the latter issue, analyses that consider phylogeny or taxonomic hierarchy are needed.
The boreal forest is a compelling ecosystem in which to characterize fungal biodiversity and niche dimensions for three reasons. First, the boreal biome has low plant diversity, suggesting that it should be possible to completely census the richness of both plants and fungi. Second, the northern boreal regions are experiencing some of the most rapid and profound changes due to climate warming, including permafrost thaw and changes in fire regime, both factors that have uncertain implications for the massive stores of carbon in boreal soils (Schuur et al. 2009, Chapin et al. 2010). As primary decomposers and plant mutualists, fungi affect rates of both carbon sequestration and emission from soil and are key players in boreal ecosystem feedbacks to climate change. Yet ecological drivers of the abundance, distribution, and function of fungal species across boreal landscapes are essentially unknown. Third, the boreal forest is one of two biomes that is circumpolar in distribution. Picea mariana (black spruce) forests account for ∼40% of forested land in Alaska, and are the most widespread vegetation type across North America. Hence, characterization of boreal aboveground–belowground relationships can be extrapolated over a wide geographic area.
Here we report the first robust estimate of both fungal and plant species richness at the same sites based on extensive molecular data from replicated and repeated sampling over a wide spatial scale. In this study, we pose the following hypotheses concerning fungal species richness, the form of aboveground–belowground linkages and axes of fungal niche partitioning:
A thorough molecular survey will reveal higher F:P ratios than have been recorded in morphology-based studies of isolates and sporocarps.
The consistent regional F:P ratio reported in prior nonmolecular studies will hold when molecular methods are applied, provided that sampling is sufficient.
Soil horizon will be the strongest measured axis of niche partitioning, based on the importance of this factor in prior studies.
Understory plant community composition will be correlated with fungal community composition due to direct interactions (e.g., mycorrhizae), indirect interactions (e.g., litter chemistry), and shared responses to other environmental drivers (e.g., soil moisture).
Closely related species will often occupy different niches with respect to environmental variables due to rapid evolution of microhabitat preferences. If found, such rapid evolution could reflect a lack of evolutionary–genetic constraints on these traits and/or strong selection favoring character displacement.
Methods
Site description
Our study was carried out in black spruce (Picea mariana) forests in the greater Fairbanks region of interior Alaska, which occurs at the western edge of the Intermontane Boreal Forest ecoregion of the North American continent (Nowacki et al. 2001). It is characterized by small mountain ranges and associated gently sloping uplands, large areas of flat lowlands, and braided rivers with broad floodplains. Interior Alaska has a relatively dry climate (286-mm annual precipitation, on average), is snow-covered for 6–9 months of the year, and much of the surface is underlain by permafrost. Black spruce stands are the most abundant vegetation type, covering roughly 40% of the landscape (Van Cleve and Dyrness 1983), with dominance on poorly drained lowlands and cold, north-facing slopes. Interior Alaska is an area of discontinuous permafrost, with approximately 75–80% of the ground underlain with permafrost (Osterkamp and Romanovsky 1999). Most of interior Alaska, and our study area, escaped the repeated glaciations of the Quaternary; however, Illinoian and Pre-Illinoian glaciers were present at the southern edge of the study area (along the northern flanks of the Alaska Range) (Péwé et al. 1965). Interior Alaska soils show relatively little morphological development. Silt–loam upland soils have developed from loess laid down during the last glacial maximum and from underlying schist bedrock, and floodplain soils have developed from sandy or silt-textured alluvium, mostly of glacial origin (Rieger et al. 1963).
In the present study we sampled 12 of the 151 black spruce sites studied by Hollingsworth et al. (2006, 2008). All sites have a primary overstory canopy of mature black spruce, and are thus representative of the most widespread boreal vegetation type in Alaska and Canada, but vary with respect to tree density, stand age, understory vegetation, mineral soil pH, soil moisture, topography and other biotic and abiotic features. The 12 sites were visited in two successive years, and all soil samples divided into organic and mineral horizon composites; thus, 48 samples form the foundation of subsequent analyses. Site locations are shown in Appendix A: Fig. A1.
Vegetation description and measurement of environmental variables
Vegetation was classified into two communities and two subtypes related to floristic composition of the sites and correlations with environmental factors. These communities are described in detail elsewhere (Hollingsworth et al. 2006). Acidic black spruce/lichen forests are dominated by black spruce, fruticose lichens, and many species of mosses. This community type occurs in both uplands and lowlands of interior Alaska and in areas of low mineral-soil pH (<5.5). Within this community type we find wet and dry sites related to topography and presence of permafrost. Wet acidic black spruce muskeg is an open black spruce subtype that occurs on low-nutrient soils, often in lowland areas of shallow active layer. Black spruce trees are often stunted, and the subtype tends to be dominated by shrubs, sedges, and acidic Sphagnum. The nonacidic black spruce/Rose/Horsetail forest plant community is dominated more by vascular species than the acidic black spruce forest, with the occasional presence of Picea glauca (white spruce). The wet nonacidic black spruce/larch fen is restricted to minerotrophic lowlands and is usually co-dominated by larch (Larix laricina). The dry nonacidic black spruce forest subtype occurs in xeric conditions, mostly in the uplands. Like the acidic black spruce forest, this subtype is dominated by lichens, but the lichen species are different.
Soil sampling and processing for fungal communities
Sites were visited between 25 August and 26 September in 2004 and again in 2005. On each visit to a site, a total of 50 soil cores 1.8 cm in diameter × 20 cm in depth were collected along four parallel transects with approximately 10-m spacing between cores. Subsamples of the Oi (organic) and A (mineral) horizons (Soil Survey Division Staff 1993) from each core were pooled within sites and deposited in 50mL Falcon tubes (BD Biosciences, San Jose, California, USA) then stored at −80°C until lyophilization. The soils were then ground on a ball mill at −20°C, with 0.8-cm steel beads. Genomic DNA was extracted from 1–5 g of soil from each composite sample using the Mo Bio Powersoil kit (MO BIO Laboratories, Carlsbad, California, USA) following the manufacturer's instructions. The soil DNA extracts were normalized to 2.5 ng/μL after Picogreen (Molecular Probes, Eugene, Oregon, USA) quantification.
PCR and sequencing
Polymerase chain reactions (PCRs) utilized 25-μL Amersham Ready-To-Go beads, 0.5-μm primers ITS1-FL and tagged versions of TW13 (Taylor et al. 2008), and the following cycling conditions: initial denaturation at 96°C for 2 min followed by 25 cycles of denaturation at 94°C for 30 s, annealing at 57°C for 40 s and extension at 72°C for 3 min, with a final extension at 72°C for 10 min. For each soil DNA extract seven replicate PCRs were performed and pooled. We utilized a low number of cycles and a large number of replicate PCRs to reduce chimera formation and stochastic amplification biases. We applied a molecular-tagging strategy to mark PCR products from various sources with DNA tags, which were then pooled before library sequencing (Taylor et al. 2008). To minimize the cloning of primer-dimers and other short inserts, 100 μL of the pooled fragments were size-fractionated over Chroma Spin 400 columns (BD Biosciences, San Jose, California, USA), then concentrated through DNA Clean and Concentrator-5 columns (Zymo Research, Irving, California, USA). The templates were quantified on a Nanodrop spectrophotometer and normalized to 25ng/μL prior to ligation. The pooled PCR products were then cloned into the Invitrogen PCR4-TOPO (Life Technologies Corporation, Carlsbad, California, USA) vector. The resulting PCR libraries were shipped frozen to the Broad Institute, Cambridge Massachusettss (USA), where transformation, plating, colony picking, Templiphi reactions, and sequencing were carried out on automated equipment, using M13 forward and reverse vector primers. These molecular methods have been partially described elsewhere (Taylor et al. 2007, 2008, Geml et al. 2008, 2009). Site coordinates and numbers of soil cores and sequences generated are given in Appendix A: Tables A1 and A2.
Sequence analyses
Our approaches to sequence bioinformatics have also been described in detail elsewhere (Taylor et al. 2008, 2010, Geml et al. 2009, 2010, Taylor and Houston 2011). In brief, sequence data obtained for both strands were edited and assembled for each clone using CodonCode Aligner version 1.3.4 (CodonCode Corporation, Centerville, Massachusetts, USA). Primer tags were identified, bases with phred scores below 20 were masked, and clone orientations were corrected using Perl scripts (available online).6 We then removed primer and vector sequences and isolated the internal transcriber space (ITS) region of each clone by locating the positions of ITS1-F and ITS4 (White et al. 1990, Gardes et al. 1991) in multiple sequence alignments generated using ClustalW (Thomson et al. 1994). The dirty ends of ITS sequences were removed using the program EMBOSS TrimSeq (Rice et al. 2000), and sequences with greater than 2% Ns remaining (i.e., base calls of such poor quality that they are undetermined) were deleted from the data set. Sequences were grouped into operational taxonomic units (OTUs) using the single-linkage genome assembly program Cap3 (Huang and Madan 1999) with the following non-default settings: overlap percent identity = 96.9; match score factor = 5; maximum overhang percent length = 60; clipping range = 6. No simple ITS sequence identity threshold can perfectly distinguish species boundaries across diverse fungi since the molecular evolution of the ITS region varies across fungal lineages. However, a number of focused phylogenetic studies suggest that a threshold of 97% may lump discrete sister species (Kindermann et al. 1998, Horton 2002, Geml et al. 2006, 2009); on the other hand, a more rigorous threshold would begin to artificially divide species due to sequencing errors and intraspecific variation (O'Brien et al. 2005, Quince et al. 2009, Kunin et al. 2010). Thus we view 97% as the best available approximation when a single standard must be applied to large data sets spanning wide fungal diversity. Furthermore, this threshold is likely to be conservative with respect to diversity of biological species. To evaluate whether our 97% sequence identity threshold and single-linkage clustering using Cap3 accurately captured species-level boundaries and whether our OTU counts might have been inflated by pseudogenes or chimeras, we carried out detailed phylogenetic analyses of selected OTUs. We independently analyzed OTUs that fell into three distinct genus-level clusters, one representing the Basidiomycota (Cortinarius), one the Ascomycota (Leptodontidium), and one the former Zygomycota (Mortierella). For each phylum, these genera represent one of the most species-rich clusters of OTUs seen in our data set, which were chosen because these are the most likely to exhibit artifactually high numbers of OTUs. We searched GenBank for close matches to all OTUs in each of the three groups, created multiple sequence alignments using MAFFT EINSI (Katoh et al. 2002) and/or MUSCLE (Edgar 2004) and inferred evolutionary relationships using maximum likelihood with rapid bootstrap under the GTRCAT model in RAxML 7.0.4 (Stamatakis et al. 2005, 2008).
We ran all sequences through the UCHIME (Edgar et al. 2011) chimera-detection program using the de novo option and including close relatives from GenBank, as implemented on our public web tool (available online).7 All sequences with scores above 3.0 were discarded as likely chimeras, while all sequences with scores between 0.8 and 3 were manually investigated via construction of multiple alignments containing all most closely related sequences derived from separate ITS1 and ITS2 BLAST searches (Taylor and Houston 2011). Several additional sequences were discarded. In all cases, chimeric sequence types were of very low abundance (1–5 clones), all but one being recovered from a single soil sample.
The best representative sequence from each OTU was roughly identified through nucleotide BLAST (Altshul et al. 1997) searches of our local fungal ITS database (footnote 2) that is updated from the GenBank nr nucleotide database weekly (Taylor and Houston 2011), with and without environmental (unidentified) sequences included. If the match with the highest bit score was an unidentified sequence, both the top identified and unidentified accessions are given in the Supplement: file 1, Blast_IDs.csv. Otherwise, only the top identified accession is given. These identifications allowed us to assign guild membership to many OTUs. We used the following criteria to evaluate the level of identification for each OTU. A score of A, indicating identification to the species level, was given when a hit with >97% similarity was found to a well-identified fungus. A score of B was given when no match >97% was found to a well-identified fungus, but a match >97% was found to another uncultured/environmental sequence. A score of C was given when the best match to a well-identified or uncultured sequence was between 90 and 96.9%. A score of D was given, indicating a poor identification, when only matches below 90% identity were found, or only overlaps of less than 200 base pairs (bp) were found. These identification levels are listed in the Supplement: file 1, Blast_IDs.csv. The latter scenario usually indicates that BLAST has found significant similarity only to the highly conserved 5.8S region. Note that we likely overestimate the proportion of species-level matches for two reasons: first, many of the hits with >97% identity overlapped only a portion of the query sequence; second, we accepted in category A 97% hits to fungal isolates or sporocarps that had a genus, but not a species name (e.g., Phialophora sp.), and thus were not “fully identified” according to previous criteria (Nilsson et al. 2009). The representative sequence for each OTU has been submitted to GenBank under accessions KF617225–KF618373. The full OTU-by-site matrix is given in Supplement: file 2, OTU_by_site_matrix.csv.
Statistical analyses
Species-accumulation curves and bootstrap estimates of total richness were computed using EstimateS, version 7.5 (Colwell 1997). Curves are based on means of 50 randomized subsamplings. We performed additional rarefaction analyses by randomly adding each of the 12 sites 50 times and tabulating numbers of vascular plant species and fungal OTUs observed, then calculating the corresponding fungus-to-plant (F:P) ratios. Our estimate of the minimum ratio utilized the 95% confidence interval (CI) upper bound for plant species combined with the 95% CI lower bound for fungal species. The maximum ratio was calculated similarly, using the lower CI for plants and upper CI for fungi.
To compare community composition among sites and samples, we removed fungal taxa occurring in fewer than two samples, relativized the abundances by site totals, and calculated dissimilarity among the 48 samples using the abundance-based version of the Bray-Curtis index. We then tested whether fungal communities differed across soil horizon, sampling year, and the four vegetation types using multiple-response permutation procedures (MRPP) (Berry et al. 1983). This method calculates the dissimilarity among samples grouped by a categorical variable and compares this value to a distribution generated by randomly permuting species observations across samples. Relationships between individual OTUs and selected categorical environmental factors were tested using “indicator species analyses” (Dufrene and Legendre 1997). In this approach, an index that combines frequency and abundance of an OTU in a target group of samples relative to its overall frequency and abundance is compared to a randomized distribution. OTUs that are more frequent and abundant (i.e., concentrated) in a particular group of samples (e.g., organic horizon samples) than expected by chance are considered to be indicator species for that environment. To visualize relationships between fungal community composition and measured environmental variables, we ordinated the 48 site–date–horizon combinations using nonmetric multidimensional scaling (NMS) (Kruskal 1964), a method of indirect gradient analysis (Whittaker and Niering 1965) that makes relatively few assumptions about the forms of species–environment relationships (McCune et al. 2002). We started with a random configuration to estimate the minimum supported dimensionality. Using this dimensionality, a final solution was estimated via 50 runs started from random seeds and 500 Monte Carlo resamplings to test for significance. We performed a parallel ordination of plant community composition and stored the resulting plant axes as a way to summarize vegetation variation across the sites. We carried out principal-components analysis (PCA) of the 12 sites in order to reduce the 76 environmental variables measured at each site (Hollingsworth et al. 2006) to a few sets of intercorrelated variables aligned with axes that best explain the variation among sites. Correlations between fungal species ordination axes and potential drivers were evaluated in two ways. First, we examined the Kendall correlations between each individual environmental variable and fungal species ordination axes. Second, the site scores derived from the environmental PCA and vegetation NMS ordinations were used to create a second environmental matrix that was compared to fungal ordination axes, again by Kendall correlations. Calculations were carried out using PC-Ord 4.0 (McCune and Mefford 1999).
Results
Fungal diversity
We initially obtained 36 864 clone sequences, and then applied rigorous quality, length, taxonomic, and chimera filters, reducing the final data set to 28 884 clones. When clustered at 97% sequence identity across the internal transcriber space (ITS) region, these clones yielded 1002 operational taxonomic units (OTUs). Rarefaction analysis revealed a distinct inflexion with a slope that neared zero as our full sample size was reached (Fig. 1A). Further, the Chao 1 estimator, which adds a calculated number of “unseen” species to arrive at an estimate of true richness, was also asymptotic, reaching a maximum of 1044 OTUs. This saturation of the rarefaction curve indicates that we obtained a complete accounting of fungal OTU richness in these samples, which has not previously been accomplished in a forested ecosystem, to our knowledge. Additionally, only 117 of these 1002 OTUs were sequenced only once (singletons), meaning that our results are not driven by a long “tail” of exceptionally rare taxa that might represent polymerase chain reaction (PCR) and/or sequencing artifacts (Kunin et al. 2010).
figure image
Figure 1
Open in figure viewer
PowerPoint
(A) Saturation of the fungal rarefaction curve in black spruce forests of interior Alaska, USA. The x-axis represents subsets of the total number of clones sequenced (28 884) for fungi, or the number of sites sampled (12) for plants. (B) Fungus–plant ratios as a function of the number of sites. Notice that the ratio increases only slightly over the first five sites and is nearly constant when adding sites 9–12.
The observed hyperdiversity of the fungal community is in stark contrast to the low plant diversity found in interior Alaskan Picea mariana forests. Only 60 of the 97 plant species documented at the 12 sites (Appendix A: Fig. A1) are vascular, giving a fungus-to-vascular plant multiplier of approximately 17:1. This result confirms our first hypothesis that molecular methods would lead to higher F:P ratios than reported in prior nonmolecular studies. The F:P ratio varied from site to site, with a mean of 16.0 (± 6.1 SD) (Appendix A: Table A1). However, when sites were added sequentially via rarefaction, the mean ratio rises from 14.5 for a single site to 16.35 with five sites, and is essentially constant at 16.8 with eight or more sites (Fig. 1B). Hence, the estimated F:P ratio was consistent across the 200-km study area. Furthermore, our sampling of sites and DNA sequences was adequate to reach a stable plateau in the F:P ratio. These findings lend support to our second hypothesis of a consistent regional F:P ratio, at least within our study area and forest type.
Our focused phylogenetic analyses of three of the most diverse genus-level clusters of OTUs in our data set demonstrate a strong correspondence between OTUs and species-level phylogenetic groups. For each genus, nearly all of our OTUs fell into distinct clades, and many were closely related to sequences obtained in other studies. In the case of Cortinarius, a mushroom-forming ectomycorrhizal basidiomycete, most OTUs fell into clusters together with sequences derived from sporocarps, with each cluster containing a different taxonomically defined species (Fig. 2). In Leptodontium, a group of root-associated dark septate endophytes (DSE) in the Ascomycota, several of our OTUs were closely related to well-characterized strains, but many other sequences were closely related only to other uncultured/environmental sequences (Appendix B). Similar patterns were seen in our analysis of Mortierella, a diverse group of saprotrophic soil “sugar fungi” that have historically been placed in the zygomycetes (Appendix B). With only two exceptions across 89 OTUs, different OTUs did not belong to a shared species-group, with terminal species-group defined by (1) branch length equivalent to 3% or greater from the nearest taxon, and/or (2) greater than 70% bootstrap support. Thus, pseudogenes, oversplitting, or chimeric artifacts appear unlikely to explain the great species richness reported in this study.
figure image
Figure 2
Open in figure viewer
PowerPoint
Phylogenetic tree of Cortinarius-like OTUs (operational taxonomic units). This maximum-likelihood tree was estimated with RAxML 7.04 (Stamatakis et al. 2005, 2008); bootstrap support values above 70% based on 1000 replicates are shown. Notice that most Cortinarius-like OTUs (boldface type) are close matches to sequences reported from other studies, and nearly every OTU falls out in a distinct clade defined by a sporocarp sequence with a different specific epithet (i.e., a different species).
figure image
Figure 2
Open in figure viewer
PowerPoint
Continued.
The large majority (67%) of the 1002 OTUs had a best BLAST match, based on bit scores, to an uncultured/environmental sequence, rather than a well-identified sequence from a fungal isolate or sporocarp. Only 28% of the OTUs could be considered matched to the species level (score A). Another 20% of the OTUs had a close match, but only to another uncultured/environmental sequence (score B). A further 40% were only moderately related to any known sequence (score C), and 12% were distantly related to any known sequence (score D). Based on the best BLAST matches, we assigned each OTU to its most likely phylum. The Ascomycota and Basidiomycota dominated the data set, and were nearly equal in abundance. The Ascomycota accounted for 55.2% of the OTUs and 48.4% of the clones, while the Basidiomycota accounted for 39.4% of the OTUs and 48.3% of the clones (Table 1). The Glomeromycota and Mortierellomycotina accounted for 2% and <1% of the clones, respectively. The Helotiales, including many putative ericoid mycorrhizal fungi and dark septate endophytes (DSE), were the most abundant and diverse order of Ascomycota, accounting for 180 OTUs and 34.5% of Ascomycota clones. Similarly, among the Basidiomycota, the majority of OTUs were related to putative ectomycorrhizal taxa. Summing across both phyla, OTUs with top matches that are suspected to be ectomycorrhizal comprised 12 312 clones, or 42.6% of the data set. Thus, the majority of taxa uncovered in this study appear to be plant associated in some capacity. We note that typical wood and litter decay basidiomycetes that are known from sporocarps in interior Alaska (e.g., Agaricus, Clitocybe, Marasmius, Mycena, Pholiota, Pleurotus, and Pluteus) were present at low abundances in the data set.
Table 1. Representation of fungal phyla in this study in black spruce forests of interior Alaska, USA.
table image
Structure of fungal communities
The coexistence of so many fungal species in these sites motivates investigation of fungal habitat preferences within and among the study sites. We investigated how the soil fungal communities are structured with respect to the aboveground plant communities and a suite of environmental variables using nonmetric multidimensional scaling (NMS). Ordinations and multiple response permutation procedure (MRPP) analyses failed to detect a difference between fungal communities in 2004 compared to 2005. In the MRPP tests, soil horizon had by far the largest effect size with respect to community composition (Table 2), in agreement with our third hypothesis. The strong distinction in community composition between horizons was also seen in the NMS ordination of sites (Fig. 3). Because the fungal communities of these two soil horizons are so distinct, we carried out further analyses to investigate their structure independently.
figure image
Figure 3
Open in figure viewer
PowerPoint
Strong horizon partitioning of fungal communities illustrated by NMS (nonmetric multidimensional scaling) ordination. This analysis included all 48 samples, representing two collection years and two soil horizons from each of the 12 sites. Several examples of the same site and horizon sampled in successive years are indicated with ovals, illustrating the year-to-year consistency of fungal communities within a site.
Table 2. Multiple-response permutation procedure tests for differentiation of fungal communities.
table image
Considering soil chemistry and site factors, the occurrence and abundance of fungal taxa in the organic horizon were most strongly related to soil pH (Appendix C). A suite of soil and site factors co-vary with pH and were thus strongly related to soil PCA axis 2, including concentrations of Na, Mg, and Ca, the percentages of clay, sand and silt, elevation and topographic position (Appendix C). Vegetation variables that were strongly correlated with organic horizon fungi included tree density, the proportions of seedlings vs. trees, and the percentage of graminoid species. In contrast, mineral-horizon fungal community structure was more strongly correlated with site moisture and its covariates on PCA soil axis 1, including percentage C, organic soil depth, cation exchange capacity, and bulk density (Appendix C). Parameters describing tree size and density (tree height, diameter at breast height, and stand basal area) were the vegetation factors most strongly related to mineral-horizon fungal composition. These aspects of stand structure are highly correlated with site moisture.
We directly compared the composition of plant and fungal communities using Mantel tests of correlations between community Bray-Curtis dissimilarity matrices. The test was significant for organic-horizon fungi (P = 0.004), with a correlation of r = 0.43 between the plant and fungal matrices, but was weakly correlated and nonsignificant for mineral-horizon fungi (r = 0.09, P = 0.29). The Mantel tests are consistent with the MRPP tests, in which the four site classes based on vegetation types were a significant predictor only for the organic-horizon fungal community (Table 2). Hence, our results demonstrate horizon-dependent variation in the nature of linkages between structures of aboveground plant and belowground fungal communities.
More surprisingly, indicator-species analyses showed that it is often species belonging to the same genus and broad functional guild (e.g., ectomycorrhizal, saprotrophic, pathogenic) that have divergent preferences with respect to soil horizon or plant community type (Table 3). The most abundant taxon in the entire data set, OTU221 (Cortinarius collinitus, an ectomycorrhizal species), was identified in 1321 clones from 25 of the 48 site–year–horizon combinations, but was found almost exclusively in the organic horizon (indicator value = 92.3, P = 0.0002). In contrast, another Cortinarius species, OTU442, was a strong indicator for the mineral horizon (indicator value = 25, P = 0.019) (Appendix C). Many additional examples of divergent habitat preferences among closely related taxa were apparent, such as Piloderma OTU320 in comparison to OTU322 (both ectomycorrhizal; wet nonacidic vs. dry acidic preferences), Leptodontidium OTU173 vs. Leptodontidium OTU177 (both DSE; mineral vs. organic horizon preferences) and Cryptococcus OTU246 in comparison to OTU247 (saprotrophic; organic- vs. mineral-horizon preferences). Note that this pattern of niche differentiation spans major functional guilds, including several types of mycorrhizae, saprotrophs, and parasites. Niche differentiation among closely related species was also seen in each of the three predominant phyla recovered in this study, the Basidiomycota, Ascomycota, and the “basal fungal lineages” (primarily Mortierellomycotina).
Table 3. Indicator species analyses, showing habitat preferences for selected taxa, with significant indicator values demonstrating niche differentiation among closely related taxa.
table image
Discussion
We report here a much higher fungus-to-plant (F:P) ratio than has been seen in prior nonmolecular studies, and the first molecular saturation of a rarefaction curve for soil fungi. Our data support the idea that F:P ratios may be predictable on a regional scale. To the extent to which this first saturated F:P ratio can be extrapolated globally, it suggests far higher global numbers of fungal species than other recent estimates. Moreover, our estimate of the regional F:P ratio, though stable, is likely to be low because many taxa will not be represented in a soil census. We also find strong structuring of fungal communities related to environmental variables and understory plant community composition. Perhaps most surprisingly, species that show the strongest and most divergent habitat preferences are often closely related.
Expanding the global estimate of fungal diversity
Next-generation sequencing studies have typically discarded rare sequences and have often failed to distinguish artifacts such as non-fungal sequences and chimeras, and thus have generally not attempted to enumerate total fungal species richness. We have overcome these issues using an alternative molecular approach combined with rigorous bioinformatics steps and large-scale sampling. Instead of relying on next-generation sequencing of short amplicons that are less amenable to phylogenetic analyses, taxon identification (Porter and Brian Golding 2011), and chimera detection, we amplified and Sanger-sequenced a relatively long ∼1200 bp region that spans the entire highly variable nuclear ribosomal internal transcribed spacer (ITS) and extends roughly 700 bp into the large subunit (LSU). The ITS fragment is ideal for species discrimination (Schoch et al. 2012), while the LSU portion provides significant resolution for deeper-level phylogenetic analyses. This approach reduces amplification of DNA from dead organisms, since most soil DNA is quickly degraded to short fragments (Willerslev and Cooper 2005), and improves detection and exclusion of chimeras and non-fungal OTUs. Furthermore, our comparisons of OTU clustering using the genome assembly program Cap3 (Huang and Madan 1999) to phylogenetic groupings inferred from tree-building exercises in three diverse genera support the claim that clusters based on 97% sequence identity are reasonable approximations of species-level divisions. Most importantly, these analyses suggest that we are not over-splitting, and thereby overestimating fungal diversity.
Methods that have been used to extrapolate the richness of hyperdiverse groups have included body-size frequency distributions, latitudinal gradients, species–area relationships, host-specificity ratios, time–species accumulation curves, and expert opinions (Mora et al. 2011). Body-size relationships are not applicable to fungi due to their fragmented growth patterns and mostly unknown biomasses and sizes. Currently, we lack adequate data to apply latitudinal gradients, species–area relationships or host-specificity ratios to fungi. Combining opinions of taxonomic experts has been attempted for fungi (Hawksworth and Rossman 1997), but is suspect due to the bias toward species with macroscopic sporocarps. Mora et al. (2011) recently proposed a general mathematical method that extrapolates numbers of species from the more stable numbers of taxa at higher ranks, i.e., families through Kingdoms, while simultaneously modeling the accumulation of higher ranks over time (i.e., the asymptotic completeness of higher level taxonomy since Linnaeus). Their method resulted in an estimate of 616 000 species of terrestrial fungi worldwide.
There are compelling reasons why this type of extrapolation may perform poorly for fungi. The “known” species of fungi on which the extrapolation was based included no molecular data, thereby discounting numerous cryptic species. Furthermore, higher taxonomic ranks do not yet appear stable in the Eumycota, as major new classes and even phyla have recently been described (Jones et al. 2011, Rosling et al. 2011). It has been argued that using ratios of diversity between major taxonomic groups (i.e., fungi vs. plants) is a crude and unreliable approach to estimating richness of a target group (May 1991, Mora et al. 2011). However, our results support our hypothesis and contribute to growing evidence that F:P ratios are consistent at regional scales in at least some ecosystems. For example, in a meta-analysis, diversity of tree species and mushroom-forming fungi were found to be strongly correlated (Schmit et al. 2005), as were plant species richness and fungal richness in soil across 24 grassland plots in Japan (Sugiyama et al. 2008).
If F:P ratios are consistent at regional scales, the next key question is whether these ratios vary across habitats or biomes. To arrive at the 1.5 million species estimate, it was argued that the ratio observed in the temperate United Kingdom could be applied globally (Hawksworth 1991). However, there are reasons to suspect that such ratios may not be universal. For example, diversity of ectomycorrhizal fungi appears to peak in temperate and boreal regions and decline in tropical and arctic regions (Tedersoo et al. 2012). On the other hand, some guilds of fungi, especially plant endophytes, have been shown to increase in diversity from high latitudes to the tropics (Arnold and Lutzoni 2007). Whether endophyte : plant ratios vary along this gradient is not yet clear.
When our 17:1 F:P ratio is combined with the current estimate of vascular plant diversity of 352 000 (Paton et al. 2008), it would suggest a global fungal richness of 6 million species, as opposed to the 616 000 recently suggested by Mora et al. (2011). Moreover, our estimate of the fungus-to-plant ratio at our sites is conservative for several reasons. First, single-linkage clustering of fungal ITS sequences at 97% identity often lumps sequences belonging to different, closely related species (Kindermann et al. 1998, Peintner et al. 2004, Geml et al. 2009). Second, our census of soil does not encompass the numerous fungal species that occur only in aboveground habitats, such as the diverse fungal species of the Laboulbeniales (Weir and Hammond 1997) and Entomophthorales (Humber 1989), which may number as many species as certain of their host groups among the Insecta, the ascomycetous yeasts that live primarily within the digestive tracts of insects (Blackwell and Jones 1997, Suh et al. 2005) and other animal hosts, and the hyperdiverse lichens and associated lichenicolous fungi (Esslinger and Egan 1995). Even among soil fungi, our census overlooked some guilds; we did not sample coarse woody debris, fresh litter, or decaying moss, each of which is known to host specialist fungi (Gilbert and Sousa 2002, Lindahl et al. 2007, Davey et al. 2012), nor deep soil. Third, although we utilized primers and barcodes that we have previously shown to have minimal taxonomic biases (Taylor et al. 2008), there are no fungal-selective primers that do not miss some lineages (Taylor and McCormick 2008). Regardless of taxa we may have missed, our findings clearly imply that the estimate of Mora et al. (2011) is likely to be far too low.
A key issue in interpreting the high F:P ratio we report concerns the origins and identities of the fungal species we observed. Over 70% of our OTUs could not be identified to species by way of BLAST matches to known species. Our unidentified species span a wide range of precision of identification. Many appear to represent species that have not been previously sequenced, but that belong to well known genera (some class B and C identifications). However, many also appear to belong to families and even deeper taxonomic divisions that lack previously sequenced representatives. There are few known families without a sequenced representative, implying that some of our novel OTUs may be members of previously unknown higher taxa. Some of the poorly identified class D OTUs were also rare, which might suggest a higher likelihood that they represent artifacts rather than real taxa. However, this may not necessarily be the case. We studied in considerable detail a highly novel sequence type for which we have only found two clones out of tens of thousands sequenced at the same sites. Our analyses of functional attributes of these ribosomal sequences suggest that they represent a legitimate taxon (Glass et al. 2013). Moreover, many of our class B and C sequences are now known to belong to the new class Archaeorhizomyces (Rosling et al. 2013). Lastly, some of the class C and D identifications were derived for high abundance OTUs, making an artifactual origin unlikely.
As expected, well-studied macroscopic basidiomycete genera such as Cortinarius (Fig. 2), Tricholoma, Russula, Leccinum, and Alnicola had high proportions of species matches, as did frequently isolated “weedy” fungi in the genera Neurospora, Penicillium, Trichoderma, and Verticillium. Counter to these trends, however, were several OTUs that appear to belong to the Agaricomycotina, yet were distantly related to any previously sequenced fungi. The top matches for some of these OTUs belonged to the genera Clavulina, Exidiopsis, Sebacina, Sistotrema, Piloderma, Mycena, Hydnopolyporus, Peni-ophora, Inocybe, Hydnum, Lentinula, Hygrophorus, and Hebeloma. Some of these taxa have microscopic and/or resupinate fruitbodies—groups that are noted for incomplete taxonomic knowledge (Gardes and Bruns 1996, Kõljalg et al. 2003, Smith et al. 2007). On the other hand, the latter four genera are surprising members of the list, given that they produce relatively large and well-known sporocarps. Inocybe is similar in this regard, but has previously been shown to have extremely high species diversity at high latitudes and a rapidly evolving ITS locus (Ryberg et al. 2008, Geml et al. 2012, Timling and Taylor 2012).
The immense fungal diversity reported here appears to be due to novel (unsequenced) diversity at all taxonomic levels, but is unlikely to be due principally to a “rare biosphere” or sequence artifacts. Many taxonomically described fungi have no corresponding molecular barcode sequence available (Hibbett et al. 2011). Furthermore, new species descriptions continue to accelerate for fungi (Hawksworth 2012), despite a dearth of trained taxonomists. These observations call for increasing allocation of resources to taxonomic work on fungi, as well as support for standardized barcode submission to go along with all new species descriptions.
Structure of fungal communities
All our sites were completely dominated by a Picea mariana overstory, yet we found strong structuring of fungal communities in relation to nearly every abiotic and biotic variable examined. In our view, the consistency of community composition from year to year in the same site, juxtaposed with strong distinctions among sites, principally in accord with site pH and moisture status, argues against either technical issues (e.g., barcode biases) or stochastic spore dispersal processes in driving the patterns we found. Instead, our results strongly support our hypothesis that soil horizon would be an important environmental variable driving fungal community composition. The partitioning by soil horizon is so strong that the arrays of fungi present in two samples collected over 100 km apart, but from the same horizon, are more similar than are the arrays of fungi in different horizons collected only centimeters apart, well within the distance that can be spanned by mycelium from a single fungal individual. Thus, these strong species horizon preferences demonstrate partitioning of the alpha niche (Silvertown 2004), i.e., well within the spatial scale of single populations.
Other environmental variables were measured at the site level, providing contrasts among sites, which is of more relevance to beta niches. Our findings of strong community correlations with pH and site moisture agree with other studies of fungal communities in soil (O'Dell et al. 2000, Toljander et al. 2006). Furthermore, these factors are also strongly correlated with the composition of the understory plant communities at these sites (Hollingsworth et al. 2006). Some of these factors likely also influence fungal community composition and niche partitioning at much finer spatial scales, but this cannot be assessed with the present data set due to our within-site pooling protocol, which was required to achieve the sampling intensity and resultant species saturation that were the overriding objectives.
Our data suggest strong yet complex aboveground–belowground linkages. Studies that enumerate the F:P ratio at a particular site implicitly average across plant species that host many fungi and those that host few fungi, as well as fungi that are narrow host specialists and those that are broad host generalists. We found an overwhelming dominance of fungal OTUs that are likely to interact strongly with plants as mycorrhizae, endophytes, or pathogens. Yet the patterns of linkage differed between the two soil horizons. Because these elements are confounded, we cannot clearly infer direct or indirect effects of plants on fungi or vice versa. It is interesting, however, that organic-horizon fungi were more closely correlated with understory vegetation than were mineral-horizon fungi. Fine-root density is orders of magnitude greater in the organic horizon of Picea mariana forests in interior Alaska (Ruess et al. 1996, 2006). One plausible scenario, then, is that the roots or litter from the understory vegetation exert a stronger effect on fungi in the organic horizon. These understories are dominated by ericaceous shrubs (Ledum and Vacinnium spp.), and a significant fraction of the Helotialean ascomycete diversity we uncovered may form ericoid mycorrhizas and/or dark septate endophytes. On the other hand, it is likely that strong plant–fungal interactions continue into the mineral horizon, despite the lack of Mantel correlations in community matrices, as some of the mineral-horizon indicator species are ectomycorrhizal or ericoid mycorrhizal species. Ectomycorrhizal Picea mariana roots obviously occur at all sites, so this factor may not structure differences among sites, even while exerting strong influences on fungal ecology. Upland sites that lack Picea mariana host yet a different constellation of fungal communities (Taylor et al. 2010); it is important to recognize that conclusions about the relative importance of various structuring agents are scale dependent.
Our data reveal a novel pattern in the striking degree to which co-occurring, closely related taxa belonging to the same broad guild differ in preference for soil horizon or plant community type (e.g., acidic vs. nonacidic, wet vs. dry). This pattern is seen across trophic guilds, including various root associates, parasites, and saprotrophs (Table 2). Silvertown (2004) proposes four criteria to evaluate the role of niche partitioning in species coexistence: first, establishing that within-species competition is stronger than among-species competition; second, establishing with tests against a null model that species segregate along environmental gradients; third, demonstration of trade-offs in performance related to proposed niche specialization; and, fourth, demonstration of a change in niche occupancy when putative competitors are added or removed. Our evidence simply hints that the second criterion may be fulfilled, i.e., that species are segregated with respect to resources and other environmental variables. While there are challenges with respect to feasibility of field studies, efforts to test these criteria might return rich rewards in the study of fungal community ecology. Another productive avenue might be phylogenetic analyses of community assembly. For example, documentation of phylogenetic clustering of habitat preferences can be interpreted as resulting from the influence of habitat filters, whereas phylogenetic overdispersion of habitat preferences can be interpreted as resulting from the influence of competitive exclusion (Webb et al. 2002). However, other interpretations have also been suggested (Mayfield and Levine 2010). While much additional work is warranted, our findings strongly suggest that individual fungal species often carry out different functional roles in the ecosystem, despite their enormous diversity. Furthermore, ours is a minimal estimate of niche partitioning, since only soil horizon and plant community type were considered here. Because fungi must acquire food from their environment by growing into or onto their preferred substrate (e.g., wood), the distinction between environmental requirements and functional roles becomes clouded in fungi. For example, by growing into dead wood of a preferred host species, fungi are able to decompose the wood and radically alter the ecosystem. Our results point to fine-scale variation in fungal-niche axes that relate primarily to environmental requirements (e.g., pH, moisture), but several that also imply functional roles, particularly the axes of soil horizon and vegetation community type.
Conclusions
With accelerating extinction and reorganization of biodiversity on the planet, our ignorance of even its approximate magnitude is cause for concern. Fungal biodiversity hotspots are currently unknown, and we cannot predict either patterns of extinction or new epidemics. In this work, we demonstrate a consistent F:P ratio in the boreal forest that is much higher than prior non-molecular estimates. A critical need is to carry out comparable, thorough, and rigorous studies in other biomes to quantify patterns of variation in fungus-to-plant ratios and to better understand global drivers of fungal distribution and community assembly. Our results suggest that <2% of fungal species have been described, implying that the Fungi are equaled only by the Insecta with respect to Eukaryote diversity, and that closely related fungi differ in niche axes related to their roles in the environment.
Acknowledgments
We thank József Geml, Ian Herriott, and Ina Timling for technical assistance, and Sarah Youssof, Meredith Blackwell, and John Taylor for helpful comments on the manuscript. Lisa Green and Scott Anderson assisted with sequencing at Broad. Shawn Houston, and the UAF Life Sciences Informatics core provided critical computational support. This study was supported by the National Science Foundation through awards EF-0333308, ARC-0632332, and DEB-0620579 and by the USDA Forest Service, Pacific Northwest Research Station, through agreement number RJVA-PNW-01-JV-11261952-231. This paper is dedicated to the memory of Michael G. Booth, an inspiring scientist and friend.
Supplemental Material
Appendix A
Site coordinates, sampling information, and F:P ratios across sites (Ecological Archives M084-001-A1).
Appendix B
Phylogenetic trees of Leptodontidium and Mortierella OTUs (Ecological Archives M084-001-A2).
Appendix C
Graphical and tabular views of fungal habitat preferences as revealed by ordinations and environmental correlations (Ecological Archives M084-001-A3).
Supplement
Three files containing BLAST identifications for fungal OTUs and abundances across samples (Ecological Archives M084-001-S1).
Supporting Information
Literature Cited
www.borealfungi.alaska.edu
www.borealfungi.uaf.edu
Corresponding Editor: D. M. Rizzo.
Citing Literature
Volume84, Issue1
February 2014
Pages 3-20
Figures
References
Related
Information
Recommended
Integrating species traits into species pools
Marko J. Spasojevic,
Christopher P. Catano,
Joseph A. LaManna,
Jonathan A. Myers
Ecology
Evidence of dispersal limitation in soil microorganisms: Isolation reduces species richness on mycorrhizal tree islands
Kabir G. Peay,
Matteo Garbelotto,
Thomas D. Bruns
Ecology
Experimental plant communities develop phylogenetically overdispersed abundance distributions during assembly
Eric Allan,
Tania Jenkins,
Alexander J. F. Fergus,
Christiane Roscher,
Markus Fischer,
Jana Petermann,
Wolfgang W. Weisser,
Bernhard Schmid
Ecology
The Fungi: 1, 2, 3 … 5.1 million species?
Meredith Blackwell
American Journal of Botany
Evolutionary history of mycorrhizal symbioses and global host plant diversity
Mark C. Brundrett,
Leho Tedersoo
New Phytologist
Download PDF
Back
The Ecological Society of America Logo
© 2024 Ecological Society of America. All rights reserved.
ADVERTISING
MEDIA KIT
ABOUT THE ESA
ESA Headquarters
1990 M Street, NW
Suite 700
Washington, DC 20036
phone 202-833-8773
email: [email protected]
Additional links
ABOUT WILEY ONLINE LIBRARY
Privacy Policy
Terms of Use
About Cookies
Manage Cookies
Accessibility
Wiley Research DE&I Statement and Publishing Policies
HELP & SUPPORT
Contact Us
Training and Support
DMCA & Reporting Piracy
OPPORTUNITIES
Subscription Agents
Advertisers & Corporate Partners
CONNECT WITH WILEY
The Wiley Network
Wiley Press Room
Copyright © 1999-2024 John Wiley & Sons, Inc or related companies. All rights reserved, including rights for text and data mining and training of artificial technologies or similar technologies.
Wiley Home Page
| biology | 927912 | https://sv.wikipedia.org/wiki/Vit%20silkesapa | Vit silkesapa | Vit silkesapa (Callithrix jacchus), vitörad marmosett eller marmosettapa som den också kallas, är en art i släktet silkesapor, (Callithrix), i familjen kloapor (Callitrichidae).
Kännetecken
Den vita silkesapan blir 16 till 21 centimeter lång (huvud och bål) och svansen är med en längd av 24 till 31 centimeter betydlig längre. Den genomsnittliga vikten anges i olika källor med 236 eller 322 gram för honor och med 256 eller 318 gram för hannar. Pälsen färg är huvudsakligen spräcklig gråbrun med några svarta hår och på djurets rygg finns ljusare gulaktiga tvärstrimmor. Huvudet är brunaktig och vid öronen finns karakteristiska vita tofsar. Ansiktet saknar hår och ovanför ögonen finns en vit fläck. På svansen förekommer flera ljusa ringar. Extremiteterna är jämförelsevis korta. Liksom alla andra kloapor har djuret klor vid tårna istället för naglar (undantag tummen).
Utbredning och habitat
Arten levde ursprungligen bara i Brasiliens nordöstra delar. Utbredningsområdet sträckte sig från delstaterna Maranhão och Piauí till floden Rio São Franciscos norra strandlinje. Idag förekommer djuret även i andra regioner i Brasilien. Habitatet utgörs av olika sorters skogar. De lever bland annat i den stäppliknande buskskogsbeväxta caatingan som förekommer även i blöta skogar längs Atlantens kustlinje. De har lätt att anpassa sig och finns även på odlingsmark eller i stadsparker.
Levnadssätt
Vit silkesapa är aktiv på dagen. På natten vilar den i trädets håligheter eller i täta ansamlingar av klätterväxter i trädens krona. Den går på fyra fötter över grenar eller hoppar.
De lever i grupper om 6 upp till 11 djur. Grupperna består oftast av ett dominant par samt några andra unga eller vuxna individer. De icke dominanta honorna i gruppen saknar förmåga att föda ungdjur då äggcellerna blockeras i äggstocken. Det antas att feromoner spelar en viktig roll.
Gruppens revir är med 2,2 till 5 hektar jämförelsevis litet. Territoriet markeras med vätska från körtlarna vid bröstet och vid djurets anus. Flocken vandrar varje dag 1000 till 1300 meter.
För att kommunicera använder sig vit silkesapa av olika läten, kroppsställningar och ansiktsuttryck.
Föda
Artens föda utgörs huvudsakligen av trädvätskor som naturgummi och insekter. Liksom andra silkesapor har de speciella tänder för att göra hål i trädens bark. På så sätt behöver de inte lika stora revir som andra primater. Födan består bara till 25 eller 30 procent av insekter. I viss mån äter de även frukter, blommor, frön, svampar, snäckor, mindre ryggradsdjur och fågelägg.
Fortplantning
I fångenskap förekommer nästan uteslutande monogama par. Det antas att fortplantningssättet i naturen är mer varierad, så att den dominanta honan parar sig med flera hannar.
Efter dräktigheten som varar i ungefär fem månader (omkring 150 dagar) föder honan vanligen tvillingar. Ungarna är stora med ungefär en fjärde del av moderns vikt. Hannar och andra medlemmar av gruppen är delaktig i ungarnas uppfostring. Efter tre månader sluter honan att ge di. Ungdjur är efter två år könsmogna.
De lever upp till 10 i naturen och upp till 16 år i fångenskap.
Vit silkesapa och människor
Sedan 1960-talet fångas individer som försöksdjur och för att göra de till sällskapsdjur. I laboratorium hör de idag till de vanligaste primaterna. De avlas idag främst i fångenskap istället för att fånga nya individer i naturen. Dessutom hotas arten i vissa regioner genom förstöringen av levnadsområdet.
IUCN listar arten som livskraftig (LC).
Artepitet i det vetenskapliga namnet kommer från den grekiska mytologin. Det är antingen ett synonym till Dionysos eller karaktären Iakkhos från en annan legend. Carl von Linné hämtade flera artnamn från mytologier utan att de behövde ha någon koppling till djuret.
Referenser
Thomas Geissmann: Vergleichende Primatologie, Springer-Verlag 2003,
D. E. Wilson & D. M. Reeder: Mammal Species of the World. Johns Hopkins University Press, 2005.
Noter
Externa länkar
Kloapor
Däggdjur i neotropiska regionen | swedish | 0.827413 |
extinction_fungus/globalextinctionrate.txt | Close
/
← →
Search Search
# [ Yale Environment 360 ](https://e360.yale.edu/)
## Published at the [ Yale School of the Environment
](http://environment.yale.edu/)
* [ Explore ](/topics)
* [ Search ](/search)
* [ About E360 ](/about)
# Global Extinction Rates: Why Do Estimates Vary So Wildly?
Is it 150 species a day or 24 a day or far less than that? Prominent
scientists cite dramatically different numbers when estimating the rate at
which species are going extinct. Why is that?
By [ Fred Pearce ](/authors/fred-pearce) • August 17, 2015
* [ Facebook ](https://www.facebook.com/sharer/sharer.php?u=https%3A%2F%2Fe360.yale.edu%2Ffeatures%2Fglobal_extinction_rates_why_do_estimates_vary_so_wildly)
* [ Twitter ](https://twitter.com/intent/tweet?url=https%3A%2F%2Fe360.yale.edu%2Ffeatures%2Fglobal_extinction_rates_why_do_estimates_vary_so_wildly&text=Global%20Extinction%20Rates%3A%20Why%20Do%20Estimates%20Vary%20So%20Wildly%3F&via=YaleE360)
* [ Email ](mailto:?subject=Yale%20E360%20-%20Global%20Extinction%20Rates%3A%20Why%20Do%20Estimates%20Vary%20So%20Wildly%3F&body=Global%20Extinction%20Rates%3A%20Why%20Do%20Estimates%20Vary%20So%20Wildly%3F%0D%0ABy Fred%20Pearce%0D%0Ahttps%3A%2F%2Fe360.yale.edu%2Ffeatures%2Fglobal_extinction_rates_why_do_estimates_vary_so_wildly%0D%0A)
Most ecologists believe that we are in the midst of the sixth mass extinction.
Humanity’s impact on nature, they say, is now comparable to the five previous
catastrophic events over the past 600 million years, during which up to 95
percent of the planet’s species disappeared. We may very well be. But recent
studies have cited extinction rates that are extremely fuzzy and vary wildly.
The Millennium Ecosystem Assessment, which involved more than a thousand
experts, estimated an extinction rate that was later calculated at up to 8,700
species a year, or 24 a day. More recently, scientists at the [ U.N.
Convention on Biological Diversity concluded
](https://www.cbd.int/doc/speech/2007/sp-2007-05-22-es-en.pdf) that: “Every
day, up to 150 species are lost.” That could be as much as 10 percent a
decade.

The golden toad, once abundant in parts of Costa Rica, was declared extinct in
2007. WIKIMEDIA COMMONS
But nobody knows whether such estimates are anywhere close to reality. They
are based on computer modeling, and documented losses are tiny by comparison.
Only about 800 extinctions have been documented in the past 400 years,
according to data held by the International Union for the Conservation of
Nature (IUCN). Out of some 1.9 million recorded current or recent species on
the planet, that represents less than a tenth of one percent.
Nor is there much documented evidence of accelerating loss. In its latest
update, released in June, the IUCN reported “no new extinctions,” although
last year it reported the loss of an earwig on the island of St. Helena and a
Malaysian snail. And some species once thought extinct have turned out to be
still around, like the [ Guadalupe fur seal
](http://www.nmfs.noaa.gov/pr/species/mammals/pinnipeds/guadalupefurseal.htm)
, which “died out” a century ago, but now numbers over 20,000.
Moreover, the majority of documented extinctions have been on small islands,
where species with small gene pools have usually succumbed to human hunters.
That may be an ecological tragedy for the islands concerned, but most species
live in continental areas and, ecologists agree, are unlikely to prove so
vulnerable.
But the documented losses may be only the tip of the iceberg. That’s because
the criteria adopted by the IUCN and others for declaring species extinct are
very stringent, requiring targeted research. It’s also because we often simply
don’t know what is happening beyond the world of vertebrate animals that make
up perhaps 1 percent of known species.
> One recent report noted that current extinctions were ‘up to 100 times
> higher than the background rate.’
One way to fill the gap is by extrapolating from the known to the unknown. In
June, Gerardo Ceballos at the National Autonomous University of Mexico — in
collaboration with luminaries such as Paul Ehrlich of Stanford and Anthony
Barnosky of the University of California, Berkeley — got headlines around the
world when he used this approach to [ estimate that current global extinctions
](http://advances.sciencemag.org/content/advances/1/5/e1400253.full.pdf) were
“up to 100 times higher than the background rate.”
Ceballos looked at the recorded loss since 1900 of 477 species of vertebrates.
That represented a loss since the start of the 20th century of around 1
percent of the 45,000 known vertebrate species. He compared this loss rate
with the likely long-term natural “background” extinction rate of vertebrates
in nature, which one of his co-authors, Anthony Barnosky of UC Berkeley
recently put at two per 10,000 species per 100 years. This background rate
would predict around nine extinctions of vertebrates in the past century, when
the actual total was between one and two orders of magnitude higher.
Ceballos went on to assume that this accelerated loss of vertebrate species
would apply across the whole of nature, leading him to conclude that
extinction rates today are “up to a hundred times higher” than background.
A few days earlier, Claire Regnier, of the National Museum of Natural History
in Paris, had put the spotlight on invertebrates, which make up the majority
of known species but which, she said, currently “languish in the shadows.”
Regnier looked at one group of invertebrates with comparatively good records —
land snails. And to get around the problem of under-reporting, she threw away
the IUCN’s rigorous methodology and relied instead on expert assessments of
the likelihood of extinction. Thus, she figured that _Amastra baldwiniana_ , a
land snail endemic to the Hawaiian island of Maui, was no more because its
habitat has declined and it has not been seen for several decades. In this
way, she estimated that probably 10 percent of the 200 or so known land snails
were now extinct — a loss seven times greater than IUCN records indicate.
> ‘Marine populations tend to be better connected [so] the extinction threat
> is likely to be lower.’
Extrapolated to the wider world of invertebrates, and making allowances for
the preponderance of endemic land snail species on small islands, she
concluded that “we have probably already lost 7 percent of described living
species.” That could mean, she said, that perhaps 130,000 of recorded
invertebrates have gone.
Several leading analysts applauded the estimation technique used by Regnier.
But others have been more cautious about reading across taxa. They say it is
dangerous to assume that other invertebrates are suffering extinctions at a
similar rate to land snails. Mark Costello, a marine biologist of the
University of Auckland in New Zealand, warned that land snails may be at
greater risk than insects, which make up the majority of invertebrates.
“Because most insects fly, they have wide dispersal, which mitigates against
extinction,” he told me.
The same should apply to marine species that can swim the oceans, says Alex
Rogers of Oxford University. Only 24 marine extinctions are recorded by the
IUCN, including just 15 animal species and none in the past five decades. Some
think this reflects a [ lack of research
](http://www.sciencemag.org/content/347/6219/1255641) . But [ Rogers says
](http://www.cell.com/current-biology/abstract/S0960-9822\(14\)01624-8) :
“Marine populations tend to be better connected [so] the extinction threat is
likely to be lower.”
Whatever the drawbacks of such extrapolations, it is clear that a huge number
of species are under threat from lost habitats, climate change, and other
human intrusions. And while the low figures for recorded extinctions look like
underestimates of the full tally, that does not make the high estimates right.
> Can we really be losing thousands of species for every loss that is
> documented?
Can we really be losing thousands of species for every loss that is
documented? Some ecologists believe the high estimates are inflated by basic
misapprehensions about what drives species to extinction. So where do these
big estimates come from?
Mostly, they go back to the 1980s, when forest biologists proposed that
extinctions were driven by the “species-area relationship.” This relationship
holds that the number of species in a given habitat is determined by the area
of that habitat. The biologists argued, therefore, that the massive loss and
fragmentation of pristine tropical rainforests — which are thought to be home
to around half of all land species — will inevitably lead to a pro-rata loss
of forest species, with dozens, if not hundreds, of species being silently
lost every day. The presumed relationship also underpins assessments that as
much as a third of all species are at risk of extinction in the coming decades
as a result of habitat loss, including from climate change.
But, as rainforest ecologist Nigel Stork, then at the University of Melbourne,
pointed out in a [ groundbreaking paper in 2009
](http://www.griffith.edu.au/__data/assets/pdf_file/0009/351729/Stork-Biod-
Cons-2009.pdf) , if the formula worked as predicted, up to half the planet’s
species would have disappeared in the past 40 years. And they haven’t. “There
are almost no empirical data to support estimates of current extinctions of
100, or even one, species a day,” he concluded.
He is not alone. In 2011, ecologist Stephen Hubbell of UC Los Angeles
concluded, from a [ study of forest plots around the world
](http://www.nature.com/nature/journal/v473/n7347/full/nature09985.html) run
by the Smithsonian Institution, that as forests were lost, “more species
always remained than were expected from the species-area relationship.” Nature
is proving more adaptable than previously supposed, he said. It seems that [
most species don’t simply die out
](http://www.sciencemag.org/content/339/6118/413) if their usual habitats
disappear. Instead they hunker down in their diminished refuges, or move to
new habitats.
Claude Martin, former director of the environment group WWF International — an
organization that in his time often promoted many of the high scenarios of
future extinctions — now agrees that the “pessimistic projections” are not
playing out. In his new book, [ _On The Edge_
](http://www.clubofrome.org/?p=8465) , he points out that El Salvador has lost
90 percent of its forests but only three of its 508 forest bird species.
Meanwhile, the island of Puerto Rico has lost 99 percent of its forests but
just seven native bird species, or 12 percent.
> Some researchers now question the widely held view that most species remain
> to be described.
Some ecologists believe that this is a temporary stay of execution, and that
thousands of species are living on borrowed time as their habitat disappears.
But with more than half the world’s former tropical forests removed, most of
the species that once populated them live on. If nothing else, that gives time
for ecological restoration to stave off the losses, Stork suggests.
But we are still swimming in a sea of unknowns. For one thing, there is no
agreement on the number of species on the planet. Researchers have described
an estimated 1.9 million species (estimated, because of the risk of double-
counting). But, allowing for those so far unrecorded, researchers have put the
real figure at anywhere from two million to 100 million.
Last year Julian Caley of the Australian Institute of Marine Sciences in
Townsville, Queensland, [ complained that
](http://www.sciencedirect.com/science/article/pii/S0169534714000263) “after
more than six decades, estimates of global species richness have failed to
converge, remain highly uncertain, and in many cases are logically
inconsistent.”
That may be a little pessimistic. Some semblance of order is at least emerging
in the area of recorded species. In March, the [ World Register of Marine
Species ](http://www.marinespecies.org/about.php) , a global research network,
pruned the number of known marine species from 418,000 to 228,000 by [
eliminating double-counting ](http://www.lifewatch.be/en/2015.03.12-WoRMS-
LifeWatch-press-release) . Embarrassingly, they discovered that until recently
one species of sea snail, the rough periwinkle, had been masquerading under no
fewer than 113 different scientific names.
Costello says double-counting elsewhere could reduce the real number of known
species from the current figure of 1.9 million overall to 1.5 million. That
still leaves open the question of how many unknown species are out there
waiting to be described. But here too some researchers are starting to draw
down the numbers.
Back in the 1980s, after analyzing beetle biodiversity in a small patch of
forest in Panama, Terry Erwin of the Smithsonian Institution calculated that
the world might be home to 30 million insect species alone — a far higher
figure than previously estimated. His numbers became the received wisdom. But
new analyses of beetle taxonomy have raised questions about them.
In June, Stork used a collection of some 9,000 beetle species held at London’s
Natural History Museum to [ conduct a reassessment
](http://www.pnas.org/content/early/2015/05/27/1502408112.full.pdf) . He
analyzed patterns in how collections from particular places grow, with larger
specimens found first, and concluded that the likely total number of beetle
species in the world might be 1.5 million. From this, he judged that a likely
figure for the total number of species of arthropods, including insects, was
between 2.6 and 7.8 million.
Some researchers now question the widely held view that most species remain to
be described — and so could potentially become extinct even before we know
about them. Costello thinks that [ perhaps only a third of species
](http://www.ncbi.nlm.nih.gov/pubmed/23349283) are yet to be described, and
that “most will be named before they go extinct.”
Does all this argument about numbers matter? Yes, it does, says Stork.
“Success in planning for conservation … can only be achieved if we know what
species there are, how many need protection and where. Otherwise, we have no
baseline against which to measure our successes.” Or indeed to measure our
failures.
None of this means humans are off the hook, or that extinctions cease to be a
serious concern. Extinction [ rates remain high
](http://www.rivm.nl/bibliotheek/digitaaldepot/20040108nature.pdf) . And, even
if some threats such as hunting may be diminished, others such as climate
change have barely begun. Moreover, if there are fewer species, that only
makes each one more valuable.
But Stork raises another issue. He warns that, by concentrating on global
biodiversity, we may be missing a bigger and more immediate threat — the loss
of local biodiversity. That may have a more immediate and profound effect on
the survival of nature and the services it provides, he says.
Ecosystems are profoundly local, based on individual interactions of
individual organisms. It may be debatable how much it matters to nature how
many species there are on the planet as a whole. But it is clear that local
biodiversity matters a very great deal.
* [ Facebook ](https://www.facebook.com/sharer/sharer.php?u=https%3A%2F%2Fe360.yale.edu%2Ffeatures%2Fglobal_extinction_rates_why_do_estimates_vary_so_wildly)
* [ Twitter ](https://twitter.com/intent/tweet?url=https%3A%2F%2Fe360.yale.edu%2Ffeatures%2Fglobal_extinction_rates_why_do_estimates_vary_so_wildly&text=Global%20Extinction%20Rates%3A%20Why%20Do%20Estimates%20Vary%20So%20Wildly%3F&via=YaleE360)
* [ Email ](mailto:?subject=Yale%20E360%20-%20Global%20Extinction%20Rates%3A%20Why%20Do%20Estimates%20Vary%20So%20Wildly%3F&body=Global%20Extinction%20Rates%3A%20Why%20Do%20Estimates%20Vary%20So%20Wildly%3F%0D%0ABy Fred%20Pearce%0D%0Ahttps%3A%2F%2Fe360.yale.edu%2Ffeatures%2Fglobal_extinction_rates_why_do_estimates_vary_so_wildly%0D%0A)
[ 
](https://e360.yale.edu/authors/fred-pearce)
**Fred Pearce** is a freelance author and journalist based in the U.K. He is a
contributing writer for _Yale Environment 360_ and is the author of numerous
books, including [ _The Land Grabbers_ ](https://www.amazon.com/Land-Grabbers-
Fight-over-Earth/dp/0807003417) _,[ Earth Then and Now: Amazing Images of Our
Changing World ](https://www.amazon.com/Earth-Then-Now-Amazing-
Changing/dp/1554074215) _ , and _[ The Climate Files: The Battle for the Truth
About Global Warming. ](https://www.amazon.com/Climate-Files-Battle-Global-
Warming/dp/0852652291) _ [ More about Fred Pearce →
](https://e360.yale.edu/authors/fred-pearce)
## Topics
* [ Climate ](https://e360.yale.edu/topics/climate)
* [ Biodiversity ](https://e360.yale.edu/topics/biodiversity)
* [ Oceans ](https://e360.yale.edu/topics/oceans)
* [ Solutions ](https://e360.yale.edu/topics/solutions)
* [ Policy ](https://e360.yale.edu/topics/policy)
## Regions
* [ Asia ](/regions/asia)
Join the conversation: Global Extinction Rates: Why Do Estimates Vary So
Wildly? Show comments →
[ Never miss a feature! Sign up for the E360 Newsletter → ](/newsletter)
### More From E360
* ##### [ Oceans ](https://e360.yale.edu/features/european-eels)
#### [ Dire Straits: Can a Fishing Ban Save the Elusive European Eel?
](https://e360.yale.edu/features/european-eels)
* ##### [ Climate ](https://e360.yale.edu/features/mcdr-marine-carbon-dioxide-removal)
#### [ Scientists Are Trying to Coax the Ocean to Absorb More CO2
](https://e360.yale.edu/features/mcdr-marine-carbon-dioxide-removal)
* ##### [ INTERVIEW ](https://e360.yale.edu/features/marina-silva-interview)
#### [ Marina Silva on Brazil’s Fight to Turn the Tide on Deforestation
](https://e360.yale.edu/features/marina-silva-interview)
* ##### [ Solutions ](https://e360.yale.edu/features/solomon-islands-sirebe-carbon-credits)
#### [ Solomon Islands Tribes Sell Carbon Credits, Not Their Trees
](https://e360.yale.edu/features/solomon-islands-sirebe-carbon-credits)
* ##### [ INTERVIEW ](https://e360.yale.edu/features/christine-figgener-interview)
#### [ With Sea Turtles in Peril, a Call for New Strategies to Save Them
](https://e360.yale.edu/features/christine-figgener-interview)
* ##### [ RIVERS ](https://e360.yale.edu/features/jared-kushner-albania-vjose-vjosa-river-hotel)
#### [ Jared Kushner Has Big Plans for Delta of Europe’s Last Wild River
](https://e360.yale.edu/features/jared-kushner-albania-vjose-vjosa-river-
hotel)
* ##### [ Energy ](https://e360.yale.edu/features/us-uranium-mining-nuclear-power)
#### [ A Nuclear Power Revival Is Sparking a Surge in Uranium Mining
](https://e360.yale.edu/features/us-uranium-mining-nuclear-power)
* ##### [ OPINION ](https://e360.yale.edu/features/anthropocene-denied)
#### [ Despite Official Vote, the Evidence of the Anthropocene Is Clear
](https://e360.yale.edu/features/anthropocene-denied)
* ##### [ INTERVIEW ](https://e360.yale.edu/features/sonam-wangchuk-interview)
#### [ At 11,500 Feet, a ‘Climate Fast’ to Save the Melting Himalaya
](https://e360.yale.edu/features/sonam-wangchuk-interview)
* ##### [ Oceans ](https://e360.yale.edu/features/octopus-intelligence-conservation)
#### [ Octopuses Are Highly Intelligent. Should They Be Farmed for Food?
](https://e360.yale.edu/features/octopus-intelligence-conservation)
* ##### [ Climate ](https://e360.yale.edu/features/undercounted-emissions-un-climate-change)
#### [ Nations Are Undercounting Emissions, Putting UN Goals at Risk
](https://e360.yale.edu/features/undercounted-emissions-un-climate-change)
* ##### [ Solutions ](https://e360.yale.edu/features/direct-air-capture)
#### [ As Carbon Air Capture Ramps Up, Major Hurdles Remain
](https://e360.yale.edu/features/direct-air-capture)
# [ E360 ](https://e360.yale.edu/)
* [ About E360 ](/about#about-e360)
* [ Reprints ](/about#reprints)
* [ Contact ](/about#contact)
* [ Support E360 ](/about#support-e360)
* [ Privacy Policy ](/about#privacy-policy)
* [ Submission Guidelines ](/about#submission-guidelines)
* [ Newsletter ](https://e360.yale.edu/newsletter)
## Published at the [ Yale School of the Environment
](http://environment.yale.edu/)
| biology | 1571159 | https://sv.wikipedia.org/wiki/Nattpapegoja | Nattpapegoja | Nattpapegoja (Pezoporus occidentalis) är en liten nattaktiv papegoja inom ordningen papegojfåglar. Tidigare placerades den i det egna släktet Geopsittacus, men idag placeras den oftast i släktet Pezoporus tillsammans med markpapegojan. Den kategoriseras som starkt hotad.
Utseende
Nattpapegojan mäter cirka 23 centimeter. Den har klargrön ovansida som är svart-, och gulvattrad, och gul undersidan. Stjärten är ganska kort och spetsig. Könen är lika till utseendet och det fortfarande är okänt hur den ser ut som ungfågel.
Utbredning och fyndstatistik
Dess ursprungliga utbredningsområde var mycket stort och sträckte sig över inlandet i västra och centrala Australien. Idag förekommer den mycket lokalt i sitt forna utbredningsområde i Australiens torra inland. Det första belagda fyndet gjordes 1845 i närheten av den ofta uttorkade Cooper Creek i allra nordöstligaste South Australia. Typexemplaret samlades in 1854 i det inre av Western Australia och beskrevs av John Gould 1861.
Fram till 1912 samlades det in cirka 20 exemplar, varav sexton ifrån Gawler Ranges i South Australia. 1979 rapporterade den kände ornitologen Shane Parker att han observerat fyra stycken i Cooper Creek. I oktober 1990 fann tre ornitologer ett dött och torkat exemplar i västra Queensland i närheten av orten Boulia. I september 2006 hittades ännu ett dött exemplar, nu i Diamantina National Park.
En natt 2009 lyckades ornitologen John Young spela in ett papegojläte som han trodde kunde vara från nattpapegojan. Med hjälp av detta läte lyckades han en natt i maj 2013, i västra Queensland, tillsammans med ytterligare en ornitolog, att locka fram en nattpapegoja som de även lyckades fotografera, vilket är de första fotografierna som tagits av arten.
Den 4 april 2015 lyckades ornitologen Steve Murphy och hans partner Rachel Barr fånga ett levande exemplar och förse den med radiosändare. Ett naturreservat skapades i centrala västra Queensland för att skydda arten. I oktober 2016 rapporterades att forskare hittat ännu en population i den angränsande nationalparken Diamantina och i november 2016 observerades där den första flygande årsungen på över hundra år. Fler populationer har därefter bekräftats i områdena East Murchison och Great Sandy Desert i Western Australia.
Ekologi
På grund av de fåtaliga observationerna är mycket kring dess ekologi okänt. Som många andra ökenlevande djur är den nattaktiv. Det spekuleras att arten kan vara nomadisk och då följer de sällsynta och oregelbundna regnen som resulterar i riklig frösättning. Den verkar vara knuten till biotoper med specifika grässorter och annan låg växtlighet. Två bofynd från 1800-talet indikerar att den häckar i jordhålor som den själv gräver ut i tuvor av ett specifikt hög grässort. Sådana tunnlar kan kanske även ge svalka under heta dagar. Den lever förmodligen av gräsfrön och vätskebehovet tillgodoses förmodligen genom födan.
Status och hot
Inga observationer gjordes av den mycket skygga papegojan under åren 1912 till 1967, vilket ledde till att man trodde den var utdöd. Under 1979-2015 har det gjort ett mycket litet antal observationer varför populationsstorleken är okänd. Under 2016 gjordes ett flertal observationer, däribland av en flygande årsunge. De främsta hoten mot arten utgörs av habitatförstöring och introducerade arter. IUCN kategoriserade arten som akut hotad (CR) fram till 2019, då den nedgraderades till den lägre hotnivån starkt hotad (EN). Sedan 2022 kategoriseras den återigen som akut hotad. Beståndet uppskattas till mellan 40 och 500 vuxna individer.
Referenser
Externa länkar
Fåglar i australiska regionen
Endemiska fågelarter i Australien
Östpapegojor | swedish | 0.892457 |
extinction_fungus/sciadv1400253.txt |
NEWS
CAREERS
COMMENTARY
JOURNALS
ScienceScience
LOG IN
logo
Current Issue
First release papers
Archive
About
HOME
SCIENCE ADVANCES
VOL. 1, NO. 5
ACCELERATED MODERN HUMAN–INDUCED SPECIES LOSSES: ENTERING THE SIXTH MASS EXTINCTION
OPEN ACCESS
RESEARCH ARTICLE
ENVIRONMENTAL SCIENCES
Share on
Accelerated modern human–induced species losses: Entering the sixth mass extinction
GERARDO CEBALLOS , PAUL R. EHRLICH, ANTHONY D. BARNOSKY, ANDRÉS GARCÍA, ROBERT M. PRINGLE, AND TODD M. PALMERAuthors Info & Affiliations
SCIENCE ADVANCES
19 Jun 2015
Vol 1, Issue 5
DOI: 10.1126/sciadv.1400253
127,999
33
Metrics
Total Downloads
127,999
Last 6 Months
29,962
Last 12 Months
49,736
Total Citations
33
Last 6 Months
3
Last 12 Months
5
Abstract
INTRODUCTION
RESULTS
DISCUSSION
MATERIALS AND METHODS
Acknowledgments
Supplementary Material
REFERENCES AND NOTES
eLetters (4)
Information & Authors
Metrics & Citations
View Options
References
Media
Tables
Share
Abstract
The oft-repeated claim that Earth’s biota is entering a sixth “mass extinction” depends on clearly demonstrating that current extinction rates are far above the “background” rates prevailing between the five previous mass extinctions. Earlier estimates of extinction rates have been criticized for using assumptions that might overestimate the severity of the extinction crisis. We assess, using extremely conservative assumptions, whether human activities are causing a mass extinction. First, we use a recent estimate of a background rate of 2 mammal extinctions per 10,000 species per 100 years (that is, 2 E/MSY), which is twice as high as widely used previous estimates. We then compare this rate with the current rate of mammal and vertebrate extinctions. The latter is conservatively low because listing a species as extinct requires meeting stringent criteria. Even under our assumptions, which would tend to minimize evidence of an incipient mass extinction, the average rate of vertebrate species loss over the last century is up to 100 times higher than the background rate. Under the 2 E/MSY background rate, the number of species that have gone extinct in the last century would have taken, depending on the vertebrate taxon, between 800 and 10,000 years to disappear. These estimates reveal an exceptionally rapid loss of biodiversity over the last few centuries, indicating that a sixth mass extinction is already under way. Averting a dramatic decay of biodiversity and the subsequent loss of ecosystem services is still possible through intensified conservation efforts, but that window of opportunity is rapidly closing.
SIGN UP FOR THE SCIENCE ADVANCES eTOC
Get the latest table of contents from Science Advances delivered right to you!
INTRODUCTION
The loss of biodiversity is one of the most critical current environmental problems, threatening valuable ecosystem services and human well-being (1–7). A growing body of evidence indicates that current species extinction rates are higher than the pre-human background rate (8–15), with hundreds of anthropogenic vertebrate extinctions documented in prehistoric and historic times (16–23). For example, in the islands of tropical Oceania, up to 1800 bird species (most described in the last few decades from subfossil remains) are estimated to have gone extinct in the ~2000 years since human colonization (24). Written records of extinctions of large mammals, birds, and reptiles date back to the 1600s and include species such as the dodo (Raphus cucullatus, extinguished in the 17th century), Steller’s sea cow (Hydrodamalis gigas, extinguished in the 18th century), and the Rodrigues giant tortoise (Cylindraspis peltastes, extinguished in the 19th century). More species extinction records date from the 19th century and include numerous species of mammals and birds. Records of extinction for reptiles, amphibians, freshwater fishes, and other organisms have mainly been documented since the beginning of the 20th century (14, 17). Moreover, even in species that are not currently threatened, the extirpation of populations is frequent and widespread, with losses that far outstrip species-level extinctions (18, 25). Population-level extinction directly threatens ecosystem services and is the prelude to species-level extinction (18).
Here, we analyze the modern rates of vertebrate species extinction and compare them with a recently computed background rate for mammals (7). We specifically addressed the following questions: (i) Are modern rates of mammal and vertebrate extinctions higher than the highest empirically derived background rates? (ii) How have modern extinction rates in mammals and vertebrates changed through time? (iii) How many years would it have taken for species that went extinct in modern times to have been lost if the background rate had prevailed? These are important issues because the uncertainties about estimates of species loss have led skeptics to question the magnitude of anthropogenic extinctions (26) and because understanding the magnitude of the extinction crisis is relevant for conservation, maintenance of ecosystem services, and public policy.
Until recently, most studies of modern extinction rates have been based on indirect estimates derived, for example, on the rates of deforestation and on species-area relationships (11, 14). Problems related to estimating extinction since 1500 AD (that is, modern extinctions) have been widely discussed, and the literature reflects broad agreement among environmental scientists that biases lead to underestimating the number of species that have gone extinct in the past few centuries—the period during which Homo sapiens truly became a major force on the biosphere (1–4, 6–8, 14, 15). However, direct evaluation is complicated by uncertainties in estimating the incidence of extinction in historical time and by methodological difficulties in comparing contemporary extinctions with past ones.
Less discussed are assumptions underlying the estimation of background extinction rates. The lower these estimates, the more dramatic current extinction rates will appear by comparison. In nearly all comparisons of modern versus background extinction rates, the background rate has been assumed to be somewhere between 0.1 and 1 species extinction per 10,000 species per 100 years (equal to 0.1 to 1 species extinction per million species per year, a widely used metric known as E/MSY). Those estimates reflect the state of knowledge available from the fossil record in the 1990s (7, 9–13). In a recent analysis, which charted the stratigraphic ranges of thousands of mammal species, extinction rates were measured over intervals ranging from single years to millions of years, and the mean extinction rate and variance were computed for each span of time (7). In this way, the background extinction rate estimated for mammals was estimated at 1.8 E/MSY, here rounded upward conservatively to 2 E/MSY (that is, 2 extinctions per 100 years per 10,000 species). This is double the highest previous rough estimate.
Those previously estimated background rates were primarily derived from marine invertebrate fossils, which are likely to have greater species longevity than vertebrates (10, 15). Data deficiencies make it impossible to conduct empirical analyses (as was done for mammals) for non-mammal terrestrial vertebrates; therefore, we assume the background rates of other vertebrates to be similar to those of mammals. This supposition leads to a more conservative assessment of differences between current and past extinction rates for the vertebrates as a whole, compared with using the very low background extinction rate derived from marine invertebrates.
The analysis we present here avoids using assumptions such as loss of species predicted from species-area relationships, which can suggest very high extinction rates, and which have raised the possibility that scientists are “alarmists” seeking to exaggerate the impact of humans on the biosphere (26). Here, we ascertain whether even the lowest estimates of the difference between background and contemporary extinction rates still justify the conclusion that people are precipitating a global spasm of biodiversity loss.
RESULTS
Modern and background rates of vertebrate extinctions
Modern rates of vertebrate extinction were much higher than a background extinction rate of 2 E/MSY. Among the vertebrate taxa evaluated by the International Union of Conservation of Nature (IUCN), 338 extinctions have been documented since 1500 [“extinct” (EX), Table 1]. An additional 279 species have become either “extinct in the wild” (EW) or listed as “possibly extinct” (PE), totaling 617 vertebrate species summed over the three categories. Most extinctions have occurred in the last 114 years (that is, since 1900; Table 1). Our estimated “highly conservative” (that is, using data for EX species only) and “conservative” (that is, by including EX, EW, and PE) modern extinction rates for vertebrates varied from 8 to 100 times higher than the background rate (Table 2). This means, for example, that under the 2 E/MSY background rate, 9 vertebrate extinctions would have been expected since 1900; however, under the conservative rate, 468 more vertebrates have gone extinct than would have if the background rate had persisted across all vertebrates under that period. Specifically, these 468 species include 69 mammal species, 80 bird species, 24 reptiles, 146 amphibians, and 158 fish.
Vertebrate taxon No. of species, IUCN 2014.3
Highly conservative
rates (EX) Conservative rates
(EX + EW + PE) No. of species
evaluated by IUCN
Since 1500 Since 1900 Since 1500 Since 1900
Vertebrates 338 198 617 477 59% (39,223)
Mammals 77 35 111 69 100% (5,513)
Birds 140 57 163 80 100% (10,425)
Reptiles 21 8 37 24 44% (4,414)
Amphibians 34 32 148 146 88% (6,414)
Fishes 66 66 158 158 38% (12,457)
Table 1 Numbers of species used in the Table 2 calculations of “highly conservative” and “conservative” modern extinction rates based on the IUCN Red List (17).
For the highly conservative rates, only species verified as “extinct” (EX) were included; for the conservative extinction rates, species in the categories “extinct in the wild” (EW) and “possibly extinct” (PE) were also included.
Animal group Elevation of modern rates with
respect to expected rates
Highly conservative Conservative
Since 1500 Since 1900 Since 1500 Since 1900
Vertebrates 8 22 15 53
Mammals 14 28 20 55
Birds 13 24 15 34
Reptiles 5 8 8 24
Amphibians 5 22 22 100
Fishes 5 23 12 56
Table 2 Elevation of “highly conservative” and “conservative” modern vertebrate extinction rates above background rate of 2 E/MSY (see table S2 for calculations).
For each assessment category, two periods are shown: extinction rates computed from 1500 to the present, and from 1900 to the present.
Variation in modern extinction rates through time
Modern extinction rates have increased sharply over the past 200 years (corresponding to the rise of industrial society) and are considerably higher than background rates (Fig. 1). Rates of modern extinctions vary among vertebrate groups (Fig. 1). For example, amphibians, comprising of ~7300 species, show an accelerating rate of extinction: only 34 extinctions have been documented with a high level of certainty since 1500, yet >100 species have likely disappeared since 1980 (17, 23). This may not only reflect real trends but also a shortage of data for groups for which most species are not yet evaluated, such as reptiles and fish (21, 22).
Fig. 1 Cumulative vertebrate species recorded as extinct or extinct in the wild by the IUCN (2012).
Graphs show the percentage of the number of species evaluated among mammals (5513; 100% of those described), birds (10,425; 100%), reptiles (4414; 44%), amphibians (6414; 88%), fishes (12,457; 38%), and all vertebrates combined (39,223; 59%). Dashed black curve represents the number of extinctions expected under a constant standard background rate of 2 E/MSY. (A) Highly conservative estimate. (B) Conservative estimate.
Modern extinctions if background rate had prevailed
Our results indicate that modern vertebrate extinctions that occurred since 1500 and 1900 AD would have taken several millennia to occur if the background rate had prevailed. The total number of vertebrate species that went extinct in the last century would have taken about 800 to 10,000 years to disappear under the background rate of 2 E/MSY (Fig. 2). The particularly high losses in the last several decades accentuate the increasing severity of the modern extinction crisis.
Fig. 2 Number of years that would have been required for the observed vertebrate species extinctions in the last 114 years to occur under a background rate of 2 E/MSY.
Red markers, highly conservative scenario; blue markers, conservative scenario. Note that for all vertebrates, the observed extinctions would have taken between 800 to 10,000 years to disappear, assuming 2 E/MSY. Different classes of vertebrates all show qualitatively similar trends.
DISCUSSION
Arguably the most serious aspect of the environmental crisis is the loss of biodiversity—the other living things with which we share Earth. This affects human well-being by interfering with crucial ecosystem services such as crop pollination and water purification and by destroying humanity’s beautiful, fascinating, and culturally important living companions (4, 5, 15, 27–30).
Our analysis shows that current extinction rates vastly exceed natural average background rates, even when (i) the background rate is considered to be double previous estimates and when (ii) data on modern vertebrate extinctions are treated in the most conservative plausible way. We emphasize that our calculations very likely underestimate the severity of the extinction crisis because our aim was to place a realistic “lower bound” on humanity’s impact on biodiversity. Therefore, although biologists cannot say precisely how many species there are, or exactly how many have gone extinct in any time interval, we can confidently conclude that modern extinction rates are exceptionally high, that they are increasing, and that they suggest a mass extinction under way—the sixth of its kind in Earth’s 4.5 billion years of history.
A final important point is that we focus exclusively on species, ignoring the extirpation of populations—the units relevant to ecological functioning and the delivery of ecosystem services (4, 5, 29). Population extinction cannot be reliably assessed from the fossil record, precluding any analysis along the lines of that presented here. Also, although it is clear that there are high rates of population extinction (18), existing data are much less reliable and far harder to obtain than those for species, which will remain true for the foreseeable future. Likewise, we have not considered animals other than vertebrates because of data deficiencies.
The evidence is incontrovertible that recent extinction rates are unprecedented in human history and highly unusual in Earth’s history. Our analysis emphasizes that our global society has started to destroy species of other organisms at an accelerating rate, initiating a mass extinction episode unparalleled for 65 million years. If the currently elevated extinction pace is allowed to continue, humans will soon (in as little as three human lifetimes) be deprived of many biodiversity benefits. On human time scales, this loss would be effectively permanent because in the aftermath of past mass extinctions, the living world took hundreds of thousands to millions of years to rediversify. Avoiding a true sixth mass extinction will require rapid, greatly intensified efforts to conserve already threatened species and to alleviate pressures on their populations—notably habitat loss, overexploitation for economic gain, and climate change (31–33). All of these are related to human population size and growth, which increases consumption (especially among the rich), and economic inequity (6). However, the window of opportunity is rapidly closing.
MATERIALS AND METHODS
To estimate modern extinction rates, we compiled data on the total number of described species and the number of extinct and possibly extinct vertebrate species from the 2014 IUCN Red List (17). In the IUCN’s list, extinct species can be viewed as the minimum number of actual extinctions during recent human history (that is, since 1500) because it lists species known to be extinct (EX), extinct in the wild (EW), and possibly extinct (PE, a subcategory within “critically endangered” reserved for species thought to be extinct, but not confirmed) (17) (table S1). We used the IUCN data to calculate modern extinction rates in two ways: (i) we estimate a “highly conservative modern extinction rate” by using the data exclusively on species listed as EX, and (ii) we calculate a “conservative extinction rate” by including also both EW and PE species (table S2). Including these latter two categories recognizes that there is only a slim chance that most of the species in those categories can reestablish viable populations in their native habitats. In terms of biological impact and the provision of ecosystem services, we consider EW and PE species to be functionally equivalent to EX species: even if some individuals still exist, their abundances are not sufficient to have a substantial influence on ecological function and processes.
The IUCN’s list is considered the authoritative, albeit likely conservative, assessment of the conservation status of plant and animal species. About 1.8 million species have been described since 1758 (when the current nomenclature system was developed), of which 1.3 million are animals (3, 17). Of these animal species, about 39,223 (of the currently counted 66,178) vertebrate species have been formally assessed and reported in the 2014 IUCN Red List (17). In the IUCN sample, mammals, birds, and amphibians have had between 88 and 100% of their known species evaluated, whereas only 44% of reptiles and 38% of fish species have been assessed (Table 1). We focus our comparisons on vertebrates because they are the group for which the most reliable data exist, both fossil and modern.
To produce conservative comparisons with modern extinctions, we assumed a background extinction rate of 2 E/MSY as the highest likely baseline average background extinction rate (7); that is, we should expect 2 extinctions per 10,000 vertebrate species per 100 years. That background extinction rate was empirically determined using the exceptionally good fossil records of mammals, combining extinction counts from paleontological databases and published literature on the fossil, subfossil, and historical records (7). Using the resulting high background extinction rate provides a stringent test for assessing whether current modern extinction rates indicate that a mass extinction event is under way. Previous estimates of background extinction rates for other taxa are invariably lower than the mammal-derived estimate of 2 E/MSY used here.
Acknowledgments
We would like to thank B. Young for helping us with the data on “possibly extinct species” published by IUCN. J. Soberon, C. Mendenhall, and J. Pacheco gave valuable suggestions on the manuscript. Funding: This work has been supported by the Programa de apoyo a proyectos de investigación e innovación tecnológica from UNAM. Competing interests: The authors declare that they have no competing interests.
Supplementary Material
Summary
Table S1. Definitions of IUCN categories (17) used to assess modern extinction rates.
Table S2. Estimation of modern extinction rates since 1500 and 1900.
Resources
File (1400253_sm.pdf)
DOWNLOAD
1.20 MB
REFERENCES AND NOTES
1
G. Ceballos, A. Garcia, P. R. Ehrlich, The sixth extinction crisis: Loss of animal populations and species. J. Cosmology 8, 1821–1831 (2010).
GOOGLE SCHOLAR
2
R. Dirzo, P. H. Raven, Global state of biodiversity and loss. Annu. Rev. Environ. Resour. 28, 137–167 (2003).
CROSSREF
ISI
GOOGLE SCHOLAR
3
G. Mace, K. Norris, A. Fitter, Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 27, 19–26 (2012).
GO TO REFERENCE
GOOGLE SCHOLAR
4
G. Mace, C. Revenga, E. Ken, Biodiversity, in Ecosystems and Human Well-Being: Current State and Trends, G. Ceballos, G. Orians, S. L. Pacala, Eds. (Island Press, Washington, DC, 2005), chap. 4, pp. 77–121.
GOOGLE SCHOLAR
(4)
eLetters
eLetters is a forum for ongoing peer review. eLetters are not edited, proofread, or indexed, but they are screened. eLetters should provide substantive and scholarly commentary on the article. Embedded figures cannot be submitted, and we discourage the use of figures within eLetters in general. If a figure is essential, please include a link to the figure within the text of the eLetter. Please read our Terms of Service before submitting an eLetter.
JUN. 29, 2015
RE: Accelerated modern human-induced species losses: Enteringthe sixth mass extinction
JOHN C. BRIGGS Professor Emeritus Department of Fisheries and Wildlife, Oregon State University, Corvallis, OR 97333, USA
Is a Sixth Global Mass Extinction Underway?
John C. Briggs*
In their recent article, published in Sciences Advances, Ceballos et al. (19 June 2015) endeavored to demonstrate that species extinctions by humans have caused the world to enter into its sixth mass extinction. As evidence, they compared extinction rates, among terrestrial vertebrates for the past few centuries, to the historical rate from the geological record. However, the historical rate was calculated only from data on mammal extinctions. The authors assumed that the historical mammal rate could be applied to all the other vertebrates (birds, reptiles, amphibians, and fishes). Using the mammal data, the authors determined that, without human influence, only nine vertebrate extinctions would have been expected since the year 1900.
According to the Red List (IUCN, 2014), 198 extinctions were recorded for all terrestrial vertebrates since the year 1900. Compared to the expected nine extinctions during that time (Caballos et al., 2015), an increase of 189 appears to indicate a disastrous human influence. Extending back to the year 1500, a total of 338 were documented by the Red List. In addition to these numbers, the authors recognized 279 more species under "extinct in the wild" and "possibly extinct" categories. These figures produced a grand total 617 vertebrate species lost or presumed lost since 1500. These impressive numbers persuaded the authors to declare that the evidence is "incontrovertible", that the recent extinction rates are unprecedented, and that the rate is highly unusual in Earth history. Therefore, they concluded that a mass extinction episode had been initiated.
The unfortunate aspect of this work is that no reference was made to other estimates of global extinctions using other faunal groups. Extinctions recorded in the marine environment were not considered, and no mention was made about concurrent gains in species diversity. About 90% of the terrestrial world's species are insects. A recent estimate by an entomologist (Dunn, 2005) indicates a species diversity of about 3.4 million. While many insect species are still undescribed, certain groups, (butterflies, tiger beetles, dragonflies, and damselflies) are well known and their extinctions during the past 500 years have been documented. Within those groups, which total 25,260 species, only two species have become extinct (Briggs, 2014). In the marine environment, which covers about 71% of the Earth's surface, the diversity of multicellular species is about 2.21 million (Mora et al., 2011); of this number, only 20 have been recorded to be extinct (Dulvy et al., 2009). Furthermore, a recent, uncited paleontological work on extinctions (MacLeod, 2013) provides valuable information on Pleistocene and modern extinction rates. Finally, there is good evidence that points to continual gains in diversity due to species invasions and rapid ecological speciation (Briggs, 2014).
Aside from the neglect of the published information cited above, the attempt to forecast a global extinction on the basis of only terrestrial vertebrate animals (Ceballos et al., 2015), is unconvincing when comparable information on other faunal groups is not utilized. Also, in dealing with the numbers of vertebrate extinctions, the authors failed to mention that most of them took place on oceanic islands. In regard to the mammals and birds, more than 95% of all extinctions during the past 500 years were island species (Loehle and Eschenbach, 2012). If we use the mammals and birds as surrogates for all the vertebrates, extinctions on the world's continents must have been very low. For example, only three of the 61 mammal extinctions took place on continents. The loss of isolated island species had little or no effect on continental ecology.
This is the latest in a series of books and papers, published over the past 40 years, devoted to the frightening vision of a human-caused mass extinction. Its use of colorful graphs, showing vertebrate species dying off by the hundreds, presents a doomsday scenario that many are likely to believe, unless they consider the global extinction evidence.
References
Briggs, J.C. (2014) Global biodiversity gain is concurrent with declining population sizes. Biodiversity Journal, 5, 447-452.
Ceballos, G., Ehrlich, P.R., Barnosky, A.D., García, A., Pringle, R.M. & Palmer, T.M. (2015) Accelerated modern human-induced species losses: entering the sixth mass extinction. Science Advances. 1:e 1400253.
Dulvy, N.K., Pinnegar, J.K., & Reynolds, J.D. (2009) Holocene extinctions in the sea. In: Turvey, S.T. (Ed.) Holocene extinctions. Oxford University Press, Oxford, pp. 129-150.
Dunn, R.R. (2005) Modern insect extinctions: the neglected majority. Conservation Biology, 19, 1030-1036.
IUCN (2014) www.iucnredlist.org (March 20, 2015).
Loehle, C. & Eschenbach, W. (2012) Historic bird and terrestrial mammal extinction: rates and causes. Diversity and Distribution, 18, 84-91.
MacLeod, N. (2013) The great extinctions. Firefly Books, Buffalo, NY.
Mora, C., Tittensor, D.P., Adl, S., Simpson, A.G.B. & Worm, B. (2011) How many species are there on Earth and in the ocean? PLoS Biol. 9, e21001127.
……………………………………………………………………………………………………………………………………………….
*Department of Fisheries and Wildlife, Oregon State University, Corvallis, OR 97333, USA; e-mail: [email protected] Current address: 2320 Guerneville Rd., Santa Rosa, CA 95403,
JUN. 23, 2015
Are we nearing "Dooms Day"?
AMIT KUMAR BANERJEE Researcher CSIR-Indian Institute of Chemical Technology, Hyderbad, India
NEELIMA ARORA Postdoctoral Fellow Alumnous, CSIR-Indian Institute of Chemical Technology, Hyderbad, India
Is human species who considered himself at the pedestal of superiority is inviting its own end? This is suggested in the present study by Ceballos et al. where they attempted to identify the next mass extinction. The pace at which species are getting extinct is alarming. Though there were reports earlier about the high rate of extinction of species in post industrialization era, yet lack of conclusive parameters and estimates always diluted the effect it should have on the policy makers and public. Comparison of "highly conservative modern extinction rate" and "background extinction rate" in this study provides a clear picture of rapid extinction of species. Man's consumerist approach has been responsible for bringing on the possibility of sixth mass extinction that could very well have been avoided by adopting small measures in light of knowledge of ecology and modern science.
Various anthropogenic factors like deforestation, aggressive exploitation of natural resources, mining and mindless dumping toxic wastes in natural environment may have plausible role for massive extinction. While we are trying to explore life on other planets, forgetting to concentrate to take measures to save our own planet. Man has lost the touch with nature and has become oblivious of his role of just a co-habitant on Earth not its master. Small steps like supporting the local animals and birds and separating solid wastes can go a long way in this direction. If we learn to appreciate the role of biodiversity for our well being, progress and future survival and found ways to nurture and conserve it in a sustainable manner, we can at least delay the mass extinction if not avoid it.
Consortium development and effective data collection and further analysis will lead to more awareness about the issue and conservation at global scale.
JUN. 22, 2015
RE: Accelerated modern human–induced species losses: Entering the sixth mass extinction
THOMAS CROFT Youth sports coordinator. PSA
The article brings some interesting statistics showing the increase rate of extinctions among mammal and vertebrate. The over population of humans is obviously a great problem and has been for years. The population of humans in the early 1960s was around 3 billion and today over 7 billion and projected to be around 10 billion by 2020. No matter what mankind does that kind of overpopulation will put a stress on earth's ecosystems and animals and bio-systems. I remember scientists in the 1970s warning of overpopulation.. it is the fundamental basis for all the environmental problems. This article, if the stats are correct, bring this more to light.
However this article's very important point looses credibility with statements like: "All of these are related to human population size and growth, which increases consumption (especially among the rich), and economic inequity". Over population increases consumption - period. Consumption of resources is world wide and is effected by population increases world wide, not just the rich ( and I assume you meant rich countries). And population increases are happening more rapidly in the less "rich" countries. "Overexploitation for economic gain" is minor compared to just the sheer volume of people living on the earth that consume air and food and resources and create waste. The destruction of the rain forest in South America since the 1940s to make cheap farming land for the poor has probably done as much damage to species extinction as other factors. That rain forest is huge role in photosynthesis as well as maintaining millions of animal lives.
While this article is important in its statistics on animal extinctions, it barely touches on how the loss directly is caused by man or effects man or any solutions (which I understand that is not what this article was for). The article ended sounding of some sort of opinionated agenda which lessen its scientific credibility. Still very important.
JUN. 19, 2015
RE: Figure 2 legend reversed?
KENNETH DANIEL HOOPER Electrical Engineer
Possibly it is my lack of understanding, but should the MORE conservative figure result in a smaller equivalent time to extinction?
Looking at Table S2 "1900 to 2014 Highly Conservative" F/E for Amphibians (for example) shows '22'
Looking at Table S2 "1900 to 2014 Conservative" F/E for Amphibians shows '100'.
This is reversed from Figure 2, which shows "Very Conservative" for Amphibians at 10,000 years and "Conservative" for Amphibians at 2,200 years.
Recommended articles from TrendMD
Collapse of the world’s largest herbivores
William J. Ripple et al., Sci Adv, 2015
Parasite biodiversity faces extinction and redistribution in a changing climate
Colin J. Carlson et al., Sci Adv, 2017
Estimating the global conservation status of more than 15,000 Amazonian tree species
Hans ter Steege et al., Sci Adv, 2015
The commonness of rarity: Global and future distribution of rarity across land plants
Brian J. Enquist et al., Sci Adv, 2019
Twentieth century turnover of Mexican endemic avifaunas: Landscape change versus climate drivers
A. Townsend Peterson et al., Sci Adv, 2015
Inhibiting the inhibitors: Development of the IAP inhibitor xevinapant for the treatment of locally advanced squamous cell carcinoma of the head and neck
Robert L. Ferris et al., Cancer Treatment Reviews, 2023
Powered by
CURRENT ISSUE
Science Advances cover image
Dual and opposing roles for the kinesin-2 motor, KIF17, in Hedgehog-dependent cerebellar development
BY BRIDGET WAAS BRANDON S. CARPENTER ET AL.
An osteoarthritis pathophysiological continuum revealed by molecular biomarkers
BY VIRGINIA BYERS KRAUS SHUMING SUN ET AL.
Monolithic integration of perovskite heterojunction on TFT backplanes through vapor deposition for sensitive and stable x-ray imaging
BY LIQI LI LITING TAO ET AL.
LATEST NEWS
NEWS FEATURE29 APR 2024
Which wild animals carry the COVID-19 virus? An ambitious U.S. project aims to find out
NEWS26 APR 2024
All dog noses—whether a pug’s or a bloodhound’s—are created equal
SCIENCEINSIDER26 APR 2024
What’s in your food? A new research effort intends to find out
NEWS26 APR 2024
Injectable antibody drug protects children from malaria in Mali trial
NEWS26 APR 2024
What is a presumed sign of life doing on a dead comet?
NEWS26 APR 2024
AI transcription tools ‘hallucinate,’ too
RECOMMENDED
REPORTSMAY 2002
Mammal Population Losses and the Extinction Crisis
LETTERJUNE 2018
The misunderstood sixth mass extinction
REPORTMAY 2015
Paleontological baselines for evaluating extinction risk in the modern oceans
REPORTSMARCH 2004
Comparative Losses of British Butterflies, Birds, and Plants and the Global Extinction Crisis
View full text|Download PDF
Skip slideshow
NEWS
All News
ScienceInsider
News Features
Subscribe to News from Science
News from Science FAQ
About News from Science
CAREERS
Careers Articles
Find Jobs
Employer Hubs
COMMENTARY
Opinion
Analysis
Blogs
JOURNALS
Science
Science Advances
Science Immunology
Science Robotics
Science Signaling
Science Translational Medicine
Science Partner Journals
AUTHORS & REVIEWERS
Information for Authors
Information for Reviewers
LIBRARIANS
Manage Your Institutional Subscription
Library Admin Portal
Request a Quote
Librarian FAQs
ADVERTISERS
Advertising Kits
Custom Publishing Info
Post a Job
RELATED SITES
AAAS.org
AAAS Communities
EurekAlert!
Science in the Classroom
ABOUT US
Leadership
Work at AAAS
Prizes and Awards
HELP
FAQs
Access and Subscriptions
Order a Single Issue
Reprints and Permissions
TOC Alerts and RSS Feeds
Contact Us
FOLLOW US
AAAS logo
© 2024 American Association for the Advancement of Science. All rights reserved. AAAS is a partner of HINARI, AGORA, OARE, CHORUS, CLOCKSS, CrossRef and COUNTER. Science Advances eISSN 2375-2548.
Terms of Service Privacy Policy Accessibility
PDF
Help
Reference #1
| biology | 998 | https://da.wikipedia.org/wiki/Art | Art | Arten (species, forkortet sp., flertal: spp.) er den grundlæggende systematiske enhed inden for biologien. Arten defineres ofte som en naturlig gruppe af populationer, hvor udveksling af gener finder sted (eller kan finde sted) og som i forhold til forplantning er isoleret fra andre grupper. Det vil sige at kun individer inden for samme art kan parre sig og få forplantningsdygtigt afkom. Dette kaldes det biologiske artsbegreb. For organismer, der formerer sig ukønnet eller ved selvbestøvning, må arter afgrænses ud fra ligheder og forskelle mellem forskellige individer. Nogle dyrearter kan i fangenskab hybridisere og få fertilt afkom, men da dette ikke vil ske i naturen, selv om de mødes her, betragtes de som forskellige arter.
Eksempel
To heste kan parre sig og få et føl, der igen kan få føl med andre heste – hestene tilhører derfor samme art. En hest og et æsel kan også parre sig og deres unger kaldes enten muldyr eller mulæsel, afhængig af hvem der er moren, men muldyret eller mulæselet kan (normalt) ikke få unger, da de oftest er sterile. Af den grund regnes hest og æsel som to forskellige arter. Det samme princip gælder også for planterne. Denne naturskabte afgrænsning mellem to arter kaldes en artsbarriere. Den kan af og til gennembrydes, når ellers sterile krydsninger spontant eller kunstigt får gennemført en kromosomfordobling. Se f.eks. Vadegræs (Spartina pectinata).
Arter over for hybrider
Man kan dog godt komme ud for, at arter kan krydses og får blandet afkom, men hybriden vil kun kunne bestå på steder, hvor ingen af forældrearterne kan klare sig. Dette er et særligt udpræget problem med Rododendron (Rhododendron) og Tjørn (Crataegus), fordi disse slægter breder sig voldsomt efter skovbrand eller stormfald. Da hybriderne bliver frugtbare i en yngre alder end arterne, kan de dominere i en periode, men når skoven lukker sig, så fortrænges hybriderne og kun de specialiserede arter kan overleve i skovens dybe skygge eller ude i lyset i sumpe, på ur og i kalksten, m.m.
Flere artsbegreber
Fordi det biologiske artsbegreb kan være besværligt at anvende i praksis, er der efterhånden skabt en række andre artsbegreber:
Morfologisk artsbegreb Arterne adskiller sig fra hinanden ved deres bygning. Dette begreb er blevet meget anvendt gennem tiden.
Økologisk artsbegreb Definerer en art som en gruppe af organismer, der udfylder samme niche. Krydsninger mellem to nærtstående arter vil ikke være optimalt tilpasset til forældrearternes nicher og vil ikke klare sig i konkurrencen.
Evolutionære artsbegreb Også kaldet det kladistiske eller fylogenetiske artsbegreb. Naturen er dynamisk, ikke statisk - alle arter ændrer sig med tiden og bliver, hvis de ikke uddør som følge af konkurrence, naturkatastrofer m.v., til én eller flere nye arter. Det evolutionære artsbegreb minder om det biologiske, men inddrager tidsdimensionen, det vil sige at en art udvikler sig over tid og at nye arter opstår ved artsdannelse. Individer der fylogenetisk har samme stamfader tilhører samme art.
Pluralistisk artsbegreb En art er et samfund af populationer, der formerer sig og lever inden for en bestemt niche i naturen.
Se også
Systematik
Evolutionsteori
Kilder
Lars Skipper: Hvad er en art? Citat: "...Arten er den eneste [klassifikations-kategori] der eksisterer i virkeligheden, alle andre (slægter, familier, ordener m.v.) er indført for overskuelighedens skyld..."
Eksterne henvisninger
2003-12-31, ScienceDaily: Working On The 'Porsche Of Its Time': New Model For Species Determination Offered Citat: "...two species of dinosaur that are members of the same genera varied from each other by just 2.2 percent. Translation of the percentage into an actual number results in an average of just three skeletal differences out of the total 338 bones in the body. Amazingly, 58 percent of these differences occurred in the skull alone. "This is a lot less variation than I'd expected," said Novak..."
2003-08-08, ScienceDaily: Cross-species Mating May Be Evolutionarily Important And Lead To Rapid Change, Say Indiana University Researchers Citat: "...the sudden mixing of closely related species may occasionally provide the energy to impel rapid evolutionary change..."
2004-01-09 ScienceDaily: Mayo Researchers Observe Genetic Fusion Of Human, Animal Cells; May Help Explain Origin Of AIDS Citat: "...The researchers have discovered conditions in which pig cells and human cells can fuse together in the body to yield hybrid cells that contain genetic material from both species..."What we found was completely unexpected," says Jeffrey Platt, M.D..."
2000-09-18, ScienceDaily: Scientists Unravel Ancient Evolutionary History Of Photosynthesis Citat: "...gene-swapping was common among ancient bacteria early in evolution..."
2004-06-07, Sciencedaily: Parting Genomes: University Of Arizona Biologists Discover Seeds Of Speciation Citat: "...There's a huge amount of biodiversity out there, and we don't know where it comes from. Evolutionary biologists are excited to figure out what causes what we see out there--the relative forces of selection and drift--whether things are adapting to their environment or variation is random..."
2005-07-05, Sciencedaily: Trees, Vines And Nets -- Microbial Evolution Changes Its Face Citat: "... EBI researchers have changed our view of 4 billion years of microbial evolution...In all, more than 600,000 vertical transfers are observed, coupled with 90,000 gene loss events and approximately 40,000 horizontal gene transfers...A few species, including beneficial nitrogen-fixing soil bacteria, appear to be 'champions'of horizontal gene transfer; "it's entirely possible that apparently harmless organisms are quietly spreading antibiotic resistance under our feet," concludes Christos Ouzounis..."
2005-11-11, Sciencedaily: Lateral Thinking Produces First Map Of Gene Transmission Citat: "...Their results clearly show genetic modification of organisms by lateral transfer is a widespread natural phenomenon, and it can occur even between distantly related organisms... it was assumed that transfer of genes could only be vertical, i.e. from parents to offspring..."
Økologi
Biologi | danish | 0.791178 |
extinction_fungus/plantextinction.txt | Skip to main content
* [ MyUW ](https://my.uw.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=myuw)
* [ Calendar ](//uw.edu/calendar/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=calendar)
* [ Directories ](https://directory.uw.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=directories)
* [ Libraries ](https://lib.uw.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=libraries)
* [ UW Medicine ](https://www.uwmedicine.org/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=uwmedicine)
* [ Maps ](//uw.edu/maps/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=maps)
* [ UW News ](//uw.edu/news/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=uwnews)
### Helpful Links
* [ Computing/IT ](https://itconnect.uw.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=computingit)
* [ Workday HCM ](https://isc.uw.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=iscworkday)
* [ Husky Card ](https://hfs.uw.edu/Husky-Card-Services/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=huskycard)
* [ UW Bothell ](https://www.uwb.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=uwbothell)
* [ UW Tacoma ](https://www.tacoma.uw.edu/?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=uwtacoma)
* [ UW Facebook ](https://www.facebook.com/UofWA?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=uwfacebook)
* [ UW Twitter ](https://twitter.com/UW?utm_source=quicklinks&utm_medium=click&utm_campaign=quicklinks&utm_term=uwtwitter)
[ University of Washington University of Washington ](http://uw.edu
"University of Washington Home")
* [ Students ](http://uw.edu/studentlife "Students")
* [ Parents ](http://uw.edu/parents "Parents")
* [ Faculty & Staff ](http://uw.edu/facultystaff "Faculty & Staff")
* [ Alumni ](http://uw.edu/alumni "Alumni")
Quick Links
* [ About ](http://www.washington.edu/news/about "About")
* [ Stories ](http://www.washington.edu/news/the-latest-news-from-the-uw/ "Stories") __
* [ All stories ](http://www.washington.edu/news/the-latest-news-from-the-uw/ "All stories")
* [ News releases ](http://www.washington.edu/news/category/news-releases/ "News releases")
* [ UW News blog ](http://www.washington.edu/news/category/blog/ "UW News blog")
* [ UW Notebook ](http://www.washington.edu/news/category/notebook/ "UW Notebook")
* [ For Washington ](https://www.washington.edu/forwashington/ "For Washington")
* [ Official notices ](http://www.washington.edu/news/category/official-notices/ "Official notices")
* [ UW in the media ](https://www.washington.edu/news/uw-in-the-media/ "UW in the media")
* [ Multimedia ](http://washington.edu/news/video/?utm_source=UW%20News&utm_medium=nav&utm_campaign=multimedia "Multimedia") __
* [ Video stories ](http://washington.edu/news/video/?utm_source=UW%20News&utm_medium=nav&utm_campaign=video "Video stories")
* [ Podcasts ](https://www.washington.edu/news/podcasts/ "Podcasts")
* [ Soundbites/b-roll ](http://washington.edu/news/for-journalists/b-roll?utm_source=UW%20News&utm_medium=nav&utm_campaign=Soundbites "Soundbites/b-roll")
* [ Interactives ](https://www.washington.edu/news/category/interactive/?utm_source=UW%20News&utm_medium=nav&utm_campaign=interactive "Interactives")
* [ Experts ](http://www.washington.edu/news/experts/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Experts "Experts") __
* [ Directory ](http://www.washington.edu/news/experts/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Experts "Directory")
* [ Expert quotes ](https://www.washington.edu/news/category/expert-quotes/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Expert-quotes "Expert quotes")
* [ COVID-19 experts ](https://www.washington.edu/news/for-journalists/uw-experts-on-novel-coronavirus-covid-19/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Covid19_experts "COVID-19 experts")
* [ Speakers Bureau ](https://www.washington.edu/news/speakers-bureau/?utm_source=UW%2520News&utm_medium=nav&utm_campaign=speakers "Speakers Bureau")
* [ Media contacts ](http://www.washington.edu/news/for-journalists/media-contacts/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Media_Contacts "Media contacts")
* [ For Journalists ](http://washington.edu/news/for-journalists?utm_source=UW%20News&utm_medium=nav&utm_campaign=For_Journalists "For Journalists")
* [ For researchers ](https://www.washington.edu/news/faculty-staff/ "For researchers") __
* [ Media Training ](https://www.washington.edu/news/faculty-staff/media-training/ "Media Training")
[ UW News ](https://www.washington.edu/news/ "UW News") Menu
* [ About ](http://www.washington.edu/news/about)
* [ Stories ](http://www.washington.edu/news/the-latest-news-from-the-uw/)
* [ All stories ](http://www.washington.edu/news/the-latest-news-from-the-uw/)
* [ News releases ](http://www.washington.edu/news/category/news-releases/)
* [ UW News blog ](http://www.washington.edu/news/category/blog/)
* [ UW Notebook ](http://www.washington.edu/news/category/notebook/)
* [ For Washington ](https://www.washington.edu/forwashington/)
* [ Official notices ](http://www.washington.edu/news/category/official-notices/)
* [ UW in the media ](https://www.washington.edu/news/uw-in-the-media/)
* [ Multimedia ](http://washington.edu/news/video/?utm_source=UW%20News&utm_medium=nav&utm_campaign=multimedia)
* [ Video stories ](http://washington.edu/news/video/?utm_source=UW%20News&utm_medium=nav&utm_campaign=video)
* [ Podcasts ](https://www.washington.edu/news/podcasts/)
* [ Soundbites/b-roll ](http://washington.edu/news/for-journalists/b-roll?utm_source=UW%20News&utm_medium=nav&utm_campaign=Soundbites)
* [ Interactives ](https://www.washington.edu/news/category/interactive/?utm_source=UW%20News&utm_medium=nav&utm_campaign=interactive)
* [ Experts ](http://www.washington.edu/news/experts/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Experts)
* [ Directory ](http://www.washington.edu/news/experts/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Experts)
* [ Expert quotes ](https://www.washington.edu/news/category/expert-quotes/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Expert-quotes)
* [ COVID-19 experts ](https://www.washington.edu/news/for-journalists/uw-experts-on-novel-coronavirus-covid-19/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Covid19_experts)
* [ Speakers Bureau ](https://www.washington.edu/news/speakers-bureau/?utm_source=UW%2520News&utm_medium=nav&utm_campaign=speakers)
* [ Media contacts ](http://www.washington.edu/news/for-journalists/media-contacts/?utm_source=UW%20News&utm_medium=nav&utm_campaign=Media_Contacts)
* [ For Journalists ](http://washington.edu/news/for-journalists?utm_source=UW%20News&utm_medium=nav&utm_campaign=For_Journalists)
* [ For researchers ](https://www.washington.edu/news/faculty-staff/)
* [ Media Training ](https://www.washington.edu/news/faculty-staff/media-training/)
* [ Home ](http://uw.edu "University of Washington")
* [ UW News ](https://www.washington.edu/news/ "UW News")
* * [ Research ](http://www.washington.edu/news/category/research/ "Research")
* Study shows plant extinction is more common than previously realized
[ Research ](http://www.washington.edu/news/category/research/) | [ Science
](http://www.washington.edu/news/category/science/) | [ UW News blog
](http://www.washington.edu/news/category/blog/)
November 24, 2020
# Study shows plant extinction is more common than previously realized
[ Andrea Godinez ](https://www.washington.edu/news/author/andrea-godinez/
"Posts by Andrea Godinez")
Burke Museum
A new study reveals that 65 plant species have gone extinct in the continental
United States and Canada since European settlement, more extinctions than any
previous scientific study has ever documented.
Led by Wesley Knapp of the North Carolina Natural Heritage Program, a group of
16 experts from across the United States — including [ Richard Olmstead
](https://www.biology.washington.edu/people/profile/richard-olmstead) , a
University of Washington professor of biology and curator of the UW’s [ Burke
Museum Herbarium ](https://www.burkemuseum.org/collections-and-
research/biology/plants-and-fungi) — collaborated on this first-of-its-kind
project to document the extinct plants of the continental United States and
Canada. Their findings [ were published
](https://conbio.onlinelibrary.wiley.com/doi/full/10.1111/cobi.13621) Aug. 28
in Conservation Biology.
[ 
](https://uw-s3-cdn.s3.us-west-2.amazonaws.com/wp-
content/uploads/sites/6/2020/11/24105237/Astragalus-kentrophyta-
var.-douglasii_Courtesy-NY-Botanic-Gardens.jpg)
_Astragalus kentrophyta var. douglasii_ , or thistle milk-vetch, is one of two
Washington-specific extinct species identified in this study. New York
Botanical Garden
The team found that most plant extinctions occurred in the western United
States, where the vegetation was minimally documented before widespread
European settlement. Since many extinctions likely occurred before scientists
analyzed an area, it is likely the 65 documented extinctions underestimate the
actual number of plant species that have been lost. Previous studies
documented far fewer plant extinctions on the North American continent.
In Washington state, the team found two confirmed extinctions: the thistle
milk-vetch, or [ _Astragalus kentrophyta_ var. _douglasii_
](https://biology.burke.washington.edu/herbarium/imagecollection/taxon.php?Taxon=Astragalus%20kentrophyta)
, and the pale bugseed, or [ _Corispermum pallidum_
](https://biology.burke.washington.edu/herbarium/imagecollection/taxon.php?Taxon=Corispermum%20pallidum)
. While neither of these Eastern Washington species were ever abundant, their
disappearance is likely due to the human impact of changing land use. This has
also dramatically reduced the populations of countless other species, many of
which are likely to follow these into extinction, unless efforts to protect
what remains of native habitat are stepped up.
[ 
](https://uw-s3-cdn.s3.us-west-2.amazonaws.com/wp-
content/uploads/sites/6/2020/11/24105547/Corispermum-pallidum.jpg)
_Corispermum pallidum_ , or pale bugseed, is one of two Washington-specific
extinct species identified in this study. Burke Museum
“Preventing extinction is the lowest bar for conservation success we can set,
yet we are not always successful,” said Knapp. “This study started as an
academic question but later developed into an opportunity to learn from what
we have lost. By studying the trends and patterns of plants that have already
gone extinct, hopefully we can learn how to prevent plant extinction going
forward.”
Of the 65 documented extinctions in the report, 64% were known only from a
single location. While conservation often focuses on protecting entire
landscapes, this finding points to the importance of small-scale site
protection to prevent extinctions. Extinct species are still being described
from old herbarium specimens, underscoring the importance of continued
documentation of the flora and supporting museum collections like the [ Burke
Herbarium ](https://www.burkemuseum.org/news/plant-extinction-more-common-
previously-realized) . _Corispermum pallidum_ , one of the species extinct in
Washington, was first collected in 1893. Yet the species wasn’t formally
“discovered” until much later when it was first described as a new species
posthumously in 1995. Only a handful of herbarium specimens exist today. The
Burke has three, including one of the two collections from 1893 and the last
known collection from 1931.
“There is no living memory of either of these species today,” Olmstead said.
“Herbarium collections record our flora as it was historically and provide
documentation of change, and loss, over time, whether through direct human
impact, or indirectly through climate change and the impact it will have on
plants. Documenting extinction is difficult, but herbarium records can say
where and when a species was last observed and guide the search for surviving
members.”
This work also highlights the need for collaborative science in addressing
large-scale conservation issues. The team of 16 botanists from across the
United States includes experts with state and federal government agencies,
numerous botanical gardens, museums, nonprofit organizations, regional
conservation groups and academic institutions. To answer the overarching
question of what exists and where, the team of experts cross-checked thousands
of records to ensure accuracy, discovering that botanical gardens occasionally
harbored the last of an extremely rare species and may not have been aware of
it.
Plants serve as the foundation for most terrestrial ecosystems. The predicted
rise of extinction rates over the next century adds even greater urgency to
the need to document plant extinctions. Anne Frances, lead botanist at
NatureServe, said, “In most cases, we can stop plants from going extinct, we
just need the resources and commitment to do so.”
_For high-resolution images, videos and interviews, contact_ [
[email protected]_ ](mailto:[email protected]) _._
_Adapted from a_ [ _press release_ ](https://www.ncdcr.gov/news/press-
releases/2020/08/31/study-led-nc-botanist-shows-plant-extinction-more-common-
previously) _by the North Carolina Department of Natural and Cultural
Resources._
[
](https://www.addtoany.com/add_to/facebook?linkurl=https%3A%2F%2Fwww.washington.edu%2Fcms%2Fnews%2F2020%2F11%2F24%2Fplant-
extinction%2F&linkname=Study%20shows%20plant%20extinction%20is%20more%20common%20than%20previously%20realized
"Facebook") [
](https://www.addtoany.com/add_to/twitter?linkurl=https%3A%2F%2Fwww.washington.edu%2Fcms%2Fnews%2F2020%2F11%2F24%2Fplant-
extinction%2F&linkname=Study%20shows%20plant%20extinction%20is%20more%20common%20than%20previously%20realized
"Twitter") [
](https://www.addtoany.com/add_to/reddit?linkurl=https%3A%2F%2Fwww.washington.edu%2Fcms%2Fnews%2F2020%2F11%2F24%2Fplant-
extinction%2F&linkname=Study%20shows%20plant%20extinction%20is%20more%20common%20than%20previously%20realized
"Reddit") [
](https://www.addtoany.com/add_to/email?linkurl=https%3A%2F%2Fwww.washington.edu%2Fcms%2Fnews%2F2020%2F11%2F24%2Fplant-
extinction%2F&linkname=Study%20shows%20plant%20extinction%20is%20more%20common%20than%20previously%20realized
"Email") [
](https://www.addtoany.com/add_to/print?linkurl=https%3A%2F%2Fwww.washington.edu%2Fcms%2Fnews%2F2020%2F11%2F24%2Fplant-
extinction%2F&linkname=Study%20shows%20plant%20extinction%20is%20more%20common%20than%20previously%20realized
"Print") [ ](https://www.addtoany.com/share)
Tag(s): [ Burke Museum of Natural History & Culture
](https://www.washington.edu/news/tag/burke-museum-of-natural-history-
culture/) • [ College of Arts & Sciences
](https://www.washington.edu/news/tag/college-of-arts-sciences/) • [
conservation ](https://www.washington.edu/news/tag/conservation/) • [
Department of Biology ](https://www.washington.edu/news/tag/department-of-
biology/) • [ Richard Olmstead ](https://www.washington.edu/news/tag/richard-
olmstead/)
* * *
### [ UW News Blog ](https://www.washington.edu/news/category/blog/)
Read more from the UW News Blog
[ Blog ](https://www.washington.edu/news/category/blog/)
## Search UW News
Search for:
## UW Experts
* ##### [ Weather ](https://www.washington.edu/news/experts/?expertise=weather-climate-weather)
* ##### [ Artificial intelligence ](https://www.washington.edu/news/experts/?expertise=artificial-intelligence-2)
* ##### [ Flooding and landslides ](https://www.washington.edu/news/experts/?expertise=flooding-and-landslides)
[ Full directory
](http://www.washington.edu/news/experts/?utm_source=UW%20News&utm_medium=experts&utm_campaign=homepage_widget)
## Categories
Browse Administrative affairs Arts and entertainment Buildings and grounds
Education Engineering Environment For UW employees Health and medicine
Honors and awards Interactive Learning News releases News roundups
Official notices Politics and government Population Health Profiles
Research Science Social science Technology UW and the community UW
Notebook UW Today blog
## Latest news releases
* [  [ New circuit boards can be repeatedly recycled
3 days ago
](https://www.washington.edu/news/2024/04/26/recyclable-circuit-boards-
vitrimer-pcb-e-waste/ "New circuit boards can be repeatedly recycled")
](https://www.washington.edu/news/2024/04/26/recyclable-circuit-boards-
vitrimer-pcb-e-waste/ "New circuit boards can be repeatedly recycled")
* [  [ Q&A: How TikTok’s ‘black box’ algorithm and design shape user behavior
5 days ago
](https://www.washington.edu/news/2024/04/24/tiktok-black-box-algorithm-and-
design-user-behavior-recommendation/ "Q&A: How TikTok’s ‘black box’ algorithm
and design shape user behavior")
](https://www.washington.edu/news/2024/04/24/tiktok-black-box-algorithm-and-
design-user-behavior-recommendation/ "Q&A: How TikTok’s ‘black box’ algorithm
and design shape user behavior")
* [  [ Daniel James Brown to address UW’s Class of 2024
6 days ago
](https://www.washington.edu/news/2024/04/23/daniel-james-brown-to-address-
uws-class-of-2024/ "Daniel James Brown to address UW’s Class of 2024")
](https://www.washington.edu/news/2024/04/23/daniel-james-brown-to-address-
uws-class-of-2024/ "Daniel James Brown to address UW’s Class of 2024")
[ More ](https://www.washington.edu/news/?cat=23)
## Connect
[ ](https://www.facebook.com/uwnews "Facebook") [ ](https://twitter.com/UWnews
"Twitter")
## UW Today Newsletter
#### Subscribe
UW Today Daily
UW Today Week in Review
* * *
#### For UW employees
[ Submission guidelines ](https://www.washington.edu/news/uw-today-submission-
guidelines/) [ Submission form
](https://form.asana.com/?k=lY4uuTrGcroevKe_0emL1Q&d=941506444463212)
[ University of Washington ](http://www.washington.edu) [
### Be boundless
](http://www.washington.edu/boundless/)
#### Connect with us:
* [ Facebook ](http://www.facebook.com/UofWA)
* [ Twitter ](http://twitter.com/UW)
* [ Instagram ](http://instagram.com/uofwa)
* [ YouTube ](http://www.youtube.com/user/uwhuskies)
* [ LinkedIn ](http://www.linkedin.com/company/university-of-washington)
* [ Pinterest ](http://www.pinterest.com/uofwa/)
* [ Accessibility ](http://www.uw.edu/accessibility)
* [ Contact Us ](http://uw.edu/contact)
* [ Jobs ](http://www.washington.edu/jobs)
* [ Campus Safety ](http://www.washington.edu/safety)
* [ My UW ](http://my.uw.edu/)
* [ Rules Docket ](http://www.washington.edu/rules/wac)
* [ Privacy ](http://www.washington.edu/online/privacy/)
* [ Terms ](http://www.washington.edu/online/terms/)
* [ Newsletter ](http://www.washington.edu/newsletter/)
© 2024 University of Washington | Seattle, WA
| biology | 145075 | https://da.wikipedia.org/wiki/Urt%20%28flertydig%29 | Urt (flertydig) | Urt har flere betydninger:
Urt – plantekategori.
Urt – ingrediens ved ølbrygning.
Urt – populærvidenskabeligt tidsskrift om botanik. | danish | 1.253406 |
evolution_not_make_our_life_longer/Evolution.txt |
Evolution is the change in the heritable characteristics of biological populations over successive generations. Evolution occurs when evolutionary processes such as natural selection and genetic drift act on genetic variation, resulting in certain characteristics becoming more or less common within a population over successive generations. The process of evolution has given rise to biodiversity at every level of biological organisation.
The theory of evolution by natural selection was conceived independently by Charles Darwin and Alfred Russel Wallace in the mid-19th century as an explanation for why organisms are adapted to their physical and biological environments. The theory was first set out in detail in Darwin's book On the Origin of Species. Evolution by natural selection is established by observable facts about living organisms: (1) more offspring are often produced than can possibly survive; (2) traits vary among individuals with respect to their morphology, physiology, and behaviour; (3) different traits confer different rates of survival and reproduction (differential fitness); and (4) traits can be passed from generation to generation (heritability of fitness). In successive generations, members of a population are therefore more likely to be replaced by the offspring of parents with favourable characteristics for that environment.
In the early 20th century, competing ideas of evolution were refuted and evolution was combined with Mendelian inheritance and population genetics to give rise to modern evolutionary theory. In this synthesis the basis for heredity is in DNA molecules that pass information from generation to generation. The processes that change DNA in a population include natural selection, genetic drift, mutation, and gene flow.
All life on Earth—including humanity—shares a last universal common ancestor (LUCA), which lived approximately 3.5–3.8 billion years ago. The fossil record includes a progression from early biogenic graphite to microbial mat fossils to fossilised multicellular organisms. Existing patterns of biodiversity have been shaped by repeated formations of new species (speciation), changes within species (anagenesis), and loss of species (extinction) throughout the evolutionary history of life on Earth. Morphological and biochemical traits tend to be more similar among species that share a more recent common ancestor, which historically was used to reconstruct phylogenetic trees, although direct comparison of genetic sequences is a more common method today.
Evolutionary biologists have continued to study various aspects of evolution by forming and testing hypotheses as well as constructing theories based on evidence from the field or laboratory and on data generated by the methods of mathematical and theoretical biology. Their discoveries have influenced not just the development of biology but also other fields including agriculture, medicine, and computer science.
Evolution in organisms occurs through changes in heritable characteristics—the inherited characteristics of an organism. In humans, for example, eye colour is an inherited characteristic and an individual might inherit the "brown-eye trait" from one of their parents. Inherited traits are controlled by genes and the complete set of genes within an organism's genome (genetic material) is called its genotype.
The complete set of observable traits that make up the structure and behaviour of an organism is called its phenotype. Some of these traits come from the interaction of its genotype with the environment while others are neutral. Some observable characteristics are not inherited. For example, suntanned skin comes from the interaction between a person's genotype and sunlight; thus, suntans are not passed on to people's children. The phenotype is the ability of the skin to tan when exposed to sunlight. However, some people tan more easily than others, due to differences in genotypic variation; a striking example are people with the inherited trait of albinism, who do not tan at all and are very sensitive to sunburn.
Heritable characteristics are passed from one generation to the next via DNA, a molecule that encodes genetic information. DNA is a long biopolymer composed of four types of bases. The sequence of bases along a particular DNA molecule specifies the genetic information, in a manner similar to a sequence of letters spelling out a sentence. Before a cell divides, the DNA is copied, so that each of the resulting two cells will inherit the DNA sequence. Portions of a DNA molecule that specify a single functional unit are called genes; different genes have different sequences of bases. Within cells, each long strand of DNA is called a chromosome. The specific location of a DNA sequence within a chromosome is known as a locus. If the DNA sequence at a locus varies between individuals, the different forms of this sequence are called alleles. DNA sequences can change through mutations, producing new alleles. If a mutation occurs within a gene, the new allele may affect the trait that the gene controls, altering the phenotype of the organism. However, while this simple correspondence between an allele and a trait works in some cases, most traits are influenced by multiple genes in a quantitative or epistatic manner.
Evolution can occur if there is genetic variation within a population. Variation comes from mutations in the genome, reshuffling of genes through sexual reproduction and migration between populations (gene flow). Despite the constant introduction of new variation through mutation and gene flow, most of the genome of a species is very similar among all individuals of that species. However, discoveries in the field of evolutionary developmental biology have demonstrated that even relatively small differences in genotype can lead to dramatic differences in phenotype both within and between species.
An individual organism's phenotype results from both its genotype and the influence of the environment it has lived in. The modern evolutionary synthesis defines evolution as the change over time in this genetic variation. The frequency of one particular allele will become more or less prevalent relative to other forms of that gene. Variation disappears when a new allele reaches the point of fixation—when it either disappears from the population or replaces the ancestral allele entirely.
Mutations are changes in the DNA sequence of a cell's genome and are the ultimate source of genetic variation in all organisms. When mutations occur, they may alter the product of a gene, or prevent the gene from functioning, or have no effect.
About half of the mutations in the coding regions of protein-coding genes are deleterious — the other half are neutral. A small percentage of the total mutations in this region confer a fitness benefit. Some of the mutations in other parts of the genome are deleterious but the vast majority are neutral. A few are beneficial.
Mutations can involve large sections of a chromosome becoming duplicated (usually by genetic recombination), which can introduce extra copies of a gene into a genome. Extra copies of genes are a major source of the raw material needed for new genes to evolve. This is important because most new genes evolve within gene families from pre-existing genes that share common ancestors. For example, the human eye uses four genes to make structures that sense light: three for colour vision and one for night vision; all four are descended from a single ancestral gene.
New genes can be generated from an ancestral gene when a duplicate copy mutates and acquires a new function. This process is easier once a gene has been duplicated because it increases the redundancy of the system; one gene in the pair can acquire a new function while the other copy continues to perform its original function. Other types of mutations can even generate entirely new genes from previously noncoding DNA, a phenomenon termed de novo gene birth.
The generation of new genes can also involve small parts of several genes being duplicated, with these fragments then recombining to form new combinations with new functions (exon shuffling). When new genes are assembled from shuffling pre-existing parts, domains act as modules with simple independent functions, which can be mixed together to produce new combinations with new and complex functions. For example, polyketide synthases are large enzymes that make antibiotics; they contain up to 100 independent domains that each catalyse one step in the overall process, like a step in an assembly line.
One example of mutation is wild boar piglets. They are camouflage coloured and show a characteristic pattern of dark and light longitudinal stripes. However, mutations in the melanocortin 1 receptor (MC1R) disrupt the pattern. The majority of pig breeds carry MC1R mutations disrupting wild-type colour and different mutations causing dominant black colouring.
In asexual organisms, genes are inherited together, or linked, as they cannot mix with genes of other organisms during reproduction. In contrast, the offspring of sexual organisms contain random mixtures of their parents' chromosomes that are produced through independent assortment. In a related process called homologous recombination, sexual organisms exchange DNA between two matching chromosomes. Recombination and reassortment do not alter allele frequencies, but instead change which alleles are associated with each other, producing offspring with new combinations of alleles. Sex usually increases genetic variation and may increase the rate of evolution.
The two-fold cost of sex was first described by John Maynard Smith. The first cost is that in sexually dimorphic species only one of the two sexes can bear young. This cost does not apply to hermaphroditic species, like most plants and many invertebrates. The second cost is that any individual who reproduces sexually can only pass on 50% of its genes to any individual offspring, with even less passed on as each new generation passes. Yet sexual reproduction is the more common means of reproduction among eukaryotes and multicellular organisms. The Red Queen hypothesis has been used to explain the significance of sexual reproduction as a means to enable continual evolution and adaptation in response to coevolution with other species in an ever-changing environment. Another hypothesis is that sexual reproduction is primarily an adaptation for promoting accurate recombinational repair of damage in germline DNA, and that increased diversity is a byproduct of this process that may sometimes be adaptively beneficial.
Gene flow is the exchange of genes between populations and between species. It can therefore be a source of variation that is new to a population or to a species. Gene flow can be caused by the movement of individuals between separate populations of organisms, as might be caused by the movement of mice between inland and coastal populations, or the movement of pollen between heavy-metal-tolerant and heavy-metal-sensitive populations of grasses.
Gene transfer between species includes the formation of hybrid organisms and horizontal gene transfer. Horizontal gene transfer is the transfer of genetic material from one organism to another organism that is not its offspring; this is most common among bacteria. In medicine, this contributes to the spread of antibiotic resistance, as when one bacteria acquires resistance genes it can rapidly transfer them to other species. Horizontal transfer of genes from bacteria to eukaryotes such as the yeast Saccharomyces cerevisiae and the adzuki bean weevil Callosobruchus chinensis has occurred. An example of larger-scale transfers are the eukaryotic bdelloid rotifers, which have received a range of genes from bacteria, fungi and plants. Viruses can also carry DNA between organisms, allowing transfer of genes even across biological domains.
Large-scale gene transfer has also occurred between the ancestors of eukaryotic cells and bacteria, during the acquisition of chloroplasts and mitochondria. It is possible that eukaryotes themselves originated from horizontal gene transfers between bacteria and archaea.
Some heritable changes cannot be explained by changes to the sequence of nucleotides in the DNA. These phenomena are classed as epigenetic inheritance systems. DNA methylation marking chromatin, self-sustaining metabolic loops, gene silencing by RNA interference and the three-dimensional conformation of proteins (such as prions) are areas where epigenetic inheritance systems have been discovered at the organismic level. Developmental biologists suggest that complex interactions in genetic networks and communication among cells can lead to heritable variations that may underlay some of the mechanics in developmental plasticity and canalisation. Heritability may also occur at even larger scales. For example, ecological inheritance through the process of niche construction is defined by the regular and repeated activities of organisms in their environment. This generates a legacy of effects that modify and feed back into the selection regime of subsequent generations. Other examples of heritability in evolution that are not under the direct control of genes include the inheritance of cultural traits and symbiogenesis.
From a neo-Darwinian perspective, evolution occurs when there are changes in the frequencies of alleles within a population of interbreeding organisms, for example, the allele for black colour in a population of moths becoming more common. Mechanisms that can lead to changes in allele frequencies include natural selection, genetic drift, and mutation bias.
Evolution by natural selection is the process by which traits that enhance survival and reproduction become more common in successive generations of a population. It embodies three principles:
More offspring are produced than can possibly survive, and these conditions produce competition between organisms for survival and reproduction. Consequently, organisms with traits that give them an advantage over their competitors are more likely to pass on their traits to the next generation than those with traits that do not confer an advantage. This teleonomy is the quality whereby the process of natural selection creates and preserves traits that are seemingly fitted for the functional roles they perform. Consequences of selection include nonrandom mating and genetic hitchhiking.
The central concept of natural selection is the evolutionary fitness of an organism. Fitness is measured by an organism's ability to survive and reproduce, which determines the size of its genetic contribution to the next generation. However, fitness is not the same as the total number of offspring: instead fitness is indicated by the proportion of subsequent generations that carry an organism's genes. For example, if an organism could survive well and reproduce rapidly, but its offspring were all too small and weak to survive, this organism would make little genetic contribution to future generations and would thus have low fitness.
If an allele increases fitness more than the other alleles of that gene, then with each generation this allele has a higher probability of becoming common within the population. These traits are said to be "selected for." Examples of traits that can increase fitness are enhanced survival and increased fecundity. Conversely, the lower fitness caused by having a less beneficial or deleterious allele results in this allele likely becoming rarer—they are "selected against."
Importantly, the fitness of an allele is not a fixed characteristic; if the environment changes, previously neutral or harmful traits may become beneficial and previously beneficial traits become harmful. However, even if the direction of selection does reverse in this way, traits that were lost in the past may not re-evolve in an identical form. However, a re-activation of dormant genes, as long as they have not been eliminated from the genome and were only suppressed perhaps for hundreds of generations, can lead to the re-occurrence of traits thought to be lost like hindlegs in dolphins, teeth in chickens, wings in wingless stick insects, tails and additional nipples in humans etc. "Throwbacks" such as these are known as atavisms.
Natural selection within a population for a trait that can vary across a range of values, such as height, can be categorised into three different types. The first is directional selection, which is a shift in the average value of a trait over time—for example, organisms slowly getting taller. Secondly, disruptive selection is selection for extreme trait values and often results in two different values becoming most common, with selection against the average value. This would be when either short or tall organisms had an advantage, but not those of medium height. Finally, in stabilising selection there is selection against extreme trait values on both ends, which causes a decrease in variance around the average value and less diversity. This would, for example, cause organisms to eventually have a similar height.
Natural selection most generally makes nature the measure against which individuals and individual traits, are more or less likely to survive. "Nature" in this sense refers to an ecosystem, that is, a system in which organisms interact with every other element, physical as well as biological, in their local environment. Eugene Odum, a founder of ecology, defined an ecosystem as: "Any unit that includes all of the organisms...in a given area interacting with the physical environment so that a flow of energy leads to clearly defined trophic structure, biotic diversity, and material cycles (i.e., exchange of materials between living and nonliving parts) within the system...." Each population within an ecosystem occupies a distinct niche, or position, with distinct relationships to other parts of the system. These relationships involve the life history of the organism, its position in the food chain and its geographic range. This broad understanding of nature enables scientists to delineate specific forces which, together, comprise natural selection.
Natural selection can act at different levels of organisation, such as genes, cells, individual organisms, groups of organisms and species. Selection can act at multiple levels simultaneously. An example of selection occurring below the level of the individual organism are genes called transposons, which can replicate and spread throughout a genome. Selection at a level above the individual, such as group selection, may allow the evolution of cooperation.
Genetic drift is the random fluctuation of allele frequencies within a population from one generation to the next. When selective forces are absent or relatively weak, allele frequencies are equally likely to drift upward or downward in each successive generation because the alleles are subject to sampling error. This drift halts when an allele eventually becomes fixed, either by disappearing from the population or by replacing the other alleles entirely. Genetic drift may therefore eliminate some alleles from a population due to chance alone. Even in the absence of selective forces, genetic drift can cause two separate populations that begin with the same genetic structure to drift apart into two divergent populations with different sets of alleles.
According to the neutral theory of molecular evolution most evolutionary changes are the result of the fixation of neutral mutations by genetic drift. In this model, most genetic changes in a population are thus the result of constant mutation pressure and genetic drift. This form of the neutral theory has been debated since it does not seem to fit some genetic variation seen in nature. A better-supported version of this model is the nearly neutral theory, according to which a mutation that would be effectively neutral in a small population is not necessarily neutral in a large population. Other theories propose that genetic drift is dwarfed by other stochastic forces in evolution, such as genetic hitchhiking, also known as genetic draft. Another concept is constructive neutral evolution (CNE), which explains that complex systems can emerge and spread into a population through neutral transitions due to the principles of excess capacity, presuppression, and ratcheting, and it has been applied in areas ranging from the origins of the spliceosome to the complex interdependence of microbial communities.
The time it takes a neutral allele to become fixed by genetic drift depends on population size; fixation is more rapid in smaller populations. The number of individuals in a population is not critical, but instead a measure known as the effective population size. The effective population is usually smaller than the total population since it takes into account factors such as the level of inbreeding and the stage of the lifecycle in which the population is the smallest. The effective population size may not be the same for every gene in the same population.
It is usually difficult to measure the relative importance of selection and neutral processes, including drift. The comparative importance of adaptive and non-adaptive forces in driving evolutionary change is an area of current research.
Mutation bias is usually conceived as a difference in expected rates for two different kinds of mutation, e.g., transition-transversion bias, GC-AT bias, deletion-insertion bias. This is related to the idea of developmental bias. Haldane and Fisher argued that, because mutation is a weak pressure easily overcome by selection, tendencies of mutation would be ineffectual except under conditions of neutral evolution or extraordinarily high mutation rates. This opposing-pressures argument was long used to dismiss the possibility of internal tendencies in evolution, until the molecular era prompted renewed interest in neutral evolution.
Noboru Sueoka and Ernst Freese proposed that systematic biases in mutation might be responsible for systematic differences in genomic GC composition between species. The identification of a GC-biased E. coli mutator strain in 1967, along with the proposal of the neutral theory, established the plausibility of mutational explanations for molecular patterns, which are now common in the molecular evolution literature.
For instance, mutation biases are frequently invoked in models of codon usage. Such models also include effects of selection, following the mutation-selection-drift model, which allows both for mutation biases and differential selection based on effects on translation. Hypotheses of mutation bias have played an important role in the development of thinking about the evolution of genome composition, including isochores. Different insertion vs. deletion biases in different taxa can lead to the evolution of different genome sizes. The hypothesis of Lynch regarding genome size relies on mutational biases toward increase or decrease in genome size.
However, mutational hypotheses for the evolution of composition suffered a reduction in scope when it was discovered that (1) GC-biased gene conversion makes an important contribution to composition in diploid organisms such as mammals and (2) bacterial genomes frequently have AT-biased mutation.
Contemporary thinking about the role of mutation biases reflects a different theory from that of Haldane and Fisher. More recent work showed that the original "pressures" theory assumes that evolution is based on standing variation: when evolution depends on events of mutation that introduce new alleles, mutational and developmental biases in the introduction of variation (arrival biases) can impose biases on evolution without requiring neutral evolution or high mutation rates.
Several studies report that the mutations implicated in adaptation reflect common mutation biases though others dispute this interpretation.
Recombination allows alleles on the same strand of DNA to become separated. However, the rate of recombination is low (approximately two events per chromosome per generation). As a result, genes close together on a chromosome may not always be shuffled away from each other and genes that are close together tend to be inherited together, a phenomenon known as linkage. This tendency is measured by finding how often two alleles occur together on a single chromosome compared to expectations, which is called their linkage disequilibrium. A set of alleles that is usually inherited in a group is called a haplotype. This can be important when one allele in a particular haplotype is strongly beneficial: natural selection can drive a selective sweep that will also cause the other alleles in the haplotype to become more common in the population; this effect is called genetic hitchhiking or genetic draft. Genetic draft caused by the fact that some neutral genes are genetically linked to others that are under selection can be partially captured by an appropriate effective population size.
A special case of natural selection is sexual selection, which is selection for any trait that increases mating success by increasing the attractiveness of an organism to potential mates. Traits that evolved through sexual selection are particularly prominent among males of several animal species. Although sexually favoured, traits such as cumbersome antlers, mating calls, large body size and bright colours often attract predation, which compromises the survival of individual males. This survival disadvantage is balanced by higher reproductive success in males that show these hard-to-fake, sexually selected traits.
Evolution influences every aspect of the form and behaviour of organisms. Most prominent are the specific behavioural and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding predators or attracting mates. Organisms can also respond to selection by cooperating with each other, usually by aiding their relatives or engaging in mutually beneficial symbiosis. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed. These outcomes of evolution are distinguished based on time scale as macroevolution versus microevolution. Macroevolution refers to evolution that occurs at or above the level of species, in particular speciation and extinction; whereas microevolution refers to smaller evolutionary changes within a species or population, in particular shifts in allele frequency and adaptation. Macroevolution the outcome of long periods of microevolution. Thus, the distinction between micro- and macroevolution is not a fundamental one—the difference is simply the time involved. However, in macroevolution, the traits of the entire species may be important. For instance, a large amount of variation among individuals allows a species to rapidly adapt to new habitats, lessening the chance of it going extinct, while a wide geographic range increases the chance of speciation, by making it more likely that part of the population will become isolated. In this sense, microevolution and macroevolution might involve selection at different levels—with microevolution acting on genes and organisms, versus macroevolutionary processes such as species selection acting on entire species and affecting their rates of speciation and extinction.
A common misconception is that evolution has goals, long-term plans, or an innate tendency for "progress", as expressed in beliefs such as orthogenesis and evolutionism; realistically however, evolution has no long-term goal and does not necessarily produce greater complexity. Although complex species have evolved, they occur as a side effect of the overall number of organisms increasing and simple forms of life still remain more common in the biosphere. For example, the overwhelming majority of species are microscopic prokaryotes, which form about half the world's biomass despite their small size, and constitute the vast majority of Earth's biodiversity. Simple organisms have therefore been the dominant form of life on Earth throughout its history and continue to be the main form of life up to the present day, with complex life only appearing more diverse because it is more noticeable. Indeed, the evolution of microorganisms is particularly important to evolutionary research, since their rapid reproduction allows the study of experimental evolution and the observation of evolution and adaptation in real time.
Adaptation is the process that makes organisms better suited to their habitat. Also, the term adaptation may refer to a trait that is important for an organism's survival. For example, the adaptation of horses' teeth to the grinding of grass. By using the term adaptation for the evolutionary process and adaptive trait for the product (the bodily part or function), the two senses of the word may be distinguished. Adaptations are produced by natural selection. The following definitions are due to Theodosius Dobzhansky:
Adaptation may cause either the gain of a new feature, or the loss of an ancestral feature. An example that shows both types of change is bacterial adaptation to antibiotic selection, with genetic changes causing antibiotic resistance by both modifying the target of the drug, or increasing the activity of transporters that pump the drug out of the cell. Other striking examples are the bacteria Escherichia coli evolving the ability to use citric acid as a nutrient in a long-term laboratory experiment, Flavobacterium evolving a novel enzyme that allows these bacteria to grow on the by-products of nylon manufacturing, and the soil bacterium Sphingobium evolving an entirely new metabolic pathway that degrades the synthetic pesticide pentachlorophenol. An interesting but still controversial idea is that some adaptations might increase the ability of organisms to generate genetic diversity and adapt by natural selection (increasing organisms' evolvability).
Adaptation occurs through the gradual modification of existing structures. Consequently, structures with similar internal organisation may have different functions in related organisms. This is the result of a single ancestral structure being adapted to function in different ways. The bones within bat wings, for example, are very similar to those in mice feet and primate hands, due to the descent of all these structures from a common mammalian ancestor. However, since all living organisms are related to some extent, even organs that appear to have little or no structural similarity, such as arthropod, squid and vertebrate eyes, or the limbs and wings of arthropods and vertebrates, can depend on a common set of homologous genes that control their assembly and function; this is called deep homology.
During evolution, some structures may lose their original function and become vestigial structures. Such structures may have little or no function in a current species, yet have a clear function in ancestral species, or other closely related species. Examples include pseudogenes, the non-functional remains of eyes in blind cave-dwelling fish, wings in flightless birds, the presence of hip bones in whales and snakes, and sexual traits in organisms that reproduce via asexual reproduction. Examples of vestigial structures in humans include wisdom teeth, the coccyx, the vermiform appendix, and other behavioural vestiges such as goose bumps and primitive reflexes.
However, many traits that appear to be simple adaptations are in fact exaptations: structures originally adapted for one function, but which coincidentally became somewhat useful for some other function in the process. One example is the African lizard Holaspis guentheri, which developed an extremely flat head for hiding in crevices, as can be seen by looking at its near relatives. However, in this species, the head has become so flattened that it assists in gliding from tree to tree—an exaptation. Within cells, molecular machines such as the bacterial flagella and protein sorting machinery evolved by the recruitment of several pre-existing proteins that previously had different functions. Another example is the recruitment of enzymes from glycolysis and xenobiotic metabolism to serve as structural proteins called crystallins within the lenses of organisms' eyes.
An area of current investigation in evolutionary developmental biology is the developmental basis of adaptations and exaptations. This research addresses the origin and evolution of embryonic development and how modifications of development and developmental processes produce novel features. These studies have shown that evolution can alter development to produce new structures, such as embryonic bone structures that develop into the jaw in other animals instead forming part of the middle ear in mammals. It is also possible for structures that have been lost in evolution to reappear due to changes in developmental genes, such as a mutation in chickens causing embryos to grow teeth similar to those of crocodiles. It is now becoming clear that most alterations in the form of organisms are due to changes in a small set of conserved genes.
Interactions between organisms can produce both conflict and cooperation. When the interaction is between pairs of species, such as a pathogen and a host, or a predator and its prey, these species can develop matched sets of adaptations. Here, the evolution of one species causes adaptations in a second species. These changes in the second species then, in turn, cause new adaptations in the first species. This cycle of selection and response is called coevolution. An example is the production of tetrodotoxin in the rough-skinned newt and the evolution of tetrodotoxin resistance in its predator, the common garter snake. In this predator-prey pair, an evolutionary arms race has produced high levels of toxin in the newt and correspondingly high levels of toxin resistance in the snake.
Not all co-evolved interactions between species involve conflict. Many cases of mutually beneficial interactions have evolved. For instance, an extreme cooperation exists between plants and the mycorrhizal fungi that grow on their roots and aid the plant in absorbing nutrients from the soil. This is a reciprocal relationship as the plants provide the fungi with sugars from photosynthesis. Here, the fungi actually grow inside plant cells, allowing them to exchange nutrients with their hosts, while sending signals that suppress the plant immune system.
Coalitions between organisms of the same species have also evolved. An extreme case is the eusociality found in social insects, such as bees, termites and ants, where sterile insects feed and guard the small number of organisms in a colony that are able to reproduce. On an even smaller scale, the somatic cells that make up the body of an animal limit their reproduction so they can maintain a stable organism, which then supports a small number of the animal's germ cells to produce offspring. Here, somatic cells respond to specific signals that instruct them whether to grow, remain as they are, or die. If cells ignore these signals and multiply inappropriately, their uncontrolled growth causes cancer.
Such cooperation within species may have evolved through the process of kin selection, which is where one organism acts to help raise a relative's offspring. This activity is selected for because if the helping individual contains alleles which promote the helping activity, it is likely that its kin will also contain these alleles and thus those alleles will be passed on. Other processes that may promote cooperation include group selection, where cooperation provides benefits to a group of organisms.
Speciation is the process where a species diverges into two or more descendant species.
There are multiple ways to define the concept of "species." The choice of definition is dependent on the particularities of the species concerned. For example, some species concepts apply more readily toward sexually reproducing organisms while others lend themselves better toward asexual organisms. Despite the diversity of various species concepts, these various concepts can be placed into one of three broad philosophical approaches: interbreeding, ecological and phylogenetic. The Biological Species Concept (BSC) is a classic example of the interbreeding approach. Defined by evolutionary biologist Ernst Mayr in 1942, the BSC states that "species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups." Despite its wide and long-term use, the BSC like other species concepts is not without controversy, for example, because genetic recombination among prokaryotes is not an intrinsic aspect of reproduction; this is called the species problem. Some researchers have attempted a unifying monistic definition of species, while others adopt a pluralistic approach and suggest that there may be different ways to logically interpret the definition of a species.
Barriers to reproduction between two diverging sexual populations are required for the populations to become new species. Gene flow may slow this process by spreading the new genetic variants also to the other populations. Depending on how far two species have diverged since their most recent common ancestor, it may still be possible for them to produce offspring, as with horses and donkeys mating to produce mules. Such hybrids are generally infertile. In this case, closely related species may regularly interbreed, but hybrids will be selected against and the species will remain distinct. However, viable hybrids are occasionally formed and these new species can either have properties intermediate between their parent species, or possess a totally new phenotype. The importance of hybridisation in producing new species of animals is unclear, although cases have been seen in many types of animals, with the gray tree frog being a particularly well-studied example.
Speciation has been observed multiple times under both controlled laboratory conditions and in nature. In sexually reproducing organisms, speciation results from reproductive isolation followed by genealogical divergence. There are four primary geographic modes of speciation. The most common in animals is allopatric speciation, which occurs in populations initially isolated geographically, such as by habitat fragmentation or migration. Selection under these conditions can produce very rapid changes in the appearance and behaviour of organisms. As selection and drift act independently on populations isolated from the rest of their species, separation may eventually produce organisms that cannot interbreed.
The second mode of speciation is peripatric speciation, which occurs when small populations of organisms become isolated in a new environment. This differs from allopatric speciation in that the isolated populations are numerically much smaller than the parental population. Here, the founder effect causes rapid speciation after an increase in inbreeding increases selection on homozygotes, leading to rapid genetic change.
The third mode is parapatric speciation. This is similar to peripatric speciation in that a small population enters a new habitat, but differs in that there is no physical separation between these two populations. Instead, speciation results from the evolution of mechanisms that reduce gene flow between the two populations. Generally this occurs when there has been a drastic change in the environment within the parental species' habitat. One example is the grass Anthoxanthum odoratum, which can undergo parapatric speciation in response to localised metal pollution from mines. Here, plants evolve that have resistance to high levels of metals in the soil. Selection against interbreeding with the metal-sensitive parental population produced a gradual change in the flowering time of the metal-resistant plants, which eventually produced complete reproductive isolation. Selection against hybrids between the two populations may cause reinforcement, which is the evolution of traits that promote mating within a species, as well as character displacement, which is when two species become more distinct in appearance.
Finally, in sympatric speciation species diverge without geographic isolation or changes in habitat. This form is rare since even a small amount of gene flow may remove genetic differences between parts of a population. Generally, sympatric speciation in animals requires the evolution of both genetic differences and nonrandom mating, to allow reproductive isolation to evolve.
One type of sympatric speciation involves crossbreeding of two related species to produce a new hybrid species. This is not common in animals as animal hybrids are usually sterile. This is because during meiosis the homologous chromosomes from each parent are from different species and cannot successfully pair. However, it is more common in plants because plants often double their number of chromosomes, to form polyploids. This allows the chromosomes from each parental species to form matching pairs during meiosis, since each parent's chromosomes are represented by a pair already. An example of such a speciation event is when the plant species Arabidopsis thaliana and Arabidopsis arenosa crossbred to give the new species Arabidopsis suecica. This happened about 20,000 years ago, and the speciation process has been repeated in the laboratory, which allows the study of the genetic mechanisms involved in this process. Indeed, chromosome doubling within a species may be a common cause of reproductive isolation, as half the doubled chromosomes will be unmatched when breeding with undoubled organisms.
Speciation events are important in the theory of punctuated equilibrium, which accounts for the pattern in the fossil record of short "bursts" of evolution interspersed with relatively long periods of stasis, where species remain relatively unchanged. In this theory, speciation and rapid evolution are linked, with natural selection and genetic drift acting most strongly on organisms undergoing speciation in novel habitats or small populations. As a result, the periods of stasis in the fossil record correspond to the parental population and the organisms undergoing speciation and rapid evolution are found in small populations or geographically restricted habitats and therefore rarely being preserved as fossils.
Extinction is the disappearance of an entire species. Extinction is not an unusual event, as species regularly appear through speciation and disappear through extinction. Nearly all animal and plant species that have lived on Earth are now extinct, and extinction appears to be the ultimate fate of all species. These extinctions have happened continuously throughout the history of life, although the rate of extinction spikes in occasional mass extinction events. The Cretaceous–Paleogene extinction event, during which the non-avian dinosaurs became extinct, is the most well-known, but the earlier Permian–Triassic extinction event was even more severe, with approximately 96% of all marine species driven to extinction. The Holocene extinction event is an ongoing mass extinction associated with humanity's expansion across the globe over the past few thousand years. Present-day extinction rates are 100–1000 times greater than the background rate and up to 30% of current species may be extinct by the mid 21st century. Human activities are now the primary cause of the ongoing extinction event; global warming may further accelerate it in the future. Despite the estimated extinction of more than 99% of all species that ever lived on Earth, about 1 trillion species are estimated to be on Earth currently with only one-thousandth of 1% described.
The role of extinction in evolution is not very well understood and may depend on which type of extinction is considered. The causes of the continuous "low-level" extinction events, which form the majority of extinctions, may be the result of competition between species for limited resources (the competitive exclusion principle). If one species can out-compete another, this could produce species selection, with the fitter species surviving and the other species being driven to extinction. The intermittent mass extinctions are also important, but instead of acting as a selective force, they drastically reduce diversity in a nonspecific manner and promote bursts of rapid evolution and speciation in survivors.
Concepts and models used in evolutionary biology, such as natural selection, have many applications.
Artificial selection is the intentional selection of traits in a population of organisms. This has been used for thousands of years in the domestication of plants and animals. More recently, such selection has become a vital part of genetic engineering, with selectable markers such as antibiotic resistance genes being used to manipulate DNA. Proteins with valuable properties have evolved by repeated rounds of mutation and selection (for example modified enzymes and new antibodies) in a process called directed evolution.
Understanding the changes that have occurred during an organism's evolution can reveal the genes needed to construct parts of the body, genes which may be involved in human genetic disorders. For example, the Mexican tetra is an albino cavefish that lost its eyesight during evolution. Breeding together different populations of this blind fish produced some offspring with functional eyes, since different mutations had occurred in the isolated populations that had evolved in different caves. This helped identify genes required for vision and pigmentation.
Evolutionary theory has many applications in medicine. Many human diseases are not static phenomena, but capable of evolution. Viruses, bacteria, fungi and cancers evolve to be resistant to host immune defences, as well as to pharmaceutical drugs. These same problems occur in agriculture with pesticide and herbicide resistance. It is possible that we are facing the end of the effective life of most of available antibiotics and predicting the evolution and evolvability of our pathogens and devising strategies to slow or circumvent it is requiring deeper knowledge of the complex forces driving evolution at the molecular level.
In computer science, simulations of evolution using evolutionary algorithms and artificial life started in the 1960s and were extended with simulation of artificial selection. Artificial evolution became a widely recognised optimisation method as a result of the work of Ingo Rechenberg in the 1960s. He used evolution strategies to solve complex engineering problems. Genetic algorithms in particular became popular through the writing of John Henry Holland. Practical applications also include automatic evolution of computer programmes. Evolutionary algorithms are now used to solve multi-dimensional problems more efficiently than software produced by human designers and also to optimise the design of systems.
The Earth is about 4.54 billion years old. The earliest undisputed evidence of life on Earth dates from at least 3.5 billion years ago, during the Eoarchean Era after a geological crust started to solidify following the earlier molten Hadean Eon. Microbial mat fossils have been found in 3.48 billion-year-old sandstone in Western Australia. Other early physical evidence of a biogenic substance is graphite in 3.7 billion-year-old metasedimentary rocks discovered in Western Greenland as well as "remains of biotic life" found in 4.1 billion-year-old rocks in Western Australia. Commenting on the Australian findings, Stephen Blair Hedges wrote: "If life arose relatively quickly on Earth, then it could be common in the universe." In July 2016, scientists reported identifying a set of 355 genes from the last universal common ancestor (LUCA) of all organisms living on Earth.
More than 99% of all species, amounting to over five billion species, that ever lived on Earth are estimated to be extinct. Estimates on the number of Earth's current species range from 10 million to 14 million, of which about 1.9 million are estimated to have been named and 1.6 million documented in a central database to date, leaving at least 80% not yet described.
Highly energetic chemistry is thought to have produced a self-replicating molecule around 4 billion years ago, and half a billion years later the last common ancestor of all life existed. The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions. The beginning of life may have included self-replicating molecules such as RNA and the assembly of simple cells.
All organisms on Earth are descended from a common ancestor or ancestral gene pool. Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events. The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits. Fourth, organisms can be classified using these similarities into a hierarchy of nested groups, similar to a family tree.
Due to horizontal gene transfer, this "tree of life" may be more complicated than a simple branching tree, since some genes have spread independently between distantly related species. To solve this problem and others, some authors prefer to use the "Coral of life" as a metaphor or a mathematical model to illustrate the evolution of life. This view dates back to an idea briefly mentioned by Darwin but later abandoned.
Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record. By comparing the anatomies of both modern and extinct species, palaeontologists can infer the lineages of those species. However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.
More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and amino acids. The development of molecular genetics has revealed the record of evolution left in organisms' genomes: dating when species diverged through the molecular clock produced by mutations. For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 98% of their genomes and analysing the few areas where they differ helps shed light on when the common ancestor of these species existed.
Prokaryotes inhabited the Earth from approximately 3–4 billion years ago. No obvious changes in morphology or cellular organisation occurred in these organisms over the next few billion years. The eukaryotic cells emerged between 1.6 and 2.7 billion years ago. The next major change in cell structure came when bacteria were engulfed by eukaryotic cells, in a cooperative association called endosymbiosis. The engulfed bacteria and the host cell then underwent coevolution, with the bacteria evolving into either mitochondria or hydrogenosomes. Another engulfment of cyanobacterial-like organisms led to the formation of chloroplasts in algae and plants.
The history of life was that of the unicellular eukaryotes, prokaryotes and archaea until about 610 million years ago when multicellular organisms began to appear in the oceans in the Ediacaran period. The evolution of multicellularity occurred in multiple independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime moulds and myxobacteria. In January 2016, scientists reported that, about 800 million years ago, a minor genetic change in a single molecule called GK-PID may have allowed organisms to go from a single cell organism to one of many cells.
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over approximately 10 million years, in an event called the Cambrian explosion. Here, the majority of types of modern animals appeared in the fossil record, as well as unique lineages that subsequently became extinct. Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.
About 500 million years ago, plants and fungi colonised the land and were soon followed by arthropods and other animals. Insects were particularly successful and even today make up the majority of animal species. Amphibians first appeared around 364 million years ago, followed by early amniotes and birds around 155 million years ago (both from "reptile"-like lineages), mammals around 129 million years ago, Homininae around 10 million years ago and modern humans around 250,000 years ago. However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to be highly successful and dominate the Earth, with the majority of both biomass and species being prokaryotes.
The proposal that one type of organism could descend from another type goes back to some of the first pre-Socratic Greek philosophers, such as Anaximander and Empedocles. Such proposals survived into Roman times. The poet and philosopher Lucretius followed Empedocles in his masterwork De rerum natura (lit. 'On the Nature of Things').
In contrast to these materialistic views, Aristotelianism had considered all natural things as actualisations of fixed natural possibilities, known as forms. This became part of a medieval teleological understanding of nature in which all things have an intended role to play in a divine cosmic order. Variations of this idea became the standard understanding of the Middle Ages and were integrated into Christian learning, but Aristotle did not demand that real types of organisms always correspond one-for-one with exact metaphysical forms and specifically gave examples of how new types of living things could come to be.
A number of Arab Muslim scholars wrote about evolution, most notably Ibn Khaldun, who wrote the book Muqaddimah in 1377 AD, in which he asserted that humans developed from "the world of the monkeys", in a process by which "species become more numerous".
The "New Science" of the 17th century rejected the Aristotelian approach. It sought to explain natural phenomena in terms of physical laws that were the same for all visible things and that did not require the existence of any fixed natural categories or divine cosmic order. However, this new approach was slow to take root in the biological sciences: the last bastion of the concept of fixed natural types. John Ray applied one of the previously more general terms for fixed natural types, "species", to plant and animal types, but he strictly identified each type of living thing as a species and proposed that each species could be defined by the features that perpetuated themselves generation after generation. The biological classification introduced by Carl Linnaeus in 1735 explicitly recognised the hierarchical nature of species relationships, but still viewed species as fixed according to a divine plan.
Other naturalists of this time speculated on the evolutionary change of species over time according to natural laws. In 1751, Pierre Louis Maupertuis wrote of natural modifications occurring during reproduction and accumulating over many generations to produce new species. Georges-Louis Leclerc, Comte de Buffon, suggested that species could degenerate into different organisms, and Erasmus Darwin proposed that all warm-blooded animals could have descended from a single microorganism (or "filament"). The first full-fledged evolutionary scheme was Jean-Baptiste Lamarck's "transmutation" theory of 1809, which envisaged spontaneous generation continually producing simple forms of life that developed greater complexity in parallel lineages with an inherent progressive tendency, and postulated that on a local level, these lineages adapted to the environment by inheriting changes caused by their use or disuse in parents. (The latter process was later called Lamarckism.) These ideas were condemned by established naturalists as speculation lacking empirical support. In particular, Georges Cuvier insisted that species were unrelated and fixed, their similarities reflecting divine design for functional needs. In the meantime, Ray's ideas of benevolent design had been developed by William Paley into the Natural Theology or Evidences of the Existence and Attributes of the Deity (1802), which proposed complex adaptations as evidence of divine design and which was admired by Charles Darwin.
The crucial break from the concept of constant typological classes or types in biology came with the theory of evolution through natural selection, which was formulated by Charles Darwin and Alfred Wallace in terms of variable populations. Darwin used the expression "descent with modification" rather than "evolution". Partly influenced by An Essay on the Principle of Population (1798) by Thomas Robert Malthus, Darwin noted that population growth would lead to a "struggle for existence" in which favourable variations prevailed as others perished. In each generation, many offspring fail to survive to an age of reproduction because of limited resources. This could explain the diversity of plants and animals from a common ancestry through the working of natural laws in the same way for all types of organism. Darwin developed his theory of "natural selection" from 1838 onwards and was writing up his "big book" on the subject when Alfred Russel Wallace sent him a version of virtually the same theory in 1858. Their separate papers were presented together at an 1858 meeting of the Linnean Society of London. At the end of 1859, Darwin's publication of his "abstract" as On the Origin of Species explained natural selection in detail and in a way that led to an increasingly wide acceptance of Darwin's concepts of evolution at the expense of alternative theories. Thomas Henry Huxley applied Darwin's ideas to humans, using paleontology and comparative anatomy to provide strong evidence that humans and apes shared a common ancestry. Some were disturbed by this since it implied that humans did not have a special place in the universe.
The mechanisms of reproductive heritability and the origin of new traits remained a mystery. Towards this end, Darwin developed his provisional theory of pangenesis. In 1865, Gregor Mendel reported that traits were inherited in a predictable manner through the independent assortment and segregation of elements (later known as genes). Mendel's laws of inheritance eventually supplanted most of Darwin's pangenesis theory. August Weismann made the important distinction between germ cells that give rise to gametes (such as sperm and egg cells) and the somatic cells of the body, demonstrating that heredity passes through the germ line only. Hugo de Vries connected Darwin's pangenesis theory to Weismann's germ/soma cell distinction and proposed that Darwin's pangenes were concentrated in the cell nucleus and when expressed they could move into the cytoplasm to change the cell's structure. De Vries was also one of the researchers who made Mendel's work well known, believing that Mendelian traits corresponded to the transfer of heritable variations along the germline. To explain how new variants originate, de Vries developed a mutation theory that led to a temporary rift between those who accepted Darwinian evolution and biometricians who allied with de Vries. In the 1930s, pioneers in the field of population genetics, such as Ronald Fisher, Sewall Wright and J. B. S. Haldane set the foundations of evolution onto a robust statistical philosophy. The false contradiction between Darwin's theory, genetic mutations, and Mendelian inheritance was thus reconciled.
In the 1920s and 1930s, the modern synthesis connected natural selection and population genetics, based on Mendelian inheritance, into a unified theory that included random genetic drift, mutation, and gene flow. This new version of evolutionary theory focused on changes in allele frequencies in population. It explained patterns observed across species in populations, through fossil transitions in palaeontology.
Since then, further syntheses have extended evolution's explanatory power in the light of numerous discoveries, to cover biological phenomena across the whole of the biological hierarchy from genes to populations.
The publication of the structure of DNA by James Watson and Francis Crick with contribution of Rosalind Franklin in 1953 demonstrated a physical mechanism for inheritance. Molecular biology improved understanding of the relationship between genotype and phenotype. Advances were also made in phylogenetic systematics, mapping the transition of traits into a comparative and testable framework through the publication and use of evolutionary trees. In 1973, evolutionary biologist Theodosius Dobzhansky penned that "nothing in biology makes sense except in the light of evolution", because it has brought to light the relations of what first seemed disjointed facts in natural history into a coherent explanatory body of knowledge that describes and predicts many observable facts about life on this planet.
One extension, known as evolutionary developmental biology and informally called "evo-devo," emphasises how changes between generations (evolution) act on patterns of change within individual organisms (development). Since the beginning of the 21st century, some biologists have argued for an extended evolutionary synthesis, which would account for the effects of non-genetic inheritance modes, such as epigenetics, parental effects, ecological inheritance and cultural inheritance, and evolvability.
In the 19th century, particularly after the publication of On the Origin of Species in 1859, the idea that life had evolved was an active source of academic debate centred on the philosophical, social and religious implications of evolution. Today, the modern evolutionary synthesis is accepted by a vast majority of scientists. However, evolution remains a contentious concept for some theists.
While various religions and denominations have reconciled their beliefs with evolution through concepts such as theistic evolution, there are creationists who believe that evolution is contradicted by the creation myths found in their religions and who raise various objections to evolution. As had been demonstrated by responses to the publication of Vestiges of the Natural History of Creation in 1844, the most controversial aspect of evolutionary biology is the implication of human evolution that humans share common ancestry with apes and that the mental and moral faculties of humanity have the same types of natural causes as other inherited traits in animals. In some countries, notably the United States, these tensions between science and religion have fuelled the current creation–evolution controversy, a religious conflict focusing on politics and public education. While other scientific fields such as cosmology and Earth science also conflict with literal interpretations of many religious texts, evolutionary biology experiences significantly more opposition from religious literalists.
The teaching of evolution in American secondary school biology classes was uncommon in most of the first half of the 20th century. The Scopes Trial decision of 1925 caused the subject to become very rare in American secondary biology textbooks for a generation, but it was gradually re-introduced later and became legally protected with the 1968 Epperson v. Arkansas decision. Since then, the competing religious belief of creationism was legally disallowed in secondary school curricula in various decisions in the 1970s and 1980s, but it returned in pseudoscientific form as intelligent design (ID), to be excluded once again in the 2005 Kitzmiller v. Dover Area School District case. The debate over Darwin's ideas did not generate significant controversy in China.
Heredity
Further information: Introduction to genetics, Genetics, and Heredity
DNA structure. Bases are in the centre, surrounded by phosphate–sugar chains in a double helix.
Evolution in organisms occurs through changes in heritable characteristics—the inherited characteristics of an organism. In humans, for example, eye colour is an inherited characteristic and an individual might inherit the "brown-eye trait" from one of their parents. Inherited traits are controlled by genes and the complete set of genes within an organism's genome (genetic material) is called its genotype.
The complete set of observable traits that make up the structure and behaviour of an organism is called its phenotype. Some of these traits come from the interaction of its genotype with the environment while others are neutral. Some observable characteristics are not inherited. For example, suntanned skin comes from the interaction between a person's genotype and sunlight; thus, suntans are not passed on to people's children. The phenotype is the ability of the skin to tan when exposed to sunlight. However, some people tan more easily than others, due to differences in genotypic variation; a striking example are people with the inherited trait of albinism, who do not tan at all and are very sensitive to sunburn.
Heritable characteristics are passed from one generation to the next via DNA, a molecule that encodes genetic information. DNA is a long biopolymer composed of four types of bases. The sequence of bases along a particular DNA molecule specifies the genetic information, in a manner similar to a sequence of letters spelling out a sentence. Before a cell divides, the DNA is copied, so that each of the resulting two cells will inherit the DNA sequence. Portions of a DNA molecule that specify a single functional unit are called genes; different genes have different sequences of bases. Within cells, each long strand of DNA is called a chromosome. The specific location of a DNA sequence within a chromosome is known as a locus. If the DNA sequence at a locus varies between individuals, the different forms of this sequence are called alleles. DNA sequences can change through mutations, producing new alleles. If a mutation occurs within a gene, the new allele may affect the trait that the gene controls, altering the phenotype of the organism. However, while this simple correspondence between an allele and a trait works in some cases, most traits are influenced by multiple genes in a quantitative or epistatic manner.
Sources of variation
Main article: Genetic variation
Further information: Genetic diversity and Population genetics
White peppered mothBlack morph in peppered moth evolution
Evolution can occur if there is genetic variation within a population. Variation comes from mutations in the genome, reshuffling of genes through sexual reproduction and migration between populations (gene flow). Despite the constant introduction of new variation through mutation and gene flow, most of the genome of a species is very similar among all individuals of that species. However, discoveries in the field of evolutionary developmental biology have demonstrated that even relatively small differences in genotype can lead to dramatic differences in phenotype both within and between species.
An individual organism's phenotype results from both its genotype and the influence of the environment it has lived in. The modern evolutionary synthesis defines evolution as the change over time in this genetic variation. The frequency of one particular allele will become more or less prevalent relative to other forms of that gene. Variation disappears when a new allele reaches the point of fixation—when it either disappears from the population or replaces the ancestral allele entirely.
Mutation
Main article: Mutation
Duplication of part of a chromosome
Mutations are changes in the DNA sequence of a cell's genome and are the ultimate source of genetic variation in all organisms. When mutations occur, they may alter the product of a gene, or prevent the gene from functioning, or have no effect.
About half of the mutations in the coding regions of protein-coding genes are deleterious — the other half are neutral. A small percentage of the total mutations in this region confer a fitness benefit. Some of the mutations in other parts of the genome are deleterious but the vast majority are neutral. A few are beneficial.
Mutations can involve large sections of a chromosome becoming duplicated (usually by genetic recombination), which can introduce extra copies of a gene into a genome. Extra copies of genes are a major source of the raw material needed for new genes to evolve. This is important because most new genes evolve within gene families from pre-existing genes that share common ancestors. For example, the human eye uses four genes to make structures that sense light: three for colour vision and one for night vision; all four are descended from a single ancestral gene.
New genes can be generated from an ancestral gene when a duplicate copy mutates and acquires a new function. This process is easier once a gene has been duplicated because it increases the redundancy of the system; one gene in the pair can acquire a new function while the other copy continues to perform its original function. Other types of mutations can even generate entirely new genes from previously noncoding DNA, a phenomenon termed de novo gene birth.
The generation of new genes can also involve small parts of several genes being duplicated, with these fragments then recombining to form new combinations with new functions (exon shuffling). When new genes are assembled from shuffling pre-existing parts, domains act as modules with simple independent functions, which can be mixed together to produce new combinations with new and complex functions. For example, polyketide synthases are large enzymes that make antibiotics; they contain up to 100 independent domains that each catalyse one step in the overall process, like a step in an assembly line.
One example of mutation is wild boar piglets. They are camouflage coloured and show a characteristic pattern of dark and light longitudinal stripes. However, mutations in the melanocortin 1 receptor (MC1R) disrupt the pattern. The majority of pig breeds carry MC1R mutations disrupting wild-type colour and different mutations causing dominant black colouring.
Sex and recombination
Further information: Sexual reproduction, Genetic recombination, and Evolution of sexual reproduction
In asexual organisms, genes are inherited together, or linked, as they cannot mix with genes of other organisms during reproduction. In contrast, the offspring of sexual organisms contain random mixtures of their parents' chromosomes that are produced through independent assortment. In a related process called homologous recombination, sexual organisms exchange DNA between two matching chromosomes. Recombination and reassortment do not alter allele frequencies, but instead change which alleles are associated with each other, producing offspring with new combinations of alleles. Sex usually increases genetic variation and may increase the rate of evolution.
This diagram illustrates the twofold cost of sex. If each individual were to contribute to the same number of offspring (two), (a) the sexual population remains the same size each generation, where the (b) Asexual reproduction population doubles in size each generation.
The two-fold cost of sex was first described by John Maynard Smith. The first cost is that in sexually dimorphic species only one of the two sexes can bear young. This cost does not apply to hermaphroditic species, like most plants and many invertebrates. The second cost is that any individual who reproduces sexually can only pass on 50% of its genes to any individual offspring, with even less passed on as each new generation passes. Yet sexual reproduction is the more common means of reproduction among eukaryotes and multicellular organisms. The Red Queen hypothesis has been used to explain the significance of sexual reproduction as a means to enable continual evolution and adaptation in response to coevolution with other species in an ever-changing environment. Another hypothesis is that sexual reproduction is primarily an adaptation for promoting accurate recombinational repair of damage in germline DNA, and that increased diversity is a byproduct of this process that may sometimes be adaptively beneficial.
Gene flow
Further information: Gene flow
Gene flow is the exchange of genes between populations and between species. It can therefore be a source of variation that is new to a population or to a species. Gene flow can be caused by the movement of individuals between separate populations of organisms, as might be caused by the movement of mice between inland and coastal populations, or the movement of pollen between heavy-metal-tolerant and heavy-metal-sensitive populations of grasses.
Gene transfer between species includes the formation of hybrid organisms and horizontal gene transfer. Horizontal gene transfer is the transfer of genetic material from one organism to another organism that is not its offspring; this is most common among bacteria. In medicine, this contributes to the spread of antibiotic resistance, as when one bacteria acquires resistance genes it can rapidly transfer them to other species. Horizontal transfer of genes from bacteria to eukaryotes such as the yeast Saccharomyces cerevisiae and the adzuki bean weevil Callosobruchus chinensis has occurred. An example of larger-scale transfers are the eukaryotic bdelloid rotifers, which have received a range of genes from bacteria, fungi and plants. Viruses can also carry DNA between organisms, allowing transfer of genes even across biological domains.
Large-scale gene transfer has also occurred between the ancestors of eukaryotic cells and bacteria, during the acquisition of chloroplasts and mitochondria. It is possible that eukaryotes themselves originated from horizontal gene transfers between bacteria and archaea.
Epigenetics
Further information: Epigenetics
Some heritable changes cannot be explained by changes to the sequence of nucleotides in the DNA. These phenomena are classed as epigenetic inheritance systems. DNA methylation marking chromatin, self-sustaining metabolic loops, gene silencing by RNA interference and the three-dimensional conformation of proteins (such as prions) are areas where epigenetic inheritance systems have been discovered at the organismic level. Developmental biologists suggest that complex interactions in genetic networks and communication among cells can lead to heritable variations that may underlay some of the mechanics in developmental plasticity and canalisation. Heritability may also occur at even larger scales. For example, ecological inheritance through the process of niche construction is defined by the regular and repeated activities of organisms in their environment. This generates a legacy of effects that modify and feed back into the selection regime of subsequent generations. Other examples of heritability in evolution that are not under the direct control of genes include the inheritance of cultural traits and symbiogenesis.
Evolutionary forces
Mutation followed by natural selection results in a population with darker colouration.
From a neo-Darwinian perspective, evolution occurs when there are changes in the frequencies of alleles within a population of interbreeding organisms, for example, the allele for black colour in a population of moths becoming more common. Mechanisms that can lead to changes in allele frequencies include natural selection, genetic drift, and mutation bias.
Natural selection
Main article: Natural selection
See also: Dollo's law of irreversibility
Evolution by natural selection is the process by which traits that enhance survival and reproduction become more common in successive generations of a population. It embodies three principles:
Variation exists within populations of organisms with respect to morphology, physiology and behaviour (phenotypic variation).
Different traits confer different rates of survival and reproduction (differential fitness).
These traits can be passed from generation to generation (heritability of fitness).
More offspring are produced than can possibly survive, and these conditions produce competition between organisms for survival and reproduction. Consequently, organisms with traits that give them an advantage over their competitors are more likely to pass on their traits to the next generation than those with traits that do not confer an advantage. This teleonomy is the quality whereby the process of natural selection creates and preserves traits that are seemingly fitted for the functional roles they perform. Consequences of selection include nonrandom mating and genetic hitchhiking.
The central concept of natural selection is the evolutionary fitness of an organism. Fitness is measured by an organism's ability to survive and reproduce, which determines the size of its genetic contribution to the next generation. However, fitness is not the same as the total number of offspring: instead fitness is indicated by the proportion of subsequent generations that carry an organism's genes. For example, if an organism could survive well and reproduce rapidly, but its offspring were all too small and weak to survive, this organism would make little genetic contribution to future generations and would thus have low fitness.
If an allele increases fitness more than the other alleles of that gene, then with each generation this allele has a higher probability of becoming common within the population. These traits are said to be "selected for." Examples of traits that can increase fitness are enhanced survival and increased fecundity. Conversely, the lower fitness caused by having a less beneficial or deleterious allele results in this allele likely becoming rarer—they are "selected against."
Importantly, the fitness of an allele is not a fixed characteristic; if the environment changes, previously neutral or harmful traits may become beneficial and previously beneficial traits become harmful. However, even if the direction of selection does reverse in this way, traits that were lost in the past may not re-evolve in an identical form. However, a re-activation of dormant genes, as long as they have not been eliminated from the genome and were only suppressed perhaps for hundreds of generations, can lead to the re-occurrence of traits thought to be lost like hindlegs in dolphins, teeth in chickens, wings in wingless stick insects, tails and additional nipples in humans etc. "Throwbacks" such as these are known as atavisms.
These charts depict the different types of genetic selection. On each graph, the x-axis variable is the type of phenotypic trait and the y-axis variable is the number of organisms. Group A is the original population and Group B is the population after selection. · Graph 1 shows directional selection, in which a single extreme phenotype is favoured. · Graph 2 depicts stabilizing selection, where the intermediate phenotype is favoured over the extreme traits. · Graph 3 shows disruptive selection, in which the extreme phenotypes are favoured over the intermediate.
Natural selection within a population for a trait that can vary across a range of values, such as height, can be categorised into three different types. The first is directional selection, which is a shift in the average value of a trait over time—for example, organisms slowly getting taller. Secondly, disruptive selection is selection for extreme trait values and often results in two different values becoming most common, with selection against the average value. This would be when either short or tall organisms had an advantage, but not those of medium height. Finally, in stabilising selection there is selection against extreme trait values on both ends, which causes a decrease in variance around the average value and less diversity. This would, for example, cause organisms to eventually have a similar height.
Natural selection most generally makes nature the measure against which individuals and individual traits, are more or less likely to survive. "Nature" in this sense refers to an ecosystem, that is, a system in which organisms interact with every other element, physical as well as biological, in their local environment. Eugene Odum, a founder of ecology, defined an ecosystem as: "Any unit that includes all of the organisms...in a given area interacting with the physical environment so that a flow of energy leads to clearly defined trophic structure, biotic diversity, and material cycles (i.e., exchange of materials between living and nonliving parts) within the system...." Each population within an ecosystem occupies a distinct niche, or position, with distinct relationships to other parts of the system. These relationships involve the life history of the organism, its position in the food chain and its geographic range. This broad understanding of nature enables scientists to delineate specific forces which, together, comprise natural selection.
Natural selection can act at different levels of organisation, such as genes, cells, individual organisms, groups of organisms and species. Selection can act at multiple levels simultaneously. An example of selection occurring below the level of the individual organism are genes called transposons, which can replicate and spread throughout a genome. Selection at a level above the individual, such as group selection, may allow the evolution of cooperation.
Genetic drift
Further information: Genetic drift and Effective population size
Simulation of genetic drift of 20 unlinked alleles in populations of 10 (top) and 100 (bottom). Drift to fixation is more rapid in the smaller population.
Genetic drift is the random fluctuation of allele frequencies within a population from one generation to the next. When selective forces are absent or relatively weak, allele frequencies are equally likely to drift upward or downward in each successive generation because the alleles are subject to sampling error. This drift halts when an allele eventually becomes fixed, either by disappearing from the population or by replacing the other alleles entirely. Genetic drift may therefore eliminate some alleles from a population due to chance alone. Even in the absence of selective forces, genetic drift can cause two separate populations that begin with the same genetic structure to drift apart into two divergent populations with different sets of alleles.
According to the neutral theory of molecular evolution most evolutionary changes are the result of the fixation of neutral mutations by genetic drift. In this model, most genetic changes in a population are thus the result of constant mutation pressure and genetic drift. This form of the neutral theory has been debated since it does not seem to fit some genetic variation seen in nature. A better-supported version of this model is the nearly neutral theory, according to which a mutation that would be effectively neutral in a small population is not necessarily neutral in a large population. Other theories propose that genetic drift is dwarfed by other stochastic forces in evolution, such as genetic hitchhiking, also known as genetic draft. Another concept is constructive neutral evolution (CNE), which explains that complex systems can emerge and spread into a population through neutral transitions due to the principles of excess capacity, presuppression, and ratcheting, and it has been applied in areas ranging from the origins of the spliceosome to the complex interdependence of microbial communities.
The time it takes a neutral allele to become fixed by genetic drift depends on population size; fixation is more rapid in smaller populations. The number of individuals in a population is not critical, but instead a measure known as the effective population size. The effective population is usually smaller than the total population since it takes into account factors such as the level of inbreeding and the stage of the lifecycle in which the population is the smallest. The effective population size may not be the same for every gene in the same population.
It is usually difficult to measure the relative importance of selection and neutral processes, including drift. The comparative importance of adaptive and non-adaptive forces in driving evolutionary change is an area of current research.
Mutation bias
Mutation bias is usually conceived as a difference in expected rates for two different kinds of mutation, e.g., transition-transversion bias, GC-AT bias, deletion-insertion bias. This is related to the idea of developmental bias. Haldane and Fisher argued that, because mutation is a weak pressure easily overcome by selection, tendencies of mutation would be ineffectual except under conditions of neutral evolution or extraordinarily high mutation rates. This opposing-pressures argument was long used to dismiss the possibility of internal tendencies in evolution, until the molecular era prompted renewed interest in neutral evolution.
Noboru Sueoka and Ernst Freese proposed that systematic biases in mutation might be responsible for systematic differences in genomic GC composition between species. The identification of a GC-biased E. coli mutator strain in 1967, along with the proposal of the neutral theory, established the plausibility of mutational explanations for molecular patterns, which are now common in the molecular evolution literature.
For instance, mutation biases are frequently invoked in models of codon usage. Such models also include effects of selection, following the mutation-selection-drift model, which allows both for mutation biases and differential selection based on effects on translation. Hypotheses of mutation bias have played an important role in the development of thinking about the evolution of genome composition, including isochores. Different insertion vs. deletion biases in different taxa can lead to the evolution of different genome sizes. The hypothesis of Lynch regarding genome size relies on mutational biases toward increase or decrease in genome size.
However, mutational hypotheses for the evolution of composition suffered a reduction in scope when it was discovered that (1) GC-biased gene conversion makes an important contribution to composition in diploid organisms such as mammals and (2) bacterial genomes frequently have AT-biased mutation.
Contemporary thinking about the role of mutation biases reflects a different theory from that of Haldane and Fisher. More recent work showed that the original "pressures" theory assumes that evolution is based on standing variation: when evolution depends on events of mutation that introduce new alleles, mutational and developmental biases in the introduction of variation (arrival biases) can impose biases on evolution without requiring neutral evolution or high mutation rates.
Several studies report that the mutations implicated in adaptation reflect common mutation biases though others dispute this interpretation.
Genetic hitchhiking
Further information: Genetic hitchhiking, Hill–Robertson effect, and Selective sweep
Recombination allows alleles on the same strand of DNA to become separated. However, the rate of recombination is low (approximately two events per chromosome per generation). As a result, genes close together on a chromosome may not always be shuffled away from each other and genes that are close together tend to be inherited together, a phenomenon known as linkage. This tendency is measured by finding how often two alleles occur together on a single chromosome compared to expectations, which is called their linkage disequilibrium. A set of alleles that is usually inherited in a group is called a haplotype. This can be important when one allele in a particular haplotype is strongly beneficial: natural selection can drive a selective sweep that will also cause the other alleles in the haplotype to become more common in the population; this effect is called genetic hitchhiking or genetic draft. Genetic draft caused by the fact that some neutral genes are genetically linked to others that are under selection can be partially captured by an appropriate effective population size.
Sexual selection
Further information: Sexual selection
Male moor frogs become blue during the height of mating season. Blue reflectance may be a form of intersexual communication. It is hypothesised that males with brighter blue coloration may signal greater sexual and genetic fitness.
A special case of natural selection is sexual selection, which is selection for any trait that increases mating success by increasing the attractiveness of an organism to potential mates. Traits that evolved through sexual selection are particularly prominent among males of several animal species. Although sexually favoured, traits such as cumbersome antlers, mating calls, large body size and bright colours often attract predation, which compromises the survival of individual males. This survival disadvantage is balanced by higher reproductive success in males that show these hard-to-fake, sexually selected traits.
Natural outcomes
A visual demonstration of rapid antibiotic resistance evolution by E. coli growing across a plate with increasing concentrations of trimethoprim
Evolution influences every aspect of the form and behaviour of organisms. Most prominent are the specific behavioural and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding predators or attracting mates. Organisms can also respond to selection by cooperating with each other, usually by aiding their relatives or engaging in mutually beneficial symbiosis. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed. These outcomes of evolution are distinguished based on time scale as macroevolution versus microevolution. Macroevolution refers to evolution that occurs at or above the level of species, in particular speciation and extinction; whereas microevolution refers to smaller evolutionary changes within a species or population, in particular shifts in allele frequency and adaptation. Macroevolution the outcome of long periods of microevolution. Thus, the distinction between micro- and macroevolution is not a fundamental one—the difference is simply the time involved. However, in macroevolution, the traits of the entire species may be important. For instance, a large amount of variation among individuals allows a species to rapidly adapt to new habitats, lessening the chance of it going extinct, while a wide geographic range increases the chance of speciation, by making it more likely that part of the population will become isolated. In this sense, microevolution and macroevolution might involve selection at different levels—with microevolution acting on genes and organisms, versus macroevolutionary processes such as species selection acting on entire species and affecting their rates of speciation and extinction.
A common misconception is that evolution has goals, long-term plans, or an innate tendency for "progress", as expressed in beliefs such as orthogenesis and evolutionism; realistically however, evolution has no long-term goal and does not necessarily produce greater complexity. Although complex species have evolved, they occur as a side effect of the overall number of organisms increasing and simple forms of life still remain more common in the biosphere. For example, the overwhelming majority of species are microscopic prokaryotes, which form about half the world's biomass despite their small size, and constitute the vast majority of Earth's biodiversity. Simple organisms have therefore been the dominant form of life on Earth throughout its history and continue to be the main form of life up to the present day, with complex life only appearing more diverse because it is more noticeable. Indeed, the evolution of microorganisms is particularly important to evolutionary research, since their rapid reproduction allows the study of experimental evolution and the observation of evolution and adaptation in real time.
Adaptation
Further information: Adaptation
Homologous bones in the limbs of tetrapods. The bones of these animals have the same basic structure, but have been adapted for specific uses.
Adaptation is the process that makes organisms better suited to their habitat. Also, the term adaptation may refer to a trait that is important for an organism's survival. For example, the adaptation of horses' teeth to the grinding of grass. By using the term adaptation for the evolutionary process and adaptive trait for the product (the bodily part or function), the two senses of the word may be distinguished. Adaptations are produced by natural selection. The following definitions are due to Theodosius Dobzhansky:
Adaptation is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats.
Adaptedness is the state of being adapted: the degree to which an organism is able to live and reproduce in a given set of habitats.
An adaptive trait is an aspect of the developmental pattern of the organism which enables or enhances the probability of that organism surviving and reproducing.
Adaptation may cause either the gain of a new feature, or the loss of an ancestral feature. An example that shows both types of change is bacterial adaptation to antibiotic selection, with genetic changes causing antibiotic resistance by both modifying the target of the drug, or increasing the activity of transporters that pump the drug out of the cell. Other striking examples are the bacteria Escherichia coli evolving the ability to use citric acid as a nutrient in a long-term laboratory experiment, Flavobacterium evolving a novel enzyme that allows these bacteria to grow on the by-products of nylon manufacturing, and the soil bacterium Sphingobium evolving an entirely new metabolic pathway that degrades the synthetic pesticide pentachlorophenol. An interesting but still controversial idea is that some adaptations might increase the ability of organisms to generate genetic diversity and adapt by natural selection (increasing organisms' evolvability).
A baleen whale skeleton. Letters a and b label flipper bones, which were adapted from front leg bones, while c indicates vestigial leg bones, both suggesting an adaptation from land to sea.
Adaptation occurs through the gradual modification of existing structures. Consequently, structures with similar internal organisation may have different functions in related organisms. This is the result of a single ancestral structure being adapted to function in different ways. The bones within bat wings, for example, are very similar to those in mice feet and primate hands, due to the descent of all these structures from a common mammalian ancestor. However, since all living organisms are related to some extent, even organs that appear to have little or no structural similarity, such as arthropod, squid and vertebrate eyes, or the limbs and wings of arthropods and vertebrates, can depend on a common set of homologous genes that control their assembly and function; this is called deep homology.
During evolution, some structures may lose their original function and become vestigial structures. Such structures may have little or no function in a current species, yet have a clear function in ancestral species, or other closely related species. Examples include pseudogenes, the non-functional remains of eyes in blind cave-dwelling fish, wings in flightless birds, the presence of hip bones in whales and snakes, and sexual traits in organisms that reproduce via asexual reproduction. Examples of vestigial structures in humans include wisdom teeth, the coccyx, the vermiform appendix, and other behavioural vestiges such as goose bumps and primitive reflexes.
However, many traits that appear to be simple adaptations are in fact exaptations: structures originally adapted for one function, but which coincidentally became somewhat useful for some other function in the process. One example is the African lizard Holaspis guentheri, which developed an extremely flat head for hiding in crevices, as can be seen by looking at its near relatives. However, in this species, the head has become so flattened that it assists in gliding from tree to tree—an exaptation. Within cells, molecular machines such as the bacterial flagella and protein sorting machinery evolved by the recruitment of several pre-existing proteins that previously had different functions. Another example is the recruitment of enzymes from glycolysis and xenobiotic metabolism to serve as structural proteins called crystallins within the lenses of organisms' eyes.
An area of current investigation in evolutionary developmental biology is the developmental basis of adaptations and exaptations. This research addresses the origin and evolution of embryonic development and how modifications of development and developmental processes produce novel features. These studies have shown that evolution can alter development to produce new structures, such as embryonic bone structures that develop into the jaw in other animals instead forming part of the middle ear in mammals. It is also possible for structures that have been lost in evolution to reappear due to changes in developmental genes, such as a mutation in chickens causing embryos to grow teeth similar to those of crocodiles. It is now becoming clear that most alterations in the form of organisms are due to changes in a small set of conserved genes.
Coevolution
Further information: Coevolution
The common garter snake has evolved resistance to the defensive substance tetrodotoxin in its amphibian prey.
Interactions between organisms can produce both conflict and cooperation. When the interaction is between pairs of species, such as a pathogen and a host, or a predator and its prey, these species can develop matched sets of adaptations. Here, the evolution of one species causes adaptations in a second species. These changes in the second species then, in turn, cause new adaptations in the first species. This cycle of selection and response is called coevolution. An example is the production of tetrodotoxin in the rough-skinned newt and the evolution of tetrodotoxin resistance in its predator, the common garter snake. In this predator-prey pair, an evolutionary arms race has produced high levels of toxin in the newt and correspondingly high levels of toxin resistance in the snake.
Cooperation
Further information: Co-operation (evolution)
Not all co-evolved interactions between species involve conflict. Many cases of mutually beneficial interactions have evolved. For instance, an extreme cooperation exists between plants and the mycorrhizal fungi that grow on their roots and aid the plant in absorbing nutrients from the soil. This is a reciprocal relationship as the plants provide the fungi with sugars from photosynthesis. Here, the fungi actually grow inside plant cells, allowing them to exchange nutrients with their hosts, while sending signals that suppress the plant immune system.
Coalitions between organisms of the same species have also evolved. An extreme case is the eusociality found in social insects, such as bees, termites and ants, where sterile insects feed and guard the small number of organisms in a colony that are able to reproduce. On an even smaller scale, the somatic cells that make up the body of an animal limit their reproduction so they can maintain a stable organism, which then supports a small number of the animal's germ cells to produce offspring. Here, somatic cells respond to specific signals that instruct them whether to grow, remain as they are, or die. If cells ignore these signals and multiply inappropriately, their uncontrolled growth causes cancer.
Such cooperation within species may have evolved through the process of kin selection, which is where one organism acts to help raise a relative's offspring. This activity is selected for because if the helping individual contains alleles which promote the helping activity, it is likely that its kin will also contain these alleles and thus those alleles will be passed on. Other processes that may promote cooperation include group selection, where cooperation provides benefits to a group of organisms.
Speciation
Main article: Speciation
Further information: Assortative mating and Panmixia
The four geographic modes of speciation
Speciation is the process where a species diverges into two or more descendant species.
There are multiple ways to define the concept of "species." The choice of definition is dependent on the particularities of the species concerned. For example, some species concepts apply more readily toward sexually reproducing organisms while others lend themselves better toward asexual organisms. Despite the diversity of various species concepts, these various concepts can be placed into one of three broad philosophical approaches: interbreeding, ecological and phylogenetic. The Biological Species Concept (BSC) is a classic example of the interbreeding approach. Defined by evolutionary biologist Ernst Mayr in 1942, the BSC states that "species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups." Despite its wide and long-term use, the BSC like other species concepts is not without controversy, for example, because genetic recombination among prokaryotes is not an intrinsic aspect of reproduction; this is called the species problem. Some researchers have attempted a unifying monistic definition of species, while others adopt a pluralistic approach and suggest that there may be different ways to logically interpret the definition of a species.
Barriers to reproduction between two diverging sexual populations are required for the populations to become new species. Gene flow may slow this process by spreading the new genetic variants also to the other populations. Depending on how far two species have diverged since their most recent common ancestor, it may still be possible for them to produce offspring, as with horses and donkeys mating to produce mules. Such hybrids are generally infertile. In this case, closely related species may regularly interbreed, but hybrids will be selected against and the species will remain distinct. However, viable hybrids are occasionally formed and these new species can either have properties intermediate between their parent species, or possess a totally new phenotype. The importance of hybridisation in producing new species of animals is unclear, although cases have been seen in many types of animals, with the gray tree frog being a particularly well-studied example.
Speciation has been observed multiple times under both controlled laboratory conditions and in nature. In sexually reproducing organisms, speciation results from reproductive isolation followed by genealogical divergence. There are four primary geographic modes of speciation. The most common in animals is allopatric speciation, which occurs in populations initially isolated geographically, such as by habitat fragmentation or migration. Selection under these conditions can produce very rapid changes in the appearance and behaviour of organisms. As selection and drift act independently on populations isolated from the rest of their species, separation may eventually produce organisms that cannot interbreed.
The second mode of speciation is peripatric speciation, which occurs when small populations of organisms become isolated in a new environment. This differs from allopatric speciation in that the isolated populations are numerically much smaller than the parental population. Here, the founder effect causes rapid speciation after an increase in inbreeding increases selection on homozygotes, leading to rapid genetic change.
The third mode is parapatric speciation. This is similar to peripatric speciation in that a small population enters a new habitat, but differs in that there is no physical separation between these two populations. Instead, speciation results from the evolution of mechanisms that reduce gene flow between the two populations. Generally this occurs when there has been a drastic change in the environment within the parental species' habitat. One example is the grass Anthoxanthum odoratum, which can undergo parapatric speciation in response to localised metal pollution from mines. Here, plants evolve that have resistance to high levels of metals in the soil. Selection against interbreeding with the metal-sensitive parental population produced a gradual change in the flowering time of the metal-resistant plants, which eventually produced complete reproductive isolation. Selection against hybrids between the two populations may cause reinforcement, which is the evolution of traits that promote mating within a species, as well as character displacement, which is when two species become more distinct in appearance.
Geographical isolation of finches on the Galápagos Islands produced over a dozen new species.
Finally, in sympatric speciation species diverge without geographic isolation or changes in habitat. This form is rare since even a small amount of gene flow may remove genetic differences between parts of a population. Generally, sympatric speciation in animals requires the evolution of both genetic differences and nonrandom mating, to allow reproductive isolation to evolve.
One type of sympatric speciation involves crossbreeding of two related species to produce a new hybrid species. This is not common in animals as animal hybrids are usually sterile. This is because during meiosis the homologous chromosomes from each parent are from different species and cannot successfully pair. However, it is more common in plants because plants often double their number of chromosomes, to form polyploids. This allows the chromosomes from each parental species to form matching pairs during meiosis, since each parent's chromosomes are represented by a pair already. An example of such a speciation event is when the plant species Arabidopsis thaliana and Arabidopsis arenosa crossbred to give the new species Arabidopsis suecica. This happened about 20,000 years ago, and the speciation process has been repeated in the laboratory, which allows the study of the genetic mechanisms involved in this process. Indeed, chromosome doubling within a species may be a common cause of reproductive isolation, as half the doubled chromosomes will be unmatched when breeding with undoubled organisms.
Speciation events are important in the theory of punctuated equilibrium, which accounts for the pattern in the fossil record of short "bursts" of evolution interspersed with relatively long periods of stasis, where species remain relatively unchanged. In this theory, speciation and rapid evolution are linked, with natural selection and genetic drift acting most strongly on organisms undergoing speciation in novel habitats or small populations. As a result, the periods of stasis in the fossil record correspond to the parental population and the organisms undergoing speciation and rapid evolution are found in small populations or geographically restricted habitats and therefore rarely being preserved as fossils.
Extinction
Further information: Extinction
Tyrannosaurus rex. Non-avian dinosaurs died out in the Cretaceous–Paleogene extinction event at the end of the Cretaceous period.
Extinction is the disappearance of an entire species. Extinction is not an unusual event, as species regularly appear through speciation and disappear through extinction. Nearly all animal and plant species that have lived on Earth are now extinct, and extinction appears to be the ultimate fate of all species. These extinctions have happened continuously throughout the history of life, although the rate of extinction spikes in occasional mass extinction events. The Cretaceous–Paleogene extinction event, during which the non-avian dinosaurs became extinct, is the most well-known, but the earlier Permian–Triassic extinction event was even more severe, with approximately 96% of all marine species driven to extinction. The Holocene extinction event is an ongoing mass extinction associated with humanity's expansion across the globe over the past few thousand years. Present-day extinction rates are 100–1000 times greater than the background rate and up to 30% of current species may be extinct by the mid 21st century. Human activities are now the primary cause of the ongoing extinction event; global warming may further accelerate it in the future. Despite the estimated extinction of more than 99% of all species that ever lived on Earth, about 1 trillion species are estimated to be on Earth currently with only one-thousandth of 1% described.
The role of extinction in evolution is not very well understood and may depend on which type of extinction is considered. The causes of the continuous "low-level" extinction events, which form the majority of extinctions, may be the result of competition between species for limited resources (the competitive exclusion principle). If one species can out-compete another, this could produce species selection, with the fitter species surviving and the other species being driven to extinction. The intermittent mass extinctions are also important, but instead of acting as a selective force, they drastically reduce diversity in a nonspecific manner and promote bursts of rapid evolution and speciation in survivors.
Applications
Main articles: Applications of evolution, Selective breeding, and Evolutionary computation
Concepts and models used in evolutionary biology, such as natural selection, have many applications.
Artificial selection is the intentional selection of traits in a population of organisms. This has been used for thousands of years in the domestication of plants and animals. More recently, such selection has become a vital part of genetic engineering, with selectable markers such as antibiotic resistance genes being used to manipulate DNA. Proteins with valuable properties have evolved by repeated rounds of mutation and selection (for example modified enzymes and new antibodies) in a process called directed evolution.
Understanding the changes that have occurred during an organism's evolution can reveal the genes needed to construct parts of the body, genes which may be involved in human genetic disorders. For example, the Mexican tetra is an albino cavefish that lost its eyesight during evolution. Breeding together different populations of this blind fish produced some offspring with functional eyes, since different mutations had occurred in the isolated populations that had evolved in different caves. This helped identify genes required for vision and pigmentation.
Evolutionary theory has many applications in medicine. Many human diseases are not static phenomena, but capable of evolution. Viruses, bacteria, fungi and cancers evolve to be resistant to host immune defences, as well as to pharmaceutical drugs. These same problems occur in agriculture with pesticide and herbicide resistance. It is possible that we are facing the end of the effective life of most of available antibiotics and predicting the evolution and evolvability of our pathogens and devising strategies to slow or circumvent it is requiring deeper knowledge of the complex forces driving evolution at the molecular level.
In computer science, simulations of evolution using evolutionary algorithms and artificial life started in the 1960s and were extended with simulation of artificial selection. Artificial evolution became a widely recognised optimisation method as a result of the work of Ingo Rechenberg in the 1960s. He used evolution strategies to solve complex engineering problems. Genetic algorithms in particular became popular through the writing of John Henry Holland. Practical applications also include automatic evolution of computer programmes. Evolutionary algorithms are now used to solve multi-dimensional problems more efficiently than software produced by human designers and also to optimise the design of systems.
Evolutionary history of life
Life timelineThis box: viewtalkedit−4500 —–—–−4000 —–—–−3500 —–—–−3000 —–—–−2500 —–—–−2000 —–—–−1500 —–—–−1000 —–—–−500 —–—–0 — Water Single-celled life Photosynthesis Eukaryotes Multicellular life Plants Arthropods MolluscsFlowersDinosaurs MammalsBirdsPrimatesHadeanArcheanProterozoicPhanerozoic ←Earth formed←Earliest water←LUCA←Earliest fossils←LHB meteorites←Earliest oxygen←Pongola glaciation*←Atmospheric oxygen←Huronian glaciation*←Sexual reproduction←Earliest multicellular life←Earliest fungi←Earliest plants←Earliest animals←Cryogenian ice age*←Ediacaran biota←Cambrian explosion←Andean glaciation*←Earliest tetrapods←Karoo ice age*←Earliest apes / humans←Quaternary ice age*(million years ago)*Ice Ages
Main article: Evolutionary history of life
See also: Timeline of the evolutionary history of life
Origin of life
Further information: Abiogenesis, Earliest known life forms, Panspermia, and RNA world hypothesis
The Earth is about 4.54 billion years old. The earliest undisputed evidence of life on Earth dates from at least 3.5 billion years ago, during the Eoarchean Era after a geological crust started to solidify following the earlier molten Hadean Eon. Microbial mat fossils have been found in 3.48 billion-year-old sandstone in Western Australia. Other early physical evidence of a biogenic substance is graphite in 3.7 billion-year-old metasedimentary rocks discovered in Western Greenland as well as "remains of biotic life" found in 4.1 billion-year-old rocks in Western Australia. Commenting on the Australian findings, Stephen Blair Hedges wrote: "If life arose relatively quickly on Earth, then it could be common in the universe." In July 2016, scientists reported identifying a set of 355 genes from the last universal common ancestor (LUCA) of all organisms living on Earth.
More than 99% of all species, amounting to over five billion species, that ever lived on Earth are estimated to be extinct. Estimates on the number of Earth's current species range from 10 million to 14 million, of which about 1.9 million are estimated to have been named and 1.6 million documented in a central database to date, leaving at least 80% not yet described.
Highly energetic chemistry is thought to have produced a self-replicating molecule around 4 billion years ago, and half a billion years later the last common ancestor of all life existed. The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions. The beginning of life may have included self-replicating molecules such as RNA and the assembly of simple cells.
Common descent
Further information: Common descent and Evidence of common descent
All organisms on Earth are descended from a common ancestor or ancestral gene pool. Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events. The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits. Fourth, organisms can be classified using these similarities into a hierarchy of nested groups, similar to a family tree.
The hominoids are descendants of a common ancestor.
Due to horizontal gene transfer, this "tree of life" may be more complicated than a simple branching tree, since some genes have spread independently between distantly related species. To solve this problem and others, some authors prefer to use the "Coral of life" as a metaphor or a mathematical model to illustrate the evolution of life. This view dates back to an idea briefly mentioned by Darwin but later abandoned.
Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record. By comparing the anatomies of both modern and extinct species, palaeontologists can infer the lineages of those species. However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.
More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and amino acids. The development of molecular genetics has revealed the record of evolution left in organisms' genomes: dating when species diverged through the molecular clock produced by mutations. For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 98% of their genomes and analysing the few areas where they differ helps shed light on when the common ancestor of these species existed.
Evolution of life
Main articles: Evolutionary history of life and Timeline of evolutionary history of life
Evolutionary tree showing the divergence of modern species from their common ancestor in the centre. The three domains are coloured, with bacteria blue, archaea green and eukaryotes red.
Prokaryotes inhabited the Earth from approximately 3–4 billion years ago. No obvious changes in morphology or cellular organisation occurred in these organisms over the next few billion years. The eukaryotic cells emerged between 1.6 and 2.7 billion years ago. The next major change in cell structure came when bacteria were engulfed by eukaryotic cells, in a cooperative association called endosymbiosis. The engulfed bacteria and the host cell then underwent coevolution, with the bacteria evolving into either mitochondria or hydrogenosomes. Another engulfment of cyanobacterial-like organisms led to the formation of chloroplasts in algae and plants.
The history of life was that of the unicellular eukaryotes, prokaryotes and archaea until about 610 million years ago when multicellular organisms began to appear in the oceans in the Ediacaran period. The evolution of multicellularity occurred in multiple independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime moulds and myxobacteria. In January 2016, scientists reported that, about 800 million years ago, a minor genetic change in a single molecule called GK-PID may have allowed organisms to go from a single cell organism to one of many cells.
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over approximately 10 million years, in an event called the Cambrian explosion. Here, the majority of types of modern animals appeared in the fossil record, as well as unique lineages that subsequently became extinct. Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.
About 500 million years ago, plants and fungi colonised the land and were soon followed by arthropods and other animals. Insects were particularly successful and even today make up the majority of animal species. Amphibians first appeared around 364 million years ago, followed by early amniotes and birds around 155 million years ago (both from "reptile"-like lineages), mammals around 129 million years ago, Homininae around 10 million years ago and modern humans around 250,000 years ago. However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to be highly successful and dominate the Earth, with the majority of both biomass and species being prokaryotes.
History of evolutionary thought
Main article: History of evolutionary thought
Further information: History of speciation
Lucretius
Alfred Russel Wallace
Thomas Robert Malthus
In 1842, Charles Darwin penned his first sketch of On the Origin of Species.
Classical antiquity
The proposal that one type of organism could descend from another type goes back to some of the first pre-Socratic Greek philosophers, such as Anaximander and Empedocles. Such proposals survived into Roman times. The poet and philosopher Lucretius followed Empedocles in his masterwork De rerum natura (lit. 'On the Nature of Things').
Middle Ages
In contrast to these materialistic views, Aristotelianism had considered all natural things as actualisations of fixed natural possibilities, known as forms. This became part of a medieval teleological understanding of nature in which all things have an intended role to play in a divine cosmic order. Variations of this idea became the standard understanding of the Middle Ages and were integrated into Christian learning, but Aristotle did not demand that real types of organisms always correspond one-for-one with exact metaphysical forms and specifically gave examples of how new types of living things could come to be.
A number of Arab Muslim scholars wrote about evolution, most notably Ibn Khaldun, who wrote the book Muqaddimah in 1377 AD, in which he asserted that humans developed from "the world of the monkeys", in a process by which "species become more numerous".
Pre-Darwinian
The "New Science" of the 17th century rejected the Aristotelian approach. It sought to explain natural phenomena in terms of physical laws that were the same for all visible things and that did not require the existence of any fixed natural categories or divine cosmic order. However, this new approach was slow to take root in the biological sciences: the last bastion of the concept of fixed natural types. John Ray applied one of the previously more general terms for fixed natural types, "species", to plant and animal types, but he strictly identified each type of living thing as a species and proposed that each species could be defined by the features that perpetuated themselves generation after generation. The biological classification introduced by Carl Linnaeus in 1735 explicitly recognised the hierarchical nature of species relationships, but still viewed species as fixed according to a divine plan.
Other naturalists of this time speculated on the evolutionary change of species over time according to natural laws. In 1751, Pierre Louis Maupertuis wrote of natural modifications occurring during reproduction and accumulating over many generations to produce new species. Georges-Louis Leclerc, Comte de Buffon, suggested that species could degenerate into different organisms, and Erasmus Darwin proposed that all warm-blooded animals could have descended from a single microorganism (or "filament"). The first full-fledged evolutionary scheme was Jean-Baptiste Lamarck's "transmutation" theory of 1809, which envisaged spontaneous generation continually producing simple forms of life that developed greater complexity in parallel lineages with an inherent progressive tendency, and postulated that on a local level, these lineages adapted to the environment by inheriting changes caused by their use or disuse in parents. (The latter process was later called Lamarckism.) These ideas were condemned by established naturalists as speculation lacking empirical support. In particular, Georges Cuvier insisted that species were unrelated and fixed, their similarities reflecting divine design for functional needs. In the meantime, Ray's ideas of benevolent design had been developed by William Paley into the Natural Theology or Evidences of the Existence and Attributes of the Deity (1802), which proposed complex adaptations as evidence of divine design and which was admired by Charles Darwin.
Darwinian revolution
The crucial break from the concept of constant typological classes or types in biology came with the theory of evolution through natural selection, which was formulated by Charles Darwin and Alfred Wallace in terms of variable populations. Darwin used the expression "descent with modification" rather than "evolution". Partly influenced by An Essay on the Principle of Population (1798) by Thomas Robert Malthus, Darwin noted that population growth would lead to a "struggle for existence" in which favourable variations prevailed as others perished. In each generation, many offspring fail to survive to an age of reproduction because of limited resources. This could explain the diversity of plants and animals from a common ancestry through the working of natural laws in the same way for all types of organism. Darwin developed his theory of "natural selection" from 1838 onwards and was writing up his "big book" on the subject when Alfred Russel Wallace sent him a version of virtually the same theory in 1858. Their separate papers were presented together at an 1858 meeting of the Linnean Society of London. At the end of 1859, Darwin's publication of his "abstract" as On the Origin of Species explained natural selection in detail and in a way that led to an increasingly wide acceptance of Darwin's concepts of evolution at the expense of alternative theories. Thomas Henry Huxley applied Darwin's ideas to humans, using paleontology and comparative anatomy to provide strong evidence that humans and apes shared a common ancestry. Some were disturbed by this since it implied that humans did not have a special place in the universe.
Pangenesis and heredity
The mechanisms of reproductive heritability and the origin of new traits remained a mystery. Towards this end, Darwin developed his provisional theory of pangenesis. In 1865, Gregor Mendel reported that traits were inherited in a predictable manner through the independent assortment and segregation of elements (later known as genes). Mendel's laws of inheritance eventually supplanted most of Darwin's pangenesis theory. August Weismann made the important distinction between germ cells that give rise to gametes (such as sperm and egg cells) and the somatic cells of the body, demonstrating that heredity passes through the germ line only. Hugo de Vries connected Darwin's pangenesis theory to Weismann's germ/soma cell distinction and proposed that Darwin's pangenes were concentrated in the cell nucleus and when expressed they could move into the cytoplasm to change the cell's structure. De Vries was also one of the researchers who made Mendel's work well known, believing that Mendelian traits corresponded to the transfer of heritable variations along the germline. To explain how new variants originate, de Vries developed a mutation theory that led to a temporary rift between those who accepted Darwinian evolution and biometricians who allied with de Vries. In the 1930s, pioneers in the field of population genetics, such as Ronald Fisher, Sewall Wright and J. B. S. Haldane set the foundations of evolution onto a robust statistical philosophy. The false contradiction between Darwin's theory, genetic mutations, and Mendelian inheritance was thus reconciled.
The 'modern synthesis'
Main article: Modern synthesis (20th century)
In the 1920s and 1930s, the modern synthesis connected natural selection and population genetics, based on Mendelian inheritance, into a unified theory that included random genetic drift, mutation, and gene flow. This new version of evolutionary theory focused on changes in allele frequencies in population. It explained patterns observed across species in populations, through fossil transitions in palaeontology.
Further syntheses
Since then, further syntheses have extended evolution's explanatory power in the light of numerous discoveries, to cover biological phenomena across the whole of the biological hierarchy from genes to populations.
The publication of the structure of DNA by James Watson and Francis Crick with contribution of Rosalind Franklin in 1953 demonstrated a physical mechanism for inheritance. Molecular biology improved understanding of the relationship between genotype and phenotype. Advances were also made in phylogenetic systematics, mapping the transition of traits into a comparative and testable framework through the publication and use of evolutionary trees. In 1973, evolutionary biologist Theodosius Dobzhansky penned that "nothing in biology makes sense except in the light of evolution", because it has brought to light the relations of what first seemed disjointed facts in natural history into a coherent explanatory body of knowledge that describes and predicts many observable facts about life on this planet.
One extension, known as evolutionary developmental biology and informally called "evo-devo," emphasises how changes between generations (evolution) act on patterns of change within individual organisms (development). Since the beginning of the 21st century, some biologists have argued for an extended evolutionary synthesis, which would account for the effects of non-genetic inheritance modes, such as epigenetics, parental effects, ecological inheritance and cultural inheritance, and evolvability.
Social and cultural responses
Further information: Social effects of evolutionary theory, 1860 Oxford evolution debate, Rejection of evolution by religious groups, Objections to evolution, and Evolution in fiction
As evolution became widely accepted in the 1870s, caricatures of Charles Darwin with an ape or monkey body symbolised evolution.
In the 19th century, particularly after the publication of On the Origin of Species in 1859, the idea that life had evolved was an active source of academic debate centred on the philosophical, social and religious implications of evolution. Today, the modern evolutionary synthesis is accepted by a vast majority of scientists. However, evolution remains a contentious concept for some theists.
While various religions and denominations have reconciled their beliefs with evolution through concepts such as theistic evolution, there are creationists who believe that evolution is contradicted by the creation myths found in their religions and who raise various objections to evolution. As had been demonstrated by responses to the publication of Vestiges of the Natural History of Creation in 1844, the most controversial aspect of evolutionary biology is the implication of human evolution that humans share common ancestry with apes and that the mental and moral faculties of humanity have the same types of natural causes as other inherited traits in animals. In some countries, notably the United States, these tensions between science and religion have fuelled the current creation–evolution controversy, a religious conflict focusing on politics and public education. While other scientific fields such as cosmology and Earth science also conflict with literal interpretations of many religious texts, evolutionary biology experiences significantly more opposition from religious literalists.
The teaching of evolution in American secondary school biology classes was uncommon in most of the first half of the 20th century. The Scopes Trial decision of 1925 caused the subject to become very rare in American secondary biology textbooks for a generation, but it was gradually re-introduced later and became legally protected with the 1968 Epperson v. Arkansas decision. Since then, the competing religious belief of creationism was legally disallowed in secondary school curricula in various decisions in the 1970s and 1980s, but it returned in pseudoscientific form as intelligent design (ID), to be excluded once again in the 2005 Kitzmiller v. Dover Area School District case. The debate over Darwin's ideas did not generate significant controversy in China.
See also
Devolution (biology) – Notion that species can revert to primitive forms | biology | 998 | https://da.wikipedia.org/wiki/Art | Art | Arten (species, forkortet sp., flertal: spp.) er den grundlæggende systematiske enhed inden for biologien. Arten defineres ofte som en naturlig gruppe af populationer, hvor udveksling af gener finder sted (eller kan finde sted) og som i forhold til forplantning er isoleret fra andre grupper. Det vil sige at kun individer inden for samme art kan parre sig og få forplantningsdygtigt afkom. Dette kaldes det biologiske artsbegreb. For organismer, der formerer sig ukønnet eller ved selvbestøvning, må arter afgrænses ud fra ligheder og forskelle mellem forskellige individer. Nogle dyrearter kan i fangenskab hybridisere og få fertilt afkom, men da dette ikke vil ske i naturen, selv om de mødes her, betragtes de som forskellige arter.
Eksempel
To heste kan parre sig og få et føl, der igen kan få føl med andre heste – hestene tilhører derfor samme art. En hest og et æsel kan også parre sig og deres unger kaldes enten muldyr eller mulæsel, afhængig af hvem der er moren, men muldyret eller mulæselet kan (normalt) ikke få unger, da de oftest er sterile. Af den grund regnes hest og æsel som to forskellige arter. Det samme princip gælder også for planterne. Denne naturskabte afgrænsning mellem to arter kaldes en artsbarriere. Den kan af og til gennembrydes, når ellers sterile krydsninger spontant eller kunstigt får gennemført en kromosomfordobling. Se f.eks. Vadegræs (Spartina pectinata).
Arter over for hybrider
Man kan dog godt komme ud for, at arter kan krydses og får blandet afkom, men hybriden vil kun kunne bestå på steder, hvor ingen af forældrearterne kan klare sig. Dette er et særligt udpræget problem med Rododendron (Rhododendron) og Tjørn (Crataegus), fordi disse slægter breder sig voldsomt efter skovbrand eller stormfald. Da hybriderne bliver frugtbare i en yngre alder end arterne, kan de dominere i en periode, men når skoven lukker sig, så fortrænges hybriderne og kun de specialiserede arter kan overleve i skovens dybe skygge eller ude i lyset i sumpe, på ur og i kalksten, m.m.
Flere artsbegreber
Fordi det biologiske artsbegreb kan være besværligt at anvende i praksis, er der efterhånden skabt en række andre artsbegreber:
Morfologisk artsbegreb Arterne adskiller sig fra hinanden ved deres bygning. Dette begreb er blevet meget anvendt gennem tiden.
Økologisk artsbegreb Definerer en art som en gruppe af organismer, der udfylder samme niche. Krydsninger mellem to nærtstående arter vil ikke være optimalt tilpasset til forældrearternes nicher og vil ikke klare sig i konkurrencen.
Evolutionære artsbegreb Også kaldet det kladistiske eller fylogenetiske artsbegreb. Naturen er dynamisk, ikke statisk - alle arter ændrer sig med tiden og bliver, hvis de ikke uddør som følge af konkurrence, naturkatastrofer m.v., til én eller flere nye arter. Det evolutionære artsbegreb minder om det biologiske, men inddrager tidsdimensionen, det vil sige at en art udvikler sig over tid og at nye arter opstår ved artsdannelse. Individer der fylogenetisk har samme stamfader tilhører samme art.
Pluralistisk artsbegreb En art er et samfund af populationer, der formerer sig og lever inden for en bestemt niche i naturen.
Se også
Systematik
Evolutionsteori
Kilder
Lars Skipper: Hvad er en art? Citat: "...Arten er den eneste [klassifikations-kategori] der eksisterer i virkeligheden, alle andre (slægter, familier, ordener m.v.) er indført for overskuelighedens skyld..."
Eksterne henvisninger
2003-12-31, ScienceDaily: Working On The 'Porsche Of Its Time': New Model For Species Determination Offered Citat: "...two species of dinosaur that are members of the same genera varied from each other by just 2.2 percent. Translation of the percentage into an actual number results in an average of just three skeletal differences out of the total 338 bones in the body. Amazingly, 58 percent of these differences occurred in the skull alone. "This is a lot less variation than I'd expected," said Novak..."
2003-08-08, ScienceDaily: Cross-species Mating May Be Evolutionarily Important And Lead To Rapid Change, Say Indiana University Researchers Citat: "...the sudden mixing of closely related species may occasionally provide the energy to impel rapid evolutionary change..."
2004-01-09 ScienceDaily: Mayo Researchers Observe Genetic Fusion Of Human, Animal Cells; May Help Explain Origin Of AIDS Citat: "...The researchers have discovered conditions in which pig cells and human cells can fuse together in the body to yield hybrid cells that contain genetic material from both species..."What we found was completely unexpected," says Jeffrey Platt, M.D..."
2000-09-18, ScienceDaily: Scientists Unravel Ancient Evolutionary History Of Photosynthesis Citat: "...gene-swapping was common among ancient bacteria early in evolution..."
2004-06-07, Sciencedaily: Parting Genomes: University Of Arizona Biologists Discover Seeds Of Speciation Citat: "...There's a huge amount of biodiversity out there, and we don't know where it comes from. Evolutionary biologists are excited to figure out what causes what we see out there--the relative forces of selection and drift--whether things are adapting to their environment or variation is random..."
2005-07-05, Sciencedaily: Trees, Vines And Nets -- Microbial Evolution Changes Its Face Citat: "... EBI researchers have changed our view of 4 billion years of microbial evolution...In all, more than 600,000 vertical transfers are observed, coupled with 90,000 gene loss events and approximately 40,000 horizontal gene transfers...A few species, including beneficial nitrogen-fixing soil bacteria, appear to be 'champions'of horizontal gene transfer; "it's entirely possible that apparently harmless organisms are quietly spreading antibiotic resistance under our feet," concludes Christos Ouzounis..."
2005-11-11, Sciencedaily: Lateral Thinking Produces First Map Of Gene Transmission Citat: "...Their results clearly show genetic modification of organisms by lateral transfer is a widespread natural phenomenon, and it can occur even between distantly related organisms... it was assumed that transfer of genes could only be vertical, i.e. from parents to offspring..."
Økologi
Biologi | danish | 0.791178 |
evolution_not_make_our_life_longer/Disposable_soma_theory_of_aging.txt | The disposable soma theory of aging states that organisms age due to an evolutionary trade-off between growth, reproduction, and DNA repair maintenance. Formulated by Thomas Kirkwood, the disposable soma theory explains that an organism only has a limited amount of resources that it can allocate to its various cellular processes. Therefore, a greater investment in growth and reproduction would result in reduced investment in DNA repair maintenance, leading to increased cellular damage, shortened telomeres, accumulation of mutations, compromised stem cells, and ultimately, senescence. Although many models, both animal and human, have appeared to support this theory, parts of it are still controversial.
Specifically, while the evolutionary trade-off between growth and aging has been well established,
the relationship between reproduction and aging is still without scientific consensus, and the cellular mechanisms largely undiscovered.
Background and history[edit]
British biologist Thomas Kirkwood first proposed the disposable soma theory of aging in a 1977 Nature review article. The theory was inspired by Leslie Orgel's Error Catastrophe Theory of Aging, which was published fourteen years earlier, in 1963. Orgel believed that the process of aging arose due to mutations acquired during the replication process, and Kirkwood developed the disposable soma theory in order to mediate Orgel's work with evolutionary genetics.
Principles[edit]
The disposable soma theory of aging posits that there is a trade-off in resource allocation between somatic maintenance and reproductive investment. Too low an investment in self-repair would be evolutionarily unsound, as the organism would likely die before reproductive age. However, too high an investment in self-repair would also be evolutionarily unsound due to the fact that one's offspring would likely die before reproductive age. Therefore, there is a compromise and resources are partitioned accordingly. However, this compromise is thought to damage somatic repair systems, which can lead to progressive cellular damage and senescence. Repair costs can be categorized into three groups: (1) the costs of increased durability of nonrenewable parts; (2) the costs of maintenance involving cell renewal, and (3) the costs of intracellular maintenance. In a nutshell, aging and decline is essentially a trade-off for increased reproductive robustness in youth.
Mechanisms[edit]
The IGF-1 pathway, which represses FOXO, thus preventing gene expression of longevity-inducing proteins
Growth and somatic maintenance[edit]
Main article: Mechanistic target of rapamycin § Aging
Much research has been done on the antagonistic effects of increased growth on lifespan. Specifically, the hormone insulin-like growth factor 1 (IGF-1), binds to a cell receptor, leading to a phosphorylation cascade. This cascade results in kinases phosphorylating forkhead transcription factor (FOXO), deactivating it. Deactivation of FOXO results in an inability to express genes involved in responding to oxidative stress response, such as antioxidants, chaperones, and heat-shock proteins.
Additionally, uptake of IGF-1 stimulates the mTOR pathway, which activates protein synthesis (and therefore growth) through upregulation of the translation-promoting S6K1, and also inhibits autophagy, a process necessary for recycling of damaged cellular products. Decline of autophagy causes neurodegeneration, protein aggregation and premature aging. Lastly, studies have also indicated that the mTOR pathway also alters immune responses and stimulates cyclin-dependent kinase (CDK) inhibitors such as p16 and p21. This leads to alteration of the stem-cell niche and results in stem cell exhaustion, another theorized mechanism of aging.
Reproduction and somatic maintenance[edit]
While reproduction inhibits lifespan with regard to multicellular organisms, the precise mechanism responsible for this effect remains unclear. Although many models do illustrate an inverse relationship, and the theory makes sense from an evolutionary perspective, the cellular mechanisms have yet to be explored. However, with regards to cellular replication, the progressive shortening of telomeres is a mechanism which limits the amount of generations of a single cell may undergo. Furthermore, in unicellular organisms like Saccharomyces cerevisiae, the formation of extrachromosomal rDNA circles (ERCs) in mother cells (but not daughter cells) upon every subsequent division is an identifiable type of DNA damage that is associated with replication. These ERCs accumulate over time and eventually trigger replicative senescence and death of the mother cell.
Evidence[edit]
Growth and aging[edit]
There is a large body of evidence indicating the negative effects of growth on longevity across many species. As a general rule, individuals of a smaller size generally live longer than larger individuals of the same species.
Animal models[edit]
In dwarf models of mice, such Snell or Ames mice, mutations have arisen, either rendering them incapable of producing IGF-1 or unable to have adequate receptors for IGF-1 uptake. Furthermore, mice injected with growth hormone have been shown to have progressive weight loss, roughing of the coat, curvature of the spine, enlargement of the organs, kidney lesions and increased cancer risk. This effect is also seen in different breeds of dogs, where smaller breeds of dogs typically live significantly longer compared to their larger counterparts. Selectively bred for their small size, smaller dog breeds like the Chihuahua (average lifespan of 15–20 years) have the B/B genotype for the IGF-1 haplotype, reducing the amount of IGF-1 produced. Conversely, large dogs like the Great Dane (average lifespan of 6–8 years) are homozygous for the IGF-1 I allele, which increases the amount of IGF-1 production.
Human models[edit]
Initially, it was believed that growth hormone actually prolonged lifespan due to a 1990 study that indicated that injection of growth hormone to men over 60 years of age appeared to reverse various biomarkers implicated in aging, such as decreased muscle mass, bone density, skin thickness, and increased adipose tissue. However, a 1999 study found that administering growth hormone also significantly increased mortality rate. Recent genomic studies have confirmed that the genes involved in growth hormone uptake and signaling are largely conserved across a plethora of species, such as yeast, nematodes, fruit flies, mice and humans. These studies have also shown that individuals with Laron syndrome, an autosomal recessive disorder resulting in dwarfism due to defects in growth hormone receptors, have increased lifespan. Additionally, these individuals have much lower incidences of age-related diseases such as type 2 diabetes and cancer. Lastly, human centenarians around the world are disproportionately of short stature, and have low levels of IGF-1.
Reproduction and aging[edit]
Numerous studies have found that lifespan is inversely correlated with both the total amount of offspring birthed, as well as the age at which females first gives birth, also known as primiparity. Additionally, it has been found that reproduction is a costly mechanism that alters the metabolism of fat. Lipids invested in reproduction would be unable to be allocated to support mechanisms involved in somatic maintenance.
Animal models[edit]
The disposable soma theory has been consistent with the majority of animal models. It was found in numerous animal studies that castration or genetic deformities of reproduction organs was correlated with increased lifespan. Moreover, in red squirrels, it was found that females with an early primiparity achieved the highest immediate and lifetime reproductive success. However, it was also found that these same individuals had a decreased median and maximum lifespan. Specifically squirrels who mated earlier had a 22.4% rate of mortality until two years of age compared to a 16.5% rate of mortality in late breeders. In addition, these squirrels had an average maximum lifespan of 1035 days compared to an average maximum lifespan of 1245 days for squirrels that bred later.
In another study, researchers selectively bred fruit flies over three years to develop two different strains, an early-reproducing strain and a late-reproducing strain. The late-reproducing line had a significantly longer lifespan than the early-reproducing line. Even more telling was that when the researchers introduced a mutation in the ovarian-associated gene ovoD1, resulting in defective oogenesis, the differences in lifespan between the two lines disappeared. The researchers in this case concluded that "aging has evolved primarily because of the damaging effects of reproduction earlier in life".
Prominent aging researcher Steven Austad also performed a large-scale ecological study on the coast of Georgia in 1993. Austad isolated two opossum populations, one from the predator-infested mainland and one from the predator-absent nearby island of Sapelo. According to the disposable soma theory, a genetically isolated population subject to low environmentally-induced mortality would evolve delayed reproduction and aging. This is because without the pressure of predation, it would be evolutionarily advantageous to allocate more resources to somatic maintenance than reproduction, as early offspring mortality would be low. As predicted, even after controlling for predation, the isolated population had a longer lifespan, delayed primiparity, and reduced aging biomarkers such as tail collagen cross-linking.
Human models[edit]
In general, only a few studies exist in human models. It was found that castrated men live longer than their fertile counterparts. Further studies found that in British women, primiparity was earliest in women who died early and latest in women who died at the oldest ages. Furthermore, increased number of children birthed was associated with a decreased lifespan. A final study found that female centenarians were more likely to have children in later life compared average, especially past the age of 40. The researchers discovered that 19.2% of female centenarians had their first child after the age of 40, compared to 5.5% of the rest of the female population.
Relationship between cell damage and aging[edit]
Main article: Free radical theory
Main article: DNA damage theory of aging
The naked mole rat has a disproportionately long life of 30 years through efficient cellular repair mechanisms.
There are numerous studies that support cellular damage, often due to a lack of somatic maintenance mechanisms, as a primary determinant for aging, and these studies have given rise to the free radical theory of aging and the DNA damage theory of aging. One study found that the cells of short-living rodents in vitro show much greater mutation rates and a general lack of genome surveillance compared to human cells and are far more susceptible to oxidative stress.
Other studies have been conducted on the naked mole rat, a rodent species with remarkable longevity (30 years), capable of outliving the brown rat (3 years) by ten-fold. Additionally, almost no incidence of cancer has ever been detected in naked mole rats. Nearly all of the differences found between these two organisms, which are otherwise rather genetically similar, was in somatic maintenance. Naked mole rats were found to have higher levels of superoxide dismutase, a reactive oxygen species clearing antioxidant. In addition, naked mole rats had higher levels of base excision repair, DNA damage response signaling, homologous recombination repair, mismatch repair, nucleotide excision repair, and non-homologous end joining. In fact, many of these processes were near or exceeded human levels. Proteins from naked mole rats were also more resistant to oxidation, misfolding, ubiquitination, and had increased translational fidelity.
Further studies have been conducted on patients with Hutchinson-Gilford Progeria Syndrome (HGPS), a condition that leads to premature aging. Patients with HGPS typically age about seven times faster than average and usually succumb to the disease in their early teens. Patients with HGPS have cellular defects, specifically in the lamin proteins, which regulate the organization of the lamina and nuclear envelope for mitosis. A-type lamins promote genetic stability by maintaining levels of proteins that have key roles in the repair processes of non-homologous end joining and homologous recombination. Mouse cells deficient for maturation of prelamin A show increased DNA damage and chromosome aberrations and have increased sensitivity to DNA damaging agents.
Lastly, as mentioned previously, it has been found that the suppression of autophagy is associated with reduced lifespan, while stimulation is associated with extended lifespan. Activated in times of caloric restriction, autophagy is a process that prevents cellular damage through clearance and recycling of damaged proteins and organelles.
Criticism[edit]
One of the main weaknesses of the disposable soma theory is that it does not postulate any specific cellular mechanisms to which an organism shifts energy to somatic repair over reproduction. Instead, it only offers an evolutionary perspective on why aging may occur due to reproduction. Therefore, parts of it are rather limited outside of the field of evolutionary biology.
Caloric restriction[edit]
Main article: Calorie restriction § Sirtuin-mediated mechanism
Schematic showing the reallocation of energy investment towards self-repair during caloric restriction
Critics have pointed out the supposed inconsistencies of the disposable soma theory due to the observed effects of caloric restriction, which is correlated with increased lifespan. Although it activates autophagy, according to classical disposable soma principles, with less caloric intake, there would less total energy to be distributed towards somatic maintenance, and decreased lifespan would be observed (or at least the positive autophagic effects would be balanced out). However, Kirkwood, alongside his collaborator Darryl P. Shanley, assert that caloric restriction triggers an adaptive mechanism which causes the organism to shift a higher proportion of resources to somatic maintenance, away from reproduction. This theory is supported by multiple studies, which show that caloric restriction typically results in impaired fertility, but leave an otherwise healthy organism. Evolutionarily, an organism would want to delay reproduction to when resources were more plentiful. During a resource-barren period, it would evolutionarily unwise to invest resources in progeny that would be unlikely to survive in famine. Mechanistically, the NAD-dependent deacetylase Sirtuin 1 (SIRT-1) is upregulated during low-nutrient periods. SIRT-1 increases insulin sensitivity, decreases the amount of inflammatory cytokines, stimulates autophagy, and activates FOXO, the aforementioned protein involved in activating stress response genes. SIRT-1 is also found to result in decreased fertility.
In additional to differential partitioning of energy allocation during caloric restriction, less caloric intake would result in less metabolic waste in the forms of free radicals like hydrogen peroxide, superoxide and hydroxyl radicals, which damage important cellular components, particularly mitochondria. Elevated levels of free radicals in mice has been correlated with neurodegeneration, myocardial injury, severe anemia, and premature death.
No changes were observed in the spontaneous chromosomal mutation frequency of dietary restricted mice (aged 6 and 12 months) compared to ad libitum fed control mice. Thus dietary restriction appears to have no appreciable effect on spontaneous mutation in chromosomal DNA, and the increased longevity of dietary restricted mice apparently is not attributable to reduced chromosomal mutation frequency.
The grandmother hypothesis[edit]
Main article: Menopause § Evolutionary rationale
Another primary criticism of the disposable soma theory is that it fails to account for why women tend to live longer than their male counterparts. Afterall, females invest significantly more resources into reproduction and according to the classical disposable soma principles, this would compromise energy diverted to somatic maintenance. However, this can be reconciled with the grandmother hypothesis. The Grandmother Hypothesis states that menopause comes about into older women in order to restrict the time of reproduction as a protective mechanism. This would allow women to live longer and increase the amount of care they could provide to their grandchildren, increasing their evolutionary fitness. And so, although women do invest a greater proportion of resources into reproduction during their fertile years, their overall reproductive period is significantly shorter than men, who are able of reproduction during and even beyond middle age. Additionally, males invest more resources into growth compare to females, which is correlated with decreased lifespan. Other variables such as increased testosterone levels in males are not accounted for. Increased testosterone is often associated with reckless behaviour, which may lead to a high accidental death rate.
Contradicting models[edit]
A few contradicting animal models weaken the validity of the disposable soma theory. This includes studies done on the aforementioned naked mole rats. In these studies, it was found that reproductive naked mole rats actually show significantly increased lifespans compared to non-reproductive individuals, which contradicts the principles of disposable soma. However, although these naked mole rats are mammalian, they are highly atypical in terms of aging studies and may not serve as the best model for humans. For example, naked mole rats have a disproportionately high longevity quotient and live in eusocial societies, where breeding is usually designated to a queen.
Sex biases and environment[edit]
The disposable soma theory is tested disproportionately on female organisms for the relationship between reproduction and aging, as females carry a greater burden in reproduction. Additionally, for the relationship between growth and aging, studies are disproportionately conducted on males, to minimize the hormonal fluctuations that occur with menstrual cycling. Lastly, genetic and environmental factors, rather than reproductive patterns, may explain the variations in human lifespan. For example, studies have shown that poorer individuals, to whom nutritious food and medical care is less accessible, typically have higher birth rates and earlier primiparity. | biology | 998 | https://da.wikipedia.org/wiki/Art | Art | Arten (species, forkortet sp., flertal: spp.) er den grundlæggende systematiske enhed inden for biologien. Arten defineres ofte som en naturlig gruppe af populationer, hvor udveksling af gener finder sted (eller kan finde sted) og som i forhold til forplantning er isoleret fra andre grupper. Det vil sige at kun individer inden for samme art kan parre sig og få forplantningsdygtigt afkom. Dette kaldes det biologiske artsbegreb. For organismer, der formerer sig ukønnet eller ved selvbestøvning, må arter afgrænses ud fra ligheder og forskelle mellem forskellige individer. Nogle dyrearter kan i fangenskab hybridisere og få fertilt afkom, men da dette ikke vil ske i naturen, selv om de mødes her, betragtes de som forskellige arter.
Eksempel
To heste kan parre sig og få et føl, der igen kan få føl med andre heste – hestene tilhører derfor samme art. En hest og et æsel kan også parre sig og deres unger kaldes enten muldyr eller mulæsel, afhængig af hvem der er moren, men muldyret eller mulæselet kan (normalt) ikke få unger, da de oftest er sterile. Af den grund regnes hest og æsel som to forskellige arter. Det samme princip gælder også for planterne. Denne naturskabte afgrænsning mellem to arter kaldes en artsbarriere. Den kan af og til gennembrydes, når ellers sterile krydsninger spontant eller kunstigt får gennemført en kromosomfordobling. Se f.eks. Vadegræs (Spartina pectinata).
Arter over for hybrider
Man kan dog godt komme ud for, at arter kan krydses og får blandet afkom, men hybriden vil kun kunne bestå på steder, hvor ingen af forældrearterne kan klare sig. Dette er et særligt udpræget problem med Rododendron (Rhododendron) og Tjørn (Crataegus), fordi disse slægter breder sig voldsomt efter skovbrand eller stormfald. Da hybriderne bliver frugtbare i en yngre alder end arterne, kan de dominere i en periode, men når skoven lukker sig, så fortrænges hybriderne og kun de specialiserede arter kan overleve i skovens dybe skygge eller ude i lyset i sumpe, på ur og i kalksten, m.m.
Flere artsbegreber
Fordi det biologiske artsbegreb kan være besværligt at anvende i praksis, er der efterhånden skabt en række andre artsbegreber:
Morfologisk artsbegreb Arterne adskiller sig fra hinanden ved deres bygning. Dette begreb er blevet meget anvendt gennem tiden.
Økologisk artsbegreb Definerer en art som en gruppe af organismer, der udfylder samme niche. Krydsninger mellem to nærtstående arter vil ikke være optimalt tilpasset til forældrearternes nicher og vil ikke klare sig i konkurrencen.
Evolutionære artsbegreb Også kaldet det kladistiske eller fylogenetiske artsbegreb. Naturen er dynamisk, ikke statisk - alle arter ændrer sig med tiden og bliver, hvis de ikke uddør som følge af konkurrence, naturkatastrofer m.v., til én eller flere nye arter. Det evolutionære artsbegreb minder om det biologiske, men inddrager tidsdimensionen, det vil sige at en art udvikler sig over tid og at nye arter opstår ved artsdannelse. Individer der fylogenetisk har samme stamfader tilhører samme art.
Pluralistisk artsbegreb En art er et samfund af populationer, der formerer sig og lever inden for en bestemt niche i naturen.
Se også
Systematik
Evolutionsteori
Kilder
Lars Skipper: Hvad er en art? Citat: "...Arten er den eneste [klassifikations-kategori] der eksisterer i virkeligheden, alle andre (slægter, familier, ordener m.v.) er indført for overskuelighedens skyld..."
Eksterne henvisninger
2003-12-31, ScienceDaily: Working On The 'Porsche Of Its Time': New Model For Species Determination Offered Citat: "...two species of dinosaur that are members of the same genera varied from each other by just 2.2 percent. Translation of the percentage into an actual number results in an average of just three skeletal differences out of the total 338 bones in the body. Amazingly, 58 percent of these differences occurred in the skull alone. "This is a lot less variation than I'd expected," said Novak..."
2003-08-08, ScienceDaily: Cross-species Mating May Be Evolutionarily Important And Lead To Rapid Change, Say Indiana University Researchers Citat: "...the sudden mixing of closely related species may occasionally provide the energy to impel rapid evolutionary change..."
2004-01-09 ScienceDaily: Mayo Researchers Observe Genetic Fusion Of Human, Animal Cells; May Help Explain Origin Of AIDS Citat: "...The researchers have discovered conditions in which pig cells and human cells can fuse together in the body to yield hybrid cells that contain genetic material from both species..."What we found was completely unexpected," says Jeffrey Platt, M.D..."
2000-09-18, ScienceDaily: Scientists Unravel Ancient Evolutionary History Of Photosynthesis Citat: "...gene-swapping was common among ancient bacteria early in evolution..."
2004-06-07, Sciencedaily: Parting Genomes: University Of Arizona Biologists Discover Seeds Of Speciation Citat: "...There's a huge amount of biodiversity out there, and we don't know where it comes from. Evolutionary biologists are excited to figure out what causes what we see out there--the relative forces of selection and drift--whether things are adapting to their environment or variation is random..."
2005-07-05, Sciencedaily: Trees, Vines And Nets -- Microbial Evolution Changes Its Face Citat: "... EBI researchers have changed our view of 4 billion years of microbial evolution...In all, more than 600,000 vertical transfers are observed, coupled with 90,000 gene loss events and approximately 40,000 horizontal gene transfers...A few species, including beneficial nitrogen-fixing soil bacteria, appear to be 'champions'of horizontal gene transfer; "it's entirely possible that apparently harmless organisms are quietly spreading antibiotic resistance under our feet," concludes Christos Ouzounis..."
2005-11-11, Sciencedaily: Lateral Thinking Produces First Map Of Gene Transmission Citat: "...Their results clearly show genetic modification of organisms by lateral transfer is a widespread natural phenomenon, and it can occur even between distantly related organisms... it was assumed that transfer of genes could only be vertical, i.e. from parents to offspring..."
Økologi
Biologi | danish | 0.791178 |
evolution_not_make_our_life_longer/Antagonistic_pleiotropy_hypothesis.txt | The antagonistic pleiotropy hypothesis was first proposed by George C. Williams in 1957 as an evolutionary explanation for senescence. Pleiotropy is the phenomenon where one gene controls more than one phenotypic trait in an organism. A gene is considered to possess antagonistic pleiotropy if it controls more than one trait, where at least one of these traits is beneficial to the organism's fitness early on in life and at least one is detrimental to the organism's fitness later on due to a decline in the force of natural selection. The theme of G. C. William's idea about antagonistic pleiotropy was that if a gene caused both increased reproduction in early life and aging in later life, then senescence would be adaptive in evolution. For example, one study suggests that since follicular depletion in human females causes both more regular cycles in early life and loss of fertility later in life through menopause, it can be selected for by having its early benefits outweigh its late costs.
As a constraint on perfection[edit]
Antagonistic pleiotropy is one of the several reasons evolutionary biologists give for organisms never being able to reach perfection through natural selection. Antagonistically pleiotropic genes are the explanation for fitness trade-offs. This means that genes that are pleiotropic control for some beneficial traits and some detrimental traits; thus, if they happen to persist through natural selection, this will prevent organisms from reaching perfection because if they possess the benefits of the gene, they must also possess the imperfections or faults. An example of this would be female rodents that live in a nest with other females and may end up feeding young that are not theirs due to their intense parental drive. This strong parental drive will be selected for, but the organisms will still make the mistake of feeding young that are not theirs and misallocating their resources.
Benefits and costs[edit]
Antagonistic pleiotropy has several negative consequences. It results in delayed adaptation, an altered path of evolution, and reduced adaptation of other traits. In addition, the overall benefit of alleles is cut down significantly (by about half) by pleiotropy. Still, antagonistic pleiotropy has some evolutionary benefits. In fact, the conservation of genes is directly related to the pleiotropic character of an organism. This implies that genes that control for multiple traits, even if the traits have different implications for the organism's fitness, have more staying power in an evolutionary context.
Role in sexual selection[edit]
It is generally accepted that the evolution of secondary sexual characteristics persists until the relative costs of survival outweigh the benefits of reproductive success. At the level of genes, this means a trade-off between variation and expression of selected traits. Strong, persistent sexual selection should result in decreased genetic variation for these traits. However, higher levels of variation have been reported in sexually-selected traits compared to non-sexually selected traits. This phenomenon is especially clear in lek species, where males' courtship behavior confers no immediate advantage to the female. Female choice presumably depends on correlating male displays (secondary sexual characteristics) with overall genetic quality. If such directional sexual selection depletes variation in males, why would female choice continue to exist? Rowe and Houle answer this question (the lek paradox) using the notion of genetic capture, which couples the sexually-selected traits with the overall condition of the organism. They posit that the genes for secondary sexual characteristics must be pleiotropically linked to condition, a measure of the organism's fitness. In other words, the genetic variation in secondary sexual characteristics is maintained due to variation in the organism's condition.
Role in disease[edit]
The survival of many serious genetic disorders in our long evolutionary history has led researchers to reassess the role of antagonistic pleiotropy in disease. If genetic disorders are defined by the existence of deleterious alleles, then natural selection acting over evolutionary time would result in a lower frequency of mutations than are currently observed. In a recent article, Carter and Nguyen identify several genetic disorders, arguing that far from being a rare phenomenon, antagonistic pleiotropy might be a fundamental mechanism for the survival of these non-optimal alleles.
In one of these studies, 99 individuals with Laron syndrome (a rare form of dwarfism) were monitored alongside their non-dwarf kin for a period of ten years. Patients with Laron syndrome possess one of three genotypes for the growth hormone receptor gene (GHR). Most patients have an A->G splice site mutation in position 180 in exon 6. Some others possess a nonsense mutation (R43X), while the rest are heterozygous for the two mutations. Laron syndrome patients experienced a lower incidence of cancer mortality and diabetes compared to their non-dwarf kin. This suggests a role for antagonistic pleiotropy, whereby a deleterious mutation is preserved in a population because it still confers some survival benefit.
Another instance of antagonistic pleiotropy is manifested in Huntington's disease, a rare neurodegenerative disorder characterized by a high number of CAG repeats within the Huntingtin gene. The onset of Huntington's is usually observed post-reproductive age and generally involves involuntary muscle spasms, cognitive difficulties and psychiatric problems. Incidentally, the high number of CAG repeats is associated with increased activity of p53, a tumor suppressing protein that participates in apoptosis. It has been hypothesized that this explains the lower rates of cancer among Huntington's patients. Huntington's disease is also correlated with high fecundity.
Additionally, it was found that individuals with a higher pro-inflammatory ratio TNFα/IL-10 had a significantly higher incidence of death due to cardiovascular disease in old age. Yet, it was hypothesized that this genotype was prevalent because higher ratios of TNFα/IL-10 allow individuals to more effectively combat infection during reproductive years.
Sickle cell anemia, Beta-thalassemia, and cystic fibrosis are some other examples of the role antagonistic pleiotropy may play in genetic disorders.
Ubiquity[edit]
Although there are so many negative effects related to genes that are antagonistically pleiotropic, it is still present among most forms of life. Indeed, pleiotropy is one of the most common traits possessed by genes overall. In addition to that, pleiotropy is under strong stabilizing selection. In one experiment with mice and the morphology of the mandible, 1/5 of the loci had effects of pleiotropy for the entire mandible. One other example was in the Russian biologist Dmitry K. Belyaev's study on the domestication of the fox. In Dmitry K. Belyaev's farm-fox experiment, wild foxes were bred for docile behavior alone. After 40 generations, other physiological changes had surfaced including shortened tails, floppy ears, a white star in the forehead, rolled tails, shorter legs. Since the only thing being selected for was behavior, this leads scientists to believe that these secondary characteristics were controlled by the same gene or genes as docile behavior.
Adaptivity and senescence[edit]
An antagonistically pleiotropic gene can be selected for if it has beneficial effects in early life while having its negative effects in later life because genes tend to have larger impacts on fitness in an organism's prime than in their old age. An example of this is testosterone levels in male humans. Higher levels of this hormone lead to increased fitness in early life, while causing decreased fitness in later life due to a higher risk for prostate cancer. This is an example of antagonistic pleiotropy being an explanation for senescence. Senescence is the act of ageing in individuals; it's the failure over time of the individual's life processes by natural causes. Williams's theory has been the motivation for many of the experimental studies on the reasons for aging in the last 25 years. However, there is more than one theory out there for aging. The competing model to explain senescence is Medawar's "mutation accumulation" hypothesis, saying that "over evolutionary time, late-acting mutations will accumulate at a much faster rate than early-acting mutation. These late-acting mutations will thus lead to declining viability and/or fertility as an organism ages." Medawar's theory is based around the older concept of selection shadow that had been discussed throughout the early 1900s and led to Medawar's theory after discussions with J. B. S. Haldane in the 1940s.
Potential Examples[edit]
DNA repair[edit]
DNA Damage Theory of Aging[edit]
A prominent explanation for aging at the molecular level is the DNA damage theory of aging. It has been proposed that genetic elements that regulate DNA repair in somatic cells may constitute an important example of age-dependent pleiotropic "genes". As pointed out by Vijg, genome repair and maintenance is beneficial early in life by swiftly eliminating DNA damage or damaged cells. However, studies of DNA repair in the brain and in muscle indicate that the transition from mitotic cell division to the post-mitotic condition that occurs early in life is accompanied by a reduction in DNA repair. The reduced expression of DNA repair is presumably part of an evolutionary adaptation for diverting the resources of the cell that were previously used for DNA repair, as well as for replication and cell division, to more essential neuronal and muscular functions.
The harmful effect of this genetically controlled reduction in expression is to allow increased accumulation of DNA damage. Reduced DNA repair causes increased impairment of transcription and progressive loss of cell and tissue function. However, these harmful effects of DNA damage are cumulative and most severe in chronologically older individuals whose numbers diminish with time (by causes of death that can be independent of senescence). As a consequence, the beneficial effects of the genetic elements that control the reduction of DNA repair early in life would predominate. Thus regulatory genetic elements that reduce expression of DNA repair genes in post-mitotic cells appear to be important examples of the postulated pleiotropic "genes" that are beneficial in youth but deleterious at an older age.
Telomere Theory[edit]
Another example related to aging is the Telomere theory. Telomere theory proposes that telomeres shorten with repeated cell division which attribute to cell senescence and tissue damage. The end replication problem explains the mechanism behind the inability of DNA polymerase to commence the RNA primer to perform its function in completing the lagging strand due to the shortening of DNA. Telomere shortening is common in somatic cells. However, germ line and stem cells prevent the end replication problem with the help of telomerase. Telomerase elongates the 3’ end that is then formed into a t-loop to prevent the cell from entering the G0 phase and cell senescence.
Inflammation and damage to tissue are the underlying problems due to increased senescent cells. In several studies shortened telomeres have been associated with age related sarcopenia, atherosclerotic cardiovascular disease, and cancer. However, there is still the question whether telomere length causes these diseases or if the diseases cause shortened telomeres. Hence, the shortening of telomeres complies with antagonistic pleiotropy theory. The trade-off exists as the cell benefits from telomerase which prevents permanent growth arrest but telomere shortening is associated with functional loss.
Free Radical Theory[edit]
Another example related to aging is the Free Radical theory. Free Radical Theory suggests that the free radicals, which are being produced by aerobic respiration, are causing oxidative stress to be put on the body. This oxidative stress will result in aging and lead to death. Oxygen centered radicals are very reactive and can cause the accumulation of damage on lipids, nucleic acids as well as proteins within the body. This accumulation of damage on the biological molecules changes the framework and leads to a reduction in the molecules' activity levels. Lipid peroxides accumulate in the membrane phospholipids, which in turn diminishes the mitochondrial membrane's effectiveness as a barrier. The process of DNA transcription and translation also acquires oxidative damage. The result is alterations in the base pairings of the DNA sequence. Research has found that DNA mutations from free radical damage are highly uncommon but would still lead to the build up of damaged proteins as well as decreased biological activity.
However, a large body of research has shown that antioxidants (compounds that defuse free radicals) do not slow aging or age-related diseases, putting the popular free radical theory into doubt.
See also[edit]
Evolution of ageing | biology | 1118638 | https://sv.wikipedia.org/wiki/Neutral%20evolution | Neutral evolution | Neutral evolution förklarar nedärvningen av egenskaper/mutationer som inte har en direkt inverkan på en organisms fitness (reproduktionsframgång). Teoretiskt sett kan en neutral egenskap, med hjälp av genetisk drift, sprida sig till dess att alla individer i en population bär på den. Detta är däremot mindre sannolikt än att egenskapen helt försvinner efter ett tag.
Ett vanligt missförstånd är att neutral evolution skulle motsäga processen av naturligt urval. Så är inte fallet. De två teorierna kompletterar faktiskt varandra. "The theory does not deny the role of natural selection in determining the course of adaptive evolution" (Kimura, 1986).
Neutral molekylär evolution
Det är sedan länge känt att det, mer eller mindre ofta, sker mutationer i alla organismers genom som till stor del är helt neutrala. De har alltså ingen påverkan på individens fitness och ger i många fall genprodukter som är analoga till den tidigare genotypens. En skillnad i ett baspar i en gen ger ofta samma produkt när genen transkriberats och translaterats. Ofta kan även ett protein med en eller ett par aminosyrasubstitutioner (se kodon) fortfarande bibehålla sin funktion förutsatt att strukturen inte ändrats nämnvärt, då gäller det framförallt den/de aktiva delarna av proteinet.
Själva idén av neutral molekylär evolution har blygsamt antytts, av bland annat Sueoka (1962), men Motoo Kimura var den som först framförde en konkret, sammanhängande teori 1968.
Första förutsättningen som teorin, framförd av Kimura, bygger på är att man vid jämförelse av genomen hos nu existerande organismer funnit att en övervägande stor del av de genetiska skillnaderna är selektivt neutrala. De påverkar inte organismernas fitness. Dessa skillnader betraktas som opåverkade av den naturliga selektionen och därför kan deras existens inte förklaras med principerna för naturligt urval. Teorin bygger, ifrån början, på de konsekventa skillnaderna i proteiners aminosyrasekvenser i olika organismer och hypotesen att dessa till största del är neutrala (även om aminosyrasekvenserna är lite olika har proteinerna identiska funktioner).
Andra förutsättningen är mer av en hypotes och bygger på att större delen av alla evolutionära förändringar beror på genetisk drift som agerar på neutrala alleler (neutrala mutationer ger ”nya” neutrala alleler). Förutsättningarna för fortlevnaden av en neutral mutation är olika för encelliga, asexuella organismer och sexuella organismer. När en neutral mutation sker i en encellig organism har det automatiskt bildats en ”ny” allel i populationen som kommer spridas vidare till dottercellerna när cellen delar sig. I sexuella organismer måste mutationen uppstå i en könscell som i sin tur måste vara delaktig i bildandet av en ny individ. Först när detta skett kan den ”nya” allelen spridas i populationen.
Tillämpning
Teorin om neutral evolution kan numera ses som allmänt accepterad och på så vis även användas. Huvudsakliga användningsområdet är molekylära klockor som tillämpas för att uppskatta hur länge olika organismer varit evolutionärt skilda från varandra. När man ser på olika populationer kan man anta att antalet skillnader i neutrala basparsmutationer har ökat i samma hastighet som individer med nya mutationer föds (detta enligt den genetiska driftens matematik). Anledningen till att man antar att antalet neutrala mutationer som uppstår är samma för olika organismer är att man utgår ifrån frekvensen fel som uppstår under DNA-replikationen. De enzymer som genomför DNA-replikation i olika organismer är otroligt konserverade och lika varandra, de gör alltså fel lika ofta.
Mer avancerade molekylära klockor har senare designats för att ta varierande mutationshastigheter i beaktande.
Finns det en konflikt mellan neutral evolution och selektion?
När Kimura först föreslog sin teori blev det en hel del heta diskussioner mellan biologer. Vissa tyckte att det inte kunde vara möjligt att större delen av alla evolutionära förändringar är neutrala. Däremot bör man vara mycket medveten om att neutral evolution aldrig setts som en motsägelse mot selektionen på något sätt, de är kompatibla med varandra. Poängen som görs är bara den att neutrala mutationer är vanligare än negativa och positiva mutationer (mutationer som ger sänkt eller ökad fitness).
Kimura argumenterade att på molekylär nivå (genotypen) är neutrala mutationer vanligare, medan större delen av de mutationer som syns i fenotypen troligtvis beror på selektion snarare än genetisk drift.
Vad är vanligast? Neutrala eller selektiva alleler?
Det var först när Kimuras elev Tomoko Ohta 1978 framförde en generalisering som debatten svalnade något. Hon förde in begreppet ”near-neutrality” i teorin, som förespråkar att man måste undersöka om gener påverkas mest av drift eller av populationsstorlekar.
Andreas Wagner har nyligen publicerat en serie artiklar som fört de två sidorna av debatten närmare varandra. Han demonstrerar hur evolutionära skillnader kan uppstå genom flera enskilda mutationer som sker stegvis. Om man enbart applicerar ren selektion skulle detta vara omöjligt eftersom varje steg, teoretiskt sett, måste uppstå helt enskilt och gynnas för att det ska konserveras i genomet. Wagner använder sig av uttrycket ”neutralist regime”. Under en sådan regim ackumuleras neutrala mutationer och den genetiska diversiteten ökar. När en fenotyp med högre fitness dyker upp kommer den att sprida sig i populationen och fixera genotypen, detta sker under en så kallad ”Selectionist regime”.
Reflektioner
Att neutral evolution sker kan nog betraktas som ett faktum. Men hur stor del av evolutionen som sker med hjälp av neutrala mutationer är ännu oklart. Diskussionen om detta lär fortgå ett tag till.
Värt att tänka på är ändå, precis som Wagner visat, att förekomsten av neutrala mutationer styrs av selektionen. Neutrala mutationer har endast en framtid i en population om organismen, där mutationen finns, har en fitness över 0 (organismen måste få avkomma). Ju högre fitness organismen har desto starkare blir driften mot fixering av allelen i populationen.
Man bör även tänka på att samma fenotyp kan ha olika genetisk härkomst och att olika fenotyper kan ha samma fitness i en viss miljö och olika i en annan. Evolutionen är, som så mycket annat, dynamisk.
Se även
Neutral theory of molecular evolution
Molekylär klocka
Naturligt urval
Referenser
Evolution
Evolutionsbiologi | swedish | 0.591042 |
evolution_not_make_our_life_longer/Life.txt |
Life is a quality that distinguishes matter that has biological processes, such as signaling and self-sustaining processes, from matter that does not, and is defined descriptively by the capacity for homeostasis, organisation, metabolism, growth, adaptation, response to stimuli, and reproduction. Many philosophical definitions of living systems have been proposed, such as self-organizing systems. Viruses in particular make definition difficult as they replicate only in host cells. Life exists all over the Earth in air, water, and soil, with many ecosystems forming the biosphere. Some of these are harsh environments occupied only by extremophiles.
Life has been studied since ancient times, with theories such as Empedocles's materialism asserting that it was composed of four eternal elements, and Aristotle's hylomorphism asserting that living things have souls and embody both form and matter. Life originated at least 3.5 billion years ago, resulting in a universal common ancestor. This evolved into all the species that exist now, by way of many extinct species, some of which have left traces as fossils. Attempts to classify living things, too, began with Aristotle. Modern classification began with Carl Linnaeus's system of binomial nomenclature in the 1740s.
Living things are composed of biochemical molecules, formed mainly from a few core chemical elements. All living things contain two types of large molecule, proteins and nucleic acids, the latter usually both DNA and RNA: these carry the information needed by each species, including the instructions to make each type of protein. The proteins, in turn, serve as the machinery which carries out the many chemical processes of life. The cell is the structural and functional unit of life. Smaller organisms, including prokaryotes (bacteria and archaea), consist of small single cells. Larger organisms, mainly eukaryotes, can consist of single cells or may be multicellular with more complex structure. Life is only known to exist on Earth but extraterrestrial life is thought probable. Artificial life is being simulated and explored by scientists and engineers.
Definitions
Challenge
The definition of life has long been a challenge for scientists and philosophers. This is partially because life is a process, not a substance. This is complicated by a lack of knowledge of the characteristics of living entities, if any, that may have developed outside Earth. Philosophical definitions of life have also been put forward, with similar difficulties on how to distinguish living things from the non-living. Legal definitions of life have been debated, though these generally focus on the decision to declare a human dead, and the legal ramifications of this decision. At least 123 definitions of life have been compiled.
Descriptive
Further information: Organism
Since there is no consensus for a definition of life, most current definitions in biology are descriptive. Life is considered a characteristic of something that preserves, furthers or reinforces its existence in the given environment. This implies all or most of the following traits:
Homeostasis: regulation of the internal environment to maintain a constant state; for example, sweating to reduce temperature.
Organisation: being structurally composed of one or more cells – the basic units of life.
Metabolism: transformation of energy, used to convert chemicals into cellular components (anabolism) and to decompose organic matter (catabolism). Living things require energy for homeostasis and other activities.
Growth: maintenance of a higher rate of anabolism than catabolism. A growing organism increases in size and structure.
Adaptation: the evolutionary process whereby an organism becomes better able to live in its habitat.
Response to stimuli: such as the contraction of a unicellular organism away from external chemicals, the complex reactions involving all the senses of multicellular organisms, or the motion of the leaves of a plant turning toward the sun (phototropism), and chemotaxis.
Reproduction: the ability to produce new individual organisms, either asexually from a single parent organism or sexually from two parent organisms.
Physics
Further information: Entropy and life
From a physics perspective, an organism is a thermodynamic system with an organised molecular structure that can reproduce itself and evolve as survival dictates. Thermodynamically, life has been described as an open system which makes use of gradients in its surroundings to create imperfect copies of itself. Another way of putting this is to define life as "a self-sustained chemical system capable of undergoing Darwinian evolution", a definition adopted by a NASA committee attempting to define life for the purposes of exobiology, based on a suggestion by Carl Sagan. This definition, however, has been widely criticized because according to it, a single sexually reproducing individual is not alive as it is incapable of evolving on its own. The reason for this potential flaw is that "NASA's definition" refers to life as a phenomenon, not a living individual, which makes it incomplete. Alternative definitions based on the notion of life as a phenomenon and a living individual have been proposed as continuum of a self-maintainable information, and a distinct element of this continuum, respectively. A major strength of this approach is that it defines life in terms of mathematics and physics, avoiding biological vocabulary.
Living systems
Main article: Living systems
Others take a living systems theory viewpoint that does not necessarily depend on molecular chemistry. One systemic definition of life is that living things are self-organizing and autopoietic (self-producing). Variations of this include Stuart Kauffman's definition as an autonomous agent or a multi-agent system capable of reproducing itself, and of completing at least one thermodynamic work cycle. This definition is extended by the evolution of novel functions over time.
Death
Main article: Death
Animal corpses, like this African buffalo, are recycled by the ecosystem, providing energy and nutrients for living organisms.
Death is the termination of all vital functions or life processes in an organism or cell.
One of the challenges in defining death is in distinguishing it from life. Death would seem to refer to either the moment life ends, or when the state that follows life begins. However, determining when death has occurred is difficult, as cessation of life functions is often not simultaneous across organ systems. Such determination, therefore, requires drawing conceptual lines between life and death. This is problematic because there is little consensus over how to define life. The nature of death has for millennia been a central concern of the world's religious traditions and of philosophical inquiry. Many religions maintain faith in either a kind of afterlife or reincarnation for the soul, or resurrection of the body at a later date.
Viruses
Main article: Virus
Adenoviruses as seen under an electron microscope
Whether or not viruses should be considered as alive is controversial. They are most often considered as just gene coding replicators rather than forms of life. They have been described as "organisms at the edge of life" because they possess genes, evolve by natural selection, and replicate by making multiple copies of themselves through self-assembly. However, viruses do not metabolise and they require a host cell to make new products. Virus self-assembly within host cells has implications for the study of the origin of life, as it may support the hypothesis that life could have started as self-assembling organic molecules.
History of study
Materialism
Main article: Materialism
Some of the earliest theories of life were materialist, holding that all that exists is matter, and that life is merely a complex form or arrangement of matter. Empedocles (430 BC) argued that everything in the universe is made up of a combination of four eternal "elements" or "roots of all": earth, water, air, and fire. All change is explained by the arrangement and rearrangement of these four elements. The various forms of life are caused by an appropriate mixture of elements.
Democritus (460 BC) was an atomist; he thought that the essential characteristic of life was having a soul (psyche), and that the soul, like everything else, was composed of fiery atoms. He elaborated on fire because of the apparent connection between life and heat, and because fire moves.
Plato, in contrast, held that the world was organized by permanent forms, reflected imperfectly in matter; forms provided direction or intelligence, explaining the regularities observed in the world. The mechanistic materialism that originated in ancient Greece was revived and revised by the French philosopher René Descartes (1596–1650), who held that animals and humans were assemblages of parts that together functioned as a machine. This idea was developed further by Julien Offray de La Mettrie (1709–1750) in his book L'Homme Machine. In the 19th century the advances in cell theory in biological science encouraged this view. The evolutionary theory of Charles Darwin (1859) is a mechanistic explanation for the origin of species by means of natural selection. At the beginning of the 20th century Stéphane Leduc (1853–1939) promoted the idea that biological processes could be understood in terms of physics and chemistry, and that their growth resembled that of inorganic crystals immersed in solutions of sodium silicate. His ideas, set out in his book La biologie synthétique was widely dismissed during his lifetime, but has incurred a resurgence of interest in the work of Russell, Barge and colleagues.
Hylomorphism
Main article: Hylomorphism
The structure of the souls of plants, animals, and humans, according to Aristotle
Hylomorphism is a theory first expressed by the Greek philosopher Aristotle (322 BC). The application of hylomorphism to biology was important to Aristotle, and biology is extensively covered in his extant writings. In this view, everything in the material universe has both matter and form, and the form of a living thing is its soul (Greek psyche, Latin anima). There are three kinds of souls: the vegetative soul of plants, which causes them to grow and decay and nourish themselves, but does not cause motion and sensation; the animal soul, which causes animals to move and feel; and the rational soul, which is the source of consciousness and reasoning, which (Aristotle believed) is found only in man. Each higher soul has all of the attributes of the lower ones. Aristotle believed that while matter can exist without form, form cannot exist without matter, and that therefore the soul cannot exist without the body.
This account is consistent with teleological explanations of life, which account for phenomena in terms of purpose or goal-directedness. Thus, the whiteness of the polar bear's coat is explained by its purpose of camouflage. The direction of causality (from the future to the past) is in contradiction with the scientific evidence for natural selection, which explains the consequence in terms of a prior cause. Biological features are explained not by looking at future optimal results, but by looking at the past evolutionary history of a species, which led to the natural selection of the features in question.
Spontaneous generation
Main article: Spontaneous generation
Spontaneous generation was the belief that living organisms can form without descent from similar organisms. Typically, the idea was that certain forms such as fleas could arise from inanimate matter such as dust or the supposed seasonal generation of mice and insects from mud or garbage.
The theory of spontaneous generation was proposed by Aristotle, who compiled and expanded the work of prior natural philosophers and the various ancient explanations of the appearance of organisms; it was considered the best explanation for two millennia. It was decisively dispelled by the experiments of Louis Pasteur in 1859, who expanded upon the investigations of predecessors such as Francesco Redi. Disproof of the traditional ideas of spontaneous generation is no longer controversial among biologists.
Vitalism
Main article: Vitalism
Vitalism is the belief that there is a non-material life-principle. This originated with Georg Ernst Stahl (17th century), and remained popular until the middle of the 19th century. It appealed to philosophers such as Henri Bergson, Friedrich Nietzsche, and Wilhelm Dilthey, anatomists like Xavier Bichat, and chemists like Justus von Liebig. Vitalism included the idea that there was a fundamental difference between organic and inorganic material, and the belief that organic material can only be derived from living things. This was disproved in 1828, when Friedrich Wöhler prepared urea from inorganic materials. This Wöhler synthesis is considered the starting point of modern organic chemistry. It is of historical significance because for the first time an organic compound was produced in inorganic reactions.
During the 1850s Hermann von Helmholtz, anticipated by Julius Robert von Mayer, demonstrated that no energy is lost in muscle movement, suggesting that there were no "vital forces" necessary to move a muscle. These results led to the abandonment of scientific interest in vitalistic theories, especially after Eduard Buchner's demonstration that alcoholic fermentation could occur in cell-free extracts of yeast. Nonetheless, belief still exists in pseudoscientific theories such as homoeopathy, which interprets diseases and sickness as caused by disturbances in a hypothetical vital force or life force.
Development
Life timelineThis box: viewtalkedit−4500 —–—–−4000 —–—–−3500 —–—–−3000 —–—–−2500 —–—–−2000 —–—–−1500 —–—–−1000 —–—–−500 —–—–0 — Water Single-celled life Photosynthesis Eukaryotes Multicellular life Plants Arthropods MolluscsFlowersDinosaurs MammalsBirdsPrimatesHadeanArcheanProterozoicPhanerozoic ←Earth formed←Earliest water←LUCA←Earliest fossils←LHB meteorites←Earliest oxygen←Pongola glaciation*←Atmospheric oxygen←Huronian glaciation*←Sexual reproduction←Earliest multicellular life←Earliest fungi←Earliest plants←Earliest animals←Cryogenian ice age*←Ediacaran biota←Cambrian explosion←Andean glaciation*←Earliest tetrapods←Karoo ice age*←Earliest apes / humans←Quaternary ice age*(million years ago)*Ice Ages
Origin of life
Main article: Abiogenesis
The age of Earth is about 4.54 billion years. Life on Earth has existed for at least 3.5 billion years, with the oldest physical traces of life dating back 3.7 billion years. Estimates from molecular clocks, as summarized in the TimeTree public database, place the origin of life around 4.0 billion years ago. Hypotheses on the origin of life attempt to explain the formation of a universal common ancestor from simple organic molecules via pre-cellular life to protocells and metabolism. In 2016, a set of 355 genes from the last universal common ancestor was tentatively identified.
The biosphere is postulated to have developed, from the origin of life onwards, at least some 3.5 billion years ago. The earliest evidence for life on Earth includes biogenic graphite found in 3.7 billion-year-old metasedimentary rocks from Western Greenland and microbial mat fossils found in 3.48 billion-year-old sandstone from Western Australia. More recently, in 2015, "remains of biotic life" were found in 4.1 billion-year-old rocks in Western Australia. In 2017, putative fossilised microorganisms (or microfossils) were announced to have been discovered in hydrothermal vent precipitates in the Nuvvuagittuq Belt of Quebec, Canada that were as old as 4.28 billion years, the oldest record of life on Earth, suggesting "an almost instantaneous emergence of life" after ocean formation 4.4 billion years ago, and not long after the formation of the Earth 4.54 billion years ago.
Evolution
Main article: Evolution
Evolution is the change in heritable characteristics of biological populations over successive generations. It results in the appearance of new species and often the disappearance of old ones. Evolution occurs when evolutionary processes such as natural selection (including sexual selection) and genetic drift act on genetic variation, resulting in certain characteristics increasing or decreasing in frequency within a population over successive generations. The process of evolution has given rise to biodiversity at every level of biological organisation.
Fossils
Main article: Fossils
Fossils are the preserved remains or traces of organisms from the remote past. The totality of fossils, both discovered and undiscovered, and their placement in layers (strata) of sedimentary rock is known as the fossil record. A preserved specimen is called a fossil if it is older than the arbitrary date of 10,000 years ago. Hence, fossils range in age from the youngest at the start of the Holocene Epoch to the oldest from the Archaean Eon, up to 3.4 billion years old.
Extinction
Main article: Extinction
Extinction is the process by which a species dies out. The moment of extinction is the death of the last individual of that species. Because a species' potential range may be very large, determining this moment is difficult, and is usually done retrospectively after a period of apparent absence. Species become extinct when they are no longer able to survive in changing habitat or against superior competition. Over 99% of all the species that have ever lived are now extinct. Mass extinctions may have accelerated evolution by providing opportunities for new groups of organisms to diversify.
Environmental conditions
Cyanobacteria dramatically changed the composition of life forms on Earth by leading to the near-extinction of oxygen-intolerant organisms.
The diversity of life on Earth is a result of the dynamic interplay between genetic opportunity, metabolic capability, environmental challenges, and symbiosis. For most of its existence, Earth's habitable environment has been dominated by microorganisms and subjected to their metabolism and evolution. As a consequence of these microbial activities, the physical-chemical environment on Earth has been changing on a geologic time scale, thereby affecting the path of evolution of subsequent life. For example, the release of molecular oxygen by cyanobacteria as a by-product of photosynthesis induced global changes in the Earth's environment. Because oxygen was toxic to most life on Earth at the time, this posed novel evolutionary challenges, and ultimately resulted in the formation of Earth's major animal and plant species. This interplay between organisms and their environment is an inherent feature of living systems.
Biosphere
Main article: Biosphere
Deinococcus geothermalis, a bacterium that thrives in geothermal springs and deep ocean subsurfaces.
The biosphere is the global sum of all ecosystems. It can also be termed as the zone of life on Earth, a closed system (apart from solar and cosmic radiation and heat from the interior of the Earth), and largely self-regulating. Organisms exist in every part of the biosphere, including soil, hot springs, inside rocks at least 19 km (12 mi) deep underground, the deepest parts of the ocean, and at least 64 km (40 mi) high in the atmosphere. For example, spores of Aspergillus niger have been detected in the mesosphere at an altitude of 48 to 77 km. Under test conditions, life forms have been observed to thrive in the near-weightlessness of space and to survive in the vacuum of space. Life forms thrive in the deep Mariana Trench, and inside rocks up to 580 m (1,900 ft; 0.36 mi) below the sea floor under 2,590 m (8,500 ft; 1.61 mi) of ocean off the coast of the northwestern United States, and 2,400 m (7,900 ft; 1.5 mi) beneath the seabed off Japan. In 2014, life forms were found living 800 m (2,600 ft; 0.50 mi) below the ice of Antarctica. Expeditions of the International Ocean Discovery Program found unicellular life in 120 °C sediment 1.2 km below seafloor in the Nankai Trough subduction zone. According to one researcher, "You can find microbes everywhere—they're extremely adaptable to conditions, and survive wherever they are."
Range of tolerance
The inert components of an ecosystem are the physical and chemical factors necessary for life—energy (sunlight or chemical energy), water, heat, atmosphere, gravity, nutrients, and ultraviolet solar radiation protection. In most ecosystems, the conditions vary during the day and from one season to the next. To live in most ecosystems, then, organisms must be able to survive a range of conditions, called the "range of tolerance." Outside that are the "zones of physiological stress," where the survival and reproduction are possible but not optimal. Beyond these zones are the "zones of intolerance," where survival and reproduction of that organism is unlikely or impossible. Organisms that have a wide range of tolerance are more widely distributed than organisms with a narrow range of tolerance.
Extremophiles
Further information: Extremophile
Deinococcus radiodurans is an extremophile that can resist extremes of cold, dehydration, vacuum, acid, and radiation exposure.
To survive, some microorganisms have evolved to withstand freezing, complete desiccation, starvation, high levels of radiation exposure, and other physical or chemical challenges. These extremophile microorganisms may survive exposure to such conditions for long periods. They excel at exploiting uncommon sources of energy. Characterization of the structure and metabolic diversity of microbial communities in such extreme environments is ongoing.
Classification
Main article: Biological classification
Antiquity
Main article: Aristotle's biology
The first classification of organisms was made by the Greek philosopher Aristotle (384–322 BC), who grouped living things as either plants or animals, based mainly on their ability to move. He distinguished animals with blood from animals without blood, which can be compared with the concepts of vertebrates and invertebrates respectively, and divided the blooded animals into five groups: viviparous quadrupeds (mammals), oviparous quadrupeds (reptiles and amphibians), birds, fishes and whales. The bloodless animals were divided into five groups: cephalopods, crustaceans, insects (which included the spiders, scorpions, and centipedes), shelled animals (such as most molluscs and echinoderms), and "zoophytes" (animals that resemble plants). This theory remained dominant for more than a thousand years.
Linnaean
In the late 1740s, Carl Linnaeus introduced his system of binomial nomenclature for the classification of species. Linnaeus attempted to improve the composition and reduce the length of the previously used many-worded names by abolishing unnecessary rhetoric, introducing new descriptive terms and precisely defining their meaning.
The fungi were originally treated as plants. For a short period Linnaeus had classified them in the taxon Vermes in Animalia, but later placed them back in Plantae. Herbert Copeland classified the Fungi in his Protoctista, including them with single-celled organisms and thus partially avoiding the problem but acknowledging their special status. The problem was eventually solved by Whittaker, when he gave them their own kingdom in his five-kingdom system. Evolutionary history shows that the fungi are more closely related to animals than to plants.
As advances in microscopy enabled detailed study of cells and microorganisms, new groups of life were revealed, and the fields of cell biology and microbiology were created. These new organisms were originally described separately in protozoa as animals and protophyta/thallophyta as plants, but were united by Ernst Haeckel in the kingdom Protista; later, the prokaryotes were split off in the kingdom Monera, which would eventually be divided into two separate groups, the Bacteria and the Archaea. This led to the six-kingdom system and eventually to the current three-domain system, which is based on evolutionary relationships. However, the classification of eukaryotes, especially of protists, is still controversial.
As microbiology developed, viruses, which are non-cellular, were discovered. Whether these are considered alive has been a matter of debate; viruses lack characteristics of life such as cell membranes, metabolism and the ability to grow or respond to their environments. Viruses have been classed into "species" based on their genetics, but many aspects of such a classification remain controversial.
The original Linnaean system has been modified many times, for example as follows:
Linnaeus1735
Haeckel1866
Chatton1925
Copeland1938
Whittaker1969
Woese et al.1990
Cavalier-Smith1998, 2015
2 kingdoms
3 kingdoms
2 empires
4 kingdoms
5 kingdoms
3 domains
2 empires,6/7 kingdoms
(not treated)
Protista
Prokaryota
Monera
Monera
Bacteria
Bacteria
Archaea
Archaea (2015)
Eukaryota
Protoctista
Protista
Eucarya
"Protozoa"
"Chromista"
Vegetabilia
Plantae
Plantae
Plantae
Plantae
Fungi
Fungi
Animalia
Animalia
Animalia
Animalia
Animalia
The attempt to organise the Eukaryotes into a small number of kingdoms has been challenged. The Protozoa do not form a clade or natural grouping, and nor do the Chromista (Chromalveolata).
Metagenomic
The ability to sequence large numbers of complete genomes has allowed biologists to take a metagenomic view of the phylogeny of the whole tree of life. This has led to the realisation that the majority of living things are bacteria, and that all have a common origin.
Phylogenetic tree based on rRNA genes data (Woese et al., 1990) showing the 3 life domains, with the last universal common ancestor (LUCA) at its root
A 2016 metagenomic representation of the tree of life, unrooted, using ribosomal protein sequences. Bacteria are at top (left and right); Archaea at bottom; Eukaryotes in green at bottom right.
Composition
Chemical elements
All life forms require certain core chemical elements for their biochemical functioning. These include carbon, hydrogen, nitrogen, oxygen, phosphorus, and sulfur—the elemental macronutrients for all organisms. Together these make up nucleic acids, proteins and lipids, the bulk of living matter. Five of these six elements comprise the chemical components of DNA, the exception being sulfur. The latter is a component of the amino acids cysteine and methionine. The most abundant of these elements in organisms is carbon, which has the desirable attribute of forming multiple, stable covalent bonds. This allows carbon-based (organic) molecules to form the immense variety of chemical arrangements described in organic chemistry.
Alternative hypothetical types of biochemistry have been proposed that eliminate one or more of these elements, swap out an element for one not on the list, or change required chiralities or other chemical properties.
DNA
Main article: DNA
Deoxyribonucleic acid or DNA is a molecule that carries most of the genetic instructions used in the growth, development, functioning and reproduction of all known living organisms and many viruses. DNA and RNA are nucleic acids; alongside proteins and complex carbohydrates, they are one of the three major types of macromolecule that are essential for all known forms of life. Most DNA molecules consist of two biopolymer strands coiled around each other to form a double helix. The two DNA strands are known as polynucleotides since they are composed of simpler units called nucleotides. Each nucleotide is composed of a nitrogen-containing nucleobase—either cytosine (C), guanine (G), adenine (A), or thymine (T)—as well as a sugar called deoxyribose and a phosphate group. The nucleotides are joined to one another in a chain by covalent bonds between the sugar of one nucleotide and the phosphate of the next, resulting in an alternating sugar-phosphate backbone. According to base pairing rules (A with T, and C with G), hydrogen bonds bind the nitrogenous bases of the two separate polynucleotide strands to make double-stranded DNA. This has the key property that each strand contains all the information needed to recreate the other strand, enabling the information to be preserved during reproduction and cell division. Within cells, DNA is organised into long structures called chromosomes. During cell division these chromosomes are duplicated in the process of DNA replication, providing each cell its own complete set of chromosomes. Eukaryotes store most of their DNA inside the cell nucleus.
Cells
Main article: Cell (biology)
Cells are the basic unit of structure in every living thing, and all cells arise from pre-existing cells by division. Cell theory was formulated by Henri Dutrochet, Theodor Schwann, Rudolf Virchow and others during the early nineteenth century, and subsequently became widely accepted. The activity of an organism depends on the total activity of its cells, with energy flow occurring within and between them. Cells contain hereditary information that is carried forward as a genetic code during cell division.
There are two primary types of cells, reflecting their evolutionary origins. Prokaryote cells lack a nucleus and other membrane-bound organelles, although they have circular DNA and ribosomes. Bacteria and Archaea are two domains of prokaryotes. The other primary type is the eukaryote cell, which has a distinct nucleus bound by a nuclear membrane and membrane-bound organelles, including mitochondria, chloroplasts, lysosomes, rough and smooth endoplasmic reticulum, and vacuoles. In addition, their DNA is organised into chromosomes. All species of large complex organisms are eukaryotes, including animals, plants and fungi, though with a wide diversity of protist microorganisms. The conventional model is that eukaryotes evolved from prokaryotes, with the main organelles of the eukaryotes forming through endosymbiosis between bacteria and the progenitor eukaryotic cell.
The molecular mechanisms of cell biology are based on proteins. Most of these are synthesised by the ribosomes through an enzyme-catalyzed process called protein biosynthesis. A sequence of amino acids is assembled and joined based upon gene expression of the cell's nucleic acid. In eukaryotic cells, these proteins may then be transported and processed through the Golgi apparatus in preparation for dispatch to their destination.
Cells reproduce through a process of cell division in which the parent cell divides into two or more daughter cells. For prokaryotes, cell division occurs through a process of fission in which the DNA is replicated, then the two copies are attached to parts of the cell membrane. In eukaryotes, a more complex process of mitosis is followed. However, the result is the same; the resulting cell copies are identical to each other and to the original cell (except for mutations), and both are capable of further division following an interphase period.
Multicellular structure
Multicellular organisms may have first evolved through the formation of colonies of identical cells. These cells can form group organisms through cell adhesion. The individual members of a colony are capable of surviving on their own, whereas the members of a true multi-cellular organism have developed specialisations, making them dependent on the remainder of the organism for survival. Such organisms are formed clonally or from a single germ cell that is capable of forming the various specialised cells that form the adult organism. This specialisation allows multicellular organisms to exploit resources more efficiently than single cells. About 800 million years ago, a minor genetic change in a single molecule, the enzyme GK-PID, may have allowed organisms to go from a single cell organism to one of many cells.
Cells have evolved methods to perceive and respond to their microenvironment, thereby enhancing their adaptability. Cell signalling coordinates cellular activities, and hence governs the basic functions of multicellular organisms. Signaling between cells can occur through direct cell contact using juxtacrine signalling, or indirectly through the exchange of agents as in the endocrine system. In more complex organisms, coordination of activities can occur through a dedicated nervous system.
Extraterrestrial
Main articles: Extraterrestrial life, Astrobiology, and Astroecology
Though life is confirmed only on Earth, many think that extraterrestrial life is not only plausible, but probable or inevitable. Other planets and moons in the Solar System and other planetary systems are being examined for evidence of having once supported simple life, and projects such as SETI are trying to detect radio transmissions from possible alien civilisations. Other locations within the Solar System that may host microbial life include the subsurface of Mars, the upper atmosphere of Venus, and subsurface oceans on some of the moons of the giant planets.
Investigation of the tenacity and versatility of life on Earth, as well as an understanding of the molecular systems that some organisms utilise to survive such extremes, is important for the search for extraterrestrial life. For example, lichen could survive for a month in a simulated Martian environment.
Beyond the Solar System, the region around another main-sequence star that could support Earth-like life on an Earth-like planet is known as the habitable zone. The inner and outer radii of this zone vary with the luminosity of the star, as does the time interval during which the zone survives. Stars more massive than the Sun have a larger habitable zone, but remain on the Sun-like "main sequence" of stellar evolution for a shorter time interval. Small red dwarfs have the opposite problem, with a smaller habitable zone that is subject to higher levels of magnetic activity and the effects of tidal locking from close orbits. Hence, stars in the intermediate mass range such as the Sun may have a greater likelihood for Earth-like life to develop. The location of the star within a galaxy may also affect the likelihood of life forming. Stars in regions with a greater abundance of heavier elements that can form planets, in combination with a low rate of potentially habitat-damaging supernova events, are predicted to have a higher probability of hosting planets with complex life. The variables of the Drake equation are used to discuss the conditions in planetary systems where civilisation is most likely to exist, within wide bounds of uncertainty. A "Confidence of Life Detection" scale (CoLD) for reporting evidence of life beyond Earth has been proposed.
Artificial
Main articles: Artificial life and Synthetic biology
Artificial life is the simulation of any aspect of life, as through computers, robotics, or biochemistry. Synthetic biology is a new area of biotechnology that combines science and biological engineering. The common goal is the design and construction of new biological functions and systems not found in nature. Synthetic biology includes the broad redefinition and expansion of biotechnology, with the ultimate goals of being able to design and build engineered biological systems that process information, manipulate chemicals, fabricate materials and structures, produce energy, provide food, and maintain and enhance human health and the environment.
See also
Biology, the study of life
Biosignature
Carbon-based life
Central dogma of molecular biology
History of life
Lists of organisms by population
Viable system theory
Notes
^ Viruses are strongly believed not to descend from a common ancestor, with each realm corresponding to separate instances of a virus coming into existence. | biology | 4504220 | https://sv.wikipedia.org/wiki/Phyllodium%20elegans | Phyllodium elegans | Phyllodium elegans är en ärtväxtart som först beskrevs av João de Loureiro, och fick sitt nu gällande namn av Nicaise Auguste Desvaux. Phyllodium elegans ingår i släktet Phyllodium och familjen ärtväxter. IUCN kategoriserar arten globalt som livskraftig. Inga underarter finns listade i Catalogue of Life.
Källor
Externa länkar
Ärtväxter
elegans | swedish | 0.958866 |
evolution_not_make_our_life_longer/Mutation_accumulation_theory.txt | The mutation accumulation theory of aging was first proposed by Peter Medawar in 1952 as an evolutionary explanation for biological aging and the associated decline in fitness that accompanies it. Medawar used the term 'senescence' to refer to this process. The theory explains that, in the case where harmful mutations are only expressed later in life, when reproduction has ceased and future survival is increasingly unlikely, then these mutations are likely to be unknowingly passed on to future generations. In this situation the force of natural selection will be weak, and so insufficient to consistently eliminate these mutations. Medawar posited that over time these mutations would accumulate due to genetic drift and lead to the evolution of what is now referred to as aging.
Background and history[edit]
Despite Charles Darwin's completion of his theory of biological evolution in the 19th century, the modern logical framework for evolutionary theories of aging wouldn't emerge until almost a century later. Though August Weismann did propose his theory of programmed death, it was met with criticism and never gained mainstream attention. It wasn't until 1930 that Ronald Fisher first noted the conceptual insight which prompted the development of modern aging theories. This concept, namely that the force of natural selection on an individual decreases with age, was analysed further by J. B. S. Haldane, who suggested it as an explanation for the relatively high prevalence of Huntington's disease despite the autosomal dominant nature of the mutation. Specifically, as Huntington's only presents after the age of 30, the force of natural selection against it would have been relatively low in pre-modern societies. It was based on the ideas of Fisher and Haldane that Peter Medawar was able to work out the first complete model explaining why aging occurs, which he presented in a lecture in 1951 and then published in 1952
Mechanism of action[edit]
(a) The survival rate within a population decreases with age, while the reproduction rate remains constant. (b) The reproduction probability peaks early in life, at sexual maturity, and then steadily decreases as an individual ages, with the remaining share of the population decreasing with age as they enter the selection shadow.
Amongst almost all populations, the likelihood that an individual will reproduce is related directly to their age. Starting at 0 at birth, the probability increases to its maximum in young adulthood once sexual maturity has been reached, before gradually decreasing with age. This decrease is caused by the increasing likelihood of death due to external pressures such as predation or illness, as well as the internal pressures inherent to organisms that experience senescence. In such cases deleterious mutations which are expressed early on are strongly selected against due to their major impact on the number of offspring produced by that individual. Mutations that present later in life, by contrast, are relatively unaffected by selective pressure, as their carriers have already passed on their genes, assuming they survive long enough for the mutation to be expressed at all. The result, as predicted by Medawar, is that deleterious late-life mutations will accumulate and result in the evolution of aging as it is known colloquially. This concept is portrayed graphically by Medawar through the concept of a "selection shadow". The shaded region represents the 'shadow' of time during which selective pressure has no effect. Mutations that are expressed within this selection shadow will remain as long as reproductive probability within that age range remains low.
Evidence supporting the mutation accumulation theory[edit]
Predation and Delayed Senescence[edit]
In populations where extrinsic mortality is low, the drop in reproductive probability after maturity is less severe than in other cases. The mutation accumulation theory therefore predicts that such populations would evolve delayed senescence. One such example of this scenario can be seen when comparing birds to organisms of equivalent size. It has been suggested that their ability to fly, and therefore lower relative risk of predation, is the cause of their longer than expected life span. The implication that flight, and therefore lower predation, increases lifespan is further born out by the fact that bats live on average 3 times longer than similarly sized mammals with comparable metabolic rates. Providing further evidence, insect populations are known to experience very high rates of extrinsic mortality, and as such would be expected to experience rapid senescence and short life spans. The exception to this rule, however, is found in the longevity of eusocial insect queens. As expected when applying the mutation accumulation theory, established queens are at almost no risk of predation or other forms of extrinsic mortality, and consequently age far more slowly than others of their species.
Age-specific reproductive success of Drosophila Melanogaster[edit]
In the interest of finding specific evidence for the mutation accumulation theory, separate from that which also supports the similar antagonistic pleiotropy hypothesis, an experiment was conducted involving the breeding of successive generations of Drosophila Melanogaster. Genetic models predict that, in the case of mutation accumulation, elements of fitness, such as reproductive success and survival, will show age-related increases in dominance, homozygous genetic variance and additive variance. Inbreeding depression will also increase with age. This is because these variables are proportional to the equilibrium frequencies of deleterious alleles, which are expected to increase with age under mutation accumulation but not under the antagonistic pleiotropy hypothesis. This was tested experimentally by measuring age specific reproductive success in 100 different genotypes of Drosophila Melanogaster, with findings ultimately supporting the mutation accumulation theory of aging.
Criticisms of the mutation accumulation theory[edit]
Under most assumptions, the mutation accumulation theory predicts that mortality rates will reach close to 100% shortly after reaching post-reproductive age. Experimental populations of Drosophila Melanogaster, and other organisms, however, exhibit age-specific mortality rates that plateau well before reaching 100%, making mutation accumulation alone an insufficient explanation. It is suggested instead that mutation accumulation is only one factor among many, which together form the cause of aging. In particular, the mutation accumulation theory, the antagonistic pleiotropy hypothesis and the disposable soma theory of aging are all believed to contribute in some way to senescence. | biology | 1118638 | https://sv.wikipedia.org/wiki/Neutral%20evolution | Neutral evolution | Neutral evolution förklarar nedärvningen av egenskaper/mutationer som inte har en direkt inverkan på en organisms fitness (reproduktionsframgång). Teoretiskt sett kan en neutral egenskap, med hjälp av genetisk drift, sprida sig till dess att alla individer i en population bär på den. Detta är däremot mindre sannolikt än att egenskapen helt försvinner efter ett tag.
Ett vanligt missförstånd är att neutral evolution skulle motsäga processen av naturligt urval. Så är inte fallet. De två teorierna kompletterar faktiskt varandra. "The theory does not deny the role of natural selection in determining the course of adaptive evolution" (Kimura, 1986).
Neutral molekylär evolution
Det är sedan länge känt att det, mer eller mindre ofta, sker mutationer i alla organismers genom som till stor del är helt neutrala. De har alltså ingen påverkan på individens fitness och ger i många fall genprodukter som är analoga till den tidigare genotypens. En skillnad i ett baspar i en gen ger ofta samma produkt när genen transkriberats och translaterats. Ofta kan även ett protein med en eller ett par aminosyrasubstitutioner (se kodon) fortfarande bibehålla sin funktion förutsatt att strukturen inte ändrats nämnvärt, då gäller det framförallt den/de aktiva delarna av proteinet.
Själva idén av neutral molekylär evolution har blygsamt antytts, av bland annat Sueoka (1962), men Motoo Kimura var den som först framförde en konkret, sammanhängande teori 1968.
Första förutsättningen som teorin, framförd av Kimura, bygger på är att man vid jämförelse av genomen hos nu existerande organismer funnit att en övervägande stor del av de genetiska skillnaderna är selektivt neutrala. De påverkar inte organismernas fitness. Dessa skillnader betraktas som opåverkade av den naturliga selektionen och därför kan deras existens inte förklaras med principerna för naturligt urval. Teorin bygger, ifrån början, på de konsekventa skillnaderna i proteiners aminosyrasekvenser i olika organismer och hypotesen att dessa till största del är neutrala (även om aminosyrasekvenserna är lite olika har proteinerna identiska funktioner).
Andra förutsättningen är mer av en hypotes och bygger på att större delen av alla evolutionära förändringar beror på genetisk drift som agerar på neutrala alleler (neutrala mutationer ger ”nya” neutrala alleler). Förutsättningarna för fortlevnaden av en neutral mutation är olika för encelliga, asexuella organismer och sexuella organismer. När en neutral mutation sker i en encellig organism har det automatiskt bildats en ”ny” allel i populationen som kommer spridas vidare till dottercellerna när cellen delar sig. I sexuella organismer måste mutationen uppstå i en könscell som i sin tur måste vara delaktig i bildandet av en ny individ. Först när detta skett kan den ”nya” allelen spridas i populationen.
Tillämpning
Teorin om neutral evolution kan numera ses som allmänt accepterad och på så vis även användas. Huvudsakliga användningsområdet är molekylära klockor som tillämpas för att uppskatta hur länge olika organismer varit evolutionärt skilda från varandra. När man ser på olika populationer kan man anta att antalet skillnader i neutrala basparsmutationer har ökat i samma hastighet som individer med nya mutationer föds (detta enligt den genetiska driftens matematik). Anledningen till att man antar att antalet neutrala mutationer som uppstår är samma för olika organismer är att man utgår ifrån frekvensen fel som uppstår under DNA-replikationen. De enzymer som genomför DNA-replikation i olika organismer är otroligt konserverade och lika varandra, de gör alltså fel lika ofta.
Mer avancerade molekylära klockor har senare designats för att ta varierande mutationshastigheter i beaktande.
Finns det en konflikt mellan neutral evolution och selektion?
När Kimura först föreslog sin teori blev det en hel del heta diskussioner mellan biologer. Vissa tyckte att det inte kunde vara möjligt att större delen av alla evolutionära förändringar är neutrala. Däremot bör man vara mycket medveten om att neutral evolution aldrig setts som en motsägelse mot selektionen på något sätt, de är kompatibla med varandra. Poängen som görs är bara den att neutrala mutationer är vanligare än negativa och positiva mutationer (mutationer som ger sänkt eller ökad fitness).
Kimura argumenterade att på molekylär nivå (genotypen) är neutrala mutationer vanligare, medan större delen av de mutationer som syns i fenotypen troligtvis beror på selektion snarare än genetisk drift.
Vad är vanligast? Neutrala eller selektiva alleler?
Det var först när Kimuras elev Tomoko Ohta 1978 framförde en generalisering som debatten svalnade något. Hon förde in begreppet ”near-neutrality” i teorin, som förespråkar att man måste undersöka om gener påverkas mest av drift eller av populationsstorlekar.
Andreas Wagner har nyligen publicerat en serie artiklar som fört de två sidorna av debatten närmare varandra. Han demonstrerar hur evolutionära skillnader kan uppstå genom flera enskilda mutationer som sker stegvis. Om man enbart applicerar ren selektion skulle detta vara omöjligt eftersom varje steg, teoretiskt sett, måste uppstå helt enskilt och gynnas för att det ska konserveras i genomet. Wagner använder sig av uttrycket ”neutralist regime”. Under en sådan regim ackumuleras neutrala mutationer och den genetiska diversiteten ökar. När en fenotyp med högre fitness dyker upp kommer den att sprida sig i populationen och fixera genotypen, detta sker under en så kallad ”Selectionist regime”.
Reflektioner
Att neutral evolution sker kan nog betraktas som ett faktum. Men hur stor del av evolutionen som sker med hjälp av neutrala mutationer är ännu oklart. Diskussionen om detta lär fortgå ett tag till.
Värt att tänka på är ändå, precis som Wagner visat, att förekomsten av neutrala mutationer styrs av selektionen. Neutrala mutationer har endast en framtid i en population om organismen, där mutationen finns, har en fitness över 0 (organismen måste få avkomma). Ju högre fitness organismen har desto starkare blir driften mot fixering av allelen i populationen.
Man bör även tänka på att samma fenotyp kan ha olika genetisk härkomst och att olika fenotyper kan ha samma fitness i en viss miljö och olika i en annan. Evolutionen är, som så mycket annat, dynamisk.
Se även
Neutral theory of molecular evolution
Molekylär klocka
Naturligt urval
Referenser
Evolution
Evolutionsbiologi | swedish | 0.591042 |
organism_learn/Learning.txt |
Main menu Main menu move to sidebar hide Navigation Main page Contents Current events Random article About Wikipedia Contact us Donate Contribute Help Learn to edit Community portal Recent changes Upload file Search Search Create account Log in Personal tools Create account Log in Pages for logged out editors learn more Contributions Talk
Main menu Main menu move to sidebar hide Navigation Main page Contents Current events Random article About Wikipedia Contact us Donate Contribute Help Learn to edit Community portal Recent changes Upload file
Main menu Main menu move to sidebar hide Navigation Main page Contents Current events Random article About Wikipedia Contact us Donate Contribute Help Learn to edit Community portal Recent changes Upload file
Main menu move to sidebar hide Navigation Main page Contents Current events Random article About Wikipedia Contact us Donate Contribute Help Learn to edit Community portal Recent changes Upload file
Main menu move to sidebar hide Navigation Main page Contents Current events Random article About Wikipedia Contact us Donate Contribute Help Learn to edit Community portal Recent changes Upload file
Main menu move to sidebar hide Navigation Main page Contents Current events Random article About Wikipedia Contact us Donate Contribute Help Learn to edit Community portal Recent changes Upload file
Search Search Create account Log in Personal tools Create account Log in Pages for logged out editors learn more Contributions Talk
Contents move to sidebar hide (Top) 1 Types Toggle Types subsection 1.1 Non-associative learning 1.1.1 Habituation 1.1.2 Sensitization 1.2 Active learning 1.3 Associative learning 1.3.1 Operant conditioning 1.3.2 Classical conditioning 1.3.3 Observational learning 1.3.4 Imprinting 1.4 Play 1.5 Enculturation 1.6 Episodic learning 1.7 Multimedia learning 1.8 E-learning and augmented learning 1.9 Rote learning 1.10 Meaningful learning 1.11 Evidence-based learning 1.12 Formal learning 1.13 Non-formal learning 1.14 Informal learning 1.15 Nonformal learning and combined approaches 1.16 Tangential learning 1.17 Dialogic learning 1.18 Incidental learning 2 Domains 3 Transfer 4 Factors affecting learning Toggle Factors affecting learning subsection 4.1 Genetics 4.2 Socioeconomic and physical conditions 4.3 Psychological factors and teaching style 4.4 Epigenetic factors 4.5 General cognition-related factors 4.6 Adult learning vs children's learning 5 In animal evolution Toggle In animal evolution subsection 5.1 Costs and benefits of learned and innate knowledge 6 In plants 7 Machine learning Toggle Machine learning subsection 7.1 Types 7.2 Phases 8 See also Toggle See also subsection 8.1 Information theory 8.2 Types of education 9 References 10 Notes 11 Further reading 12 External links Toggle the table of contents Learning 84 languages Afrikaans Ænglisc العربية Asturianu Aymar aru Azərbaycanca বাংলা 閩南語 / Bân-lâm-gú Беларуская Български Bosanski Català Čeština ChiShona Dansk Deutsch Eesti Ελληνικά Español Esperanto Euskara فارسی Français Galego 한국어 हिन्दी Hrvatski Ido Bahasa Indonesia IsiZulu Íslenska Italiano עברית Jawa ಕನ್ನಡ ქართული Қазақша Kreyòl ayisyen Latviešu Македонски Bahasa Melayu Na Vosa Vakaviti Nederlands 日本語 Norsk bokmål Oʻzbekcha / ўзбекча ਪੰਜਾਬੀ پښتو ភាសាខ្មែរ Polski Português Română Runa Simi Русский संस्कृतम् Scots Sesotho Shqip සිංහල Simple English Slovenčina Slovenščina Српски / srpski Srpskohrvatski / српскохрватски Sunda Suomi Svenska Tagalog தமிழ் Татарча / tatarça తెలుగు ไทย Тоҷикӣ Türkçe Українська اردو Tiếng Việt Walon West-Vlams Winaray 吴语 ייִדיש 粵語 中文 Edit links Article Talk English Read Edit View history Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote From Wikipedia, the free encyclopedia Process of acquiring new knowledge For the album by Perfume Genius, see Learning (album) . "Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) . American students learning how to make and roll sushi Cognitive psychology Perception Visual perception Object recognition Face recognition Pattern recognition Attention Memory Aging and memory Emotional memory Learning Long-term memory Metacognition Language Metalanguage Thinking Cognition Concept Reasoning Decision making Problem solving Numerical cognition Numerosity adaptation effect Approximate number system Parallel individuation system v t e Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved. Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development . Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity. Types [ edit ] See also: Learning styles and Machine learning § Types of problems and tasks There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory. Non-associative learning [ edit ] Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury. Non-associative learning can be divided into habituation and sensitization . Habituation [ edit ] Main article: Habituation Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization. Sensitization [ edit ] Main article: Sensitization Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism. Active learning [ edit ] Main article: Active learning Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ). Associative learning [ edit ] Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus. Operant conditioning [ edit ] Main article: Operant conditioning Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior. Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning. Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc. Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life. Classical conditioning [ edit ] Main article: Classical conditioning The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants. Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles. Observational learning [ edit ] Main article: Observational learning Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings. Imprinting [ edit ] Main article: Imprinting (psychology) Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds. Play [ edit ] Main article: Play (activity) Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness . Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children. There are five types of play: Sensorimotor play aka functional play, characterized by the repetition of an activity Roleplay occurs starting at the age of three Rule-based play where authoritative prescribed codes of conduct are primary Construction play involves experimentation and building Movement play aka physical play These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others. Enculturation [ edit ] Main article: Enculturation Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one. Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults. Episodic learning [ edit ] Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to. Multimedia learning [ edit ] Main article: Multimedia learning Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory . E-learning and augmented learning [ edit ] Main article: Electronic learning Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones . When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education . Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning: Learner–learner (i.e. communication between and among peers with or without the teacher present), Learner–instructor (i.e. student-teacher communication), and Learner–content (i.e. intellectually interacting with content that results in changes in learners' understanding, perceptions, and cognitive structures). In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance). Rote learning [ edit ] Main article: Rote learning Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion. Meaningful learning [ edit ] See also: Deeper learning Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned. Evidence-based learning [ edit ] Main article: Evidence-based learning Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns. Formal learning [ edit ] Main article: Education A depiction of the world's oldest continually operating university, the University of Bologna , Italy Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition. Non-formal learning [ edit ] Main article: Nonformal learning Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity. Informal learning [ edit ] Main article: Informal learning Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience. Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself. Nonformal learning and combined approaches [ edit ] The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc. To learn a skill, such as solving a Rubik's Cube quickly, several factors come into play at once: Reading directions helps a player learn the patterns that solve the Rubik's Cube. Practicing the moves repeatedly helps build " muscle memory " and speed. Thinking critically about moves helps find shortcuts, which speeds future attempts. Observing the Rubik's Cube's six colors help anchor solutions in the mind. Revisiting the cube occasionally helps retain the skill. Tangential learning [ edit ] Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning. Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay. Dialogic learning [ edit ] Main article: Dialogic learning Dialogic learning is a type of learning based on dialogue. Incidental learning [ edit ] In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set. Here are some steps most commonly used in incidental teaching: An instructor will arrange the learning environment so that necessary materials are within the student's sight, but not within his reach, thus impacting his motivation to seek out those materials. An instructor waits for the student to initiate engagement. An instructor prompts the student to respond if needed. An instructor allows access to an item/activity contingent on a correct response from the student. The instructor fades out the prompting process over a period of time and subsequent trials. Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically. Domains [ edit ] Future school (1901 or 1910) Benjamin Bloom has suggested three domains of learning in his taxonomy which are: Cognitive : To recall, calculate, discuss, analyze, problem solve, etc. Psychomotor : To dance, swim, ski, dive, drive a car, ride a bike, etc. Affective : To like something or someone, love, appreciate, fear, hate, worship, etc. These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain). Transfer [ edit ] Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction. Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote. A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content. Factors affecting learning [ edit ] Main article: Evidence-based learning Genetics [ edit ] Main article: Heritability of IQ Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning. Socioeconomic and physical conditions [ edit ] Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions. The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school. Psychological factors and teaching style [ edit ] Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning . The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive. Certain techniques can increase long-term retention: The spacing effect means that lessons or studying spaced out over time ( spaced repetition ) are better than cramming Teaching material to other people "Self-explaining" (paraphrasing material to oneself) rather than passive reading Low-stakes quizzing Epigenetic factors [ edit ] Further information: Epigenetics in learning and memory The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins . During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation. General cognition-related factors [ edit ] This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging Adult learning vs children's learning [ edit ] See also: Aging brain This section needs expansion . You can help by adding to it . ( January 2023 ) Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is. Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development . What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities. In animal evolution [ edit ] Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species. Costs and benefits of learned and innate knowledge [ edit ] In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information. Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning. However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live. In plants [ edit ] In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors. Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways. During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning. While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight. Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning". Machine learning [ edit ] This section needs expansion . You can help by adding to it . ( February 2020 ) Robots can learn to cooperate. Main article: Machine learning Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output. Types [ edit ] Phases [ edit ] See also [ edit ] 21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling Information theory [ edit ] Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence Types of education [ edit ] Autodidacticism – Independent education without the guidance of teachers Andragogy – Methods and principles in adult education Pedagogy – Theory and practice of education References [ edit ] ^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 . Notes [ edit ] Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 . Further reading [ edit ] Library resources about Learning Resources in your library Ulrich Boser (2019). Learn Better: Mastering the Skills for Success in Life, Business, and School, or How to Become an Expert in Just About Anything . Rodale Books. ISBN 978-0593135310 . External links [ edit ] Wikimedia Commons has media related to Learning . Wikiquote has quotations related to Learning . How People Learn: Brain, Mind, Experience, and School (expanded edition) published by the National Academies Press Applying Science of Learning in Education: Infusing Psychological Science into the Curriculum published by the American Psychological Association Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition Authority control databases : National Spain France BnF data Germany Israel United States Czech Republic Retrieved from " https://en.wikipedia.org/w/index.php?title=Learning&oldid=1220741482 " Categories : Learning Memorization Cognitive science Developmental psychology Intelligence Neuropsychological assessment Systems science Hidden categories: Webarchive template wayback links Articles with Encyclopædia Britannica links Articles with short description Short description is different from Wikidata Wikipedia articles needing clarification from January 2011 All articles with unsourced statements Articles with unsourced statements from March 2016 Articles with unsourced statements from March 2022 Wikipedia articles needing factual verification from March 2022 Articles with excerpts Articles to be expanded from January 2023 All articles to be expanded Articles using small message boxes Articles to be expanded from February 2020 Pages displaying wikidata descriptions as a fallback via Module:Annotated link Commons category link from Wikidata Articles with BNE identifiers Articles with BNF identifiers Articles with BNFdata identifiers Articles with GND identifiers Articles with J9U identifiers Articles with LCCN identifiers Articles with NKC identifiers Articles containing video clips This page was last edited on 25 April 2024, at 16:57 (UTC) . Text is available under the Creative Commons Attribution-ShareAlike License 4.0 ; additional terms may apply. By using this site, you agree to the Terms of Use and Privacy Policy . Wikipedia® is a registered trademark of the Wikimedia Foundation, Inc. , a non-profit organization. Privacy policy About Wikipedia Disclaimers Contact Wikipedia Code of Conduct Developers Statistics Cookie statement Mobile view
Contents move to sidebar hide (Top) 1 Types Toggle Types subsection 1.1 Non-associative learning 1.1.1 Habituation 1.1.2 Sensitization 1.2 Active learning 1.3 Associative learning 1.3.1 Operant conditioning 1.3.2 Classical conditioning 1.3.3 Observational learning 1.3.4 Imprinting 1.4 Play 1.5 Enculturation 1.6 Episodic learning 1.7 Multimedia learning 1.8 E-learning and augmented learning 1.9 Rote learning 1.10 Meaningful learning 1.11 Evidence-based learning 1.12 Formal learning 1.13 Non-formal learning 1.14 Informal learning 1.15 Nonformal learning and combined approaches 1.16 Tangential learning 1.17 Dialogic learning 1.18 Incidental learning 2 Domains 3 Transfer 4 Factors affecting learning Toggle Factors affecting learning subsection 4.1 Genetics 4.2 Socioeconomic and physical conditions 4.3 Psychological factors and teaching style 4.4 Epigenetic factors 4.5 General cognition-related factors 4.6 Adult learning vs children's learning 5 In animal evolution Toggle In animal evolution subsection 5.1 Costs and benefits of learned and innate knowledge 6 In plants 7 Machine learning Toggle Machine learning subsection 7.1 Types 7.2 Phases 8 See also Toggle See also subsection 8.1 Information theory 8.2 Types of education 9 References 10 Notes 11 Further reading 12 External links Toggle the table of contents Learning 84 languages Afrikaans Ænglisc العربية Asturianu Aymar aru Azərbaycanca বাংলা 閩南語 / Bân-lâm-gú Беларуская Български Bosanski Català Čeština ChiShona Dansk Deutsch Eesti Ελληνικά Español Esperanto Euskara فارسی Français Galego 한국어 हिन्दी Hrvatski Ido Bahasa Indonesia IsiZulu Íslenska Italiano עברית Jawa ಕನ್ನಡ ქართული Қазақша Kreyòl ayisyen Latviešu Македонски Bahasa Melayu Na Vosa Vakaviti Nederlands 日本語 Norsk bokmål Oʻzbekcha / ўзбекча ਪੰਜਾਬੀ پښتو ភាសាខ្មែរ Polski Português Română Runa Simi Русский संस्कृतम् Scots Sesotho Shqip සිංහල Simple English Slovenčina Slovenščina Српски / srpski Srpskohrvatski / српскохрватски Sunda Suomi Svenska Tagalog தமிழ் Татарча / tatarça తెలుగు ไทย Тоҷикӣ Türkçe Українська اردو Tiếng Việt Walon West-Vlams Winaray 吴语 ייִדיש 粵語 中文 Edit links Article Talk English Read Edit View history Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote From Wikipedia, the free encyclopedia Process of acquiring new knowledge For the album by Perfume Genius, see Learning (album) . "Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) . American students learning how to make and roll sushi Cognitive psychology Perception Visual perception Object recognition Face recognition Pattern recognition Attention Memory Aging and memory Emotional memory Learning Long-term memory Metacognition Language Metalanguage Thinking Cognition Concept Reasoning Decision making Problem solving Numerical cognition Numerosity adaptation effect Approximate number system Parallel individuation system v t e Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved. Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development . Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity. Types [ edit ] See also: Learning styles and Machine learning § Types of problems and tasks There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory. Non-associative learning [ edit ] Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury. Non-associative learning can be divided into habituation and sensitization . Habituation [ edit ] Main article: Habituation Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization. Sensitization [ edit ] Main article: Sensitization Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism. Active learning [ edit ] Main article: Active learning Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ). Associative learning [ edit ] Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus. Operant conditioning [ edit ] Main article: Operant conditioning Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior. Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning. Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc. Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life. Classical conditioning [ edit ] Main article: Classical conditioning The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants. Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles. Observational learning [ edit ] Main article: Observational learning Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings. Imprinting [ edit ] Main article: Imprinting (psychology) Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds. Play [ edit ] Main article: Play (activity) Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness . Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children. There are five types of play: Sensorimotor play aka functional play, characterized by the repetition of an activity Roleplay occurs starting at the age of three Rule-based play where authoritative prescribed codes of conduct are primary Construction play involves experimentation and building Movement play aka physical play These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others. Enculturation [ edit ] Main article: Enculturation Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one. Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults. Episodic learning [ edit ] Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to. Multimedia learning [ edit ] Main article: Multimedia learning Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory . E-learning and augmented learning [ edit ] Main article: Electronic learning Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones . When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education . Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning: Learner–learner (i.e. communication between and among peers with or without the teacher present), Learner–instructor (i.e. student-teacher communication), and Learner–content (i.e. intellectually interacting with content that results in changes in learners' understanding, perceptions, and cognitive structures). In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance). Rote learning [ edit ] Main article: Rote learning Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion. Meaningful learning [ edit ] See also: Deeper learning Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned. Evidence-based learning [ edit ] Main article: Evidence-based learning Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns. Formal learning [ edit ] Main article: Education A depiction of the world's oldest continually operating university, the University of Bologna , Italy Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition. Non-formal learning [ edit ] Main article: Nonformal learning Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity. Informal learning [ edit ] Main article: Informal learning Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience. Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself. Nonformal learning and combined approaches [ edit ] The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc. To learn a skill, such as solving a Rubik's Cube quickly, several factors come into play at once: Reading directions helps a player learn the patterns that solve the Rubik's Cube. Practicing the moves repeatedly helps build " muscle memory " and speed. Thinking critically about moves helps find shortcuts, which speeds future attempts. Observing the Rubik's Cube's six colors help anchor solutions in the mind. Revisiting the cube occasionally helps retain the skill. Tangential learning [ edit ] Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning. Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay. Dialogic learning [ edit ] Main article: Dialogic learning Dialogic learning is a type of learning based on dialogue. Incidental learning [ edit ] In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set. Here are some steps most commonly used in incidental teaching: An instructor will arrange the learning environment so that necessary materials are within the student's sight, but not within his reach, thus impacting his motivation to seek out those materials. An instructor waits for the student to initiate engagement. An instructor prompts the student to respond if needed. An instructor allows access to an item/activity contingent on a correct response from the student. The instructor fades out the prompting process over a period of time and subsequent trials. Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically. Domains [ edit ] Future school (1901 or 1910) Benjamin Bloom has suggested three domains of learning in his taxonomy which are: Cognitive : To recall, calculate, discuss, analyze, problem solve, etc. Psychomotor : To dance, swim, ski, dive, drive a car, ride a bike, etc. Affective : To like something or someone, love, appreciate, fear, hate, worship, etc. These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain). Transfer [ edit ] Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction. Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote. A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content. Factors affecting learning [ edit ] Main article: Evidence-based learning Genetics [ edit ] Main article: Heritability of IQ Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning. Socioeconomic and physical conditions [ edit ] Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions. The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school. Psychological factors and teaching style [ edit ] Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning . The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive. Certain techniques can increase long-term retention: The spacing effect means that lessons or studying spaced out over time ( spaced repetition ) are better than cramming Teaching material to other people "Self-explaining" (paraphrasing material to oneself) rather than passive reading Low-stakes quizzing Epigenetic factors [ edit ] Further information: Epigenetics in learning and memory The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins . During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation. General cognition-related factors [ edit ] This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging Adult learning vs children's learning [ edit ] See also: Aging brain This section needs expansion . You can help by adding to it . ( January 2023 ) Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is. Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development . What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities. In animal evolution [ edit ] Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species. Costs and benefits of learned and innate knowledge [ edit ] In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information. Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning. However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live. In plants [ edit ] In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors. Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways. During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning. While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight. Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning". Machine learning [ edit ] This section needs expansion . You can help by adding to it . ( February 2020 ) Robots can learn to cooperate. Main article: Machine learning Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output. Types [ edit ] Phases [ edit ] See also [ edit ] 21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling Information theory [ edit ] Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence Types of education [ edit ] Autodidacticism – Independent education without the guidance of teachers Andragogy – Methods and principles in adult education Pedagogy – Theory and practice of education References [ edit ] ^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 . Notes [ edit ] Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 . Further reading [ edit ] Library resources about Learning Resources in your library Ulrich Boser (2019). Learn Better: Mastering the Skills for Success in Life, Business, and School, or How to Become an Expert in Just About Anything . Rodale Books. ISBN 978-0593135310 . External links [ edit ] Wikimedia Commons has media related to Learning . Wikiquote has quotations related to Learning . How People Learn: Brain, Mind, Experience, and School (expanded edition) published by the National Academies Press Applying Science of Learning in Education: Infusing Psychological Science into the Curriculum published by the American Psychological Association Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition Authority control databases : National Spain France BnF data Germany Israel United States Czech Republic Retrieved from " https://en.wikipedia.org/w/index.php?title=Learning&oldid=1220741482 " Categories : Learning Memorization Cognitive science Developmental psychology Intelligence Neuropsychological assessment Systems science Hidden categories: Webarchive template wayback links Articles with Encyclopædia Britannica links Articles with short description Short description is different from Wikidata Wikipedia articles needing clarification from January 2011 All articles with unsourced statements Articles with unsourced statements from March 2016 Articles with unsourced statements from March 2022 Wikipedia articles needing factual verification from March 2022 Articles with excerpts Articles to be expanded from January 2023 All articles to be expanded Articles using small message boxes Articles to be expanded from February 2020 Pages displaying wikidata descriptions as a fallback via Module:Annotated link Commons category link from Wikidata Articles with BNE identifiers Articles with BNF identifiers Articles with BNFdata identifiers Articles with GND identifiers Articles with J9U identifiers Articles with LCCN identifiers Articles with NKC identifiers Articles containing video clips This page was last edited on 25 April 2024, at 16:57 (UTC) . Text is available under the Creative Commons Attribution-ShareAlike License 4.0 ; additional terms may apply. By using this site, you agree to the Terms of Use and Privacy Policy . Wikipedia® is a registered trademark of the Wikimedia Foundation, Inc. , a non-profit organization. Privacy policy About Wikipedia Disclaimers Contact Wikipedia Code of Conduct Developers Statistics Cookie statement Mobile view
Contents move to sidebar hide (Top) 1 Types Toggle Types subsection 1.1 Non-associative learning 1.1.1 Habituation 1.1.2 Sensitization 1.2 Active learning 1.3 Associative learning 1.3.1 Operant conditioning 1.3.2 Classical conditioning 1.3.3 Observational learning 1.3.4 Imprinting 1.4 Play 1.5 Enculturation 1.6 Episodic learning 1.7 Multimedia learning 1.8 E-learning and augmented learning 1.9 Rote learning 1.10 Meaningful learning 1.11 Evidence-based learning 1.12 Formal learning 1.13 Non-formal learning 1.14 Informal learning 1.15 Nonformal learning and combined approaches 1.16 Tangential learning 1.17 Dialogic learning 1.18 Incidental learning 2 Domains 3 Transfer 4 Factors affecting learning Toggle Factors affecting learning subsection 4.1 Genetics 4.2 Socioeconomic and physical conditions 4.3 Psychological factors and teaching style 4.4 Epigenetic factors 4.5 General cognition-related factors 4.6 Adult learning vs children's learning 5 In animal evolution Toggle In animal evolution subsection 5.1 Costs and benefits of learned and innate knowledge 6 In plants 7 Machine learning Toggle Machine learning subsection 7.1 Types 7.2 Phases 8 See also Toggle See also subsection 8.1 Information theory 8.2 Types of education 9 References 10 Notes 11 Further reading 12 External links
Contents move to sidebar hide (Top) 1 Types Toggle Types subsection 1.1 Non-associative learning 1.1.1 Habituation 1.1.2 Sensitization 1.2 Active learning 1.3 Associative learning 1.3.1 Operant conditioning 1.3.2 Classical conditioning 1.3.3 Observational learning 1.3.4 Imprinting 1.4 Play 1.5 Enculturation 1.6 Episodic learning 1.7 Multimedia learning 1.8 E-learning and augmented learning 1.9 Rote learning 1.10 Meaningful learning 1.11 Evidence-based learning 1.12 Formal learning 1.13 Non-formal learning 1.14 Informal learning 1.15 Nonformal learning and combined approaches 1.16 Tangential learning 1.17 Dialogic learning 1.18 Incidental learning 2 Domains 3 Transfer 4 Factors affecting learning Toggle Factors affecting learning subsection 4.1 Genetics 4.2 Socioeconomic and physical conditions 4.3 Psychological factors and teaching style 4.4 Epigenetic factors 4.5 General cognition-related factors 4.6 Adult learning vs children's learning 5 In animal evolution Toggle In animal evolution subsection 5.1 Costs and benefits of learned and innate knowledge 6 In plants 7 Machine learning Toggle Machine learning subsection 7.1 Types 7.2 Phases 8 See also Toggle See also subsection 8.1 Information theory 8.2 Types of education 9 References 10 Notes 11 Further reading 12 External links
Contents move to sidebar hide (Top) 1 Types Toggle Types subsection 1.1 Non-associative learning 1.1.1 Habituation 1.1.2 Sensitization 1.2 Active learning 1.3 Associative learning 1.3.1 Operant conditioning 1.3.2 Classical conditioning 1.3.3 Observational learning 1.3.4 Imprinting 1.4 Play 1.5 Enculturation 1.6 Episodic learning 1.7 Multimedia learning 1.8 E-learning and augmented learning 1.9 Rote learning 1.10 Meaningful learning 1.11 Evidence-based learning 1.12 Formal learning 1.13 Non-formal learning 1.14 Informal learning 1.15 Nonformal learning and combined approaches 1.16 Tangential learning 1.17 Dialogic learning 1.18 Incidental learning 2 Domains 3 Transfer 4 Factors affecting learning Toggle Factors affecting learning subsection 4.1 Genetics 4.2 Socioeconomic and physical conditions 4.3 Psychological factors and teaching style 4.4 Epigenetic factors 4.5 General cognition-related factors 4.6 Adult learning vs children's learning 5 In animal evolution Toggle In animal evolution subsection 5.1 Costs and benefits of learned and innate knowledge 6 In plants 7 Machine learning Toggle Machine learning subsection 7.1 Types 7.2 Phases 8 See also Toggle See also subsection 8.1 Information theory 8.2 Types of education 9 References 10 Notes 11 Further reading 12 External links
Contents move to sidebar hide (Top) 1 Types Toggle Types subsection 1.1 Non-associative learning 1.1.1 Habituation 1.1.2 Sensitization 1.2 Active learning 1.3 Associative learning 1.3.1 Operant conditioning 1.3.2 Classical conditioning 1.3.3 Observational learning 1.3.4 Imprinting 1.4 Play 1.5 Enculturation 1.6 Episodic learning 1.7 Multimedia learning 1.8 E-learning and augmented learning 1.9 Rote learning 1.10 Meaningful learning 1.11 Evidence-based learning 1.12 Formal learning 1.13 Non-formal learning 1.14 Informal learning 1.15 Nonformal learning and combined approaches 1.16 Tangential learning 1.17 Dialogic learning 1.18 Incidental learning 2 Domains 3 Transfer 4 Factors affecting learning Toggle Factors affecting learning subsection 4.1 Genetics 4.2 Socioeconomic and physical conditions 4.3 Psychological factors and teaching style 4.4 Epigenetic factors 4.5 General cognition-related factors 4.6 Adult learning vs children's learning 5 In animal evolution Toggle In animal evolution subsection 5.1 Costs and benefits of learned and innate knowledge 6 In plants 7 Machine learning Toggle Machine learning subsection 7.1 Types 7.2 Phases 8 See also Toggle See also subsection 8.1 Information theory 8.2 Types of education 9 References 10 Notes 11 Further reading 12 External links
Toggle the table of contents Learning 84 languages Afrikaans Ænglisc العربية Asturianu Aymar aru Azərbaycanca বাংলা 閩南語 / Bân-lâm-gú Беларуская Български Bosanski Català Čeština ChiShona Dansk Deutsch Eesti Ελληνικά Español Esperanto Euskara فارسی Français Galego 한국어 हिन्दी Hrvatski Ido Bahasa Indonesia IsiZulu Íslenska Italiano עברית Jawa ಕನ್ನಡ ქართული Қазақша Kreyòl ayisyen Latviešu Македонски Bahasa Melayu Na Vosa Vakaviti Nederlands 日本語 Norsk bokmål Oʻzbekcha / ўзбекча ਪੰਜਾਬੀ پښتو ភាសាខ្មែរ Polski Português Română Runa Simi Русский संस्कृतम् Scots Sesotho Shqip සිංහල Simple English Slovenčina Slovenščina Српски / srpski Srpskohrvatski / српскохрватски Sunda Suomi Svenska Tagalog தமிழ் Татарча / tatarça తెలుగు ไทย Тоҷикӣ Türkçe Українська اردو Tiếng Việt Walon West-Vlams Winaray 吴语 ייִדיש 粵語 中文 Edit links Article Talk English Read Edit View history Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote From Wikipedia, the free encyclopedia Process of acquiring new knowledge For the album by Perfume Genius, see Learning (album) . "Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) . American students learning how to make and roll sushi Cognitive psychology Perception Visual perception Object recognition Face recognition Pattern recognition Attention Memory Aging and memory Emotional memory Learning Long-term memory Metacognition Language Metalanguage Thinking Cognition Concept Reasoning Decision making Problem solving Numerical cognition Numerosity adaptation effect Approximate number system Parallel individuation system v t e Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved. Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development . Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity. Types [ edit ] See also: Learning styles and Machine learning § Types of problems and tasks There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory. Non-associative learning [ edit ] Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury. Non-associative learning can be divided into habituation and sensitization . Habituation [ edit ] Main article: Habituation Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization. Sensitization [ edit ] Main article: Sensitization Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism. Active learning [ edit ] Main article: Active learning Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ). Associative learning [ edit ] Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus. Operant conditioning [ edit ] Main article: Operant conditioning Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior. Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning. Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc. Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life. Classical conditioning [ edit ] Main article: Classical conditioning The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants. Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles. Observational learning [ edit ] Main article: Observational learning Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings. Imprinting [ edit ] Main article: Imprinting (psychology) Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds. Play [ edit ] Main article: Play (activity) Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness . Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children. There are five types of play: Sensorimotor play aka functional play, characterized by the repetition of an activity Roleplay occurs starting at the age of three Rule-based play where authoritative prescribed codes of conduct are primary Construction play involves experimentation and building Movement play aka physical play These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others. Enculturation [ edit ] Main article: Enculturation Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one. Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults. Episodic learning [ edit ] Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to. Multimedia learning [ edit ] Main article: Multimedia learning Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory . E-learning and augmented learning [ edit ] Main article: Electronic learning Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones . When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education . Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning: Learner–learner (i.e. communication between and among peers with or without the teacher present), Learner–instructor (i.e. student-teacher communication), and Learner–content (i.e. intellectually interacting with content that results in changes in learners' understanding, perceptions, and cognitive structures). In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance). Rote learning [ edit ] Main article: Rote learning Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion. Meaningful learning [ edit ] See also: Deeper learning Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned. Evidence-based learning [ edit ] Main article: Evidence-based learning Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns. Formal learning [ edit ] Main article: Education A depiction of the world's oldest continually operating university, the University of Bologna , Italy Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition. Non-formal learning [ edit ] Main article: Nonformal learning Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity. Informal learning [ edit ] Main article: Informal learning Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience. Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself. Nonformal learning and combined approaches [ edit ] The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc. To learn a skill, such as solving a Rubik's Cube quickly, several factors come into play at once: Reading directions helps a player learn the patterns that solve the Rubik's Cube. Practicing the moves repeatedly helps build " muscle memory " and speed. Thinking critically about moves helps find shortcuts, which speeds future attempts. Observing the Rubik's Cube's six colors help anchor solutions in the mind. Revisiting the cube occasionally helps retain the skill. Tangential learning [ edit ] Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning. Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay. Dialogic learning [ edit ] Main article: Dialogic learning Dialogic learning is a type of learning based on dialogue. Incidental learning [ edit ] In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set. Here are some steps most commonly used in incidental teaching: An instructor will arrange the learning environment so that necessary materials are within the student's sight, but not within his reach, thus impacting his motivation to seek out those materials. An instructor waits for the student to initiate engagement. An instructor prompts the student to respond if needed. An instructor allows access to an item/activity contingent on a correct response from the student. The instructor fades out the prompting process over a period of time and subsequent trials. Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically. Domains [ edit ] Future school (1901 or 1910) Benjamin Bloom has suggested three domains of learning in his taxonomy which are: Cognitive : To recall, calculate, discuss, analyze, problem solve, etc. Psychomotor : To dance, swim, ski, dive, drive a car, ride a bike, etc. Affective : To like something or someone, love, appreciate, fear, hate, worship, etc. These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain). Transfer [ edit ] Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction. Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote. A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content. Factors affecting learning [ edit ] Main article: Evidence-based learning Genetics [ edit ] Main article: Heritability of IQ Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning. Socioeconomic and physical conditions [ edit ] Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions. The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school. Psychological factors and teaching style [ edit ] Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning . The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive. Certain techniques can increase long-term retention: The spacing effect means that lessons or studying spaced out over time ( spaced repetition ) are better than cramming Teaching material to other people "Self-explaining" (paraphrasing material to oneself) rather than passive reading Low-stakes quizzing Epigenetic factors [ edit ] Further information: Epigenetics in learning and memory The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins . During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation. General cognition-related factors [ edit ] This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging Adult learning vs children's learning [ edit ] See also: Aging brain This section needs expansion . You can help by adding to it . ( January 2023 ) Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is. Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development . What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities. In animal evolution [ edit ] Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species. Costs and benefits of learned and innate knowledge [ edit ] In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information. Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning. However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live. In plants [ edit ] In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors. Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways. During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning. While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight. Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning". Machine learning [ edit ] This section needs expansion . You can help by adding to it . ( February 2020 ) Robots can learn to cooperate. Main article: Machine learning Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output. Types [ edit ] Phases [ edit ] See also [ edit ] 21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling Information theory [ edit ] Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence Types of education [ edit ] Autodidacticism – Independent education without the guidance of teachers Andragogy – Methods and principles in adult education Pedagogy – Theory and practice of education References [ edit ] ^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 . Notes [ edit ] Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 . Further reading [ edit ] Library resources about Learning Resources in your library Ulrich Boser (2019). Learn Better: Mastering the Skills for Success in Life, Business, and School, or How to Become an Expert in Just About Anything . Rodale Books. ISBN 978-0593135310 . External links [ edit ] Wikimedia Commons has media related to Learning . Wikiquote has quotations related to Learning . How People Learn: Brain, Mind, Experience, and School (expanded edition) published by the National Academies Press Applying Science of Learning in Education: Infusing Psychological Science into the Curriculum published by the American Psychological Association Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition Authority control databases : National Spain France BnF data Germany Israel United States Czech Republic Retrieved from " https://en.wikipedia.org/w/index.php?title=Learning&oldid=1220741482 " Categories : Learning Memorization Cognitive science Developmental psychology Intelligence Neuropsychological assessment Systems science Hidden categories: Webarchive template wayback links Articles with Encyclopædia Britannica links Articles with short description Short description is different from Wikidata Wikipedia articles needing clarification from January 2011 All articles with unsourced statements Articles with unsourced statements from March 2016 Articles with unsourced statements from March 2022 Wikipedia articles needing factual verification from March 2022 Articles with excerpts Articles to be expanded from January 2023 All articles to be expanded Articles using small message boxes Articles to be expanded from February 2020 Pages displaying wikidata descriptions as a fallback via Module:Annotated link Commons category link from Wikidata Articles with BNE identifiers Articles with BNF identifiers Articles with BNFdata identifiers Articles with GND identifiers Articles with J9U identifiers Articles with LCCN identifiers Articles with NKC identifiers Articles containing video clips
84 languages Afrikaans Ænglisc العربية Asturianu Aymar aru Azərbaycanca বাংলা 閩南語 / Bân-lâm-gú Беларуская Български Bosanski Català Čeština ChiShona Dansk Deutsch Eesti Ελληνικά Español Esperanto Euskara فارسی Français Galego 한국어 हिन्दी Hrvatski Ido Bahasa Indonesia IsiZulu Íslenska Italiano עברית Jawa ಕನ್ನಡ ქართული Қазақша Kreyòl ayisyen Latviešu Македонски Bahasa Melayu Na Vosa Vakaviti Nederlands 日本語 Norsk bokmål Oʻzbekcha / ўзбекча ਪੰਜਾਬੀ پښتو ភាសាខ្មែរ Polski Português Română Runa Simi Русский संस्कृतम् Scots Sesotho Shqip සිංහල Simple English Slovenčina Slovenščina Српски / srpski Srpskohrvatski / српскохрватски Sunda Suomi Svenska Tagalog தமிழ் Татарча / tatarça తెలుగు ไทย Тоҷикӣ Türkçe Українська اردو Tiếng Việt Walon West-Vlams Winaray 吴语 ייִדיש 粵語 中文 Edit links
Afrikaans Ænglisc العربية Asturianu Aymar aru Azərbaycanca বাংলা 閩南語 / Bân-lâm-gú Беларуская Български Bosanski Català Čeština ChiShona Dansk Deutsch Eesti Ελληνικά Español Esperanto Euskara فارسی Français Galego 한국어 हिन्दी Hrvatski Ido Bahasa Indonesia IsiZulu Íslenska Italiano עברית Jawa ಕನ್ನಡ ქართული Қазақша Kreyòl ayisyen Latviešu Македонски Bahasa Melayu Na Vosa Vakaviti Nederlands 日本語 Norsk bokmål Oʻzbekcha / ўзбекча ਪੰਜਾਬੀ پښتو ភាសាខ្មែរ Polski Português Română Runa Simi Русский संस्कृतम् Scots Sesotho Shqip සිංහල Simple English Slovenčina Slovenščina Српски / srpski Srpskohrvatski / српскохрватски Sunda Suomi Svenska Tagalog தமிழ் Татарча / tatarça తెలుగు ไทย Тоҷикӣ Türkçe Українська اردو Tiếng Việt Walon West-Vlams Winaray 吴语 ייִדיש 粵語 中文 Edit links
Afrikaans Ænglisc العربية Asturianu Aymar aru Azərbaycanca বাংলা 閩南語 / Bân-lâm-gú Беларуская Български Bosanski Català Čeština ChiShona Dansk Deutsch Eesti Ελληνικά Español Esperanto Euskara فارسی Français Galego 한국어 हिन्दी Hrvatski Ido Bahasa Indonesia IsiZulu Íslenska Italiano עברית Jawa ಕನ್ನಡ ქართული Қазақша Kreyòl ayisyen Latviešu Македонски Bahasa Melayu Na Vosa Vakaviti Nederlands 日本語 Norsk bokmål Oʻzbekcha / ўзбекча ਪੰਜਾਬੀ پښتو ភាសាខ្មែរ Polski Português Română Runa Simi Русский संस्कृतम् Scots Sesotho Shqip සිංහල Simple English Slovenčina Slovenščina Српски / srpski Srpskohrvatski / српскохрватски Sunda Suomi Svenska Tagalog தமிழ் Татарча / tatarça తెలుగు ไทย Тоҷикӣ Türkçe Українська اردو Tiếng Việt Walon West-Vlams Winaray 吴语 ייִדיש 粵語 中文 Edit links
Article Talk English Read Edit View history Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
Article Talk English Read Edit View history Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
Read Edit View history Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
Tools Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
Tools move to sidebar hide Actions Read Edit View history General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item Print/export Download as PDF Printable version In other projects Wikimedia Commons Wikiquote
General What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item
What links here Related changes Upload file Special pages Permanent link Page information Cite this page Get shortened URL Download QR code Wikidata item
From Wikipedia, the free encyclopedia Process of acquiring new knowledge For the album by Perfume Genius, see Learning (album) . "Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) . American students learning how to make and roll sushi Cognitive psychology Perception Visual perception Object recognition Face recognition Pattern recognition Attention Memory Aging and memory Emotional memory Learning Long-term memory Metacognition Language Metalanguage Thinking Cognition Concept Reasoning Decision making Problem solving Numerical cognition Numerosity adaptation effect Approximate number system Parallel individuation system v t e Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved. Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development . Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity. Types [ edit ] See also: Learning styles and Machine learning § Types of problems and tasks There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory. Non-associative learning [ edit ] Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury. Non-associative learning can be divided into habituation and sensitization . Habituation [ edit ] Main article: Habituation Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization. Sensitization [ edit ] Main article: Sensitization Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism. Active learning [ edit ] Main article: Active learning Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ). Associative learning [ edit ] Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus. Operant conditioning [ edit ] Main article: Operant conditioning Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior. Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning. Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc. Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life. Classical conditioning [ edit ] Main article: Classical conditioning The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants. Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles. Observational learning [ edit ] Main article: Observational learning Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings. Imprinting [ edit ] Main article: Imprinting (psychology) Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds. Play [ edit ] Main article: Play (activity) Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness . Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children. There are five types of play: Sensorimotor play aka functional play, characterized by the repetition of an activity Roleplay occurs starting at the age of three Rule-based play where authoritative prescribed codes of conduct are primary Construction play involves experimentation and building Movement play aka physical play These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others. Enculturation [ edit ] Main article: Enculturation Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one. Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults. Episodic learning [ edit ] Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to. Multimedia learning [ edit ] Main article: Multimedia learning Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory . E-learning and augmented learning [ edit ] Main article: Electronic learning Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones . When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education . Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning: Learner–learner (i.e. communication between and among peers with or without the teacher present), Learner–instructor (i.e. student-teacher communication), and Learner–content (i.e. intellectually interacting with content that results in changes in learners' understanding, perceptions, and cognitive structures). In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance). Rote learning [ edit ] Main article: Rote learning Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion. Meaningful learning [ edit ] See also: Deeper learning Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned. Evidence-based learning [ edit ] Main article: Evidence-based learning Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns. Formal learning [ edit ] Main article: Education A depiction of the world's oldest continually operating university, the University of Bologna , Italy Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition. Non-formal learning [ edit ] Main article: Nonformal learning Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity. Informal learning [ edit ] Main article: Informal learning Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience. Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself. Nonformal learning and combined approaches [ edit ] The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc. To learn a skill, such as solving a Rubik's Cube quickly, several factors come into play at once: Reading directions helps a player learn the patterns that solve the Rubik's Cube. Practicing the moves repeatedly helps build " muscle memory " and speed. Thinking critically about moves helps find shortcuts, which speeds future attempts. Observing the Rubik's Cube's six colors help anchor solutions in the mind. Revisiting the cube occasionally helps retain the skill. Tangential learning [ edit ] Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning. Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay. Dialogic learning [ edit ] Main article: Dialogic learning Dialogic learning is a type of learning based on dialogue. Incidental learning [ edit ] In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set. Here are some steps most commonly used in incidental teaching: An instructor will arrange the learning environment so that necessary materials are within the student's sight, but not within his reach, thus impacting his motivation to seek out those materials. An instructor waits for the student to initiate engagement. An instructor prompts the student to respond if needed. An instructor allows access to an item/activity contingent on a correct response from the student. The instructor fades out the prompting process over a period of time and subsequent trials. Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically. Domains [ edit ] Future school (1901 or 1910) Benjamin Bloom has suggested three domains of learning in his taxonomy which are: Cognitive : To recall, calculate, discuss, analyze, problem solve, etc. Psychomotor : To dance, swim, ski, dive, drive a car, ride a bike, etc. Affective : To like something or someone, love, appreciate, fear, hate, worship, etc. These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain). Transfer [ edit ] Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction. Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote. A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content. Factors affecting learning [ edit ] Main article: Evidence-based learning Genetics [ edit ] Main article: Heritability of IQ Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning. Socioeconomic and physical conditions [ edit ] Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions. The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school. Psychological factors and teaching style [ edit ] Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning . The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive. Certain techniques can increase long-term retention: The spacing effect means that lessons or studying spaced out over time ( spaced repetition ) are better than cramming Teaching material to other people "Self-explaining" (paraphrasing material to oneself) rather than passive reading Low-stakes quizzing Epigenetic factors [ edit ] Further information: Epigenetics in learning and memory The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins . During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation. General cognition-related factors [ edit ] This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging Adult learning vs children's learning [ edit ] See also: Aging brain This section needs expansion . You can help by adding to it . ( January 2023 ) Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is. Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development . What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities. In animal evolution [ edit ] Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species. Costs and benefits of learned and innate knowledge [ edit ] In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information. Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning. However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live. In plants [ edit ] In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors. Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways. During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning. While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight. Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning". Machine learning [ edit ] This section needs expansion . You can help by adding to it . ( February 2020 ) Robots can learn to cooperate. Main article: Machine learning Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output. Types [ edit ] Phases [ edit ] See also [ edit ] 21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling Information theory [ edit ] Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence Types of education [ edit ] Autodidacticism – Independent education without the guidance of teachers Andragogy – Methods and principles in adult education Pedagogy – Theory and practice of education References [ edit ] ^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 . Notes [ edit ] Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 . Further reading [ edit ] Library resources about Learning Resources in your library Ulrich Boser (2019). Learn Better: Mastering the Skills for Success in Life, Business, and School, or How to Become an Expert in Just About Anything . Rodale Books. ISBN 978-0593135310 . External links [ edit ] Wikimedia Commons has media related to Learning . Wikiquote has quotations related to Learning . How People Learn: Brain, Mind, Experience, and School (expanded edition) published by the National Academies Press Applying Science of Learning in Education: Infusing Psychological Science into the Curriculum published by the American Psychological Association Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition Authority control databases : National Spain France BnF data Germany Israel United States Czech Republic Retrieved from " https://en.wikipedia.org/w/index.php?title=Learning&oldid=1220741482 " Categories : Learning Memorization Cognitive science Developmental psychology Intelligence Neuropsychological assessment Systems science Hidden categories: Webarchive template wayback links Articles with Encyclopædia Britannica links Articles with short description Short description is different from Wikidata Wikipedia articles needing clarification from January 2011 All articles with unsourced statements Articles with unsourced statements from March 2016 Articles with unsourced statements from March 2022 Wikipedia articles needing factual verification from March 2022 Articles with excerpts Articles to be expanded from January 2023 All articles to be expanded Articles using small message boxes Articles to be expanded from February 2020 Pages displaying wikidata descriptions as a fallback via Module:Annotated link Commons category link from Wikidata Articles with BNE identifiers Articles with BNF identifiers Articles with BNFdata identifiers Articles with GND identifiers Articles with J9U identifiers Articles with LCCN identifiers Articles with NKC identifiers Articles containing video clips
Process of acquiring new knowledge For the album by Perfume Genius, see Learning (album) . "Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) . American students learning how to make and roll sushi Cognitive psychology Perception Visual perception Object recognition Face recognition Pattern recognition Attention Memory Aging and memory Emotional memory Learning Long-term memory Metacognition Language Metalanguage Thinking Cognition Concept Reasoning Decision making Problem solving Numerical cognition Numerosity adaptation effect Approximate number system Parallel individuation system v t e Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved. Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development . Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity. Types [ edit ] See also: Learning styles and Machine learning § Types of problems and tasks There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory. Non-associative learning [ edit ] Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury. Non-associative learning can be divided into habituation and sensitization . Habituation [ edit ] Main article: Habituation Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization. Sensitization [ edit ] Main article: Sensitization Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism. Active learning [ edit ] Main article: Active learning Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ). Associative learning [ edit ] Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus. Operant conditioning [ edit ] Main article: Operant conditioning Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior. Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning. Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc. Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life. Classical conditioning [ edit ] Main article: Classical conditioning The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants. Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles. Observational learning [ edit ] Main article: Observational learning Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings. Imprinting [ edit ] Main article: Imprinting (psychology) Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds. Play [ edit ] Main article: Play (activity) Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness . Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children. There are five types of play: Sensorimotor play aka functional play, characterized by the repetition of an activity Roleplay occurs starting at the age of three Rule-based play where authoritative prescribed codes of conduct are primary Construction play involves experimentation and building Movement play aka physical play These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others. Enculturation [ edit ] Main article: Enculturation Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one. Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults. Episodic learning [ edit ] Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to. Multimedia learning [ edit ] Main article: Multimedia learning Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory . E-learning and augmented learning [ edit ] Main article: Electronic learning Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones . When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education . Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning: Learner–learner (i.e. communication between and among peers with or without the teacher present), Learner–instructor (i.e. student-teacher communication), and Learner–content (i.e. intellectually interacting with content that results in changes in learners' understanding, perceptions, and cognitive structures). In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance). Rote learning [ edit ] Main article: Rote learning Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion. Meaningful learning [ edit ] See also: Deeper learning Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned. Evidence-based learning [ edit ] Main article: Evidence-based learning Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns. Formal learning [ edit ] Main article: Education A depiction of the world's oldest continually operating university, the University of Bologna , Italy Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition. Non-formal learning [ edit ] Main article: Nonformal learning Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity. Informal learning [ edit ] Main article: Informal learning Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience. Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself. Nonformal learning and combined approaches [ edit ] The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc. To learn a skill, such as solving a Rubik's Cube quickly, several factors come into play at once: Reading directions helps a player learn the patterns that solve the Rubik's Cube. Practicing the moves repeatedly helps build " muscle memory " and speed. Thinking critically about moves helps find shortcuts, which speeds future attempts. Observing the Rubik's Cube's six colors help anchor solutions in the mind. Revisiting the cube occasionally helps retain the skill. Tangential learning [ edit ] Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning. Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay. Dialogic learning [ edit ] Main article: Dialogic learning Dialogic learning is a type of learning based on dialogue. Incidental learning [ edit ] In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set. Here are some steps most commonly used in incidental teaching: An instructor will arrange the learning environment so that necessary materials are within the student's sight, but not within his reach, thus impacting his motivation to seek out those materials. An instructor waits for the student to initiate engagement. An instructor prompts the student to respond if needed. An instructor allows access to an item/activity contingent on a correct response from the student. The instructor fades out the prompting process over a period of time and subsequent trials. Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically. Domains [ edit ] Future school (1901 or 1910) Benjamin Bloom has suggested three domains of learning in his taxonomy which are: Cognitive : To recall, calculate, discuss, analyze, problem solve, etc. Psychomotor : To dance, swim, ski, dive, drive a car, ride a bike, etc. Affective : To like something or someone, love, appreciate, fear, hate, worship, etc. These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain). Transfer [ edit ] Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction. Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote. A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content. Factors affecting learning [ edit ] Main article: Evidence-based learning Genetics [ edit ] Main article: Heritability of IQ Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning. Socioeconomic and physical conditions [ edit ] Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions. The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school. Psychological factors and teaching style [ edit ] Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning . The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive. Certain techniques can increase long-term retention: The spacing effect means that lessons or studying spaced out over time ( spaced repetition ) are better than cramming Teaching material to other people "Self-explaining" (paraphrasing material to oneself) rather than passive reading Low-stakes quizzing Epigenetic factors [ edit ] Further information: Epigenetics in learning and memory The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins . During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation. General cognition-related factors [ edit ] This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging Adult learning vs children's learning [ edit ] See also: Aging brain This section needs expansion . You can help by adding to it . ( January 2023 ) Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is. Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development . What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities. In animal evolution [ edit ] Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species. Costs and benefits of learned and innate knowledge [ edit ] In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information. Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning. However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live. In plants [ edit ] In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors. Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways. During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning. While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight. Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning". Machine learning [ edit ] This section needs expansion . You can help by adding to it . ( February 2020 ) Robots can learn to cooperate. Main article: Machine learning Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output. Types [ edit ] Phases [ edit ] See also [ edit ] 21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling Information theory [ edit ] Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence Types of education [ edit ] Autodidacticism – Independent education without the guidance of teachers Andragogy – Methods and principles in adult education Pedagogy – Theory and practice of education References [ edit ] ^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 . Notes [ edit ] Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 . Further reading [ edit ] Library resources about Learning Resources in your library Ulrich Boser (2019). Learn Better: Mastering the Skills for Success in Life, Business, and School, or How to Become an Expert in Just About Anything . Rodale Books. ISBN 978-0593135310 . External links [ edit ] Wikimedia Commons has media related to Learning . Wikiquote has quotations related to Learning . How People Learn: Brain, Mind, Experience, and School (expanded edition) published by the National Academies Press Applying Science of Learning in Education: Infusing Psychological Science into the Curriculum published by the American Psychological Association Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition Authority control databases : National Spain France BnF data Germany Israel United States Czech Republic Retrieved from " https://en.wikipedia.org/w/index.php?title=Learning&oldid=1220741482 "
Process of acquiring new knowledge For the album by Perfume Genius, see Learning (album) . "Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) . American students learning how to make and roll sushi Cognitive psychology Perception Visual perception Object recognition Face recognition Pattern recognition Attention Memory Aging and memory Emotional memory Learning Long-term memory Metacognition Language Metalanguage Thinking Cognition Concept Reasoning Decision making Problem solving Numerical cognition Numerosity adaptation effect Approximate number system Parallel individuation system v t e Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved. Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development . Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity. Types [ edit ] See also: Learning styles and Machine learning § Types of problems and tasks There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory. Non-associative learning [ edit ] Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury. Non-associative learning can be divided into habituation and sensitization . Habituation [ edit ] Main article: Habituation Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization. Sensitization [ edit ] Main article: Sensitization Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism. Active learning [ edit ] Main article: Active learning Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ). Associative learning [ edit ] Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus. Operant conditioning [ edit ] Main article: Operant conditioning Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior. Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning. Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc. Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life. Classical conditioning [ edit ] Main article: Classical conditioning The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants. Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles. Observational learning [ edit ] Main article: Observational learning Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings. Imprinting [ edit ] Main article: Imprinting (psychology) Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds. Play [ edit ] Main article: Play (activity) Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness . Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children. There are five types of play: Sensorimotor play aka functional play, characterized by the repetition of an activity Roleplay occurs starting at the age of three Rule-based play where authoritative prescribed codes of conduct are primary Construction play involves experimentation and building Movement play aka physical play These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others. Enculturation [ edit ] Main article: Enculturation Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one. Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults. Episodic learning [ edit ] Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to. Multimedia learning [ edit ] Main article: Multimedia learning Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory . E-learning and augmented learning [ edit ] Main article: Electronic learning Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones . When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education . Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning: Learner–learner (i.e. communication between and among peers with or without the teacher present), Learner–instructor (i.e. student-teacher communication), and Learner–content (i.e. intellectually interacting with content that results in changes in learners' understanding, perceptions, and cognitive structures). In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance). Rote learning [ edit ] Main article: Rote learning Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion. Meaningful learning [ edit ] See also: Deeper learning Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned. Evidence-based learning [ edit ] Main article: Evidence-based learning Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns. Formal learning [ edit ] Main article: Education A depiction of the world's oldest continually operating university, the University of Bologna , Italy Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition. Non-formal learning [ edit ] Main article: Nonformal learning Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity. Informal learning [ edit ] Main article: Informal learning Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience. Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself. Nonformal learning and combined approaches [ edit ] The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc. To learn a skill, such as solving a Rubik's Cube quickly, several factors come into play at once: Reading directions helps a player learn the patterns that solve the Rubik's Cube. Practicing the moves repeatedly helps build " muscle memory " and speed. Thinking critically about moves helps find shortcuts, which speeds future attempts. Observing the Rubik's Cube's six colors help anchor solutions in the mind. Revisiting the cube occasionally helps retain the skill. Tangential learning [ edit ] Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning. Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay. Dialogic learning [ edit ] Main article: Dialogic learning Dialogic learning is a type of learning based on dialogue. Incidental learning [ edit ] In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set. Here are some steps most commonly used in incidental teaching: An instructor will arrange the learning environment so that necessary materials are within the student's sight, but not within his reach, thus impacting his motivation to seek out those materials. An instructor waits for the student to initiate engagement. An instructor prompts the student to respond if needed. An instructor allows access to an item/activity contingent on a correct response from the student. The instructor fades out the prompting process over a period of time and subsequent trials. Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically. Domains [ edit ] Future school (1901 or 1910) Benjamin Bloom has suggested three domains of learning in his taxonomy which are: Cognitive : To recall, calculate, discuss, analyze, problem solve, etc. Psychomotor : To dance, swim, ski, dive, drive a car, ride a bike, etc. Affective : To like something or someone, love, appreciate, fear, hate, worship, etc. These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain). Transfer [ edit ] Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction. Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote. A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content. Factors affecting learning [ edit ] Main article: Evidence-based learning Genetics [ edit ] Main article: Heritability of IQ Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning. Socioeconomic and physical conditions [ edit ] Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions. The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school. Psychological factors and teaching style [ edit ] Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning . The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive. Certain techniques can increase long-term retention: The spacing effect means that lessons or studying spaced out over time ( spaced repetition ) are better than cramming Teaching material to other people "Self-explaining" (paraphrasing material to oneself) rather than passive reading Low-stakes quizzing Epigenetic factors [ edit ] Further information: Epigenetics in learning and memory The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins . During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation. General cognition-related factors [ edit ] This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging Adult learning vs children's learning [ edit ] See also: Aging brain This section needs expansion . You can help by adding to it . ( January 2023 ) Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is. Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development . What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities. In animal evolution [ edit ] Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species. Costs and benefits of learned and innate knowledge [ edit ] In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information. Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning. However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live. In plants [ edit ] In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors. Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways. During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning. While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight. Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning". Machine learning [ edit ] This section needs expansion . You can help by adding to it . ( February 2020 ) Robots can learn to cooperate. Main article: Machine learning Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output. Types [ edit ] Phases [ edit ] See also [ edit ] 21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling Information theory [ edit ] Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence Types of education [ edit ] Autodidacticism – Independent education without the guidance of teachers Andragogy – Methods and principles in adult education Pedagogy – Theory and practice of education References [ edit ] ^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 . Notes [ edit ] Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 . Further reading [ edit ] Library resources about Learning Resources in your library Ulrich Boser (2019). Learn Better: Mastering the Skills for Success in Life, Business, and School, or How to Become an Expert in Just About Anything . Rodale Books. ISBN 978-0593135310 . External links [ edit ] Wikimedia Commons has media related to Learning . Wikiquote has quotations related to Learning . How People Learn: Brain, Mind, Experience, and School (expanded edition) published by the National Academies Press Applying Science of Learning in Education: Infusing Psychological Science into the Curriculum published by the American Psychological Association Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition Authority control databases : National Spain France BnF data Germany Israel United States Czech Republic
"Learn" and "Learned" redirect here. For other uses, see Learn (disambiguation) and Learned (disambiguation) .
Learning is the process of acquiring new understanding , knowledge , behaviors , skills , values , attitudes , and preferences . The ability to learn is possessed by humans , non-human animals , and some machines ; there is also evidence for some kind of learning in certain plants . Some learning is immediate, induced by a single event (e.g. being burned by a hot stove ), but much skill and knowledge accumulate from repeated experiences. The changes induced by learning often last a lifetime, and it is hard to distinguish learned material that seems to be "lost" from that which cannot be retrieved.
Human learning starts at birth (it might even start before in terms of an embryo's need for both interaction with, and freedom within its environment within the womb. ) and continues until death as a consequence of ongoing interactions between people and their environment. The nature and processes involved in learning are studied in many established fields (including educational psychology , neuropsychology , experimental psychology , cognitive sciences , and pedagogy ), as well as emerging fields of knowledge (e.g. with a shared interest in the topic of learning from safety events such as incidents/accidents , or in collaborative learning health systems ). Research in such fields has led to the identification of various sorts of learning. For example, learning may occur as a result of habituation , or classical conditioning , operant conditioning or as a result of more complex activities such as play , seen only in relatively intelligent animals. Learning may occur consciously or without conscious awareness. Learning that an aversive event cannot be avoided or escaped may result in a condition called learned helplessness . There is evidence for human behavioral learning prenatally , in which habituation has been observed as early as 32 weeks into gestation , indicating that the central nervous system is sufficiently developed and primed for learning and memory to occur very early on in development .
Play has been approached by several theorists as a form of learning. Children experiment with the world, learn the rules, and learn to interact through play. Lev Vygotsky agrees that play is pivotal for children's development, since they make meaning of their environment through playing educational games. For Vygotsky, however, play is the first form of learning language and communication, and the stage where a child begins to understand rules and symbols. This has led to a view that learning in organisms is always related to semiosis , and is often associated with representational systems/activity.
There are various functional categorizations of memory which have developed. Some memory researchers distinguish memory based on the relationship between the stimuli involved (associative vs non-associative) or based to whether the content can be communicated through language (declarative/explicit vs procedural/implicit). Some of these categories can, in turn, be parsed into sub-types. For instance, declarative memory comprises both episodic and semantic memory.
Non-associative learning refers to "a relatively permanent change in the strength of response to a single stimulus due to repeated exposure to that stimulus." This definition exempts the changes caused by sensory adaptation , fatigue , or injury.
Habituation is an example of non-associative learning in which one or more components of an innate response (e.g., response probability, response duration) to a stimulus diminishes when the stimulus is repeated. Thus, habituation must be distinguished from extinction , which is an associative process. In operant extinction, for example, a response declines because it is no longer followed by a reward. An example of habituation can be seen in small song birds—if a stuffed owl (or similar predator ) is put into the cage, the birds initially react to it as though it were a real predator. Soon the birds react less, showing habituation. If another stuffed owl is introduced (or the same one removed and re-introduced), the birds react to it again as though it were a predator, demonstrating that it is only a very specific stimulus that is habituated to (namely, one particular unmoving owl in one place). The habituation process is faster for stimuli that occur at a high rather than for stimuli that occur at a low rate as well as for the weak and strong stimuli, respectively. Habituation has been shown in essentially every species of animal, as well as the sensitive plant Mimosa pudica and the large protozoan Stentor coeruleus . This concept acts in direct opposition to sensitization.
Sensitization is an example of non-associative learning in which the progressive amplification of a response follows repeated administrations of a stimulus . This is based on the notion that a defensive reflex to a stimulus such as withdrawal or escape becomes stronger after the exposure to a different harmful or threatening stimulus. An everyday example of this mechanism is the repeated tonic stimulation of peripheral nerves that occurs if a person rubs their arm continuously. After a while, this stimulation creates a warm sensation that can eventually turn painful. This pain results from a progressively amplified synaptic response of the peripheral nerves. This sends a warning that the stimulation is harmful. Sensitization is thought to underlie both adaptive as well as maladaptive learning processes in the organism.
Active learning occurs when a person takes control of his/her learning experience. Since understanding information is the key aspect of learning, it is important for learners to recognize what they understand and what they do not. By doing so, they can monitor their own mastery of subjects. Active learning encourages learners to have an internal dialogue in which they verbalize understandings. This and other meta-cognitive strategies can be taught to a child over time. Studies within metacognition have proven the value in active learning, claiming that the learning is usually at a stronger level as a result. In addition, learners have more incentive to learn when they have control over not only how they learn but also what they learn. Active learning is a key characteristic of student-centered learning . Conversely, passive learning and direct instruction are characteristics of teacher-centered learning (or traditional education ).
Associative learning is the process by which a person or animal learns an association between two stimuli or events. In classical conditioning , a previously neutral stimulus is repeatedly paired with a reflex-eliciting stimulus until eventually the neutral stimulus elicits a response on its own. In operant conditioning, a behavior that is reinforced or punished in the presence of a stimulus becomes more or less likely to occur in the presence of that stimulus.
Operant conditioning is a way in which behavior can be shaped or modified according to the desires of the trainer or head individual. Operant conditioning uses the thought that living things seek pleasure and avoid pain, and that an animal or human can learn through receiving either reward or punishment at a specific time called trace conditioning. Trace conditioning is the small and ideal period of time between the subject performing the desired behavior, and receiving the positive reinforcement as a result of their performance. The reward needs to be given immediately after the completion of the wanted behavior.
Operant conditioning is different from classical conditioning in that it shapes behavior not solely on bodily reflexes that occur naturally to a specific stimulus, but rather focuses on the shaping of wanted behavior that requires conscious thought, and ultimately requires learning.
Punishment and reinforcement are the two principal ways in which operant conditioning occurs. Punishment is used to reduce unwanted behavior, and ultimately (from the learner's perspective) leads to avoidance of the punishment, not necessarily avoidance of the unwanted behavior. Punishment is not an appropriate way to increase wanted behavior for animals or humans. Punishment can be divided into two subcategories, positive punishment and negative punishment. Positive punishment is when an aversive aspect of life or thing is added to the subject, for this reason it is called positive punishment. For example, the parent spanking their child would be considered a positive punishment, because a spanking was added to the child. Negative punishment is considered the removal of something loved or desirable from the subject. For example, when a parent puts his child in time out, in reality, the child is losing the opportunity to be with friends, or to enjoy the freedom to do as he pleases. In this example, negative punishment is the removal of the child's desired rights to play with his friends etc.
Reinforcement on the other hand is used to increase a wanted behavior either through negative reinforcement or positive reinforcement. Negative reinforcement is defined by removing an undesirable aspect of life, or thing. For example, a dog might learn to sit as the trainer scratches his ears, which ultimately is removing his itches (undesirable aspect). Positive reinforcement is defined by adding a desirable aspect of life or thing. For example, a dog might learn to sit if he receives a treat. In this example the treat was added to the dog's life.
The typical paradigm for classical conditioning involves repeatedly pairing an unconditioned stimulus (which unfailingly evokes a reflexive response) with another previously neutral stimulus (which does not normally evoke the response). Following conditioning, the response occurs both to the unconditioned stimulus and to the other, unrelated stimulus (now referred to as the "conditioned stimulus"). The response to the conditioned stimulus is termed a conditioned response . The classic example is Ivan Pavlov and his dogs. Pavlov fed his dogs meat powder, which naturally made the dogs salivate—salivating is a reflexive response to the meat powder. Meat powder is the unconditioned stimulus (US) and the salivation is the unconditioned response (UR). Pavlov rang a bell before presenting the meat powder. The first time Pavlov rang the bell, the neutral stimulus, the dogs did not salivate, but once he put the meat powder in their mouths they began to salivate. After numerous pairings of bell and food, the dogs learned that the bell signaled that food was about to come, and began to salivate when they heard the bell. Once this occurred, the bell became the conditioned stimulus (CS) and the salivation to the bell became the conditioned response (CR). Classical conditioning has been demonstrated in many species. For example, it is seen in honeybees, in the proboscis extension reflex paradigm. It was recently also demonstrated in garden pea plants.
Another influential person in the world of classical conditioning is John B. Watson . Watson's work was very influential and paved the way for B.F. Skinner 's radical behaviorism. Watson's behaviorism (and philosophy of science) stood in direct contrast to Freud and other accounts based largely on introspection. Watson's view was that the introspective method was too subjective and that we should limit the study of human development to directly observable behaviors. In 1913, Watson published the article "Psychology as the Behaviorist Views", in which he argued that laboratory studies should serve psychology best as a science. Watson's most famous, and controversial, experiment was " Little Albert ", where he demonstrated how psychologists can account for the learning of emotion through classical conditioning principles.
Observational learning is learning that occurs through observing the behavior of others. It is a form of social learning which takes various forms, based on various processes. In humans, this form of learning seems to not need reinforcement to occur, but instead, requires a social model such as a parent, sibling, friend, or teacher with surroundings.
Imprinting is a kind of learning occurring at a particular life stage that is rapid and apparently independent of the consequences of behavior. In filial imprinting, young animals, particularly birds, form an association with another individual or in some cases, an object, that they respond to as they would to a parent. In 1935, the Austrian Zoologist Konrad Lorenz discovered that certain birds follow and form a bond if the object makes sounds.
Play generally describes behavior with no particular end in itself, but that improves performance in similar future situations. This is seen in a wide variety of vertebrates besides humans, but is mostly limited to mammals and birds . Cats are known to play with a ball of string when young, which gives them experience with catching prey. Besides inanimate objects, animals may play with other members of their own species or other animals, such as orcas playing with seals they have caught. Play involves a significant cost to animals, such as increased vulnerability to predators and the risk of injury and possibly infection . It also consumes energy , so there must be significant benefits associated with play for it to have evolved. Play is generally seen in younger animals, suggesting a link with learning. However, it may also have other benefits not associated directly with learning, for example improving physical fitness .
Play, as it pertains to humans as a form of learning is central to a child's learning and development. Through play, children learn social skills such as sharing and collaboration. Children develop emotional skills such as learning to deal with the emotion of anger, through play activities. As a form of learning, play also facilitates the development of thinking and language skills in children.
These five types of play are often intersecting. All types of play generate thinking and problem-solving skills in children. Children learn to think creatively when they learn through play. Specific activities involved in each type of play change over time as humans progress through the lifespan. Play as a form of learning, can occur solitarily, or involve interacting with others.
Enculturation is the process by which people learn values and behaviors that are appropriate or necessary in their surrounding culture . Parents, other adults, and peers shape the individual's understanding of these values. If successful, enculturation results in competence in the language, values, and rituals of the culture. This is different from acculturation , where a person adopts the values and societal rules of a culture different from their native one.
Multiple examples of enculturation can be found cross-culturally. Collaborative practices in the Mazahua people have shown that participation in everyday interaction and later learning activities contributed to enculturation rooted in nonverbal social experience. As the children participated in everyday activities, they learned the cultural significance of these interactions. The collaborative and helpful behaviors exhibited by Mexican and Mexican-heritage children is a cultural practice known as being "acomedido". Chillihuani girls in Peru described themselves as weaving constantly, following behavior shown by the other adults.
Episodic learning is a change in behavior that occurs as a result of an event. For example, a fear of dogs that follows being bitten by a dog is episodic learning. Episodic learning is so named because events are recorded into episodic memory , which is one of the three forms of explicit learning and retrieval, along with perceptual memory and semantic memory . Episodic memory remembers events and history that are embedded in experience and this is distinguished from semantic memory, which attempts to extract facts out of their experiential context or – as some describe – a timeless organization of knowledge. For instance, if a person remembers the Grand Canyon from a recent visit, it is an episodic memory. He would use semantic memory to answer someone who would ask him information such as where the Grand Canyon is. A study revealed that humans are very accurate in the recognition of episodic memory even without deliberate intention to memorize it. This is said to indicate a very large storage capacity of the brain for things that people pay attention to.
Multimedia learning is where a person uses both auditory and visual stimuli to learn information. This type of learning relies on dual-coding theory .
Electronic learning or e-learning is computer-enhanced learning. A specific and always more diffused e-learning is mobile learning (m-learning), which uses different mobile telecommunication equipment, such as cellular phones .
When a learner interacts with the e-learning environment, it is called augmented learning . By adapting to the needs of individuals, the context-driven instruction can be dynamically tailored to the learner's natural environment. Augmented digital content may include text, images, video, audio (music and voice). By personalizing instruction, augmented learning has been shown to improve learning performance for a lifetime. See also minimally invasive education .
Moore (1989) purported that three core types of interaction are necessary for quality, effective online learning:
In his theory of transactional distance, Moore (1993) contented that structure and interaction or dialogue bridge the gap in understanding and communication that is created by geographical distances (known as transactional distance).
Rote learning is memorizing information so that it can be recalled by the learner exactly the way it was read or heard. The major technique used for rote learning is learning by repetition , based on the idea that a learner can recall the material exactly (but not its meaning) if the information is repeatedly processed. Rote learning is used in diverse areas, from mathematics to music to religion.
Meaningful learning is the concept that learned knowledge (e.g., a fact) is fully understood to the extent that it relates to other knowledge. To this end, meaningful learning contrasts with rote learning in which information is acquired without regard to understanding. Meaningful learning, on the other hand, implies there is a comprehensive knowledge of the context of the facts learned.
Evidence-based learning is the use of evidence from well designed scientific studies to accelerate learning. Evidence-based learning methods such as spaced repetition can increase the rate at which a student learns.
Formal learning is a deliberate way attaining of knowledge, which takes place within a teacher-student environment, such as in a school system or work environment. The term formal learning has nothing to do with the formality of the learning, but rather the way it is directed and organized. In formal learning, the learning or training departments set out the goals and objectives of the learning and oftentimes learners will be awarded with a diploma, or a type of formal recognition.
Non-formal learning is organized learning outside the formal learning system. For example, learning by coming together with people with similar interests and exchanging viewpoints, in clubs or in (international) youth organizations, and workshops. From the organizer's point of reference, non-formal learning does not always need a main objective or learning outcome. From the learner's point of view, non-formal learning, although not focused on outcomes, often results in an intentional learning opportunity.
Informal learning is less structured than "non-formal learning". It may occur through the experience of day-to-day situations (for example, one would learn to look ahead while walking because of the possible dangers inherent in not paying attention to where one is going). It is learning from life, during a meal at the table with parents, during play , and while exploring etc.. For the learner, informal learning is most often an experience of happenstance, and not a deliberately planned experience. Thus this does not require enrolment into any class. Unlike formal learning, informal learning typically does not lead to accreditation. Informal learning begins to unfold as the learner ponders his or her situation. This type of learning does not require a professor of any kind, and learning outcomes are unforeseen following the learning experience.
Informal learning is self-directed and because it focuses on day-to-day situations, the value of informal learning can be considered high. As a result, information retrieved from informal learning experiences will likely be applicable to daily life. Children with informal learning can at times yield stronger support than subjects with formal learning in the topic of mathematics. Daily life experiences take place in the workforce, family life, and any other situation that may arise during one's lifetime. Informal learning is voluntary from the learner's viewpoint, and may require making mistakes and learning from them. Informal learning allows the individual to discover coping strategies for difficult emotions that may arise while learning. From the learner's perspective, informal learning can become purposeful, because the learner chooses which rate is appropriate to learn and because this type of learning tends to take place within smaller groups or by oneself.
The educational system may use a combination of formal, informal, and nonformal learning methods. The UN and EU recognize these different forms of learning (cf. links below). In some schools, students can get points that count in the formal-learning systems if they get work done in informal-learning circuits. They may be given time to assist international youth workshops and training courses, on the condition they prepare, contribute, share, and can prove this offered valuable new insight, helped to acquire new skills, a place to get experience in organizing, teaching , etc.
Tangential learning is the process by which people self-educate if a topic is exposed to them in a context that they already enjoy. For example, after playing a music-based video game, some people may be motivated to learn how to play a real instrument, or after watching a TV show that references Faust and Lovecraft, some people may be inspired to read the original work. Self-education can be improved with systematization. According to experts in natural learning, self-oriented learning training has proven an effective tool for assisting independent learners with the natural phases of learning.
Extra Credits writer and game designer James Portnow was the first to suggest games as a potential venue for "tangential learning". Mozelius et al. points out that intrinsic integration of learning content seems to be a crucial design factor, and that games that include modules for further self-studies tend to present good results. The built-in encyclopedias in the Civilization games are presented as an example – by using these modules gamers can dig deeper for knowledge about historical events in the gameplay. The importance of rules that regulate learning modules and game experience is discussed by Moreno, C., in a case study about the mobile game Kiwaka . In this game, developed by Landka in collaboration with ESA and ESO , progress is rewarded with educational content, as opposed to traditional education games where learning activities are rewarded with gameplay.
In incidental teaching learning is not planned by the instructor or the student, it occurs as a byproduct of another activity — an experience, observation, self-reflection, interaction, unique event (e.g. in response to incidents/accidents), or common routine task. This learning happens in addition to or apart from the instructor's plans and the student's expectations. An example of incidental teaching is when the instructor places a train set on top of a cabinet. If the child points or walks towards the cabinet, the instructor prompts the student to say "train". Once the student says "train", he gets access to the train set.
Incidental learning is an occurrence that is not generally accounted for using the traditional methods of instructional objectives and outcomes assessment. This type of learning occurs in part as a product of social interaction and active involvement in both online and onsite courses. Research implies that some un-assessed aspects of onsite and online learning challenge the equivalency of education between the two modalities. Both onsite and online learning have distinct advantages with traditional on-campus students experiencing higher degrees of incidental learning in three times as many areas as online students. Additional research is called for to investigate the implications of these findings both conceptually and pedagogically.
These domains are not mutually exclusive. For example, in learning to play chess , the person must learn the rules (cognitive domain)—but must also learn how to set up the chess pieces and how to properly hold and move a chess piece (psychomotor). Furthermore, later in the game the person may even learn to love the game itself, value its applications in life, and appreciate its history (affective domain).
Transfer of learning is the application of skill, knowledge or understanding to resolve a novel problem or situation that happens when certain conditions are fulfilled. Research indicates that learning transfer is infrequent; most common when "... cued, primed, and guided..." and has sought to clarify what it is, and how it might be promoted through instruction.
Over the history of its discourse, various hypotheses and definitions have been advanced. First, it is speculated that different types of transfer exist, including: near transfer, the application of skill to solve a novel problem in a similar context; and far transfer, the application of skill to solve a novel problem presented in a different context. Furthermore, Perkins and Salomon (1992) suggest that positive transfer in cases when learning supports novel problem solving, and negative transfer occurs when prior learning inhibits performance on highly correlated tasks, such as second or third-language learning. Concepts of positive and negative transfer have a long history; researchers in the early 20th century described the possibility that "...habits or mental acts developed by a particular kind of training may inhibit rather than facilitate other mental activities". Finally, Schwarz, Bransford and Sears (2005) have proposed that transferring knowledge into a situation may differ from transferring knowledge out to a situation as a means to reconcile findings that transfer may both be frequent and challenging to promote.
A significant and long research history has also attempted to explicate the conditions under which transfer of learning might occur. Early research by Ruger, for example, found that the "level of attention", "attitudes", "method of attack" (or method for tackling a problem), a "search for new points of view", a "careful testing of hypothesis" and "generalization" were all valuable approaches for promoting transfer. To encourage transfer through teaching, Perkins and Salomon recommend aligning ("hugging") instruction with practice and assessment, and "bridging", or encouraging learners to reflect on past experiences or make connections between prior knowledge and current content.
Some aspects of intelligence are inherited genetically, so different learners to some degree have different abilities with regard to learning and speed of learning.
Problems like malnutrition , fatigue , and poor physical health can slow learning, as can bad ventilation or poor lighting at home, and unhygienic living conditions.
The design, quality, and setting of a learning space , such as a school or classroom, can each be critical to the success of a learning environment . Size, configuration, comfort—fresh air, temperature, light, acoustics, furniture—can all affect a student's learning. The tools used by both instructors and students directly affect how information is conveyed, from the display and writing surfaces (blackboards, markerboards, tack surfaces) to digital technologies. For example, if a room is too crowded, stress levels rise, student attention is reduced, and furniture arrangement is restricted. If furniture is incorrectly arranged, sightlines to the instructor or instructional material are limited and the ability to suit the learning or lesson style is restricted. Aesthetics can also play a role, for if student morale suffers, so does motivation to attend school.
Intrinsic motivation , such as a student's own intellectual curiosity or desire to experiment or explore, has been found to sustain learning more effectively than extrinsic motivations such as grades or parental requirements. Rote learning involves repetition in order to reinforce facts in memory, but has been criticized as ineffective and " drill and kill " since it kills intrinsic motivation. Alternatives to rote learning include active learning and meaningful learning .
The speed, accuracy, and retention, depend upon aptitude , attitude , interest, attention, energy level, and motivation of the students. Praising students who answer a question properly or give good results should be praised. This encouragement increases their ability and helps them produce better results. Certain attitudes, such as always finding fault in a student's answer or provoking or embarrassing the student in front of a class are counterproductive.
The underlying molecular basis of learning appears to be dynamic changes in gene expression occurring in brain neurons that are introduced by epigenetic mechanisms. Epigenetic regulation of gene expression involves, most notably, chemical modification of DNA or DNA-associated histone proteins. These chemical modifications can cause long-lasting changes in gene expression. Epigenetic mechanisms involved in learning include the methylation and demethylation of neuronal DNA as well as methylation , acetylation and deacetylation of neuronal histone proteins .
During learning, information processing in the brain involves induction of oxidative modification in neuronal DNA followed by the employment of DNA repair processes that introduce epigenetic alterations. In particular, the DNA repair processes of non-homologous end joining and base excision repair are employed in learning and memory formation.
This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ] The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging
This section is an excerpt from Development of the nervous system in humans § Adult neural development . [ edit ]
The nervous system continues to develop during adulthood until brain death . For example: physical exercise has neurobiological effects the consumption of foods (or nutrients ), obesity, alterations of the microbiome , drinks, dietary supplements , recreational drugs and medications may possibly also have effects on the development of the nervous system various diseases, such as COVID-19 , have effects on the development of the nervous system For example, several genes have been identified as to being associated with changes in brain structure over lifetime and are potential Alzheimer's disease therapy-targets. psychological events such as mental trauma and resilience-building exposure to environmental pollution and toxins such as air pollution may have effects on the further development of the nervous system other activities may also have effects on the development of the nervous system, such as lifelong learning , retraining , and types of media- and economic activities broadly, brain aging
Learning is often more efficient in children and takes longer or is more difficult with age . A study using neuroimaging identified rapid neurotransmitter GABA boosting as a major potential explanation-component for why that is.
Children's brains contain more " silent synapses " that are inactive until recruited as part of neuroplasticity and flexible learning or memories . Neuroplasticity is heightened during critical or sensitive periods of brain development, mainly referring to brain development during child development .
What humans learn at the early stages, and what they learn to apply, sets humans on course for life or has a disproportional impact. Adults usually have a higher capacity to select what they learn, to what extent and how. For example, children may learn the given subjects and topics of school curricula via classroom blackboard-transcription handwriting , instead of being able to choose specific topics/skills or jobs to learn and the styles of learning. For instance, children may not have developed consolidated interests, ethics, interest in purpose and meaningful activities, knowledge about real-world requirements and demands, and priorities.
Animals gain knowledge in two ways. First is learning—in which an animal gathers information about its environment and uses this information. For example, if an animal eats something that hurts its stomach, it learns not to eat that again. The second is innate knowledge that is genetically inherited. An example of this is when a horse is born and can immediately walk. The horse has not learned this behavior; it simply knows how to do it. In some scenarios, innate knowledge is more beneficial than learned knowledge. However, in other scenarios the opposite is true—animals must learn certain behaviors when it is disadvantageous to have a specific innate behavior. In these situations, learning evolves in the species.
In a changing environment, an animal must constantly gain new information to survive. However, in a stable environment, this same individual needs to gather the information it needs once, and then rely on it for the rest of its life. Therefore, different scenarios better suit either learning or innate knowledge. Essentially, the cost of obtaining certain knowledge versus the benefit of already having it determines whether an animal evolved to learn in a given situation, or whether it innately knew the information. If the cost of gaining the knowledge outweighs the benefit of having it, then the animal does not evolve to learn in this scenario—but instead, non-learning evolves. However, if the benefit of having certain information outweighs the cost of obtaining it, then the animal is far more likely to evolve to have to learn this information.
Non-learning is more likely to evolve in two scenarios. If an environment is static and change does not or rarely occurs, then learning is simply unnecessary. Because there is no need for learning in this scenario—and because learning could prove disadvantageous due to the time it took to learn the information—non-learning evolves. Similarly, if an environment is in a constant state of change, learning is also disadvantageous, as anything learned is immediately irrelevant because of the changing environment. The learned information no longer applies. Essentially, the animal would be just as successful if it took a guess as if it learned. In this situation, non-learning evolves. In fact, a study of Drosophila melanogaster showed that learning can actually lead to a decrease in productivity, possibly because egg-laying behaviors and decisions were impaired by interference from the memories gained from the newly learned materials or because of the cost of energy in learning.
However, in environments where change occurs within an animal's lifetime but is not constant, learning is more likely to evolve. Learning is beneficial in these scenarios because an animal can adapt to the new situation, but can still apply the knowledge that it learns for a somewhat extended period of time. Therefore, learning increases the chances of success as opposed to guessing. An example of this is seen in aquatic environments with landscapes subject to change. In these environments, learning is favored because the fish are predisposed to learn the specific spatial cues where they live.
In recent years, plant physiologists have examined the physiology of plant behavior and cognition. The concepts of learning and memory are relevant in identifying how plants respond to external cues, a behavior necessary for survival. Monica Gagliano, an Australian professor of evolutionary ecology, makes an argument for associative learning in the garden pea, Pisum sativum . The garden pea is not specific to a region, but rather grows in cooler, higher altitude climates. Gagliano and colleagues' 2016 paper aims to differentiate between innate phototropism behavior and learned behaviors. Plants use light cues in various ways, such as to sustain their metabolic needs and to maintain their internal circadian rhythms. Circadian rhythms in plants are modulated by endogenous bioactive substances that encourage leaf-opening and leaf-closing and are the basis of nyctinastic behaviors.
Gagliano and colleagues constructed a classical conditioning test in which pea seedlings were divided into two experimental categories and placed in Y-shaped tubes. In a series of training sessions, the plants were exposed to light coming down different arms of the tube. In each case, there was a fan blowing lightly down the tube in either the same or opposite arm as the light. The unconditioned stimulus (US) was the predicted occurrence of light and the conditioned stimulus (CS) was the wind blowing by the fan. Previous experimentation shows that plants respond to light by bending and growing towards it through differential cell growth and division on one side of the plant stem mediated by auxin signaling pathways.
During the testing phase of Gagliano's experiment, the pea seedlings were placed in different Y-pipes and exposed to the fan alone. Their direction of growth was subsequently recorded. The 'correct' response by the seedlings was deemed to be growing into the arm where the light was "predicted" from the previous day. The majority of plants in both experimental conditions grew in a direction consistent with the predicted location of light based on the position of the fan the previous day. For example, if the seedling was trained with the fan and light coming down the same arm of the Y-pipe, the following day the seedling grew towards the fan in the absence of light cues despite the fan being placed in the opposite side of the Y-arm. Plants in the control group showed no preference to a particular arm of the Y-pipe. The percentage difference in population behavior observed between the control and experimental groups is meant to distinguish innate phototropism behavior from active associative learning.
While the physiological mechanism of associative learning in plants is not known, Telewski et al. describes a hypothesis that describes photoreception as the basis of mechano-perception in plants. One mechanism for mechano-perception in plants relies on MS ion channels and calcium channels. Mechanosensory proteins in cell lipid bilayers, known as MS ion channels, are activated once they are physically deformed in response to pressure or tension. Ca2+ permeable ion channels are "stretch-gated" and allow for the influx of osmolytes and calcium, a well-known second messenger, into the cell. This ion influx triggers a passive flow of water into the cell down its osmotic gradient, effectively increasing turgor pressure and causing the cell to depolarize. Gagliano hypothesizes that the basis of associative learning in Pisum sativum is the coupling of mechanosensory and photosensory pathways and is mediated by auxin signaling pathways. The result is directional growth to maximize a plant's capture of sunlight.
Gagliano et al. published another paper on habituation behaviors in the mimosa pudica plant whereby the innate behavior of the plant was diminished by repeated exposure to a stimulus. There has been controversy around this paper and more generally around the topic of plant cognition. Charles Abrahmson, a psychologist and behavioral biologist, says that part of the issue of why scientists disagree about whether plants have the ability to learn is that researchers do not use a consistent definition of "learning" and "cognition". Similarly, Michael Pollan, an author, and journalist, says in his piece The Intelligent Plant that researchers do not doubt Gagliano's data but rather her language, specifically her use of the term "learning" and "cognition" with respect to plants. A direction for future research is testing whether circadian rhythms in plants modulate learning and behavior and surveying researchers' definitions of "cognition" and "learning".
Machine learning, a branch of artificial intelligence , concerns the construction and study of systems that can learn from data. For example, a machine learning system could be trained on email messages to learn to distinguish between spam and non-spam messages. Most of the Machine Learning models are based on probabilistic theories where each input (e.g. an image ) is associated with a probability to become the desired output.
21st century skills – Skills identified as being required for success in the 21st century Anticipatory socialization – Process in which people take on the values of groups that they aspire to join Epistemology – Branch of philosophy concerning knowledge Implicit learning – in learning psychology Pages displaying wikidata descriptions as a fallback Instructional theory – Theory that offers explicit guidance on how to better help people learn and develop Learning sciences – Interdisciplinary field to further scientific understanding of learning Lifelong learning – Ongoing, voluntary, and self-motivated pursuit of knowledge Living educational theory Media psychology – Area of psychology Subgoal labeling
Algorithmic information theory – Subfield of information theory and computer science Algorithmic probability – mathematical method of assigning a prior probability to a given observation Pages displaying wikidata descriptions as a fallback Bayesian inference – Method of statistical inference Inductive logic programming – learning logic programs from data Pages displaying wikidata descriptions as a fallback Inductive probability – Determining the probability of future events based on past events Information theory – Scientific study of digital information Minimum description length – Model selection principle Minimum message length – Formal information theory restatement of Occam's Razor Occam's razor – Philosophical problem-solving principle Solomonoff's theory of inductive inference – mathematical formalization of Occam's razor that, assuming the world is generated by a computer program, the most likely one is the shortest, using Bayesian inference Pages displaying wikidata descriptions as a fallback AIXI – Mathematical formalism for artificial general intelligence
^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 .
^ Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 . ^ Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine . ^ Lakoff, G., & Johnson, M. (2008). Metaphors we live by . University of Chicago press. ^ Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 . ^ OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 . ^ Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York. ^ Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121. ^ Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 . ^ "Jungle Gyms: The Evolution of Animal Play" . Archived from the original on October 11, 2007. ^ "What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 . ^ Learned helplessness at the Encyclopædia Britannica ^ Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 . ^ Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 . ^ Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381. ^ Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49. ^ Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 . ^ "Non-associative Learning" (PDF) . Archived from the original (PDF) on 2014-01-03 . Retrieved 2013-08-09 . ^ Pear, Joseph (2014). The Science of Learning . London: Psychology Press. p. 15. ISBN 978-1-317-76280-5 . ^ Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 . ^ Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 . ^ Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior (2nd ed.). New York: Oxford. ^ Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 . ^ Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 . ^ Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 . ^ Bransford, 2000, pp. 15–20 ^ J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16. ^ Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 . ^ Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 . ^ "Reflex Definition & Meaning | Britannica Dictionary" . www.britannica.com . Retrieved 2023-06-30 . ^ Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 . ^ Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 . ^ Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 . ^ Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 . ^ Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing. ^ Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 . ^ Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547. ^ Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 . ^ Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology . ^ Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 . ^ Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc. ^ Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd. ^ Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 . ^ Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 . ^ Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 . ^ ( Mayer 2001 ) ^ ( Paivio 1971 ) ^ Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning ^ Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 . ^ Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge ^ Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 . ^ Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 . ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 . ^ "Glossary" . CEDEFOP . Retrieved 2023-06-24 . ^ Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999 ^ "What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 . ^ Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 . ^ Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 . ^ Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29 ^ Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 . ^ J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19. ^ Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 . ^ Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017). ^ Moreno, Carlos (2014). "Kiwaka | Kiwaka Story (by LANDKA ®)" (PDF) . LifePlay . 3 . ^ European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 . ^ Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 . ^ "What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 . ^ Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 . ^ "Bloom's Taxonomy" . www.businessballs.com . Retrieved 4 May 2018 . ^ Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 . ^ Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26. ^ Perkins, D.N.; Salomon, G. (1992). "Transfer of Learning". International Encyclopedia of Education . 2 . ^ Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 . ^ Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15. ^ Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2). ^ Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 . ^ Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 . ^ New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19 ^ Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 . ^ Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 . ^ The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser) ^ Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . ^ Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 . ^ Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 . ^ Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 . ^ Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 . ^ "Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 . ^ Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 . ^ "Brain scans shed light on how kids learn faster than adults" . UPI . Retrieved 17 December 2022 . ^ Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 . ^ Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 . ^ Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 . ^ Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 . ^ Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 . ^ < Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . > ^ Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 . ^ Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x . ^ Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar. ^ Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 . ^ Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 . ^ Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 . ^ Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 .
Richard Gross, Psychology: The Science of Mind and Behaviour Archived 2022-12-31 at the Wayback Machine 6E, Hachette UK, ISBN 978-1-4441-6436-7 .
Karban, R. (2015). Plant Learning and Memory. In: Plant Sensing and Communication . Chicago and London: The University of Chicago Press, pp. 31–44, [1] Archived 2022-12-31 at the Wayback Machine .
Daniel L. Schacter; Daniel T. Gilbert; Daniel M. Wegner (2011) [2009]. Psychology, 2nd edition . Worth Publishers. p. 264 . ISBN 978-1-4292-3719-2 .
OECD (2007). Understanding the Brain: The Birth of a Learning Science . OECD Publishing. p. 165. ISBN 978-92-64-02913-2 .
Chapter 2: The Montessori philosophy. From Lillard, P. P. Lillard (1972). Montessori: A Modern Approach. Schocken Books, New York.
Sujan, M. A., Huang, H., & Braithwaite, J. (2017). Learning from incidents in health care: critique from a Safety-II perspective. Safety Science , 99 , 115–121.
Hartley, David M.; Seid, Michael (2021). "Collaborative learning health systems: Science and practice" . Learning Health Systems . 5 (3): e10286. doi : 10.1002/lrh2.10286 . PMC 8278439 . PMID 34277947 .
"What behavior can we expect of octopuses?" . www.thecephalopodpage.org . The Cephalopod Page. Archived from the original on 5 October 2017 . Retrieved 4 May 2018 .
Sandman, Wadhwa; Hetrick, Porto; Peeke (1997). "Human fetal heart rate dishabituation between thirty and thirty-two weeks gestation". Child Development . 68 (6): 1031–1040. doi : 10.1111/j.1467-8624.1997.tb01982.x . PMID 9418223 .
Sheridan, Mary; Howard, Justine; Alderson, Dawn (2010). Play in Early Childhood: From Birth to Six Years . Oxon: Routledge. ISBN 978-1-136-83748-7 .
Campbell, Cary; Olteanu, Alin; Kull, Kalevi 2019. Learning and knowing as semiosis: Extending the conceptual apparatus of semiotics Archived 2022-04-09 at the Wayback Machine . Sign Systems Studies 47(3/4): 352–381.
Hutchins, E., 2014. The cultural ecosystem of human cognition. Philosophical Psychology 27(1), 34–49.
Fuentes, Agustín (2017). The International Encyclopedia of Primatology, 3 Volume Set . Malden, MA: Wiley Blackwell. p. 712. ISBN 978-0-470-67337-9 .
Gagliano, M.; et al. (2014). "Experience teaches plants to learn faster and forget slower in environments where it matters". Oecologia . 175 (1): 63–72. Bibcode : 2014Oecol.175...63G . doi : 10.1007/s00442-013-2873-7 . PMID 24390479 . S2CID 5038227 .
Wood, D.C. (1988). "Habituation in Stentor produced by mechanoreceptor channel modification" . Journal of Neuroscience . 8 (7): 2254–8. doi : 10.1523/JNEUROSCI.08-07-02254.1988 . PMC 6569508 . PMID 3249223 .
Galizia, Giovanni; Lledo, Pierre-Marie (2013). Neurosciences – From Molecule to Behavior . Heidelberg: Springer Spektrum. p. 578. ISBN 978-3-642-10768-9 .
Woolf, Clifford J. (2018-02-27). "Pain amplification-A perspective on the how, why, when, and where of central sensitization" . Journal of Applied Biobehavioral Research . 23 (2): e12124. doi : 10.1111/jabr.12124 . ISSN 1071-2089 .
Bonne, Omer; Grillon, Christian; Vythilingam, Meena; Neumeister, Alexander; Charney, Dennis S (March 2004). "Adaptive and maladaptive psychobiological responses to severe psychological stress: implications for the discovery of novel pharmacotherapy" . Neuroscience & Biobehavioral Reviews . 28 (1): 65–94. doi : 10.1016/j.neubiorev.2003.12.001 . ISSN 0149-7634 . PMID 15036934 . S2CID 23745725 .
J. Scott Armstrong (2012). "Natural Learning in Higher Education" . Encyclopedia of the Sciences of Learning . Archived from the original on 2014-09-16.
Plotnik, Rod; Kouyomdijan, Haig (2012). Discovery Series: Introduction to Psychology . Belmont, CA: Wadsworth Cengage Learning. p. 208. ISBN 978-1-111-34702-4 .
Bangasser, Debra A.; Waxler, David E.; Santollo, Jessica; Shors, Tracey J. (2006-08-23). "Trace Conditioning and the Hippocampus: The Importance of Contiguity" . The Journal of Neuroscience . 26 (34): 8702–8706. doi : 10.1523/JNEUROSCI.1742-06.2006 . ISSN 0270-6474 . PMC 3289537 . PMID 16928858 .
Pryor, Karen (1999-08-03). Don't Shoot the Dog: The New Art of Teaching and Training (Revised ed.). New York: Bantam. ISBN 978-0-553-38039-2 .
Chance, Paul; Furlong, Ellen (2022-03-16). Learning and Behavior: Active Learning Edition (8th ed.). Boston, MA: Cengage Learning. ISBN 978-0-357-65811-6 .
Bitterman; et al. (1983). "Classical Conditioning of Proboscis Extension in Honeybees ( Apis mellifera )". J. Comp. Psychol . 97 (2): 107–119. doi : 10.1037/0735-7036.97.2.107 . PMID 6872507 .
Gagliano, Monica; Vyazovskiy, Vladyslav V.; Borbély, Alexander A.; Grimonprez, Mavra; Depczynski, Martial (2016-12-02). "Learning by Association in Plants" . Scientific Reports . 6 (1): 38427. Bibcode : 2016NatSR...638427G . doi : 10.1038/srep38427 . ISSN 2045-2322 . PMC 5133544 . PMID 27910933 .
Lillemyr, O.F. (2009). Taking play seriously. Children and play in early childhood education: an exciting challenge. Charlotte, NC: Information Age Publishing.
Whitebread, D.; Coltman, P.; Jameson, H.; Lander, R. (2009). "Play, cognition and self-regulation: What exactly are children learning when they learn through play?" . Educational and Child Psychology . 26 (2): 40–52. doi : 10.53841/bpsecp.2009.26.2.40 . S2CID 150255306 .
Grusec, Joan E.; Hastings, Paul D. "Handbook of Socialization: Theory and Research", 2007, Guilford Press; ISBN 1-59385-332-7 , 978-1-59385-332-7 ; at p. 547.
Paradise, Ruth (1994). "Interactional Style and Nonverbal Meaning: Mazahua Children Learning How to Be Separate-But-Together". Anthropology & Education Quarterly . 25 (2): 156–172. doi : 10.1525/aeq.1994.25.2.05x0907w . S2CID 146505048 .
Lopez, Angelica; Najafi, Behnosh; Rogoff, Barbara; Mejia-Arauz, Rebeca (2012). "Collaboration and helping as cultural practices". The Oxford Handbook of Culture and Psychology .
Bolin, Inge (2006). Growing Up in a Culture of Respect: Childrearing in highland Peru (2 ed.). Austin: University of Texas. pp. 90–99. ISBN 978-0-292-71298-0 .
Terry, W.S. (2006). Learning and Memory: Basic principles, processes, and procedures. Boston: Pearson Education, Inc.
Baars, B.J. & Gage, N.M. (2007). Cognition, Brain, and Consciousness: Introduction to cognitive neuroscience. London: Elsevier Ltd.
Lovett, Marsha; Schunn, Christian; Lebiere, Christian; Munro, Paul (2004). Sixth International Conference on Cognitive Modeling: ICCM – 2004 . Mahwah, NJ: Lawrence Erlbaum Associates Publishers. p. 220. ISBN 978-0-8058-5426-8 .
Chrisley, Ronald; Begeer, Sander (2000). Artificial Intelligence: Critical Concepts, Volume 1 . London: Routledge. p. 48. ISBN 978-0-415-19332-0 .
Gage, Nicole; Baars, Bernard (2018). Fundamentals of Cognitive Neuroscience: A Beginner's Guide . London: Academic Press. p. 219. ISBN 978-0-12-803813-0 .
Augmented Learning Archived 2020-03-13 at the Wayback Machine , Augmented Learning: Context-Aware Mobile Augmented Reality Architecture for Learning
Moore, M (1989). "Three types of interaction". American Journal of Distance Education . 3 (2): 1–6. CiteSeerX 10.1.1.491.4800 . doi : 10.1080/08923648909526659 .
Moore, M.G. (1993). Theory of transactional distance. In D. Keegan (Ed.), Theoretical principles of distance education (pp. 22–38). London and New York: Routledge
Hassard, Jack. "Backup of Meaningful Learning Model" . Archived from the original on 29 October 2011 . Retrieved 30 November 2011 .
Smolen, Paul; Zhang, Yili; Byrne, John H. (25 January 2016). "The right time to learn: mechanisms and optimization of spaced learning" . Nature Reviews Neuroscience . 17 (2): 77–88. arXiv : 1606.08370 . Bibcode : 2016arXiv160608370S . doi : 10.1038/nrn.2015.18 . PMC 5126970 . PMID 26806627 .
"What is the difference between "informal" and "non formal" learning?" . 2014-10-15. Archived from the original on 2014-10-15 . Retrieved 2023-05-03 .
Bell, J., and Dale, M., " Informal Learning in the Workplace" Archived 2013-01-21 at the Wayback Machine , Department for Education and Employment Research Report No. 134. London, England: Department for Education and Employment , August 1999
"What is the difference between "informal" and "non formal" learning?" . 2014-10-16. Archived from the original on 2014-10-16 . Retrieved 2023-06-22 .
Kyndt, Eva; Baert, Herman (June 2013). "Antecedents of Employees' Involvement in Work-Related Learning: A Systematic Review" . Review of Educational Research . 83 (2): 273–313. doi : 10.3102/0034654313478021 . ISSN 0034-6543 . S2CID 145446612 .
Decius, Julian; Schaper, Niclas; Seifert, Andreas (December 2019). "Informal workplace learning: Development and validation of a measure" . Human Resource Development Quarterly . 30 (4): 495–535. doi : 10.1002/hrdq.21368 . ISSN 1044-8004 . S2CID 201376378 .
Dunst, Carl J.; Hamby, Deborah W.; Wilkie, Helen; Dunst, Kerran Scott (2017), Phillipson, Sivanes; Gervasoni, Ann; Sullivan, Peter (eds.), "Meta-Analysis of the Relationship Between Home and Family Experiences and Young Children's Early Numeracy Learning" , Engaging Families as Children's First Mathematics Educators , Early Mathematics Learning and Development, Singapore: Springer Singapore, pp. 105–125, doi : 10.1007/978-981-10-2553-2_7 , ISBN 978-981-10-2551-8 , retrieved 2023-06-29
Tangential Learning "Penny Arcade – PATV – Tangential Learning" . Archived from the original on 2012-01-04 . Retrieved 2012-01-31 .
J. Scott Armstrong (1979). "The Natural Learning Project" . Journal of Experiential Learning and Simulation . 1 : 5–12. Archived from the original on 2014-10-19.
Robert, Rath (2015-01-22). "Game Criticism as Tangential Learning Facilitator: The Case of Critical Intel" . Journal of Games Criticism . 2 (1). Archived from the original on 2023-04-19 . Retrieved 2018-06-08 .
Mozelius; et al. "Motivating Factors and Tangential Learning for Knowledge Acquisition in Educational Games" (PDF) . The Electronic Journal of e-Learning . 15 (4 2017).
European Southern Observatory. "New App Kiwaka Features ESO Material" . www.eso.org . Retrieved 2018-06-10 .
Landka (2014). "Kiaka Press Release" (PDF) . landka.com/documents/10/Kiwaka-PressRelease.pdf . Archived from the original (PDF) on 2020-08-03 . Retrieved 2018-06-10 .
"What is incidental teaching?" . North Shore Pediatric Therapy, Illinois . 2017. Archived from the original on August 29, 2017 . Retrieved August 29, 2017 .
Konetes, George (2011). The Effects of Distance Education and Student Involvement on Incidental Learning (PDF) (PhD dissertation). Indiana University of Pennsylvania. p. 115. ERIC ED535973 ProQuest 909895728 . Archived from the original (PDF) on 2014-07-14 . Retrieved 2014-07-12 .
Perkins, D.N.; Salomon, G. (Jan–Feb 1989). "Are Cognitive Skills Context-Bound?". Educational Researcher . 18 (1): 16–25 [19]. doi : 10.3102/0013189x018001016 . S2CID 15890041 .
Committee on Developments in the Science of Learning with additional material from the Committee on Learning Research (2000). Chapter 3. Learning and Transfer. How People Learn: Brain, Mind, Experience, and School: Expanded Edition . The National Academies Press. doi : 10.17226/9853 . ISBN 978-0-309-07036-2 . Archived from the original on 2013-04-26.
Rogers, Agnes L. (1916). "The Bearing of the New Psychology upon the Teaching of Mathematics". Teacher's College Record . 17 (4): 344–352. doi : 10.1177/016146811601700413 . S2CID 251487440 .
Schwartz, Daniel L.; Bransford, John D.; Sears, David (2005). "Efficiency and innovation in transfer". Transfer of Learning from a Modern Multidisciplinary Perspective : 1–15.
Ruger, Henry Alfred (1910). "The psychology of efficiency: an experimental study of the processes involved in the solution of mechanical puzzles and in the acquisition of skill in their manipulation". Science Press . 19 (2).
Mangal, S.K. (2007). Essentials of Educational Psychology . PHI Learning Pvt. Ltd. p. 736. ISBN 978-81-203-3055-9 .
Aggarwal, J.C (2009). Essentials Of Educational Psychology (Second ed.). Vikas Publishing House Pvt Ltd. p. 596. ISBN 978-81-259-2292-6 .
New Teachers: Designing Learning Environments , May 7, 2015 Archived March 28, 2016, at the Wayback Machine . Retrieved 2016-03-19
A Place for Learning: The Physical Environment of Classrooms , Mark Phillips, May 20, 2014 Archived March 13, 2016, at the Wayback Machine . Retrieved 2016-03-19
Mangal, SK (2002). Advanced Educational Psychology (Second ed.). PHI Learning Pvt. Ltd. p. 536. ISBN 978-81-203-2038-3 .
Bhatia, H.R (1973). Elements Of Educational Psychology . Orient Blackswan. p. 558. ISBN 978-81-250-0029-7 .
The Science Of Learning Archived 2022-05-17 at the Wayback Machine – April 11, 2017 (podcast interview with Ulrich Boser)
Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 . Erratum in: Li, X; Marshall, PR; Leighton, LJ; Zajaczkowski, EL; Wang, Z; Madugalle, SU; Yin, J; Bredy, TW; Wei, W (2019). "The DNA Repair-Associated Protein Gadd45γ Regulates the Temporal Coding of Immediate Early Gene Expression within the Prelimbic Prefrontal Cortex and Is Required for the Consolidation of Associative Fear Memory" . J Neurosci . 39 (6): 970–983. doi : 10.1523/JNEUROSCI.2024-18.2018 . PMC 6363930 . PMID 30545945 .
Brito, David V.C.; Kupke, Janina; Gulmez Karaca, Kubra; Zeuch, Benjamin; Oliveira, Ana M.M. (2020). "Mimicking Age-Associated Gadd45γ Dysregulation Results in Memory Impairments in Young Adult Mice" . J Neurosci . 40 (6): 1197–1210. doi : 10.1523/JNEUROSCI.1621-19.2019 . PMC 7002144 . PMID 31826946 .
Dye, Louise; Boyle, Neil Bernard; Champ, Claire; Lawton, Clare (November 2017). "The relationship between obesity and cognitive health and decline" . The Proceedings of the Nutrition Society . 76 (4): 443–454. doi : 10.1017/S0029665117002014 . ISSN 1475-2719 . PMID 28889822 . S2CID 34630498 .
Spindler, Carolin; Mallien, Louisa; Trautmann, Sebastian; Alexander, Nina; Muehlhan, Markus (27 January 2022). "A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder" . Translational Psychiatry . 12 (1): 40. doi : 10.1038/s41398-022-01809-0 . ISSN 2158-3188 . PMC 8795454 . PMID 35087021 . S2CID 246292525 .
Wollman, Scott C.; Alhassoon, Omar M.; Hall, Matthew G.; Stern, Mark J.; Connors, Eric J.; Kimmel, Christine L.; Allen, Kenneth E.; Stephan, Rick A.; Radua, Joaquim (September 2017). "Gray matter abnormalities in opioid-dependent patients: A neuroimaging meta-analysis". The American Journal of Drug and Alcohol Abuse . 43 (5): 505–517. doi : 10.1080/00952990.2016.1245312 . ISSN 1097-9891 . PMID 27808568 . S2CID 4775912 .
"Genetic 'hotspots' that speed up and slow down brain aging could provide new targets for Alzheimer's drugs" . University of Southern California . Retrieved 15 May 2022 .
Brouwer, Rachel M.; Klein, Marieke; Grasby, Katrina L.; Schnack, Hugo G.; et al. (April 2022). "Genetic variants associated with longitudinal changes in brain structure across the lifespan" . Nature Neuroscience . 25 (4): 421–432. doi : 10.1038/s41593-022-01042-4 . ISSN 1546-1726 . PMC 10040206 . PMID 35383335 . S2CID 247977288 .
Frank, Sebastian M.; Becker, Markus; Qi, Andrea; Geiger, Patricia; Frank, Ulrike I.; Rosedahl, Luke A.; Malloni, Wilhelm M.; Sasaki, Yuka; Greenlee, Mark W.; Watanabe, Takeo (5 December 2022). "Efficient learning in children with rapid GABA boosting during and after training" . Current Biology . 32 (23): 5022–5030.e7. bioRxiv 10.1101/2022.01.02.474022 . doi : 10.1016/j.cub.2022.10.021 . ISSN 0960-9822 . PMID 36384138 . S2CID 253571891 .
Lloreda, Claudia López (16 December 2022). "Adult mouse brains are teeming with 'silent synapses' " . Science News . Retrieved 18 December 2022 .
Vardalaki, Dimitra; Chung, Kwanghun; Harnett, Mark T. (December 2022). "Filopodia are a structural substrate for silent synapses in adult neocortex" . Nature . 612 (7939): 323–327. Bibcode : 2022Natur.612..323V . doi : 10.1038/s41586-022-05483-6 . ISSN 1476-4687 . PMID 36450984 . S2CID 254122483 . University press release: Trafton, Anne. "Silent synapses are abundant in the adult brain" . Massachusetts Institute of Technology via medicalxpress.com . Retrieved 18 December 2022 .
Ismail, Fatima Yousif; Fatemi, Ali; Johnston, Michael V. (1 January 2017). "Cerebral plasticity: Windows of opportunity in the developing brain". European Journal of Paediatric Neurology . 21 (1): 23–48. doi : 10.1016/j.ejpn.2016.07.007 . ISSN 1090-3798 . PMID 27567276 .
Buxton, Alex (10 February 2016). "What Happens in the Brain When Children Learn?" . Neuroscience News . Retrieved 11 January 2023 .
< Aimee Sue Dunlap-Lehtilä. Change and Reliability in the Evolution of Learning and Memory (PDF) (PhD). University of Minnesota. Archived from the original (PDF) on 2013-11-13 . Retrieved 2013-12-15 . >
Mery, Frederic; Kawecki, Tadeusz J. (2004). "An operating cost of learning in Drosophila melanogaster" (PDF) . Animal Behaviour . 68 (3): 589–598. doi : 10.1016/j.anbehav.2003.12.005 . S2CID 53168227 .
Odling-Smee, L.; Braithwaite, V.A. (2003). "The role of learning in fish orientation". Fish and Fisheries . 4 (3): 235–246. doi : 10.1046/j.1467-2979.2003.00127.x .
Ueda, Minoru (2007). "Endogenous factors involved in the regulation of movement and "memory" in plants" (PDF) . Pure Appl. Chem . 79 (4): 519–527. doi : 10.1351/pac200779040519 . S2CID 35797968 . Archived from the original (PDF) on 2019-06-06 – via Semantic Scholar.
Liscum, Emmanuel (January 2014). "Phototropism: Growing towards an Understanding of Plant Movement" . Plant Cell . 1 (1): 38–55. doi : 10.1105/tpc.113.119727 . PMC 3963583 . PMID 24481074 .
Telewski, FW (October 2006). "A unified hypothesis of mechanoreception in plants" . American Journal of Botany . 93 (10): 1466–76. doi : 10.3732/ajb.93.10.1466 . PMID 21642094 .
Abramson, Charles I.; Chicas-Mosier, Ana M. (2016-03-31). "Learning in Plants: Lessons from Mimosa pudica" . Frontiers in Psychology . 7 : 417. doi : 10.3389/fpsyg.2016.00417 . ISSN 1664-1078 . PMC 4814444 . PMID 27065905 .
Pollan, Michael (2013-12-16). "The Intelligent Plant" . The New Yorker . ISSN 0028-792X . Retrieved 2019-06-06 .
Mayer, R.E. (2001). Multimedia learning . New York: Cambridge University Press. ISBN 978-0-521-78749-9 . Paivio, A. (1971). Imagery and verbal processes . New York: Holt, Rinehart, and Winston. ISBN 978-0-03-085173-5 .
Links to related articles v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition
v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition
v t e Ethology Branches Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology Ethologists Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman Societies Association for the Study of Animal Behaviour International Society for Applied Ethology Journals Animal Behaviour Animal Cognition Animal Welfare Behavioral Ecology Behaviour Category
Animal cognition Animal communication Animal consciousness Animal culture Animal sexual behaviour Animal welfare science Anthrozoology Bee learning and communication Behavioural ecology Behavioral endocrinology Behavioural genetics Breed Cognitive ethology Comparative psychology Emotion in animals Evolutionary neuroscience Feeding Hover Human ethology Instinct Learning Neuroethology Pain in animals Philosophical ethology Sociobiology Stereotypy Structures Hive Nest Instinct Swarm Tool use by non-humans Zoosemiotics Zoomusicology
Patrick Bateson Marc Bekoff Donald Broom John B. Calhoun Charles Darwin Marian Dawkins Richard Dawkins Irenäus Eibl-Eibesfeldt Dian Fossey Karl von Frisch Jane Goodall Heini Hediger Julian Huxley Konrad Lorenz Desmond Morris Thomas Sebeok William Homan Thorpe Nikolaas Tinbergen Jakob von Uexküll Wolfgang Wickler E. O. Wilson Solly Zuckerman
v t e Human intelligence topics Types Collective Emotional Intellectual Linguistic Multiple Social Spatial ( visuospatial ) Abilities, traits, and constructs Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing Models and theories Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory Areas of research Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex Outline of human intelligence / thought
Cognition Cognitive liberty Communication Creativity Fluid and crystallized intelligence g factor Intelligence quotient Knowledge Learning Memory Problem solving Reasoning Thought ( abstraction ) Understanding Visual processing
Cattell–Horn–Carroll theory Fluid and crystallized intelligence Multiple-intelligences theory PASS theory Three-stratum theory Triarchic theory
Evolution of human intelligence Heritability of IQ Psychometrics Intelligence and environment / fertility / height / health / longevity / neuroscience / personality / race / sex
v t e Learning Non-associative learning Habituation Sensitization Associative learning Classical conditioning Imprinting Observational learning Operant conditioning Insight learning Abductive reasoning Deductive reasoning Inductive reasoning
v t e Mental processes Cognition Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving Perception Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual Memory Consolidation Encoding Storage Recall Other Attention Higher nervous activity Intention Learning Mental fatigue Mental set Thinking Volition
Awareness Cognitive liberty Comprehension Consciousness Critical thinking Decision-making Imagination Intuition Problem solving
Amodal Color RGB model Depth Form Haptic ( Touch ) Perception as interpretation Peripheral Social Sound Harmonics Pitch Speech Visual
Categories : Learning Memorization Cognitive science Developmental psychology Intelligence Neuropsychological assessment Systems science Hidden categories: Webarchive template wayback links Articles with Encyclopædia Britannica links Articles with short description Short description is different from Wikidata Wikipedia articles needing clarification from January 2011 All articles with unsourced statements Articles with unsourced statements from March 2016 Articles with unsourced statements from March 2022 Wikipedia articles needing factual verification from March 2022 Articles with excerpts Articles to be expanded from January 2023 All articles to be expanded Articles using small message boxes Articles to be expanded from February 2020 Pages displaying wikidata descriptions as a fallback via Module:Annotated link Commons category link from Wikidata Articles with BNE identifiers Articles with BNF identifiers Articles with BNFdata identifiers Articles with GND identifiers Articles with J9U identifiers Articles with LCCN identifiers Articles with NKC identifiers Articles containing video clips
Categories : Learning Memorization Cognitive science Developmental psychology Intelligence Neuropsychological assessment Systems science
Hidden categories: Webarchive template wayback links Articles with Encyclopædia Britannica links Articles with short description Short description is different from Wikidata Wikipedia articles needing clarification from January 2011 All articles with unsourced statements Articles with unsourced statements from March 2016 Articles with unsourced statements from March 2022 Wikipedia articles needing factual verification from March 2022 Articles with excerpts Articles to be expanded from January 2023 All articles to be expanded Articles using small message boxes Articles to be expanded from February 2020 Pages displaying wikidata descriptions as a fallback via Module:Annotated link Commons category link from Wikidata Articles with BNE identifiers Articles with BNF identifiers Articles with BNFdata identifiers Articles with GND identifiers Articles with J9U identifiers Articles with LCCN identifiers Articles with NKC identifiers Articles containing video clips
This page was last edited on 25 April 2024, at 16:57 (UTC) . Text is available under the Creative Commons Attribution-ShareAlike License 4.0 ; additional terms may apply. By using this site, you agree to the Terms of Use and Privacy Policy . Wikipedia® is a registered trademark of the Wikimedia Foundation, Inc. , a non-profit organization. Privacy policy About Wikipedia Disclaimers Contact Wikipedia Code of Conduct Developers Statistics Cookie statement Mobile view | biology | 85278 | https://da.wikipedia.org/wiki/Taksonomi%20for%20indl%C3%A6ringsm%C3%A5l | Taksonomi for indlæringsmål | En taksonomi for indlæringsmål er en klassifikation af indlæringsmål/læringsmål. Den mest kendte taksonomi blev oprindeligt foreslået af Benjamin Bloom, som arbejdede med affektive, psykomotoriske og kognitive områder.
Andre skelner mellem faglige, generiske og holdningsmæssige læringsmål.
Affektiv
Det affektive område handler om, hvorledes folk reagerer følelsesmæssigt – om evnen til at føle andres smerte eller glæde, men også om motivation, holdninger og værdier.
Psykomotorisk
Det psykomotoriske område handler om evnen til at agere fysisk – eksempelvis at bruge en hammer eller gennemføre gymnastiske øvelser.
Han inddeler i 5 niveauer, hvor 1 er det laveste
Imitation (copy)
Manipulation (follow instructions)
Develop Precision
Articulation (combine, integrate related skills)
Naturalization (automate, become expert)
Kognitiv
Det kognitive område handler om evnen til at "tænke et område igennem". Det kognitive kredser om viden og forståelse for et emne.
Blooms taksonomi arbejder med seks klasser, hvor de højere niveauer inkorporerer de lavere.
Lorin Anderson (en tidligere student hos Bloom) opdaterede Blooms taksonomi.
Læg mærke til skiftet fra navne- til udsagnsord. (OG dermed fra produkter til processer)
Remembering: can the student recall or remember the information? – define, duplicate, list, memorize, recall, repeat, reproduce state
Understanding: can the student explain ideas or concepts? – classify, describe, discuss, explain, identify, locate, recognize, report, select, translate, paraphrase
Applying: can the student use the information in a new way? – choose, demonstrate, dramatize, employ, illustrate, interpret, operate, schedule, sketch, solve, use, write.
Analysing: can the student distinguish between the different parts? – appraise, compare, contrast, criticize, differentiate, discriminate, distinguish, examine, experiment, question, test.
Evaluating: can the student justify a stand or decision? – appraise, argue, defend, judge, select, support, value, evaluate
Creating: can the student create new product or point of view? – assemble, construct, create, design, develop, formulate, write.
Læg mærke til at de to nederste niveauer (evaluating og creating) er byttet om i forhold til den gamle version.
Henvisninger
Generelle
Taxonomy of Educational Objectives: The Classification of Educational Goals, B. S. Bloom (Ed.) David McKay Company, Inc. 1956.
Ledelse og uddannelse – Grundbog, Forsvarskommandoen 1998.
Pædagogik | danish | 0.819379 |
Subsets and Splits
No community queries yet
The top public SQL queries from the community will appear here once available.